Four-State Dairy Nutrition and Management Conference
|
|
|
- Rolf Bruce
- 6 years ago
- Views:
Transcription


1 Four-State Dairy Nutrition and Management Conference June 9 & 10, 2010 Dubuque, Iowa Cooperative Extension for: Iowa State University University of Illinois University of Minnesota University of Wisconsin
2 Table of Contents Sponsors and Speakers Getting Cows Pregnant and Keeping Them Pregnant in Coordination with Practical Omega Nutrition Economic Returns of Improved Reproductive Performance in Dairy Cattle Where Do All These Fatty Acids Come From And What Do They Do To My Cow? The Compatibility between Diary Productivity and Carbon Footprint Feeding Economics For State Dairy Nutrition & Management Conference Coordination of Reproductive and Nutritional Management of Lactating Dairy Cows Demystifying the Environmental Sustainability of Food Production The Use of Records to Evaluate and Improve Transition Cow Performance Comparison of Immune Function, Uterine Health and More in Holstein and Crossbred Transition Cows Manureology Troubleshooting Silage Yeast, Mold and VFA Problems Improving Feeding Consistency Through TMR Audits The Chemistry of High Moisture Corn Improving Starch Digestibility in Dairy Cows: Opportunities with Reduced-Starch Diets FARM Dairy Well-Being Program Dairy calf liquid feeding strategies to cope with new antibiotic use regulations Improving the Value of Cull Cows The Wisconsin Dairy Feed Cost Evaluator Improving Reproductive Efficiency using Double Ovsynch Food Economics and Consumer Choice
3 Sponsors and Speakers The program committee deeply appreciates the following for their support and commitment to strengthening the Midwest dairy industry. Platinum Co-Sponsors Elanco Animal Health Virtus Nutrition Gold Co-Sponsors Analab Dairyland Laboratories Diamond V Mills Kemin Industries Nutriad, Inc. Pioneer, A DuPont Business Quali Tech, Inc. Vi-COR Silver Co-Sponsors Adisseo Ag Processing, Inc./Amino Plus AgSource Cooperative Services Alltech, Inc. AMTS Arm & Hammer Animal Nutrition Balchem Corporation BASF Plant Science BIOMIN USA Biozyme Incorporated Byron Seeds, LLC Central Life Sciences Citura Cumberland Valley Analytical Services Digi-Star LLC Galdwin A Read Co. Milk Specialties Global Animal Health MIN-AD Inc. Mycogen Seeds Novus International, Inc. Papillon Agricultural Company Prince Agri Products, Inc. Quality Roasting Inc. Rock River Laboratory Inc. SoyBest SoyPLUS/Soy Chlor Supervisor Systems Trouw Nutrition USA Zeeland Farm Services, Inc. Zinpro Corp. Bronze Co-Sponsors Agri-Nutrition Consulting Inc. Calf-Tel By Hampel CHR Hansen Lallemand Specialties Speakers: Bill Thatcher University of Florida Mike Overton University of Georgia Tom Jenkins Clemson University Jude Capper Washington State Mike Hutjens University of Illinois Jim Barmore GPS Dairy Consulting Ed Kreykes Dairy Health Services Doug Scheider Scheidairy Farms Inc. Gary Sipiorski Vita Plus Corp. Ricardo Chebel University of Minnesota Bill Mahanna Pioneer Rob Aukerman Elanco Animal Health Tom Oelberg Diamond V Mills Pat Hoffman University of Wisconsin Randy Shaver University of Wisconsin Marcia Endres University of Minnesota Noah Litherland University of Minnesota Dick Wallace University of Illinois Victor Cabrera University of Wisconsin Extension Milo Wiltbank University of Wisconsin
4 Getting Cows Pregnant and Keeping Them Pregnant in Coordination with Practical Omega Nutrition W.W. Thatcher, C.R. Staples and J.E.P. Santos Department of Animal Sciences, University of Florida Introduction Intensive genetic selection for milk production without attention to reproductive performance has contributed to an inverse relationship between milk production and reproduction (Lucy, 2001). Inclusion of Productive Life, Daughter Pregnancy Rate, and more recently the availability of Sire Conception Rate, as a measurement of phenotypic service-sire fertility, appear to have reduced the rate of decline in fertility in the USA (Norman et al., 2007). Reproductive management of the lactating dairy cow has been a challenge because of poor expression of estrus and low fertility to insemination at a detected estrus. The duration of estrus is reduced as milk production increases, and the frequency of double ovulations and subsequent occurrence of twins also is increased in cows with high levels of milk production at the time of the breeding period (Lopez et al., 2005) The high producing dairy cow of today expresses estrus for approximately 7 hours during which time an average of 6.5 standing events takes place with an accumulative period of standing of 20 seconds (i.e., 3 seconds per standing event; Lopez et al., 2004). Pregnancy rate over a 21 day period for the national herd of dairy cows in the United States is approximately 16.2%. The component parts of pregnancy rate are the rate of estrus detection and conception rate. Technology is available for systems to detect estrus accurately, but a major issue is that lactating dairy cows don t display strong symptoms of estrus. Expression of estrus has been affected adversely by high milk production and associated metabolism of hormones, as well as housing facilities (e.g., concrete floors) that reduce the cow s willingness to be sexually active. An additional challenge is the high occurrence of non-ovulatory dairy cows that either have re-occurring follicle waves without ovulation or development of ovarian cysts. A major advance in reproductive management that has addressed how to improve pregnancy rate has been development of timed artificial insemination (TAI) programs based on development of systems to control or program optimal development of ovarian follicles, induce ovulation, and develop a corpus luteum (CL) capable of supporting pregnancy (Moore and Thatcher 2006). The component pharmaceutical agents available to the dairy industry in many countries for use with dairy cattle are GnRH, luteolytic prostaglandins, and intravaginal progesterone (using a controlled internal drugreleasing insert, CIDR, or similar device). These are pharmaceuticals that mimic the actions of the cow s endogenous hormones, are physiological, and pose no health hazard to the cow. The original TAI protocol is the Ovsynch procedure (Pursley et al., 1997a). This protocol has been in use for approximately 12 years. During this period, both basic and applied research has lead to major advancements in optimizing the system. As a consequence, pregnancy responses have increased, the system has been extended to resynchronization of nonpregnant cows, and programs have been developed for TAI in dairy heifers. The dynamics of various cow factors such as body condition score, parity, and health status in the transitionalperiparturient period have been shown to influence pregnancy rates to the controlled breeding program. The present objective is to update major advancements in ìgetting cows pregnant and keeping them pregnantî and coordination of reproductive management with practical omega nutrition. Lactating Dairy Cattle It is essential that nutritionists, producers and veterinarians understand the physiological underlying reasons why certain components of the reproductive management program are able to improve reproductive performance or conversely why a misunderstanding of the program can lead to catastrophic pregnancy results. No one reproductive breeding program is practical and economically optimal for all dairy production units due to differences in available facilities, size of the unit, labor that places reproduction as a high priority, and a functionally dynamic record system. Optimizing stage of the estrous cycle at onset of Ovsynch The original Ovsynch program involved two injections of GnRH administered 7 days before and 1
 at the onset of the Ovsynch protocol is important to achieve a subsequent synchronized ovulation at the second GnRH preceding the TAI (Figure 1).")
5 48 hours after an injection of PGF2a, and cows are inseminated h after the second injection of GnRH (Pursley et al., 1997a). If TAI in the OvSynch protocol is performed at the same time as the second GnRH injection than the protocol is referred to as Co- Synch.Optimization of stage of the estrous cycle (i.e., days 5 to 9) at the onset of the Ovsynch protocol is important to achieve a subsequent synchronized ovulation at the second GnRH preceding the TAI (Figure 1). Programming the stage of the estrous cycle at the time the Ovsynch protocol is implemented (e.g., Days 5-9 of estrous cycle) insures there is progesterone availability throughout the period between the first injection of GnRH and injection of PGF2a, and that there is a CL to respond to the luteolytic injection of PGF2a (Figure 1). The continual exposure to progesterone is important for sequentially programming the brain, oviduct and uterus with the appropriate changes in hormones, receptors and secretions leading to an induced ovulation, fertilization and development of an embryo capable of maintaining a pregnancy with minimal embryonic and fetal losses. Programming the start of the Ovsynch protocol to occur between Days 5 to 9 of the estrous cycle increases the probability that the first injection of GnRH will induce ovulation of the first wave follicle and recruitment of a new follicle wave (Figure 1), which upon induction of ovulation to the second GnRH increases the probability of producing a viable oocyte for fertilization and a robust CL. Indeed ovulation of the first follicle wave results in presence of both the original CL and an accessory CL, induced by the GnRH injection, which are responsive to the injection of PGF2a. The Ovsynch protocol preceded by a PGF2a presynchronization program (Presync-Ovsynch) has become the nucleus program for reproductive management in the industry (Moreira et al., 2001; El- Zarkouny et al., 2004). Successful use of such a program is dependent highly upon obtaining good compliance in implementing all component parts of the protocol. The original Presynch-Ovsynch program entailed two injections of PGF2a given 14 days apart with the Ovsynch protocol initiated 12 days after the second injection of PGF2a for presynchronization (Figure 2). This system increased pregnancy rates compared to Ovsynch alone for the reasons outlined above, when the Ovsynch protocol is initiated in early diestrus (Table 1). Dairy producers were keen to extend the period when Ovsynch was initiated to a 14 day interval such that four of the five sequential hormonal injections would be given on the same day of week. Field experiences indicate that 60% of detected estruses occur on days 3-6 after the second injection of PGF2a of presynchronization. A recent study indicated that an 11 day interval after presynchronization (i.e., cows would be predominately 5-8 days of the estrous cycle) is better than a 14 day interval to begin the TAI protocol (Galv o et al., 2007). The overall ovulation rate to the first injection of GnRH was greater for an 11 day than a 14 day interval (62% > 44.7%). This was attributed to GnRH being given at 11 days when the first wave follicle will ovulate whereas the 14 day interval increased the proportion of cows injected early in the second follicle wave at a time the follicle was developed insufficiently to ovulate in response to GnRH. The latter follicle would continue to develop and be slightly more aged and/or dominant compared to the newly recruited follicle from the day 11 injection interval for GnRH. Indeed Pregnancy per TAI was 6.6% greater for the 11 day interval (40.1% > 33.5% at day 38 after TAI; Table 1). Thus subtle changes in presynchronization protocols can cause substantial increases in pregnancy rate and the optimal period to start the Ovsynch protocol is 10 to 12 days after the second PGF2a injection of presynchronization (Figure 2). 2
6 Table 1. Pregnancy rates for lactating dairy cows receiving various reproductive management systems for timed insemination Interval from PGF2a to ovulatory injection of GnRH and timing of AI It has been well documented that cows should be inseminated 8 to 16 hours after the onset of estrus for an optimal conception rate (Pursley et al., 1997a). The preovulatory surge of LH occurs very close to the onset of estrus with ovulation occurring approximately 28 h after the LH surge. It is important to recognize that the second injection of GnRH of an Ovsynch program is analogous to the onset of estrus since an LH surge is induced immediately. Indeed maximal rate of pregnancies per AI was achieved when a timed insemination was made at 16 h after the injection of GnRH. In contrast percent pregnant to AI was decreased when inseminations were made at the time of GnRH injection or 28 h later. Producers often favor the convenience of carrying out a TAI at the time of GnRH injection (i.e., referred to as a Co-synch program) to reduce the number of times cows need to be held up. Alternatively, some producers prefer to perform TAI on the following day at approximately h after the GnRH injection for convenience. Either option likely will reduce percent pregnant to AI. The importance of the correct timing is indicated by a study completed at the University of Wisconsin (Brusveen et al., 2008). All cows were presynchronized with two injections of PGF2a, and the Ovsynch protocol was started 11 days later. The optimal timing program was to inject GnRH 56 h after injection of PGF2a and inseminate the cows 16 h after GnRH, which was 72 h after the injection of PGF2a (e.g., Figure 2). Percent pregnant to AI was 36.1% compared to a Co-synch 48h (26.7%) or 72 h (27.3%) programs. The latter two programs injected GnRH and TAI concurrently at 48 h or 72 h, respectively. Clearly, subtle changes in timing of the GnRH injection and time of insemination result in substantial differences in percent pregnant to AI responses. If a Co-synch program is to be followed, one needs to understand the physiology of the injection sequence so that functionally active ovarian follicles are at an optimal stage analogous to a follicle in the close periestrus period when GnRH/TAI is performed. Application of a Presynchronization/ 5-Day Co- Synch Program in Lactating Dairy Cows High-producing lactating dairy cows have a greater incidence of two waves of follicle growth during the estrous cycle compared with growing heifers that are more likely to have three follicular waves (Savio et al.,1988). The interval from follicle emergence to estrus is ~3.5 days greater for cows with two follicular waves than for those with three follicular waves (Bleach et al., 2004). In a comparison of Ovsynch programs initiated at day 3 or 6 of the estrous cycle, Cerri et al. (2009) reported that ovulatory follicles with shorter length of dominance (5 to 6 days) yielded a greater proportion of grades 1 and 2 embryos, whereas cows with a longer period of dominance had increased proportion of poor quality and degenerated embryos. Fertilization rate remained unaffected by the period of dominance. One means of reducing the period of ovulatory follicle dominance is to shorten the interval from follicle recruitment to luteal regression (i.e., implement a 5 day interval between GnRH and PGF2a injection) to possibly increase pregnancy per TAI in lactating dairy cows. Following two injections of PGF2a at 36 and 50 d in milk, Santos et al. (2010) randomly assigned 933 cows to a Cosynch 72 h protocol (CoS72: d 61 GnRH, d 68 PGF2a, d 71 GnRH) or to a 5d-Cosynch 72 h with two injections of PGF2a (5dCoS2: d 61 GnRH, d 66 and 67 PGF2a, d 69 GnRH). Blood was sampled for progesterone analyses at the first GnRH, first PGF2a, second GnRH and 7 day after timed AI. Regression of CL was lesser (91.5 vs. 96.3%) and P/AI was greater (39.3 vs. 33.9%) for 5dCoS2 than CoS72, respectively. It was essential to inject two doses of PGF2a given 24 h apart (i.e., d 66 and 67) to insure complete regression of the CL, whereas in dairy heifers described above CL regression was completed with one injection of PGF2a. Following two injections of PGF2a at 46 and 60 days in milk, Bisinotto R. and Santos J.E.P (Unpublished observations, 2010) randomly assigned 1227 lactating dairy cows to a 5-day OVS56h or to a 5- daycosynch72h as depicted in Figure 3. Pregnancy/TAI did not differ between groups when evaluated at either d32 or d 60 after TAI (Table 2). Indeed, overall mean Pregnancy/TAI of 45.9% (n=1227) at d32 is excellent with overall pregnancy loss estimated d 60 to be 13.1% (n=557). Thus, the 5- daycosynch 72 h program with two injections of PGF2a is very efficient in getting cows pregnant. 3
7 Re-synchronization of nonpregnant cows following first service A reproductive management challenge following first service is to re-inseminate cows that did not conceive as quickly as possible (Bartolome et al., 2005). The same principles to optimize the Presynch- Ovsynch program are applicable to development of a resynchronization program. However, a resynchronization system is somewhat constrained in that programming nonpregnant cows to ovulate must be done in a manner that will not interfere with cows that are pregnant to first service. Thus, accurate identification of non-pregnant and pregnant cows is important, and timing of the diagnosis is dependent upon the technology applied (i.e., rectal palpation at days, ultrasound diagnosis at days, blood pregnancy test 27, 28 to 30 days [measurement of Pregnancy Associated Glycoprotein; PAG]; Silva et al., 2007). To some degree, there is a natural presynchronization of non-pregnant cows because those detected in estrus have a median return to estrus interval of 22 days in which 64.3% show estrus within days after first service. Thus, initation of Ovsynch at 30 days after first service would mean most cows would be at approximately Day 8 of the cycle. GnRH injection would induce ovulation of a first wave follicle and initiate recruitment of a new follicular wave under a high progesterone environment. At 37 days after first service, a decision can be made to inject PGF2a in cows diagnosed nonpregnant (e.g., rectal palpation). These cows would then be injected with GnRH and TAI at 56 h and 72 h after the PGF2a injection, respectively. Several days after first service (days 19, 26 and 33) have been examined to begin a resynchronization of nonpregnant cows with Ovsynch (Fricke et al., 2003). Starting the resynchronization on day 33 resulted in the highest pregnancy rate for the second service. Ultrasound technology was used for detection of non pregnant cows at 26 or 33 days after first service for the day and 33 resynchronization groups, respectively. Hypothetically, the timing of GnRH at day 26 would tend to target the majority of cows too early in their follicle wave (i.e., day 4 of the wave) to induce follicle turnover, whereas at day 33 they would be ovulating potential first or second wave follicles and a sustained progesterone environment would be present for cows potentially returning to estrus between 17 to 24 days after first service. Experimental results clearly document that fertility was increased for the Day 33 resynchronization group (i.e., 33.7%) compared to the Day 19 and 26 groups (27.1% and 26.6%, respectively). The benefit of the day 33 resynchronization on pregnancy per TAI compared to the day 26 resynchronization group was repeated (39.4% > 28.6%; Table 1) with the benefit most apparent in primiparous cows (Sterry et al., 2006). In the latter study, insertion of a CIDR insert in cows without a CL improved pregnancy rate per TAI in the multiparous cows to a level comparable to that of primiparous with or without a CL. An alternative resynchronization strategy is a more conventional system based solely on pregnancy diagnosis per rectal palpation at day 38 (Figure 4; Dewey et al., 2009). In this scenario, an Ovsynch 72 h Co-synch (GnRH, 7 d later PGF, and 72 h later GnRH and TAI) was initiated at day 38 after first service in three groups of non-pregnant cows (Group 1 control, GnRH/Group 2 received a GnRH injection on Day 31 at 7 days before pregnancy diagnosis, Group 3/CIDR received a CIDR insert on D 38 that was removed at the time of PGF2a injection; Figure 4). Pregnancy rate per TAI was and tended to be greater for GnRH/Group 2 (33.6%) and Group3/CIDR (31.3%) cows, respectively, than Group 1 (24.6%) cows. It is likely that presynchronization with a single injection of GnRH at Day 31 programmed a new follicle wave and increased the occurrence of a CL at the beginning of the Ovsynch 72 h Cosynch protocol. Insertion of a CIDR insert likely improved the synchronization of ovulation associated with the 72 h Co-synch response because it held ovarian follicles from ovulating prematurely in cows that were in late diestrus at the time the Ovsynch 72h program was started. 4
8 It is clear that several alternatives are available for resynchronization of lactating dairy cows. With the acquisition of new technology for a cow side diagnosis of nonpregnant cows early after insemination (e.g., 18, 27, 28 or 30 days), it will be possible to implement even earlier resynchronization systems for TAI within 3 days (e.g., d 21, 30, 31 or 33) after the diagnosis of a nonpregnancy. This would offer a further reduction interval to re-insemination of 9.5 to 17 days compared to the promising systems described above. Improving Embryo Survival: Induction of accessory CL with hcg after insemination It is clear that lactating dairy cows have lower plasma concentrations of progesterone due to greater progesterone metabolism by the liver. Associated with high dry matter intake during lactation and associated increases of liver blood flow steroid metabolism is increased. Strategies to increase progesterone production are a challenge. For example insertion of a CIDR device causes only a slight chronic increase in progesterone of approximately 1 ng/ml far less than concentrations of progesterone achieved with a functional CL. On Day 5 of the estrous cycle, granulosa cells of the dominant follicle contan LH receptors such that hcg will induce ovulation and formation of an accessory CL (Diaz et al., 1998; Schmitt et al., 1996a,b). Therefore, administration of hcg 5 d after AI has the potential to increase progesterone secretion during early pregnancy, and to alter ovarian follicular dynamics so that cows have three follicular waves within the period approaching the time of CL maintenance (Diaz et al., 1998). Injection of 3,300 IU hcg in lactating cows on Day 5 after AI increased the number of CL and substantially elevated plasma progesterone concentrations (Santos etal., 2001). Conception rates on Days 28, 42, and 90 were increased by hcg treatment, but late embryonic and fetal losses remained unaltered. Therefore, the positive effect of hcg on conception rates was mediated by reducing early embryonic losses. In addition, most of the benefit of hcg treatment was observed in lactating dairy cows that were losing body condition during the breeding period and were likely those cows producing more milk. Furthermore, use of hcg should be targeted to populations of cows that are subfertile such as the high producing, lactating cow that is losing body condition. It is important to recognize that use of GnRH, as opposed to hcg, is associated with a shorter duration of LH exposure, with the induction of an accessory CL that is less responsive to LH in vitro, and a substantially lower increase in plasma progesterone concentrations during the subsequent luteal phase (Schmitt et al., 1996b). Recent study from Kansas (Stevenson et al., 2007) reported that hcg effectively induced ovulation between 4 to 9 days after insemination, increased number of CL and serum P4, and increased conception rates but only in some herds. In contrast GnRH failed to increase progesterone or alter conception rate. Integration of Reproductive Management with Omega Nutrition Collectively, the above experimental approaches, which have fine tuned the dynamics of follicle development, CL development and regression, and timing of ovulation and insemination, result in good on farm responses in getting cows pregnant. Development of such programs provides a platform to address what sources of variation are contributing to animals that do not conceive following the periparturient and postpartum periods. A major nutritional focus has been undertaken in lactating dairy cows involving the feeding of supplemental fats and has been reviewed extensively (Palmquist, 2009). We have recently, developed a program of differential feeding of supplemental fats that are proinflammatory during the transition period and antiinflammatory during the transition subsequent breeding period. Rationale and Design Feeding supplemental fatty acids (e.g., C18:2n-6 linoleic acid) during the transition period is a strategy to increase the energy density of the diet, but fatty acids could act also as a precursor for the biosynthesis of prostaglandins of the 2 series that exert a pro-inflammatory effect that may benefit postpartum health of the cow. Later on after 30 days postpartum period, it may be reasonable to feed fatty acids (e.g.,c20:5n-3 eicosapentaenoic) that lead to suppression in the the biosynthesis of inflammatory molecules. This would reduce possible residual inflammatory responses in the uterus associated with carryover effects of subclinical endometritis or as reported in repeat breeder cows, reduce the potential luteolytic peaks at the time that the conceptus is 5

9 suppressing PGF2a secretion in order to maintain the corpus luteum for pregnancy maintenance and amplify localized immune-suppression responses associated with early pregnancy. In a recent Florida study, Silvestre et al. (2008a, 2008b and 2008c) randomly allocated cows (n = 1,582) into two experimental transition diets beginning at approximately 30 days before the expected date of parturition and continued until 30 dpp. After 30 dpp cows within each transition diet were allocated randomly into the experimental breeding diets that were fed until 160 dpp. Experimental transition and breeding diets differed only in the source of supplemental FA. Transition diets consisted of CS of palm oil (PO; EnerGII) or CS of safflower oil (SO; Prequel 21) and breeding diets consisted of CS of PO (EnerGII) or CS of fish oil (FO, StrataG). All CS of FAs were manufactured by Virtus Nutrition (Corcoran, CA, USA) and supplemented at 1.5% of dry matter. Diets were formulated to meet or exceed NRC (2001) nutrient requirements for net energy of lactation (NE L ), crude protein (CP), fiber, mineral and vitamins and fed to obtain intakes of 200 and 400 g/d of CS of FAs, for pre- and postpartum cows, respectively. Diets were fed as a total mixed ration twice daily targeting 5% orts. Cows at 43 dpp began a Presynch protocol with two injections of PGF2a (25 mg, dinoprost tromethamine, i.m., Lutalyse ; Sterile Solution; Pfizer Animal Health, New York, NY) injected 14 days apart. The Ovsynch protocol was initiated 14 days after the second injection of PGF2a of the Presynch with a GnRH injection (100 µg; gonadorelin diacetate tetrahydrate, i.m., Cystorelin, Merial Ltd., Athens, GA) followed 7 days later by an injection of PGF2a and a final injection of GnRH 56 hours later. Timed artificial insemination (TAI) for first service was performed 16 hours after the second GnRH injection of the Ovsynch protocol. second service. All cows diagnosed pregnant after first and second services were re-examined by perrectum ultrasonography at 60 days after insemination to determine pregnancy losses. Pregnancy per AI, pregnancy losses, and cumulative proportion of pregnant cows after two services were analyzed using pre-determined statistical contrasts to test the effects of the transition diets (PO-PO + PO-FO vs. SO-PO + SO-FO), breeding diets (PO-PO + SO-PO vs. PO-FO + SO-FO) and the interaction of transition and breeding diets (PO-PO + SO-FO vs. PO-FO + SO- PO) accordingly with the experimental feeding design described above (Silvestre, 2008a, 2008c). Reproductive and Lactation Responses Transition, breeding and interaction of diets did not affect pregnancy per AI at 32 and 60 days after TAI for first service (Table 3). However, pregnancy loss from day 32 to day 60 after the first TAI was less (P < 0.05) in FO compared to PO supplemented cows during the breeding period (Table 3). For second service, breeding diet altered (P < 0.05) the 32 day estimates of pregnancy per AI and a tendency (P < 0.10) for an interaction was detected between transition and breeding diets (Table 3). The increase in day 32 pregnancy per AI caused by FO was greater in cows fed the SO transition diet, whereas there was no increase in pregnancy per AI in cows fed the PO breeding diet regardless of transition diet (Table 3). Both breeding diet and a transition by breeding diet interaction (P < 0.05) were detected for the 60 day pregnancy per AI response in which FO stimulated pregnancy rate per AI but the response to FO was greater in cows fed the SO transition diet (Table 3). Table 3. First and second services pregnancies per AI at 32 and 60 days after insemination and pregnancy loss for experimental diets All cows received a controlled internal drug-releasing device (CIDR, EAZI-BREED; Pfizer Animal Health, New York, NY) containing 1.38 g of progesterone at 18 days after the first TAI followed 7 days later by removal of the CIDR device and an 100 µg injection of GnRH. At 32 days after first TAI, cows were examined for pregnancy by per-rectum ultrasonography to identify presence of an embryo and an embryonic heart beat. Non-pregnant cows were injected with 25 mg of PGF2a and then injected with 100 µg of GnRH 56 hours later. A TAI was performed 16 hours after the last GnRH for the second service. Cows were examined for pregnancy by per-rectum ultrasonography at 32 days after 6
10 Milk weights were recorded once a month for all cows. The single measurement of milk production for each month was considered as the average for the month. Data from the first 5 months of lactation were used. Average milk yield for combinations of transition and breeding diets were PO-PO (41.1 ± 0.6 Kg/day; n = 295), PO-FO (41.3 ± 0.7 Kg/day; n = 280), SO-PO (41.7 ± 0.6 Kg/day; n= 302) and SO-FO (42.1 ± 0.7 Kg/day; n = 289). Average milk yield was affected (P = 0.02) by transition diets such that cows supplemented with SO (41.9 ± 0.4 Kg/day; n = 591) during the transition period had a greater average milk yield for the 5 months postpartum compared with cow fed PO (41.2 ± 0.4 Kg/day; n = 575). Average milk yield was not affected by breeding diet or the interaction between transition and breeding diets. The integration of the disciplines of ruminant nutrition, reproductive physiology, immunology and clinical medicine has the potential to provide useful alternatives to improve postpartum health and fertility in dairy cows in a scenario of increasing milk production. For example sequential feeding of diets rich in linoleic acid followed by diets rich in eicosapentaenoic and docosahexaenoic fatty acids during the peri-parturient and breeding periods, respectively, impacted fatty acid composition of tissues, altered immune-responses and fertility (Silvestre et al. 2008a, 2008b and 2008c). Conclusions Tremendous advances have been made for improving milk production, but have in turn, resulted in an overall decline in reproductive efficiency for the dairy industry. Problems associated with the cow include inability to properly express estrus and altered hormonal profiles resulting in low conception rates and increased early embryonic death. Coordinated systems of reproductive management offer means to improve herd reproductive performance and major advances have been made for synchronization of ovulation in both lactating dairy cows and dairy heifers. Such systems are predicated on a greater understanding of the factors controlling follicle development, ovulation and CL development. Systems of reproductive management need to consider: efficient systems of presynchronization, induction of a new follicle wave in a high proportion of animals, optimization of the period of follicle dominance, sustained presence of a CL during the period of follicle recruitment, effective induction of CL regression, optimal timing of ovulation and timing of insemination. The programs as described require the producer, veterinarian and reproductive management staff to understand the programs and make the effort to obtain a high level of compliance. The platforms used for controlling first service and resynchronized subsequent services in cows that do not conceive provide valuable platforms to implement new technology such as the use of sexed semen, embryo transfer and cow side chemical diagnosis of nonpregnant cows. Functional and efficient computer record programs are essential to implement such reproductive management programs. The integration of the disciplines of ruminant nutrition, reproductive physiology, immunology and clinical medicine has the potential to provide useful alternatives to improve postpartum health and fertility in dairy cows in a scenario of increasing milk production. We propose that sequential feeding of diets rich in LN followed by diets rich in EPA and DHA during the peri-parturient and breeding periods, respectively can benefit overall cow performance and fertility. Such feeding strategies warrant economic analyses to evaluate cost-benefit. With the advent of new technologies to precisely manipulate reproductive function in lactating dairy cows, dairy producers are presented with a new opportunity. Coordination of management strategies to maximize both milk production and reproductive performance may optimize the economical return of dairy herds, and allow for the industry to take complete advantage of the genetic potential to improve milk production through artificial insemination. References Bartolome JA, Sozzi, A., McHale J, Melendez P, Arteche A C M, Silvestre FT, Kelbert D, Swift K, Archbald LF, Thatcher WW. (2005). Resynchronization of ovulation and timed insemination in lactating dairy cows, II: assigning protocols according to stages of the estrous cycle, or presence of ovarian cysts or anestrus. Theriogenology 63, Bleach ECL, Glencross RG, Knight P.G. (2004). Association between ovarian follicle development and pregnancy rates in dairy cows undergoing spontaneous oestrous cycle. Reproduction 127: Brusveen DJ, Cunha AP, Silva CD, Cunha PM, Sterry RA, Silva EPB, Guenther JN, Wiltbank MC. (2008). Altering the time of the second gonadotropin-releasing hormone injection and artificial insemination (AI) during Ovsynch affects pregnancies per AI in lactating dairy cows. J. Dairy Sci. 91, Cerri RLA, Rutigliano HM, Chebel RC, Bruno RGS, Santos JEP. (2009). Period of dominance of the ovulatory follicle during synchronization programs influences embryo quality. Reproduction 137: Dewey ST, Mendonça LG, Lopes Jr G, Rivera FA, Guagnini F, Chebel RC and Bilby TR. (2009). Resynchronization strategies to improve fertility in lactating dairy cows utilizing a presynchronization injection of GnRH or supplemental progesterone: I. Pregnancy rates and ovarian responses. J. Dairy Sci. 92: E-Suppl. 1 (Abstract): 267. [RTF bookmark start: }_Ref
11 Diaz T, Schmitt EJ-P, De La Sota RL, Thatcher M-J, Thatcher WW. (1998). Human chorionic gonadotropin-induced alterations in ovarian follicular dynamics during the estrous cycle of heifers. J. Animal Sci. 1998;76: [RTF bookmark end: }_Ref El-Zarkouny SZ, Cartmill JA, Hensley BA, Stevenson JS. (2004). Pregnancy in dairy cows after synchronized ovulation regimens with or without presynchronization and progesterone. J. Dairy Sci. 87, Fricke PM, Caraviello DZ, Weigel KA, Welle ML. (2003). Fertility of dairy cows after resynchronization of ovulation at three intervals following first timed insemination. J. Dairy Sci. 86, Galv o KN, SaFilho MF, Santos JEP. (2007). Reducing the interval from presynchronization to initiation of timed artificial insemination improves fertility in dairy cows. J. Dairy Sci. 90, Lopez H, Caraviello DZ, Satter LD, Fricke PM, Wiltbank MC. (2005). Relationship between level of milk production and multiple ovulations in lactating dairy cows. J. Dairy Sci. 88, Lopez H, Satter LD, Wiltbank MC. (2004). Relationship between level of milk production and estrous behavior of lactating dairy cows. Anim. Reprod. Sci. 81, Lucy MC. (2001). Reproductive loss in high-producing dairy cattle: where will it end? J. Dairy Sci. 84, Moore K, Thatcher WW. (2006). Major advances associated with reproduction in dairy cattle. J. Dairy Sci. 89, Moreira F, Orlandi C, Risco CA, Mattos R, Lopes F, Thatcher WW. (2001). Effects of presynchronization and bovine somatotropin on pregnancy rates to a timed artificial insemination protocol in lactating dairy cows. J. Dairy Sci. 84, Norman HD, Wright JR, Hubbard SM, Kuhn MT, Miller RH. (2007). Genetic Selection for Reproduction: Current Reproductive Status of the National Herd; Application of Selection Indexes for Dairy Producers. In: Proc. Dairy Cattle Reproductive Conference, Eds. W.W. Thatcher and E.R. Jordan, Dairy Cattle Reproductive Council, Hartland, WI, pp Palmquist DL. (2009). Omega-3 fatty acids in metabolism, health, and nutrition and for modified animal product foods. The Professional Animal Scientist 25: Pursley JR, Kosorok MR, Wiltbank MC. (1997a). Reproductive management of lactating dairy cows using synchronization of ovulation. J. Dairy Sci. 80, Santos JEP, Narciso CD, Rivera F, Chebel RC, Thatcher WW. (2010). Effect of reducing the period of follicle dominance in a timed AI protocol on reproduction of dairy cows. J. Dairy Sci. (In Press). Santos JEP, Thatcher WW, Pool L, Overton MW. (2001). Effect of human chorionic gonadotropin on luteal function and reproductive performance of high-producing lactating Holstein dairy cows. J. Animal Sci.:79: Savio JD, Keenan L, Boland M P, Roche JF. (1988). Pattern of growth of dominant follicles during the oestrous cycle of heifers. J. Reprod. Fertil. 83, Schmitt EJ-P, Barros CM, Fields PA, Fields MJ, Diaz T, Kluge JM, Thatcher WW. A cellular and endocrine characterization of the original and induced CL after administration of a gonadotropin-releasing hormone agonist or human chorionic gonadotropin on day five of the estrous cycle. J Anim Sci 1996a;74: [RTF bookmark start: }_Ref [RTF bookmark end: }_Ref Schmitt, E.J.-P., T. Diaz, C.M. Barros, R.L. de la Sota, M. Drost, E.W. Fredriksson, C.R. Staples, R. Thorner and W.W. Thatcher. 1996b. Differential response of the luteal phase and fertility in cattle following ovulation of the first-wave follicle with human chorionic gonadotropin or an agonist of gonadotropin-releasing hormone. J. Anim. Sci. 74: Silva E, Sterry RA, Kolb D, Mathialagan N, McGrath MF, Ballam JM and Fricke PM. (2007). Accuracy of a pregnancyassociated glycoprotein ELISA to determine pregnancy status of lactating dairy cows twenty-seven days after timed artificial insemination. J. Dairy Sci. 90, Silvestre F. T., T. S. M. Carvalho, N. Francisco, J. E. P. Santos, C. R. Staples, W.W.Thatcher. Effects of Differential Supplementation of Calcium Salts of Fatty Acids (CSFAs) on Dairy Cows. J. Dairy Sci., 91:(suppl. 1), p. 76, 2008a. Silvestre F.T., T.S.M. Carvalho, C. Crawford, J.E.P. Santos, C.R. Staples, W.W. Thatcher. 2008b. Effects of Differential Supplementation of Calcium Salts of Fatty Acids (CSFAs) to Lactating Dairy Cows on Plasma Acute Phase Proteins and Leukocyte Responses: Phagocytic and Oxidative Burst, CD62L and CD18 Expression and Cytokine Production. Biol. Reprod., special issue, p.158. Silvestre, F.T. 2008c. Nutraceutial and Hormonal Regulation of Immunity, Uterine Health, Fertility, and Milk Production of Postpartum Dairy Cows. Doctorate Dissertation. Univ. of Florida, Gainesville. Sterry RA, Welle ML and Fricke PM. (2006). Effect of interval from timed artificial insemination to initiation of resynchronization of ovulation on fertility of lactating dairy cows. J. Dairy Sci. 89, Stevenson JS, M. A. Portaluppi, D. E. Tenhouse, A. Lloyd, D. R. Eborn, S. Kacuba, and J. M. DeJarnette Interventions After Artificial Insemination: Conception Rates, Pregnancy Survival, and Ovarian Responses to Gonadotropin-Releasing Hormone, Human Chorionic Gonadotropin, and Progesterone. J. Dairy Sci. 90:
12 Economic Returns of Improved Reproductive Performance in Dairy Cattle Introduction Poor reproductive efficiency is a significant source of economic loss affecting many dairies. Reproductive inefficiency results in economic losses through a variety of ways. Cows that fail to become pregnant during the breeding period are culled once milk production has declined below economically viable levels, forcing the replacement of otherwise healthy animals. Cattle that eventually become pregnant, but at a much later time than desired, spend a disproportionate amount of their lactation at lower levels of milk production, costing the dairy lost opportunity for addition marginal milk. Historically, dairy managers and consultants have used calving interval or days open as indices for evaluation of reproductive performance and the optimal calving interval for most cows has been considered to be in the range of 12 to 13 months, yielding a lactation length of approximately 10.5 to 11 months. This belief has been supported by other economic models examining the effect of days open and reproductive success on economic returns of dairies. 1-3 However, often, these assumptions do not fully consider the transition related culling risk, the potential differences in lactation curve shape of today s modern dairy cow, or the true cost associated with cows that fail to become pregnant and thus are removed from the herd. The use of calving interval or average days open are biased estimates of reproductive performance since they only reveal information about the successful cows and fail to consider the cows that never conceive, and thus, are removed from the herd as non-pregnant cows. Average days open only gives the interval from calving to conception for cows that have successfully conceived. In most cases, days open gives no information regarding the status of the non-pregnant animals other than potentially their current days in lactation or a projected minimum days open. Calving interval is even more limited in that it only considers the cows that became pregnant and maintained that pregnancy throughout a full gestation. Pregnancy rate (PR), defined as the historical rate at which eligible cows became M. W. Overton DVM, MPVM University of Georgia, College of Veterinary Medicine Department of Population Health Rhodes Center - ADS 425 River Road Athens, GA moverton@uga.edu 9 pregnant each 21-d cycle (# pregnant/ # of eligible cow-cycles) and examined over a sequence of 21-d cycles, is the preferred parameter for evaluating reproductive performance. To be considered as eligible or at risk for PR, a cow must be past the voluntary waiting period (VWP), not already pregnant at the start of the 21-day period of interest, must have a known outcome at the end of the period under consideration, and must be present and eligible for pregnancy for at least half of the 21-day period. A do-not-breed (DNB) is a code that is assigned to cows that are destined to be culled. These cows are no longer considered eligible for breeding or pregnancy and are removed from the future reproductive statistics. PR is more sensitive to detecting recent changes in reproductive performance and provides useful information since both pregnant and non pregnant cows that meet the eligibility criteria are considered in the calculation. 4 Based on database surveys as reported by Steve Stewart, Bruce Clark, Don Niles, Stephen LeBlanc and David Galligan (personal communications), PR in the U.S and Canada appears to average between 14-16%. Yet, many dairy advisors consider a PR of 25-30% to be the ultimate goal for optimum reproductive management. With the large difference between the average PR and the goal for PR, there is significant room for improvement. There are many ways of improving herd reproductive efficiency and many non-breeding factors that dramatically influence reproductive outcomes, but essentially it comes down to improving insemination risk (IR), conception risk (CR), or preferably, both. Insemination risk is defined as the number of animals inseminated divided by the number of animals eligible (same eligibility definition as discussed previously for PR). For IR, it does not matter whether the animal was inseminated via estrus detection or by using some form of timed AI. Conception risk is defined as the number of animals that were found pregnant divided by the total number of cows (inseminations) either pregnant or open; hence, to calculate conception risk, the outcome must be known. The objective of this paper is to
13 explore the potential value generated as a result of improving PR via a variety of methods and to examine the various drivers of economic returns associated with changes in PR, as well as illustrating how much a dairy may be able to spend in order to try and achieve an improved PR. Model Building and Assumptions The original simulation model was built using Excel spreadsheets simulation software and has been modified numerous times in order to refine it and to examine new factors. Distributions describing CR and IR (almost exclusively from estrus detection) were fit from data obtained from approximately 95 herds representing approximately 150,000 cows (Niles, et al. and other California dairy herds) and are used to mimic the normal variation seen between and within dairies. Daily milk and 305 day mature equivalent milk production estimates were also obtained from a variety of dairies and used to fit lactation persistency curves based on day in milk (DIM). Milk price estimates, cull cow values, market cow values, labor estimates, and other key inputs were derived from either published work or adapted from actual herd data. Culling risks over the entire lactation period were obtained from actual herd Dairy Comp 305 records and mathematically adjusted from 30-day to 21-day intervals to be consistent with the breeding cycles. All values of change in PR are obtained by comparison with a baseline program. The baseline breeding program is a simple estrus detection-based reproductive program with CR and IR distributions at each 21-day interval following a 50-day voluntary waiting period. The potential breeding period is day cycles for a total of 252 days of breeding. In other words, cows are eligible for breeding from 50 days in milk until 302 days in milk. Simulated PR s are obtained by multiplying randomly generated samples from the CR distribution and IR distribution. The user can manipulate the baseline PR by applying a multiplier to the sampled CR, IR or both, throughout the program, resulting in the desired PR for comparison. There are 3 breeding programs for comparison. The first is called an improved baseline program (IBP). This program is designed to mimic the changes that may be obtained as a consequence of improving CR, IR or both over the course of the breeding period. No synchronization protocols are included in this particular approach. This particular program was designed to estimate the value that may be obtained by simply doing a better job with a traditional estrus detection-based program. The second program that is used for comparison is a total timed AI program (TAI) and is based upon a Presync-Ovsynch with day-32 Resynch. Briefly, this approach includes an injection of prostaglandin F2 at 36+/- 3 days in milk, followed in 14 days with a second injection. After an additional 14 days, cows received an injection of GnRH to start the Ovsynch portion. In 7 days, another prostaglandin is given, followed in 56 hours with the final GnRH injection and a timed insemination hours later. No estrus detection is used. All cows are given an injection of GnRH at 32 days post-breeding. In 7 days, cows are examined via palpation per rectum and non-pregnant animals are given a prostaglandin injection and then proceed to complete the Resynch (Ovsynch) portion of the TAI. Following this schedule, all non-pregnant cows are re-inseminated every 42 days until the breeding period is concluded. The final program for comparison is a combination of estrus detection and TAI and is referred to as the modified Presynch program (MPS). Cows that follow this protocol receive a prostaglandin injection at 36+/- 3 days in milk and again at 50+/- 3 days in milk. Cows that are observed in estrus after the second injection are inseminated per normal farm routine. Cows that are not observed within 14 days start the Ovsynch program as previously described. Afterwards, all inseminations for the remainder of the breeding period are performed using estrus detection. Thus, the second breeding cycle is composed of two groups of cows - those that are inseminated via estrus detection and those that are inseminated by TAI, depending upon whether estrus was detected in the first 21 days following Presynch or not. In both the TAI and the MPS programs, all cows are assumed to incur the cost of the injections, as per the schedule, but due to less than perfect on-farm compliance, only 85-90% of cows initially enrolled are actually inseminated, depending on the compliance factor input into the model. The CR for each of these two programs is modeled as a function of the farm s baseline conception risk, the estimated proportion of cows that are truly cycling, expected distribution within the estrous cycle at the start of the program, and published reports involving TAI The PR from traditional breeding is obtained by taking the product of random samples from CR and IR for each cycle. All cycles are exactly 21 days long, all cows calve at the same time, and they are followed prospectively. There was no attempt to model the impact of abortion or seasonal effects on reproduction except as demonstrated by the impact from the original data set on CR and IR distributions. (It is assumed that CR results used in the model already reflect some of the expected embryonic 10
14 deaths since most herds are not palpated for pregnancy until approximately 40 days postinsemination and a large proportion of pregnancy wastage occurs by days.) Timed AI programs are assumed to have no effect on subsequent conception or estrus detection risk in non-pregnant cows and it is assumed that there are no differences in reproductive efficiency for any of the programs between parities of lactating cows, i.e., the results are expressed for the blended population on a per cow slot basis. The voluntary waiting period is 50 days and as cows move from the first cycle to the second, the proportion of cows expected to be cycling increases (the model decreases the proportion that are anestrus by 33% of the original proportion). Cows are palpated for pregnancy at approximately 39+/- 3 days post-breeding. Milk production, price of milk, and other economic values remain the same throughout the year. Milk production level may be utilized as a discrete variable to determine the impact at a certain 305ME milk production level, or stochastically (sampled from a distributions) to determine the average impact (and expected range of impacts) over many dairies. Reproductive performance for each of the 3 alternative breeding approaches is then compared to the original baseline program. Herd specific data that may influence on-farm profitability, including dry period length, calf death losses, culling risk across time, milk production, milk price, pharmaceutical costs, labor costs, and feed costs are entered. The model s inputs, herd-specific data, and pre-set distributions are linked to tables for each reproductive intervention and are used to estimate the average pregnancy rate over 252 days of potential breeding. The input table, herd-specific data, and pregnancy rate projections are linked to partial budgets (modifications of original work by Wolf and Dartt) to compare predicted economic returns resulting from changes in daily milk yield as a result of changes in reproductive performance. 12 Cows that are ultimately culled as non-pregnant, but that are milked successfully until then, are removed from the dairy at 600 to 750 days in milk. Stochastic modeling simulation software utilizes Monte Carlo sampling of the pre-set distributions and runs 1000 iterations. Results are then displayed as probability distributions, with a mean and 90% confidence interval. Annual herd turnover or culling risk may be dramatically impacted by changes in reproductive performance. Herds that get more cows pregnant have fewer cows that must be removed due to a failure to become pregnant. However, these same herds are also producing more female calves, and assuming proportional mortality risk across time, will have more replacement animals available to either sell, expand the herd, or replace a less profitable animal in the herd. If the latter option is chosen, the herd s culling risk will increase. In the model, all calves are sold as newborn calves and purchased back as needed. When reproductive performance improves, there are fewer cows that are forcibly removed due to reproductive failure. Consequently, the herd s apparent culling risk decreases. However, in the model, I assumed that the dairy would save on involuntary culling by retaining cows equal in number to one half of the increased number of pregnancies accrued. The other half would allow the dairy to cull some poor producers from the herd. As a consequence, the herd s culling risk would change as a consequence of the changes in reproductive performance. The economic value of the change in PR is estimated by use of simple partial budgeting approaches. Each new program is compared to the baseline program by transferring the various outputs into its own partial budget. The sources of revenue include predicted milk per cow per day over a year (as determined by the modeled herd s estimated average days in milk and the herd s lactation curve), the annualized value of the calves produced, and the annualized value of the culled cows. Subtracted from the revenues are a variety of expenses that include any additional replacement costs, the marginal feed consumed by cows to produce the marginal milk, additional feed consumed by additional non-lactating cows, the additional costs for housing, labor, and medical expenses, as well as any additional costs due to the change in reproductive management approach. Finally, the difference is adjusted for the time value of money. Since money received in the future is worth less than money received today, future returns have to be adjusted for when the returns actually occur. All of the revenues and expenses, and thus the net returns, are reported as dollars gained (or lost) per lactating cow slot on the dairy per year. Results and Conclusions The predicted results of 1000 model iterations comparing the benefits of improving PR by increasing IR by 10% (through improved estrus detection) over baseline are shown in figure 1. The starting average PR was 16% and the improved program s PR was approximately 18%. 11
 over 1000 iterations.")
, market cow value ($621), replacement heifer cost ($1700), heifer calf value ($250) and bull calf value ($15).")
15 Figure 1. Distribution of PR outcomes for improved program as a consequence of improving insemination risk (in this case, estrus detection) by 10% as compared to the baseline program. Figure 2 displays the predicted economic value of the PR improvement (average of approximately 2 units of PR change) over 1000 iterations. These results were taken from the fully stochastic model with the following input distributions for the baseline program (and their expected or average value): milk price ($12), CR (30%), IR (57%), herd level of milk production (23,000 lbs), market cow value ($621), replacement heifer cost ($1700), heifer calf value ($250) and bull calf value ($15). Overall, the improvement in IR resulted in a net of approximately $9 per unit change in PR, or a total return of approximately $18 per cow slot per year. Figure 2. Distribution of Predicted Returns per Unit Change in PR As a Consequence of Improving Insemination Risk by 10% (Mean = $18) yield a value of about $18 per cow slot per year as a result of the improved insemination risk. The total TAI approach also yields an improved PR, although the apparent increase is minor relative to the other approaches, but a word of caution is due here. Total TAI, as modeled in this scenario, yields a PR of almost 19% when using a VWP of 70 days (when the breeding actually starts) but for comparison sake, I maintained the same VWP as the other programs of 50 days. Hence, the lower than expected PR of only 16% is due in large part to the lack of breeding during the first potential cycle at DIM. These non-breeding days are used to presynchronize cows, yielding an improved CR. However, it comes at the cost of a delay to first service. Despite the increased cost of this approach, there is still a positive return of about $15 as compared to the baseline approach. The final approach was the backdoor Ovsynch protocol which included a Presynch series for all cows followed by a one-time use of Ovsynch only for cows that failed to be inseminated via estrus detection in the first cycle. This program incurs a larger cost with a lower rate of return as compared to the others, but is still an improvement compared to the baseline. The expected value of this approach would be greater had the program continued with additional Ovsynch-based breedings in later cycles. However, this hybrid approach was not modeled in this set of iterations. Figure 3. Estimated results and associated values of three different approaches to improving reproductive performance as compared to the baseline program. As mentioned previously, the model also allows for comparison of different approaches to improving reproductive efficiency. Each of these new approaches is compared with the original baseline program to estimate the value of the change after also considering the cost of implementing the new programs. The results are shown below in figure 3. Each program is expected to yield an improved PR but the magnitude of the improvement and the value of the change is different for each one. First, the improved baseline shows a similar result as above with the new PR of 17.5% which is predicted to Figure 4 illustrates a very important concept to remember when evaluating the economic returns of improving reproductive performance predicted returns follow a curvilinear relationship. In other words, the average return associated with improving PR varies depending upon the relative success of the starting point. In this series of scenarios, I calculated the predicted returns of increasing PR starting at a 10% baseline PR. The baseline PR of 10% was compared to improved PR across a range of values, derived by changing the CR and IR. At 10% PR, the value of improving PR by 2 unit (ie. 10% to 12% PR) is predicted to be worth approximately $54 per cow slot per year when milk was valued at $12/ cwt. Conversely, going from 18 to 20% is worth 12

16 approximately $14 and from 26 to 28% is worth only $2. Figure 4. Model Results of Value of Changes in PR over Ranges of PR of milk production = 25,000 lbs, market cow value = $0.46/ lb, replacement heifer cost = $1600, TMR cost = $210/ ton, heifer calf = $250 and bull calf = $15. For this set of scenarios, a 50-day voluntary waiting period was used and other than additional semen and insemination fees associated with an increased insemination risk, no additional reproductive management costs were assumed. As the value of milk increases, the value of the reproductive change increases within a range of PR change. For example, at $14 milk, increasing PR from 18% to 20% is predicted to yield an economic return of approximately $24 per cow slot or roughly about $12/ unit change in PR/ per cow slot per year. One of the benefits of using stochastic simulation models is the ability to perform sensitivity analyses of the major effect modifiers. Within the distributional ranges used in the model, the following variables had the largest impact on the economic value associated with reproductive performance change and each of the variables is listed in descending order of impact: 1) Insemination risk As insemination risk increased in the baseline model, the value of improving PR decreased. In other words, if the starting reproductive performance was high, there was less to be gained from further improvements. 2) Conception risk As conception risk increased in the baseline model, the value of each incremental change decreased just as with insemination risk. 3) Milk price As milk price increased, the predicted value of the change in reproductive performance increased. 4) Feed cost As feed cost increased, the value of change in PR decreased due to the reduced profit margin associated with the marginal milk produced. 5) Level of milk production As the herd level of milk production increased, the predicted value due to improving reproductive performance increased. 6) Replacement cost As the price for replacement heifers increased, the value realized by improving reproductive performance increased. 7) Market cow price As the value of the market cow increased, the value realized by improving reproductive performance decreased. The primary economic driver of improving reproductive performance is the value of the additional marginal milk produced as a consequence of decreasing days in milk for the herd. Figure 5 demonstrates the impact that milk price may have on the economic returns. For this example, each of the following inputs was entered as specific values and the only one to vary was the price of milk: herd level The concept of diminishing returns is nothing new. We see similar patterns in many biological systems. In the case of reproductive management, one must keep this issue in mind relative to making recommendations to clients. Herds that are already doing a good job reproductively have less potential economic value to be gained by improving performance even further. Likewise, if a herd s performance is already good with a baseline breeding program, one should carefully consider whether additional input costs will legitimately improve PR and return a profit to the dairy. In general, herds should work to improve basic semen handling and estrus detection prior to jumping on a TAI program. Other management issues such as compliance to protocol are also critical to the success of any program. Most herds have much to gain by improving reproductive performance. Improving PR should result in higher milk production, more pregnant cows, more calves, and reduced reproductive-based culling. Sensitivity analyses of model results reveal that insemination intensity, whether by estrus detection, timed AI or a combination, has the largest impact on reproductive performance. Efforts at improving reproductive success should first focus on maximizing the herd s basic estrus detection efficiency, due to its large impact on reproductive success and because it is more easily improved as compared to conception rate. Herds with very poor reductive efficiency have the most to gain by 13
17 improving PR, and within a given level of PR, the price of milk has the greatest effect on the value of the economic change, followed by the herd s level of milk production. Consequently, our emphasis in reproductive management should continue to be placed on improving insemination risk while at least maintaining conception risk. Although almost any herd can potentially benefit from synchronization programs, herds with poor reproductive performance are expected to realize the greatest potential return from improving reproductive performance, especially those herds with higher levels of milk production. References: 1. DeLorenzo MA, Spreen TH, Bryan GR, et al. Optimizing model: insemination, replacement, seasonal production, and cash flow. J Dairy Sci 1992;75: Hady PJ, Lloyd JW, Kaneene JB, et al. Partial Budget Model for Reproductive Programs of Dairy Farm Businesses. J Dairy Sci 1994;77: Meadows C, Rajala-Schultz PJ, Frazer GS. A spreadsheet-based model demonstrating the nonuniform economic effects of varying reproductive performance in Ohio dairy herds. J Dairy Sci 2005;88: Galligan DT, Ferguson JD, Dijkhuizen AA, et al. Optimal Economic Goals in Reproduction: Underlying Concepts from Semen to Culling in Proceedings of the National Reproduction Symposium 1994; Bartolome JA, Archbald LF, Morresey P, et al. Comparison of synchronization of ovulation and induction of estrus as therapeutic strategies for bovine ovarian cysts in the dairy cow. Theriogenology 2000;53: Cartmill JA, El Zarkouny SZ, Hensley BA, et al. Stage of cycle, incidence, and timing of ovulation, and pregnancy rates in dairy cattle after three timed breeding protocols. J Dairy Sci 2001;84: Keister ZO, Denise SK, Armstrong DV, et al. Pregnancy outcomes in two commercial dairy herds following hormonal scheduling programs. Theriogenology 1999;51: Moreira F, Orlandi C, Risco CA, et al. Effects of presynchronization and bovine somatotropin on pregnancy rates to a timed artificial insemination protocol in lactating dairy cows. J Dairy Sci 2001;84: Pursley JR, Mee MO, Wiltbank MC. Synchronization of Ovulation in Dairy Cows Using PGF2Alpha and GnRH. Theriogenology 1995;44: Risco CA, Drost M, Archbald L, et al. Timed Artificial Insemination in Dairy Cattle - Parts I and II. The Compendium 1998;20:s280-s287 (issue 210)-and (issue 1211). 11. Stevenson JS, Kobayashi Y, Thompson KE. Reproductive performance of dairy cows in various programmed breeding systems including OvSynch and combinations of gonadotropin-releasing hormone and prostaglandin F2 alpha. J Dairy Sci 1999;82: Wolf CA. Analyzing reproductive management strategies on dairy farms. Staff Paper 99-23, Department of Ag Econ, Michigan State University

18 Where Do All These Fatty Acids Come From And What Do They Do to My Cow? What are Fatty Acids? Fatty acids are chains of carbons that end in an acid group, or carboxyl group as it is referred to in biochemistry. An example of a common fatty acid is stearic acid with 18 carbons and no double bonds (Figure 1). Tom Jenkins Clemson University tjnkns@clemson.edu acid in animal fats and some plant oils, such as canola oil Linoleic acid is the predominant fatty acid in many plant oils, including cottonseed oil, soybean oil, and corn oil. Linolenic acid, with three double bonds, is the primary fatty acid in most pasture species and in linseed oil from flax. Fatty acids, such as stearic acid, are referred to as saturated because all the carbons are holding the maximum number of hydrogens possible, or the fatty acid is saturated with hydrogen. Stearic acid is low in plant oils, but present in higher amounts in animal fats, particularly in fats obtained from ruminant species such as beef tallow. Oleic acid and linoleic acid are examples of unsaturated fatty acids containing one or more double bonds (Figure 2). Oleic acid has a single double bond between carbons 9 and 10, and is referred to as a monounsaturated fatty acid. Linoleic acid is a polyunsaturated fatty acid containing two double bonds between carbons 9 and 10, and between carbons 12 and 13. Oleic acid is the predominant fatty Sources of Fatty Acids Grain and forage lipids The fatty acid content of most cereal seeds and forages typically ranges from 10 to 30 g/kg DM, with the majority of the fatty acids classified as unsaturated (predominately oleic, linoleic, and linolenic acids). Among the unsaturated fatty acids, linolenic acid is the predominant fatty acid in most forage species followed by linoleic acid (Hatfield et al., 2007). In the cereal seeds, fatty acids are comprised mainly of linoleic acid followed by oleic acid. Fatty acid concentrations in some pasture can exceed 50 g/kg DM, depending on plant species, stage of maturity, environment, etc. Fatty acid content of annual ryegrass pasture that was clipped in the field, immediately immersed in liquid nitrogen, and then freeze dried contained as much as 68 g/kg DM total fatty acids (Freeman-Pounders et al., 2009). CPM- Dairy lists 116 g/kg DM fatty acids in perennial ryegrass pasture. Cattle grazing some species of immature pasture, in effect, may be consuming a high fat diet. Much lower concentrations are usually seen in hay and silage prepared from the same plant species. This is partially due to loss of plant leaves where chloroplast lipid is concentrated, but also due to plant metabolism of stored energy sources. Plant enzymes can continue to function in dried forage containing as little as 5 to 10% moisture. Plant maturity has a definite impact on both fatty acid content and fatty acid composition. Fatty acid content (g/kg DM) generally is highest in the spring and fall seasons and lowest in the summer months. For example, fresh perennial ryegrass contained 32 g/kg DM total fatty acids during primary growth in May, but only 12 g/kg DM at the beginning of second regrowth (Bauchart et al., 1984). Linolenic acid follows a similar seasonal pattern (Bauchart et al., 1984). As linolenic acid declines over the summer months, percentages of palmitic and linoleic acid increases. 15
19 Fat supplements Most plant oils contain 100% ether extract with a high percentage of fatty acids. The impurities extracted, such as water and pigments, are removed during refining leaving the commercial plant (soybean oil, canola oil, corn oil, etc) and animal (tallow, grease, etc) fats with mainly triglycerides consisting of 90-93% fatty acids. The remaining 7-10% is mainly glycerol. Glycerol is readily utilized as an energy source, but only contains the energy of carbohydrates. Caution is advised when obtaining fats from unknown vendors to be sure that considerable impurities do not still remain in the product that lower the fatty acid and energy content. Rather than guessing, it pays to have a sample of the fat analyzed for fatty acid content and profile. The ether extract values listed in NRC (2001) for dairy cattle exceed 99% for all fat categories (vegetable oil, tallow, hydrolyzed tallow fatty acids, and partially hydrogenated tallow) except for calcium soaps. Ether extract for the vegetable oil, tallow, and partially hydrogenated tallow are assumed to contain 90% fatty acid and 10% glycerol. Ether extract for the hydrolyzed tallow fatty acids is assumed to contain 100% fatty acids. Ether extract for calcium soaps is listed as 84.5% in NRC (2001) for dairy cattle, which is assumed to be all fatty acid. Fatty Acid Transformations in the Rumen Feed consumed by ruminants first passes through the largest of the four stomach compartments or rumen, which acts like a fermentation vat. Countless numbers of bacteria, protozoa, and fungi in the rumen ferment the feed releasing end products that are utilized by the host animal for maintenance and growth of body tissues. The microbial population in the rumen also is responsible for extensive transformation of dietary lipid. Lipid transformations include lipolysis to release free fatty acids from complex plant lipids, and biohydrogenation to convert unsaturated fatty acids in plant matter to more saturated lipid end products. Lipids entering the rumen are first transformed by microbial lipases in a process called lipolysis. The microbial lipases hydrolyze the ester linkages in complex lipids causing release of fatty acids and glycerol. The glycerol produced is fermented yielding mostly volatile fatty acids.the main types of lipids entering the rumen are triglycerides, phospholipids, and galactolipids from forages and concentrates in the diet. Rapid hydrolysis of triglycerides occurs by microbial enzymes. Linseed oil incubated with ruminal contents of sheep at 1.0 g/100 ml resulted in greater than 75% of the total lipid recovered in the form of free fatty acids. Phospholipids and galactolipids also undergo rapid and extensive breakdown in the rumen as a result of the enzyme activity of ruminal microorganisms. Some evidence suggests hydrolysis of triglycerides and galactolipids from pasture grass was due primarily to plant enzyme activity. Dawson et al. (1977) autoclaved 14 C- labelled grass to inactivate plant lipolytic enzymes. The grass was then administered intraruminally to a sheep and the galactolipids were rapidly hydrolyzed. Additionally, in an in vitro trial, grass was homogenized with rumen fluid taken from sheep that had been given autoclaved grass for 7 d and the grass galactolipids were rapidly hydrolyzed. It was assumed that the ruminal contents and grass were devoid of plant lipases. In the absence of plant lipases the grass galactolipids were rapidly hydrolyzed. Grass was also homogenized with boiled ruminal fluid. In the absence of microbial lipases the galactolipids were not metabolized. Therefore, it was concluded that lipases produced by ruminal microorganisms are mainly responsible for the breakdown of ingested plant lipids (Dawson et al., 1977). The biohydrogenation of linoleic acid in the rumen (Figure 3) begins with its conversion to CLA. In this initial step, the number of double bonds remains the same but one of the double bonds is shifted to a new position by microbial enzymes. Normally, the double bonds in linoleic acid are separated by two single bonds, but in CLA, the double bonds are only separated by one single bond. Many types of CLA are produced in the rumen of dairy cows (Bauman and Lock, 2006), but a common CLA produced from biohydrogenation of linoleic acid is cis-9, trans-11 C18:2. As biohydrogenation progresses, double bonds in the CLA intermediates are then hydrogenated further to trans fatty acids having only one double bond. Trans 16

20 double bonds only differ from cis double bonds in the placement of the hydrogens. The hydrogens are located on opposite sides of the double bond for trans fatty acids, but on the same side of the double bond for cis fatty acids. Although the difference in structure between trans and cis fatty acids appears small, it causes significant differences in their physical and metabolic properties. A final hydrogenation step by the ruminal microbes eliminates the last double bond yielding stearic acid as the final end product. As a result of biohydrogenation, there is extensive loss of unsaturated fatty acids from the mouth to the duodenum of the animal. In cows on a typical forage diet, the major trans C18:1 present in ruminal contents is trans-11 C18:1. Most of the remaining isomers have double bonds distributed equally among carbons 12 through 16 (Bickerstaffe et al., 1972). The exact pathways for the production of these positional isomers are not known. Linoleic and linolenic acids are converted to several trans C18:1 and C18:2 intermediates during biohydrogenation. Protection of Ruminal Fermentation Biohydrogenation can be argued as an evolutionary adaptation of the microbial population in the rumen as a self-protection device against the toxic effects of unsaturated fatty acids. Elevating fatty acid concentration in ruminal contents may cause a number of changes in ruminal fermentation characteristics and microbial population distribution. Ruminal changes are the result of the antimicrobial nature of unsaturated fatty acids, where fatty acids adsorb onto the cell membrane of selected microbial species, and then penetrate into the membrane causing disorganization of phospholipids and eventual cytological damage (Jenkins, 2002). Because some bacterial species are more susceptible than others, the result is a microbial shift in the rumen. The fatty acid-induced microbial shift can disrupt fermentation of carbohydrate digestion causing a drop in the acetate to propionate ratio and possibly a reduction in fiber digestion (Jenkins, 2002). Two factors that affect the antibacterial activity of lipids are fatty acid structure and concentration. Free fatty acids generally disrupt fermentation more than triglycerides and antibacterial activity of free fatty acids can be enhanced by increasing the number of double bonds (Chalupa et al., 1984). Growth of some bacterial species is stimulated by low concentrations of fatty acids, but inhibited at higher concentrations (Maczulak et al., 1981). In attempting to predict ruminal fermentation changes caused by dietary lipid, it is often assumed that the fat load is contributed only by the fat supplement and that free fatty acid concentration is low. Both assumptions can be wrong. Fatty acids from the grain and forage can significantly contribute to total rumen fat load, for example when animals are consuming immature pasture. Also, free fatty acid concentration may be elevated in some feed ingredients such as whole cottonseed stored in warm, humid conditions (Cooke et al., 2007), or in forages resulting from hydrolytic cleavage of esterified lipids during hay-making (Yang and Fujita, 1997). Fatty Acid Outflow from the Rumen and Animal Performance Meeting Essential Fatty Acid Demands Omega fatty acids belong to one of three families, the w-9, w-6, or w-3 family. Each family has a parent fatty acid that is converted to other biologically-active acids within the same omega family (Figure 4). The only parent fatty acid that can be made by body tissues is oleic acid. The w-6 and w-3 parent compounds (linoleic and linolenic acids) cannot be synthesized by body tissues and, therefore, must be supplied in the diet. Thus, linoleic and linolenic acids are regarded as essential because they are required for normal tissue function but cannot be synthesized by body tissues. A typical total mixed ration of grains and forages generally contains adequate essential fatty acids to meet the needs of the animal. However, the majority of the dietary essential fatty acids are destroyed by microorganisms through biohydrogenation. Part of the interest in omega fatty acids in dairy cattle is to enhance their concentration in milk for valueadded opportunities, and part is to enhance their concentration in body tissues of the cow to enhance production and health. Omega fatty acids in milk are increased to improve manufacturing properties and to increase fatty acid nutraceuticals known to enhance human health. Increasing omega fatty acids in tissues of the cow has potential benefits on reproductive performance, immunity and disease resistance, and positive hormonal shifts. In a few studies, feeding fat to lactating dairy cows has improved reproductive performance implying possible benefits on lifetime production potential. Reported improvements of reproductive performance from added fat include higher conception rates 17
21 (Schneider et al., 1988; Sklan et al., 1989), increased pregnancy rates (Schneider et al., 1988; Sklan et al., 1991), and reduced open days (Sklan et al., 1991). However, supplemental fat has had little or no benefit on reproductive efficiency in other studies (Carroll et al., 1990). The mechanism of how fat supplements alter reproductive performance is not clear. Fat may function in one capacity by providing additional energy during early lactation to support improved productive functions, including reproduction. Negative energy balance delays ovulation and the initiation of the first normal luteal phase (Butler et al., 1981). However, recent studies also suggest that the mechanism involves an energy independent response to fat. Immune System CLA decreased the growth rate in chicks and rats after they were injected with endotoxin (lipopolysaccharide; LPS). This probably was caused by release of cytokines and the prevention of the catabolic effects (Cook et al., 1993). Miller et al. (1994) examined endotoxin-induced growth suppression in mice fed with 0.5 % fish oil and CLA. The fish oil fedgroup lost twice as much body weight after the inoculation with endotoxin than the CLA-fed groups. These researchers found that the CLA in the endotoxin injection inhibited anorexia (a decreased sensation of appetite) and increased splenocyte blastogenesis, concluding that it might inhibit arachidonic acid synthesis, thus preventing the catabolism of tissue by removing eicosanoid precursors. In addition, Bontempo et al. (2004) examined the effects of CLA on the immunological variables of lactating sows and piglets fed with a 0.5 % CLA diet. They found that CLA-fed sows exhibited increased colostrum IgG and serum leptin, and IgG and lysozyme. Nursing piglets of CLA-fed sows also exhibited higher levels of IgG and lysozyme. As these results show, dietary CLA enhanced the effect of immunological variables in lactating sows and piglets. Effect on Body Weight and Fat Park et al. (1997) found that CLA contributed to a reduction in body fat mass and an increase in lean body mass. Mice fed a 0.5 % CLA augumented diet exhibited % lower body fat and 5-14 % increased lean body mass than the controls. They also found that the total carnitine palmitoyltransferase activity was enhanced with dietary CLA supplementation. Moreover, in cultured 3T3-L1 adipocytes, CLA treatment reduced the intracellular heparin-releasable lipoprotein lipase activity and triacylglyceride and glycerol concentration. In addition, DeLany et al. (1999) suggested that CLA played a key role in reducing body fat content and increasing protein accumulation in mice. In this study, the diets of mice were supplemented with CLA (0.0, 0.25, 0.50, 0.75, and 1.0 % by weight). In the 0.50, 0.75, and 1.0 % CLA feeding groups, body fat was significantly lower than in the control, however, body energy was not depressed by any CLA dose. As these results suggested, dietary CLA reduces fat deposition and increases lipolysis in adipocytes, coupled with enhanced fatty acid oxidation. In a human trial, Blankson et al. (2000) conducted a double blind study with 60 overweight or obese volunteers (body mass index kg/m 2 ). The CLA doses given varied from 1.7, 3.4, 5.1, or 6.8 g/day for 12 weeks. The reduction of body fat mass was significant for the 3.4, 5.1, and 6.8 g CLA groups. No significant differences in lean body mass, body mass index, blood safety variables or blood lipids were observed in the groups. Long-term supplementation of CLA studied by Gaullier et al. (2007) reduced body fat mass and increased lean body mass. They conducted a double blind placebo-controlled study with 118 healthy, overweight, and obese adults. The CLA dose was 3.4 g/day. During the six-month period of study, the CLA significantly decreased the body fat mass (-3.4 %) located primary in the legs. Two families of transcription factors may be involved in the intracellular signaling mechanisms of CLA according to Tsuboyama-Kasaoka et al., (2000); peroxisome proliferator activated receptors (PPAR) and sterol regulatory element binding-protein (SREBP). However, the mechanisms by which CLA alters lipid metabolism in the human body are still unclear. Initially, CLA, especially trans-10 cis-12 CLA, enters the cell membrane through unknown transport mechanisms before being shuttled into various regulatory compartments. Brown and Mclntosh (2003) suggested that subsequently three mechanisms in adipose tissue depressed the triglyceride. The first esterified CLA into triglyceride-rich oil droplets. This mechanism did not have much effect on the regulation as a whole, even increasing triglyceride stores. The second mechanism esterified CLA into the membrane-bound phospholipids bilayer, where it changed the fluidity of the membranes associated with a signal transduction cascade. The last mechanism modified the activity of a currently unidentified transcription factor (TFX), resulting in less regulation of PPAR and the depression of lipoprotein lipase, an acyl-coa-binding protein, an adipocyte fatty acid binding protein, a glucose transporter, and leptin. In addition, CLA reduced the acitivity of steroyl-coa desaturase and acetyl-coa carboxylase by altering the activity of TFX. By inhibiting these factors, CLA attenuated insulinstimulated glucose uptake, malonyl-coa synthesis, 18
22 and oleate synthesis, collectively decreasing de novo fatty acid synthesis. CLA also inhibited the expression of SREBP, by enhancing SREBP transcript decay as well as by inhibiting the proteolytic cleavage, releasing the activated nuclear SREBP fragment. Milk Fat Depression The discovery of major events in rumen lipid metabolism occurred decades ago, with very little new information available on the details of its biochemistry and regulation until the discovery of the anticarcinogenic properties of cis-9, trans-11 CLA. This discovery made it apparent that even minute quantities of biohydrogenation intermediates in the rumen could have dramatic effects on health and metabolism of both the host animal and the humans consuming animal-food products. Eventually it was discovered that a second CLA, namely the trans-10, cis-12 isomer, was closely associated with milk fat depression (MFD). This led to the biohydrogenation theory of MFD that suggested feeding management was linked to an abnormal ruminal fermentation causing accumulation of the trans-10, cis-12 isomer. Feed ingredients containing appreciable concentrations of fat often cause MFD, an effect that can be explained by the biohydrogenation theory of MFD. Predicting when fat sources will lead to MFD is complex and is a function of total unsaturated fatty acid supply to the rumen, how fatty acid supply will impact the pathways of lipid biohydrogenation and its interaction with other feed ingredients. An improved understanding of these events will provide the critical framework with which to better troubleshoot MFD. The biohydrogenation theory represents a unifying concept to explain the basis for diet-induced MFD where unusual intermediates of ruminal fatty acid biohydrogenation accumulate in the rumen and eventually reduce milk fat synthesis in the mammary gland. Under certain dietary situations the rumen environment is altered and a portion of biohydrogenation occurs via a pathway that produces trans-10, cis-12 CLA and trans-10 18:1 (Figure 5). Bifidobacterium, Propionibacterium, Lactococcus, Streptococcus, and Lactobacillus isolates from other habitats have been reported to produce trans-10, cis- 12-CLA. As these genera occur in the rumen, although generally at rather low numbers, they may contribute to biohydrogenation and specifically to trans-10, cis-12-cla formation in the rumen. Propionibacterium, Streptococcus, and Lactobacillus are also more numerous in the rumen with concentrate diets (Jenkins et al., 2008), which would again be consistent with greater trans-10, cis-12 CLA production with concentrate diets. Therefore, dietary situations causing MFD alter the pathways of rumen biohydrogenation resulting in changes in the specific trans-18:1 and CLA isomers available for uptake by the mammary gland and incorporation into milk fat. As shown in Figure 5, this trans-10 shift in biohydrogenation pathways, and the associated increase in the trans-10 18:1 content of milk fat, is indicative of the complex changes in ruminal biohydrogenation pathways characteristic of MFD. Although trans-10 18:1 does not directly inhibit mammary synthesis of milk fat (Lock et al., 2007), it is relatively easy to analyze compared to trans 10, cis-12 CLA and other CLA isomers. Therefore, in general, this fatty acid can serve as a marker for the type of alterations in rumen biohydrogenation that characterize diet-induced MFD. References Bauchart, D., R. Verite, and B. Remond Long-chain fatty acid digestion in lactating cows fed fresh grass from spring to autumn. Can. J. Anim. Sci. 64: Bauman, D. E. and A. L. Lock Concepts in lipid digestion and metabolism in dairy cows. Proc. Tri-State Dairy Nutr. Conf. pp Available at: Blankson, H., J. A. Stakkestad, H. Fagertun, E. Thom, J. Wadstein, and O. Gudmundsen Conjugated linoleic acid reduces body fat mass in overweight and obese humans. J. Nutr. 130: Bontempo, V., D. Sciannimanico, G. Pastorelli, R. Rossi, F. Rosi, and C. Corino Dietary conjugated linoleic acid positively affects immunologic variables in lactating sows and piglets. J. Nutr. 134: Brown, J. M., and M. K. Mclntosh Conjugated linoleic acid in humans: regulation of adiposity and insulin sensitivity. J. Nutr. 133: Butler, W. R., R. W. Everett and C. E. Coppock The relationships between energy balance, milk production and ovulation in postpartum Holstein cows. J. Anim. Sci. 53:742. Carroll, D. J., M. J. Jerred, R. R. Grummer, D. K. Combs, R. A. Pierson and E. R. Hauser Effects of fat supplementation and immature alfalfa to concentrate ratio on plasma progesterone, energy balance, and reproductive traits of dairy cattle. J. Dairy Sci. 73:
23 Chalupa, W., B. Rickabaugh, D. S. Kronfeld, and D. Sklan Rumen fermentation in vitro as influenced by long chain fatty J. Dairy Sci. 67: Cook, M. E., C. C. Miller, Y. Park, and M. Pariza Immune modulation by altered nutrient metabolism: nutritional control of immune-induced growth depression. Poult. Sci. 72: Cooke, K. M., J. K. Bernard, C. D. Wildman, J. W. West, and A. H. Parks Performance and ruminal fermentation of dairy cows fed whole cottonseed with elevated concentrations of free fatty acids in the oil. J. Dairy Sci. 90: Dawson, R. M. C., N. Hemington, and G. P. Hazlewood On the role of higher plant and microbial lipases in the ruminal hydrolysis of grass lipids. Br. J. Nutr. 38: DeLany, J. P., F. Blohm, A. A. Truett, J. A. Scimeca, and D. B. West Conjugated linoleic acid rapidly reduces body fat content in mice without affecting energy intake. Am. J. Physiol. 276:R Gaullier, J. M., J. Halse, H. O. Høivik, K. Høye, C. Syvertsen, M. Nurminiemi, C. Hassfeld, A. Einerhand, M. O shea, and O. Gudmundsen Six months supplementation with conjugated linoleic acid induces regional-specific fat mass decreases in overweight and obese. Brit. J. Nutr. 97: Hatfied, R., H. G. Jung, G. Broderick, and T. C.Jenkins Nutritional chemistry of forages. In Forages, Volume 2. The Science of Grassland Agriculture, Sixth Edition. R. F. Barnes (Ed.), Blackwell Publishing, Iowa State Press, Ames. Jenkins, T. C Lipid transformations by the rumen microbial ecosystem and their impact on fermentative capacity. pp in Gastrointestinal Microbiology in Animals, S. A. Martin (Ed.), Research Signpost, Kerala, India. Jenkins, T. C., and W. C. Bridges, Jr Protection of fatty acids against ruminal biohydrogenation in cattle. Eur. J. Lipid Sci. Technol. 109: Jenkins, T. C., R. J. Wallace, P. J. Moate, and E. E. Mosley Board-invited review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J. Anim. Sci. 86: Lock, A. L., C. Tyburczy, D. A. Dwyer, K. J. Harvatine, F. Destaillats, Z. Mouloungui, L. Candy, and D. E. Bauman Trans-10 octadecenoic acid does not reduce milk fat synthesis in dairy cows. J. Nutr. 137: Maczulak, A. E., B. A. Dehority, and D. L. Palmquist Effects of long chain fatty acids on growth of bacteria. Appl. Environ. Microbiol. 42: Miller, C. C., Y. Park, M. W. Pariza, and M. E. Cook Feeding conjugated linoleic acid to animals partially overcomes catabolic responses due to endotoxin injection. Biochem. Ciophys. Res. Commun. 198: Park, Y., K. J. Albright, W. Liu, J. M. Storkson, M. E. Cook, M. W. Pariza Effect of conjugated linoleic acid on body composition in mice. Lipids. 32: Freeman-Pounders, S. J., D. W. Hancock, J. A. Bertrand, T. C. Jenkins, and B. W. Pinkerton The fatty acid profile of rye and annual ryegrass pasture changes during their growth cycle. Forage and Grazinglands 30 January, Schneider, P., D. Sklan, W. Chalupa, and D. S. Kronfeld Feeding calcium salts of fatty acids to lactating cows. J. Dairy Sci. 71: Sklan, D., E. Bogin, Y. Avidar and S. Gur-Arie Feeding calcium soaps of fatty acids to lactating cows: effect on production, body condition and blood lipids. J. Dairy Res. 56: Sklan, D., U. Moallem and Y. Folman Effect of feeding calcium soaps of fatty acids on production and reproduction responses in high producing lactating cows. J. Dairy Sci. 74: Tsuboyama-Kasaoka, N., M. Takahashi, K. Tanemura, H. J. Kim, T. Tange, H. Okuyama, M. Kasai, S. Ikemoto, and O. Ezaki Conjugated linoleic acid supplementation reduces adipose tissue by apoptosis and develops lipodystrophy in mice. Diabetes. 49: Yang, U. M., and H. Fujita Changes in grass lipid fractions and fatty acid composition attributed to hay making. Grassl. Sci. 42: Kellens, M. J., H. L. Goderis, and P. P. Tobback Biohydrogenation of unsaturated fatty acids by a mixed culture of rumen microorganisms. Biotechnol. Bioeng. 28:

24 The Compatibility Between Dairy Productivity and Carbon Footprint Jude Capper Washington State University 21

25 22
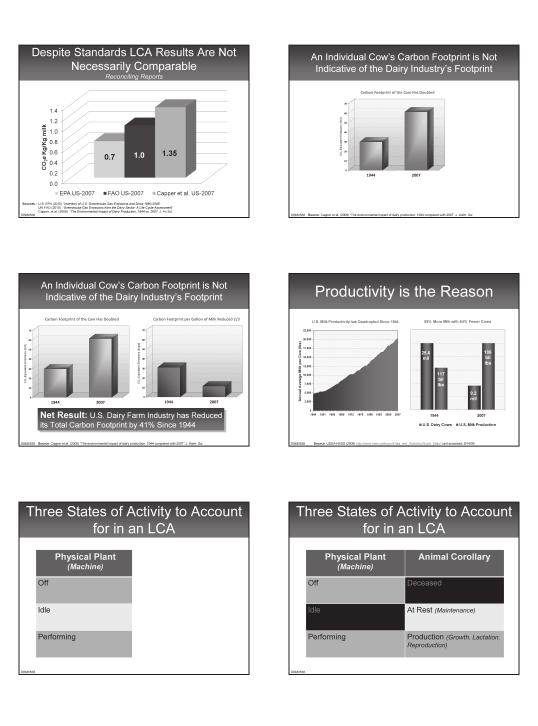
26 23

27 24
28 25
29 26
30 Feeding Economics For 2010 When reviewing the 2009 dairy industry and economics, no one could imagine the steep drop in milk prices at the farm gate (over 40 percent), the quickness of the drop in milk prices (less than one month), and the length of time (10 months below breakeven milk prices). Another change was Midwest states had higher milk increases varying from four to six percent in 2009 on the same number of cows while many other states reported less milk with fewer cows. What may have happened in Illinois and the Midwest allows us to look back to look ahead and prepare for the next milk price challenge which started in February, Dairy managers will need to plan for variable shifts in milk prices, feed prices, and profit margins. Michael Hutjens University of Illinois 232 Animal Sciences Lab 1207 West Gregory Drive, Urbana, IL Illinois dairy producers had ample supplies of forages and corn produced on their farms which did not have to be purchased in This advantage reflects the agronomic skills of Illinois dairy producers, but they did not capture higher market prices and income if they sold feed. This strategy avoided out-of-pocket costs which bankers encouraged/demanded of dairy managers. Illinois producers did not expand and/or paid down debt. Bank and interest payment could be minimized. Stable land prices allowed credit to be extended compared to dairy cow equity that declined 25 to 50 percent. Making Correct and Incorrect Management Decisions Thirty two dairy nutritionists, nine veterinarians, and ten educators responded to a field survey to the list the three best or correct decisions dairy managers made in 2009 and the three incorrect changes made in 2009 in ranked order. The top ranked item was assigned three points, the second ranked item was assigned two points, and third ranked decision was assigned one point. Table 1 summarizes positive or correct decisions by each group. The survey points out the role of forage quality, level, and type (corn silage specifically) and keep focused on correct decision with $12 or $20 per one hundred pounds of milk. Other interesting correct points included maintaining high morale and positive attitude with family members and hired labor, feed contracting, amino acid balancing reduced protein purchases, grouping cows, and bringing heifers back to the farm to reduce costs. Table 2 summarizes negative or incorrect dairy manager decisions by each group in the survey. The main concern was reducing or removing feed intake, nutrients, minerals, vitamins, and/or additives. Other interesting points were not entering the CWT program, lame and broken cows, major expansions in 2007 and 2008, and not using professionals (AI technicians, calf and heifer growers, hoof trimmer, and/or veterinarians). Providing Economic Flexibility Illinois dairy managers can have several advantages compared to other dairy managers facing the 2009 low milk prices with high feed prices. Illinois dairy farms (average herd size of 106 cows) use family labor as a resource. Dairy farm families did not draw $50,000 (Minnesota guideline for a dairy farm family with two children) in the short term. Because the average Illinois herd size is fewer than 150 cows, dairy farm managers could qualify for the MILC program providing $1 to $2 per cwt month added income. A rbst payment for not using this technology resulted in forty to sixty cents per cwt added income. One Illinois cooperative provided one dollar advanced patronage per cwt payment for several months to assist financially stressed dairy farms. Making Correct Decisions An important decision was not to make short decisions to save a nickel while leading to a long term loss of a dollar ( staying the course ). Examples of potential decisions and choices are outlined below. Reducing or removing minerals and vitamins can save six cents (heifers) and twenty cents (lactating cows) a day. Because minerals do not immediately reduce milk yield, dairy managers reasoned this may be a prudent move. However, when mineral deficiencies occur six months later (reduce immunity, slower growth, and declining fertility), it resulted in large negative economic impacts which may be difficult to pinpoint and recover later in lactation or growth phases. 27
31 Holstein heifers must gain over 1.7 pounds per day if they are expected to calve at 23 to 24 months of age, weigh 1250 pounds after calving, and produce lactations yields above herd average. The cost of delayed calving is $2 per day (reflects only added feed costs). An increase in somatic cell count due to reduced immunity and health (removal of organic trace minerals, less vitamin E, and/or energy shortage for example) will lead to a loss of 2 to 2 1/2 pounds per cow per day due to a change in one linear somatic cell count score. An increase in days open will cost $2 per day (each day over 120 days open) to $8 per day (each day over 180 days open) based on Wisconsin data which could be related to negative energy balance due to removal of fat or effective feed additives. Dropping an accelerated calf feeding program can reduce milk yield in the first lactation by 1100 pounds due it impact on mammary gland development. This decision is a long term investment of $30 to $50 added feed costs not recovered for nearly two years in higher milk yield based on IL and NY data. Feed additives must be purchased (an out-ofpocket cost) which can lead to removing additives than return 3 to 10 times the cost of the feed additive (for example a buffer returns 30 cents in added milk production for a six cent investment). The feed additives listed are ranked: 1 st choice monensin (an ionophore); 2 nd choice yeast-based products; 3 rd choice silage inoculants; 4 th choice organic trace minerals; 5 th choice rumen buffers; and 6 th choice biotin. Shifting from a one group TMR to multiple TMRs may be an alternative to lower feed costs. Feeding a ration higher in forages to lower producing cows can save 40 to 75 cents or more per day. Consider low producing cows may consume 4 to 6 pounds less dry matter which can reduce the estimated savings. High producing cows may need more nutrients to replace lost body weight in late lactation. Heifers may need added nutrients to grow reaching their mature weight. Another economic consideration is if the one group TMR contains expensive nutrient sources (such as inert fat, amino acids, added fat/oil, or high quality RUP protein sources). Recent research from Michigan State University suggest metabolic reasons for grouping cows (high group cows requiring more glucose, more dry matter intake, has lower insulin sensitivity, and higher levels of natural BST compared to low producing cows). Monitoring Feed Changes From Cow Responses When dairy managers make changes, lactating cows will respond (cows talk to you). Monitor the following cow measurements to determine if your change led to lost income or health. MUN (target 8 to 14 mg /dl to avoid nitrogen lose while maintaining milk protein levels) Milk protein and milk fat test (meet or exceed breed averages) Management level milk or 150 day milk (should increase or maintain herd values) Fecal scores (range from 2.5 to 3.5) Changes in feed benchmarks Herd feed efficiency from 1.5 to 1.7 pounds of 3.5% milk per pound of dry matter with each change in 0.1 point worth 25 to 35 cents per cow per day. Feed cost per pound of dry matter at 9 to 10 cents per pound of dry matter reflects the cost of feed ingredients selected when building and balancing the ration. Feed cost per cwt ($5 to $6 per cwt) reflects the cost per pound of dry matter, amount of dry matter offered including weigh backs, and milk yield. Milk yield is the key factor. Income over feed costs represents margin (dollars available) for fix, variable, labor, and return to management. Milk price is a key factor in this value. Feeding Strategies That Worked Forage quality is a key solution. Consider increasing corn silage levels in rations as feed cost per cow per day may drop 15 to 30 cents. Evaluate the use of low lignin forages and forages high in NDFD (neutral detergent fiber digestibility). Use of computer modeling programs allows for fine-tuning rations. Lower levels of protein based on amino acid balancing and rumen microbial estimation can be reduce feed costs while optimizing production. Determine if starch levels and utilization are optimal. Lower levels of starch (20 to 22 percent) can maintain milk production with high quality 28
32 forage, rumen fermentable fiber, adding sugar, and/or feeding an ionophore. Plant or kernel processing of corn silage and processing corn grain can increase starch availability in the rumen and reduce fecal losses of starch. If fecal starch is over 5 to 7 percent, examine sources and processing reducing starch utilization. By-product feeds can be an economical nutrient source. Distillers grain and wet brewers grain can reduce protein costs while corn gluten feed, soy hulls, and wheat midds can maintain energy levels while reducing feed costs (Table 3). Review shrink losses. Managing and monitoring weigh backs can increase profitability. One guideline is to target 1 to 2 percent weigh back per cow per day. Bunk management may allow feeding to an empty bunk reducing feed refusals saving 1 to 3 pounds of dry matter per cow per day (9 to 27 cents a day). Table 3. Breakeven prices for selected by-product feeds using Feed Val 3 and Sesame with recommended levels in lactating cow rations. Feed prices used to calculate breakeven prices with FeedVal3 were soybean meal at $350 a ton, shelled corn at $3.50 a bushel, tallow at 30 cents a pound, dicalcium phosphate at $25 per cwt, and limestone at $10 per cwt. Sesame calculations were based on 30 reference feeds on April 22, By-product Breakeven price Level Feed Val 3 Sesame (% ration DM) - ($ per ton) Soy hulls Cottonseed, fuzzy Corn gluten feed to 25 Brewers grain (30% DM) 66 na 15 to 20 Corn hominy to 20 Corn distillers grain (> 10% oil) 20 (< 10% oil) Table 1. Summary of field responses to correct dairy management decisions made with low milk prices in Decision Nutritionists Veterinarians Educator Total Points Forage program aspects Staying the course Ration balancing/nutrients Strategic culling Milk components and quality Financial adjustments Use of by-product feeds Labor management Use of rbst Contracting milk Table 2 Summary of field responses to incorrect dairy management decisions made with low milk prices in Decision Nutritionists Veterinarians Educator Total Points Removing feed Pulling min, vit, and additives Not staying the course Low forage quality Avoiding financial support Reduce hoof care Poor cow comfort Reduced rbst use Overfeeding of distillers grain Reducing health Incorrect culling decisions Not using records

33 4-State Dairy Nutrition & Management Conference 30
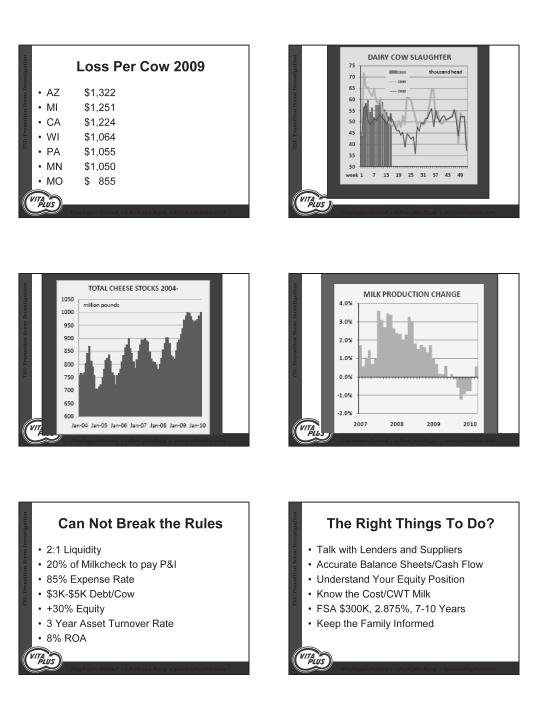
34 31

35 Coordination of Reproductive and Nutritional Management of Lactating Dairy Cows Bill Thatcher University of Florida 32

36 33
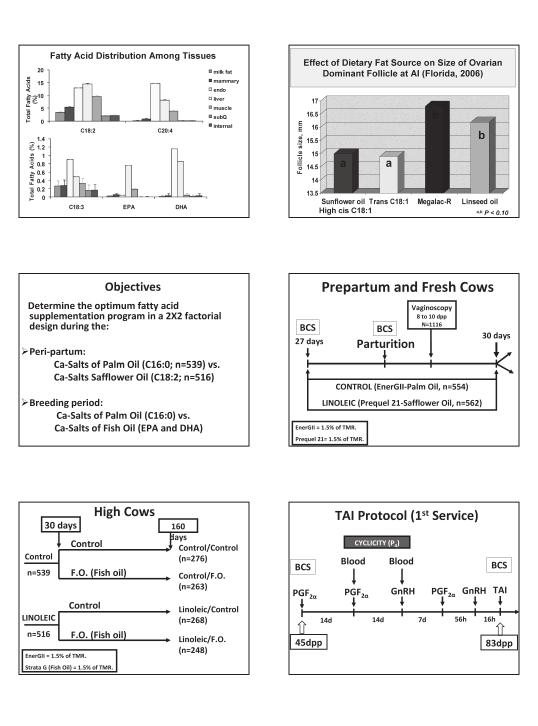
37 34
38 35
39 36
40 37

41 Demystifying the Environmental Sustainability of Food Production Jude Capper Washington State University 38
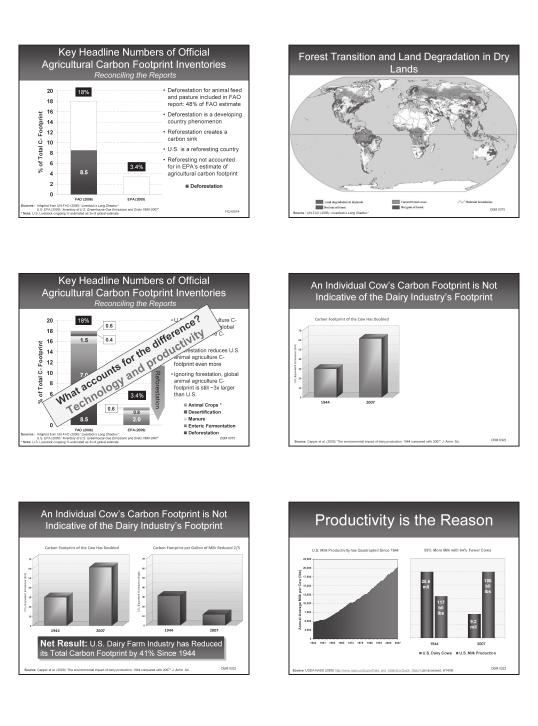
42 39
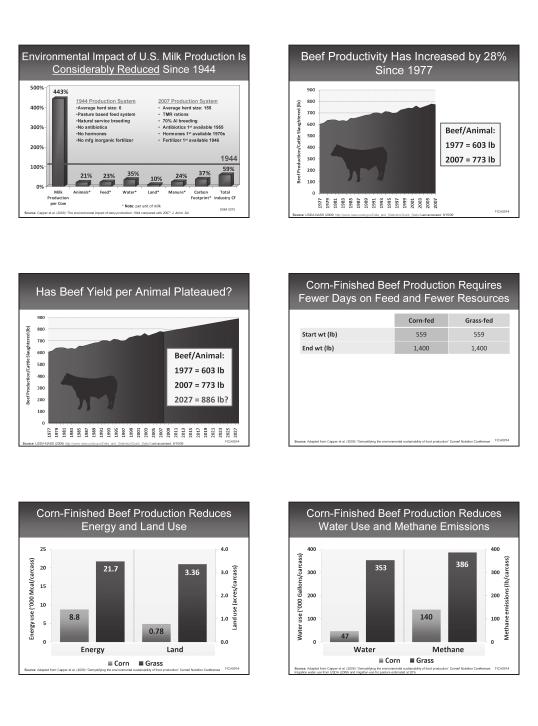
43 40
44 41

45 42
46 The Use of Records to Evaluate and Improve Transition Cow Performance M. W. Overton DVM, MPVM, and I. J. Smith, DVM University of Georgia, College of Veterinary Medicine Department of Population Health Rhodes Center - ADS 425 River Road, Athens, GA moverton@uga.edu Introduction The transition period, which extends from approximately 3 weeks prior to calving until approximately 3 weeks post-calving, is a high-risk time in a cow s life and proper management of this period is critical to the success of any dairy. During this time, cows experience many physiological and metabolic changes and must successfully adapt to the changing demands. There is a large increase in nutrient requirements that cannot be met through reliance on dry matter intake and thus, cows must mobilize body tissue to meet the energy and protein demands. During late gestation, the fetus is growing rapidly and the cow begins producing colostrum. As a consequence, there is a large increase in the amount of glucose and amino acids that she needs to produce in order to meet the demands for energy and growth of the fetus and for the production of colostrum. Despite these increasing needs, feed intake declines during the last 7-14 days prior to calving. Cows experience large changes in hormone levels including increases in estradiol, growth hormone, cortisol, adrenaline, endorphins, and oxytocin and decreases in progesterone. There is also a large increase in the demand for calcium, first for colostrum and then for lactation. With the onset of lactation following parturition, glucose and amino acid demands increase by approximately 3-fold as compared to the already high demands of the previously gravid uterus. In order the meet the energy needs of this transition period, cattle shift the ways in which their bodies use glucose (essentially sparing glucose for the mammary gland and fetus at the sacrifice of other body tissues such as muscle) via changes in insulin sensitivity and increases in growth hormone levels. These other tissues derive their energy from mobilized fatty acids. If a successful transition and adaptation to the changing demands is made, cows will experience mild increases in the levels of nonesterified fatty acids during the last 10 days of gestation that carry over into early lactation. Cows that successfully transition into lactation will not experience appreciable hyperketonemia prior to calving, although the levels of -hydroxybutyrate will moderately increase in early lactation but generally remain less than 1,000 Ìmol/L. However, if cows fail to successfully manage the demands of the transition period, excessive mobilization of fatty acids from their body stores will occur leading to a fatty liver, poor immune response, poor glucose production, hyperketonemia above 1,400 Ìmol/L, increased risk of disease, increased risk of premature culling, and decreased reproductive performance. Key components of successful transition include: 1) the implementation of herd management approaches that focus on prevention of periparturient problems, optimization of feed intake, and the removal of stressors, 2) real time monitoring of key processes that impact the risk of periparturient disease problems, and 3) finally, the monitoring of success or failure in key outcomes of interest. Each of these components will be discussed in part in the remainder of the paper. When proper preventative management is combined with real-time monitoring, the result should be an improved transition program and increased lactation and reproductive success. Management Keys Grouping, Housing and Pen Movement The goal of proper grouping is to reduce the social, environmental, and metabolic stressors on a population of cows by minimizing the number of pen changes a cow is forced to make while also working to fit the management needs of the dairy. Strive to avoid unnecessary pen changes, as each pen move is likely to result in a drop in DMI and elevated cortisol levels both of which may negatively impact immune function and overall health and productivity. When pen moves are necessary, decrease the impact of pen changes by moving animals once weekly and move in groups of 10 or more animals, if possible. Avoid moving cows into new pens during the last 10 d prior to calving. Ideally, cows should spend at least 14 d in the close-up pen. Due to the inevitable variation around calving dates, strive to have at least 90 % of cows spend at least 10 d in close-up pens by modifying the move dates for cows going from the far-dry to the closeup pens. Change the move dates during summer heat stress and for cows carrying twins as each of these conditions usually result in a gestation length that is 5-7 d shorter than expected. 43
47 Separate heifers and older cows, if possible, since heifers have been shown to have longer resting times and higher DMI when separated from mature cows. Some researchers feel that heifers need higher levels of protein during the close-up period ( > 15 % CP or g of metabolizable protein) as compared to mature dry cows. Feeding dietary cation-anion difference (DCAD) diets to springing heifers is not usually required, since they are not as susceptible to clinical hypocalcemia and are less affected by subclinical hypocalcemia than mature cows; but some herds report improved performance with DCAD diets for heifers. Maintain the stocking density at less than 100 %, based on both feed bunk space and resting area. Provide ~ 30 in of bunk space per animal or, in pens with lock-ups spaced at 24 in, populate the pen at % of the number of lock-ups. To avoid confusion, instead of relying on a count of lock-ups, strive to always provide 30 to 36 in of feed bunk access per cow in the close-up and fresh cow pens. Maintain a clean, dry environment with adequate areas for animals to rest. Mud and heat stress increase metabolic needs, but decrease feed intake. Wet, mucky conditions also increase the risk of mastitis that may not appear until later in the fresh pen. Cows calving in wet conditions may experience higher risks of metritis. If using maternity pens, these areas should be bedded with clean, dry material and changed frequently. The frequency of rebedding will depend on a variety of issues such as stocking density, bedding type, weather conditions, etc. Striving to maintain a clean, dry area results in cows maintaining good hygiene scores. Following calving, cows should be housed in a colostrum pen, instead of a hospital pen, if feasible, until milk is free of dry cow antibiotic residues and legal for sale. If this additional pen is not possible, strive to understock the hospital pen, both in terms of feed bunk space and resting area. Minimize distance walked in these tired and sore fresh cows by placing the pre- and post-fresh pens close to the parlor, if possible. Ideally, the close-up pens should be located close enough to allow frequent observation, but not in the midst of noisy, high traffic areas that might stress the cows and interfere with feeding or calving. Nutrition and Feed Delivery The primary feeding management goals during the periparturient period is to minimize the inevitable drop in DMI that occurs prior to calving, meet the energy and amino acid demands without overfeeding energy, and to maximize the rise in intake postpartum. Feed intake, energy balance, and the magnitude of change during the periparturient period are associated with changes in immune function, risk of developing retained placenta and metritis, and postpartum feed intake. Energy and protein requirements during the last week of gestation are estimated to be approximately 15 Mcals NE L and 1100 g of metabolizable protein/d, respectively. It is beyond the scope of this paper to adequately describe the various strategies and guidelines for balancing rations, but there are a few basics worth mentioning: Ensure an adequate level of fiber intake by feeding 7 to 7.5 lb of forage ADF. Make sure cows are actually consuming the ration provided by using a Penn State Particle Separator to evaluate both the fresh feed and the refusals. Increase metabolizable protein to approximately 1100 g/d (corresponds to a positive balance of 400 to 450 g of metabolizable protein in some ration balancing programs) Be careful with fermentable carbohydrate levels keep total NFC to less than 30 to 32 % and starch at approximately %. If using DCAD diets for close-up cows, select forages, grains, and grain by-products that are low in potassium to minimize the amount of anionic salts needed. Feed additional vitamin E to close-up and fresh cows. Vitamin E has been shown to improve immune function and decrease the risk of retained placenta, metritis, and mastitis in fresh cows. Specific levels to feed depend upon type of diet and feed ingredients but many consultants recommend levels of 1800 to 3000 IU/d in these high risk cows. Energy and protein requirements during early lactation change dramatically as milk production increases. After the prescribed withdrawal time, move cows from the colostrum pen to a fresh cow pen for ~ 10 to 21 d. Duration of time in fresh pen is dependent upon preferred feeding strategy, ability to feed a special fresh cow ration, 44
48 and calving pressure. Once again, it is beyond the scope of this paper to adequately describe the various strategies and guidelines for balancing rations, but there are a few basics depending on the management/ feeding option chosen: Option 1 short duration in fresh pen (10 to 14 d) with more aggressive protein feeding followed by move to normal high cow ration at 10 to 14 d in milk. Ensure an adequate level of fiber intake by feeding approximately 7 lb of forage ADF with total NDF levels at approximately 32 %. Increase metabolizable protein balance to a positive 500 to 600 g. Maintain correct blend of carbohydrates to drive propionate production, but keep total NFC at 35 to 38 %. (Note, some programs will report a higher level of NFC on the same diet as compared to other ration balancing programs. Check on the suggested requirements for the program you are using, but in general, most feed a slightly lower level of NFC to these postpartum cows and then increase the level as they move along in lactation.) Option 2 move cows onto regular high cow ration. Ensure an adequate level of fiber intake by feeding 7 to 7.5 lb of forage ADF with total NDF levels at approximately 30 to 33 %. Shoot for metabolizable protein balance of positive 250 to 400 g through first 100 d in milk (DIM). Feed a balanced carbohydrate blend of approximately % starch, 4.5 to 5.5 % sugars, and 9.5 to 11 % soluble fiber. (Note: Ration balancing programs may estimate and report specific ration components differently. The aforementioned comments are meant as general guidelines, but may not be directly comparable across different programs.) In both scenarios, the goal is to ensure an adequate level of fiber intake to maintain rumen health while still providing the proper mix of fermentable substrate and nitrogenous sources (protein) to increase microbial numbers and propionate, the driver behind glucose/ lactose production and subsequently, milk production. Fat cows ( 4.0 BCS) are at increased risk of ketosis and often benefit from oral drenching. Consider 8 to 10 oz propylene glycol drench/cow/d at calving and again in 24 hr. General feeding principles: Ensure uniform feed intake by all animals. Monitor particle size using a particle separator. Maintain a moisture content of the ration between 50-60% to help reduce sorting and increase palatability (may need to add water to some rations). Monitor manure for fiber length, grain particles, gas bubbles and consistency across cows within a pen. Pre-batch mix/ chop hays to control length to approximately 2-3 in (i.e., less than the width of a cow s muzzle) in order to reduce sorting. Use high quality, highly palatable hays free of mold and mycotoxins. Use high quality, highly palatable silages free of clostridial or butyric acid fermentation problems and mycotoxins. Do not feed silage from top and sides of the silo to transition animals. In general, limit silages to no more than ~ 40 to 50 % of forage needs in pre-fresh cows if possible. Clean out feed bunks daily for both close-up and fresh cows to minimize risk of feed intake depression from moldy or heated feeds. Facilities and Cow Comfort Good cow comfort to promote more lying time and to minimize additional metabolic needs associated with excessive standing and or walking is critical, especially in fresh cows since these animals are at an increased risk of lameness/ laminitis due to the influence of periparturient hormonal changes that may negatively impact foot and leg tissues and due to pen, ration, and feed intake changes. Cows need sufficient space for resting in a clean, dry area. This need can be accomplished using well-designed free stalls, dry lots, bedded packs or in other ways. Space requirements: ~ 100 sq ft/ cow in bedded packs; 45
49 ~ 500 to 600 sq ft/ cow of loafing area and sq ft shade area/ cow in open corrals; or A minimum of 1 properly bedded and maintained freestall/cow, if using freestall housing, is important. Heat stress abatement is critical in both prefresh and fresh cows. Provide soaker lines on lock-ups during heat stress conditions that cycle once every 15 min from 70 to 79 F, once every 10 min from 80 to 88 F, and once every 5 min above 88 F with 0.33 gal of water/cow/cycle. Water is a critical nutrient and should not be overlooked. Strive to provide a minimum of two locations per pen and a minimum of 3 linear in. per cow. Acclimate heifers to lock-ups/stanchions and concrete feeding aprons, if possible, prior to entering the close-up pen. Monitoring Real Time and Other General Items Monitoring is the regular observation and recording of activities, events and yields that occur for the purposes of observing and evaluating the degree of change, intended or unintended, positive or negative, within a system. It should include a systematic approach to data collection, evaluation, and provision of feedback about the changes detected. The objectives of monitoring include: 1) to recognize normal performance, 2) to test the impact of intentional change in some area of management or performance, 3) to discover unintended drifts or declines in procedures or performance, and 4) to determine potential causes of abnormal performance. However, before getting into specific monitors, reports or interpretation, there are some general concepts, concerns and terminology that must be considered. Goals are target levels of performance toward which producers are trying to achieve and are typically related to profitability. For instance, a farm might have a goal of higher milk production, better reproductive performance, or lower somatic cell counts. Metrics are any type of measurement or set of measurements that quantify results and are used to gauge some quantifiable component of dairy performance, i.e., Is the herd meeting its goals?. Metrics that are monitored in dairy production are typically numbers that represent some type of process and are always important in achieving a goal, but are not synonymous with the goal themselves. Goals are great to establish for herds, but it is rarely a good idea to use the goal itself as a monitor. An example might be the average age-at-freshening of replacement heifers. If the average age has been 27 months, it might be a good goal to lower this age by a few months. Age-at-freshening is an appropriate goal, but a horrible monitor since it is the result of many processes that are involved in achievement of the goal such as appropriate feeding, housing, vaccination, breeding, etc. The process of monitoring involves the routine and systematic collection and evaluation of information (monitoring parameters) from a dairy in an attempt to detect change in the process. In order to do this, it is critical to monitor as close to the process(es) as possible, as opposed to simply measuring the outcome. Close-up urine ph, stocking density and feed intake are often some of the best predictors of future fresh cow problems and can be monitored on a daily basis. These specific real-time monitors will be discussed below as well as other general items that can be used to evaluate transition performance. Monitoring feed intake is one of the simplest monitors of change in performance (and predictors of future performance), but yet is most often overlooked. Feed intake should be weighed daily, both the amount of fresh feed delivered and the amount leftover, as both a monitor and predictor of fresh cow performance. Target a 5 % refusal (or more) on a daily basis and ensure that the ration is not easily sortable by grinding hays to 2 3 in. and adding water if necessary. Based on a typical 21 to 24/d average for cows in the close-up pen, strive to achieve a DMI of at least 26 lb for mature Holsteins and at least 23 lb for Holstein heifers when these animals are housed in separate pens. In fresh pens that range from 2 21 DIM, strive to achieve at least 35 lbs of DMI for first lactation Holsteins and at least 43 lbs of DMI for mature Holsteins. In mixed parity fresh pens, DMI should be at least 38 lb for Holsteins. If using DCAD diets for close-up cows, urine ph s should be closely monitored. Monitor urine ph once weekly from 10 to 15 cows while feeding a DCAD diet or more frequently if feed ingredients change. In general, cows must be on the new diet for at least 48 hrs in order to accurately assess the DCAD balance using urine ph. The goal is to have all cows at a ph of 6.0 to 6.9. Many people monitor the average ph, but the 46
50 average can be very misleading, especially in situations where cows are sorting the ration and some animals have a high ph while others are too low. Overacidification (urine ph < 5.8) may result in depressed DMI and perhaps compromised immune function; while inadequate acidification (urine ph > 7.2) can lead to severe, non-responsive downer animals following calving. Either scenario can also result in an increase in retained placentas. All lactating cows are expected to lose some weight post-calving. Normal weight loss during the first 30 DIM should be < 0.75 BCS or ~90 lb (1 BCS ~ 120 lb of fat and protein). If possible, monitor BCS for close-up and just fresh cows and then again at approximately 2 months of lactation. First service conception risk may be reduced by 50 % when BCS decreases by more than 1.0 BCS during the first 60 DIM and the risk of a prolonged anovulatory condition (failure to cycle) increases in animals whose BCS falls below 2.75 or who lose excessive condition during the early postparturient period. Use some form of a fresh cow monitoring and treatment program custom designed with your veterinarian to fit your farm s needs. No one program fits all herds, but most herds benefit from some sort of evaluation program to assess appetite, attitude, and appearance of every cow in the fresh pen every day. Depending on the amount of labor available, as well as the quality of the labor, some herds need a rigorous fresh cow monitoring program to prevent cows from falling through the cracks. If some form of a 10-d monitoring program is utilized, careful attention should be paid to ensure that fresh cows are not locked up for more than 30 to 45 min/d. Other herds that have very high quality herdsmen and fewer fresh cow issues may actually perform better with a prompted assessment approach instead of individually examining every fresh cow every day for the first 10 d of lactation. Many herd owners and consultants like to monitor fresh cow culling risk (sold and died are often calculated separately) during the first 30 and 60 days in lactation and to use this as a metric for evaluating transition cow management. However, this approach is fraught with issues and in most cases, probably should not be used. First, most herds do not calve enough cows to get an accurate assessment of the true point estimate of culling risk. Second, culling risk in early lactation should not be compared between herds due to the impact of other management issues or different philosophies regarding the culling of animals in early lactation. For example, one herd may have severe reproductive challenges and must keep as many animals as possible simply to keep herd numbers up. Another herd may have good reproductive performance, an abundance of fresh heifers, and a greater ability to cull poor-doers. This better managed herd may even sell animals to another dairy for milking replacements, yet the apparent culling risk would be much higher than the first herd. Also, some herds tend to play games to make this number look good by waiting until the next 30 d window to cull poor doing animals. Thus, the use of early lactation culling is confounded by potentially unknown herd management issues, highly subject to sample size constraints, and really has very little utility within the area of transition monitoring. Instead of using culling evaluations, herds should strive to develop better disease treatment and monitoring protocols. If a subset of animals is affected severely enough to require culling, there are also likely negative effects on the remaining population of survivors that will adversely impact production and reproduction. Having a system in place to monitor changes in disease risk (i.e., metritis, mastitis, and displaced abomasum) or risk factors for disease, such as milk fever or retained placenta, allows for more timely and appropriate intervention. Record major, consistently defined fresh cow events such as milk fever, DA, RP, mastitis, metritis, and lameness in addition to freshenings. Some events such as ketosis may be too subjective or prone to detection biases and are usually not as valuable to record. However, if recorded, monthly ketosis incidence can be used to evaluate employee performance. Retained placenta risk, calculated on a weekly or monthly basis, can be a very good monitor of both preparturient feed intake as well as a predictor of future metritis risk. Typically, cows are considered retained if the placenta is still present 24 hrs after calving. Monthly risk of displaced abomasum (DA) (number of DA divided by number of fresh cows at risk) can also be helpful to indicate transition problems, but this metric suffers from more lag than RP risk. 47
51 In general, the following fresh cow event risks are achievable goals for most operations: Milk fever less than 3-5 % of mature cow calvings, Displaced abomasum less than 3-5 % of all calvings, and Retained placenta less than 8 % of all calvings. Monitor total days dry and days in the closeup pen. Very short dry periods (less than 30 days) may have negative impacts on the subsequent lactation performance. Equally important are the number of excessively long dry periods due to the cost associated with keeping nonproductive cows on the dairy. Excessively long dry periods often reflect problems such as pregnancy loss and rebreeding that results in pregnant cows that are dried prematurely relative to gestational age due to low milk production, technical error either with conception date estimation or data entry into the record system, or management decisions to move pregnant cows into the dry period (rather than culling them) prior to the normal date range due to decreased milk production. Excessively short dry periods are often the result of errors with conception date estimation or data entry into the record system, shortened gestation length due to abortion or premature delivery, or management mistakes (failed to move the cow at the correct time). In general, with weekly pregnancy evaluations in AI herds and weekly moves to the dry pen, a herd should be able to have about 85% of the dry lengths within 14 days +/- of their stated goal. In natural service herds or herds that move cows less often, the variation will be significantly greater. Most herds strive to achieve about 21 d in a close-up pen in order to modify the ration from a nutritional perspective, manipulate dietary cation anion differences to manage hypocalcemia, to decrease stocking density and to allow for closer observations. However, few herds actually monitor this item. Longer than necessary time spent in close-up is costly and occupies space that other close-up cows may use. Days in close-up of less than 10 may predispose cows to increased risk of periparturient issues. Strive to achieve at least 10 days in close-up for at least 90% of all cows. The evaluation of peak milk by parity group is an approach that has been practiced for many years, but the correlation to overall lactation performance is actually pretty low. In addition, this metric suffers from the consequences of lag, momentum and the difficulties with accurate calculations of an animal s true peak. Therefore, peak milk is a very poor monitor of transition management and its use should be discouraged. Early lactation milk production, first test milk or week 4 milk production estimates are better monitors of transition performance than waiting for peak milk. First test milk is the earliest production data that can be used to evaluate early lactation performance and the impact of transition programs. The lag for this approach is 1 to 3 mo shorter than relying on peak milk and allows for the inclusion of cows that may be culled prior to reaching true peak milk. However, this approach is subject to the impact of DIM at the first test. To correct for this confounding factor, in large herds first test milk can be limited to only evaluating animals that experience first test between 20 and 30 DIM (or some comparable range). A useful approach that has gained in popularity is the use of wk 4 milk. In DC305, an estimate of milk production during the fourth week can be calculated using item type 122 (weekly average milk on week X where X equals 4). This estimate will include data from more cows than only evaluating first test for cows that tested between 20 and 30 DIM and can be used to illustrate the impact of seasonal changes in early lactation performance, as well as showing the impact of management changes. In herds with daily milk meters, changes in milk production can also be a good monitor, but results should be interpreted with caution. In general, cows should increase in milk flow by ~ 10 %/d for the first 14 d and heifers should increase in milk flow by ~ 6 to 8 %/d for the first 14 d. One quick assessment of how parity groups are performing relative to each other is to compare p305me production for parity =1, parity =2 and parity > 2. Interpretation of the results must consider the potential that one or more groups is performing poorly, or conversely, that one or more groups is performing better than expected relative to the remainder of the herd. In most 48
52 herds, the first lactation group s p305me milk estimate will be 2 to 5% lower than older cows. This result is often due to a lower culling pressure for production being applied to these first calf animals. The third and greater lactation group, while representing the oldest genetics, should have a p305me that is at least as good as the second lactation animals or higher since these cows have had multiple lactations of culling pressure. If the performance of the first lactation group is vastly inferior to expectations, potential areas for investigation include frame size at calving, stocking density (especially in mixed parity groups), heifer mastitis challenges, and dystocia challenges unique to the heifers. Common reasons that the first lactation group might be outperforming the other two groups may be related to the purchase of higher genetic merit heifers, more aggressive culling of heifers due to an increase in supply related to the use of sexed semen or due to disease issues in mature cows such as mastitis, metabolic challenges around calving, or lameness. In expanding herds, herds that suffer higher risks for lameness or mastitis, or herds that struggle with reproductive performance, the p305me of the older cows may in fact be 5 to 15% less than the second lactation cows. Milk components can also be used at the herd level to indicate potential transition issues. Fresh cows that mobilize excessive body fat will often demonstrate higher than normal levels of butterfat. On an individual cow basis, the use of either first test fat percentage or fat:protein ratio is not very sensitive for identifying cows at increased risk of subclinical or clinical ketosis. However, at the herd level, examining the fat:protein ratio at first test can provide valuable information. Calculate fat:protein ratio for cows with DIM at first test of 10 to 40. If 40 % or more of this population has a fat:protein ratio > 1.4, further investigation is warranted. Another approach is to look at first test fat percentage alone. In this case, if > 10 % have an excessively high first test fat percent, further investigation may be warranted. Cut-points used by the authors for quick screening are: 5.0 for Holsteins and 6.0 for Jerseys. Summary Transition performance is critical to the success of any dairy. Key components of successful transition include the implementation of herd management guidelines that focus on prevention of periparturient problems, real time monitoring of key processes that impact the prepartum and the postpartum periods, and evaluating the results of the program through the examination of key outcomes. The points covered in this paper are not an exhaustive list of all possible transition management issues or of all the possible approaches to monitoring transition performance. Each herd may have specific approaches that work well for it and consultants all have their preferred approaches for evaluating performance. High quality records and their appropriate use are vital to the evaluation of how well cows are transitioning into lactation, but one must always remember there is no substitute for walking the herd and observing the cow and her environment. The combination of direct observation of housing, cow comfort, nutrition, and general cow health and condition and a careful approach to proper evaluation of records should lead to improved performance in the transition period and beyond. LITERATURE USED Bell, A.W Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation, J. Anim. Sci. 73: Bell, A. W., W. S. Burhans, and T. R. Overton Protein nutrition in late pregnancy, maternal protein reserves and lactation performance in dairy cows. Proc. Nutri. Soc. 59: Butler, W.R., and R.D. Smith Interrelationships between energy balance and postpartum reproductive function in dairy cattle. J. Dairy Sci. 72: Comin, A., D. Gerin, A. Cappa, V. Marchi, R. Renaville, M. Motta, U. Fazzini, and A. Prandi The effect of an acute energy deficit on the hormone profile of dominant follicles in dairy cows. Theriogenology 58: Contreras, L.L., C.M. Ryan, and T.R. Overton Effects of dry cow grouping strategy and prepartum body condition score on performance and health of transition dairy cows. J. Dairy Sci. 87: Drackley, J.K Biology of dairy cows during the transition period: the final frontier? J. Dairy Sci. 82: Drackley, J.K., T.R. Overton, and G.N. Douglas Adaptations of gulcose and long-chain fatty acid metabolism in liver of dairy cows during the periparturient period. J. Dairy Sci. 84:E100-E112E. Duffield, T.F., D.F. Kelton, K.E. Leslie, K.D. Lissemore, and J.H. Lumsden Use of test day milk fat and milk protein to detect subclinical ketosis in dairy cattle in Ontario. Can. Vet. Jour.-Revue Veterinaire Canadienne 38:
53 Duffield, T Subclinical ketosis in lactating dairy cattle. In: Vet. Clin..N. Am.: Food Anim. Prac., p Grant, R., and J.L. Albright. Dry matter intake influenced by cow grouping, behavior. Feedstuffs, Dec 8, p Grummer, R.R Etiology of lipid-related metabolic disorders in periparturient dairy cows. J. Dairy Sci. 76: Grummer, R.R Impact of changes in organic nutrient metabolism on feeding the transition dairy cow. J. Anim. Sci. 73: Hayirli, A., R.R. Grummer, E.V. Nordheim, and P.M. Crump Animal and dietary factors affecting feed intake during the prefresh transition period in Holsteins. J. Dairy Sci 85: Ingvartsen, K.L., and J.B. Andersen Integration of metabolism and intake regulation: a review focusing on periparturient animals. J. Dairy Sci. 83: Moreira, F., C. Risco, M.F.A. Pires, J.D. Ambrose, M. Drost, M. DeLorenzo, and W.W. Thatcher Effect of body condition on reproductive efficiency of lactating dairy cows receiving a timed insemination. Theriogenology 53: Rabelo, E., R.L. Rezende, S.J. Bertics, and R.R. Grummer Effects of transition diets varying in dietary energy density on lactation performance and ruminal parameters of dairy cows. J. Dairy Sci. 86: Reynolds, C.K., P.C. Aikman, B. Lupoli, D. J. Humphries, and D. E. Beever Splanchnic metabolism of dairy cows during the transition from late gestation through early lactation. J. Dairy Sci. 86: Santos, J.E., E.J. DePeters, P.W. Jardon, and J.T. Huber Effect of prepartum dietary protein level on performance of primigravid and multiparous Holstein dairy cows. J. Dairy Sci. 84: Santos, J.E., S.O. Juchem, K.N. Galvao, and R.L. Cerri Transition cow management to reduce metabolic diseases and improve reproductive management. Proc. Western Canadian Dairy Seminar, March 11-14, University of Alberta, Edmonton, Canada. Stokes, S.R., and J.P. Goff Evaluation of calcium propionate and propylene glycol administered into the esophagus at calving. Prof. Anim. Sci. 17: Thatcher, W.W., F. Moreira, J. Santos, and C.R. Staples Factors influencing reproductive effciency. In: Proc. Fifth Western Dairy Conf., p Vandehaar, M.J., G. Yousif, B.K. Sharma, T.H. Herdt, R.S. Emery, M.S. Allen, and J.S. Liesman Effect of energy and protein density of prepartum diets on fat and protein metabolism of dairy cattle in the periparturient period. J. Dairy Sci. 82:

54 Comparison of Immune Function, Uterine Health and More in Holstein and Crossbred Transition Cows Ricardo Chebel University of Minnesota 51

55 52
56 53
57 54
58 55
59 56
60 57

61 58
62 Manureology 101 Michael F. Hutjens University of Illinois, Urbana 232 Animal Sciences Lab 1207 West Gregory Drive Urbana, Illinois Dairy managers watch manure changes as a guide when making feed changes and evaluating rations. Fresh, undisturbed piles of feces or droppings may provide valuable clues and should be part of your tool box when evaluating the nutritional status of the dairy herd. Four aspects of manure evaluation can be considered to be part of your tool box. Washing manure samples Washing manure through a screen (6 to 8 squares to the inch) allows the dairy manager and nutritionist to quickly find or see if feed processing and digestion is optimal. Take a cup of fresh manure and wash it with a stream of warm water (cold water takes longer) through the screen removing the digested material. It typically takes about 30 seconds if your screen has sides allowing for more water pressure. Look for the following remaining feed particles. Finding pieces of barley or corn grain with white starch remaining indicates that some feed value was lost. If the seed and starch pieces are hard, additional grinding or processing may be needed to expose the starch to rumen microbial fermentation or lower gut enzymatic digestion. Corn kernels from corn silage reflect that the seed was too hard for digestion and chewing by the cow. Mature and dry corn silage can cause this observation as grain is hard. Some corn silage varieties can be selected for softer kernels allowing for more digestion. Whole cottonseeds or soybean splits (half of a soybean seed) that appear in the washed manure reflect a loss of feed nutrients. The cottonseeds are not caught in the rumen mat and do not allow for rechewing. If roasted soybean seeds are hard, they must be processed finer. Wisconsin workers suggest breaking soybeans into fourths or eighths. Forage particles over 1/2 inch long may reflect a lack of long forage particles to maintain the rumen mat and adequate cud chewing. A higher rate of passage reduces the time needed in the rumen to digest the fiber properly. The Cargill Manure Separator (NASCO Digestion Analyzer) is commercially available through NASCO (price is $195 plus shipping and handling). Users can decide to use the top screen only or two to three screens depending on time and personal bias and experience. Scoring manure Michigan workers developed a scoring system to evaluate fresh manure. Consistency is dependent on water and fiber content of the manure, type of feed, and passage rate. A scale of 1 to 5 is listed below with a score 3 optimal. Score 1. This manure is very liquid with the consistency of pea soup. The manure may actually arc from the cow. Excess protein or starch, too much mineral, or lack of fiber can lead to this score. Excess urea in the hindgut can create an osmotic gradient drawing water in the manure. Cow with diarrhea will be in this category. Score 2. This manure appears runny and does not form a distinct pile. It will measure less than on inch in height and splatters when it hits the ground or concrete. Cows on lush pasture may have this manure score. Low fiber or a lack of functional fiber can also lead to this manure score. Score 3. This is the optimal score! The manure has a porridge-like appearance, will stack up 1 1/2 to 2 inches, have several concentric rings, a small depression or dimple in the middle, make a plopping sound with it hits concrete floors, and it will stick to the toe of your shoe. Score 4. The manure is thicker and stacks up over 2 inches. Dry cows and older heifers may have this type of manure (this may reflect that low quality forages are fed and/or a shortage of protein). Adding more grain or protein can lower this manure score or improve forage quality. Score 5. This manure appears as firm fecal balls. Feeding a straw-based diet or dehydration could contribute to this score. Cows with a digestive blockage may exhibit this score. Manure scores 1 and 5 are not desirable and may reflect a health problem besides dietary limitations. Score under score 2 and over score 4 manure scores may reflect a need to rebalance the ration. As cows progress through their lactation, manure score may also shift as outlined below. Early lactation cows (2 1/2 to 3) Late lactation cows (3 to 3 1/2) 59
63 Far off dry cows (3 to 4) Close up dry cows (3 to 3 1/2) Increasing the amount of degradable, soluble, or total protein, deceasing the amount or physical form of the fiber; increasing starch level, decreasing grain particle size (such as fine grinding or steam flaking), and consuming excess minerals (especially potassium and sodium) can cause manure scores to decline. A University of Illinois study investigated relationships between dairy fecal scores and physical fiber property based on manure score, fiber particle size, and fecal dry matter content. Fecal samples were collected with 17 pooled fecal samples from 42 dairy cows based on their fecal score (1 to 5). After collecting fresh samples over three weeks, the samples were labeled, mixed, washed, and dried. The amount collected was 500 g of manure per sample. For samples less than 500 g, an adjustment was calculated to correct to 500 g. Each sample was collected based on its manure score and given a letter assigned (A-F). Samples were washed using warm water with a Cargill Manure Separator containing three stainless steel screens (3/16, 3/32, and 1/16 inch hole openings). The fibrous fraction on each screen was oven dried for two days at 45 degrees centigrade. Each sample was weighed and recorded. Individual samples and fecal score summary were listed in table 1. Total dry matter in samples increased as manure score increased. The percentage of dry matter on the top screen (the largest particle size) increased as manure score increased (Table 2) while the percent of dry matter on the bottom and middle screens decreased. The number of whole fuzzy cottonseeds found in the feces samples increased as fecal scores increased. During week two of manure collection, the ration contained 50 percent less fuzzy cottonseed (2.5 pounds compared to 5 pounds). This decrease was reflected in the number of whole fuzzy cottonseeds found in the fecal samples in week two. Manure color The color of manure is influenced by feed, amount of bile, and passage rate. Manure from cows on pasture is dark green while hay-based rations are brown. Manure from high grain-based diets is more graylike. Slower rates of passage cause the color to darken and become more ball-shaped with a shine on the surface due to mucus coating. Score 1 may be more pale due to more water and less bile content. Hemorrhage in the small intestine causes black and tar-like manure while bleeding in the rectum results in red to brown discoloration or streaks of red. Fecal starch evaluation As corn prices have increase, the need to optimize the starch digestibility in the dairy cow continues to important. Kernel or plant processing corn silage, grinding corn grain (900 to 1100 microns), invitro rumen starch fermentation values, level of prolamin (type of protein related to vitreousness), and the NRC energy values based on the rumen model program illustrate the important of total tract starch digestibility. University of Pennsylvania published a formula to predict total tract starch digestibility using fecal starch and fecal lignin along with ration starch and lignin. Lignin was used as a marker to estimate starch utilization. This approach could be useful to evaluate starch utilization on farms. The Pennsylvania data concluded that for each increase in fecal starch, the potential loss in milk yield was 0.7 pound per day with a range of 4 to 10 percent starch in group or herd values. Cumberland Valley Lab analyzed 1420 fecal samples with starch content ranging from 0.20 to 38.9 percent with 62 percent containing less than five percent starch. Rock River Lab reported 52 samples averaged 7.9 percent starch with an apparent total tract digestibility of 84.8 percent. An Illinois field study using nineteen Holstein herds in southwestern Illinois evaluated the fecal starch digestion using the Pennsylvania equation. The manure samples used in this project were taken from undisturbed, fresh pies in the cow lot. On average, four to five pies were sampled, mixed together, and placed into the quart sized containers supplied by Rock River Lab. They were refrigerated until all samples were collected and shipped by UPS to the lab using cold packs to prevent the samples from bursting out of the containers in transit. Rock River Labs conducted feed and fecal starch and NDF analysis. Results were statistically analyzed using SAS software to evaluate feed and fecal results and statistically determine which parameters were significant in predicting a prediction model for starch digestibility. Table 2 summarizes data collected at each farm illustrating variation from farm to farm. Table 3 60

64 summarizes the average, standard deviation, and range associated with each variable evaluated. Milk yield was converted to 3.5% fat-corrected milk using plant milk fat test. Feed values were calculated using rations balancing software and forages tested results. The equation from University of Pennsylvania used to estimate starch digestibility is listed below that calculated starch total tract digestibility values in Table 2. The R-squared value for the equation was 73 indicating 73 percent of the variation in starch digestibility can be explained by the equation and variable used with P < The equation must be used with care as the values are based on 19 Holstein herds, the manure samples procedure, and analytical lab procedures. Additional studies are needed to confirm and/or refine the equation. Dairy managers and consultants may want to add fecal starch and fecal lignin analysis to monitor starch utilization by their dairy herd. The cost of fecal starch analysis is $15 to $20 per sample. The recommendation is to pool fecal samples from 10 to 15 cows and submit a mixed sample requesting fecal starch. Fecal starch can be another tool to evaluate total track starch utilization. Selected References DeOndarza, M.B. and R. Ward Starch in manure can t make milk. Hoard s Dairyman Magazine. Feb 25 issue. p Lidy, D, J. S. Osorio, M.F. Hutjens, and D.W. Meyer Evaluating total tract starch digestibility. IL Dairy Report. pp Jones, J and M.F. Hutjens Relationship of fecal scores and fecal fiber levels. IL Dairy Report. pp Skidmore, A.L., A. Brand, and C.J. Sniffin Monitoring milk production: Defining preset targets and execution. Herd Health and Production Management. Wageningen Pers, Wageningen, Netherlands. pp Percent starch apparent digestibility = 1 ((% lignin in feed x % starch in feces) / (% lignin in feces x % starch in feed)) When entering all variables in Table 4, fecal starch and fecal NDFD were the two variable that were correlated with a P < 0.01 to calculated starch digestibility. Statistical trends (P > 0.05) were observed for feed NDF levels and fecal dry matter content. Based on the statistical evaluation, the following Illinois prediction equation was developed: Percent starch apparent digestibility = ( x fecal starch) + ( x fecal lignin). 61

65 Troubleshooting Silage Yeast, Mold and VFA Problems Bill Mahanna, Ph.D., Dipl. ACAN Nutritional Sciences Manager - Pioneer, A DuPont Business bill.mahanna@pioneer.com;

66 63

67 64

68 65

69 66

70 67

71 68

72 69

73 70

74 71
75 72

76 Improving Feeding Consistency Through TMR Audits Tom Oelberg, Ph.D. Diamond V toelberg@diamondv.com 73

77 74

78 75

79 76

80 77

81 78

82 79

83 80

84 81

85 82

86 83
87 The Chemistry of High Moisture Corn P.C. Hoffman, R.D. Shaver and N.M. Esser Department of Dairy Science University of Wisconsin-Madison Introduction The first trial reported in the Journal of Dairy Science evaluating the nutritive value of high moisture corn (HMC) for lactating dairy cows was published by Zogg et al., (1961). The use of HMC on commercial dairy farms has grown, with approximately 65 % of Wisconsin dairy producers now utilizing HMC (Holmes, 2010). Despite wide spread use, HMC is an enigmatic feed because HMC per se is not a homogeneous feed. A specific HMC corn fed to a lactating dairy cow can be a highly variable feedstuff. Corns of varying endosperm type can be ensiled between 20%-40 % moisture, with or without inoculants, at ambient temperatures of o F, ensiled whole or ground, treated with or without organic acids, contain cob or husk, be fermented 1 to 365 days or more, stored in bags, bunkers and oxygen limiting silos and still be classified as HMC. Variance associated with HMC production practices has been hypothesized or shown to create variance in fermentation characteristics, chemical composition, starch digestibility and milk yield in lactating dairy cows. The effects of chemical alteration of HMC on animal performance is challenging to quantify because most if not all HMC studies in the literature did not measure detailed chemical compositions of the HMC fed. As a result, general review articles (Firkins et al., 2001; Huntington, 1997, Owens et al., 1986) pertaining to starch digestibility in ruminants have focused on animal responses in trials where chemically undefined HMC was fed. Differences in ruminal, post-ruminal or total tract starch digestibilities between various grain sources and HMC can be generally categorized, but animal responses are challenging to directly link to HMC chemistry. Absent from the literature, are defining chemical mechanisms associated with fermentation, which explain altered starch digestion of HMC in ruminants. In short, altered starch digestion in lactating dairy cows fed HMC is presently binomially classified (i.e. dry vs HMC) but mechanisms explaining chemical alterations in HMC as compared to dry corn and variance of these alterations within a multitude of HMC production practices are poorly defined. This paper will focus on the chemistry of HMC during fermentation and attempt to provide inference regarding why HMC starch digestibility in ruminants is altered as compared to other grain sources such as dry corn. A greater understanding of HMC chemistry may yield a better understanding of animal performance variance associated with feeding HMC of various origins. High Moisture Corn-Starch Digestibility Reviews (Firkins et al., 2001; Huntington, 1997, Owens et al., 1986) pertaining to factors and limitations of starch digestibility in ruminants have been previously published and will not be rereviewed in this proceedings paper. From these reviews and trials on feeding HMC the following general concepts of HMC starch digestibility in lactating dairy cows can be defined (refer to Table 1). Figure 1. The effect of storage period (240 d) on hydrophobic prolamin-zein proteins in the endosperm of high moisture corn (Hoffman et al., 2010a). 1) The NE L value of HMC is estimated to be 5-10 % greater than dry corn of similar origin and particle size (Tyrrell and Varga, 1987; Wilkerson et al., 1997). Greater NE L values for HMC are primarily due to increased total tract starch digestion of HMC as compared to dry corn. 84
88 2) Ensiling of corn alters the site of starch digestion. Ruminal starch digestibility of HMC starch is commonly percentage units higher than dry corn (Firkins et al., 2001; Owens, 2005) 3) The rate of ruminal starch digestion is faster for HMC as compared to dry corn (Sniffen et al., 1992). 4) Increased ruminal starch digestion and increased starch digestion rates may decrease ruminal NDF digestion (Firkins et al., 2001, Knowlton et al., 1998, Oba and Allen, 2003). 5) Feeding high levels of HMC with high rumen fermentability may depress DMI, rumen ph or milk fat test (Owens et al., 1986, Bradford and Allen, 2007, Firkins et al., 2001). Depressions in DMI are thought to be modulated by a glucoseinsulin response effect (Bradford and Allen, 2007). Starch Digestibility From a Corn Seed Perspective Corn per se is not a feed, it is a seed, and some understanding of corn seed anatomy and physiology are required to better understand chemical factors that potentially influence starch digestibility in ruminants. The corn seed is comprised of three basic morphological parts, pericarp, germ, and endosperm. The endosperm represents approximately percent of the corn kernel by weight and is the morphological structure which contains starch. The endosperm contains primarily starch and protein but does contain small amounts of fat as phospholipids and ash. The endosperm of corn is virtually devoid of fiber (ADF or NDF). Specifically, corn endosperm contains < 4% NDF, as compared to the germ which contains 17% NDF, and the pericarp with 33% NDF (Van Kempen et.al., 2003). Corn endosperm contains storage proteins (albumins, globulins, glutelins and the abundant prolamins (zein protein) which are hydrophobic. The endosperm s biological function is to serve as the primary nutrient source for the embryo until photosynthesis is initiated upon seedling emergence (Buchanan, et al., 2000; Mohr and Schopfer, 1995). The basic morphological parts of the corn seed are not equally digestible in ruminants. The pericarp is the primary morphological structure protecting both the embryo and endosperm. In native form, the pericarp of corn is poorly digested by rumen bacteria with minimal post ruminal digestion potential. Corn pericarp is relatively resistant to rumen bacteria attachment (Huntington, 1997) and whole corn seeds with the pericarp intact are largely indigestible in the small and large intestines of ruminants (Owens et al., 1986). Digestion of starch, contained in the endosperm, by ruminants is enigmatic because starch isolated from cereal grains, regardless of source is rapidly attacked by rumen microbes and fermented rapidly (Hibberd et al., 1983). Starch in corn endosperm is, however, not in isolated form. Starch in a corn seed is protected by hydrophobic (repels water) proteins called prolamins (zein). The combination of starch, prolamins and other proteins (albumins, globulins, glutelins) in corn endosperm is often referred to as the starch-protein matrix. The starch-protein matrix binds starch granules together and the degree of binding alters the grinding efficiency of corn (Fox and Manley, 2009) and the ability of, and surface area for, rumen bacterial attachment (Huntington, 1997). In corn, prolamin proteins named zein are the primary protein in the starch protein matrix, and comprise % of the total protein in whole corn (Hamaker et al., 1995). Prolamin-zein, defines a class of hydrophobic proteins synthesized on the rough endoplasmic reticulum of the amyloplast (starch producing organelle) envelope consisting of four zein sub-classess (ά,β,γ,δ) (Buchanan, et al., 2000). Because prolamin-zein proteins are synthesized on the rough endoplasmic reticulum within the amyloplast without the presence of transit genes (Buchanan et al., 2000), prolamin-zein proteins are not intrinsic within the starch granule but are primarily surface localized on the exterior of starch granules (Mu- Forster and Wasserman, 1998). As prolamin-zein proteins enlarge and distend with advancing maturity β- and γ- zeins cross-link and ά- δ-zeins penetrate their network and occupy a more central position encapsulating starch into a starchhydrophobic protein matrix (Buchanan et al., 2000; Mu-Forster and Wasserman, 1998). Differences in the starch-protein matrix can be visibly seen in dissected kernels of yellow dent corn. The visual appearance of all or portions of the starchprotein matrix in corn endosperm have historically been given visually descriptive classifications. Starch-protein matrices appearing white are commonly given the names floury, opaque or soft endosperm. Starch-protein matrices appearing yellow, shiny or glassy are classified as, horny, translucent or vitreous (Kempten, 1921). The Starch-Protein Matrix and High Moisture Corn The starch-protein matrix in corn has been previously defined as a physio-chemical impediment to starch digestion in ruminants (Owens et al., 1986), but the role of the starch- protein matrix in the digestion of HMC starch in ruminants is not well defined. Because prolamin-zein increases with advancing maturity (Murphy and Dalby, 1971), lower prolaminzein contents in HMC at ensiling could be expected. This argument is somewhat illogical because Murphy and Dalby (1971) observed that maximum prolamin- 85
89 zein accretion occurred near black layer formation (± 30 % moisture), which is similar to typical ensiling moisture contents of HMC. In addition, HMC and dry corn are often harvested (combined) at very similar moisture contents with only post-harvest handling and storage of the corn being different thereafter. Specifically, corn is commonly combined at 25%-30 % moisture and mechanically dried thereafter yielding dry corn. A more plausible explanation for greater and more rapid starch digestion of HMC starch is that fermentation acids or proteolysis degrade prolaminzein proteins during the ensiling process. Bacterial proteolysis is an intrinsic mechanism in corn-grain fermentation which induces degradation of corn proteins (Baron et al., 1986). Philippeau and Michalet-Doreau (1998) observed that ensiling grains increased ruminal starch degradability and hypothesized that ensiling increases accessibility of starch granules to rumen microorganisms, because hydrophobic prolamin-zein proteins encapsulating starch granules were partially degraded by proteolysis. Likewise, Jurjanz and Monteils (2005) observed the effective ruminal degradability of starch to be lower in corn kernels before (70.2%) than after (92.3%) ensiling. The ensiling process improved starch degradation by significantly altering the rapidly-degradable starch fraction (80.7% versus 65.6%) and the starch degradation rate (12.4 vs 8.0 %/h). Combined, these data (Baron et al., 1986; Philippeau and Michalet-Doreau, 1998; Jurjanz and Monteils, 2005) result in a very plausible hypothesis as to why higher ruminal and total tract starch digestibility is observed for HMC as compared to dry corn (Firkins, et al., 2001). In a recent study, (Hoffman et al., 2010a) we monitored the fate of the starch-protein matrix in HMC across a long storage period (240 days). Two random HMC(s), containing 25.7% and 29.3 % moisture were ground, ensiled and stored for 0, 15, 30, 60, 120 and 240 d. At 0 and 240 d, the ά, γ, δ and β zein regions of the starch-protein matrix were profiled using high performance liquid chromatography. The effect of fermentation (storage time) on the starch-protein matrix of HMC after 240 d of storage is presented in Figure 1. Fermentation (0 vs 240 d) reduced all ά, β and δ prolamin-zein subunits of the starch-protein matrix from 10%-40 %. The degradation of the γ prolamin-zein subunits of the starch-protein matrix of HMC was more extensive with a 60 % reduction. Because γ prolamin-zeins are surface localized and primarily responsible for crosslinking starch granules together, the degradation of γ zeins in HMC would suggest that clusters of starch granules should disassociate (fall apart) as a result of fermentation since the cross links holding starch granules together are being degraded. This was confirmed by electron microscopy (photos not shown) of HMC starch granules at 0 and 240 d.. Upon fermentation and storage for 240 d, the disassociation of starch-granule clusters in HMC could be readily seen using electron microscopy. Fermentation resulted in a greater number of individual starch granules (and surface area) for potential attack by rumen bacteria. Electron micrographs also revealed no alteration in individual starch granules in HMC prior to fermentation or after 240 d of storage. Inferences from this investigation (Hoffman et al., 2010a) also suggested the proteins in the starch-protein matrix were more likely altered by bacterial proteolysis and may not have been simply solubilized by fermentation acids. In second study (Hoffman et al., 2010b), the digestibility of HMC fermented and stored for 0, 15, 30, 60, 120 and 240 d was evaluated using an in vitro gas production system. Gas production and rate (kd) of gas production by rumen bacteria during the first 12 h of incubation increased with increasing storage time, which indirectly validates the observations of greater ruminal starch digestion of HMC as compared to unfermented corn. Increases in 12 h gas production and rate (kd) of gas production increased chronically over the entire HMC storage periods suggesting that the increase in HMC (DM) digestion is not an acute event. Similar results were reported by Benton et al. (2005) who evaluated in situ DM degradation of two HMC(s) and two reconstituted HMC(s) of varying moisture content; a chronic increase in DM degradation across a 300(+) day ensiling period was observed. The observations of Benton et al. (2005) and Hoffman et al. (2010b) are presented in Figure 2. The data are similar and when 86
. B = Hoffman et al., 2010b ( = 29.3% moisture HMC, = 25.")

90 combined suggest DM that the digestion potential of HMC increases chronically as storage time increases. Figure 2. Changes in high moisture corn DM degradation across long ensiling periods. A= Benton et al., 2005 ( = 24% moisture HMC, x = 28% moisture reconstituted corn, = 30% moisture HMC, = 35% moisture reconstituted corn). B = Hoffman et al., 2010b ( = 29.3% moisture HMC, = 25.7% moisture HMC). In situ Dacron bags and in vitro gas production were used as evaluation techniques by Benton et al., (2005) and Hoffman et al., (2010b) respectively. High Moisture Corn Fermentation is Poor and Slow In our recent research (Hoffman et al., 2010a) we evaluated the nutrient composition (CP, prolamin, starch, ADF and NDF), fermentation (ph, lactate and acetate) and protein degradation markers (buffersoluble CP and NH 3 -N) of HMC at 0, 15, 30, 60, 120 and 240 d of storage. The data provided a holistic view of nutri-chemical transformations in HMC by storage time. The CP, origin prolamin, starch, ADF and NDF contents of HMC are relatively static, but ph, lactate, acetate, buffer-soluble CP and NH 3 -N contents chronically changed with advancing storage time. The chronic changes in HMC NH 3 -N content by storage time are presented in Figure 3. Ammonia (NH 3 -N) in HMC did not stop accumulating even at 240 d. Ammonia is an important marker in fermented feeds, because NH 3 -N is intrinsic to deamination of amino acids which is the terminal phase of proteolysis (protein breakdown). The data in Figure 3 suggest that proteolysis in HMC did not abate even after 240 d of storage. These data present an indication that fermentation of HMC is very slow and chemical alterations are occurring over an extended storage period. It is logical that HMC ferments slower than legume or corn silage for a number of reasons. Normally HMC, prior to ensiling, does not contain high levels of sugars or water soluble carbohydrates for conversion to volatile fatty acids. The levels of monodisaccharides and water soluble carbohydrates of legumes, whole-plant corn and corn grain prior to ensiling (Dairy One Laboratories, Ithica, NY) are presented in Figure 4. Fresh corn contains significantly less mono-disaccharides or water soluble carbohydrates than legumes or whole-plant corn. Because the level of fermentable substrate in fresh corn is lower than fresh legumes or whole-plant corn, the production of volatile fatty acids in HMC is reduced when compared to legume or corn silage (Figure 5). Figure 4. Typical contents of mono-disaccharides and water soluble carbohydrates (CHO) in legumes, corn silage and high moisture corn prior to ensiling (Dairy One Laboratories, Ithaca NY. Figure 5. Typical ph, lactate and acetate contents of legume silage, corn silage and high moisture corn (Dairy One Laboratories, Ithaca, NY). Figure 3. The effect of storage time of on NH 3 -N of four high moisture corns (Hoffman et al., 2010a) High moisture corn is also ensiled at DM contents near 70.0%. High DM contents increase the osmotic potential in silage (or HMC), which decreases the growth rate of lactic acid producing silage bacteria (Pitt et al., 1985). As a result, silage or HMC with higher DM content ferment slower than silage or 87
91 HMC with a lower DM content. Slower growth rates of silage bacteria induced by ensiling at high DM contents can be further exacerbated by ensiling at low temperatures (Pitt et al., 1985). The effect of silage mass temperature on growth of lactic acid producing bacteria is presented in Figure 6. High moisture corn is commonly ensiled when ambient temperatures are < 50 o F, which does not facilitate rapid bacterial growth. The combination of limited substrate for fermentation, high DM content, and ensiling at cooler temperatures suggest HMC fermentations are destined to be protracted. Protracted fermentations would mean a protracted proteolytic breakdown of the starch-protein matrix in the endosperm. In our recent work (Hoffman et a., 2010ab), we observed this effect; protracted fermentations and protein alterations in HMC which resulted in chronic increases over time in storage for 12 h in vitro gas production and rates of gas production. Figure 6. Effect of ensiling temperature on silage bacteria growth rate (Calculated from Pitt et al., 2005) Conclusions The starch protein matrix in HMC is significantly altered by the fermentation process, especially γ zein proteins which cross link starch granules together. Fermentation induced degradation of γ zein proteins in HMC appears to yield a general disassociation of starch granule clusters yielding more individual starch granules and (or) surface area for potential bacterial attack. The degradation of the starch protein matrix in HMC appears to be chronic and slow. Fermentation and storage time chronically increases the DM digestion potential of HMC. Traditional feed chemistry nutrients in HMC (ADF, NDF, CP and starch) are static across the storage period and do not appear well suited for determining biochemical factors that influence starch digestibility of HMC in ruminants. High moisture corn is not a static feedstuff with a fixed or book value nutrient composition. Nutrient availabilities in HMC chronically change and changes are likely dependent on physical processing, the strength of the starchprotein matrix at ensiling, fermentation conditions at ensiling (DM and temperature), and the length of the storage period. REFERENCES Baron, V.S., K.R. Stevenson, and J.G. Buchanan-Smith Proteolysis and fermentation of corn-grain ensiled at several moisture levels and under several simulated storage methods. Can. J. Anim. Sci. 66: Bradford, B.J., and M.S. Allen Depression in feed intake by a highly fermentable diet is related to plasma insulin concentration and insulin response to glucose infusion. J. Dairy Sci. 92: Benton, J.R., T. Klopfenstein, and G.E. Erickson Effects of corn moisture and length of ensiling on dry matter digestibility and rumen degradable protein. Nebraska Beef Cattle Reports: Buchanan, B.B., W. Gruissem, and R.L. Jones Biochemistry and Molecular Biology of Plants. Am. Soc. of Plant Physiol., Rockville, MD. Fox, G. and M. Manley Hardness methods for testing maize kernels. J. Agric. Food Chem. 57: Hamaker, B.R., A.A. Mohamed, J.E. Habben, C.P. Huang, and B.A. Larkins, Efficient procedure for extracting maize and sorghum kernel proteins reveals higher prolamin contents than the conventional method. Cereal Chem. 72(6): Hibberd, C.A., D.G. Wagner, R.L. Hintz, and D.D. Griffin Effect of sorghum grain variety and processing method on the site and extent of starch digestion in steers. Oklahoma Agr. Exp. Sta. MP-114:28. Hoffman, P.C., N.M. Esser, R.D. Shaver, W. Coblentz, M.P. Scott, A.L. Bodnar, R. Schmidt and B. Charely. 2010a. Influence of inoculation and storage time on alteration of the starch-protein matrix in high moisture corn. J. Dairy Sci. 93(Suppl. 1):in press. Hoffman, P.C., N.M. Esser, R.D. Shaver, W. Coblentz, M.P. Scott, A.L. Bodnar, R. Schmidt and B. Charely. 2010b. Influence of inoculation and storage time on in vitro gas production of high moisture corn. J. Dairy Sci. 93(Suppl. 1):in press. Holmes, B A field survey of high moisture corn use on Wisconsin dairy farms. University of Wisconsin- Extension (Unpublished survey: personal communication). 88
92 Huntington, G.B Starch utilization by ruminants: from basics to the bunk. J. Anim. Sci. 75: Jurjanz, S., and V. Montels Ruminal degradability of corn forages depending on the processing method employed. Anim. Res. 3: Kempten J.H Waxy endosperm in coix and sorghum. J. Heredity. 12: Knowlton, K. F., B. P. Blenn, and R. A. Erdman Performance, ruminal fermentation, and site of starch digestion in early lactation cows fed corn grain harvested and processed differently. J. Dairy Sci. 81: Mohr, D., and P. Schopfer Plant Physiology. Springer-Verlag, Berlin, Germany. Mu-Forster, Chen, and Bruce P. Wasserman Surface localization of zein storage proteins in starch granules frim maize endosperm: Proteolytic removal by thermolysin and in vitro cross-linking of granule-associated polypeptides. Plant Physiology. 116: Murphy, J.J., and A. Dalby Changes in the protein fractions of developing normal and opaque-2 maize endosperm. Cereal Chem. 48: Oba, M., and M.S. Allen Effects of corn grain conservation method on ruminal digestion kinetics for lactating dairy cows at two dietary starch concentrations. J. Dairy Sci. 86: Owens, F.N., R.A. Zinn and Y.K. Kim Limits to starch digestion in the ruminant small intestine. J. Anim. Sci. 63: Owens, F.N Corn grain processing and digestion. University of Minnesota-Extension, Philippeau, C., and B. Michalet-Doreau Influence of genotype and ensiling of corn grain on in situ degradation of starch in the rumen. J. Dairy Sci. 81: Pitt, R.E., R.E. Muck, and R.Y. Leibensperger A quantitative model of the ensilage process of lactate silages. Grass and Forage Sci. 40: Sniffen, C.J., J.D. O Connor, P.J. Van Soest, D.G. Fox and J.B. Russell A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 70: Tyrrell, H.F., and G.A. Varga Energy value for lactation of rations containing ground whole ear maize or maize meal both conserved dry or ensiled at high moisture. Eur. Assoc. Anim. Prod. 32: Van Kempen T.A., E. van Heugten, A.J. Moeser, Dehulled, degermed corn as a preferred feed ingredient for pigs North Carolina Annual Swine Report, North Carolina State University, Raleigh, N.C. Wilkerson, V.A., B.P. Glenn, and K.R. McLeod Energy and nitrogen balance in lactating dairy cows fed diets containing dry or high moisture corn in either rolled or ground form. J. Dairy Sci. 80: Zogg, C.A., R.E. Brown, K.E. Harshbarger, and K.A. Kendall Nutritive value of high moisture corn when fed with various silages to lactating dairy cows. J. Dairy Sci., 44:
93 Improving Starch Digestibility in Dairy Cows: Opportunities with Reduced-Starch Diets R.D. Shaver Department of Dairy Science 280 Animal Sciences Building 1675 Observatory Drive, Madison, WI University of Wisconsin Take Home Messages Reduced starch diets may be acceptable for lactating dairy cows when using digestible or soluble fiber to partially replace starch. Type of endosperm, particle size, maturity and moisture content, and conservation and processing methods, influence the digestibility of starch from corn grain and corn silage by dairy cows. The inclusion of exogenous enzymes with amylolytic activity in reduced-starch diets for dairy cows may play a role in the future to increase feed conversions. Introduction The optimum starch content of diets fed to lactating cows is not well defined, but 24% to 26% starch (DM basis) has been suggested (Staples, 2007). Kaiser and Shaver (2006) and Bucholtz (2006), from surveys of high producing (>13,650 kg RHA) dairy farms in Wisconsin (n = 9) and Michigan (n = 18), respectively, reported starch concentrations of diets fed to lactating cow groups ranging from 25% to 30% (DM basis). With today s higher grain prices, however, there is increased interest in feeding diets that are lower in starch content than what has been the norm. The purpose of this paper is to evaluate the potential for using digestible or soluble neutral detergent fiber (NDF) from byproduct feeds to partially replace starch from corn grain in diets fed to lactating dairy cows. Potential modifying effects of starch digestibility on lactation performance by dairy cows fed reduced starch diets will also be examined. Partial replacement of dietary starch with digestible or soluble NDF Beet pulp Voelker and Allen (2003) fed mid-lactation cows diets containing 35, 31, 27 and 18% starch (DM basis); high-moisture corn was replaced by 6, 12 and 24% pelleted beet pulp (DM basis) to formulate diets with decreasing starch content. Effects of decreasing dietary starch content were linear (P < 0.05) for dry matter intake (DMI) and quadratic (P < 0.07 and 0.03, respectively) for FCM and fat yields. Relative to the average for the 27 and 31% starch diets, feeding the 18% starch diet reduced DMI, FCM yield and fat yield by 9%, 4% and 5%, respectively; true protein (TP) content and yield were numerically reduced by 4% and 5%, respectively. Citrus pulp Broderick et al. (2002) fed mid lactation cows diets containing 31 or 20% starch (DM basis); dry cracked corn was replaced by 19% dried citrus pulp (DM basis) to formulate the low-starch diet. Feeding the low starch diet reduced DMI (P < 0.02), milk yield (P < 0.02), fat yield (P < 0.03), TP content (P < 0.01) and yield (P < 0.01) by 8%, 11%, 14%, 4% and 20%, respectively. Soy hulls For the set of trial diets which contained 50% forage comprised of 50:50 corn silage: alfalfa silage formulated for 30%, 26% and 22% starch (DM basis) by partially replacing dry ground shelled corn with soyhulls, Weiss et al. (2009) observed in mid-lactation cows across decreasing dietary starch concentrations: DMI = 24.0, 24.5 and 25.0 kg/d; milk yield = 37.9, 38.3 and 37.6 kg/d; energy-corrected milk yield = 41.7, 40.2 and 41.2 kg/d. Stone (1996) reported no differences between 25%- and 16%-starch diets fed to early lactation cows with high-moisture corn being replaced by 19% soyhulls (DM basis) to formulate the low-starch diet. Ipharraguerre et. al (2002) fed mid-lactation cows diets containing 28, 23, 17, 13 and 7% starch (DM basis); dry ground corn was replaced by 10, 20, 30 and 40% pelleted soyhulls (DM basis) to formulate diets with decreasing starch content. Decreasing dietary starch content decreased linearly DMI (P < 0.06) by 7% and increased linearly fat content (P < 0.004) and fat yield (P < 0.001) by 8% and 10%, respectively. Yield of TP tended (P < 0.09) to be reduced by 5% for the lowest-starch diet relative to the 28%-starch diet. There were no differences between the 17%- and 23%-starch diets. Comparing the average of the 7%- and 13%-starch diets to the average of the 17%-, 23%- and 28%-starch diets, 90
94 feeding the low starch diets numerically reduced DMI, milk yield and TP yield by 6%, 3% and 3%, respectively; milk fat content and yield were each numerically increased by 8%. Corn gluten feed Staples (2007), from a review of 14 trials with lactating dairy cows where corn gluten feed partially replaced grains, protein meals or forages with dietary starch concentrations ranging across the trials from 15% to 40% (DM basis), concluded that 21%-starch diets may be acceptable. High-fiber byproduct mixtures Ranathunga et al. (2010) fed mid-lactation cows diets containing 29, 26, 23, and 20% starch (DM basis); dry ground corn was partially replaced by a mixture of distillers dried grains (DDGS) and soyhulls (approximately 2:1 DDGS: soy hulls) to formulate diets with decreasing starch content. Decreasing dietary starch content decreased (P < 0.01) linearly DMI, but actual and energy-corrected milk yields were unaffected by treatment. Thus, decreasing dietary starch content increased (P < 0.06) linearly energy-corrected milk feed conversion from 1.47 to 1.61 kg/kg DMI. It should be noted that dietary crude fat concentrations increased from 4.4 to 5.5% (DM basis) from the added DDGS as dietary starch content was decreased from 29 to 20%, which may have influenced the DMI and feed conversion response observed in this study. Batajoo and Shaver (1994) fed mid-lactation cows diets containing 30, 26, 21, and 15% starch (DM basis); dry ground corn was partially replaced by 0-10% wheat midds, 3-20% brewers dried grains and 0-9% soyhulls (DM basis) to formulate diets with decreasing starch content. Decreasing dietary starch content decreased (P < 0.05) linearly DMI, TP content and TP yield by 7%, 4% and 6%, respectively, and increased (P < 0.05) linearly fat content by 3%. Adverse effects of low starch diets on DMI, TP content and TP yield were more apparent for the 15%-starch diet than the 21% starch diet. Summary Results from these relatively short-term dairy cattle feeding trials suggest that reduced-starch diets formulated by partially replacing corn grain in diets with high-fiber, low-starch byproduct feedstuffs may be feasible, but longer-term lactation trials and trials with higher producing cows are warranted. Further, the cost of using digestible or soluble NDF from byproduct feeds to partially replace starch from corn grain needs to be evaluated for the various high-fiber byproduct sources on a local basis relative to corn and protein supplement prices. Starch digestibility Total tract digestibility of starch by dairy cows is variable ranging from 70% to 100% (Firkins et al., 2001). Various factors, particle size (fine vs. coarse grind), grain processing (steam flaked vs. dry rolled), storage method (dry vs. high-moisture corn), moisture content of high-moisture corn, type of corn endosperm, and corn silage maturity and processing, influence the digestibility of starch by dairy cows (Firkins et al., 2001; Johnson et al., 1999; Nocek and Tamminga, 1991). Kernel vitreousness, the ratio of vitreous to floury endosperm, has been used to assess the type of corn endosperm (Ngonyamo-Majee et al., 2008a, b). Increased kernel vitreousness reduced ruminal in situ corn starch degradation (Correa et al., 2002; Ngonyamo-Majee et al., 2008b). Kernel vitreousness was lower and ruminal in situ starch degradation was greater for dry corn with floury or opaque endosperm than with normal dent endosperm (Ngonyamo-Majee et al., 2008a, b). Taylor and Allen (2005) reported greater ruminal and total tract starch digestibilities in ruminally and duodenally cannulated lactating dairy cows for floury (3% vitreousness) vs. normal dent (67% vitreousness) endosperm dry corn. Highly vitreous corn types contain higher concentrations of prolamin proteins than floury or opaque corn types (Hamaker et. al., 1995; Larson and Hoffman, 2008). Starch granules in the corn endosperm are surrounded by hydrophobic prolamin proteins which are slowly degraded (McAllister et al., 1993). Lopes et al. (2009) conducted an experiment to evaluate the effect of type of corn endosperm on nutrient digestibility in lactating dairy cows using near-isogenic variants of a normal dent endosperm hybrid carrying floury-2 or opaque-2 alleles. The percentage vitreous endosperm was zero for floury and opaque endosperm corns and 64% for the vitreous corn. Prolamin protein content of floury and opaque endosperm corns was 30% of the content found in vitreous corn. Starch disappearance after 8- hr ruminal in situ incubation was 32%-units on average greater, respectively, for floury and opaque endosperm corns than vitreous corn. Total-tract starch digestibility was 6.3%-units, on average, greater for cows fed diets containing floury and opaque endosperm corns than vitreous corn. The impact of the digestibility of corn grain starch on lactation performance by dairy cows was reviewed by Firkins et al. (2001). Based on regressions from the data of (Firkins et al., 2001), increasing starch digestibility increased milk and protein yields (R 2 = 0.89; P < 0.01) and reduced milk fat percentage (R 2 = 0.58; P < 0.05) but not yield. When in concurrence 91
95 with the feeding of low-starch diets, however, increasing starch digestibility may be less likely to result in reduced milk fat percentage. But, research is limited on the impact of increasing starch digestibility in reduced-starch diets on lactation performance by dairy cows. Reduced starch diets and improving starch digestibility Gencoglu et al. (2010) conducted a lactation trial to determine lactation performance responses in highproducing dairy cows to a reduced-starch versus a normal-starch diet and to the addition of exogenous amylase to the reduced-starch diet. Some exogenous enzymes are resistant to ruminal degradation (Hristov et al., 1998), and thus may offer potential for improving diet digestibility and animal performance. Klingerman et al. (2009) reported that exogenous amylase addition to a normal-starch diet (26% of DM) increased milk yield by dairy cows; positive in vitro and in vivo digestibility responses to exogenous amylase were also observed. In the trial of Gencoglu et al. (2010), 36 multiparous Holstein cows (51 ± 22 DIM at trial initiation) were randomly assigned to 1 of 3 treatments in a completely randomized design; a 3-wk covariate adjustment period with cows fed the normal-starch diet followed by a 12-wk treatment period with cows fed their assigned treatment diets. The normal-starch TMR did not contain exogenous amylase (NS-). The reduced-starch diets, formulated by partially replacing corn grain with soy hulls, were fed without (RS-) and with (RS+) exogenous amylase addition to the TMR. Starch concentrations averaged 27.1%, 21.8% and 20.7% (DM basis) for the NS-, RS- and RS+ diets, respectively. Dry matter intake for cows fed RS- was 2.4 and 3.2 kg/d greater than for cows fed NS- and RS+, respectively. Milk yield averaged 50.4 kg/d and was unaffected by treatment. Fat-corrected milk yield was 2.9 kg/d greater for cows fed RS- than for cows fed NS-. Body weight and condition score measurements were unaffected by treatment. Fat-, solids-, and energy-corrected milk feed conversions (kg / kg DMI) were 12% to 13% greater for cows fed RS+ than for cows fed RS-. Dry matter and nutrient digestibilities were lowest for cows fed NS- and greatest for cows fed RS+, and were greater for cows fed RS+ than for cows fed RS- with the exception of starch digestibility which was similar. Greater conversion of feed to milk for dairy cows fed reduced-starch diets with inclusion of exogenous amylase may offer potential for improving economic performance depending on diet and additive costs. Additional studies have recently been conducted at The Ohio State University (Weiss pers. comm., OARDC, Wooster, OH) and UW-Madison (Ferraretto and Shaver) on the effects of reduced-starch diets and exogenous amylase addition to reduced-starch diets with formulation of the reduced-starch diets by partially replacing dry shelled corn with corn silage or a whole cottonseed/wheat middlings mixture, respectively. Conclusions Lactation performance was reduced for 18% and 20% starch diets (DM basis) formulated using beet pulp and citrus pulp, respectively, to partially replace corn grain. However, lactation performance was not reduced for diets as low as 16% to 17% starch (DM basis) formulated using soy hulls to partially replace corn grain. Diets containing 21% starch (DM basis) were acceptable when high-fiber, moderate protein byproduct feeds were used to partially replace corn grain and protein supplement. The starch in lowstarch diets should be highly digestible. Various factors, including type of endosperm, particle size, maturity and moisture content, and conservation and processing methods, influence the digestibility of starch from corn grain and corn silage by dairy cows. The inclusion of exogenous enzymes with amylolytic activity in reduced-starch diets for dairy cows may play a role in the future. References Batajoo, K. K., and R. D. Shaver Impact of nonfiber carbohydrate on intake, digestion, and milk production by dairy cows. J. Dairy Sci. 77:1580:1588. Broderick, G. A., D. R. Mertens, and R. Simons Efficacy of carbohydrate sources for milk production by cows fed diets based on alfalfa silage. J. Dairy Sci. 85: Bucholtz, H Feeding practices of high-producing herds; What can we learn? Proc. Western Canadian Dairy Seminar. Red Deer, Alberta, Canada. 18: Correa, C. E. S., R. D. Shaver, M. N. Pereira, J. G. Lauer, and K. Kohn Relationship between corn vitreousness and ruminal in situ starch degradability. J. Dairy Sci. 85: Firkins, J. L., M. L. Eastridge, N. R. St-Pierre, and S. M. Noftsger Effects of grain variability and processing on starch utilization by lactating dairy cattle. J. Anim. Sci. 79(E. Suppl.): E218-E238. Gencoglu, H., R. D. Shaver, W. Steinberg, J. Ensink, L. F. Ferraretto, S. J. Bertics, J. C. Lopes, and M. S. Akins Effect of feeding a reduced-starch diet with or without amylase addition on lactation performance by dairy cows. J. Dairy Sci. 93: Hamaker, B. R., A. A. Mohamed, J. E. Habben, C. P. Huang, and B. A. Larkins, Efficient procedure for extracting maize and sorghum kernel proteins reveals higher prolamin contents than the conventional method. Cereal Chem. 72(6):
96 Hristov, A. N., T. A. McAllister, and K. J. Cheng Stability of exogenous polysaccharide-degrading enzymes in the rumen. Anim. Feed Sci. Technol. 76: Ipharraguerre, I. R., R. R. Ipharraguerre, and J. H. Clark Performance of lactating dairy cows fed varying amounts of soyhulls as a replacement for corn grain. J. Dairy Sci. 85: Johnson, L., J. H. Harrison, C. Hunt, K. Shinners, C. G. Doggett, and D. Sapienza Nutritive value of corn silage as affected by maturity and mechanical processing: A contemporary review. J. Dairy Sci. 82: Kaiser, R., and R. Shaver Benchmarking high producing herds. Proc. Western Canadian Dairy Seminar. Red Deer, Alberta, Canada. 18: Klingerman, C. M., W. Hu, E. E. McDonell, M. C. DerBedrosian, and L. Kung, Jr An evaluation of exogenous enzymes with amylolytic activity for dairy cows. J. Dairy Sci. 92: Larson, J., and P. C. Hoffman Technical Note: A method to quantify prolamin proteins in corn that are negatively related to starch digestibility in ruminants. J. Dairy Sci. 91: Lopes, J. C., R. D. Shaver, P. C. Hoffman, M. S. Akins, S. J. Bertics, H. Gencoglu, and J. G. Coors Type of corn endosperm influences nutrient digestibility in lactating dairy cows. J. Dairy Sci. 92: McAllistar, T. A., R. C. Phillippe, L. M. Rode, and K. J. Cheng Effect of the protein matrix on the digestion of cereal grains by ruminal microorganisms. J. Anim. Sci. 71: Nocek, J. E., and S. Tamminga Site of digestion of starch in the gastrointestinal tract of dairy cows and its effects on milk yield and composition. J. Dairy Sci. 74: Ngonyamo-Majee, D., R. D. Shaver, J. G. Coors, D. Sapienza, C. E. S. Correa, J. G. Lauer and P. Berzaghi. 2008a. Relationships between kernel vitreousness and dry matter degradability for diverse corn germplasm. I. Development of near-infrared refelctance spectroscopy calibrations. Anim. Feed Sci. Technol. 142: Ngonyamo-Majee, D., R. D. Shaver, J. G. Coors, D. Sapienza and J. G. Lauer. 2008b. Relationships between kernel vitreousness and dry matter degradability for diverse corn germplasm. II. Ruminal and post-ruminal degradabilities. Anim. Feed Sci. Technol. 142: Rananthunga, S. D., K. F. Kalscheur, A. R. Hippen, and D. J. Schingoethe Replacement of starch from corn with nonforage fiber from distillers grains and soyhulls in diets of lactating dairy cows. J. Dairy Sci. 93: Staples, C. R Feeding dairy cows when corn prices are high. Proc. 44 th Florida Dairy Production Conference. Gainesville, FL. Stone, W. C Applied topics in dairy cattle nutrition: Soy hulls as either forage or concentrate replacement. Ph.D. Thesis. Cornell Univ., Ithaca, NY. Taylor, C. C., and M. S. Allen Corn grain endosperm type and brown midrib 3 corn silage: Site of digestion and ruminal digestion kinetics in lactating cows. J. Dairy Sci. 88: Voelker, J. A., and M. S. Allen Pelleted beet pulp substituted for high-moisture corn: 1. Effects on feed intake, chewing behavior, and milk production of lactating dairy cows. J. Dairy Sci. 86: Weiss, W. P., N. R. St-Pierre, and L. B. Willett Varying type of forage, concentration of metabolizable protein, and source of carbohydrate affects nutrient digestibility and production by dairy cows. J. Dairy Sci. 92:
97 FARM Dairy Well-Being Program Most consumers today have no direct relationship to livestock producers. Many have a close relationship with their pets and love animals so they want to know how animals are cared for on the farm. There is a need to reassure them that producers care for their animals. There have been various recent examples of undercover footage of dairy animal abuse appearing in the news, such as ABC Nightline, and videos, e.g., one from PETA depicting poor welfare conditions at a Pennsylvania dairy farm. These are extreme cases, and definitely not the norm. Consumers can get influenced by this negative propaganda about the dairy industry. The industry is taking a proactive approach in relation to dairy welfare and in October 2009, the National Milk Producers Federation and Dairy Management Inc. announced their new National Dairy FARM (Farmers Assuring Responsible Management) Dairy Well-Being Program. Goals of this program include demonstrating to retailers and consumers that dairy producers are using best practices in animal care and providing uniformity and consistency for these practices across the U.S. Uniformity is one of the advantages of a national program. The program includes three stages: 1) education and self-assessment, 2) evaluation, and 3) third-party verification. Educational workshops will be offered in Minnesota later this year with the goal of informing dairy producers, nutritionists, veterinarians, and other dairy professionals about the FARM program and some of the areas that will be evaluated on the farm. The workshops will help producers and their advisors conduct a selfassessment of their dairy. Educational information will also be available at the FARM website. For the second stage, each farm will be evaluated once every three years by a second party evaluator, who could be the herd veterinarian or milk coop field personnel or other professionals familiar with the dairy. A statistical sample of all dairy farms on the program will be selected every year, starting in late 2011 or 2012, to have third-party verification by a representative who is not involved with the dairy. All the information collected from each farm remains confidential and only overall summaries will be available publicly. Marcia I. Endres University of Minnesota Areas assessed in the FARM program include standard operating procedures, training and record keeping, calf care, animal health, nutrition, environment and facilities, handling, movement and transportation, special needs animals, and dairy beef. The recommended best practices or guidelines are summarized in the Quick Reference User Guide, and more detail is provided in the animal care manual, both available at the FARM website: Most producers are already addressing the areas to be evaluated in the program. For example, one of the checklist items is calves receive colostrum or colostrum replacement soon after birth. This is already being done because all producers want healthy, productive calves. Another item is the dairy has a Veterinarian/Client/Patient relationship. I don t know of many dairies in Minnesota not working with a veterinarian and not having a health plan for their animals. Rations should provide the required nutrients for maintenance, growth, and lactation. Again, that is good for the cow and for the business. Freestall bedding is refreshed (remove soiled sand or other bedding material) and fresh bedding is added on a routine basis. Hopefully that is also standard protocol in all our dairies. Adequate lighting is in place to allow inspection of animals and to provide safe working conditions. With very few exceptions, all dairy barns and parlors are well lit. The list goes on. When producers do the self-assessment or have the evaluation by the second party, there might be some practices that are not being done on the dairy, maybe some standard protocols need to be developed, or better care provided to special needs animals, or maybe more training for the workers on animal handling. This provides an opportunity to further improve animal care. Some of the numbers suggested in the guide for outcome-based measurements (such as locomotion, hock lesion and hygiene scores) are only guidelines and not standards. For example, research has shown that the prevalence of lameness in freestall Minnesota and Wisconsin dairy herds averaged approximately 25% (Espejo et al., 2006; Cook et al., 2003) not 10% as it is recommended in the FARM guide. The same goes for hock lesions, that averaged for various studies we did in the Midwest at 67% for freestalls with mattresses, 47% for freestalls with deep bedded manure solids, 30% for freestalls with sand, and 11% for compost bedded pack barns. The FARM 94
98 recommendation is less than 10%. Producers don t have to fret about these suggested numbers for locomotion and hock lesion prevalence. They are guidelines that might be re-evaluated in a few years, and maybe long term goals, not requirements. FARM is not an audit program or certification program or a marketing tool. It is an evaluation opportunity and an organized and consistent approach to document across the nation that producers provide good care for their animals, so consumers and retailers can have their reassurance when purchasing dairy products. References: Cook, N. B Prevalence of lameness among dairy cattle in Wisconsin as a function of housing type and stall surface. J. Am. Vet. Med. Assoc. 223: Espejo, L.A., M.I. Endres, and J. A. Salfer Prevalence of lameness in high-producing Holstein cows housed in freestall barns in Minnesota. J. Dairy Sci. 89: If we all work together to evaluate and improve animal care everyone wins the cow, the producer, and the consumer (who wants to be assured good care is the standard on farms). It is not easy to make a national program work, but we recommend that producers participate on the FARM program, so it will work and be of benefit to the dairy industry. 95
99 Dairy calf liquid feeding strategies to cope with new antibiotic use regulations Take Home Messages It is unclear how new milk replacer antibiotic regulations will affect calf growth efficiency and health. Numerous bio-active feed additives are available that may improve calf gut health and result in improvements in efficiency of growth and calf health. Future calf research should focus on determining which combination of these bio-active feed additives offer the best health and growth benefits for milk replacer fed dairy calves Introduction Despite many years of research and improvements in calf nutrition too many producers struggle with calf health. According to the 2007 USDA-NAHMS survey, mortality rates in unweaned heifers were 7.8 percent. Of this nearly eight percent mortalities, scours (56.5%) and respiratory illness (22.5%) were the most frequently cited causes. Implementing new strategies that show benefits in human and other livestock neonates may be useful in developing nutritional strategies to improve dairy calf health. The aim of this proceedings paper is to describe new dietary modifications that may improve calf performance and health. During the nursery phase, dairy calves are susceptible to many pathogens. Traditionally antibiotics were added to milk replacer to lower calf mortality, reduce the incidence of scours, and improve feed efficiency. Calf feeders and nutritionists have expressed concern over new FDA regulations changing the continuous use of a 2:1 ratio of neomycin sulfate:oxytetracycline (NT) medicated milk replacers. Previous NT approved use in medicated milk replacers was for concentrations of g/ton neomycin and g/ton oxytetrocycline. Within this system of antibiotic use, calves were fed the 2:1 ration of NT therapy throughout the milk replacer feeding period for the treatment of bacterial enteritis (scours) on a continuous basis. As of April 1, 2010, 2:1 NT milk replacers were no longer allowed to be manufactured. Noah B. Litherland University of Minnesota Department of Animal Science St. Paul, MN 96 Under the newly approved regulation, there will be two weight based options for the medication of milk replacer feeding programs. The first option is continuous feeding of a dose of 1:1 ratio of NT at a feeding rate of 0.05 to 0.10 mg /lb of bodyweight. The lower dose concentration will be 8-16 g/ton of each antibiotic versus the previously approved g/ton neomycin and g/ton oxytetracycline. The FDA indication for this low rate of feeding protocol is labeled for increased rate of weight gain and improved feed efficiency. The second option under the new milk replacer medication standards permits manufactures to market products for the treatment of clinical bacterial enteritis (scours) caused by E. Coli and/or bacterial pneumonia caused by P. multocida. Administered on the basis of calf weight, this treatment will deliver five to ten times more antibiotic therapy than the previous 2:1 NT program. This higher concentration antibiotic program is labeled to be fed continuously for 7 to 14 days. During this therapeutic treatment period, medication will be added to the nonmedicated milk replacer so that 1:1 NT treatment (10 mg/lb of body weight) will be consumed daily. Assuming a 100 lb calf consuming 1 to 1.25 lb of milk replacer powder daily, this equates to 16 to 20 g of NT per ton of milk replacer. Producers will need to either handle two separate milk replacers (low and high antibiotic) or use an add-pack to add NT 1:1 for calves needing treatment. It will be increasingly important for producers, nutritionist and veterinarians to work together to ensure that calves are being treated correctly. Impact of new antibiotic regulations on animal performance Antibiotics in milk replacers were clearly effective in improving performance and health of calves (Morrill et al., 1977; Quigley et al., 1997; Heinrichs et al., 2003; Berge et al., 2005). Situations with poor sanitation or where environmental stressors and poor nutrition place extra burdens on calves likely benefited the most from medicated milk replacer programs (Berge et al., 2005). Concerns from producers and nutritionists have been expressed regarding lag time between identification of clinical symptoms, addition of NT to milk replacer, and effective treatment of bacterial scours. The new regulations will have the least impact on dairy producers that place a premium

100 on sanitation, bio-security, bedding quality, and feed a quality nutrition program. Calves may be more susceptible to disease during different seasons of the year depending on air quality and thermal stress abatement strategies. One important take home message is that antibiotics do not offer protection from rotavirus, coronavirus, coccidia or cryptosporidia which are the most prevalent causes of dairy calf scours. These viruses and parasites are not bacteria and therefore are not susceptible to antibiotics. Results from bacterial scours cases submitted to the University of Minnesota Veterinary Diagnostic Laboratory are summarized in Table 1. Results from these clinical cases indicate that 74% (450/606) tested positive for at least one pathogen. The most common pathogen cultured was cryptosporidium followed by rotavirus. Calf feeding and management strategies should be designed to reduce risk factors associated with all major pathogens. Table 1. University of Minnesota Veterinary Diagnostic lab 2006 case summary for fecal cultures from dairy calves with scours (Source: Dr. Jeremy Schefers). Feed additives and feeding strategies to improve efficiency of growth and health Many feed additives for milk replacer and calf starter as well as management strategies have been employed to improve calf performance and health. It is plausible that combinations of feed additives may provide an additive response to promote calf health and growth. There is a need to identify affordable and effective combinations of these compounds and determine their effectiveness within a diverse nutrition and management scenarios. Table 2 contains a partial description of some commercially available milk replacer additives, the pathogens they may have an effect on, purported benefits, and some references using these products in controlled research studies. Drackley (2008) suggested that the effectiveness of additives may be enhanced if tested in calves fed more biologically normal amounts of milk or milk replacer. Applications of feed additive technology for milk fed dairy calves will likely become more important as group feeding using automated calf feeders becomes more common. Automated calf feeders have the capability of detecting early signs of poor calf health and may increase the sensitivity of detecting calf-hood disease. Additionally automated calf feeding systems can be equipped with dosing equipment to deliver specific treatments to calves on an individual basis. This option may become a powerful tool to individually treat calves and prevent reduce the impact of disease outbreaks. Controlling enteric disease challenges in group housed calves will likely continue to become an important consideration for calf welfare. Altering the fatty acid composition of milk replacer Milk fat in milk replacer typically comes from lard, tallow, or white grease. While more affordable, the fatty acid profile of these sources of fat and milk fat have some dissimilarities. Milk replacer is higher in C16:0, C18:0, and C18:1 and lower in C4:0, C18:2, and C18:3. Several of the fatty acids that are in lower quantities in milk replacer compared to milk fat have properties related to health, antimicrobial properties, and growth. Butyric acid (C4:0) or butyrate is an important short chain fatty acid (SCFA) for development of the rumen and health and development of the lower gut. Butyrate is a naturally occurring organic acid that is present in cow s milk at a concentration of g/l and is normally released from milk triglycerides in the abomasum as a result of the action of preintestinal lipases (Bugaut, 1987). Muscato et al. (2002) found that newborn dairy calves given daily doses of autocalved rumen fluid gained more weight and had a reduced incidence of scours compared with controls. Butyrate, along with many other components, might have been responsible for these benefits. 97
101 SCFA s are produced by microbes in the rumen and in the large intestine and appear to have a role in growth, digestibility, feed efficiency, cell proliferation and differentiation, and function of the GI tract. The presence of SCFA in the intestinal lumen is important for; structural and functional development (including tight junction assembly and maintenance), nutrient absorption, and osmolarity regulation. In the lower gut, butyrate may help in modifying immune and inflammatory responses and influence intestinal micro-flora. Human research indicates that butyrate enhances the intestinal barrier by facilitating tight junction assembly (Peng et al., 2009). Hill et al. (2007) supplemented 1% sodium butyrate (4.4 g/calf/d) in milk replacer and observed a significant increase in body weight gain and a reduction in abnormal fecal scores. Researchers in France recently fed 0.3% of the diet as crystalline sodium-butyrate and observed improvements in starter intake, increased reticuorumen development, and improved fecal consistency (Gorka et al., 2009). When compared with flavomycin (antibiotic) calves supplemented with 3 g of sodium-butyrate weighed more at 145 days and had lower feed to gain from day 60 to 124 and day 13 to 145 (Guilloteau et al., 2009). Supplementing milk replacer with long-chain fatty acids is an option to deliver these fatty acids in similar proportions to that contained in milk fat. Determining the source, fatty acid profile, and amount of these supplemental fatty acids may be challenging. Work conducted to evaluate calf performance when vegetable oils are included in milk replacer has demonstrated variable calf response to fatty acid composition of supplemental fat. Graulet et al. (2000) reported similar gains for calved fed tallow or coconut oil. Jenkins et al. (1985) observed equivalent performance for tallow and coconut oils but markedly poorer results for corn oil. Huuskonen et al. (2005) demonstrated similar rates of gain for calves fed tallow or a combination of palm, coconut, and rapeseed oil compared with tallow. Canola and rapeseed oil contain considerable amount of linolenic acid (C18:3) Jenkins and Kramer, (1986) indicated that supplementing linoleic and linolenic acid, did not alter calf performance, however, the authors concluded that these fatty acids may play an important role in stressed calves. Similarly, Hill et al (2007) determined that formulating the calf milk replacer to increase the concentration of SCFA (C4:0), medium-chain (C8:0, C10, C12:0, and C14:0) and essential fatty acids (C18:3) above what is found in milk replacer containing animal fat reduced the number of days with abnormal fecal scores and improved ADG in calves. More recently, Hill et al. (2009) determined that supplementing C18:3 (linoleic acid) as a calcium salt of flax oil to starter in claves less than 3 months old resulted in increased ADG and feed efficiency. Recent advances in the area of functional foods and neutraceuticals have resulted in the generation of a number of human food products. Perhaps these fatty acids have a place in milk replacer. Lactoferrin Lactoferrin is a bioactive glycoprotein iron binding compound normally present in colostrum and milk (Masson and Heremans, 1971). Bovine colostrum contains about 2 mg/ml lactoferrin and milk contains a much lower concentration of 20 to 200 Ìg/ml (Masson and Heremans, 1971). Lactoferrin has bacteriostatic activities against E. Coli (Teraguchi et al., 1994) and rotavirus (Superti et al., 1997), which may help reduce colonization in the GI tract of calves. Since lactoferrin is present in the mammary gland as a potential antimicrobial protein, Robblee et al. (2003) hypothesized that it may serve a dual role in protecting the mammary gland and the gut of the neonatal calf. Roblee et al. (2003) noted some improvements in average daily gains, fecal scores, and reduced morbidity in preweaned calves with supplementation of 3 g/d lactoferrin. Adding lactoferrin (1 g/d) to milk replacer did not alter fecal scores or improvement performance in conventional or accelerated milk replacer programs (Cowles et al., 2006). Prebiotics Prebiotics are defined as a nondigestible food ingredient that beneficially affects the host by selectively stimulating the growth and/or activity of one or a limited number of bacteria. Gibson and Roberfroid, Published research (Heinrichs et al., 2003; Quigley et al., 2002; Donovan, 2002) have shown an inconsistent impact of prebiotics on health and performance of calves. The impact of prebiotics on immune function has not yet been clearly established. Effects of these products might be best positioned after treatment of calves following antibiotic treatment for 7 to 14 days with 10 mg/lb of body weight dose of 1:1 NT. These prebiotics may provide an opportunity for re-growth of normal intestinal flora following antibiotic treatment. Probiotics Probiotic is a general term defined as a live microbial feed supplement which beneficially affects the host animal by improving its intestinal balance. According to Yoon and Stern (1995), this term refers to microbial cultures, enzyme preparations and culture extracts. Most probiotic research has been conducted using direct fed microbials (DFM) which are a source of live naturally occurring microorganisms (Krehbiel et al., 2003). The mode of action of DFM in dairy calves is not clear; however, performance and health benefits are most likely 98
102 observed when DFM are fed to stressed calves (Lesmeister et al., 2004; Seymour et al., 1995; Galvao et al., 2005; Magalhãs et al., 2008). Probiotics likely change the micro-ecology of the gut through competitive inhibition. Research evaluating the impact of DFM on immune function in calves is limited. Timmerman et al. (2005) demonstrated that stressed calves had higher average daily gain and feed efficiency. Essential Oils Essential oils are blends of secondary metabolites obtained from the plant volatile fraction by steam distillation. (Calsmaiglia, 2007). The main mode of action of essential oils is as antimicrobials with their mode of action through effects on the cell membranes. Essential oils with antimicrobial activity include garlic, dill, paprika, cassia, juniper, tea tree, oregani, anise, rosemary, clove, thyme and ginger. Alicin, a component of garlic, is of particular interest as it inhibits bacterial growth by binding to the enzyme alcohol dehydrogenase and also had antioxidant effects. The majority of research conducted with essential oils has been conducted in vitro with limited published research available in calves. Published research (Hill et al., 2007, Bampidis et al., 2006 and Olson et al., 1998) does not strongly support the use of essential oils as alternatives to antibiotics, but does show some interesting potential benefits. Fructooligosaccharides (FOS) FOS belongs to a family of oligosaccharides consisting of several -(1-2) or -(1-6) linked fructose residues. FOS acts as a selective substrate that may stimulate the growth of bifidobacteria resulting in competition for pathogens. FOS may modify the environment of the gastrointestinal tract by increasing fermentable fiber, resulting in production of short-chain fatty acids. May et al. (2004) demonstrated that FOS prevented gut colonization of Clostridium difficile. Additionally, Webb et al. (1992) measured greater weight gains by adding FOS to milk replacer. Challenges with interpreting health data Interpreting the effects of dietary supplements on calf health can be difficult. Quigley (2002) the attributes of well designed experiments evaluating alternatives to antibiotics including; negative and positive controls, sufficient number of animals, an indication of passive immune status, an indication of the immunological challenge on the animal during the test, measure of environmental effects, measurements that can detect treatment differences, a complete description of methods, and proper interpretation of the data. Designing, conducting, and interpreting experiments with alterations in health as the outcome of interest are challenging. Most university research herds are inadequately sized to achieve an appropriate statistical power to detect differences in treatments for most health variables. Summary Significant changes in FDA regulations regarding the use of antibiotics in milk replacer offers an opportunity to reexamine milk fed calf nutrition. Our goal should be to continue to discover feeding and management strategies to optimize the efficiency of calf growth while continuing to improve health and welfare. Discovering new feed additives or feed additive combinations will offer more tools to accomplish this goal. Research should continue to test additives in university trials to understand mode of action and also in on-farm studies to determine animal response with greater calf numbers and subsequent economic impact. References Bampidis, V. A., V. Christodoulou, P. Florou-Paneri, and E. Christaki Effect of dried oregano leaves versus neomycin in treating newborn calves with colibacillosis. J. Vet Med A Physiol Clin Med. 53: Berge, A. C. B., T. E. Besser, D. A. Moore, and W. M. Sischo Evaluation of the effects of oral colostrum supplementation during the first fourteen days on health and performance of preweaned calves. J. Dairy Sci 92: Berge, A. C., P. Lindeque, D. A. Moore, and W. M. Sischo A clinical trial evaluating prophylactic and therapeutic antibiotic use on health and performance of preweaned calves. J. Dairy Sci. 88: Bielmann, V., T. J. DeVries, S. J. LeBlanc, K. Lissemore, and K. E. Leslie Evaluation of supplemental dried bovine colostrum in milk replacer fed dairy calves. J. Dairy Sci. Vol 92, E-Suppl Bugaut, M Occurrence, absorption and metabomism of short chain fatty acids in the digestive tract of mammals. Comp. Biochem Physio B. 86: Calsmaiglia, S., M. Busquet, P. W. Cardozo, L. Castillejos, A. Ferret Invited review: Essential oils as modifiers of ruminal microbial fermentation. J. Dairy Sci. 90: Cowles, K. E., R. A. White, N. L. Whitehouse, and P. S. Erickson Growth characteristics of calves fed an intensified milk replacer regimen with additional lactoferrin. J. Dairy Sci. 89: Donovan, D. C., S. T. Franklin, C. C. L. Chase, and A. R. Hippen Growth and health of Holstein calves fed milk replacers supplemented with antibiotic or enteroguard J. Dairy Sci. 85: Drackley, J. K Calf nutrition from birth to breeding. Vet. Clin. Food Anim. 24: Galvão, N. K., J. E. P. Santos, A. Coscioini, M. Villaseñor, W. M. Sischo, A. C. B. Berge Effect of feeding live yeast products to calves with failure of passive transger on performance and patterns of antibiotic resistance in fecal Escherichia coli Reprod. Nutr. Dev. 45:
103 Gorka, P. Z. M. Kowalski, P. Pietrazak, A. Kotunia, R. Kiljanczyk, J. Flaga, J. J. Holst, P. Guilloteau, and R. Zabielski Journal of Physiology and Pharmacology. 60: Graulet, B., D. Gruffat-Mounty, D. Durand, and D. Bauchart Effects of milk diets containing beef tallow or coconut oil on the fatty acid metabolism of liver slices from preruminant claves. Br. J. Nutr. 84: Guilloteau, P., R. Zabielski, J. C. David, J. W. Blum, J. A. Morisset, M. Biernat, J. Wolinski, D. Laubitz, and Y. Hamon Sodium-butyrate as a growth promoter in milk replacer formula for young calves. J. Dairy Sci. 92:1038- Heinrichs, A. J., C. M. Jones, B. S. Heinrichs Effects of mannan oligosaccharides or antibiotics in neonatal diets on health and growth of dairy calves. J. Dairy Sci. 86: Hill, T. M., H. G. Bataeman III, J. M. Aldrich, and R. L. Schlotterbeck Effects of changing the essential and functional fatty acid intake of dairy calves. J. Dairy Sci 92: Hill, T. M., J. M. Aldrich, R. L. Schlotterbeck, and H. G. Bateman, II Effects of changing the fat and fatty acid composition of milk replacers fed to neonatal calves. The Professional Animal Scientist. 23: Huuskonen, A., H. Khalili, J. Kiljala, E. Joki-Tokola, and J. Nousiainen Effects of vegetable fats versus lard in milk replacers on feed intake, digestibility, and growth in Finnish Ayrshire bulls calves. J. Dairy Sci. 88: Jenkins, K. J Factors affecting poor performance and scours in preruminant claves fed corn oil. J. Dairy Sci. 71: Jenkins, K. J. and J. K. G. Kramer Influence of low linoleic and linolenic acids in milk replacer on calf performance and lipid in blood plasma, heart, and liver. J. Dairy Sci. 69: Jenkins, K. J., J. K. G. Kramer, and D. B. Emmons Effect of lipids in milk replacer on calf performance and lipids in blood plasma, liver, and perirenal fat. J. Dairy Sci. 69: Jenkins, K. J., J. K. G. Kramer, F. D. Sauer, and D. B. Emmons Influences of triglycerides and free fatty acids in milk replacers on calf performance, blood plasma, and adipose lipids. J. Dairy Sci. 68: Jenny, B. F., H. J. Vandlijk, and J. A. Collins Performance and fecal flora of calves fed a Bacillus subtilis concentrate. J. Dairy Sci. 74: Krehbiel, C. R. S. R. Rust, G. Zhang and S. E. Gilliland Bacterial direct-fed microbials in ruminant diets: Performance response and mode of action. J. Anim. Sci. 81:E Lesmeister, K. E., A. J. Heinrichs, M. T. Gabler Effects of supplemental yeast (Saccharomyces cerevisiae) culture on rumen development, growth characteristics, and blood parameters in neonatal dairy calves. J. Dairy Sci. 87: Mason, P. L., and J. F. Heremans Lactoferrin in milk from different species. Comp. Biochem. Physop. 39B: Magãlhas, V.J., F. Susca, F.S. Lima, A. F. Branco, I. Yoon, and J. E. Santos Effect of feeding yeast culture on performance, health, and immunocompetence of dairy calves. J. Dairy Sci. 91: May, T., R. I. Mackie, G. C. Fahey, J. C. Cremin, and K. A. Garleb Effect of fiber source on short-chain fatty acid production on growth and toxin production by Cl. Difficile. Scand. J. Gastroentestinol. 29: Morrill, J L., A. D. Dayton, R. Mickelson Cultured milk and antibiotics for young calves. J Dairy Sci. 60: Muscato, T. V., L. O., Tedeschi, and J. B. Russell The effect of ruminal fluid preparations on the growth and health of newborn, milk-fed dairy calves. Olson, E. J., W. B. Epperson. D. H. Zeman Effects of an allicin-based product on cryptosporidiosis in neonatal calves. J. Am Vet Med Assoc. 212: Quigley, J. D., T. A. Wolfe, and T. H. Elsasser Effects of additional milk replacer feeding on calf health, growth, and selected blood metabolites in calves. J. Dairy Sci. 89: Quigley, J Calf note #89 Evaluating the use of antibiotic alternatives. ( Pages 1-5. Quigley, J. D. III and M. D. Drew Effects of oral antibiotics or bovine plasma on survival, health and growth in dairy calves challenged with Escherichia coli. Food and Agricultural Immunology. 12: Quigley, J. D. III, J. J. Drewery, L. M. Murray, and S. J. Ivey Body weight gain, feed efficiency, and fecal scores of dairy calves in response to galactosyl-lactose or antibiotics in milk replacer. J. Dairy Sci. 80: Peng, L. Z. R. Li, R. S. Green, I. R. Holzman, and J. Ling Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 139: Robblee, E. D., P. S. Erickson, N. L. Whitehouse, A. M. McLaughlin, C. G. Schwab, J. J. Rejman, and R. E. Rompala Supplemental lactoferrin improves health and growth of Holstein calves during the preweaning phase. J. Dairy Sci 86: Seymour, W. M., J. E. Nocek, J. Siciliano-Jones Effects of a colostrum substitute and of dietary brewer s yeast on the health and performance of dairy calves. J. Dairy Sci. 78: Superti, F., M. G. Ammendolia, P. Valenti, and L. Seganti Antirotaviral activity of milk proteins: lactoferrin prevents rotavirus infection in the enterocyte-like cell line HT-29. Med. Microbiol. Immunol. 186: Teraguchi, S., K. Ozawa, S. Yasuda, K. Shin, Y. Fukuwatari, and S. Shimamura The bacteriostatic effects of orally administered bovine lactoferrin on intestinal Enterobcteriacae of SPF mice fed bovine milk. Biosci. Biotech. Biochem. 58: Timmerman, H. M., L. Mulder, H. Everts, D. C. van Espen, E. van der Wal, G. Klaassen, S. M. G. Rouwers, R. Hartemink, F. M. Rombouts, and A. C. Beynen
104 Health and growth of veal calves fed milk replacer with or without probiotics. J. Dairy Sci. 88: Webb, P. R., D. W. Kellogg,M. W. McGahee, and Z. B. Johnson Addition of fructooligosaccharides (FOS) and sodium diacetate (SD) plus decoquinated (D) to milk replacer and starter grain fed to Holstein calves. J. Dairy Sci. 75:293. (Abstr.). Weinberg, E. D Human lactoferrin: a novel therapeutic with broad spectrum potential. J. Pharm. Pharmacol. 53: Yoon, I. K. and M. D. Stern. Effects of Saccharomyces cerevisiae and Aspergillus oryzae cultures on ruminal fermentation in dairy cows J. dairy Sci. 79:
105 Improving the Value of Cull Cows Richard L. Wallace, DVM, MS Dairy Extension Veterinarian University of Illinois Take Home Messages Culling and transporting decisions are an important part of dairy farming. Occasionally, an animal that is ambulatory on the farm may not be suitable for transport to a packing or processing facility. Consider the Top 10 Considerations for Culling and Transporting Dairy Animals to a Packing or Processing Facility to make appropriate decisions on the suitability of an animal to be shipped. All production animals eventually become market animals. Within the food production system, dairy producers wear several hats. The largest and most obvious hat is the production of raw milk shipped from the dairy operation. The quality of the raw milk produced will not improve once it has been harvested from the cows. Every effort should be made to present the highest quality, most wholesome product to the milk processor. A less obvious hat that dairy producers wear is the generation of calves and cows for the beef market. Cull cows and bull calves can represent between percent of the gross farm income. With proper management and timely marketing, the value of market cows and bull calves can be increased. This paper will focus on the considerations dairy producers must make when culling and transporting cattle from their operations. The National Milk Producers Federation has published a document outlining the Top 10 Considerations for Culling and Transporting Dairy Animals to a Packing or Processing Facility. Leaders in the dairy industry have been working on a dairy beef quality assurance program for years. The Humane Society of the United States release of the Heartland Packing Plant video showing inhumane treatment of dairy cattle propagated the release of this document. 1. Do not move non-ambulatory animals to market under any circumstances. (Only allow ambulatory animals to be shipped to market). 2. Make the decision to treat, to cull, or to euthanize animals promptly. Sick and injured animals should be segregated from the herd. 3. Delay transport of an animal that appears to be exhausted or dehydrated until the animal is rested and re-hydrated. 4. Milk all cows that are still lactating just prior to transporting to a packing or processing facility. 5. Use a transportation company that is knowledgeable about your animal care expectations and provides for the safety and comfort of the animals during transport. 6. Do not transport animals to a packing or processing facility until all proper treatment withdrawal times have been followed. 7. Do not transport animals with a poor body condition, generally a Body Condition Score of less than 2 (1-5 scale). 8. Do not transport animals that require mechanical assistance to rise and are reluctant or unable to walk, except for veterinary treatment. When using any handling device, abuse must not be tolerated. 9. Do not transport animals with bone fractures of the limbs or injuries to the spine. Animals with a recent fracture unrelated to mobility should be culled and transported directly to a packing or processing facility. 10. Do not transport animals with conditions that will not pass pre-slaughter inspection at a packing or processing facility. If unsure, consult with your veterinarian before transporting an animal to a packing or processing facility. The dairy industry needs to shift our thought processes regarding the classification of surplus animals. There are actually two classes of animals on dairy farms, production animals and market animals. Production animals would include lactating cows, dry cows, replacement heifers and potentially bulls for breeding. Market animals would include cull cows and bull calves. All production animals eventually become market animals! To that end, the dairy industry should discontinue using terms like cull, spent, salvage, junk and surplus, and begin using the term market when referring to animals that are no longer economically productive. Market cattle, non-fed beef and dairy animals supply 20 percent of the total beef produced in the United States. In 1998, 2.5 billion pounds of market cow beef was produced in the U.S. with nearly half of that amount from dairy cows. Almost three quarters of the market cow beef is destined for processed beef products. Most dairy producers assume that the major product from market cows is ground beef sold as hamburgers through fast-food restaurants. In reality, ground beef is a very important product from market cattle, but it is only one of many products. Depending upon the operation, market cow packers 102
106 utilize tenderloins, ribeyes and strip loins, particularly from younger cows as well as the hide and many other inedible products. The decision to market a cow is a complex one. When making a marketing decision, dairy producers may consider many cow factors, such as age, stage of lactation, milk production, health status, disposition, and reproductive performance. Other economic factors such as current milk price, market cow price, as well as cost and availability of replacement heifers may have a role in determining whether or not to market a cow. Dairy cow marketing decisions have an important influence on the financial success of the dairy. Marketing decisions can function as a component of genetic improvement programs designed for longterm gain and improved production efficiency (voluntary marketing). At the same time, marketing may also represent failure or limited success of health programs resulting in cows leaving the herd prematurely due to death, disease or health-related problems (involuntary marketing). Annual market rates for DHIA Holstein herd herds in Illinois and the Midwest as of September 2008 are shown below. The data is averaged for the bottom, middle and top third of herds based on rolling herd average milk production. Location/RHA < 18,000 18,001-22,000 > 22,001 Illinois 33.4 (126) 36.4 (172) 36.1 (107) Midwest 34.3 (1583) 35.2 (2508) 36.3 (2271) Marketing decisions are important from several different perspectives. Costs for replacement heifers may represent up to 20 percent of the dairy operating budget (Fetrow 1988, AABP). Negative cash flows occur when a cow is sold for beef and a heifer is added to the lactating herd as a replacement. Cows retained in the herd represent capital investments, which are subject to various forms of risk that may alter the earnings from those investments. Cows have different risks of being marketed depending on their age. Although there is a tendency for increased marketing rates with advancing age, management constraints and biases can modify this relationship. The typical cow remains in the milking herd less than 4 years even though peak milk production related to maturity ordinarily does not decline until 8 or 9 years of age. The reluctance of some producers to market first calf heifers and choosing instead to give them a second chance is an example of management bias affecting marketing policy. A recent study (Bascom and Young, JDS 1998) summarized the reasons dairies market cows and determined whether cows were marketed for multiple reasons. Dairy producers identified a secondary reason for marketing 35 percent of the time, and a tertiary reason for marketing 11 percent of the time. Unfortunately, DHIA data only provides the producer with one choice when categorizing marketing decisions. The most prevalent reason for marketing was reproduction. Producers may be unaware of the cost associated with reproductive marketing. In herds with less than optimal reproductive performance, dairy operators must find a balance between income loss caused by excessive days open and income loss caused by high marketing rates. The second most prevalent reason for marketing was mastitis. High somatic cell count (SCC) was rarely used as the reason for marketing, however, clinical mastitis was the primary reason for 15 percent of the cows marketed in this study. How producers interpret the difference between mastitis and high SCC is unknown, and marketing for mastitis may include both categories. Cows may be marketed for mastitis because they never recover from chronic infections or because of reduced milk production due to elevated SCC. In 1994, the National Cattlemen s Association performed an audit of market beef cows, market dairy cows, and market bulls. This study was conducted to determine areas for improvement in the manner by which these classes of animals were marketed. In 1999, the study was repeated to see if the concerns found five years previously had changed. Slightly over 6 million head of market cows and bulls were assessed in the 1999 audit. In general, producers did a good job of managing and marketing surplus animals. However, quality defects in only 1% of market cows and bulls indicate that thousands of cattle are below acceptable standards. On average, 3 percent of all market dairy cows are condemned at USDA packing plants. Considering only emaciated and disabled cattle, over 40 percent of these market cows are condemned. Lameness and disabled cattle represent a problem to the industry from a public perception standpoint. Many lame cattle, however, are the result of failure to market animals before feet and leg problems progress. The packer is required to remove all tissue associated with an arthritic joint. More than 7 percent of cattle had at least one arthritic joint, and nearly 4 percent had two bad joints. With an average trim loss of 40 pounds, more than 37 million pounds of product would have been lost in 1999 due to joint problems alone. Packers listed arthritis as one of their top concerns. Since lean muscle is the principal end product of market cattle, it is important that market animals do have adequate muscling and do not have excessive 103
107 amounts of fat. However, the 1999 audit suggested that over 70 percent of dairy cows were inadequately muscled. Certainly, dairy cows are not genetically designed for extreme muscling, but of greater concern is that the poor meat yield due to emaciation. Extremely thin cows amounted to 4.5 percent of dairy cows harvested in the 1999 audit. In many cases, the most valuable part of a thin cow is her hide. Emaciated cows are much more prone to bruising because they have no fat to serve as padding and they are more likely to be disabled upon arrival at the packing plant. The primary concern of packers in the 1999 audit was the high incidence of bruising. Only 11.8 percent of cow carcasses did not have a bruise (down from the 1994 audit). Minor, medium, major and extreme bruises results in 0.69, 1.42, 4.78 and 15 pounds of trim loss, respectively. Using these estimates, more than 14 million pounds of product were lost due to bruising. Unfortunately, the bruises do not just occur on the lower-valued portion of the carcass. The 1999 audit revealed that trim loss was observed in the top sirloin, loin, rib, round and chuck. When a bruise is created on an animal, it takes time for the body to heal. Handling practices at the farm are very important in minimizing bruises. It is estimated that one-third of bruises occur on the farm and the other two-thirds occur in transport and marketing. Close scrutiny of handling facilities to eliminate sharp, protruding corners can help reduce bruising. Producers should also merchandise market cattle before they become emaciated and are more susceptible to bruises. Another major concern of packers was the incidence of injection-site lesions and the potential for antibiotic residues. A recent study at Colorado State University revealed that approximately 29 percent of the rounds of market cows contain an injection-site lesion. Most of these lesions were detected in the upper portions of the hip. These lesions do not represent a food safety concern, but they are a beef quality problem. Scar tissue from intramuscular (IM) injections of antibiotics or vaccines causes the muscle tissue to be tougher, producing a product that may be unacceptable to the consumer. Producers should carefully avoid marketing cattle that have been treated with antibiotics until the specified withdrawal time has ended. The USDA currently monitors the incidence of antibiotic residues in market cattle, and a trace-back system is already in place through the use of back tags at the auction barn. In the 1994 audit, dairy cows and veal calves were the two classes with the highest level of violative antibiotic residues, 1.5 and 1.8 percent, respectively. By 1999, the violation level in dairy cows had dropped to 1.1 percent (415 positives from 37,308 cattle tested). Inspectors at packing plants identify animals to be tested both ante mortem and post mortem. Nearly 85 percent of the residue violations were from post mortem-identified high risk cattle. The leading causes of antibiotic contamination included gentamycin (39%), penicillin (25%), sulfadimethoxine (12%), streptomycin (9%), tetracyclines (6%) and several others. Gentamycin may not be as commonly used as some of these other antibiotics, but the prolonged tissue retention may explain the reason this drug is at the top of the list. When you look at the overall picture, the 1999 audit suggests that nearly $70 is lost in value for every market cow or bull that is merchandised. Most of this loss comes from merchandising thin, emaciated animals that are more susceptible to bruises and trim loss and have poor yields. Dairy Beef Quality Assurance (DBQA) addresses the day-to-day management practices that influence safety, quality, and wholesomeness of beef and beef products. Reducing the problem starts on the dairy and suggested changes involve seven steps. 1. Use the neck or shoulder as preferred injection site, when possible. 2. Read and understand injection product labels. 3. Avoid intramuscular (IM) injections when other labeled administration routes are available. 4. Products approved for subcutaneous injection should be done with the tenting technique by lifting animal hide between fingers and inject into the tent. 5. Avoid mixing products as this causes more tissue damage, reduces product efficacy, and extends withdrawal times. 6. Ask your veterinarian about comparable tissue damage from different products. 7. Encourage promotion of tissue reaction information from pharmaceutical companies that produce injectable products. The first step may be the biggest obstacle. Typically, large groups of dairy cows are not run through squeeze chutes like beef cattle. Injections in cows tossing their heads while in the stanchion can be dangerous. Is there an economic incentive from the buyer to justify the extra effort by the dairy personnel to reduce hindquarter injections? Tracing antibiotic residues is currently being pursued in dairy market cow carcasses. Tracing carcass quality (injection site lesions) back to the dairy would be needed to promote this type of quality assurance. Quality assurance is not an all or none situation and partial improvements could be beneficial. 104

108 The Wisconsin Dairy Feed Cost Evaluator Victor E. Cabrera 105

109 106

110 107

111 108

112 109
113 110
114 111
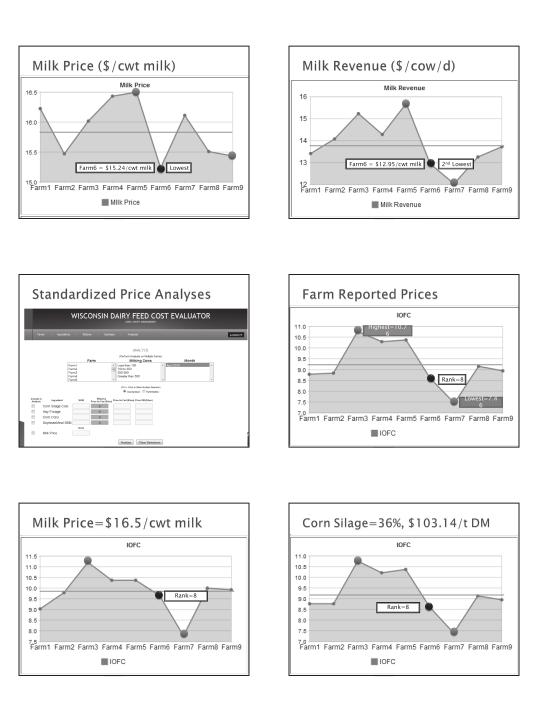
115 112
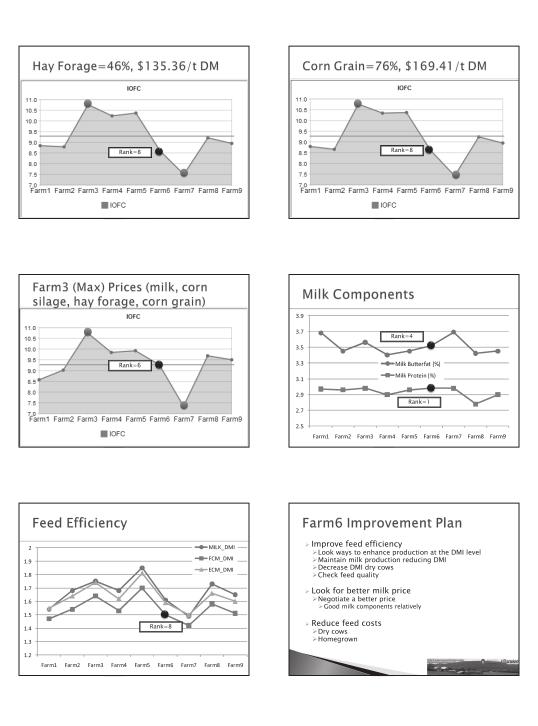
116 113

117 114
118 Improving Reproductive Efficiency using Double Ovsynch Milo C. Wiltbank, Alexandre H. Souza, Alexandre P. Cunha, Henderson Ayres, Roberto Sartori, Julio O. Giordano, Paul M. Fricke, Robb Bender, Anibal B. Nascimento, Jerry N. Guenther Department of Dairy Science, University of Wisconsin-Madison, Madison, WI Introduction High producing dairy cows have low reproductive efficiency due to reduced fertility, reduced expression of estrus, and reduced detection of estrus [1-3]. Ovsynch utilizes GnRH and prostaglandin F2a to synchronize ovulation (GnRH 7d PGF 2d- GnRH 16h AI) allowing timed AI [4]. Although all cows have AI during Ovsynch, fertility is generally not improved [5-7]. Previous experiments using lactating dairy cows [8] and dairy heifers [9] found that the ideal phase to initiate Ovsynch is from Days 5 to 12 of the estrous cycle. Therefore, researchers developed pre-synchronization systems that increase the proportion of cows in the ideal part of the estrous cycle on the day of the first GnRH of Ovsynch. For instance, Moreira and coworkers [10] reported that two PGF treatments 14 d apart, termed Presynch, increased the percentage of cows in early to mid-luteal phase and improved fertility in cycling cows when Ovsynch was initiated 12 d later. Anovular cows did not benefit from Presynch [10]. Other studies using Presynch reported improved fertility [11,12]; although, a single treatment with PGF prior to Ovsynch was not effective [13]. Anovular cows have reduced fertility to timed AI following Ovsynch [14-16]. On average 20 to 30% of cows are anovular at the time Ovsynch initiation [10,14,17]. Thus, better Presynch protocols should stimulate cyclicity in anovular cows. The CIDR during Presynch increased percentage of previously anovular cows that were cycling at the start of Ovsynch; but, did not increase fertility to timed AI [16,18]. Another limitation of the standard PGF-based Presynch protocol is that follicular and luteal stages are not precisely synchronized due to variability in time to estrus/ovulation after PGF. The G-6-G protocol [19] (PGF 2d GnRH 6d-Ovsynch) can increase ovulation to the first GnRH of Ovsynch, increase circulating progesterone at time of PGF, reduce variation in size of ovulatory follicle, and increase synchronization rates during Ovsynch[19]. Double Ovsynch (shown in Table 1) uses a complete Ovsynch protocol as a presynchronization procedure. The first Ovsynch is generally initiated on Friday with a GnRH treatment followed 7 days later by a PGF treatment. There is then a 3 day interval between the PGF treatment and the second GnRH treatment. This longer interval allows the cows to come heat and higher estradiol-17ß concentrations, potentially causing more contraction of the uterus and higher progesterone during the Ovsynch protocol. This first Ovsynch is known as the presynchronization Ovsynch. The second Ovsynch is known as the breeding Ovsynch and is done in a typical manner as the Ovsynch-56 protocol. This paper will summarize our results utilizing Double Ovsynch. Table 1. Typical calendar for Double Ovsynch. Fertility following Double Ovsynch Our first study [20] compared Double-Ovsynch to Presynch-Ovsynch in two commercial dairies. Lactating Holstein (n = 337) cows, were assigned to two treatment groups: 1) Presynch (n = 180), two injections of PGF 14 d apart, followed by the Ovsynch-timed AI protocol 12 d later; 2) Double- Ovsynch (n = 157), received GnRH, PGF 7 d later, and GnRH 3 d later followed by the Ovsynch-timed AI protocol 7 d later. All cows received the same Ovsynch-timed AI protocol: GnRH (G1) at 68 ± 3 DIM (mean ± SEM), PGF 7 d later, GnRH (G2) 56 h after PGF, and AI 16 to 20 h later. Pregnancy was diagnosed 39 to 45 d after timed AI. Double-Ovsynch increased the fertility (% pregnancy per AI) compared to Presynch-Ovsynch (49.7% vs 41.7%, P = 0.03). Parity (primiparous vs multiparous) had an effect (P = 0.05) on percentage pregnant per AI (Table 2) with primiparous (54.9%) having greater fertility than multiparous (38.5%) cows. Unexpectedly, Double- Ovsynch increased fertility only in primiparous cows (65.2% vs 45.2%; P = 0.02) with no effect of treatment observed in multiparous cows (37.5% vs 39.3%; P = 0.58). 115

119 Table 2. Effects of number of lactations on pregnancies per AI (P/AI) following presynchronization with a standard Presynch protocol or Double-Ovsynch. * P-values for comparisons of Primiparous vs Multiparous. ** P-values for comparisons of Double-Ovsynch vs Presynch. # These analyses were done with GLIMMIX and accounted for BCS in the model. There was also an effect (P = 0.04) of BCS on fertility (High = 48.3% vs Low = 28.3%). This negative impact of low BCS on fertility was numerically similar in Double-Ovsynch (20.1% decrease) and Presynch (19.5% decrease) cows. In addition, there were numerically similar effects of Double-Ovsynch on fertility in cows with both high (Double-Ovsynch 12.5% greater than Presynch; P = 0.06) and low (Double-Ovsynch 11.9% greater than Presynch; P = 0.25) BCS; although, statistical differences are clearly not detected in the small number of cows with low BCS in this study (only 46 cows in this classification). Physiology of Double Ovsynch In a subset of 87 cows [20], ovarian ultrasonography and progesterone measurements were performed at the first GnRH of the breeding Ovsynch and 7 d later (Table 3). Double-Ovsynch decreased the percentage of cows with low progesterone (< 1 ng/ml) at the first GnRH of the breeding Ovsynch (9.4 vs 33.3%) and increased the concentration of progesterone at the PGF (Table 3). Table 3. Effects of synchronization protocol on presence of CL at 1st GnRH, ovulation to first GnRH, concentration of progesterone (P4) at first GnRH, percentage of cows with P4 concentration more than 1.0 ng/ml at first GnRH, concentration of P4 at PGF, and percentage of cows with P4 concentration more than 1.0 ng/ml at PGF (mean ± SEM). In a second study (Ayres et al., unpublished) we evaluated the follicular dynamics and hormonal concentrations in a greater number of cows that were treated with Double Ovsynch compared to Presynch- Ovsynch. Lactating Holstein (n = 193) cows were assigned to two treatment groups: 1) Presynch (n = 93), two injections of PGF2a 14 days apart, followed by the Ovsynch-timed AI protocol 12 d later; or 2) Double- Ovsynch (n = 100). In this study Double-Ovsynch decreased the percentage of cows with low circulating progesterone concentration (< 0.50 ng/ml) at the first GnRH (12.0% vs. 30.1%; P = 0.003) and increased the percentage of cows with medium progesterone concentration (0.50 > P4 3.0 ng/ml) at the first GnRH (80.0% vs. 57.0%; P = ), with CL at the first GnRH (94.0% vs. 67.8%; P = ), and with high progesterone concentration (> 3.0 ng/ml) at PGF2a (88.0% vs. 76.3%; P = 0.04). Also, this protocol seemed to increase the ovulation rate at the first GnRH (80.0% vs. 69.9%; P = 0.11) and at second GnRH (98.0% vs. 93.5%; P = 0.08), the serum concentration of progesterone at PGF2a (3.52 ± 0.17 vs ± 0.21 ng/ml; P = 0.11) and the size of the dominant follicle at G2 (15.95 ± 0.31 vs ± 0.32 mm; P = 0.13). Thus, presynchronization of cows with Double-Ovsynch is probably inducing ovulation in non-cycling cows and also improving synchronization of cycling cows. Modifications of Double Ovsynch One of the potential problems with Ovsynch protocols is that there may be inadequate luteolysis following the single PGF treatment that is given prior to the final GnRH and timed AI. In one experiment [21] we evaluated whether a second PGF treatment would increase luteolysis and improve fertility (Table 4). There was a dramatic improvement in percentage of cows that had complete luteolysis by the time of the second GnRH (86.1% to 97.5%). There was not a statistically significant improvement in fertility; although there was a 5.7 % improvement in fertility with the additional PGF treatment (Table 5). This would be approximately the amount of improvement that would be expected with the improvement in luteolysis rate (i.e. 11.4% of cows that did not regress their CL regressed after the second PGF, if 50% of these cows conceive then a 5.7% improvement is expected [11.4% X 0.5 = 5.7%]). Thus, it seems likely that an additional PGF will improve fertility in Double Ovsynch; however a 4-6% increase is about the most that can be expected. 116
 at second GnRH of Ovsynch (< 0.5 ng/mll).")
120 Table 4. Experiment with Double Ovsynch to increase luteolysis by adding a second PGF treatment. Table 5. Effects of additional PGF on fertility and percentage of cows with low progesterone (P4) at second GnRH of Ovsynch (< 0.5 ng/mll). We have recently tested whether Double Ovsynch can be utilized as a resynchronization strategy, rather than just using this strategy for first AI (Giordano et al., unpublished). Table 6 shows the way that we have used Double Ovsynch for resynchronization. The first GnRH of the protocol was done at Day 22 and the pregnancy diagnosis and PGF treatment were done at Day 29. This was compared to a Resynch-32 strategy (normal Ovsynch protocol with first GnRH done at 32 days after the previous timed AI). The Resynch-32 strategy will result in AI being done at 42 days after the previous AI. The Double Ovsynch Resynch will result in AI being done at 49 days after the previous AI. In this experiment Double Ovsynch was utilized for first AI with good fertility (46.4%; 507/1092; primiparous %, multiparous %). During the Resynch experiment, Double Ovsynch had much higher (P < ) fertility (38.2%; 305/799) than Resynch-32 (29.9%; 242/810). Thus, Double Ovsynch can be utilized as a Resynch procedure to improve fertility at second and later AIs; however, it produces a longer interval between AIs and requires more hormonal treatments than Resynch-32. Table 6. Use of Double Ovsynch for resynchronization of Ovulation. Another modification that we have tested is to eliminate one of the GnRH treatments of the Double Ovsynch protocol. This has been termed the short Double Ovsynch protocol (see Table 7). This protocol was tested for two reasons. First, this may be a more practical protocol for use in Resynchronization strategies because it could produce a shorter interval between AIs (42 days rather than 49 days with Double Ovsynch Resynch). Second, we wanted to test whether the progesterone concentration during the final follicular wave would alter fertility to the timed AI. For example the short Double Ovsynch protocol would have low progesterone concentrations during this final follicular wave (only one CL present between GnRH and PGF of breeding Ovsynch); whereas, the normal Double Ovsynch protocol normally has two CL present at this time due to ovulation of a second CL at the first GnRH of the breeding Ovsynch. Table 7. protocol. Calendar for the short Double Ovsynch Some intriguing effects were observed in this comparison of the short and normal Double Ovsynch protocols. First, the normal Double Ovsynch protocol had a lower double ovulation rate than the short protocol (12.3% [32/261] vs. 23.4% [54/231]; P < 0.001). In addition, there was greater fertility with the normal Double Ovsynch protocol than short Double Ovsynch (51.0% [149/292] vs. 37.1% [101/272]). Surprisingly, there was also a reduction in pregnancy loss in cows treated with Double Ovsynch compared to short Double Ovsynch (6.8% [10/147] vs. 14.3% [14/98]; P = 0.054). The decrease in double ovulation and increase in fertility occurred in both primiparous and multiparous cows in this study. Thus, the higher progesterone concentrations that are produced by the Double Ovsynch protocol are critical for reducing double ovulation, increasing fertility, and potentially reducing pregnancy loss. Perhaps even greater progesterone concentrations would reduce double ovulation and improve fertility to an even greater extent. Conclusions In the trials that we have performed, there has been a consistently high fertility following the Double Ovsynch protocol. In field observations of fertility in herds that use Double Ovsynch there is some variability in results that have been obtained. In almost all herds that use Double Ovsynch there is much higher fertility in first lactation cows 117
121 (primiparous) than older cows (multiparous) similar to our observation in the published study on Double Ovsynch [20]. The reason(s) for this difference are not clear at this time. Further studies are needed to continue to evaluate and improve this protocol. Nevertheless, the surprisingly high fertility that is observed with the Double Ovsynch protocol, particularly in first lactation cows, indicates that enhanced timed AI protocols may be able to optimize hormonal concentrations and follicle development and apparently overcome much of the physiological barriers to high fertility in lactating dairy cows. Unfortunately, the variability between herds indicates that many other problems can also reduce fertility in lactating dairy cows (insemination technique, nutrition, disease, etc.) and therefore even the most optimized timed AI protocols are unlikely to improve fertility in all commercial dairies. References [1] Washburn SP, Silvia WJ, Brown CH, McDaniel BT, McAllister AJ. Trends in reproductive performance in southeastern Holstein and Jersey DHI herds. J Dairy Sci 2002;85: [2] Lopez H, Satter LD, Wiltbank MC. Relationship between level of milk production and estrous behavior of lactating dairy cows. Anim Reprod Sci 2004;81: [3] Wiltbank MC, Lopez H, Sartori R, Sangsritavong S, Gumen A. Changes in reproductive physiology of lactating dairy cows due to elevated steroid metabolism. Theriogenology 2006;65: [4] Pursley RJ, Mee MO, Wiltbank MC. Synchronization of ovulation in dairy cows using PGF2a and GnRH. Theriogenology 1995;44: [5] Pursley RJ, Kosorok MR, Wiltbank MC. Reproductive management of lactating dairy cows using synchronization of ovulation. J Dairy Sci 1997;80: [6] Cerri RL, Santos JE, Juchem SO, Galvao KN, Chebel RC. Timed artificial insemination with estradiol cypionate or insemination at estrus in high-producing dairy cows. J Dairy Sci 2004;87: [7] Stevenson JS, Kobayashi Y, Thompson KE. Reproductive performance of dairy cows in various programmed breeding systems including Ovsynch and combinations of gonadotropin releasing hormone and prostaglandin F2a J Dairy Sci 1999;82: [8] Vasconcelos JL, Silcox RW, Rosa GJ, Pursley RJ, Wiltbank MC. Synchronization rate, size of the ovulatory follicle, and pregnancy rate after synchronization of ovulation beginning on different days of the estrous cycle in lactating dairy cows. Theriogenology 1999;52: [9] Moreira F, De la Sota RL, Diaz T, Thatcher WW. Effect of day of the estrous cycle at the initiation of timed artificial insemination protocol on reproductive responses in dairy heifers. J Anim Sci 2000;78: [10] Moreira F, Orlandi C, Risco CA, Mattos R, Lopes F, Thatcher WW. Effects of presynchronization and bovine somatotropin on pregnancy rates to timed artificial insemination protocol in lactating dairy cows. J Dairy Sci 2001;84: [11] El-Zarkouny SZ, Cartmill JA, Hensley BA, Stevenson JS. Pregnancy in dairy cows after synchronized ovulation regimens with or without Presynchronization and progesterone. J Dairy Sci 2004;87: [12] Navanukraw C, Redmer DA, Reynolds LP, Kirsch JD, Grazul-Bilska AT, Fricke PM. A modified Presynchronization protocol improves fertility to timed artificial insemination in lactating dairy cows. J Dairy Sci 2004;87: [13] LeBlanc SJ, Leslie KE. Short communication: Presynchronization using a single injection of PGF2a before synchronized ovulation and first timed artificial insemination in dairy cows. J Dairy Sci 2003;86: [14] Gümen A, Guenther JN, Wiltbank MC. Follicular size and response to Ovsynch versus detection of estrus in anovular and ovular lactating dairy cows. J Dairy Sci 2003;86: [15] Galvão KN, Santos JEP, Juchem SO, Cerri RLA, Coscioni AC, Villaseñor M. Effect of addition of a progesterone intravaginal insert to a timed insemination protocol using estradiol cypionate on ovulation rate, pregnancy rate, and late embryonic loss in lactating dairy cows. J Anim Sci 2004;82: [16] Chebel RC, Santos JEP, Cerri RLA, Rutigliano HM, Bruno RGS. Reproduction in dairy cows following progesterone insert presynchronization and resynchronization protocols. J Dairy Sci 2006;89: [17] Lopez HD, Caraviello DZ, Satter LD, Fricke PM, Wiltbank MC. Relationship between level of milk production and multiple ovulations in lactating dairy cows. J Dairy Sci 2005;88: [18] Bicalho RC, Cheong SH, Warnick LD, Guard CL. Evaluation of progesterone supplementation in a prostaglandin F2a-based presynchronization protocol before timed insemination. J Dairy Sci 2007;90: [19] Bello NM, Steibel JP, Pursley RJ. Optimizing ovulation to first GnRH improved outcomes to each hormonal injection of Ovsynch in lactating dairy cows. J Dairy Sci 2006;89: [20] Souza AH, Ayres H, Ferreira RM, Wiltbank MC. A new presynchronization system (Double-Ovsynch) increases fertility at first postpartum timed AI in lactating dairy cows Theriogenology 2008; 70: [21] Brusveen DJ, Souza AH, Wiltbank MC, Effects of additional prostaglandin F2a and estradiol-17ß during Ovsynch in lactating dairy cows. Journal of Dairy Science 92:

122 119

123 120
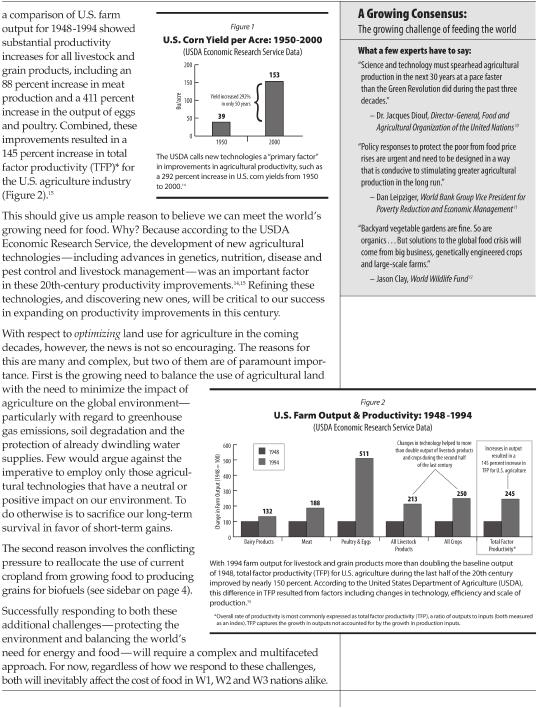
124 121
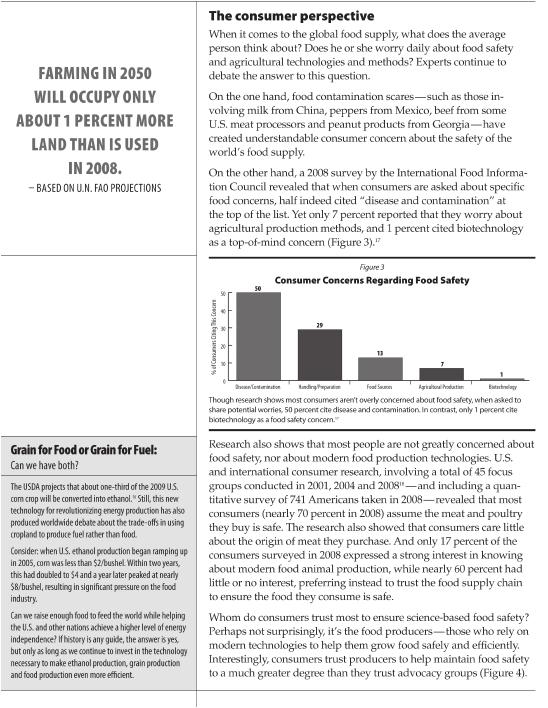
125 122
126 123

127 124
128 125

129 126

130 127

131 128
Strategies and Rationale for Resynchronization of Ovulation in Lactating Dairy Cows
 The Dairy Cattle Reproduction Council does not support one product over another and any mention herein is meant as an example, not an endorsement. Strategies and Rationale for Resynchronization of Ovulation
The Dairy Cattle Reproduction Council does not support one product over another and any mention herein is meant as an example, not an endorsement. Strategies and Rationale for Resynchronization of Ovulation
Minnesota Dairy Health Conference. May 19-20, 2010 St. Paul, Minnesota
 Minnesota Dairy Health Conference May 19-20, 2010 St. Paul, Minnesota Updates on Reproductive Management of Dairy Cows and Heifers Ricardo C. Chebel, DVM, MPVM College of Veterinary Medicine University
Minnesota Dairy Health Conference May 19-20, 2010 St. Paul, Minnesota Updates on Reproductive Management of Dairy Cows and Heifers Ricardo C. Chebel, DVM, MPVM College of Veterinary Medicine University
Setting up Lactating Dairy Cows for First Postpartum Timed AI
 Setting up Lactating Dairy Cows for First Postpartum Timed AI Professor of Dairy Science University of Wisconsin-Madison Synch Timeline 1980 1985 1990 1995 2000 2005 2010 1979 Lutalyse approved for use
Setting up Lactating Dairy Cows for First Postpartum Timed AI Professor of Dairy Science University of Wisconsin-Madison Synch Timeline 1980 1985 1990 1995 2000 2005 2010 1979 Lutalyse approved for use
Reproductive Programs for Florida Dairy Herds
 Reproductive Programs for Florida Dairy Herds J.E.P. Santos, W.W. Thatcher, R.S. Bisinotto, F.S. Lima, and E.S. Ribeiro Department of Animal Sciences, University of Florida Bldg. 499, Shealy Drive, Gainesville,
Reproductive Programs for Florida Dairy Herds J.E.P. Santos, W.W. Thatcher, R.S. Bisinotto, F.S. Lima, and E.S. Ribeiro Department of Animal Sciences, University of Florida Bldg. 499, Shealy Drive, Gainesville,
Reproductive Management of Dairy Heifers
 Reproductive Management of Dairy Heifers Carlos A. Risco University of Florida, College of Veterinary Medicine, Gainesville, FL, 32610 Email: RiscoC@vetmed.ufl.edu Take Home Messages 8 Dairy producers
Reproductive Management of Dairy Heifers Carlos A. Risco University of Florida, College of Veterinary Medicine, Gainesville, FL, 32610 Email: RiscoC@vetmed.ufl.edu Take Home Messages 8 Dairy producers
New Information on Timed Breeding Protocols for Lactating Dairy Cows
 New Information on Timed Breeding Protocols for Lactating Dairy Cows Paul M. Fricke Department of Dairy Science, University of Wisconsin - Madison, Madison, WI, USA 53706-1284 E-mail: fricke@calshp.cals.wisc.edu
New Information on Timed Breeding Protocols for Lactating Dairy Cows Paul M. Fricke Department of Dairy Science, University of Wisconsin - Madison, Madison, WI, USA 53706-1284 E-mail: fricke@calshp.cals.wisc.edu
The High Plains Dairy Conference does not support one product over another and any mention herein is meant as an example, not an endorsement.
 Managing Reproduction of Lactating Dairy Cows with Limited Use of Timed AI Programs Luís G. D. Mendonça, D.V.M., M.S. and Alexandre L. A. Scanavez, D.V.M. Department of Animal Sciences and Industry Kansas
Managing Reproduction of Lactating Dairy Cows with Limited Use of Timed AI Programs Luís G. D. Mendonça, D.V.M., M.S. and Alexandre L. A. Scanavez, D.V.M. Department of Animal Sciences and Industry Kansas
Management Strategies to Improve Fertility in Lactating Dairy Cows
 Management Strategies to Improve Fertility in Lactating Dairy Cows Paul M. Fricke and Ryan A. Sterry Department of Dairy Science University of Wisconsin-Madison Implementing a Systematic Synchronization
Management Strategies to Improve Fertility in Lactating Dairy Cows Paul M. Fricke and Ryan A. Sterry Department of Dairy Science University of Wisconsin-Madison Implementing a Systematic Synchronization
Management Strategies to Optimize Reproductive Performance of Dairy Herds
 Management Strategies to Optimize Reproductive Performance of Dairy Herds Dr. Julio O. Giordano Department of Animal Science, Cornell University, Ithaca 14853 Email: jog25@cornell.edu Take Home Messages
Management Strategies to Optimize Reproductive Performance of Dairy Herds Dr. Julio O. Giordano Department of Animal Science, Cornell University, Ithaca 14853 Email: jog25@cornell.edu Take Home Messages
Estrus Synchronization in Dairy Cattle
 Estrus Synchronization in Dairy Cattle Reproduction is one of the most important aspects of a successful dairy operation but is often overlooked. Most dairy operations focus on increasing milk production
Estrus Synchronization in Dairy Cattle Reproduction is one of the most important aspects of a successful dairy operation but is often overlooked. Most dairy operations focus on increasing milk production
APPLICATION OF THE PROGESTERONE (CIDR) INSERT IN ARTIFICIAL INSEMINATION PROGRAMS OF DAIRY CATTLE. J. S. Stevenson
 INSERT IN ARTIFICIAL INSEMINATION PROGRAMS OF DAIRY CATTLE. J. S. Stevenson") Dairy Research 007 APPLICATION OF THE PROGESTERONE () INSERT IN ARTIFICIAL INSEMINATION PROGRAMS OF DAIRY CATTLE J. S. Stevenson Summary Use of progesterone inserts (controlled interval drug release, )
Dairy Research 007 APPLICATION OF THE PROGESTERONE () INSERT IN ARTIFICIAL INSEMINATION PROGRAMS OF DAIRY CATTLE J. S. Stevenson Summary Use of progesterone inserts (controlled interval drug release, )
Systematic Synchronization and Resynchronization Systems for Reproductive Management of Lactating Dairy Cows
 Systematic Synchronization and Resynchronization Systems for Reproductive Management of Lactating Dairy Cows Paul M. Fricke, PhD Department of Dairy Science, University of Wisconsin-Madison Introduction
Systematic Synchronization and Resynchronization Systems for Reproductive Management of Lactating Dairy Cows Paul M. Fricke, PhD Department of Dairy Science, University of Wisconsin-Madison Introduction
Dairy Herd Synchronization Programs. William M. Graves Extension Dairy Scientist
 Dairy Herd Synchronization Programs William M. Graves Extension Dairy Scientist Research has shown that pregnant cows survive 420 days longer than herdmates that were open. A cow must be pregnant by 85
Dairy Herd Synchronization Programs William M. Graves Extension Dairy Scientist Research has shown that pregnant cows survive 420 days longer than herdmates that were open. A cow must be pregnant by 85
Estrous synchronization programs for the dairy herd
 College of Agricultural Sciences Cooperative Extension Estrous synchronization programs for the dairy herd Michael L. O Connor Department of Dairy and Animal Science The Pennsylvania State University 324
College of Agricultural Sciences Cooperative Extension Estrous synchronization programs for the dairy herd Michael L. O Connor Department of Dairy and Animal Science The Pennsylvania State University 324
Industry Presentation Ten Lessons That We Have Learned about Systematic Breeding Programs for Dairy Cattle
 Ten Lessons That We Have Learned about Systematic Breeding Programs for Dairy Cattle Jerry D. Olson, B.S., D.V.M, M.S. Diplomate, ACT Senior Veterinarian Pfizer Animal Health Ft. Collins, CO 80528 The
Ten Lessons That We Have Learned about Systematic Breeding Programs for Dairy Cattle Jerry D. Olson, B.S., D.V.M, M.S. Diplomate, ACT Senior Veterinarian Pfizer Animal Health Ft. Collins, CO 80528 The
Reproductive Management of Dairy Heifers
 Reproductive Management of Dairy Heifers Associate Professor Department of Dairy Science University of Wisconsin - Madison Heifer Development Age at 1 st Calving = 24 mo Birth 1 st Calving 2 nd Calving
Reproductive Management of Dairy Heifers Associate Professor Department of Dairy Science University of Wisconsin - Madison Heifer Development Age at 1 st Calving = 24 mo Birth 1 st Calving 2 nd Calving
Aggressive Management Strategies for Improving Reproductive Efficiency
 The Babcock Institute University of Wisconsin Dairy Updates Aggressive Management Strategies for Improving Reproductive Efficiency Reproduction and Genetics No. 604 Author: Dr. Paul M. Fricke 1 Take home
The Babcock Institute University of Wisconsin Dairy Updates Aggressive Management Strategies for Improving Reproductive Efficiency Reproduction and Genetics No. 604 Author: Dr. Paul M. Fricke 1 Take home
Pregnant vs. Open: Getting Cows Pregnant and the Money it Makes
 Pregnant vs. Open: Getting Cows Pregnant and the Money it Makes Paul Fricke, University of Wisconsin-Madison, Department of Dairy Science, Madison, WI Steve Stewart, University of Minnesota, College of
Pregnant vs. Open: Getting Cows Pregnant and the Money it Makes Paul Fricke, University of Wisconsin-Madison, Department of Dairy Science, Madison, WI Steve Stewart, University of Minnesota, College of
AN UPDATE ON TIMED-AI (TAI) PROGRAMS FOR DAIRY CATTLE
 PROGRAMS FOR DAIRY CATTLE") AN UPDATE ON TIMED-AI (TAI) PROGRAMS FOR DAIRY CATTLE Marcos G. Colazo Alberta Agriculture & Rural Development Edmonton, Alberta, Canada Outline of Webinar Review the physiology and endocrinology of the
AN UPDATE ON TIMED-AI (TAI) PROGRAMS FOR DAIRY CATTLE Marcos G. Colazo Alberta Agriculture & Rural Development Edmonton, Alberta, Canada Outline of Webinar Review the physiology and endocrinology of the
Department of Dairy Science, University of Wisconsin-Madison, 1675 Observatory Dr., Madison, WI 53706, U. S. A.
 Fertility associated economic losses of farms V. E. Cabrera Department of Dairy Science, University of Wisconsin-Madison, 1675 Observatory Dr., Madison, WI 53706, U. S. A. Introduction Herd net return
Fertility associated economic losses of farms V. E. Cabrera Department of Dairy Science, University of Wisconsin-Madison, 1675 Observatory Dr., Madison, WI 53706, U. S. A. Introduction Herd net return
Ovsynch, Pre-synch, the Kitchen-Synch: What s up with Synchronization Protocols? Paul M. Fricke, Ph.D.
 Ovsynch, Pre-synch, the Kitchen-Synch: What s up with Synchronization Protocols? Paul M. Fricke, Ph.D. Assistant Professor and Extension Specialist in Dairy Cattle Reproduction Department of Dairy Science
Ovsynch, Pre-synch, the Kitchen-Synch: What s up with Synchronization Protocols? Paul M. Fricke, Ph.D. Assistant Professor and Extension Specialist in Dairy Cattle Reproduction Department of Dairy Science
Estrous Synchronization Programs for Dairy Cows and Heifers
 Estrous Synchronization Programs for Dairy Cows and Heifers Jerry D. Olson, DVM, MS, DACT Technical Consultant for Pharmacia Animal Health 1808 Willow Springs Way Fort Collins, Colorado 89528 Telephone:
Estrous Synchronization Programs for Dairy Cows and Heifers Jerry D. Olson, DVM, MS, DACT Technical Consultant for Pharmacia Animal Health 1808 Willow Springs Way Fort Collins, Colorado 89528 Telephone:
TAKE HOME MESSAGES Illinois Parameter < 18,000 18,000 22,000 > 22,000
 TAKE HOME MESSAGES Average days open for the top, middle and bottom third of Illinois dairy herds on DHIA in September 2000 were 172, 186 and 204 days, respectively. At a cost of $2.00 per day open after
TAKE HOME MESSAGES Average days open for the top, middle and bottom third of Illinois dairy herds on DHIA in September 2000 were 172, 186 and 204 days, respectively. At a cost of $2.00 per day open after
THIS ARTICLE IS SPONSORED BY THE MINNESOTA DAIRY HEALTH CONFERENCE.
 THIS ARTICLE IS SPONSORED BY THE MINNESOTA DAIRY HEALTH CONFERENCE. ST. PAUL, MINNESOTA UNITED STATES OF MINNESOTA OVSYNCH: A Method For Breeding Dairy Cows Without Doing Heat Detection Dr. Brad Seguin,
THIS ARTICLE IS SPONSORED BY THE MINNESOTA DAIRY HEALTH CONFERENCE. ST. PAUL, MINNESOTA UNITED STATES OF MINNESOTA OVSYNCH: A Method For Breeding Dairy Cows Without Doing Heat Detection Dr. Brad Seguin,
Received 10 February 2004; accepted 1 July 2004
 Theriogenology 63 (2005) 1628 1642 www.journals.elsevierhealth.com/periodicals/the Resynchronization of ovulation and timed insemination in lactating dairy cows, II: assigning protocols according to stages
Theriogenology 63 (2005) 1628 1642 www.journals.elsevierhealth.com/periodicals/the Resynchronization of ovulation and timed insemination in lactating dairy cows, II: assigning protocols according to stages
Economic comparison of natural service and timed artificial insemination breeding programs in dairy cattle
 J. Dairy Sci. 93 :4404 4413 doi: 10.3168/jds.2009-2789 American Dairy Science Association, 2010. Economic comparison of natural service and timed artificial insemination breeding programs in dairy cattle
J. Dairy Sci. 93 :4404 4413 doi: 10.3168/jds.2009-2789 American Dairy Science Association, 2010. Economic comparison of natural service and timed artificial insemination breeding programs in dairy cattle
Theriogenology 81 (2014) Contents lists available at ScienceDirect. Theriogenology. journal homepage:
 Contents lists available at ScienceDirect. Theriogenology. journal homepage:") Theriogenology 81 (2014) 170 185 Contents lists available at ScienceDirect Theriogenology journal homepage: www.theriojournal.com 40 th Anniversary Special Issue The cow as an induced ovulator: Timed AI
Theriogenology 81 (2014) 170 185 Contents lists available at ScienceDirect Theriogenology journal homepage: www.theriojournal.com 40 th Anniversary Special Issue The cow as an induced ovulator: Timed AI
Proceedings, Applied Reproductive Strategies in Beef Cattle November 1 and 2, 2005, Lexington, Kentucky ESTRUS SYNCHRONIZATION SYSTEMS - COWS
 Proceedings, Applied Reproductive Strategies in Beef Cattle November 1 and 2, 2005, Lexington, Kentucky ESTRUS SYNCHRONIZATION SYSTEMS - COWS Les Anderson, Ph.D. Department of Animal and Food Science University
Proceedings, Applied Reproductive Strategies in Beef Cattle November 1 and 2, 2005, Lexington, Kentucky ESTRUS SYNCHRONIZATION SYSTEMS - COWS Les Anderson, Ph.D. Department of Animal and Food Science University
The Underlying Physiology Changing Reproduction in Lactating Dairy Cows
 The Underlying Physiology Changing Reproduction in Lactating Dairy Cows Milo Wiltbank, Hernando Lopez, Roberto Sartori, and Ahmet Gument Department of Dairy Science, University of Wisconsin-Madison Significance
The Underlying Physiology Changing Reproduction in Lactating Dairy Cows Milo Wiltbank, Hernando Lopez, Roberto Sartori, and Ahmet Gument Department of Dairy Science, University of Wisconsin-Madison Significance
IMPACTS OF ESTROUS SYNCHRONIZATION ON COWHERD PERFORMANCE
 Proceedings, The Range Beef Cow Symposium XXIV November 17, 18 and 19, Loveland, Colorado IMPACTS OF ESTROUS SYNCHRONIZATION ON COWHERD PERFORMANCE G. Cliff Lamb North Florida Research and Education Center
Proceedings, The Range Beef Cow Symposium XXIV November 17, 18 and 19, Loveland, Colorado IMPACTS OF ESTROUS SYNCHRONIZATION ON COWHERD PERFORMANCE G. Cliff Lamb North Florida Research and Education Center
FACTORS INFLUENCING THE INITIATION OF ESTROUS CYCLES AND EXPRESSION OF ESTRUS IN BEEF COWS. J.S. Stevenson
 Cattlemen s Day 2000 FACTORS INFLUENCING THE INITIATION OF ESTROUS CYCLES AND EXPRESSION OF ESTRUS IN BEEF COWS J.S. Stevenson Summary Body condition, parity, and days postpartum at the onset of the breeding
Cattlemen s Day 2000 FACTORS INFLUENCING THE INITIATION OF ESTROUS CYCLES AND EXPRESSION OF ESTRUS IN BEEF COWS J.S. Stevenson Summary Body condition, parity, and days postpartum at the onset of the breeding
Comparison of controlled internal drug release insert-based protocols to synchronize estrus in prepubertal and estrous-cycling beef heifers 1
 Published December 5, 2014 Comparison of controlled internal drug release insert-based protocols to synchronize estrus in prepubertal and estrous-cycling beef heifers 1 N. R. Leitman,* D. C. Busch,* D.
Published December 5, 2014 Comparison of controlled internal drug release insert-based protocols to synchronize estrus in prepubertal and estrous-cycling beef heifers 1 N. R. Leitman,* D. C. Busch,* D.
Cost Analysis of Implementing a Synchronization or AI Program-Using Decision-Aid Tools
 Cost Analysis of Implementing a Synchronization or AI Program-Using Decision-Aid Tools Introduction G. Cliff Lamb North Florida Research and Education Center Marianna, Florida University of Florida Estrous
Cost Analysis of Implementing a Synchronization or AI Program-Using Decision-Aid Tools Introduction G. Cliff Lamb North Florida Research and Education Center Marianna, Florida University of Florida Estrous
J. Dairy Sci. 94 : doi: /jds American Dairy Science Association, 2011.
 J. Dairy Sci. 94 :2815 2824 doi: 10.3168/jds.2010-3652 American Dairy Science Association, 2011. Effects of cloprostenol sodium at final prostaglandin F 2 of Ovsynch on complete luteolysis and pregnancy
J. Dairy Sci. 94 :2815 2824 doi: 10.3168/jds.2010-3652 American Dairy Science Association, 2011. Effects of cloprostenol sodium at final prostaglandin F 2 of Ovsynch on complete luteolysis and pregnancy
Using Resynchronization Programs
 Using Resynchronization Programs WITH ESTRUS DETECTION & ECONOMIC EVALUATION Todd Bilby, Ph.D. Associate Director, Ruminant Technical Services Merck Animal Health The Fatty Acid Forum sponsored by Achieving
Using Resynchronization Programs WITH ESTRUS DETECTION & ECONOMIC EVALUATION Todd Bilby, Ph.D. Associate Director, Ruminant Technical Services Merck Animal Health The Fatty Acid Forum sponsored by Achieving
Why AI? Accuracy Illustrated. Why AI? Why Use Estrus Synchronization? Effect of TAI on Calving Dates
 New Technologies in Artificial Insemination for Profitable Beef: WIIFM Why AI? Take advantage of superior genetics Increased selection accuracy Ted Probert Dairy Specialist University of Missouri Extension
New Technologies in Artificial Insemination for Profitable Beef: WIIFM Why AI? Take advantage of superior genetics Increased selection accuracy Ted Probert Dairy Specialist University of Missouri Extension
Introduction Materials and Methods Experiment 1 Experiment 2
 Effects of Administration of Prostaglandin F2α at Initiation of the 7-d CO- Synch+CIDR Ovulation Synchronization Protocol for Suckled Beef Cows and Replacement Beef Heifers V. R. G. Mercadante 1, L. E.
Effects of Administration of Prostaglandin F2α at Initiation of the 7-d CO- Synch+CIDR Ovulation Synchronization Protocol for Suckled Beef Cows and Replacement Beef Heifers V. R. G. Mercadante 1, L. E.
Timing of artificial insemination in postpartum beef cows following administration of the CO-Synch + controlled internal drug-release protocol 1
 Published December 5, 2014 Timing of artificial insemination in postpartum beef cows following administration of the CO-Synch + controlled internal drug-release protocol 1 D. C. Busch,* D. J. Schafer,
Published December 5, 2014 Timing of artificial insemination in postpartum beef cows following administration of the CO-Synch + controlled internal drug-release protocol 1 D. C. Busch,* D. J. Schafer,
Estrus Synchronization in Beef Cattle
 Estrus Synchronization in Beef Cattle Estrus synchronization is a reproductive management tool that is particularly useful in artificial insemination (AI) and embryo transfer (ET) programs. The estrous
Estrus Synchronization in Beef Cattle Estrus synchronization is a reproductive management tool that is particularly useful in artificial insemination (AI) and embryo transfer (ET) programs. The estrous
Received 10 February 2004; accepted 1 July 2004
 Theriogenology 63 (2005) 1617 1627 www.journals.elsevierhealth.com/periodicals/the Resynchronization of ovulation and timed insemination in lactating dairy cows I: use of the Ovsynch and Heatsynch protocols
Theriogenology 63 (2005) 1617 1627 www.journals.elsevierhealth.com/periodicals/the Resynchronization of ovulation and timed insemination in lactating dairy cows I: use of the Ovsynch and Heatsynch protocols
COST EFFECTIVENESS OF OVSYNCH PROTOCOL IN DIFFERENT COMBINATIONS FOR INDUCTION OF ESTRUS IN ANESTROUS MURRAH BUFFALOES
 Original Article COST EFFECTIVENESS OF OVSYNCH PROTOCOL IN DIFFERENT COMBINATIONS FOR INDUCTION OF ESTRUS IN ANESTROUS MURRAH BUFFALOES Sandeep Kumar 3, R.K. Malik 3, R.K. Sharma 1, Ravi Dutt 2, * and
Original Article COST EFFECTIVENESS OF OVSYNCH PROTOCOL IN DIFFERENT COMBINATIONS FOR INDUCTION OF ESTRUS IN ANESTROUS MURRAH BUFFALOES Sandeep Kumar 3, R.K. Malik 3, R.K. Sharma 1, Ravi Dutt 2, * and
Reproductive Performance in Dairy Cows Synchronized with the Ovsynch Protocol at Different Stages of the Estrus Cycle
 Reproductive Performance in Dairy Cows Synchronized with the Ovsynch Protocol at Different Stages of the Estrus Cycle Goshen, T.,, 2 * Tsitrin, K. and van Straten, M., 2 The Koret School of Veterinary
Reproductive Performance in Dairy Cows Synchronized with the Ovsynch Protocol at Different Stages of the Estrus Cycle Goshen, T.,, 2 * Tsitrin, K. and van Straten, M., 2 The Koret School of Veterinary
Effect of a second treatment with prostaglandin F during the Ovsynch protocol on luteolysis and pregnancy in dairy cows
 J. Dairy Sci. 98:8644 8654 http://dx.doi.org/10.3168/jds.2015-9353 American Dairy Science Association, 2015. Effect of a second treatment with prostaglandin F during the Ovsynch protocol on luteolysis
J. Dairy Sci. 98:8644 8654 http://dx.doi.org/10.3168/jds.2015-9353 American Dairy Science Association, 2015. Effect of a second treatment with prostaglandin F during the Ovsynch protocol on luteolysis
Strategies to Improve Economic Efficiency of the Dairy
 Strategies to Improve Economic Efficiency of the Dairy Victor E. Cabrera and Afshin S. Kalantari 1675 Observatory Dr., University of Wisconsin Madison, Department of Dairy Science, Madison, WI 53706 Email:
Strategies to Improve Economic Efficiency of the Dairy Victor E. Cabrera and Afshin S. Kalantari 1675 Observatory Dr., University of Wisconsin Madison, Department of Dairy Science, Madison, WI 53706 Email:
REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS:CIDR
 Proceedings, Applied Reproductive Strategies in Beef Cattle November 12 and 1, 25, Texas A&M University, College Station REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS:CIDR G. Cliff Lamb and Jamie E. Larson
Proceedings, Applied Reproductive Strategies in Beef Cattle November 12 and 1, 25, Texas A&M University, College Station REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS:CIDR G. Cliff Lamb and Jamie E. Larson
J. Dairy Sci. 97 : /jds American Dairy Science Association, 2014.
 J. Dairy Sci. 97 :6305 6315 http://dx.doi.org/ 10.3168/jds.2014-8222 American Dairy Science Association, 2014. Presynchronization using a modified Ovsynch protocol or a single gonadotropin-releasing hormone
J. Dairy Sci. 97 :6305 6315 http://dx.doi.org/ 10.3168/jds.2014-8222 American Dairy Science Association, 2014. Presynchronization using a modified Ovsynch protocol or a single gonadotropin-releasing hormone
Use of Intravaginal Progesterone-Releasing Device (CIDR 1 ) for Timed Artificial Insemination (AI) in Crossbred Cattle of Bos indicus Breeding 2,3
 for Timed Artificial Insemination (AI) in Crossbred Cattle of Bos indicus Breeding 2,3") Use of Intravaginal Progesterone-Releasing Device (CIDR 1 ) for Timed Artificial Insemination (AI) in Crossbred Cattle of Bos indicus Breeding 2,3 Janis Fullenwider Ray Kempfer Christin Barthle Willard
Use of Intravaginal Progesterone-Releasing Device (CIDR 1 ) for Timed Artificial Insemination (AI) in Crossbred Cattle of Bos indicus Breeding 2,3 Janis Fullenwider Ray Kempfer Christin Barthle Willard
Impact of Dry Period Length
 Rick Watters, Paul Fricke, Milo Wiltbank, and Ric Grummer Department of Dairy Science, University of Wisconsin-Madison Perry Clark Department of Dairy Science, University of Wisconsin-River Falls Take-home
Rick Watters, Paul Fricke, Milo Wiltbank, and Ric Grummer Department of Dairy Science, University of Wisconsin-Madison Perry Clark Department of Dairy Science, University of Wisconsin-River Falls Take-home
SYNCHRONIZATION OF OVULATION BASED ON STAGES OF FOLLICULAR AND CORPORA LUTEA DEVELOPMENT IN LACTATING DAIRY COWS DIAGNOSED NOT-PREGNANT.
 SYNCHRONIZATION OF OVULATION BASED ON STAGES OF FOLLICULAR AND CORPORA LUTEA DEVELOPMENT IN LACTATING DAIRY COWS DIAGNOSED NOT-PREGNANT By Megan Kron A THESIS Submitted to Michigan State University in
SYNCHRONIZATION OF OVULATION BASED ON STAGES OF FOLLICULAR AND CORPORA LUTEA DEVELOPMENT IN LACTATING DAIRY COWS DIAGNOSED NOT-PREGNANT By Megan Kron A THESIS Submitted to Michigan State University in
Comparison of protocols to synchronize estrus and ovulation in estrous-cycling and prepubertal beef heifers 1
 Published December 5, 2014 Comparison of protocols to synchronize estrus and ovulation in estrous-cycling and prepubertal beef heifers 1 N. R. Leitman,* D. C. Busch,* J. F. Bader, D. A. Mallory,* D. J.
Published December 5, 2014 Comparison of protocols to synchronize estrus and ovulation in estrous-cycling and prepubertal beef heifers 1 N. R. Leitman,* D. C. Busch,* J. F. Bader, D. A. Mallory,* D. J.
THE 30-DAY GAME CHANGER: IMPROVING REPRODUCTIVE EFFICIENCY AND PROFITABILITY OF BEEF HERDS. G. Cliff Lamb
 THE 30-DAY GAME CHANGER: IMPROVING REPRODUCTIVE EFFICIENCY AND PROFITABILITY OF BEEF HERDS G. Cliff Lamb North Florida Research and Education Center, Marianna, Florida University of Florida Introduction
THE 30-DAY GAME CHANGER: IMPROVING REPRODUCTIVE EFFICIENCY AND PROFITABILITY OF BEEF HERDS G. Cliff Lamb North Florida Research and Education Center, Marianna, Florida University of Florida Introduction
2α and CIDR+PGF 2α but not the control treat-
 Efficacy of an intravaginal progesterone insert and an injection of PGF 2α for synchronizing estrus and shortening the interval to pregnancy in postpartum beef cows, peripubertal beef heifers, and dairy
Efficacy of an intravaginal progesterone insert and an injection of PGF 2α for synchronizing estrus and shortening the interval to pregnancy in postpartum beef cows, peripubertal beef heifers, and dairy
Proceedings, Applied Reproductive Strategies in Beef Cattle September 1 and 2, 2004, North Platte, Nebraska
 Proceedings, Applied Reproductive Strategies in Beef Cattle September 1 and 2, 2004, North Platte, Nebraska PUBERTY AND ANESTRUS: DEALING WITH NON-CYCLING FEMALES Jeffrey S. Stevenson, Ph.D. Dept. of Animal
Proceedings, Applied Reproductive Strategies in Beef Cattle September 1 and 2, 2004, North Platte, Nebraska PUBERTY AND ANESTRUS: DEALING WITH NON-CYCLING FEMALES Jeffrey S. Stevenson, Ph.D. Dept. of Animal
Fertility Programs to Achieve High 21-d Pregnancy Rates in High-Producing Dairy Herds
 Fertility Programs to Achieve High 21-d Pregnancy Rates in High-Producing Dairy Herds P. M. Fricke, P. D. Carvalho, and J. O. Giordano Department of Dairy Science, University of Wisconsin - Madison 1675
Fertility Programs to Achieve High 21-d Pregnancy Rates in High-Producing Dairy Herds P. M. Fricke, P. D. Carvalho, and J. O. Giordano Department of Dairy Science, University of Wisconsin - Madison 1675
Effects of Early and Late Fall Calving of Beef Cows on Gestation Length and Pregnancy Rate
 Effects of Early and Late Fall Calving of Beef Cows on Gestation Length and Pregnancy Rate D.W. Kastner, F.J. White, I. Rubio, R.P. Wettemann, and D.L. Lalman Story in Brief Effects of early and late fall
Effects of Early and Late Fall Calving of Beef Cows on Gestation Length and Pregnancy Rate D.W. Kastner, F.J. White, I. Rubio, R.P. Wettemann, and D.L. Lalman Story in Brief Effects of early and late fall
Effect of Body Weight Gain and Bovine Somatotropin Treatment on Plasma Concentrations of IGF-I in Postpartum Beef Cows
 Effect of Body Weight Gain and Bovine Somatotropin Treatment on Plasma Concentrations of IGF-I in Postpartum Beef Cows M.J. Prado-Cooper, M.P. Davis, N.M. Long, I. Rubio, R.P. Wettemann, L.J. Spicer Story
Effect of Body Weight Gain and Bovine Somatotropin Treatment on Plasma Concentrations of IGF-I in Postpartum Beef Cows M.J. Prado-Cooper, M.P. Davis, N.M. Long, I. Rubio, R.P. Wettemann, L.J. Spicer Story
THIS ARTICLE IS SPONSORED BY THE MINNESOTA DAIRY HEALTH CONFERENCE.
 THIS ARTICLE IS SPONSORED BY THE MINNESOTA DAIRY HEALTH CONFERENCE. ST. PAUL, MINNESOTA UNITED STATES OF MINNESOTA Comparing the Economics of Reproductive Management Programs on Dairies John Fetrow VMD,
THIS ARTICLE IS SPONSORED BY THE MINNESOTA DAIRY HEALTH CONFERENCE. ST. PAUL, MINNESOTA UNITED STATES OF MINNESOTA Comparing the Economics of Reproductive Management Programs on Dairies John Fetrow VMD,
Washington Animal Agriculture Team
 Washington Animal Agriculture Team Fact Sheet # 12-21 Improving Beef Heifer Reproduction via Synchronization and Timed AI by Dr. Susan Kerr, Dr. Dale Moore, Dr. Ram Kasimanickam, and Sarah Smith Genetic
Washington Animal Agriculture Team Fact Sheet # 12-21 Improving Beef Heifer Reproduction via Synchronization and Timed AI by Dr. Susan Kerr, Dr. Dale Moore, Dr. Ram Kasimanickam, and Sarah Smith Genetic
Estrus and ovulation synchronization in beef heifers
 Estrus and ovulation synchronization in beef heifers Dr. Marcos G Colazo Research Scientist, Alberta Agriculture & Rural Development Edmonton, Alberta, Canada E-mail: marcos.colazo@gov.ab.ca; mgcolazo@yahoo.com
Estrus and ovulation synchronization in beef heifers Dr. Marcos G Colazo Research Scientist, Alberta Agriculture & Rural Development Edmonton, Alberta, Canada E-mail: marcos.colazo@gov.ab.ca; mgcolazo@yahoo.com
Dairy Reproduction Benchmarks. J.W. Smith, W.D. Gilson, L.O. Ely and W.M. Graves Animal and Dairy Science Department
 Dairy Reproduction Benchmarks J.W. Smith, W.D. Gilson, L.O. Ely and W.M. Graves Animal and Dairy Science Department Dairy Reproduction Benchmarks Table of Contents Introduction...3 Methods...3 Figure 1:
Dairy Reproduction Benchmarks J.W. Smith, W.D. Gilson, L.O. Ely and W.M. Graves Animal and Dairy Science Department Dairy Reproduction Benchmarks Table of Contents Introduction...3 Methods...3 Figure 1:
Proceedings, Applied Reproductive Strategies in Beef Cattle October 27 and 28, 2005, Reno, Nevada REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: CIDR
 Proceedings, Applied Reproductive Strategies in Beef Cattle October 27 and 28, 25, Reno, Nevada REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: CIDR G. Cliff Lamb and Jamie E. Larson North Central Research and
Proceedings, Applied Reproductive Strategies in Beef Cattle October 27 and 28, 25, Reno, Nevada REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: CIDR G. Cliff Lamb and Jamie E. Larson North Central Research and
Reproduction management and its economic value
 Reproduction management and its economic value V.E. Cabrera University of Wisconsin-Madison Dairy Science WISCONSIN UNIVERSITY WORKSHOP - PRACTICAL HIGH LEVEL DAIRY MANAGEMENT - A Case Study from a Wisconsin
Reproduction management and its economic value V.E. Cabrera University of Wisconsin-Madison Dairy Science WISCONSIN UNIVERSITY WORKSHOP - PRACTICAL HIGH LEVEL DAIRY MANAGEMENT - A Case Study from a Wisconsin
ARSBC 2016, Des Moines, IA 1
 CONTROL OF ESTRUS AND OVULATION IN BEEF HEIFERS AND COWS The next minutes..! Management considerations related to FT " Heifers and Cows! Overview of protocols " Heifers and Cows! Questions and discussion
CONTROL OF ESTRUS AND OVULATION IN BEEF HEIFERS AND COWS The next minutes..! Management considerations related to FT " Heifers and Cows! Overview of protocols " Heifers and Cows! Questions and discussion
2016 Applied Reproductive Strategies in Beef Cattle Des Moines, Iowa September 7-8,
 2016 Applied Reproductive Strategies in Beef Cattle Des Moines, Iowa September 7-8, 2016 46 Control of estrus and ovulation in beef cows 2 D.J. Patterson, J.M. Thomas, J.M. Abel, B.E. Bishop, J.W.C. Locke,
2016 Applied Reproductive Strategies in Beef Cattle Des Moines, Iowa September 7-8, 2016 46 Control of estrus and ovulation in beef cows 2 D.J. Patterson, J.M. Thomas, J.M. Abel, B.E. Bishop, J.W.C. Locke,
Beef Semen Sales. Since ARSBC, Manhattan, Kan. EVOLUTION AND APPLICATION OF ESTROUS SYNCHRONIZATION PROTOCOLS
 EVOLUTION AND APPLICATION OF ESTROUS SYNCHRONIZATION PROTOCOLS Development of Methods to Synchronize Estrus DECADE 1950 s 60 s PHASE OF DEVELOPMENT PROGESTERONE Inhibited estrus & ovulation; prolonged
EVOLUTION AND APPLICATION OF ESTROUS SYNCHRONIZATION PROTOCOLS Development of Methods to Synchronize Estrus DECADE 1950 s 60 s PHASE OF DEVELOPMENT PROGESTERONE Inhibited estrus & ovulation; prolonged
2018 Protocols for Synchronization of Estrus and Ovulation in Beef Cows and Heifers
 2018 Protocols for Synchronization of Estrus and Ovulation in Beef Cows and Heifers Biotechnology presents beef producers with an unprecedented opportunity to improve herd genetics. Producers have more
2018 Protocols for Synchronization of Estrus and Ovulation in Beef Cows and Heifers Biotechnology presents beef producers with an unprecedented opportunity to improve herd genetics. Producers have more
Fertility Factors. Factors that influence fertility in natural and synchronized breeding programs.
 Fertility Factors Factors that influence fertility in natural and synchronized breeding programs. Reproductive failure is a major source of economic loss in the beef industry. The majority of this loss
Fertility Factors Factors that influence fertility in natural and synchronized breeding programs. Reproductive failure is a major source of economic loss in the beef industry. The majority of this loss
Efficiency Comparison between Two Systematic Breeding Protocols: Target Breeding and Presynchronization Program At a Large Central Valley Dairy Farm
 Efficiency Comparison between Two Systematic Breeding Protocols: Target Breeding and Presynchronization Program At a Large Central Valley Dairy Farm A Senior Project Presented to The Faculty of the Dairy
Efficiency Comparison between Two Systematic Breeding Protocols: Target Breeding and Presynchronization Program At a Large Central Valley Dairy Farm A Senior Project Presented to The Faculty of the Dairy
ALTERED INSEMINATION TIMING IMPROVES PREGNANCY RATES AFTER A CO-SYNCH + PROGESTERONE INSERT PROTOCOL CASEY DOBBINS
 ALTERED INSEMINATION TIMING IMPROVES PREGNANCY RATES AFTER A CO-SYNCH + PROGESTERONE INSERT PROTOCOL by CASEY DOBBINS B.S., Kansas State University, 2004 A REPORT submitted in partial fulfillment of the
ALTERED INSEMINATION TIMING IMPROVES PREGNANCY RATES AFTER A CO-SYNCH + PROGESTERONE INSERT PROTOCOL by CASEY DOBBINS B.S., Kansas State University, 2004 A REPORT submitted in partial fulfillment of the
An economic decision-making support system for selection of reproductive management programs on dairy farms
 J. Dairy ci. 94 :6216 6232 doi: 10.3168/jds.2011-4376 American Dairy cience Association, 2011. An economic decision-making support system for selection of reproductive management programs on dairy farms
J. Dairy ci. 94 :6216 6232 doi: 10.3168/jds.2011-4376 American Dairy cience Association, 2011. An economic decision-making support system for selection of reproductive management programs on dairy farms
Factors To Improve Reproductive Management and Getting Cows Pregnant
 Factors To Improve Reproductive Management and Getting Cows Pregnant Jeffrey S. Stevenson, Ph.D. Dept. of Animal Sciences and Industry e-mail: jss@ksu.edu Kansas State University, Manhattan Introduction
Factors To Improve Reproductive Management and Getting Cows Pregnant Jeffrey S. Stevenson, Ph.D. Dept. of Animal Sciences and Industry e-mail: jss@ksu.edu Kansas State University, Manhattan Introduction
Nutritional Implications of Altering the Dry Period Length
 Nutritional Implications of Altering the Dry Period Length Ric R. Grummer 1 Ruminant Technical Manager, Balchem Corp. Emeritus Professor, University of Wisconsin-Madison Introduction Traditionally, the
Nutritional Implications of Altering the Dry Period Length Ric R. Grummer 1 Ruminant Technical Manager, Balchem Corp. Emeritus Professor, University of Wisconsin-Madison Introduction Traditionally, the
REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: MGA, 1
 Proceedings, Applied Reproductive Strategies in Beef Cattle November 12 and 13, 2005, Texas A&M University, College Station REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: MGA, 1 D. J. Patterson, D. J. Schafer,
Proceedings, Applied Reproductive Strategies in Beef Cattle November 12 and 13, 2005, Texas A&M University, College Station REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: MGA, 1 D. J. Patterson, D. J. Schafer,
CONTROL OF ESTRUS AND OVULATION IN BEEF COWS 1. D.J. Patterson, J.M. Thomas, J.W.C. Locke, E.R. Knickmeyer, R.C. Bonacker, and M.F.
 CONTROL OF ESTRUS AND OVULATION IN BEEF COWS 1 D.J. Patterson, J.M. Thomas, J.W.C. Locke, E.R. Knickmeyer, R.C. Bonacker, and M.F. Smith Division of Animal Sciences University of Missouri, Columbia, MO
CONTROL OF ESTRUS AND OVULATION IN BEEF COWS 1 D.J. Patterson, J.M. Thomas, J.W.C. Locke, E.R. Knickmeyer, R.C. Bonacker, and M.F. Smith Division of Animal Sciences University of Missouri, Columbia, MO
What is New in Dairy Reproduction?
 What is New in Dairy Reproduction? Carlos A. Risco, DVM, Dipl. ACT 1 College of Veterinary Medicine University of Florida, Gainesville Has the reproductive performance in your herd improved, remained the
What is New in Dairy Reproduction? Carlos A. Risco, DVM, Dipl. ACT 1 College of Veterinary Medicine University of Florida, Gainesville Has the reproductive performance in your herd improved, remained the
FERTILITY AFTER TIMED ARTIFICAL INSEMINATION IN RESPONSE TO A CONTROLLED INTERNAL DRUG RELEASE (CIDR) INSERT IN LACTATING DAIRY COWS
 INSERT IN LACTATING DAIRY COWS") FERTILITY AFTER TIMED ARTIFICAL INSEMINATION IN RESPONSE TO A CONTROLLED INTERNAL DRUG RELEASE (CIDR) INSERT IN LACTATING DAIRY COWS by CYNTHIA ANN MARTEL B. S., University of New Hampshire, Durham, 2004
FERTILITY AFTER TIMED ARTIFICAL INSEMINATION IN RESPONSE TO A CONTROLLED INTERNAL DRUG RELEASE (CIDR) INSERT IN LACTATING DAIRY COWS by CYNTHIA ANN MARTEL B. S., University of New Hampshire, Durham, 2004
ESTRUS SYNCHRONIZATION: A REPRODUCTIVE MANAGEMENT TOOL by Mel DeJarnette, Reproduction Specialist
 ESTRUS SYNCHRONIZATION: A REPRODUCTIVE MANAGEMENT TOOL by Mel DeJarnette, Reproduction Specialist Historically, estrus synchronization has been promoted as a labor saving tool for those producers who want
ESTRUS SYNCHRONIZATION: A REPRODUCTIVE MANAGEMENT TOOL by Mel DeJarnette, Reproduction Specialist Historically, estrus synchronization has been promoted as a labor saving tool for those producers who want
Effect of CIDR 12 to 19 Days After AI on Detection of Returning Estrus and Conception Rate in Dairy Cows
 Journal of Reproduction and Development, Vol. 56, No. 2, 2010, 09-134T Original Article Effect of CIDR 12 to 19 Days After AI on Detection of Returning Estrus and Conception Rate in Dairy Cows Su T. LONG
Journal of Reproduction and Development, Vol. 56, No. 2, 2010, 09-134T Original Article Effect of CIDR 12 to 19 Days After AI on Detection of Returning Estrus and Conception Rate in Dairy Cows Su T. LONG
The Livestock Event. Using breeding protocols and fertility visits to improve pregnancy rates in beef and dairy herds
 The Livestock Event Using breeding protocols and fertility visits to improve pregnancy rates in beef and dairy herds STUART ROGERS, LONGMOOR FARM PARTNERS www.longmoorfarm.com 240 cows, AYR calving, supplying
The Livestock Event Using breeding protocols and fertility visits to improve pregnancy rates in beef and dairy herds STUART ROGERS, LONGMOOR FARM PARTNERS www.longmoorfarm.com 240 cows, AYR calving, supplying
Grazing Wheat Did Not Reduce Beef Cow Pregnancy Rates
 Grazing Wheat Did Not Reduce Beef Cow Pregnancy Rates S.K. Johnson and K. Harmoney Introduction Beef producers can lower feed costs by extending the grazing period and reducing the need for harvested forages.
Grazing Wheat Did Not Reduce Beef Cow Pregnancy Rates S.K. Johnson and K. Harmoney Introduction Beef producers can lower feed costs by extending the grazing period and reducing the need for harvested forages.
USING CONTROLLED INTERNAL DRUG RELEASE (CIDR ) INSERTS FOR ESTRUS SYNCHRONIZATION IN DAIRY HEIFERS A.K. MCLEAN
 INSERTS FOR ESTRUS SYNCHRONIZATION IN DAIRY HEIFERS A.K. MCLEAN") USING CONTROLLED INTERNAL DRUG RELEASE (CIDR ) INSERTS FOR ESTRUS SYNCHRONIZATION IN DAIRY HEIFERS by A.K. MCLEAN Under the Direction of William M. Graves ABSTRACT In Experiment 1, 164 heifers were assigned
USING CONTROLLED INTERNAL DRUG RELEASE (CIDR ) INSERTS FOR ESTRUS SYNCHRONIZATION IN DAIRY HEIFERS by A.K. MCLEAN Under the Direction of William M. Graves ABSTRACT In Experiment 1, 164 heifers were assigned
Economics of Improving Reproductive Performance in Dairy Herds
 Economics of Improving Reproductive Performance in Dairy Herds Stephen LeBlanc Population Medicine, Ontario Veterinary College, University of Guelph N1G 2W1 Email: sleblanc@uoguelph.ca Take Home Messages
Economics of Improving Reproductive Performance in Dairy Herds Stephen LeBlanc Population Medicine, Ontario Veterinary College, University of Guelph N1G 2W1 Email: sleblanc@uoguelph.ca Take Home Messages
Dietary grouping strategies to improve profitability on dairy farms Introduction Materials and methods Simulation framework
 Dietary grouping strategies to improve profitability on dairy farms Victor E. Cabrera and Afshin Kalantari 1675 Observatory Dr., Madison, WI 53706, vcabrera@wisc.edu University of Wisconsin Madison, Department
Dietary grouping strategies to improve profitability on dairy farms Victor E. Cabrera and Afshin Kalantari 1675 Observatory Dr., Madison, WI 53706, vcabrera@wisc.edu University of Wisconsin Madison, Department
MSU Extension Publication Archive. Scroll down to view the publication.
 MSU Extension Publication Archive Archive copy of publication, do not use for current recommendations. Up-to-date information about many topics can be obtained from your local Extension office. Estrus
MSU Extension Publication Archive Archive copy of publication, do not use for current recommendations. Up-to-date information about many topics can be obtained from your local Extension office. Estrus
Recent studies on nutritional factors affecting reproductive efficiency in U.S. dairy herds.
 Recent studies on nutritional factors affecting reproductive efficiency in U.S. dairy herds. Milo C. Wiltbank, Kent A. Weigel, Daniel Z. Caraviello Department of Dairy Science, 1685 Observatory Drive University
Recent studies on nutritional factors affecting reproductive efficiency in U.S. dairy herds. Milo C. Wiltbank, Kent A. Weigel, Daniel Z. Caraviello Department of Dairy Science, 1685 Observatory Drive University
Outline. Heifers selection. Selection. Maternal Traits. Range Beef Cow Symposium, Dec. 3-5, /5/13. rangebeefcow.com 1
 Range Beef Cow Symposium, Dec. 3-5, 213 12/5/13 Outline POST-AI NUTRITIONAL IMPACT ON REPRODUCTION Range Beef Cow Symposium Rapid City, SD 213 Introduction Importance of Reproduction Heifer selection/management
Range Beef Cow Symposium, Dec. 3-5, 213 12/5/13 Outline POST-AI NUTRITIONAL IMPACT ON REPRODUCTION Range Beef Cow Symposium Rapid City, SD 213 Introduction Importance of Reproduction Heifer selection/management
Getting Large Numbers of Dairy Heifers Bred AI
 Getting Large Numbers of Dairy Heifers Bred AI Lessons from the Show-Me Select Program David J. Patterson Division of Animal Sciences University of Missouri - Columbia Reproductive Management Resource
Getting Large Numbers of Dairy Heifers Bred AI Lessons from the Show-Me Select Program David J. Patterson Division of Animal Sciences University of Missouri - Columbia Reproductive Management Resource
Reproductive Efficiency and Economics of Timed AI vs. Natural Service
 Reproductive Efficiency and Economics of Timed AI vs. Natural Service Carlos A. Risco 1, Fabio Lima 1, Marie-Joelle Thatcher 1, Jose Santos 2, Albert De Vries 2 and William W. Thatcher 2 University of
Reproductive Efficiency and Economics of Timed AI vs. Natural Service Carlos A. Risco 1, Fabio Lima 1, Marie-Joelle Thatcher 1, Jose Santos 2, Albert De Vries 2 and William W. Thatcher 2 University of
Proceedings, Applied Reproductive Strategies in Beef Cattle September 1 and 2, 2004, North Platte, Nebraska
 Proceedings, Applied Reproductive Strategies in Beef Cattle September 1 and 2, 2004, North Platte, Nebraska REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: MGA D. J. Patterson, F. N. Kojima, J. E. Stegner, J.
Proceedings, Applied Reproductive Strategies in Beef Cattle September 1 and 2, 2004, North Platte, Nebraska REVIEW OF ESTRUS SYNCHRONIZATION SYSTEMS: MGA D. J. Patterson, F. N. Kojima, J. E. Stegner, J.
REVIEW OF ESTROUS SYNCRHONIZATION SYSTEMS: CIDR INSERTS
 Proceedings, The Applied Reproductive Strategies in Beef Cattle Workshop, September 5-6, 2002, Manhattan, Kansas REVIEW OF ESTROUS SYNCRHONIZATION SYSTEMS: CIDR INSERTS Darrel J. Kesler, Ph.D. 1 Professor
Proceedings, The Applied Reproductive Strategies in Beef Cattle Workshop, September 5-6, 2002, Manhattan, Kansas REVIEW OF ESTROUS SYNCRHONIZATION SYSTEMS: CIDR INSERTS Darrel J. Kesler, Ph.D. 1 Professor
Reproductive Responses of Lactating Dairy Cows According to Estrus Behavior and CIDR Uses in a Heatsynch Protocol
 Acta Scientiae Veterinariae, 205. 43: 256. RESEARCH ARTICLE Pub. 256 ISSN 679-926 Reproductive Responses of Lactating Dairy Cows According to Estrus Behavior and CIDR Uses in a Heatsynch Protocol Márcio
Acta Scientiae Veterinariae, 205. 43: 256. RESEARCH ARTICLE Pub. 256 ISSN 679-926 Reproductive Responses of Lactating Dairy Cows According to Estrus Behavior and CIDR Uses in a Heatsynch Protocol Márcio
CONTROL OF ESTRUS AND OVULATION IN HEIFERS 1. D.J. Patterson, J.M. Thomas, and M.F. Smith
 CONTROL OF ESTRUS AND OVULATION IN HEIFERS 1 D.J. Patterson, J.M. Thomas, and M.F. Smith Division of Animal Sciences University of Missouri, Columbia, MO Introduction Estrus synchronization and artificial
CONTROL OF ESTRUS AND OVULATION IN HEIFERS 1 D.J. Patterson, J.M. Thomas, and M.F. Smith Division of Animal Sciences University of Missouri, Columbia, MO Introduction Estrus synchronization and artificial
Proceedings, Applied Reproductive Strategies in Beef Cattle September 11 and 12, 2007, Billings, Montana ESTRUS SYNCHRONIZATION PROTOCOLS FOR COWS
 Proceedings, Applied Reproductive Strategies in Beef Cattle September 11 and 12, 2007, Billings, Montana ESTRUS SYNCHRONIZATION PROTOCOLS FOR COWS G. Cliff Lamb, Jamie E. Larson, and Carl R. Dahlen North
Proceedings, Applied Reproductive Strategies in Beef Cattle September 11 and 12, 2007, Billings, Montana ESTRUS SYNCHRONIZATION PROTOCOLS FOR COWS G. Cliff Lamb, Jamie E. Larson, and Carl R. Dahlen North
COMPARISON OF BREEDING SYSTEM COSTS FOR ESTRUS-SYNCHRONIZATION PROTOCOLS PLUS ARTIFICIAL INSEMINATION VERSUS NATURAL SERVICE
 Cattlemen s Day 2003 COMPARISON OF BREEDING SYSTEM COSTS FOR ESTRUS-SYNCHRONIZATION PROTOCOLS PLUS ARTIFICIAL INSEMINATION VERSUS NATURAL SERVICE S. K. Johnson, S. L. Fogleman, and R. Jones Summary Breeding
Cattlemen s Day 2003 COMPARISON OF BREEDING SYSTEM COSTS FOR ESTRUS-SYNCHRONIZATION PROTOCOLS PLUS ARTIFICIAL INSEMINATION VERSUS NATURAL SERVICE S. K. Johnson, S. L. Fogleman, and R. Jones Summary Breeding
Using Estrous Synchronization in Natural-Service Breeding Situations 2.3
 livestock igrow.org SEPTEMBER 2016 SDSU DEPARTMENT OF ANIMAL SCIENCE Using Estrous Synchronization in Natural-Service Breeding Situations Taylor Grussing SDSU Extension Cow/Calf Field Specialist George
livestock igrow.org SEPTEMBER 2016 SDSU DEPARTMENT OF ANIMAL SCIENCE Using Estrous Synchronization in Natural-Service Breeding Situations Taylor Grussing SDSU Extension Cow/Calf Field Specialist George
Control of estrus and ovulation in beef heifers 1
 2016 Applied Reproductive Strategies in Beef Cattle Des Moines, Iowa September 7-8, 2016 15 Control of estrus and ovulation in beef heifers 1 D.J. Patterson, J.M. Thomas, B.E. Bishop, J.M. Abel, J.W.C.
2016 Applied Reproductive Strategies in Beef Cattle Des Moines, Iowa September 7-8, 2016 15 Control of estrus and ovulation in beef heifers 1 D.J. Patterson, J.M. Thomas, B.E. Bishop, J.M. Abel, J.W.C.
DAIRY RESEARCH. Report of Progress Kansas State University Agricultural Experiment Station and Cooperative Extension Service
 DAIRY RESEARCH 2008 Report of Progress 1002 Kansas State University Agricultural Experiment Station and Cooperative Extension Service DAIRY DAYS PROGRAM Effects of alfalfa inclusion rate in diets based
DAIRY RESEARCH 2008 Report of Progress 1002 Kansas State University Agricultural Experiment Station and Cooperative Extension Service DAIRY DAYS PROGRAM Effects of alfalfa inclusion rate in diets based
Management of Heat Stress to Improve Fertility in Dairy Cows in Israel
 5 10 15 Management of Heat Stress to Improve Fertility in Dairy Cows in Israel Israel Flamenbaum ¹ and Nadav Galon ² ¹ Advanced Cow Cooling Systems Co., Tel Aviv, Israel ² - Hachaklait Veterinary Services
5 10 15 Management of Heat Stress to Improve Fertility in Dairy Cows in Israel Israel Flamenbaum ¹ and Nadav Galon ² ¹ Advanced Cow Cooling Systems Co., Tel Aviv, Israel ² - Hachaklait Veterinary Services
Proceedings, Applied Reproductive Strategies in Beef Cattle December 2 and 3, 2008, Fort Collins, CO ESTRUS SYNCHRONIZATION PROTOCOLS FOR HEIFERS, 1
 Proceedings, Applied Reproductive Strategies in Beef Cattle December 2 and 3, 28, Fort Collins, CO ESTRUS SYNCHRONIZATION PROTOCOLS FOR HEIFERS, 1 D.J. Patterson, D.C. Busch, N.R. Leitman, D.J. Wilson,
Proceedings, Applied Reproductive Strategies in Beef Cattle December 2 and 3, 28, Fort Collins, CO ESTRUS SYNCHRONIZATION PROTOCOLS FOR HEIFERS, 1 D.J. Patterson, D.C. Busch, N.R. Leitman, D.J. Wilson,
Managing Reproduction Les Anderson
 SECTION 4 Managing Reproduction Les Anderson Reproductive efficiency is the single most important factor affecting gross returns in a cow-calf operation. Even though reproductive efficiency determines
SECTION 4 Managing Reproduction Les Anderson Reproductive efficiency is the single most important factor affecting gross returns in a cow-calf operation. Even though reproductive efficiency determines