SỰ SAO CHÉP DNA. DNA là vật chất di truyền. Thí nghiệm về biến nạp của Griffith. DNA mang tín hiệu di truyền. Thí nghiệm của Frederick Griffith (1928)
|
|
|
- Claud Stafford
- 5 years ago
- Views:
Transcription






 Chuột")
1 là vật chất di truyền SỰ SA CHÉP Thí nghiệm của Frederick Griffith (1928) mang tín hiệu di truyền Năm1944 nhóm Avery, McCarty, McLeod xác định rõ nguyên nhân gây biến nạp là gì? Tế bào S + (protease, RNAase) Chuột chết Thí nghiệm về biến nạp của Griffith 1952 Alfred Hershey và Martha Chase kết luận vật liệu di truyền của phage T2 là Tế bào S + (ase) Chuột sống là nhân tố biến nạp swald T. Avery 1

 1953 James D. Watson và Francis H. C.")
 Hai dây này đối xứng nhau, cùng song hành theo từng cặp base tương ứng, theo")
2 là vật chất di truyền Vật chất di truyền trong cơ thể sinh vật có nhiệm vụ truyền lại tính trạng từ đời trước xong đời sau, trên 3 nguyên tắc: Vật chất này phải có tính bền vững về thông tin đối với cấu trúc, chức năng, sự phát triển và sự sinh sản của tế bào. Có khả năng tự tái bản một cách chính xác sao cho tế bào con có thông tin di truyền giống như tế bào mẹ. Có khả năng thay đổi, giúp sinh vật biến dị, thích ứng, và tiến hóa. Cấu trúc xoắn kép của (Double helix structure of ) 1953 James D. Watson và Francis H. C. Crick xây dựng thành công mô hình cấu trúc phân tử Base pairing in Purine adenine (A) guanine (G) Pyrimidine thymine (T) cytosine (C) Pairing A:T 2 bonds C:G 3 bonds Đặc điểm của mô hình cấu trúc xoắn kép của Watson và Crick Phân tử có hai chuỗi dây polynucleotide quấn nhau theo chiều tay phải (right-handed fashion) Hai dây này đối xứng nhau, cùng song hành theo từng cặp base tương ứng, theo qui ước đầu 5 là gốc, đầu 3 là đuôi Dây cơ bản còn gọi là dây xương sống (The backbone) được hình thành bởi đường và photphase với những base đính hai bên trong dây. + Chuỗi xoắn kép cho phép các base purine và pirimidine có cấu trúc phẳng xếp chồng khít lên nhau ở bên trong phân tử, hạn chế sự tiếp xúc của chúng với nước. Chúng đính thẳng góc với dây xoắn. + Các nguyên tử đường và các nhóm phosphate xoay ra ngoài hình thành liên kết với nước đảm bảo tính ổn định cho phân tử Đặc điểm của mô hình cấu trúc xoắn kép của Watson và Crick Những base này ở trên hai dây đối xứng nhau được nối liền bởi cầu nối hydrogen: A-T và G-C. Cầu nối hydrogen rất dễ bị tách ra (ví dụ như nhiệt độ cao) để tạo thành hai dây đơn. Cặp base tương ứng A-T và C-G được gọi bằng thuật ngữ chuyên môn là complement base pair. Nối C-G (3 cầu nối) bền hơn nối A-T (2 cầu nối) Các cặp base cách nhau 0,34 nm trên dây xoắn. Mỗi một góc quay hoàn toàn (360o) của dây xoắn (helix) có độ dài 3,4 nm. Do đó, mỗi đoạn xoắn như vậy có tất cả 10 cặp base. Đường kính của một góc quay là 2nm. Kết quả của cấu trúc dây xoắn kép tạo ra những rãnh chính (major groove) và những rãnh phụ (minor groove). Cả hai rãnh này có kích thước đủ rộng cho phép những phân tử protein tiếp xúc với những base. 2


3 nucleotide P4 CH2 N base CH2 Deoxyribose base C H P CH 2 N base nucleotide P4 P4 CH2 base H Deoxyribose H Anti-parallel strands nucleotide P4 N base CH2 H covalent phosphodiester bonds Deoxyribose hydrogen bonds Tính ổn định và những biến động của Tính ổn định của là kết quả của hai quá trình: sao chép và sửa sai Các biến đổi của : đột biến, tái tổ hợp, các gen nhảy Tính ổn định của Cơ chế sao chép bán bảo tồn Các cơ chế sửa sai 3

: hai mạch gốc vẫn duy trì và bắt cặp trở lại sau khi tự sao chép.")
: phân tử gốc bị cắt ra thành nhiều đoạn mạch đôi hoạt động như dây nền (template) để tổng hợp")


4 Các mô hình sao chép Mô hình bán bảo tồn (Semiconservative model):vào đầu quá trình sao chép hai mạch của chuỗi xoắn kép tách rời nhau, mỗi mạch đơn được dùng làm khuôn để tổng hợp mạch mới. Kết quả là một phân tử ban đầu sẽ tạo ra hai phân tử con giống hệt nhau, mỗi phân tử con được hình thành từ một mạch cũ và một mạch mới Mô hình bảo tồn (Conservative model): hai mạch gốc vẫn duy trì và bắt cặp trở lại sau khi tự sao chép. Một trong hai phân tử mới là phân tử gốc (bao gồm hai mạch cũ), phân tử còn lại bao gồm vật liệu mới hoàn toàn. Mô hình phân tán (Dispersive model): phân tử gốc bị cắt ra thành nhiều đoạn mạch đôi hoạt động như dây nền (template) để tổng hợp ra những đoạn mạch đôi mới. Những đoạn mạch đôi như vậy hợp lại thành dây xoắn kép hoàn chỉnh, với các đoạn dây của dây gốc và dây mới xen kẽ. Các mô hình sao chép (a) Hypothesis 1: (b) Hypothesis 2: (c) Hypothesis 3: Semi-conservative replication Conservative replication Dispersive replication Intermediate molecule Thí nghiệm của Meselson và Stahl Sự sao chép của có tính chất bán bảo tồn Sự sao chép của có tính chất bán bảo tồn Isotopes of nitrogen (non-radioactive) were used in this experiment Sự sao chép ở prokaryote và eukaryote Sự sao chép ở vi khuẩn: mỗi nhiễm sắc thể là một replicon Replicon: đơn vị sao chép 4






 Polymerase III Gắn các nucleotide tự do theo")
5 SỰ KHỞI ĐẦU SA CHÉP Ở E.coli Nhóm enzyme helicase có nhiệm vụ tách mạch. Chúng bám lên một mạch đơn, cắt đứt liên kết hidro giữa 2 mạch đôi Chúng sử dụng năng lượng từ quá trình phân giải ATP 4 Sự sao chép được tiến hành đồng thời tại nhiều điểm trên phân tử của eukaryote TRÌNH TỰ KHỞI ĐẦU SA CHÉP Ở E.coli Nhóm enzyme helicase: tách mạch đôi thành mạch đơn Các protein SSB (single-stranded binding proteins) : ổn định mạch đơn đã được tách rời helicase single-stranded binding proteins NĂNG LƯỢNG Tổng hợp mạch mới (daughter ) Polymerase III Gắn các nucleotide tự do theo nguyên tắc bổ sung polymerase III CTP TTP GTP ATP CMP TMP GMP AMP ADP modified nucleotide 5





")




 Chỉ")



6 ĐẶC TÍNH HẠT ĐỘNG CỦA PLYMERASE Tổng hợp mạch mới từ mạch khuôn bằng cách gắn lần lượt các nucleotide tự do: Polymerase III no to bond Polymerase III Vào nucleotide cuối của mồi (primer) bắt cặp sẵn Polymerase III trên mạch khuôn. Theo chiều ligase Polymerase III Mạch tới và mạch chậm (Leading & Lagging strands) kazaki Replication fork / Replication bubble ki kaza ents fragm polymerase III Lagging strand ligase leading strand Leading strand lagging strand leading strand leading strand lagging strand Sự sao chép bắt đầu bằng việc tổng hợp mồi RNA (RNA Đặc tính của polymerase III primers) Chỉ có thể tổng hợp mạch mới khi đã có sẵn mồi bắt cặp trên mạch khuôn Loại bỏ mồi RNA và tổng hợp thay thế polymerase I polymerase III polymerase I polymerase III lagging strand primase RNA ligase RNA RNA primer: được tổng hợp bởi primase 6



7 Telomeres polymerase I polymerase III RNA telomerase TTAAGGGTTAAGGGTTAAGGG Telomerase and its function Telomerase Structure Reverse transcriptase with RNA template to bind to strands CHĨA BA SA CHÉP polymerase I 5 3 polymerase III ligase lagging strand primase kazaki fragments SSB CHĨA BA SA CHÉP helicase polymerase III leading strand direction of replication SSB = single-stranded binding proteins 7


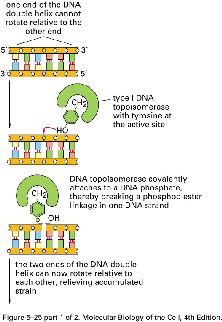
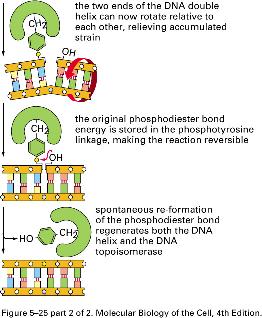
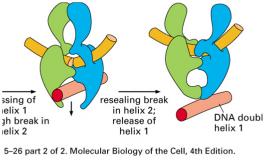
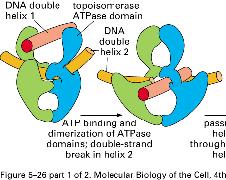
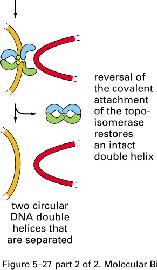
8 NÚT XẮN TẠI CHĨA BA SA CHÉP topoisomerase I topoisomerase II topoisomerase II 8