[35S] methionine, by the various types of mucous cell.
|
|
|
- Tracey King
- 5 years ago
- Views:
Transcription
 MuciNs are produced from three morphologically distinct types of cell in the mammalian stomach, viz.")
1 AUTORADIOGRAPHIC OBSERVATIONS ON THE MUCOUS CELLS OF THE STOMACH AND INTESTINE. By M. A. JENNINGS and H. W. FLOREY. From the Sir William Dunn School of Pathology, Oxford. (Received for publication 10th December 1955) MuciNs are produced from three morphologically distinct types of cell in the mammalian stomach, viz. the superficial epithelium which forms a continuous covering, the mucoid neck cells of the fundal glands, and the pyloric glands. Possibly the cardiac glands, which consist of mucus-producing cells, and are a conspicuous feature in some species, represent a fourth type of mucous cell. The duodenal or Brunner's glands are composed of mucous cells, although in some species, for instance the rabbit and horse, "serous" cells are present in addition. Goblet cells, which produce mucin, are found throughout the small and large intestines. In the small intestine they occur both in the crypts of Lieberkuhn and on the villi. The colon contains large numbers, most closely packed in the crypts but also scattered among the surface epithelium. They are relatively scarce in the csecum of the rabbit and guinea-pig, where they are found principally at the bases of the shallow crypts. Some of the mucins secreted by these various cells contain sulphur, probably in the form of ester sulphate groups. The present experiments have been concerned with observations on the incorporation and excretion of 35S, administered intravenously either as Na235SO4 or as [35S] methionine, by the various types of mucous cell. EXPERIMENTAL Comparative observations were made on mice, rats, guinea-pigs, rabbits and cats. Material was obtained from 6 mice, killed 1, 21, 3, 6, 12 and 24 hrs. respectively after the injection of radioactive sodium sulphate; from 9 rats killed 1 hr. 10 min., 2 hrs. (2 animals), 3 hrs. (4 animals) and 24 hrs. (2 animals) after sulphate; from 3 rabbits killed at 21, 61 and 24 hrs.; 2 guinea-pigs killed at 2 and 61 hrs.; and 2 cats killed at 21 and 6 hrs. after sulphate. Two of the mice (killed at 1 and 21 hrs.) and two of the rats (killed at 2 and 24 hrs.) had dilute mustard oil inserted into a loop of the colon for 4 hours to empty and stimulate the goblet cells, as previously described [Jennings and Florey, 1954]; the oil was removed immediately before sulphate was given. 131

2 132 Jennings and Florey Most of the animals were fed in the usual way before and during experiments. The 4 rats killed 3 hours after the administration of sulphate had been starved for 24 hours; 3 were fed respectively 3 hours and 1 hour before and at the time of giving sulphate; the fourth was starved throughout. In order to ensure a state of active digestion the cats were starved for 20 hours and then given food about 2 hours before the administration of sulphate. Two mice received methionine labelled with 35S and were killed 1 and 2j hours later respectively. These had both had mustard oil applied to a loop of the colon for 4 hours and removed just before the methionine was given. The weights of the animals used were: mice, g.; rats, g.; guinea-pigs, 200 g.; rabbits, 1P5 kg.; cats, 1-4 and 16 kg. At the end of experiment tissues were fixed and histological sections were prepared. Sections were so processed as to wash out any 35S present in the form of free sulphate, and were then stained by the periodic acid-schiff reaction and sometimes other stains, and were covered with photographic emulsion for the production of autoradiographs. The histological and autoradiographic techniques are described by Heatley et al. [1956]. The metachromatic reaction was investigated on ordinary sections of the same material by staining for 1 minute in 0-1 per cent toluidine blue and examining wet under the microscope. Radioactive Materials.-The radioactive materials 1 used were carrier-free Na235SO4 and DL-[35S] methionine (28 mc/m.mole) in aqueous solution. The dosage of sulphate was based on that recommended for mice by Dr. S. R. Pelc in a personal communication, and was approximately 10 mc per kg. of body weight. The dose of methionine was about half this amount. All injections were made intravenously. Interpretation of the Autoradiographs.-In an autoradiograph the position of concentrated radioactivity due to 35S in the tissues at the time the animal is killed is shown by a deposit of black particles of silver in the photographic emulsion over the area, with some scattering around the site of maximum concentration. In spite of the scattering, the elevation of the particles above the section, the partial obliteration of the section where the particles are most numerous, and the fact that full histological staining tends to obscure the particles, it is still possible to locate the main sites of radioactivity quite accurately at the cellular level. This depends on judicious staining of the section before applying the emulsion, and may be helped if there are available several autoradiographs, exposed for different lengths of time, of each tissue. Comparison with ordinary sections of the same material is occasionally helpful. The preparations under discussion might be expected to throw some light on three different aspects of the activity of mucous cells. (1) The sites where the mucus has a relatively high sulphate content might be located. This information might be correlated with histological staining reactions of the same sites, and with any chemical information available about the secretion from the area. (2) Some information about physiological activity of the cells might be obtained by comparison of preparations from animals killed at different intervals after injection of the isotope, or under different experimental conditions. 1 Obtained from the Radiochemical Centre, Amersham, Bucks.

3 Autoradiographs of Mucous Cells (3) Information about movement of labelled cells themselves along the surface of the mucosa might be forthcoming from the same comparison. For the sake of clarity these three aspects will be considered separately. RESULTS AFTER INJECTION OF Na235SO4 133 There was a striking concentration of radioactivity in certain of the mucusproducing epithelial cells. With one exception to be mentioned later, the strongest radioactivity at every site was seen in the earliest specimens examined, that is, from 1 to 3 hours after the injection of sulphate. The distribution of radioactivity will be described as it was seen at that time. RADIOACTIVITY IN MUCOIUS CELLS OF THE STOMACH AND BRUNNER'S GLAN;DS The various kinds of mucous cell in the stomach and duodenal glands varied widely in their capacity to take up Na235SO4, and the distribution of radioactivity was different in different species. Surface Epithelium of the Stomach.-The mucin in the continuous superficial epithelium which lines the stomach showed no radioactivity in any of the five species examined. Illustrations of this can be seen in figs. 1, 2, 6, 8, 9, 10, 11 and 16. Foveolar and Mucoid Neck Cells.-In all parts of the stomach the gastric pits or foveole are lined by a continuation of the surface epithelium, the cells decreasing in height and in mucin content as the pit is descended. In the fundal glands the mucoid neck cells occupy the upper parts of the glands, adjacent to the foveolse. They vary a good deal in number and position from one part of the fundus to another and from species to species. In most species they are quite distinct morphologically from the foveolar cells. Rat and Mouse.-Strong radioactivity was present in the lower parts of all the foveolwe in the rat. Mice showed the same distribution but the amount of radioactivity was less in some of the animals. The appearance is illustrated in fig. 6, which shows that radioactive mucin is being secreted in the bases of the foveolae and is emerging from the mouths of the foveolie into the cavity of the stomach. Fig. 7 shows that the main concentration of radioactivity was undoubtedly in the foveolh and not in the mucoid neck cells as has been suggested (see Discussion). The dividing line was not absolute, however, for here and there in the rat a cell adjacent to a foveola which was morphologically a mucoid neck cell showed slight activity. Cat.-In the cat the foveolae did not show any activity in the fundal region, but in the pylorus there was some in the deepest parts, adjoining the active pyloric glands (owing to oblique cutting this region is not well shown in fig. 11). The mucoid neck cells showed no radioactivity. Surprisingly, some appeared in the peptic cells in this species (fig. 8). Rabbit and Guinea-Pig.-The rabbit and guinea-pig (fig. 16) showed radioactivity in the foveole, but it was variable and less intense than in the rat, and in the rabbit seemed to be confined to the foveolae of the fundal glands close to the cardia. In the rabbit the small foveolar cells with dark-staining mucin and the larger triangular mucoid neck cells are quite distinct, and it was clear that there was radioactivity not only in the deepest foveolar cells but also in

4 134 Jennings and Florey the highest mucoid neck cells, though the main body of mucoid neck cells lying deeper in the glands showed none. In the guinea-pig the distribution of radioactivity was probably the same, but in this animal the mucoid neck cells are small and compact and look so much like the smaller foveolar cells that it was only possible to surmise from the position of the radioactivity that some of them were concerned. Cardiac Glands.-These mucous glands are extensive in some animals, e.g. the pig, but in all the species used in this work they form a trivial bunch of tubules around the cardiac orifice (or next to the squamous part of the stomach in the rat and mouse); not all sections through these regions included cardiac glands. In the cat and rabbit the cardiac glands showed radioactivity while in the guinea-pig (fig. 16) and rat they did not. In the mouse they were probably active, but too few were seen to make a firm conclusion possible. Pyloric Glands.-These mucous glands occupy a considerable area between the fundal glands and the pyloric ring. Rat, Mouse, Cat and Rabbit.-These four species all showed radioactivity in the pyloric glands. In figs. 1, 9 and 10 this region of the rat is illustrated, and the flow of mucus from glands and foveolh is seen passing on to the surface of the mucosa. Figs. 11 and 14 show the pyloric glands of the cat. Guinea-Pig.-In the guinea-pig the main body of pyloric glands was inactive (figs. 2 and 13). Brunner's glands showed strong radioactivity, and a few pyloric alveoli next to these also showed radioactivity (fig. 2). Duodenal Glands.-The duodenal or Brunner's glands begin at the pyloric ring. They lie beneath the muscularis mucosse and extend along the duodenum for a distance that differs from species to species. As will be mentioned later, it has been argued that they are an extension of the pyloric glands into the duodenum. In all the species examined these glands showed some difference from the pyloric glands in their capacity to take up 35SO42-. Rat and Mouse.-Brunner's glands in the rat (fig. 1) and mouse were inactive, except that in one of the mice examined a few of the alveoli near the pyloric ring showed some activity. The lack of radioactivity in the Brunner's gland mass contrasted sharply with the activity of the pyloric glands and goblet cells (fig. 1). Cat.-The Brunner's glands in the cat showed no radioactivity (figs. 3 and 4). Rabbit and Guinea-Pig.-In the guinea-pig (figs. 2 and 5) there was strong and in the rabbit rather weak radioactivity in some of the Brunner's gland alveoli, and a lesser amount of activity in others. In both these species the Brunner's glands extend for many inches along the duodenum between the muscularis mucosse and the main muscle layers. In the guinea-pig the most strongly radioactive alveoli were the deep alveoli next to the main muscle layer, while in the rabbit the superficial alveoli were the most active. MOVEMENT OF RADIOACTIVE MATERIAL THROUGH THE CELLS OF STOMACH AND BRUNNER'S GLANDS The appearance of "incorporated" 35S in the cells began soon after the injection of Na235SO4 (e.g. fig. 7). As far as the few animals injected showed, the amount of radioactivity in the various sites was maximal by about an hour after injection and changed little up to 3 to 6 hours after injection, but in the

5 Autoradiographs of Mucous Cells 135 next few hours fell rapidly. In the specimens taken 12 or 24 hours after injection there was usually little activity. The cells which showed weakest activity at the start lost their activity first. These generalizations apply to all the species that were examined. In the pyloric glands of the cat, and to a lesser extent in the Brunner's glands of the rabbit, radioactivity was concentrated near the deeper part of the mucin mass in each cell, i.e. in the supranuclear region, in the first specimens seen (fig. 11) and only in later specimens appeared spread diffusely through the mucin (fig. 14). In all other sites and species radioactivity was centralized over the mucin masses in the earliest specimens seen. The discharge of radioactive mucin from the cells seemed to begin soon after 35S had entered them. It was not particularly evident in specimens taken about an hour after injection, but in those taken at 2 to 3 hours radioactive mucin was often present in the lumina of glands and foveola3 and on the surface of the mucosa (e.g. fig. 16). This was true even of the pyloric glands of the cat at a time when radioactivity was still concentrated in the deeper part of the mucin mass. The cells in the bases of the foveolh of the rat showed remarkable productive activity. These cells contain only a small mass of stainable mucin, yet their radioactivity, which was centred over the mucin rather than over the cytoplasm, was intense, and highly radioactive mucin was entering the cavity of the stomach from the foveolhe. This was seen in both fundal and pyloric regions (figs. 1 and 6). Synthesis and discharge of mucin apparently go on together. From the present experiments there is no evidence that periods of discharge alternate with periods of rebuilding under physiological conditions. It appears that labelled mucin begins to be discharged soon after labelled material has entered the cell, though the last remnants of activity may not leave the cell for many hours. The fact that an apparently fairly steady level of radioactivity may be maintained in the cell for the first few hours, even though the level of radioactivity in the blood is falling steeply [Dziewiatkowski, 1949], supports the idea of an intracellular storage mechanism for SO42-, which has been suggested on chemical grounds [Kent, Whitehouse, Jennings and Florey, 1956]. Some such mechanism would seem to be particularly necessary for such cells as those of the foveolae of the rat, since they appear to maintain a secretion of radioactive mucin for at least 2 or 3 hours, though having a very small content of stainable mucin at any one time. Feeding and Starvation.-A comparison was made of radioactivity in fed and starved rats killed 3 hours after sulphate injection. In starvation there was a greater amount of radioactivity, but the distribution was the same in both, and in both radioactive mucin was passing from the foveolee into the cavity of the stomach (figs. 6 and 9). It seems that in both starvation and digestion the gastric cells manufacture mucin and, at least in the foveole, discharge it. The greater concentration of radioactivity in starvation may be due to more active uptake of S042- or, more probably, to a less vigorous stimulus to discharge from the cells. MOVEMENT OF CELLS IN THE GASTRIC MUCOSA It was of interest to see whether labelling of the contents of the cells with 35S would contribute any information about movement of the cells themselves.
. In rats killed 24 hours after the injection of sulphate some radioactivity could be seen at three sites in the fundus of the stomach.")
6 136 Jennings and Florey It is held that the gastric surface cells are entirely replaced by the sliding forward of cells from the bases of the foveolae and, with less certainty, that mucoid neck cells are replaced from the same region (see Discussion). In rats killed 24 hours after the injection of sulphate some radioactivity could be seen at three sites in the fundus of the stomach. There were welldefined though scattered areas high in the foveolae (fig. 12); other similar though less numerous patches over the mucoid neck cells; and traces of radioactivity at the bases of the foveolao, in the original position. The activity in the first and second positions, which appeared only at a late stage after sulphate injection, might represent groups of labelled foveolar cells that had travelled a certain distance in 24 hours as mitosis had proceeded in the bases of the foveole. The trace of activity at the original site might be attributed to residual activity, diluted by cell division and secretion, in cells that had not moved. No other specimens from stomach or duodenum provided any evidence of cell movement. RADIOACTIVITY IN GOBLET CELLS In all the species examined the goblet cells throughout the small and large intestines took up the radioactive material. Small Intestine. Rat, Mouse, Cat.-These species all showed a similar distribution of activity. In the earlier specimens examined the goblet cells on the villi had concentrated their content of 35S in the supranuclear region, so that in the autoradiograph the reduced silver formed a cap or small mass over or near the deeper end of the mucin in each cell (figs. 17, 20, 21, 22). As time went on the activity became spread over the whole mucin mass (fig. 18) and was seen in mucin being discharged from the cell. In the mouse there was little or no activity left in the cells on the villi at 12 hours (fig. 19) and none at 24 hours, but in the rat the loss of activity was somewhat slower. The goblet cells on the villi were thus slower in distributing radioactivity through the intracellular mucin than were most of the gastric cells. The goblet cells of the crypts, on the other hand, often showed diffuse radioactivity over the mucin in the first specimens seen, though the amount was possibly less than in the cells of the villi. This might be considered as evidence of greater metabolic activity in the cells of the crypts. The same suggestion could account for the fact that in the rat activity disappeared from the cells of the crypts before it disappeared from those on the villi. In the mouse there was still slight activity in the crypts in the last specimen showing activity on the villi, but it was diffusely spread over the cells, and did not seem to correspond with the position of the few goblet cells present in this situation. Possibly it may have been in the granules of the Paneth cells, which are abundant in the crypts of Lieberkiihn in the mouse. In most of the early specimens there was already radioactivity in the free mucin between the villi (fig. 22) and in the lumen of the intestine, but it was not usually so intense as that in some other situations (e.g. the mucin coming from the gastric foveole of the rat). Fig. 22 shows radioactive mucin between two villi whose goblet cells show activity mainly in the supranuclear region. By analogy with what was seen in the cat pylorus, it is not impossible that the radioactive free mucin might have

7 Autoradiographs of Mucous Cells 137 come from the adjacent goblet cells in spite of the deep situation of their concentrated radioactivity. It could, however, have originated from the cells in the crypts where activity was already generalized through the mucin at this time Ġuinea-Pig.-The guinea-pig showed intense radioactivity in the goblets of the small intestine. Two hours after the injection of sulphate the goblets on the villi had the activity over the deeper half of the mucin mass, while those of the crypts had diffuse or centrally placed activity. There was radioactivity over all the surfaces of the villi, corresponding with a thin, evenly spread layer of mucin. The vigour with which radioactive material was taken up by the cells of the guinea-pig had a counterpart in an apparently rapid rate of loss. In the animal killed 6j hours after sulphate injection there was much less activity than at 2 hours. The activity lay over the centres of the intracellular mucin masses and along the surfaces of the vifli (fig. 23). There was in this species no evidence of earlier loss of activity from the cells of the crypts than from those of the villi; if anything, more activity remained in the crypts than on the villi 6j hours after injection. Rabbit.-The rabbit showed a more complex picture than the other species. The distribution of radioactivity was similar at 2j and 61 hours after sulphate injection. The position of concentrated activity in the goblet cells varied a good deal. On the whole, it was centred over the mucin masses on the upper parts of the villi and in the crypts, but was concentrated at the deeper ends of the mucin masses in the middle zone of the mucosa. At this time there was more radioactivity in the cells of the villi than in those of the crypts. In the rabbit killed 24 hours after sulphate injection, the cells on the villi had lost some of their radioactivity, but most of the goblet cells towards the bases of the crypts still showed substantial activity. Movement of Mucin and of Cells in the Small Intestine.-All the goblet cells of the small intestine take up 35S042- and discharge mucin with 35S incorporated in it. The time taken for radioactivity to be lost more or less completely varied from about 12 hours in the mouse to over 24 hours in the rat and rabbit. In the mouse and rat the cells of the crypts seemed to be metabolically more active than those of the villi, distributing radioactivity throughout the intracellular mucin, and discharging it, earlier. In the rabbit, on the other hand, radioactivity was still quite strong in cells near the bases of the crypts when it was disappearing from those on the villi. From the few experiments done on the cat and guinea-pig it seemed that the cat mucosa might behave like that of the rat and mouse, and the guinea-pig mucosa like that of the rabbit. There remains the question whether the experimental results provide evidence about the supposed passage of epithelial cells by sliding from the crypts, the site of mitosis, to the tips of the villi (see Discussion). Unfortunately, labelling of the cells by 35SO42- is transient, and movement of radioactive material out of the cells into the lumen of the intestine, as well as possible exchange with unlabelled sulphate, complicates the picture. In the rat, the earlier disappearance of activity from the crypts than from the villi might be taken as evidence that the cells from the crypts had moved on to the villi, though another explanation was suggested above. In the rabbit the findings

8 138 Jennings and Florey provide no evidence of such a movement, for the cells in the crypts, at the site where mitoses were abundant, still showed considerable activity 24 hours after sulphate injection. Colon.-The goblet cells in the colon, like those in the small intestine, took up 35S042- rapidly. Rat and Mouse.-As in some other situations the cells of the rat seemed to take up more 35SO42- than those of the mouse, and to retain it for longer, though the distribution of radioactivity was the same in the two species. Up to about 3 hours after the injection of sulphate the radioactivity was gathered compactly in the supranuclear region of the cells lining the sides of the crypts (fig. 24). In cells at the bases of the crypts and in the superficial mucosa activity was distributed more generally through the intracellular mucin. The rat killed 1 hr. 10 min. after sulphate injection showed strongly radioactive free mucin on the surface of the mucosa. In the mouse killed 12 hours after injection the deeper parts of the crypts had lost most of their radioactivity, but there was still substantial activity in the superficial half of the mucosa. The activity was now centrally placed over the mucin masses (fig. 25). The rat showed a similar picture 24 hours after injection; in the mouse at this time radioactivity was still present, but weak (fig. 26). Cat.-In both the cats the mucosa of the colon showed strong radioactivity in all the goblet cells. As in the rat and mouse, the cells in the lower part of the crypts were, on the whole, the first to show generalized radioactivity, while the cells towards the surface showed activity concentrated in the supranuclear region. Free mucin in the crypts and on the surface was active. Guinea-Pig.-In the colon of the guinea-pig the intracellular mucin in the lower part of the crypts showed generalized radioactivity, while towards the surface activity was concentrated in the supranuclear region of the cells. There was, however, a markedly greater amount of radioactivity in the deeper half of the mucosa (fig. 27). The distribution was the same in the animals killed 2 hours and 6j hours after injection, but free mucin on the surface was more strongly radioactive in the latter. Rabbit.-Unlike those of any of the other species examined, the cells near the surface of the colonic mucosa were the most active in taking up 35SO42-. In the animal killed 21 hours after the injection of sulphate there was strong generalized radioactivity over all the cells in the more superficial half of the mucosa, while those towards the bases of the crypts showed less intense radioactivity and a tendency to concentration in the supranuclear region. At 6i hours (fig. 29) and 24 hours after injection there was still considerable activity in the superficial half of the mucosa, but relatively little in the deeper half. The Effect of Stimulation by Mustard Oil.-The application of an irritant such as dilute mustard oil causes the goblet cells of the colon, particularly those in the crypts, to discharge their mucin [Goldsworthy and Florey, 1930; Florey and Webb, 1931]. If the mustard oil is removed before the mucosa becomes seriously damaged, the goblets are rapidly replenished and the mucosa is restored to its usual appearance in about 24 hours [Jennings and Florey, 1954]. Fig. 31 shows the colon of a mouse 21 hours after the removal of the irritant, which was immediately followed by the injection of Na235O42-. Stainable mucin has largely been emptied from the cells, particularly from those of the

9 Autoradiographs of Mucous Cells crypts. Nevertheless, considerable radioactivity is present in all the goblet cells, corresponding in position with the small residual or newly formed masses of mucin. Apparently large amounts of 35SO2- are being taken up for the synthesis of new mucin. The surface of the colon is covered by radioactive mucin secreted from the irritated crypts. The rat colon gave the same response as that of the mouse. One rat was killed 24 hours after irritation and sulphate injection when the mucin content of the cells had largely been restored; this mucosa still contained more radioactivity than the cells of a neighbouring area. Movement of Mucin and of Cells in the Colon.-In general the goblet cells of the colon took up more 35SO42- than did the goblet cells of the small intestine, and disappearance of radioactivity was, if anything, slower than elsewhere. In the mouse the greater part of the activity had disappeared at 24 hours, as in the small intestine, but in the rat and rabbit there was still substantial radioactivity in the colon at this time. In all species, except possibly the rabbit, the cells at the bases of the crypts were the most active in passing the 35S into the general intracellular mucin mass, and in the three species that were examined late enough to show the point these were the earliest cells to lose their radioactivity. Like the crypts of Lieberkuhn, the bases of the colonic crypts may be considered to be the most active part of the epithelium in mucin formation, as in cell division. After artificial irritation of the mucosa, so that most of the mucin was discharged, all the emptied cells took over these characters, and showed much more radioactivity in proportion to their mucin content than usual. Like the cells of the foveolae of the rat stomach, the cells of the irritated colon took up large amounts of 35SO42- and discharged highly radioactive mucin even though their content of stainable mucin was small. No more than in the small intestine do the observations provide clear evidence of movement of cells from the crypts to the surface. In the rabbit, the distribution of radioactivity was much the same in an animal killed 24 hours after the injection of sulphate as in one killed 61 hours after injection, which does not suggest a rapid rate of movement. In both rabbit and rat there was still residual activity in the cells at the bases of the crypts at 24 hours, and this was especially noticeable in the mucosa which had been irritated, and which contained a particularly large number of mitotic figures. It might be accounted for by supposing it to be the last remains of material diluted by cell division. But the results must be considered unhelpful on the question of the ascent of epithelial cells in the mucosa. Ccecum.-The cecum was examined in the rabbit and guinea-pig. The goblet cells, which are scarce in the mucosa of this organ, and are largely at the bottom of the shallow crypts, showed concentration of radioactivity. The intervening columnar epithelium was extraordinary in that it developed radioactivity although histologically no mucin could be detected (fig. 15). In no other part of the intestines was this phenomenon seen. RESIrLTS WITH [35S] METHIONINE The distribution of radioactivity after the injection of [35S] methionine was strikingly different from that after Na235SO4. Concentration of radioactivity VOL. xli, NO

10 140 Jennings and Florey was less, so that longer exposure of the photographic film was needed to obtain clear pictures, and it had no relation to the presence of mucus. Of the two mice that received this substance, the one killed 1 hour later showed little radioactivity. In that killed 21 hours later most of the gastrointestinal epithelium tended to concentrate the activity, which was diffusely spread over the epithelial cells. Concentration was most marked in the epithelium of the gastric foveolae, particularly in the pyloric region, and in the crypts of Lieberkuihn and of the colonic mucosa. Brunner's glands and the pyloric glands showed little activity, but there was some over the peptic cells. A portion of colon that had been irritated by mustard oil showed strong activity over all the epithelial cells (fig. 32), and this was to some extent apparent even in the animal killed one hour after injection. DIsCUSSION The Variety of Ga8tro-intestinal Mucins It has long been known that the mucins in the various mucous cells of the gastro-intestinal epithelium do not all take histological stains in the same way, and that differences in the staining properties of the same type of cell can be observed in different species. For instance, in 1893 Bonnet showed that gastric mucous cells did not stain like the goblet cells of the small intestine, the difference being so striking that he was led to doubt whether the pyloric glands secreted mucus at all. In 1895 Cremer showed that the staining of mucin-containing cells in the gastric mucosa was different in the pig and in man, and Bensley [ ] in particular pointed out a number of differences in staining between the various mucous cells of the stomach and between corresponding cells in different species. Cremer [1895] and Mayer [1897], to whom one of the best mucin stains-mucicarmine-is owed, put forward the view that chemical differences between mucins might account for their varied histological staining reactions, and already Liebermann [ ] had suggested from chemical investigations on extracts of mucosa that there were different kinds of mucin. It is of interest that his main division of these mucins was into those that contained sulphur and those that did not. Some recent workers have suggested the division of gastro-intestinal mucins into two groups-true mucin and mucoid-on histological and histochemical grounds [Clara, 1940; Gomori, ], but Jiirvi and Lauren [1951] have pointed out that the division is by no means clear cut, and that fixation and staining methods greatly affect some of the results obtained. Apart from the periodic acid-schiff and related reactions, which stain all the gastro-intestinal mucins, the most widely used staining method for mucins which has any histochemical basis is the metachromatic reaction of toluidine blue and some other dyes, which is

11 Autoradiographs of Mucous Cells 141 produced by goblet cell mucin and sometimes by gastric mucins. This reaction is thought generally to denote the presence of sulphated groups, as suggested by Lison [1936], though other acidic groups may produce the reaction, though more feebly, and local conditions such as binding with proteins may affect its intensity [Lison, 1953; Walton and Ricketts, 1954]. Methods of fixation and the treatment of the sections after staining may also modify the result [Clara, 1940; Pearse, 1953]. There is chemical evidence that there are sulphated muco-substances among a variety of mucins in the stomach and intestine, though much of the work has been done on extracts of mucosa (which may contain mesenchymal mucins) and relatively little on secreted juice [see Werner, 1953, and Kent and Whitehouse, 1955, for reviews, and Crevier and Belanger, 1954]. Recently it has been shown that the mucin present in Brunner's gland secretion of the rabbit contains 0-8 per cent of ester sulphates [Kent et al., 1956]. From the evidence of the present work its sulphur content may be lower than that of, for instance, goblet cell mucin. Autoradiographic Work The introduction of autoradiography using 35S has permitted a further analysis of the incorporation of sulphur into gastro-intestinal mucins. By this technique it was shown that after the parenteral injecton of Na235SO4 or dilute H235SO4 there was a concentration of radioactive sulphur in the mucosa of the stomach, duodenum and ileum of the adult rat [Odeblad and Bostr6m, 1952] and of the ileum of the adult rabbit [Bostrom and Odeblad, 1953]. Bostr6m [ ] stated that the radioactivity appeared to be in the mucus, and added the mucosa ofthe descending colon to the list oftissues showing strong activity. The first evidence that 35S (injected as weak sulphuric acid) localized in certain gastro-intestinal mucous cells and not in others was provided by Belanger [1953] in autoradiographs of 10-day old rats. In the glandular stomach radioactivity was confined to a zone a little below the surface of the mucosa, and it was plain that the surface epithelium had no significant activity. This distribution was confirmed by Davies and Young [1954]. More extensive observations made and illustrated by Belanger [1954] will be referred to below. The observations recorded in the present paper have been made on 5 species of animal, and the particular technical procedures used have permitted a somewhat more detailed examination than before. Variations in Uptake of 35S in the Stomach and Duodenal Glands It is abundantly confirmed that only in certain cells of the stomach and duodenum is 35S taken up from injected 35SO42- and incorporated in the mucin produced. The cell types that do this vary from species to species.
 but, in agreement with the observations of Belanger [1953, 1954] and of Davies and Young [1954], intense activity was present in the region of the bases of the foveolae in")
12 142 Jennings and Florey Surface, Foveolar and Mucoid Neck Cells.-In no species was radiogctivity found in the superficial epithelial cells of the stomach (figs. 1, 2, 6, 8, 9, 10, 11, 16) but, in agreement with the observations of Belanger [1953, 1954] and of Davies and Young [1954], intense activity was present in the region of the bases of the foveolae in rats (fig. 6). Mice and guinea-pigs (fig. 16) showed a similar distribution, but radioactivity was less intense. It seems clear, however, that the interpretation of these authors that the radioactive mucin was in the mucoid neck cells of the fundal glands is not correct, unless there are differences between strains or between animals of different ages. Fig. 7 shows that in our rats the main mass of the mucoid neck cells were inactive while the mucous cells at the bases of the foveolwe had intense radioactivity. Some specimens showed streams of radioactive mucin pouring towards the surface of the stomach from this region (fig. 6). The picture is reminiscent of the great activity in the crypts of the colon after stimulation by mustard oil (fig. 31). In both cases cells containing relatively little stainable mucin were pouring mucin into the lumen of the pit or crypt. In the guinea-pig the morphological distinction between foveolar and mucoid neck cells was blurred, and some of both kinds probably took up the 35S. This was certainly so in the rabbit, but only in a small part of the fundal mucosa, the rest showing no radioactivity. The cat showed no activity in any of the fundal mucous cells, but 35S appeared in the peptic cells. Pyloric and Brunner's Glands.-The difference in behaviour between different species is well shown by the pyloric glands. In the rat, as observed by Belanger [1954], these glands became radioactive soon after the injection of sulphate (figs. 1, 9, 10). The mouse, rabbit and cat (figs. 11 and 14) all showed some activity in the pyloric glands, but the guinea-pig showed none except in a few alveoli close to the pyloric ring (fig. 2). Species variation was again seen in Brunner's glands. Although the pyloric glands of the rat were strongly radioactive the neighbouring duodenal glands were inactive (fig. 1), as Belanger [1954] noted. In the mouse the same was substantially true though an occasional active alveolus was seen in Brunner's glands. In the rabbit both types of gland were weakly radioactive. Many Brunner's gland alveoli of the guinea-pig were highly active (figs. 2 and 5) though the main mass of pyloric glands showed no activity (figs. 2 and 13). The duodenal glands of the cat showed no activity (figs. 3 and 4). Uptake of 35S by the Goblet Cels Belanger [1954] reported the localization of 35S in the small and large intestines of suckling rats in a pattern that seemed to correspond with

13 Autoradiographls of Mucous Cells 143 the distribution of goblet cells, though the sections were unstained. Glucksmann [1953] reported localization in the intestinal goblet cells of mice without giving details of technique. Bostr6m and Odeblad [1954] did not state whether the material from the rabbit that they used and illustrated was small or large intestine, but they found concentrations of radioactivity in unstained sections that apparently corresponded with the goblet cells. In sections from mice they were able by staining through the autoradiographic film with polychrome methylene blue to satisfy themselves that the areas of concentration corresponded with the mucus within the goblet cells. The present observations confirm this distribution for all five species examined, viz. rat, mouse, guinea-pig, rabbit and cat. Relation of Uptake of 35S to Staining Reactions It was mentioned at the beginning of this Discussion that the metachromatic reaction bears some relation to the presence of sulphate groups. Belanger [1954] stated that after the injection of Na235SO4 the presence of radioactivity as shown in autoradiographs corresponded with the presence of a metachromatic reaction, and Bostr6m and Odeblad [1954] demonstrated this for the goblet cells of mice. In our material there was a striking correspondence in the goblet cells, which uniformly showed a strong metachromatic reaction and a strong concentration of 35S. In the stomach and duodenal glands only some types of cell produced metachromasia, the distribution varying from species to species, and the staining tended to be slight and fugitive. The fainter positive reactions, whether for radioactivity or for metachromasia, were often not reflected in the other method, and there were other irregularities. But on the whole the stronger reactions corresponded fairly well. For instance, in the rat the foveolse were picked out by a long line of metachromatic staining leading down to each gland, and the pyloric glands were positive and Brunner's glands negative. In the rabbit Brunner's glands were positive, but the pyloric glands, which were weakly radioactive, were negative. In the cat the foveolm3 showed some metachromasia and this was strongest in the pyloric region, the only area where the foveole showed signs of radioactivity. The pyloric and Brunner's glands, however, showed no metachromasia, though 'the former showed moderate and the latter faint radioactivity. The peptic cells of the cat, which were radioactive, were also strongly metachromatic. On the whole the results support current views about the significance of metachromasia. The mucus throughout the stomach and small intestine, and in the colon of rat, mouse and cat, gave pink staining with the periodic acid- Schiff (P.A.S.) reaction, whether radioactive sulphate was incorporated

14 144 Jennings and Florey or not. Staining was strong in the surface cells of the stomach and in the goblet cells, but tended to be pale in the mucoid neck cells and pyloric gland cells, probably because swelling of the mucin sometimes occurs in these cells on fixation. In the colon of the guinea-pig and rabbit there was a variation in staining that could be correlated with variation in the uptake of sulphate. In the rabbit the cells towards the surface of the mucosa, which showed great and persistent radioactivity, did not stain by the P.A.S. reaction, while those at the base, which showed relatively little radioactivity, stained bright red (figs. 29 and 30). The guinea-pig showed the reverse condition-the strongest radioactivity and the weakest P.A.S. reaction were in the deeper half of the mucosa (figs. 27 and 28). Dr. P. W. Kent pointed out that chemically an increasing incorporation of sulphate is consistent with a P.A.S. reaction of diminishing intensity, which may explain the inverse relationship of the two reactions in this case. In other regions and species, however, strong P.A.S. staining and uptake of 35S were found together. Incorporation of 35S from Injected Sulphate It is possible that the washing of our sections in a solution of sodium sulphate in order to remove free Na235SO4 accounts for minor differences between our results and those of other workers, though in our preparations this extra washing made no noticeable difference. Since the 35S taken up by mucous cells is not removed by thorough washing, even with a 2 per cent solution of Na2SO4, it is fair to conclude that it is in some way incorporated in the cell contents. (1) If the sulphate were absorbed as such by the cells and then incorporated into sulphur-containing amino-acids one might expect considerable activity over all epithelial cells, but this was not seen except in the caecum of the rabbit (fig. 15) and guinea-pig, where the phenomenon awaits explanation. Bostr6m and Odeblad [1954] stated that there was weak diffuse radioactivity over the intestinal epithelium of the rabbit five days after the injection of sulphate, when the goblet cells had lost their activity. They do not make it clear whether this activity had been present from the beginning or not. Nothing corresponding to it was seen in our own shorter-term experiments. (2) If amino-acids containing sulphur arriving by the blood stream were incorporated especially into mucin-producing cells, the sulphur of injected methionine might be expected to localize in these cells. In two experiments in which methionine labelled with 35S was injected into mice there was no such localization, even in the colon where the mucous cells were highly active, having been emptied and stimulated by mustard oil (fig. 32). (3) Though autoradiographs offer no proof, it would seem likely that sulphate is a precursor of material incorporated directly into
 mucin of the rabbit, which show that following an injection of Na235SO4 radioactive sulphur is incorporated in")
15 Autoradiographs of Mucous Cells 145 mucin and a source of sulphur for the ester sulphate of the goblet cells. This view is strengthened by observations on secreted duodenal (mainly Brunner's gland) mucin of the rabbit, which show that following an injection of Na235SO4 radioactive sulphur is incorporated in the duodenal mucin and that this sulphur is present for the most part as ester sulphate [Kent et al., 1956]. Activity of Mucous Cells as shown by 35S By the use of autoradiographs Belanger [1953] showed in young rats that 1 hour after the subcutaneous injection of dilute H235SO4 there was localized radioactivity in the stomach, and that at 48 hours activity had disappeared except in the stomach contents. He later [1954] confirmed this for rats and hamsters, and observed radioactive material in the intestinal contents 2 hours after an injection. Bostr6m and Odeblad [1954], using Na235SO4 injected intravenously, showed that radioactivity was greatest in the first specimens seen, 2 and 4 hours after injection, and thereafter declined. Our results confirm these findings. An hour after the injection of Na235SO4, 35S is present in mucous cells of the stomach and intestine. As far as autoradiographs show, the amount at this time is maximal and the same level is maintained for a few hours, after which the level declines so that by 24 hours little or none is left. In the case of the duodenum of the rabbit, the cell activity traced in autoradiographs taken at successive times after the injection of Na235SO, can be correlated with changes in the juice secreted by this area after an injection of Na235SO4 [Kent et al., 1956]. In autoradiographs the radioactivity of the Brunner's glands was of the same order 2j and 6i hours after the injection of sulphate, and at 24 hours almost none was left; in secreted juice incorporated (non-dialysable) radioactivity rose for about 4 hours after injection, and then fell gradually tiu a steady level was reached at 9 hours. In certain cells, particularly the goblet cells, in which passage of radioactive material into the intracellular mucin was relatively slow, it was possible to determine that 35S first appeared in the region at the base of the theca and above the nucleus (figs. 11, 17, 20, 21, 22, 24). This observation is similar to one made by Belanger [1954] on the palatal glands of young rats. A considerable body of evidence has accumulated to suggest that the synthesis of mucin occurs in the region of the so-called Golgi apparatus [for references see Florey, 1955]. The localization of -3S in this region soon after the injection of Na235SO4 would support the view that it is a part of the cell actively engaged in incorporating sulphur into the mucin molecule. The fact that this localized zone of activity can be seen in apparently mature goblets at and near the surface of the colon and on the villi of

16 146 Jennings and Florey the small intestine suggests that these apparently fully developed cells are producing mucin continuously. Apparently they do not first discharge their contents and then again start to synthesize secretion. The goblet cells of the colon which had been emptied and stimulated by mustard oil showed intense activity in taking up 35S and elaborating and secreting radioactive mucin (fig. 31), apparently without the accumulation of any large mucin mass in the cell. This too supports the view that these cells are capable of continuous synthesis and secretion. With the passage of time the site of radioactivity moves towards the free surface of the cell (figs. 18, 23, 25) and generally little or no residual activity is to be seen by the end of 24 hours (figs. 19 and 26). It would be rash to try to correlate this movement with the rate of discharge of mucin from the cell until more is known about continuous exchange of components between tissue fluids and cells. In a colon that had been stimulated by mustard oil there was still activity over the goblet cells 24 hours after sulphate had been given, although the rate of synthesis and discharge must have been unusually high in these cells. It seems that there may be differences in the metabolic activity of cells in different parts of the mucosa. In most cases those cells that earliest became diffusely covered by reduiced silver (as contrasted with those showing localization near the nucleus at first) were the first to lose their radioactivity. But this did not always apply; for instance in the colon of the rabbit the cells near the surface were the most active by this criterion, yet they retained considerable radioactivity for 24 hours. The processes of digestion do not apparently influence the distribution of 35S. The distribution was the same in starved and fed rats, though in starvation the amount of radioactivity was greater, suggesting that a stimulus to the discharge of mucus was lacking. It should be noted that only a few animals of each species were examined, so that there should be some reserve in generalizing from what was found. As far as the observations went the findings in each species were consistent. Evidence about Movement of Cells An isotope incorporated in a secretion that is being constantly discharged is not a very efficient "label" for a cell. However, a few incidental observations on possible cell movements were made. Besides producing mucin, apparently in large amounts (fig. 6), the foveolaw are numerically the most important site of mitosis in the rat stomach [Stevens and Leblond, 1953] and are thought to be the source of new cells to replace those desquamated from the surface epithelium [Bizzozero, 1893]. There is some support for this in the fact that 24 hours after the injection of sulphate, patches of radioactivity were found in the surface epithelium of the rat (fig. 12). Cells from the foveolse may have migrated to the surface while still containing some of their

17 Autoradiographs of Mucous Cells 147 active content. Some patches of radioactivity were also seen at 24 hours over the mucoid neck cells in the rat. This would support the suggestion, which has often been made, that these cells too are replaced from foveolar cells. However, Stevens and Leblond [1953] found in rats a high rate of mitosis among the mucoid neck cells themselves. This was the only species and site in the stomach where any evidence of cell movement was obtained. There are strong reasons for believing that in a similar way the goblet cells of Lieberkuihn's and the colonic crypts are moving from the crypt fundi, where mitosis takes place, towards the surface, from which cells are constantly desquamating [see, for instance, Leblond and Stevens, 1948; Leblond, Stevens and Bogoroch, 1948]. It cannot be said that our material provides definite evidence of this movement. It is true that in mouse and rat colon the main residual activity at 12 or 24 hours was in the superficial part of the mucosa, though there were still traces in the cells of the crypts. But in the small intestine this change of pattern was less regular, while in the rabbit neither small intestine nor colon showed changes that could be interpreted to imply movement of the cells. Three factors prevent any precise interpretation of the movement of such "labelled" cells-the rate at which 35S may exchange out of or be discharged from the cells; the length of time for which it continues to be taken up as the blood level falls; and its possible dilution by partition among the progeny of dividing cells, if the mucincontaining cells take part in cell division. Functions of Foveolar and Surface Epithelium The demonstration that mucus is secreted from the foveolwe (fig. 6), as well as from mucoid neck cells and pyloric glands [in the cat: see Florey, 1955], raises again the question of the part played by the surface epithelium in the production of mucus [reviewed by Hollander in Sandweiss, 1951]. Hollander [1952] has concluded that in preserving the stomach from physical or chemical damage it both secretes mucin and, as a second line of defence, is shed-his "two-component" theory. It has never been shown, however, that the surface cells discharge any substantial amount of free mucus into the stomach, and it may be that less emphasis should be laid on their secretory function and more on their function of protection by desquamation followed by spreading out of the remaining cells and regeneration, an idea which goes back at least to Claude Bernard [1856]. A comparison may be drawn with the keratinizing cells of the skin. Relationship of Different Cell Types Observations on the uptake of 35S by the different types of cell have some bearing on the question whether Brunner's glands are an extension
![148 Jennings and Florey of the pyloric glands into the duodenum, a thesis that has been maintained by many histologists on morphological grounds [see Plenk, 1932, p. 168, for discussion].](/docs-images/90/101392864/images/18-0.jpg "As Belanger [1954] pointed out, autoradiographically there is a sharp distinction between the reactions of the two areas in the rat (fig. 1), and the same is true for the mouse, guinea-pig (fig.")
18 148 Jennings and Florey of the pyloric glands into the duodenum, a thesis that has been maintained by many histologists on morphological grounds [see Plenk, 1932, p. 168, for discussion]. As Belanger [1954] pointed out, autoradiographically there is a sharp distinction between the reactions of the two areas in the rat (fig. 1), and the same is true for the mouse, guinea-pig (fig. 2) and cat. In the rabbit there is some activity in both, but the pattern of activity is not identical. These observations lend considerable support to the existing physiological evidence that the duodenal glands and pyloric glands do not behave in the same way [Wright, Jennings, Florey and Lium, 1940; Jennings and Florey, 1940], and to the assumption that the former are not an extension of the latter. Another point of interest is the degree of resemblance between cardiac glands, pyloric glands and mucoid neck cells, which seem to have certain physiological properties in common [Jennings and Florey, 1940] and whose possible functional and histological identity has been discussed [Lim, 1922; Plenk, 1932, p. 164]. In no species did the three types of cell all behave alike in regard to the uptake of 35S; further, in no two species was the pattern of activity among the three types precisely the same. Uptake of [35S] Methionine The results with [35S] methionine agree with those of Fitzgerald, Weinstein, Batchelder and Alvizouri [1954], who briefly reported concentration of radioactivity in the epithelium of the stomach and intestine of rats. The present results make it clear that this substance has no special relationship to the mucus-producing cells (fig. 32). As far as a very few experiments on a single species could show, it seems that some parts of the epithelium concentrate the radioactivity more than others. Investigations on methionine in relation to the production of mucus by methods other than autoradiography are discussed, and some new work is reported, by Kent et al. [1956]. SUMMARY 1. Certain of the epithelial cells of the stomach and duodenum take Up 35S from intravenously injected Na235SO4 and incorporate it in the mucous secretion. The particular types of cell which do this vary from species to species. 2. The goblet cells of the small and large intestines of all the species examined take up "S from Na235SO4. The 35S is first concentrated in the supranuclear or "Golgi body"' region, before being distributed through the intracellular mucin. 3. The 35S is taken up rapidly, and radioactive mucin is being discharged within an hour or so of the injection of Na235SO4. Most of the radioactivity has disappeared from the cells in 24 hours.

19 Autoradiographs of Mucous Cells After the intravenous injection of [35S] methionine, radioactivity appears diffusely in gastro-intestinal epithelium, and shows no concentration in the mucus-producing cells. 5. Some implications of the results are discussed. We wish to acknowledge our debt to Mr. J. Kent who assisted in all the animal work, Mr. D. Jerrome who prepared the autoradiographs and devised suitable staining methods, and Mr. B. H. Glass who took the photographs. REFERENCES B]LANGER, L. F. (1953). Nature, Lond. 172, BELANGER, L. F. (1954). Anat. Rec. 54, 118. BENSLEY, R. R. ( ). Quart. J. micr. Sci. 41, 361. BERNARD, C. (1856). Le9ons de physiologie experimentale appliquee a, la medecine. Paris: Bailliere. 16e legon. BIZZOZERO, G. (1893). Arch. mikr. Anat. 42, 82. BONNET (1893). Ber. oberhess. Ges. Nat. u. Heilk. 29, 193. BOSTROM, H. ( ). Ark. Kemi, 6, 43. BOSTROM, H. and ODEBLAD, E. (1953). Anat. Rec. 115, 505. BOSTR6M, H. and ODEBLAD, E. (1954). Acta physiol. scand. 32, 124. CLARA, M. (1940). Z. mikr.-anat. Forsch. 47, 183. CREMER, W. (1895). Untersuchungen uber die chemische Natur des Schleimkorpers der Magenschleimhaut. Diss., Bonn. CREVIER, M. and B]LANGER, L. F. (1954). C.R. Soc. Biol. Paris, 148, DAVIES, D. V. and YOUNG, L. (1954). Nature, Lond. 173, 448. DZIEWIATKOWSKI, D. D. (1949). J. biol. Chem. 178, 197. FITZGERALD, P. J., WEINSTEIN, J., BATCHELDER, R. and ALVIZOURI, M. (1954). J. Histochem. Cytochem. 2, 463. FLOREY, H. (1955). Proc. roy. Soc., B, 143, 147. FLOREY, H. and WEBB, R. A. (1931). Brit. J. exp. Path. 12, 286. GLtYCKSMANN, A. (1953). British Empire Cancer Campaign Thirty-first Annual Report covering the year 1953, p GOLDSWORTHY, N. E. and FLOREY, H. (1930). Brit. J. exp. Path. 11, 192. GOMORI, G. ( ). J. nat. Cancer Inst. 13, HEATLEY, N. G., JERROME, D. W., JENNINGS, M. A. and FLOREY, H. W. (1956). Quart. J. exp. Physiol. 41, 124. HOLLANDER, F. (1951). In Peptic Ulcer. Ed. D. J. Sandweiss. Philadelphia: W. B. Saunders Company. HOLLANDER, F. (1952). J. Mt. Sinai Hosp. 19, 328. JARVI, 0. and LAURIN, P. (1951). Acta path. microbiol. scand. 29, 26. JENNINGS, M. A. and FLOREY, H. W. (1940). Quart. J. exp. Physiol. 30, 329. JENNINGS, M. A. and FLOREY, H. W. (1954). Quart. J. exp. Physiol. 39, 271. KENT, P. W. and WHITEHOUSE, M. W. (1955). Biochemistry of the Amino-sugars. London: Butterworths Scientific Publications. KENT, P. W., WHITEHOUSE, M. W., JENNINGS, M. A. and FLOREY, H. W. (1956). Quart. J. exp. Physiol. 41. [In the press.] LEBLOND, C. P. and STEVENS, C. E. (1948). Anat. Rec. 100, 357. LEBLOND, C. P., STEVENS, C. E. and BOGOROcH, R. (1948). Science, 108, 531. LIEBERMANN, L. ( ). Biol. Zbl. 7, 54. LIM, R. K. S. (1922). Quart. J. micr. Sci. 66, 187. LISON, L. (1936). Histochimie animale. Paris: Gauthier-Villars. LIsoN, L. (1953). Histochimie et cytochimie animales. Principes et m6thodes. Paris: Gauthier-Villars. MAYER, P. (1897). Mitt. zool. sta. Neapel. 12, 303. ODEBLAD, E. and BOSTROM, H. (1952). Acta path. microbiol. scand. 31, 339.
. Brit. J. exp. Path. 35, 227. WERNER, I. (1953). Acta Soc. Med. upsalien, 58, 1. WRIGHT, R. D., JENNINGS, M. A., FLOREY, H. W. and LIUM, R. (1940).")
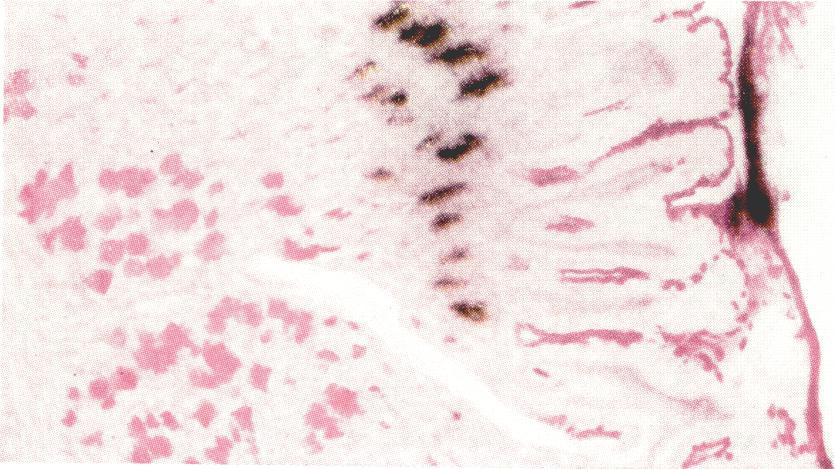
20 150 Jennings and Florey PEARSE, H. G. E. (1953). Histochemistry. London: J. & A. Churchill, Ltd. PLENK, H. (1932). Hb. mikr. anat. menschen, V/2. STEVENS, C. E. and LEBLOND, C. P. (1953). Anat. Rec. 115, 231. WALTON, K. W. and RICKETTS, C. R. (1954). Brit. J. exp. Path. 35, 227. WERNER, I. (1953). Acta Soc. Med. upsalien, 58, 1. WRIGHT, R. D., JENNINGS, M. A., FLOREY, H. W. and LIUM, R. (1940). Quart. J. exp. Physiol. 30, 73. DESCRIPTION OF PLATES The figures all show photomicrographs of histological preparations from animals injected with radioactive materials. All except figs. 28 and 30 are of autoradiographs, the black granules being silver particles in the photographic emulsion lying over the section. The granules have been focused sharply, while the mucin in the underlying cells is out of focus, though it can be recognized and located by its red colour. The exposure refers to length of time for which the section was allowed to act on the emulsion before the emulsion was developed. All sections were stained by the periodic acid-schiff method, and some in addition with Mayer's hkemalum and celestin blue, or with alum homatoxylin and metanyl yellow, or with metanyl yellow alone. PLATE I FIGc. 1.-Rat. Pylorus and duodenum. Specimen fixed 2 hrs. 10 min. after the injection of Na235SO4. Shows the contrast between the radioactivity of the pyloric glands and the lack of activity in Brunner's glands. The bases of the pyloric foveolm are strongly radioactive, the gastric surface epithelium inactive. The goblet cells are radioactive. Exposure 22 days. x 20. FIG. 2.-Guinea-pig. Pylorus and duodenum. To compare with fig. 1. Specimen fixed 2 hrs. after the intravenous injection of Na235SO4. There is great radio. activity in many of the alveoli of Brunner's glands and in some of the pyloric glands near the duodenum, but the rest of the pyloric glands show no activity. The villi of the intestine over Brunner's glands are cut obliquely; the goblet cells are radioactive. Exposure 15 days. x 20. FIG. 3.-Cat. Brunner's glands and bases of crypts of Lieberkiihn. Specimen fixed 6 hrs. after the injection of Na,35SO.. There is no radioactivity in the cells of Brunner's glands, but strong activity in the goblet cells in the crypts of Lieberkuhn. Exposure 41 days. x 120. FIG. 4.-Cat. Brunner's glands. Detail of alveoli from the same cat as that providing the section for fig. 3. There is no radioactivity in Brunner's glands. Exposure 41 days. x 480. FIG. 5.-Guinea-pig. Brunner's glands and bases of crypts of Lieberkuhn. Specimen fixed 2 hrs. after the injection of Na235SO4. Many of the cells in the alveoli of Brunner's glands show great radioactivity. Exposure 15 days. x 300. FIG. 6.-Rat. Fundus of stomach. From a starved rat. Specimen fixed 3 hrs. after the injection of Na235SO4. There is strong radioactivity in the mucous cells at the bases of the foveohe, but not in the mucoid neck cells or the superficial mucous cells of the stomach. Radioactive mucin is being discharged from the foveolae. Exposure 45 days. x 120. FIG. 7.-Rat. Mucoid neck cells. Specimen fixed 1 hr. 10 min. after the injection of Na235S04. Stained by Mayer's h?emalum and celestin blue in addition to the periodic acid-schiff reaction. There is radioactivity in the mucous cells at the bases of the foveole but none in the mucoid neck cells (contents stained red) nor in the peptic cells (contents stained blue). Exposure 70 days. x 280. FIG. 8.-Cat. Fundus of stomach. Specimen fixed 6 hrs. after the injection of Na235SO4. There is radioactivity in the peptic cells but not in the mucoid neck ceels or the mucous cells of the foveole and surface. Exposure 41 days. x 120.







21 ^ ^ ~I!t$s s t'4ir Cv i ( Al 4 * -.#he, "-WI.lw e, ti i~~~~~~~~~ 6 R 7 l- w w $t s~~~~14at F I$ /1½ c/(' ]H!/L I.)()







22 4* t it -t~ t 9' I0 I I :W i &*w: 12 tt 13 4 ii 11.4.f I *Do 0 e -* 14 lo".., +.zs + '~~~~ P1 ixi'i-'1:, I1

23 Autoradiographs of Mucous Cells 151 PLATE II FIG. 9.-Rat. Pyloric end of stomnach. The rat was fed 3 hrs. before the injection, after a period of starvation. Specimen fixed 3 hrs. after the injection of Na235SO4. Radioactive mucin is being secreted by the cells of the glands and lower ends of the foveolae, and can be seen in thelumina of the foveolae andonthesurface ofthe stomach. The superficial epithelium is free from radioactivity. Exposure 45 days. x 165. FIG. 10.-Rat. Pyloric end of stomach. Another microscopic field from the same section as fig. 9. The radioactivity of the pyloric glands is well shown. Exposure 45 days. x 165. FIG. 11.-Cat. Pyloric end of stomach. Specimen fixed 2j hrs. after the injection of Na235SO4. There is radioactivity in the gland cells and the extreme bases of the foveole, but not in the upper partspf the foveole or the superficial epithelium. The strongest activity is in the suprafhuclear region of the gland cells. Exposure 41 days. x 120. FIa. 12.-Rat. Surface of gastric mucosa. Specimen fixed 24 hrs. after the injection of Na235SO4. After the lapse of this time from the injection the residual activity is high in the foveolae, adjacent to the surface layer (cf. fig. 6). Exposure 82 days. x 480. FIG. 13.-Guinea-pig. Pyloric glands. Specimen fixed 2 hrs. after the injection of Na235SO4. There is noticeable activity in the stroma in this specimen, but no concentration over the mucous cells. Exposure 16 days. x 300. FIG. 14.-Cat. Pyloric glands. Specimen fixed 6 hrs. after the injection of Na235SO4. There is radioactivity in the mucous glands, but the peripheral arrangement of activity seen in fig. 11 has given place to general activity in the mucus in the cells. Exposure 41 days. x 300. FIG. 15.-Rabbit. Ccecum. Specimen fixed 2 hrs. after the injection of Na235SO4. There is concentrated radioactivity in the goblet cells at the bases of the crypts, but there is also diffuse activity in the apparently non-mucous cells. This contrasts with the lack of activity in epithelial cells elsewhere in the gut. Exposure 62 days. x 300. FIG. 16.-Guinea-pig. Cardiac region of stomach. Specimen fixed 2 hrs. after the injection of Na235SO4. The cardiac glands show no radioactivity. The neighbouring foveole are radioactive, and active mucin is being secreted on to the surface. Exposure 29 days. x 95.

24 152 Jennings and Florey PLATE III FIG. 17.-Mouse. Ileum. Goblet cells fixed 31 hrs. after the injection of Na236SO4. There is an accumulation of radioactivity at the base of the theca containing mucin, i.e. just above the nucleus of the cell. Exposure 24 days. x 480. FIG. 18.-Mouse. Ileum. Goblet cells fixed 6 hrs. after the injection of Na235SO4. The radioactivity is now distributed over the theca and some radioactive mucin is being discharged from the cells. Exposure 43 days. x 480. FIG. 19.-Mouse. Ileum. Goblet cells fixed 12 hrs. after the injection of Na235SO4. No radioactivity remains in the goblet cells. Exposure 43 days. x 480. FIG. 20.-Rat. Ileum. Goblet cells fixed 1 hr. 10 min. after the injection of Na235SO4. The radioactivity is principally at the base of the theca above the nucleus. Exposure 16 days. x 480. FIG. 21.-Rat. Ileum. Goblet cells fixed 3 hrs. after the injection of Na235SO4. The radioactivity is at the base of the theca above the nucleus. Exposure 45 days. x480. FIG. 22.-Cat. Ileum. Goblet cells fixed 2j hrs. after the injection of Na235SO4. The radioactivity is at the base of the theca above the nucleus. The mucin in the lumen is radioactive. Exposure 29 days. x 480. FIG. 23.-Guinea-pig. Ileum. Goblet cells fixed 61 hrs. after the injection of Na235SO4. The radioactivity is in the mass of mucin in the theca and on the surface of the epithelial cells. Exposure 14 days. x 480. FIG. 24.-Mouse. Colon. Goblet cells fixed 31 hrs. after the injection of Na235SO4. The radioactivity is at the base of the theca above the nucleus. Exposure 14 days. x 480. FIG. 25.-Mouse. Colon. Goblet cells fixed 12 hrs. after the injection of Na235SO4. The radioactivity is principally in the middle of the theca and there is radioactive mucin in the lumen of the crypt. Exposure 43 days. x 480. FIG. 26.-Mouse. Colon. Goblet cells fixed 24 hrs. after the injection of Na235SO4. There is a little residual radioactivity in the mucin in the theca. Exposure 43 days. x480. FIG. 27.-Guinea-pig. Colon. Crypts fixed 61 hrs. after the injection of Na235SO4. The goblet cells in the bases of the crypts have accumulated more radioactive sulphur than those at the surface. Exposure 14 days. x 120. FIG. 28.-Guinea-pig. Colon. Section without autoradiographic film from same block as fig. 27. Stained by the periodic acid-schiff reaction and by Mayer's hmmalum, celestin blue and tartrazine. The mucin only stains by P.A.S. in the cells near the surface, that in the cells of the crypts taking the yellow counterstain. x 120. FIG. 29.-Rabbit. Colon. Crypts fixed 61 hrs. after the injection of Na235SO4. The goblet cells in the upper parts of the crypts have accumulated more radioactive sulphur than those at the bases. Exposure 14 days. x 120. FIG. 30.-Rabbit. Colon. Section without autoradiographic film from same block as fig. 29. Stained as fig. 28. Only the mucin of the cells near the bases of the crypts stains by P.A.S. x 120. FIG. 31.-Mouse. Colon. Diluted mustard oil was applied to the surface of the colon to exhaust the goblet cells of their mucin. After 4 hrs. the mustard oil was removed and Na235SO4 was injected intravenously. The tissue was fixed 21 hrs. after this injection. There is great radioactivity in the goblet cells, in spite of their relatively small mucin content, and there is a considerable discharge of radioactive mucin on to the surface of the gut. Exposure 34 days. x 175. FIG. 32.-Mouse. Colon. Diluted mustard oil was applied to the mucosa for 4 hrs. to exhaust the goblet cells, as in the previous preparation (fig. 31). [35S] methionine was then injected and the tissue was fixed 2j hrs. later. All the epithelial cells contain radioactive material and there is no selection by the goblets. The mucus in the lumen of the gut is not radioactive. Exposure 81 days. x 175.

25 It. 'p Is 24 26, *a1r _ 4w 2 I Il \,I I,:I, I */"ot't" ph!1 52