Structural basis for CARM1 inhibition by indole and pyrazole inhibitors
|
|
|
- Theodore Parsons
- 6 years ago
- Views:
Transcription



1 Structural basis for CARM1 inhibition by indole and pyrazole inhibitors John Sack, Sandrine Thieffine, Tiziano Bandiera, Marina Fasolini, Gerald J Duke, Lata Jayaraman, Kevin F Kish, Herb E Klei, Ashok Purandare, Pamela Rosettani, et al. To cite this version: John Sack, Sandrine Thieffine, Tiziano Bandiera, Marina Fasolini, Gerald J Duke, et al.. Structural basis for CARM1 inhibition by indole and pyrazole inhibitors. Biochemical Journal, Portland Press, 2011, 436 (2), pp < /BJ >. <hal > HAL Id: hal Submitted on 13 May 2011 HAL is a multi-disciplinary open access archive for the deposit and dissemination of scientific research documents, whether they are published or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
2 Structural Basis for CARM1 Inhibition by Indole and Pyrazole Inhibitors John S. Sack *, Sandrine Thieffine, Tiziano Bandiera, Marina Fasolini, Gerald J. Duke *, Lata Jayaraman *, Kevin F. Kish *, Herbert E. Klei *, Ashok V. Purandare *, Pamela Rosettani, Sonia Troiani, Dianlin Xie * and Jay A. Bertrand 1 * Bristol-Myers Squibb Research & Development, P.O. Box 4000, Princeton, New Jersey , USA Nerviano Medical Sciences, Via Pasteur 10, Nerviano, Italy Current Address: Italian Institute of Technology, Via Morego 30, Genova, Italy 1 Corresponding author: Jay A. Bertrand, Nerviano Medical Sciences, Via Pasteur 10, Nerviano, Italy, Phone: , Fax: ; jay.bertrand@nervianoms.com Short Title: Indole and Pyrazole Inhibitors of CARM1 Keywords: Coactivator-associated arginine methyltransferase 1 (CARM1), drug discovery, indole, Protein Arginine Methyl Transferases (PRMTs), pyrazole, selective enzyme inhibition. Abbreviations: BSA, bovine serum albumin; CARM1, coactivator-associated arginine methyltransferase 1; CMPD-1, 2-{4-[3-fluoro-2-(2-methoxyphenyl)-1H-indol-5- yl]piperidin-1-yl}-n-methylethanamine; CMPD-2, N-(3-{5-[5-(1H-indol-4-yl)-1,3,4- oxadiazol-2-yl]-3-(trifluoromethyl)-1h-pyrazol-1-yl}benzyl)-l-alaninamide; CMPD-3, 1-{3-[(L-alanylamino)methyl]phenyl}-N-(naphthalen-1-ylmethyl)-3-(trifluoromethyl)- 1H-pyrazole-5-carboxamide; GAR, glycine-arginine rich; GST, glutathione S-transferase; ITC, isothermal titration calorimetry; MBP, maltose-binding protein; NHR, nuclear hormone receptor; PRMT, protein arginine methyl transferase; RMSD, root mean square deviation; SAH, S-adenosyl-L-homocyseine; SAM, S-adenosyl-L-methionine; SNF, sinefungin; TCA, trichloroacetic acid. The coordinates and structure factors for the two crystal structures have been deposited in the Protein Data Bank with the accession codes 2Y1W and 2Y1X. 1
3 SYNOPSIS Coactivator-associated arginine methyltransferase 1 (CARM1) is a protein arginine methyl transferase (PRMT) family member that catalyzes the transfer of methyl groups from S- Adenosyl-L-Methionine to the sidechain of specific arginine residues of substrate proteins. This post-translational modification of proteins regulates a variety of transcriptional events and other cellular processes. Moreover, CARM1 is a potential oncological target due to its multiple roles in transcription activation by nuclear hormone receptors and other transcription factors like p53. Here we present crystal structures of the CARM1 catalytic domain in complex with cofactors (S-adenosyl-L-homocyseine or Sinefungin) and indole or pyazole inhibitors. Analysis of the structures reveals that the inhibitors bind in the arginine-binding cavity and the surrounding pocket that exists at the interface between the N- and C-terminal domains. In addition, we show using isothermal titration calorimetry that the inhibitors bind to the CARM1 catalytic domain only in the presence of the cofactor S-adenosyl-Lhomocysteine. Furthermore, sequence differences for select residues that interact with the inhibitors may be responsible for the CARM1 selectivity versus PRMT1 and PRMT3. Together, the structural and biophysical information should aid in the design of both potent and specific inhibitors of CARM1. 2
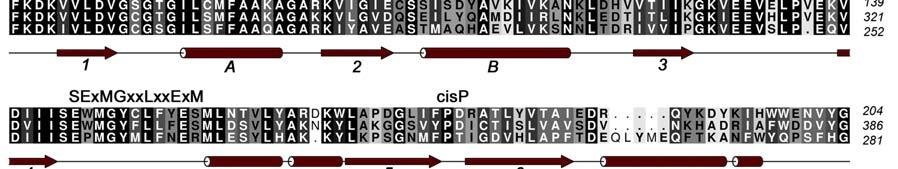
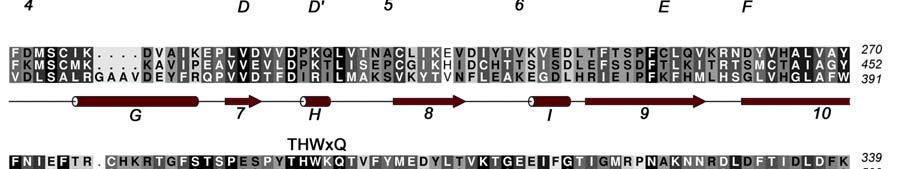
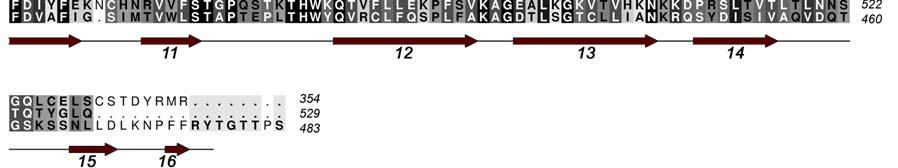
4 INTRODUCTION Arginine methylation is a common post-translational modification that in eukaryotes is catalyzed by the Protein Arginine Methyl Transferases (PRMTs), a family of proteins that transfer methyl groups from S-Adenosyl-L-Methionine (SAM) to the side chain of specific arginine residues [1]. The covalent modification of proteins by the addition of a methyl group to arginine residues can modulate their binding interactions and, as a result, provides a means of regulating their physiological function. The PRMT family, which consists of at least 9 members (PRMTs 1-9), can be classified into two main classes depending on whether they generate asymmetric (type I) or symmetric (type II) dimethyl arginine residues on substrate proteins. Coactivator-associated arginine methyltransferase 1 (CARM1), also known as PRMT4, was originally identified in a yeast two hybrid screen for proteins that associate with the p160 steroid receptor coactivator, GRIP1 [2]. CARM1 is recruited to the nuclear hormone receptor transcriptional complex by GRIP1 and this recruitment results in the methylation of other coactivators in the complex such as p300/cbp and AIB1. The recruitment of CARM1 to promoter regions of genes by the NHR complex or by transcription factors such as p53, β- catenin [3] or NF-kB [4] results in the methylation of specific arginine residues in the N- terminal tails of Histone H3. The direct consequence of these methylation events is the enhancement of gene transcription. [2, 5, 6]. Additional roles for CARM1 have been suggested in muscle differentiation [7] as well as protein repair, chromatin regulation, mrna stabilization [8] and gene splicing [9]. While full length CARM1 (608 residues) is required for co-activator function [10], the central 350 residue core region that contains both the SAM and activator binding sites is able to methylate substrate peptides in vitro. Crystal structures have been determined for the core region of CARM1 as well as for several other members of the arginine methyltranferases family that include PRMT1, PRMT3 and Hmt1 [11-15]. These proteins share a similar fold consisting of a two-domain structure; an N-terminal domain containing a mixed α/β Rossmann fold that is often referred to as the SAM binding domain and a C-terminal β-barrellike substrate-binding domain. The CARM1 N-terminal domain corresponds to the region of highly conserved sequence homology (50-60%) among the arginine methyltransferases and has been identified as responsible for binding SAM and for catalysis of the methyl transfer [14]. Three of the four PRMT signature sequences (motifs I, II and III) belong to the N-terminal domain (Figure 1). The C-terminal domain is the more structurally diverse region of the core domain with only 20-40% homology across the PRMT family. There are several divergent loops in this domain that may be involved in substrate specificity. This domain also contains the highly conserved "THWxQ loop (motif IV) that forms part of the substrate-binding groove. The β-barrel is interrupted after the first strand by a 39-residue two-helix bundle (residues ) capped by a hydrophobic tip (residues ). This dimerization arm or antenna makes extensive interactions with a second molecule to form the CARM1 dimer. CARM1 is unique among the PRMTs in that it also contains a C-terminal extension from the core region [16]. 3
5 While the full extent of CARM1 s role in cellular regulation is not yet known, it is clear that its multiple roles in transcription activation by nuclear hormone receptors [17] and other transcription factors like p53 [18] make it a likely target for the treatment of cancer. In fact, the over-expression of CARM1 has been observed in both breast and prostate cancers [19, 20]. To date, there have been a limited number of publications describing CARM1 inhibitors [21-26] and, of these, only our own efforts yielded inhibitors that were selective versus PRMT1 and PRMT3 [23, 24, 26]. Thus, in order to understand the basis for selectivity and to provide a starting point for structure-based design we determined crystal structures of the CARM1 catalytic domain in complex with cofactors and representative inhibitors of two different classes. Here, we report two crystal structures of CARM1 in complex with cofactors and inhibitors (Figure 2); one is the complex with sinefungin (SNF) and an inhibitor of the indole class (CMPD-1) and the other is the complex with S-adenosyl-L-homocyseine (SAH) and an inhibitor of the pyrazole class (CMPD-2). In addition, we characterized the binding of inhibitors from each class to CARM1 using Isothermal Titration Calorimetry (ITC). The structures of these complexes and the information obtained by ITC will help in the subsequent rounds of inhibitor design to obtain high affinity, specific inhibitors of CARM1. EXPERIMENTAL Chemical Synthesis CMPD-1 of the indole class was synthesized following the procedures reported for the analogous benzo[d]imidazole class [26] and additional information can be found in the Patent Application US 2006/ , 19 October CMPD-2 and CMPD-3 of the pyrazole class were synthesized following the procedures reported by Purandare and coworkers [23, 24]. Protein Expression, Purification and Crystallization The CARM1 catalytic domain from residues 134 to 483 was cloned into a pmal vector (New England Biolabs) obtaining by consequence a protein fused with MBP. This plasmid was transformed into the MM294(DE3) strain of E. coli. Inoculum was grown from a single colony at 37 C for 16h in LB2x media supplemented with 50 µg/ml of carbenicillin. The culture was grown at 37 C until OD 600 of 0.8, the temperature was reduced to 20 C and expression was induced using 1 mm IPTG. Cells were harvested by centrifugation after 20h and stored at 80 C. For purification, cell pellets were resuspended in Buffer A (50 mm Tris- HCl, 100 mm NaCl, 5 mm DTT, 2 mm EDTA, ph 7.5) supplemented with protease inhibitor tablets (Roche Biochemicals) and lysed by sonication. The clarified lysate was loaded onto an Amylose column and washed with Buffer B (25 mm Tris-HCl, 100 mm NaCl, 5 mm DTT, 2 mm EDTA, ph 7.5). MBP-CARM1 was eluted with maltose buffer, diluted 10 fold with 25 mm Tris-HCL, ph 7.5 and loaded onto an Ion exchange column. Next, the protein was eluted with a NaCl gradient and immediately loaded onto an Amylose column. MBP was cleaved by enterokinase overnight at 20 C on the column. After overnight cleavage, the eluted protein was loaded onto a gel filtration column pre-equilibrated with 25 mm Hepes, 100 mm NaCl, 0.5 mm TCEP, ph 7.5. Final yields were ~ 1.5 mg/l of culture. The purified CARM1 catalytic domain was ~ 95% pure as judged by SDS PAGE and electrospray mass spectrometry analysis revealed that spontaneous processing at the N-termini removes the first 4
6 6 amino acids. Thus, the resulting protein starts with Ser136-Val137-Phe138 and ends with Thr481-Pro482-Ser483. Crystals of the CARM1 catalytic domain in complex with SAH were grown using the hanging-drop vapor diffusion method from a solution of 20-30% PEG MME 2000, 0.1 M Tris-HCl ph 8.5 and 0.2 M Trimethylamine N-oxide dihydrate at 4 C. Before crystallization the CARM1 was concentrated to 2.4 mg/ml and SAH was added to the protein solution to give final concentrations of 0.25 mm. Diffraction quality crystals appeared within one week. To provide crystals with CMPD-2, crystals of CARM1 in complex with SAH were soaked in a solution of 20-30% PEG MME 2000, 0.1 M Tris-HCl ph 8.5 and 0.2 M Trimethylamine N- oxide dihydrate with 2 mm of CMPD-2 for 5 days. Crystals of the complex with sinefungin (SNF) and CMPD-1 were obtained by following the procedure described above for the crystals with SAH with some modifications. Specifically, 0.25 mm of SNF and 1 mm of CMPD-1 were added to the concentrated protein before setting up the crystallization trials. For data collection, the crystals were transferred to drops containing the equivalent mother liquor with 12.5% glycerol. Data Collection, Structure Determination and Refinement Crystals belong to the space group P (a =75.572, b = , c = ; α = β = γ = 90) with four molecules in the asymmetric unit. Diffraction data were collected at the European Synchrotron Radiation Facility (Grenoble, France) on beamlines ID14-4 and ID14-1 for CARM1 SNF CMPD-1 and CARM1 SAH CMPD-2, respectively. Indexing, integration and scaling were performed using HKL2000 [27]. Both CARM1 structures were solved by molecular replacement using the Molrep software [28]. Search models for both structures were an in-house structure of CARM1 in complex with SAH and a proprietary inhibitor that was solved by SeMet MAD. Model building was done using Coot [29] and refinement was done with CNX [30]. Crystallographic data is reported in Table 1 and SigmaA weighted difference omit maps of the inhibitors are shown in Figure 4. Structural images have been generated with PyMol [31]. Superposition of the two CARM1 structures gave RMSD values ranging from 0.2 to 0.9 Å for the 342 structurally equivalent Cα atoms. Enzymatic Activity Assay The methyltransferase activity of CARM1 was determined as described elsewhere [24] using a methylation based filter assay. Briefly, methylation reactions were performed for minutes using full-length GST-tagged CARM1 (6 nm), the substrate Histone H3 (1 µm) and tritiated SAM (0.05 µm) in methylation buffer (20 mm Tris-HCl, ph 8.0, 200 mm NaCl, 0.4 mm EDTA) both with and without inhibitors. Subsequently the reaction was stopped by the addition of TCA, the reaction mixture was precipitated with BSA overnight and the resulting mixture was filtered and washed before being read in a Top Count after the addition of MicroScint-20. PRMT1 and PRMT3 specific methylation assays were performed as described above for CARM1 but using as enzyme/substrate the combinations GST-PRMT1 (8 nm)/ Histone H4 (0.74 µm) or GST-PRMT3 (14 nm)/ GST-GAR (0.27 µm). The GST-fusion proteins (GST-CARM1, GST-PRMT1, GST-PRMT3 and GST-GAR) were expressed and purified from bacteria and the Histones H3 and H4 were both purchased from Roche Applied Science. Purity assessments using the Odyssey Imaging system with SDS-PAGE gels revealed that GST-CARM1 and GST-PRMT1 were 80 % pure and GST-PRMT3 was 85 % pure. 5
7 Peptide mass fingerprinting was used to verify the identity of the visible contaminants in the gel; all three proteins contained truncated versions of the target proteins, GST-CARM1 also contained endogenous GST (Spodoptera frugiperda) and GST-PRMT1 also contained DNAK (E. coli, at ~ 76 kda) and 60 kda chaperonin (E. coli, at ~ 65 kda). Protein concentrations were measured with the Coomassie Plus (Pierce) assay using BSA as a reference. Isothermal Titration Calorimetry The CARM1 catalytic domain was extensively dialyzed against buffer containing 25 mm Hepes ph 7.5, 100 mm NaCl, 0.5 mm TCEP. Experiments were carried out at 20 C with a VP-ITC titration calorimeter (MicroCal Inc., Northampton, MA) with protein in the cell and SAM, SAH or inhibitors in the syringe. Each titration experiment consisted of a first (5 µl) injection followed by 29 injections of 10 µl and the experiments with inhibitors were performed both in the presence and absence of 0.2 mm SAH. The SAH titration was run with 15 µm of CARM1 catalytic domain and 230 µm of SAH and all of the others with 20 µm of CARM1 catalytic domain and 200 µm of SAM, CMPD-1 or CMPD-3. Protein concentrations were measured with the Coomassie Plus (Pierce) assay using BSA as a reference after confirming that these values correspond well with those using the calculated extinction coefficient. RESULTS CMPD-1 and CMPD-2 are potent inhibitors of CARM1. A high-throughput screening effort using human full-length CARM1 in a methylation assay identified a variety of hits with modest activity and a subset of these were selected for further expansion to improve the in vitro properties [23, 24, 26]. This led to the indole and pyrazole inhibitor classes for which CMPD-1 and CMPD-2 (Figure 2), respectively, are representatives. Using a methylation based filter assay, IC50 values for the two compounds are ± and ± µm, respectively, for CARM1 and > 10 µm for PRMT1 and PRMT3. Indole and Pyrazole inhibitors bind to CARM1 in the presence of SAH. ITC experiments were done to confirm that the compounds identified and characterized with full-length CARM1 also bind to the catalytic domain under consideration for crystallographic studies. It is worth mentioning that, at the time, structures were available of PRMT1, PRMT3 and HMT1 but none of CARM1. Thus the idea was to try and crystallize a catalytic domain construct that was designed using sequence alignments in combination with structural data from the homologous proteins. In preparation for experiments with compounds, initial ITC titrations of CARM1 were done with SAM and SAH to control the integrity of the protein. SAM binds to CARM1 with a dissociation constant (Kd) of µm, a H of kcal/mol and a S of 19.1 cal/mol and SAH binds to CARM1 with a dissociation constant (Kd) of 6.90 µm, a H of 11.0 kcal/mol and a S of 13.8 cal/mol (Supplementary Figure S1). Experiments with CMPD-1 were done by titrating the compound into CARM1 both in the absence and presence of the cofactor SAH (Figure 3). Analysis of the resulting binding isotherm reveals that CMPD-1 binds to CARM1 with a dissociation constant of µm, a H of -9.0 kcal/mol and a S of -1.8 cal/mol in the presence of 0.2 mm SAH and no binding is observed in the absence of SAH. ITC experiments with CMPD-2 were not feasible due to the poor aqueous 6
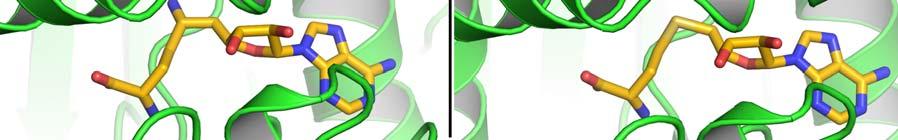
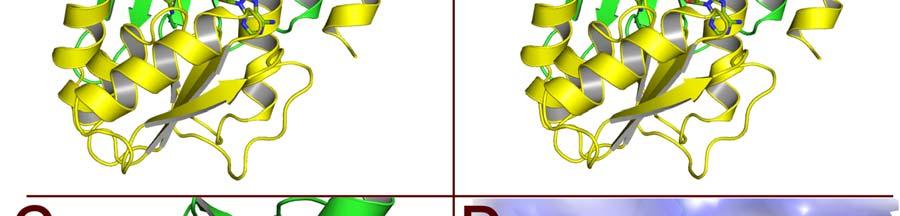
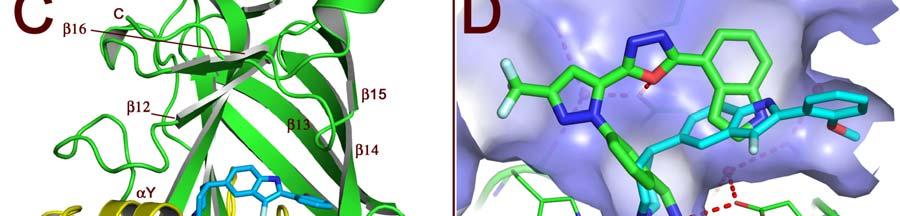
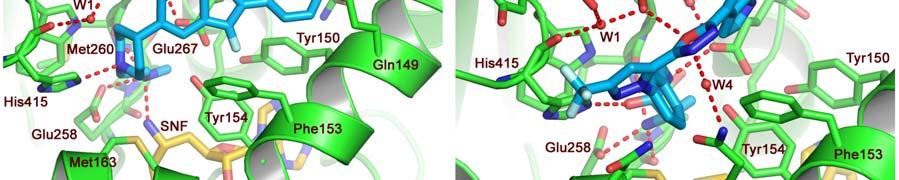
8 solubility so, instead, experiments were done with CMPD-3 (Figure 2), an analog of CMPD-2 with an IC50 of ± µm in the methylation based filter assay. Analysis of the binding isotherms reveals that CMPD-3 only binds to CARM1 in the presence of 0.2 mm SAH and, interestingly, fitting of the data with a single-site model gave poor results (data not shown) while fitting with a two-site model gave quite good results (as shown in Figure 3). Thus, CMPD-3 binds to CARM1 with a first dissociation constant (Kd1) of µm, a H of 7.9 kcal/mol, a S of 64.1 cal/mol and a stoichiometry of 0.16 and second dissociation constant (Kd2) of µm, a H of -1.0 kcal/mol, a S of -0.6 cal/mol and a stoichiometry of It s worth mentioning that although the data fits with a two-site model the combined stoichiometry equals So, a logical explanation would be that CMPD-3 binds to a single site with two different affinities. CMPD-1 and CMPD-2 occupy the substrate-binding site. The ternary complex of CARM1, CMPD-1 and SNF, a stable analog of SAM, was co-crystallized and the structure determined to 2.1 Å resolution and CMPD-2 was soaked into co-crystals of CARM1 with SAH and the structure determined to 2.4 Å resolution. Analysis of the resulting electron density maps reveals that both inhibitors bind in a rectangular-shaped pocket that exists at the interface between the two domains (Figures 4, 5 and S2). The sides of the pocket are formed by helices X and Y and the loops connecting β4 to αd, β11 to β12 and β13 to β14 (Figure 5C). At one end of the pocket, near His415, there is a deep cavity, hereafter referred to as the Arg-binding cavity, that extends towards the site of methyl transfer. Interestingly, residues from three of the PRMT signature motifs (motifs I, III and IV) help to form the pocket and cavity (Figure 1). In the case of CMPD-1, the inhibitors N-methylethanamine group is directed towards the bottom of the Arg-binding cavity and the piperidine moiety is positioned at the mouth of the cavity (Figure 6A). Interestingly, the pucker of the 6-member ring bends the inhibitor towards Ser146 and allows the indole ring system to make additional hydrophobic interactions with the residues lining the pocket. The amine nitrogen of the inhibitor makes several polar interactions with CARM1 residues in the Arg-binding cavity that include one of the side chain oxygens of Glu258, the carbonyl oxygen of Met260 and the nitrogen of SNF that mimics the methyl group of SAM. At the mouth of the cavity, the piperidine nitrogen hydrogen bonds with Nε2 of His415. Outside of the cavity, a bridging water molecule (W3) makes the only polar interaction between the protein and inhibitor, linking the indole nitrogen and Nδ2 of Asn266. Interestingly, two other water molecules help form the CMPD-1 binding-pocket; one by filling the space underneath the indole ring (W2) and the other positioned above the mouth of the Arg-binding cavity (W1). For CMPD-2, the terminal L-alaninamide is directed towards the bottom of the Arg-binding cavity and the benzyl ring is positioned at the mouth of the cavity (Figure 6B). The pyrazole moiety sits above the imidazole ring of His415 and the attached substituents point in opposite directions. Specifically, the trifluoromethyl group extends towards the solvent, passing between the side chains of Asn162 and Tyr417, while the 1,3,4-oxadiazole crosses back over the mouth of the Arg-binding cavity. The attached indole group is positioned above the Glu267 side chain and packs against the side chains of Tyr262, Pro473 and Phe475. The L- alaninamide of CMPD-2 makes several polar interactions with CARM1 in the Arg-binding 7
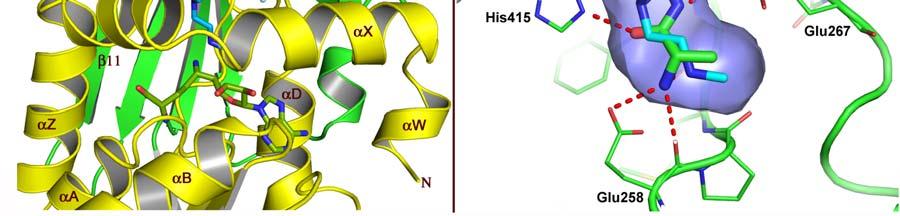
9 cavity. At the bottom of the cavity, the terminal amino nitrogen interacts with one of the side chain oxygens of Glu258 and the carbonyl oxygens of Glu258 and Met260. Then, towards the middle of the cavity the carbonyl oxygen of the L-alaninamide hydrogen bonds with Nε2 of His415 while the adjacent nitrogen hydrogen bonds with one of the side chain oxygens of Glu267. Outside of the Arg-binding cavity, the oxadiazole makes the only other polar interactions with CARM1. These include a hydrogen bond between the oxadiazole oxygen and the hydroxyl oxygen of Tyr262 and a bridging water molecule (W4) that interconnects the oxadiazole nitrogen in position 3 with Nε2 of Gln159. It s also worth noting that the three water molecules that were observed in the structure with CMPD-1 are also present in the structure with CMPD-2. DISCUSSION SAH binding to CARM1 causes large structural changes near the catalytic site that may explain the lack of inhibitor binding in the absence of SAH. To date there are three apostructures of CARM1 (PDB codes 3B3G, 3B3J and 2V7E) and two structures of the CARM1 SAH complex (PDB codes 3B3F and 2V74). Analysis of these structures by the authors [11, 12] reveals different types of structural changes that are associated with SAH binding that include major modifications to the region that contains the PRMT signature motif I and minor modifications to the other three PRMT signature motifs (motifs II, III and IV). Of the three apo-structures of CARM1, 3B3J shows the largest degree of structural reorganization with SAH binding while the other two structures are more similar to those of the CARM1 SAH complexes. In 3B3J, the segment forms a β-strand (β0) followed by a 3 10 helical turn (residues ) that leads into a long helix comprising residues Thus, the transition associated with SAH binding causes β0 to be converted into αx and a kink is introduced in the long helix comprising residues to form the helices αy and αz [11]. In addition, several residues are repositioned to properly form the Arg-binding cavity and the surrounding pocket. Interestingly, many of these residues are the same as those described above that interact with the inhibitors CMPD-1 and CMPD-2. Thus, it is not surprising that the inhibitors fail to bind in the in the absence of SAH since cofactor binding induces large structural changes that lead to the proper formation of the Arg-binding cavity and the surrounding pocket. Comparing the published CARM1 structures with SAH (PDB codes 3B3F and 2V74) and those reported here with cofactors and inhibitors reveals only slight modifications between the different structures. However, one region that varies in the different structures is the N- terminus that in our structures initiates with the helix αw that is followed by a short loop that subsequently leads into the helix αx (Figure 5C). In contrast, the structures 3B3F and 2V74 both lack the helix αw and, as a result, the ordered regions of the N-termini initiate with the helix αx [11, 12]. In our structures, αw packs against the adenine group of SAH shielding it from the solvent while in the structures 3B3F and 2V74 a cavity exists below the helix αx that exposes SAH to the solvent. Also, since there are a series of interactions between the loop connecting αw to αx and the one connecting β13 to β14 that effectively forms one end of the substrate-binding pocket, the disorder of the loop between αw to αx could impact substrate binding. This could be a partial explanation of why Yue and coworkers were unable to obtain well-ordered electron density for the CARM1 complex with SAH and the H3-peptide [12]. 8
10 Actually in the CARM1 structure with SAH reported by Yue and coworkers (2V74) the loop connecting β13 to β14 is shifted 1.7 angstroms away from the substrate-binding pocket with respect to the corresponding loops in 3B3F and the structures reported here with inhibitors. A logical explanation for the shift of the β13 to β14 loop would be because the crystallization construct used by Yue and coworkers begins with Ala147 and thus lacks all of the residues of the αw to αx loop that stabilize the loop position. Comparing the CARM1 structures with CMPD-1 and CMPD-2 reveals how the inhibitors use different regions of the Arg-binding cavity and the associated rectangular pocket that exists at the interface between the two domains (Figures 5D, 6A and 6B). Not surprisingly, both inhibitors exploit the Arg-binding cavity to make polar interactions; CMPD-1 interacts with Glu258, Met260 and His415 while CMPD-2 makes similar polar interactions plus an additional one with Glu267. The mouth of the Arg-binding cavity is a point of deviation between the two inhibitors since each one makes extensive hydrophobic interactions with a residue lining the mouth of the cavity (Glu267 and His415) but the two residues are situated on opposite sides of the mouth; CMPD-1 follows the mouth of the cavity on one side, wraps over Glu267 and extends towards Ser146 while CMPD-2 follows the mouth of the cavity on the opposite side, packs against His415 and extends towards the N-terminus of the helix αz. Actually, the binding mode of CMPD-2 is slightly more complicated since the pyrazole ring effectively directs the inhibitor in two directions with the trifluoromethyl group extending towards the N-terminus of helix αz and the oxadiazole and indole rings crossing back over the mouth of the cavity and extending towards the loop interconnecting β15 and β16. Together this gives the general effect that CMPD-2 exploits better the region of the pocket between the Arg-binding cavity and the N-terminus of the helix αz while CMPD-1 exploits better the region between the Arg-binding cavity and Ser146. The chemical expansions that started from the initial hits and progressed to CMPD-1 and CMPD-2 provide a wealth of information that can be revisited using the structures. For example, efforts were made to optimise the portion of the indole and pyrazole inhibitors that binds in the Arg-binding cavity and, in both cases, even minor modifications gave reductions in activity [24, 25, 26]. Presumably the Arg-binding cavity requires a specific shape in order for it to selectively produce asymmetric dimethyl arginine and the inhibitors optimally exploit the shape but leave little room for modifications. Outside of the Arg-binding cavity the tolerance for variation increases for both chemical classes, a result that correlates with the size of the pocket and the possibilities to expand in different directions. For example, multiple scaffolds were attempted before identifying the preferred fluorine substituted indole core of CMPD-1. In fact, the initial hit and the subsequent series that eventually led to CMPD-1 contained a methylated benzo[d]imidazole core rather than the fluorine substituted indole [26]. During the optimisation of the original pyrazole hit efforts were made to improve the permeability of the molecule by reducing the number of hydrogen bond donors. This led to the replacement of the amide that was present in the original hit with a 1,3,4-oxadiazole ring, a modification that not only improved the permeability as measured by the PAMPA assay but also improved the in vitro potency [23]. It is worth noting that CMPD-3 contains the original amide bond while CMPD-2 contains the 1,3,4-oxadiazole ring that was introduced as an amide surrogate (Figure 2). Finally, the pyrazole expansion revealed that a reasonable number of variations are permitted for the aromatic groups that are attached to the 1,3,4-oxadiazole 9
11 ring or the amide linkage [23, 24]. The positioning of the indole ring in the structure with CMPD-2 correlates well with this result since it only uses a limited amount of the space that is available between the Arg-binding cavity and Ser146. Several specific residues are believed to be responsible for the CARM1 selectivity of the inhibitors CMPD-1 and CMPD-2. These include residues coming from the helices αx and αy (Ser146, Gln149, Phe153, Gln159, Asn162), the loop between β4 and αd (Asn266), the loop between β11 and β12 (Tyr417), the loop between β13 and β14 (Arg446-Gln447-Ser448) and the C-terminal segment that contains β16 (Pro473, Phe475, Tyr477). Interestingly, the majority of these residues do not make polar interactions with the inhibitors but instead help to define the shape of the pocket (Figures 6A and 6B). For example, residues from the helices αx and αy (Ser146, Gln149, Phe153, Gln159, Asn162) form one side of rectangular-shaped pocket that exists at the interface between the two domains in CARM1. In PRMT1 and PRMT3, the corresponding residues differ in sequence thus modifying the shape of the pocket (Figures 1, 6C and 6D). For PRMT3, this difference in pocket shape can be observed with the structure 1F3L while for PRMT1 the analysis is complicated by the disorder at the N-terminus for all three of the published PRMT1 structures (1ORI, 1ORH, 1OR8). However, sequence differences between CARM1, PRMT1 and PRMT3 for helices αx and αy strongly suggest that the shape for this region of the PRMT1 pocket will be similar to that of PRMT3 and significantly different from that of CARM1. One noteworthy sequence difference that affects the shape of the pocket in this region is Phe153 in the CARM1 motif I which corresponds to Ser in both PRMT1 and PRMT3 (Ser 38 and Ser220, respectively). Since Phe153 is directed into the pocket where it makes hydrophobic interactions with the inhibitors the substitution of this residue by a hydrophilic serine will result in an overall loss of interactions. Asn266 in CARM1 is another noteworthy residue with respect to sequence differences that may affect inhibitor selectivity. In PRMT1 and PRMT3 the corresponding motif II residues are Tyr152 and Phe334, respectively, and the significant differences in residue size and character will clearly impact the shape of the pocket and the types of interactions that can be made with inhibitors. This point is highlighted by the water-mediated interaction that exists between Asn266 and the indole nitrogen of CMPD-1 since the aromatic residues of PRMT1 and PRMT3 would be unable to make the same type of interaction. Because of the different binding modes for CMPD-1 and CMPD-2, some of the specific residues may correlate with the selectivity of only one of the two inhibitors (Figures 6A and 6B). For example, the trifluoromethyl group of CMPD-2 is positioned between the side chains of Asn162 and Tyr417, two CARM1 specific residues, while none of the CMPD-1 atoms interact with these residues. In addition, since CMPD-2 extends farther out of the Arg-binding cavity than CMPD-1 it is able to make interactions with the C-terminal residues Pro473, Phe475 and Tyr477. In contrast, the methoxy-phenyl group of CMPD-1 is positioned at the other end of the rectangular pocket where it interacts with Ser146 and the loop residues Arg446-Gln447- Ser448. A surprising result was the apparent difference in affinity between the methylation-based assay and ITC results for CMPD-1 (IC50 = µm and Kd = µm) and CMPD-3 (IC50 = µm, Kd1 = µm and Kd2 = µm). The IC50 values show a 4-fold difference between the two compounds and CMPD-1 has the higher affinity while the Kd values have a significantly different pattern with a greater than 20-fold difference and CMPD- 10
12 3 has the higher affinity. Although a clear explanation concerning the source of these differences is beyond the scope of this work, several different factors could be responsible. One thing to keep in mind is that the two types of measurements are quite different since the assay measures the methylation of Histone H3 over an extended period of time (equilibrium conditions) while ITC measures inhibitor binding to CARM1 in the presence of the product cofactor SAH. Another relevant detail is that the measurements use different forms of CARM1, the IC50 measurements use full-length CARM1 while the ITC studies use the catalytic domain. Since the structural work revealed that the inhibitors bind in a cavity that is in close proximity to the N- and C-termini of the catalytic domain, it is possible that the inhibitors interact with residues from the N- and C-terminal domains of the full-length protein and these interactions impact the IC50 values. Hopefully additional insights will come from the structures of full-length CARM1 in complex with inhibitors and/or protein substrates. In conclusion, this article reports the first crystal structures of the CARM1 catalytic domain in complex with inhibitors, thus enabling structure-based drug design for this attractive oncology target. Since the indole and pyrazole inhibitors bind in the Arg-binding cavity and, to date no other structures are available with something bound in this region, they provide valuable information concerning the types of interactions that could exist with the natural substrate. Finally, analysis of the structure identified zones of the active site that are likely to be responsible for the CARM1 selectivity of the indole and pyrazole inhibitors versus PRMT1 and PRMT3. ACKNOWLEDGEMENTS John S. Sack, Gerald J. Duke, Lata Jayaraman, Kevin F. Kish, Herbert E. Klei, Ashok V. Purandare, and Dianlin Xie are employees of Bristol-Myers Squibb and Sandrine Thieffine, T. Bandiera, Marina Fasolini, Pamela Rosettani, Sonia Troiani and Jay A. Bertrand are employees of Nerviano Medical Sciences. The authors thank James Bryson, James Tamura, Michael Wittekind, Changhong Wan, Hilary Gray, and Valentina Goldfarb of Bristol-Myers Squibb Company for their helpful suggestions on expression and purification of the enzyme. 11
13 REFERENCES 1. Lee, H.W., Kim, S. and Paik, W.K. (1977). S-Adenosylmethionine:Protein-Arginine Methyltransferase. Purification and Mechainism of the enzyme. Biochem. 16, Chen D., Ma H., Hong H., Koh S.S., Huang S-M., Schurter B.T., Aswad D.W. and Stallcup M.R Regulation of Transcription by a Protein Methyltransferase. Science 284, Koh, S.S., Li, H., Lee, Y.-H., Widelitz, R.B., Chuong, C.-M. and Stallcup, M.R. (2002). Synergistic Coactivator Function by Coactivator-associated Arginine Methyltransferase (CARM) 1 and -b-catenin with Two Different Classes of DNAbinding Transcriptional Activators. J. Biol. Chem. 277, Covic, M., Hassa, P.O., Saccani, S., Buerki, C., Meier, N.I., Lombardi, C., Imhof, R., Bedford, M.T., Natoli, G., Hottiger, M.O. (2005). Arginine methyltransferease CARM1 is a promoter specific regulator of NF-kB dependent gene expression. EMBO J, 25, Bauer, U-M., Daujat, S., Nielsen, S.J., Nightingale, K. and Kouzarides, T (2002). EMBO reports. 3, Schurter, B.T., Koh, S.S., Chen, D., Bunick, G.J., Harp, J.M., Hanson, B.L., Henschen-Edman, A., Mackay, D.R., Stallcup, M.R. and Aswad D.W. (2001). Methylation of Histone H3 by Coactivator-Associated Arginine Methyltransferase 1. Biochem. 40, Chen, S.L., Loffler, K.A., Chen, D., Stallcup, M.R. and Muscat, G.E.O. (2002). The Coactivator-associated Arginine Methyltransferase Is Necessary for Muscle Differentiation. J. Biol. Chem. 277, Fugiwara, T., Mori, T., Chu, D.L., Koyama, Y., Miyata, S., Tanaka, H., Yachi, K., Kubo, T., Yoshikawa, H., and Tohyama, M (2006). CARM1 regulates proliferation of PC12 cells by methylating HuD. Mol Cell Biol 26, Cheng, D., Côté, J., Shaaban, S. and Bedford MT. (2007) The arginine methyltransferase CARM1 regulates the coupling of transcription and mrna processing. Mol Cell. 25, Teyssier, C., Chen, D. and Stallcup, M.R. (2002). Requirement for Multiple Domains of the Protein Arginine Methyltransferase CARM-1 in Its Transcriptional Coactivator Function. J. Biol.Chem. 277,
14 11. Troffer-Charlier, N., Cura, V., Hassenboehler, P., Moras, D. and Cavarelli, J. (2007). Functional insights from structures of coactivator-associated arginine methyltransferase 1 domains. EMBO J. 26, Yue, W.W., Hassler, M., Roe, S.M., Thompson-Vale, V. and Pearl, L. H. (2007). Insights into histone code syntax from structural and biochemical studies of CARM1 methyltransferase. EMBO J. 26, Zhang, X. and Cheng, X. (2003). Structure of the Predominant Protein Arginine Methyltransferase PRMT1 and Analysis of its Binding to Substrate Peptides. Structure 11, Zhang, X., Zhou, L. and Cheng, X. (2000). Crystal structure of the conserved core of protein arginine methyltransferase PRMT3. EMBO J. 19, Weiss, V.H., Mcbride, A.E., Soriano, M.A., Filman, D.J., Silver P.A. and Hogle, J.M. (2000). The Structure and Oligomerization of the Yeast Arginine Methyltransferase, Hmt1. Nat. Struct. Biol. 7, McBride, AE and Silver, PA. (2001). State of the arg: protein methylation at arginine comes of age. Cell 106, Kraus, W.L. and Wong, J. (2002). Nuclear receptor-dependent transcription with chromatin. Is it all about enzymes?. Eur. J. Biochem. 269, Woojin, A., Kim, J. and Roeder, R.G. (2004). Ordered Cooperative Functions of PRMT1, p300, and CARM-1 in Transcriptional Activation by p53. Cell 11, Hong, H., Kao, C., Jeng, M.-H., Eble, J.N., Koch, M.O., Gardner, T.A., Zhang, S., Li, L., Pan, C.-X., Hu, Z., MacLennan, G.T. and Cheng, L. (2004). Aberrant expression of CARM-1, a transcriptional coactivator of androgen receptor, in the development of prostate carcinoma and androgen-independent status. Cancer 101, El Messaoudi, S., Fabbrizio, E., Rodriguez, C., Chuchana, P., Fauquier, L., Cheng, D., Theillet, C., Vandel, L., Bedford, M. T. and Sardet, C. (2006) Coactivator-associated arginine methyltransferase 1 (CARM1) is a positive regulator of the Cyclin E1 gene. Proc. Natl. Acad. Sci. USA 103, Allan, M., Manku, S., Therrien, E., Nguyen, N., Styhler, S., Robert, M.F., Goulet, A.C., Petschner, A.J., Rahil, G., Robert Macleod, A., Déziel, R., Besterman, J.M., Nguyen, H., and Wahhab, A. (2009). N-Benzyl-1-heteroaryl-3-(trifluoromethyl)-1Hpyrazole-5-carboxamides as inhibitors of co-activator associated arginine methyltransferase 1 (CARM1). Bioorg Med Chem Lett. 19, Castellano, S., Milite, C., Ragno, R., Simeoni, S., Mai, A., Limongelli, V., Novellino, E., Bauer, I., Brosch, G., Spannhoff, A., Cheng, D., Bedford, M.T. and Sbardella, G. 13
15 (2010). Design, synthesis and biological evaluation of carboxy analogues of arginine methyltransferase inhibitor 1 (AMI-1). ChemMedChem. 5, Huynh, T., Chen, Z., Pang, S., Geng, J., Bandiera, T., Bindi, S., Vianello, P., Roletto, F., Thieffine, S., Galvani, A., Vaccaro, W., Poss, M.A., Trainor, G.L., Lorenzi, M.V., Gottardis, M., Jayaraman, L. and Purandare, A.V. (2009). Optimization of pyrazole inhibitors of Coactivator Associated Arginine Methyltransferase 1 (CARM1). Bioorg Med Chem Lett. 19, Purandare, A.V., Chen, Z., Huynh, T., Pang, S., Geng, J., Vaccaro, W., Poss, M.A., Oconnell, J., Nowak, K. and Jayaraman, L. (2008). Pyrazole inhibitors of coactivator associated arginine methyltransferase 1 (CARM1). Bioorg Med Chem Lett. 18, Therrien, E., Larouche, G., Manku, S., Allan, M., Nguyen, N., Styhler, S., Robert, M.F., Goulet, A.C., Besterman, J.M., Nguyen, H. and Wahhab, A. (2009). 1,2- Diamines as inhibitors of co-activator associated arginine methyltransferase 1 (CARM1). Bioorg Med Chem Lett. 19, Wan, H., Huynh, T., Pang, S., Geng, J., Vaccaro, W., Poss, M.A., Trainor, G.L., Lorenzi, M.V., Gottardis, M., Jayaraman, L. and Purandare, A.V. (2009). Benzo[d]imidazole inhibitors of Coactivator Associated Arginine Methyltransferase 1 (CARM1)--Hit to Lead studies. Bioorg Med Chem Lett. 19, Otwinowski, Z. and Minor, W. (1997) Processing of X-ray Diffraction Data Collected in Oscillation Mode, Methods in Enzymology, Volume 276: Macromolecular Crystallography, part A, p , C.W. Carter, Jr. & R. M. Sweet, Eds. 28. Vagin, A.A. and Teplyakov, A. (1997). MOLREP: an automated program for molecular replacement. J. Appl. Cryst. 30, Emsley, P. and Cowtan, K. (2004). Coot: model-building tools for molecular graphics. Acta Cryst. D60, Brünger, A.T., Adams, P.D., Clore, G.M., DeLano, W.L., Gros, P., Grosse-Kunstleve, R.W., Jiang, J.S., Kuszewski, J., Nilges, M., Pannu, N.S., Read, R.J., Rice, L.M., Simonson, T. and Warren, G. L. (1998) Crystallography and NMR System: a new software suite for macromolecular structure determination. Acta Cryst. D54, Delano, W. L. (2008) The PyMol Molecular Graphics System, DeLano Scientific LLC, Palo Alto, CA ( 14
16 Table 1 Crystal structure data and refinement statistics CARM1 complex CMPD-1 CMPD-2 PDB ID 2Y1W 2Y1X Data collection Space Group P P Cell Parameters (Å) a b c X-ray source ID14-4 ESRF ID14-1 ESRF Resolution (Å) No. of observations Total 282, ,993 Unique 77,014 61,230 Completeness (%) 85.4 (30.0) 99.4 (99.9) Rsym (0.493) (0.598) I/σ I 14.1 (1.7) 9.4 (2.1) Refinement Resolution (Å) No. of reflections Working set (%) 73,115 (80.2) 58,154 (94.4) Test set (%) 3,876 (5.0) 3,057 (5.0) R cryst / R free 0.205/ / R.m.s. deviations Bond lengths (Å) Bond angles ( ) Ramachandran plot (%) Most favored Allowed region Outlier region
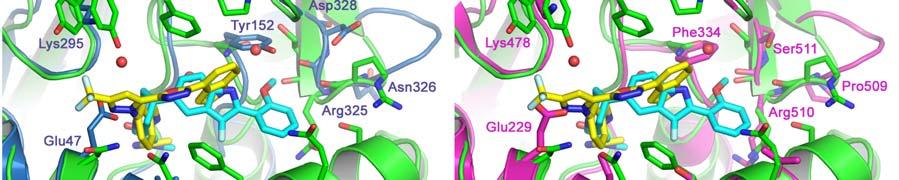
17 FIGURE CAPTIONS Figure 1. Sequence alignment for human PRMT1, PRMT3 and CARM1. Secondary structure elements corresponding to the CARM1 structure are shown below the sequences and the four characteristic PRMT motifs are shown above the sequences. Figure 2. Chemical structures of CMPD-1, CMPD-2 and CMPD-3. Figure 3. ITC results for titrations of CARM1 with CMPD-1 and CMPD-3. Titrations with CMPD-1 and CMPD-3 were done in the absence and presence of 0.2 mm SAH and binding was observed only in the presence of SAH. From left to right there are the titrations of CMPD-1 without SAH, CMPD-1 with SAH, CMPD-2 without SAH and CMPD-2 with SAH, respectively. Figure 4. Electron density associated with CMPD-1 and CMPD-2 in the CARM1 structures. Diagrams showing the SigmaA weighted difference omit maps associated with the inhibitors CMPD-1 (Left Image) and CMPD-2 (Right Image) bound in the CARM1 active site contoured at the 3 σ level. Inhibitors are shown with light-blue carbon atoms and cofactors (SNF or SAH) are shown with orange carbon atoms. Figure 5. Ribbon and surface diagrams of CARM1 with cofactors and inhibitors. (A) Ribbon diagram showing the overall architecture of CARM1 with CMPD-1 (light-blue carbon atoms) and SNF (orange carbon atoms) bound in the active site. The N-terminal domain is shown in yellow and C-terminal domain is shown in green. (B) Ribbon diagram showing the overall architecture of CARM1 with CMPD-2 (light-blue carbon atoms) and SAH (orange carbon atoms) bound in the active site. (C) Close up view of CARM1 SNF CMPD-1 with labels for select elements of secondary structure. The β4 strand is not visible in the diagram since it is hidden behind αb. (D) Comparison of the binding modes of CMPD-1 and CMPD-2 that result from the superposition of CARM1 SNF CMPD-1 onto CARM1 SAH CMPD-2. The CARM1 residues and the surface of the Arg-binding cavity are from the structure of CARM1 SAH CMPD-2. For clarity, the front part of the protein has been removed to provide an unobstructed view of the Arg-binding cavity. Figure 6. Views of CMPD-1 and CMPD-2 bound in the active site of CARM1. (A) Close-up view of CMPD-1 (light-blue carbon atoms) and SNF (orange carbon atoms) bound in the active site of CARM1. Ordered water molecules are shown as red spheres and hydrogen bonds as dashed lines. (B) Close-up view of CMPD-2 (light-blue carbon atoms) and SAH (orange carbon atoms) bound in the active site of CARM1. (C) Diagram showing the active sites of PRMT1 (blue carbon atoms) and CARM1 (green carbon atoms) resulting from the superposition of PRMT1 (PDB code 1OR8) onto CARM1 SAH CMPD-2. The residues shown are believed to be responsible for the CARM1 selectivity of CMPD-1 (cyan carbon atoms) and CMPD-2 (yellow carbon atoms). When present the labels indicate the identity of the corresponding PRMT1 residues. Several of the PRMT1 residues are partially or totally disordered in the 1OR8 structure. Ordered water molecules from the CARM1 SAH CMPD-2 16
18 structure are shown as red spheres. (D) Diagram showing the active sites of PRMT3 (magenta carbon atoms) and CARM1 (green carbon atoms) resulting from the superposition of PRMT3 (PDB code 1F3L) onto CARM1 SAH CMPD-2. The residues shown are believed to be responsible for the CARM1 selectivity of CMPD-1 (cyan carbon atoms) and CMPD-2 (yellow carbon atoms). The labels indicate the identity of the corresponding PRMT3 residues. Ordered water molecules from the CARM1 SAH CMPD-2 structure are shown as red spheres. 17

19 Figure 1 Figure 2 18


20 Figure 3 Figure 4 19

21 Figure 5 20



22 Figure 6 21
Structural basis for CARM1 inhibition by indole and pyrazole inhibitors
 Biochem. J. (2011) 436, 331 339 (Printed in Great Britain) doi:10.1042/bj20102161 331 Structural basis for CARM1 inhibition by indole and pyrazole inhibitors John S. SACK*, Sandrine THIEFFINE, Tiziano
Biochem. J. (2011) 436, 331 339 (Printed in Great Britain) doi:10.1042/bj20102161 331 Structural basis for CARM1 inhibition by indole and pyrazole inhibitors John S. SACK*, Sandrine THIEFFINE, Tiziano
Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid
 Amino acid") Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and
 Materials and Methods Immunoprecipitation and immunoblot analysis Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and immunoblot analyses with corresponding
Materials and Methods Immunoprecipitation and immunoblot analysis Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and immunoblot analyses with corresponding
Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the
 Supplementary information, Data S1 Methods Clones and protein preparation Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the Saccharomyces cerevisiae genomic DNA by polymerase chain
Supplementary information, Data S1 Methods Clones and protein preparation Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the Saccharomyces cerevisiae genomic DNA by polymerase chain
Protein expression and purification from Hi5 insect cells is as described (1, 2).
.") Materials & methods Protein expression and purification Protein expression and purification from Hi5 insect cells is as described (1, 2). The purified complex exhibited a 2:2:2 stoichiometry as determined
Materials & methods Protein expression and purification Protein expression and purification from Hi5 insect cells is as described (1, 2). The purified complex exhibited a 2:2:2 stoichiometry as determined
Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination
 201 Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination Acknowlegements. The structure of the D-8-Ad:P450cam conjugate was determined by Anna-Maria A. Hays.
201 Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination Acknowlegements. The structure of the D-8-Ad:P450cam conjugate was determined by Anna-Maria A. Hays.
The Skap-hom Dimerization and PH Domains Comprise
 Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
Supplemental Material
 Supplemental Material Molecular basis for oncohistone H3 recognition by SETD2 methyltransferase Shuang Yang, 1, 2 Xiangdong Zheng, 1, 2, 3 Chao Lu, 4 Guo-Min Li, 2 C. David Allis, 4 1, 2, 3, 5* and Haitao
Supplemental Material Molecular basis for oncohistone H3 recognition by SETD2 methyltransferase Shuang Yang, 1, 2 Xiangdong Zheng, 1, 2, 3 Chao Lu, 4 Guo-Min Li, 2 C. David Allis, 4 1, 2, 3, 5* and Haitao
Supplementary Note 1. Enzymatic properties of the purified Syn BVR
 Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Supplemental Information. The structural basis of R Spondin recognition by LGR5 and RNF43
 Supplemental Information The structural basis of R Spondin recognition by LGR5 and RNF43 Po Han Chen 1, Xiaoyan Chen 1, Deyu Fang 2, Xiaolin He 1* 1 Department of Molecular Pharmacology and Biological
Supplemental Information The structural basis of R Spondin recognition by LGR5 and RNF43 Po Han Chen 1, Xiaoyan Chen 1, Deyu Fang 2, Xiaolin He 1* 1 Department of Molecular Pharmacology and Biological
Molecular design principles underlying β-strand swapping. in the adhesive dimerization of cadherins
 Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%)
") 1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%) DESCRIPTION Nickel NTA Magnetic Agarose Beads are products that allow rapid and easy small-scale purification of
1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%) DESCRIPTION Nickel NTA Magnetic Agarose Beads are products that allow rapid and easy small-scale purification of
Solutions to 7.02 Quiz II 10/27/05
 Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
Lecture 7: Affinity Chromatography-II
 Lecture 7: Affinity Chromatography-II We have studied basics of affinity purification during last lecture. The current lecture is continuation of last lecture and we will cover following: 1. Few specific
Lecture 7: Affinity Chromatography-II We have studied basics of affinity purification during last lecture. The current lecture is continuation of last lecture and we will cover following: 1. Few specific
Supplemental Online Material
 Supplemental Online Material Supplemental Figures Supplemental Figure 1. A) Crosslinking efficiencies of 5 -biotin tagged oligonucleotides screened with the FASTDXL method 1. Reactive and nonreactive cysteine
Supplemental Online Material Supplemental Figures Supplemental Figure 1. A) Crosslinking efficiencies of 5 -biotin tagged oligonucleotides screened with the FASTDXL method 1. Reactive and nonreactive cysteine
BACTERIAL PRODUCTION EXPRESSION METHOD OVERVIEW: PEF # GENE NAME EXPRESSION VECTOR MOLECULAR WEIGHT kda (full-length) 34.
 34.") BACTERIAL PRODUCTION PEF # GENE NAME EXPRESSION VECTOR MOLECULAR WEIGHT 2015-XXXX XXXX pet-32a 50.9 kda (full-length) 34.0 kda (cleaved) EXPRESSION METHOD OVERVIEW: Plasmid DNA was transformed into BL21
BACTERIAL PRODUCTION PEF # GENE NAME EXPRESSION VECTOR MOLECULAR WEIGHT 2015-XXXX XXXX pet-32a 50.9 kda (full-length) 34.0 kda (cleaved) EXPRESSION METHOD OVERVIEW: Plasmid DNA was transformed into BL21
Figure S2, related to Figure 1. Stereo images of the CarD/RNAP complex and. electrostatic potential surface representation of the CarD/RNAP interface
 Structure, Volume 21 Supplemental Information Structure of the Mtb CarD/RNAP -Lobes Complex Reveals the Molecular Basis of Interaction and Presents a Distinct DNA-Binding Domain for Mtb CarD Gulcin Gulten
Structure, Volume 21 Supplemental Information Structure of the Mtb CarD/RNAP -Lobes Complex Reveals the Molecular Basis of Interaction and Presents a Distinct DNA-Binding Domain for Mtb CarD Gulcin Gulten
SUPPLEMENTARY INFORMATION
 Structure of a tyrosyl-trna synthetase splicing factor bound to a group I intron RNA Paul J. Paukstelis 1, Jui-Hui Chen 2, Elaine Chase 2, Alan M. Lambowitz 1,*, and Barbara L. Golden 2,*,. 1 Institute
Structure of a tyrosyl-trna synthetase splicing factor bound to a group I intron RNA Paul J. Paukstelis 1, Jui-Hui Chen 2, Elaine Chase 2, Alan M. Lambowitz 1,*, and Barbara L. Golden 2,*,. 1 Institute
BMB/Bi/Ch 170 Fall 2017 Problem Set 1: Proteins I
 BMB/Bi/Ch 170 Fall 2017 Problem Set 1: Proteins I Please use ray-tracing feature for all the images you are submitting. Use either the Ray button on the right side of the command window in PyMOL or variations
BMB/Bi/Ch 170 Fall 2017 Problem Set 1: Proteins I Please use ray-tracing feature for all the images you are submitting. Use either the Ray button on the right side of the command window in PyMOL or variations
Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein
 Cell Wall Recycling Protein") Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
Molekulare Mechanismen der Signaltransduktion
 Molekulare Mechanismen der Signaltransduktion 12 Mechanism of auxin perception slides: http://tinyurl.com/modul-mms doi: 10.1038/nature05731 previous model http://www.plantcell.org/content/17/9/2425/f1.large.jpg
Molekulare Mechanismen der Signaltransduktion 12 Mechanism of auxin perception slides: http://tinyurl.com/modul-mms doi: 10.1038/nature05731 previous model http://www.plantcell.org/content/17/9/2425/f1.large.jpg
Purification: Step 1. Protein and Peptide Chemistry. Lecture 11. Big Problem: Crude extract is not the natural environment. Cells: Break them open!
 Lecture 11 Protein and Peptide Chemistry Margaret A. Daugherty Fall 2003 Purification: Step 1 Cells: Break them open! Crude Extract Total contents of cell Big Problem: Crude extract is not the natural
Lecture 11 Protein and Peptide Chemistry Margaret A. Daugherty Fall 2003 Purification: Step 1 Cells: Break them open! Crude Extract Total contents of cell Big Problem: Crude extract is not the natural
Purification: Step 1. Lecture 11 Protein and Peptide Chemistry. Cells: Break them open! Crude Extract
 Purification: Step 1 Lecture 11 Protein and Peptide Chemistry Cells: Break them open! Crude Extract Total contents of cell Margaret A. Daugherty Fall 2003 Big Problem: Crude extract is not the natural
Purification: Step 1 Lecture 11 Protein and Peptide Chemistry Cells: Break them open! Crude Extract Total contents of cell Margaret A. Daugherty Fall 2003 Big Problem: Crude extract is not the natural
The YTH domain (residues ) of human YTHDF2 (NP_ ) was subcloned
 of human YTHDF2 (NP_ ) was subcloned") Supplementary information, Data S1 Materials and Methods Protein Expression, Purification and Crystallization The YTH domain (residues 383-553) of human YTHDF2 (NP_057342.2) was subcloned into a modified
Supplementary information, Data S1 Materials and Methods Protein Expression, Purification and Crystallization The YTH domain (residues 383-553) of human YTHDF2 (NP_057342.2) was subcloned into a modified
Supporting Information
 Supporting Information Slep et al. 10.1073/pnas.0801569105 SI Materials and Methods Mouse RGS16, residues 53 180, was subcloned into a modified pgex-2t vector (GE Healthcare) to yield a thrombin-cleavable
Supporting Information Slep et al. 10.1073/pnas.0801569105 SI Materials and Methods Mouse RGS16, residues 53 180, was subcloned into a modified pgex-2t vector (GE Healthcare) to yield a thrombin-cleavable
Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1.
 GR24 (μm) 0 20 0 20 GST-ShHTL7 anti-gst His-MAX2 His-COI1 PVDF staining Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1. Pull-down assays using GST-ShHTL7
GR24 (μm) 0 20 0 20 GST-ShHTL7 anti-gst His-MAX2 His-COI1 PVDF staining Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1. Pull-down assays using GST-ShHTL7
Protein Structure/Function Relationships
 Protein Structure/Function Relationships W. M. Grogan, Ph.D. OBJECTIVES 1. Describe and cite examples of fibrous and globular proteins. 2. Describe typical tertiary structural motifs found in proteins.
Protein Structure/Function Relationships W. M. Grogan, Ph.D. OBJECTIVES 1. Describe and cite examples of fibrous and globular proteins. 2. Describe typical tertiary structural motifs found in proteins.
Secondary structure of hvps4b (above) and sequence alignments of VPS4 proteins from
 and sequence alignments of VPS4 proteins from") Supplemental Figure 1. Secondary structure of hvps4b (above) and sequence alignments of VPS4 proteins from different species as well as four other representative members of the meiotic clade of AAA ATPases.
Supplemental Figure 1. Secondary structure of hvps4b (above) and sequence alignments of VPS4 proteins from different species as well as four other representative members of the meiotic clade of AAA ATPases.
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction
 X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
Lecture 8: Affinity Chromatography-III
 Lecture 8: Affinity Chromatography-III Key words: Chromatography; Affinity chromatography; Protein Purification During this lecture, we shall be studying few more examples of affinity chromatography. The
Lecture 8: Affinity Chromatography-III Key words: Chromatography; Affinity chromatography; Protein Purification During this lecture, we shall be studying few more examples of affinity chromatography. The
Purification of (recombinant) proteins. Pekka Lappalainen, Institute of Biotechnology, University of Helsinki
 proteins. Pekka Lappalainen, Institute of Biotechnology, University of Helsinki") Purification of (recombinant) proteins Pekka Lappalainen, Institute of Biotechnology, University of Helsinki Physical properties of proteins that can be applied for purification -size -charge (isoelectric
Purification of (recombinant) proteins Pekka Lappalainen, Institute of Biotechnology, University of Helsinki Physical properties of proteins that can be applied for purification -size -charge (isoelectric
Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport
 Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport Ivan Laponogov 1,2, Dennis A. Veselkov 1, Isabelle M-T. Crevel
Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport Ivan Laponogov 1,2, Dennis A. Veselkov 1, Isabelle M-T. Crevel
Hmwk # 8 : DNA-Binding Proteins : Part II
 The purpose of this exercise is : Hmwk # 8 : DNA-Binding Proteins : Part II 1). to examine the case of a tandem head-to-tail homodimer binding to DNA 2). to view a Zn finger motif 3). to consider the case
The purpose of this exercise is : Hmwk # 8 : DNA-Binding Proteins : Part II 1). to examine the case of a tandem head-to-tail homodimer binding to DNA 2). to view a Zn finger motif 3). to consider the case
Supplementary Information for. Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction
 Supplementary Information for Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction Takamasa Teramoto, Yukari Fujikawa, Yoshirou Kawaguchi, Katsuhisa
Supplementary Information for Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction Takamasa Teramoto, Yukari Fujikawa, Yoshirou Kawaguchi, Katsuhisa
Stabilization of a virus-like particle and its application as a nanoreactor at physiological conditions
 Supporting Information Stabilization of a virus-like particle and its application as a nanoreactor at physiological conditions Lise Schoonen, b Sjors Maassen, b Roeland J. M. Nolte b and Jan C. M. van
Supporting Information Stabilization of a virus-like particle and its application as a nanoreactor at physiological conditions Lise Schoonen, b Sjors Maassen, b Roeland J. M. Nolte b and Jan C. M. van
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Domain architecture and conformational states of the decapping complex, as revealed by structural studies. (a) Domain organization of Schizosaccharomyces pombe (Sp) and Saccharomyces
Supplementary Figure 1 Domain architecture and conformational states of the decapping complex, as revealed by structural studies. (a) Domain organization of Schizosaccharomyces pombe (Sp) and Saccharomyces
SUMOstar Gene Fusion Technology
 Gene Fusion Technology NEW METHODS FOR ENHANCING FUNCTIONAL PROTEIN EXPRESSION AND PURIFICATION IN INSECT CELLS White Paper June 2007 LifeSensors Inc. 271 Great Valley Parkway Malvern, PA 19355 www.lifesensors.com
Gene Fusion Technology NEW METHODS FOR ENHANCING FUNCTIONAL PROTEIN EXPRESSION AND PURIFICATION IN INSECT CELLS White Paper June 2007 LifeSensors Inc. 271 Great Valley Parkway Malvern, PA 19355 www.lifesensors.com
SUPPLEMENTARY INFORMATION Figures. Supplementary Figure 1 a. Page 1 of 30. Nature Chemical Biology: doi: /nchembio.2528
 SUPPLEMENTARY INFORMATION Figures Supplementary Figure 1 a b c Page 1 of 0 11 Supplementary Figure 1: Biochemical characterisation and binding validation of the reversible USP inhibitor 1. a, Biochemical
SUPPLEMENTARY INFORMATION Figures Supplementary Figure 1 a b c Page 1 of 0 11 Supplementary Figure 1: Biochemical characterisation and binding validation of the reversible USP inhibitor 1. a, Biochemical
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor
 Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Focus concept Purification of a novel seed storage protein allows sequence analysis and determination of the protein
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Focus concept Purification of a novel seed storage protein allows sequence analysis and determination of the protein
From mechanism to medicne
 From mechanism to medicne a look at proteins and drug design Chem 342 δ δ δ+ M 2009 δ+ δ+ δ M Drug Design - an Iterative Approach @ DSU Structural Analysis of Receptor Structural Analysis of Ligand-Receptor
From mechanism to medicne a look at proteins and drug design Chem 342 δ δ δ+ M 2009 δ+ δ+ δ M Drug Design - an Iterative Approach @ DSU Structural Analysis of Receptor Structural Analysis of Ligand-Receptor
SUPPLEMENTARY INFORMATION. Design and Characterization of Bivalent BET Inhibitors
 SUPPLEMENTARY INFORMATION Design and Characterization of Bivalent BET Inhibitors Minoru Tanaka 1,2,#, Justin M. Roberts 1,#, Hyuk-Soo Seo 3, Amanda Souza 1, Joshiawa Paulk 1, Thomas G. Scott 1, Stephen
SUPPLEMENTARY INFORMATION Design and Characterization of Bivalent BET Inhibitors Minoru Tanaka 1,2,#, Justin M. Roberts 1,#, Hyuk-Soo Seo 3, Amanda Souza 1, Joshiawa Paulk 1, Thomas G. Scott 1, Stephen
Index 1. Product Description 2. Purification Procedure 3. Troubleshooting 4. Ordering Information
 High Affinity Ni-Charged Resin Cat. No. L00223 Technical Manual No. TM0217 Version 07132010 Index 1. Product Description 2. Purification Procedure 3. Troubleshooting 4. Ordering Information 1. Product
High Affinity Ni-Charged Resin Cat. No. L00223 Technical Manual No. TM0217 Version 07132010 Index 1. Product Description 2. Purification Procedure 3. Troubleshooting 4. Ordering Information 1. Product
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 29 September 2005
 Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 9 September 005 Focus concept Purification of a novel seed storage protein allows sequence analysis and
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 9 September 005 Focus concept Purification of a novel seed storage protein allows sequence analysis and
Supporting Online Material. Av1 and Av2 were isolated and purified under anaerobic conditions according to
 Supporting Online Material Materials and Methods Av1 and Av2 were isolated and purified under anaerobic conditions according to published protocols (S1). Crystals of nf-, pcp- and adp-av2:av1 complexes
Supporting Online Material Materials and Methods Av1 and Av2 were isolated and purified under anaerobic conditions according to published protocols (S1). Crystals of nf-, pcp- and adp-av2:av1 complexes
Introduction to Protein Purification
 Introduction to Protein Purification 1 Day 1) Introduction to Protein Purification. Input for Purification Protocol Development - Guidelines for Protein Purification Day 2) Sample Preparation before Chromatography
Introduction to Protein Purification 1 Day 1) Introduction to Protein Purification. Input for Purification Protocol Development - Guidelines for Protein Purification Day 2) Sample Preparation before Chromatography
BC 367, Exam 2 November 13, Part I. Multiple Choice (3 pts each)- Please circle the single best answer.
- Please circle the single best answer.") Name BC 367, Exam 2 November 13, 2008 Part I. Multiple Choice (3 pts each)- Please circle the single best answer. 1. The enzyme pyruvate dehydrogenase catalyzes the following reaction. What kind of enzyme
Name BC 367, Exam 2 November 13, 2008 Part I. Multiple Choice (3 pts each)- Please circle the single best answer. 1. The enzyme pyruvate dehydrogenase catalyzes the following reaction. What kind of enzyme
Enhancers. Activators and repressors of transcription
 Enhancers Can be >50 kb away from the gene they regulate. Can be upstream from a promoter, downstream from a promoter, within an intron, or even downstream of the final exon of a gene. Are often cell type
Enhancers Can be >50 kb away from the gene they regulate. Can be upstream from a promoter, downstream from a promoter, within an intron, or even downstream of the final exon of a gene. Are often cell type
K Vinodhini, K Srinivasan
 Morphological Changes of α-lactose Monohydrate (α-lm) Single Crystals under Different Crystallization Conditions Using Polar Protic and Aprotic Solvents K Vinodhini, K Srinivasan To cite this version:
Morphological Changes of α-lactose Monohydrate (α-lm) Single Crystals under Different Crystallization Conditions Using Polar Protic and Aprotic Solvents K Vinodhini, K Srinivasan To cite this version:
High-Affinity Ni-NTA Resin
 High-Affinity Ni-NTA Resin Technical Manual No. 0217 Version 20070418 I Description.... 1 II Key Features... 1 III His-Tagged Fusion Protein Purification Procedure.. 1 IV Resin Regeneration. 4 V Troubleshooting...
High-Affinity Ni-NTA Resin Technical Manual No. 0217 Version 20070418 I Description.... 1 II Key Features... 1 III His-Tagged Fusion Protein Purification Procedure.. 1 IV Resin Regeneration. 4 V Troubleshooting...
SUPPLEMENTARY INFORMATION
 Molecular basis of RNA-dependent RNA polymerase II activity Elisabeth Lehmann, Florian Brueckner, and Patrick Cramer Gene Center Munich and Center for integrated Protein Science CiPS M, Department of Chemistry
Molecular basis of RNA-dependent RNA polymerase II activity Elisabeth Lehmann, Florian Brueckner, and Patrick Cramer Gene Center Munich and Center for integrated Protein Science CiPS M, Department of Chemistry
High-Affinity Ni-NTA Resin
 High-Affinity Ni-NTA Resin Technical Manual No. 0237 Version 20070418 I Description.. 1 II Key Features... 1 III His-Tagged Fusion Protein Purification Procedure 1 IV Resin Regeneration. 4 V Troubleshooting...
High-Affinity Ni-NTA Resin Technical Manual No. 0237 Version 20070418 I Description.. 1 II Key Features... 1 III His-Tagged Fusion Protein Purification Procedure 1 IV Resin Regeneration. 4 V Troubleshooting...
Clamping down on pathogenic bacteria how to shut down a key DNA polymerase complex
 Clamping down on pathogenic bacteria how to shut down a key DNA polymerase complex Bacterial DNA-replication machinery Pathogenic bacteria that are resistant to the current armoury of antibiotics are an
Clamping down on pathogenic bacteria how to shut down a key DNA polymerase complex Bacterial DNA-replication machinery Pathogenic bacteria that are resistant to the current armoury of antibiotics are an
Glutathione Agarose Resin User s Guide
 Glutathione Agarose Resin User s Guide DESCRIPTION Glutathione Agarose Resin is used to purify recombinant derivatives of glutathione S-transferases or glutathione binding proteins. Resins are products
Glutathione Agarose Resin User s Guide DESCRIPTION Glutathione Agarose Resin is used to purify recombinant derivatives of glutathione S-transferases or glutathione binding proteins. Resins are products
Structure formation and association of biomolecules. Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München
 Technische Universität München") Structure formation and association of biomolecules Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München Motivation Many biomolecules are chemically synthesized
Structure formation and association of biomolecules Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München Motivation Many biomolecules are chemically synthesized
5.36 Biochemistry Laboratory Spring 2009
 MIT OpenCourseWare http://ocw.mit.edu 5.36 Biochemistry Laboratory Spring 2009 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. Laboratory Manual for URIECA
MIT OpenCourseWare http://ocw.mit.edu 5.36 Biochemistry Laboratory Spring 2009 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. Laboratory Manual for URIECA
1) The penicillin family of antibiotics, discovered by Alexander Fleming in 1928, has the following general structure: O O
 The penicillin family of antibiotics, discovered by Alexander Fleming in 1928, has the following general structure: O O") ame: TF ame: LS1a Fall 06 Problem Set #3 Due Friday 10/13 at noon in your TF s drop box on the 2 nd floor of the Science Center All questions including the (*extra*) ones should be turned in 1) The penicillin
ame: TF ame: LS1a Fall 06 Problem Set #3 Due Friday 10/13 at noon in your TF s drop box on the 2 nd floor of the Science Center All questions including the (*extra*) ones should be turned in 1) The penicillin
Protein analysis. Dr. Mamoun Ahram Summer semester, Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5
 Protein analysis Dr. Mamoun Ahram Summer semester, 2015-2016 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5 Bases of protein separation Proteins can be purified on the basis Solubility
Protein analysis Dr. Mamoun Ahram Summer semester, 2015-2016 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5 Bases of protein separation Proteins can be purified on the basis Solubility
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION A human XRCC4-XLF complex bridges DNA ends. Sara N. Andres 1, Alexandra Vergnes 2, Dejan Ristic 3, Claire Wyman 3, Mauro Modesti 2,4, and Murray Junop 2,4 1 Department of Biochemistry
SUPPLEMENTARY INFORMATION A human XRCC4-XLF complex bridges DNA ends. Sara N. Andres 1, Alexandra Vergnes 2, Dejan Ristic 3, Claire Wyman 3, Mauro Modesti 2,4, and Murray Junop 2,4 1 Department of Biochemistry
SERVA Ni-NTA Magnetic Beads
 INSTRUCTION MANUAL SERVA Ni-NTA Magnetic Beads Magnetic beads for Affinity Purification of His-Tag Fusion Proteins (Cat. No. 42179) SERVA Electrophoresis GmbH - Carl-Benz-Str. 7-69115 Heidelberg Phone
INSTRUCTION MANUAL SERVA Ni-NTA Magnetic Beads Magnetic beads for Affinity Purification of His-Tag Fusion Proteins (Cat. No. 42179) SERVA Electrophoresis GmbH - Carl-Benz-Str. 7-69115 Heidelberg Phone
Nature Structural & Molecular Biology: doi: /nsmb.3018
 Supplementary Figure 1 Validation of genetic complementation assay in Bmal1 / Per2 Luc fibroblasts. (a) Only Bmal1, not Bmal2, rescues circadian rhythms from cells. Cells expressing various Bmal constructs
Supplementary Figure 1 Validation of genetic complementation assay in Bmal1 / Per2 Luc fibroblasts. (a) Only Bmal1, not Bmal2, rescues circadian rhythms from cells. Cells expressing various Bmal constructs
pt7ht vector and over-expressed in E. coli as inclusion bodies. Cells were lysed in 6 M
 Supplementary Methods MIG6 production, purification, inhibition, and kinase assays MIG6 segment 1 (30mer, residues 334 364) peptide was synthesized using standard solid-phase peptide synthesis as described
Supplementary Methods MIG6 production, purification, inhibition, and kinase assays MIG6 segment 1 (30mer, residues 334 364) peptide was synthesized using standard solid-phase peptide synthesis as described
STRUCTURAL BIOLOGY. α/β structures Closed barrels Open twisted sheets Horseshoe folds
 STRUCTURAL BIOLOGY α/β structures Closed barrels Open twisted sheets Horseshoe folds The α/β domains Most frequent domain structures are α/β domains: A central parallel or mixed β sheet Surrounded by α
STRUCTURAL BIOLOGY α/β structures Closed barrels Open twisted sheets Horseshoe folds The α/β domains Most frequent domain structures are α/β domains: A central parallel or mixed β sheet Surrounded by α
human Cdc45 Figure 1c. (c)
") 1 Details of the refined crystallographic model of human Cdc45 and comparison of its active-site region with that of bacterial RecJ. (a) Stereo view of a representative example of the final 2F o -F c electron
1 Details of the refined crystallographic model of human Cdc45 and comparison of its active-site region with that of bacterial RecJ. (a) Stereo view of a representative example of the final 2F o -F c electron
Supplementary information
 Supplementary information The E3 ligase RNF8 regulates KU80 removal and NHEJ repair Lin Feng 1, Junjie Chen 1 1 Department of Experimental Radiation Oncology, The University of Texas M. D. Anderson Cancer
Supplementary information The E3 ligase RNF8 regulates KU80 removal and NHEJ repair Lin Feng 1, Junjie Chen 1 1 Department of Experimental Radiation Oncology, The University of Texas M. D. Anderson Cancer
Supplementary Information
 Supplementary Information Supplementary Figures Figure S1. Study of mgtl translation in vitro. (A) Detection of 5 LR RNA using wild-type and anti-sd (91-95) substituted templates in a transcription-translation
Supplementary Information Supplementary Figures Figure S1. Study of mgtl translation in vitro. (A) Detection of 5 LR RNA using wild-type and anti-sd (91-95) substituted templates in a transcription-translation
Algorithms in Bioinformatics ONE Transcription Translation
 Algorithms in Bioinformatics ONE Transcription Translation Sami Khuri Department of Computer Science San José State University sami.khuri@sjsu.edu Biology Review DNA RNA Proteins Central Dogma Transcription
Algorithms in Bioinformatics ONE Transcription Translation Sami Khuri Department of Computer Science San José State University sami.khuri@sjsu.edu Biology Review DNA RNA Proteins Central Dogma Transcription
Supplemental Information
 Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Supplemental Information Experimental Procedures Cloning, expression, purification and mutagenesis
Electronic Supplementary Material (ESI) for RSC Advances. This journal is The Royal Society of Chemistry 2014 Supplemental Information Experimental Procedures Cloning, expression, purification and mutagenesis
BIOC 463A Expt. 4: Column Chromatographic Methods Column Chromatography
 Column Chromatography Chromatography is the process use to separate molecules based on SOME physical property of the molecule: Mass (i.e. size) Charge Affinity for ligands or substrates Hydrophobic interactions
Column Chromatography Chromatography is the process use to separate molecules based on SOME physical property of the molecule: Mass (i.e. size) Charge Affinity for ligands or substrates Hydrophobic interactions
High Purity Chromium Metal Oxygen Distribution (Determined by XPS and EPMA)
") High Purity Chromium Metal Oxygen Distribution (Determined by XPS and EPMA) K. Suzuki, H. Tomioka To cite this version: K. Suzuki, H. Tomioka. High Purity Chromium Metal Oxygen Distribution (Determined
High Purity Chromium Metal Oxygen Distribution (Determined by XPS and EPMA) K. Suzuki, H. Tomioka To cite this version: K. Suzuki, H. Tomioka. High Purity Chromium Metal Oxygen Distribution (Determined
pd861- NH Rham- His- ORF, Ecoli- El bp
 IP-Free E. coli Inducible Expression Vectors E. coli expression vectors are available with the following promoters: T5 or T7 (IPTG-inducible), rhabad (rhamnose-inducible), ara (arabinose and IPTG-inducible)
IP-Free E. coli Inducible Expression Vectors E. coli expression vectors are available with the following promoters: T5 or T7 (IPTG-inducible), rhabad (rhamnose-inducible), ara (arabinose and IPTG-inducible)
His-Spin Protein Miniprep
 INSTRUCTIONS His-Spin Protein Miniprep Catalog No. P2001 (10 purifications) and P2002 (50 purifications). Highlights Fast 5 minute protocol to purify His-tagged proteins from cell-free extracts Screen
INSTRUCTIONS His-Spin Protein Miniprep Catalog No. P2001 (10 purifications) and P2002 (50 purifications). Highlights Fast 5 minute protocol to purify His-tagged proteins from cell-free extracts Screen
Innovation Moléculaire et Thérapeutique, Université François Rabelais, Tours, France *For correspondence:
 Expression and Purification of the Eukaryotic MBP-MOS1 Transposase from sf21 Insect Cells Jérôme Jaillet, Audrey Dussaussois-Montagne, Sylvaine Renault and Corinne Augé-Gouillou * Innovation Moléculaire
Expression and Purification of the Eukaryotic MBP-MOS1 Transposase from sf21 Insect Cells Jérôme Jaillet, Audrey Dussaussois-Montagne, Sylvaine Renault and Corinne Augé-Gouillou * Innovation Moléculaire
(Refer Slide Time: 00:16)
") (Refer Slide Time: 00:16) Proteins and Gel-Based Proteomics Professor Sanjeeva Srivastava Department of Biosciences and Bioengineering Indian Institute of Technology, Bombay Mod 02 Lecture Number 5 Welcome
(Refer Slide Time: 00:16) Proteins and Gel-Based Proteomics Professor Sanjeeva Srivastava Department of Biosciences and Bioengineering Indian Institute of Technology, Bombay Mod 02 Lecture Number 5 Welcome
Solutions to Problem Set 1
 MIT Department of Biology 7.014 Introductory Biology, Spring 004 Question 1 Solutions to 7.014 Problem Set 1 a) Describe the conditions of the atmosphere on prebiotic earth and how these conditions differ
MIT Department of Biology 7.014 Introductory Biology, Spring 004 Question 1 Solutions to 7.014 Problem Set 1 a) Describe the conditions of the atmosphere on prebiotic earth and how these conditions differ
Proteomics. Manickam Sugumaran. Department of Biology University of Massachusetts Boston, MA 02125
 Proteomics Manickam Sugumaran Department of Biology University of Massachusetts Boston, MA 02125 Genomic studies produced more than 75,000 potential gene sequence targets. (The number may be even higher
Proteomics Manickam Sugumaran Department of Biology University of Massachusetts Boston, MA 02125 Genomic studies produced more than 75,000 potential gene sequence targets. (The number may be even higher
Drug DNA interaction. Modeling DNA ligand interaction of intercalating ligands
 Drug DNA interaction DNA as carrier of genetic information is a major target for drug interaction because of the ability to interfere with transcription (gene expression and protein synthesis) and DNA
Drug DNA interaction DNA as carrier of genetic information is a major target for drug interaction because of the ability to interfere with transcription (gene expression and protein synthesis) and DNA
Structural bases for N-glycan processing by mannosidephosphorylase
 Supporting information Volume 71 (2015) Supporting information for article: Structural bases for N-glycan processing by mannosidephosphorylase Simon Ladevèze, Gianluca Cioci, Pierre Roblin, Lionel Mourey,
Supporting information Volume 71 (2015) Supporting information for article: Structural bases for N-glycan processing by mannosidephosphorylase Simon Ladevèze, Gianluca Cioci, Pierre Roblin, Lionel Mourey,
TECHNICAL BULLETIN. Ni-CAM HC Resin High Capacity Nickel Chelate Affinity Matrix. Product No. N 3158 Storage Temperature 2 8 C
 Ni-CAM HC Resin High Capacity Nickel Chelate Affinity Matrix Product No. N 3158 Storage Temperature 2 8 C TECHNICAL BULLETIN Product Description Ni-CAM affinity resin (Ni-CAM) is an immobilized metal-ion
Ni-CAM HC Resin High Capacity Nickel Chelate Affinity Matrix Product No. N 3158 Storage Temperature 2 8 C TECHNICAL BULLETIN Product Description Ni-CAM affinity resin (Ni-CAM) is an immobilized metal-ion
Scanning the protein surface to uncover new small molecule. binding sites using fragments. Gregg Siegal ZoBio
 Scanning the protein surface to uncover new small molecule binding sites using fragments Gregg Siegal ZoBio Increasing Interest in Alternative Small Molecule Binding Sites 1. Enhanced specificity 2. Adress
Scanning the protein surface to uncover new small molecule binding sites using fragments Gregg Siegal ZoBio Increasing Interest in Alternative Small Molecule Binding Sites 1. Enhanced specificity 2. Adress
Specificity: Induced Fit
 Specificity: Induced Fit Conformational changes may occur upon ligand binding (Daniel Koshland in 1958) This adaptation is called the induced fit Induced fit allows for tighter binding of the ligand Induced
Specificity: Induced Fit Conformational changes may occur upon ligand binding (Daniel Koshland in 1958) This adaptation is called the induced fit Induced fit allows for tighter binding of the ligand Induced
INSTRUCTIONS The resins are adapted to work mainly in native conditions like denaturing.
 1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE Nickel NTA Agarose Beads DESCRIPTION Resins are products that allow batch or column purifications. This product is supplied as a suspension in 50% aqueous
1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE Nickel NTA Agarose Beads DESCRIPTION Resins are products that allow batch or column purifications. This product is supplied as a suspension in 50% aqueous
Kinetics Review. Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets)
") Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
Supplementary Figure S1. Binding of HSA mutants to hfcrn. (a) The levels of titrated amounts of HSA
 The levels of titrated amounts of HSA") Supplementary Figure S1. Binding of HSA mutants to hfcrn. (a) The levels of titrated amounts of HSA variants (5.0-0.002 μg/ml) directly coated in the wells at ph 6.0 were controlled using a horseradish
Supplementary Figure S1. Binding of HSA mutants to hfcrn. (a) The levels of titrated amounts of HSA variants (5.0-0.002 μg/ml) directly coated in the wells at ph 6.0 were controlled using a horseradish
NPTEL VIDEO COURSE PROTEOMICS PROF. SANJEEVA SRIVASTAVA
 LECTURE-06 PROTEIN PURIFICATION AND PEPTIDE ISOLATION USING CHROMATOGRAPHY TRANSCRIPT Welcome to the proteomics course. Today, we will talk about protein purification and peptide isolation using chromatography
LECTURE-06 PROTEIN PURIFICATION AND PEPTIDE ISOLATION USING CHROMATOGRAPHY TRANSCRIPT Welcome to the proteomics course. Today, we will talk about protein purification and peptide isolation using chromatography
Analysing protein protein interactions using a GST-fusion protein to pull down the interacting target from the cell lysate Hong Wang and Xin Zeng
 Analysing protein protein interactions using a GST-fusion protein to pull down the interacting target from the cell lysate Hong Wang and Xin Zeng Department of Molecular Genetics, Biochemistry and Microbiology,
Analysing protein protein interactions using a GST-fusion protein to pull down the interacting target from the cell lysate Hong Wang and Xin Zeng Department of Molecular Genetics, Biochemistry and Microbiology,
Figure A summary of spontaneous alterations likely to require DNA repair.
 DNA Damage Figure 5-46. A summary of spontaneous alterations likely to require DNA repair. The sites on each nucleotide that are known to be modified by spontaneous oxidative damage (red arrows), hydrolytic
DNA Damage Figure 5-46. A summary of spontaneous alterations likely to require DNA repair. The sites on each nucleotide that are known to be modified by spontaneous oxidative damage (red arrows), hydrolytic
AFFINITY GST PURIFICATION
 DESCRIPTION Glutathione Agarose Resin is used to purify recombinant derivatives of glutathione S-transferases or glutathione binding proteins. are products that allow batch or column purifications. Purification
DESCRIPTION Glutathione Agarose Resin is used to purify recombinant derivatives of glutathione S-transferases or glutathione binding proteins. are products that allow batch or column purifications. Purification
The Study of NF-κB Peptide Mimics and How Proteins Bind DNA
 Georgia Southern University Digital Commons@Georgia Southern University Honors Program Theses Student Research Papers 2016 The Study of NF-κB Peptide Mimics and How Proteins Bind DNA Allee M. Murray Georgia
Georgia Southern University Digital Commons@Georgia Southern University Honors Program Theses Student Research Papers 2016 The Study of NF-κB Peptide Mimics and How Proteins Bind DNA Allee M. Murray Georgia
nature methods Enabling IMAC purification of low abundance recombinant proteins from E. coli lysates
 nature methods Enabling IMAC purification of low abundance recombinant proteins from E. coli lysates Audur Magnusdottir, Ida Johansson, Lars-Göran Dahlgren, Pär Nordlund & Helena Berglund Supplementary
nature methods Enabling IMAC purification of low abundance recombinant proteins from E. coli lysates Audur Magnusdottir, Ida Johansson, Lars-Göran Dahlgren, Pär Nordlund & Helena Berglund Supplementary
Protocol for in vitro transcription
 Protocol for in vitro transcription Assemble the reaction at room temperature in the following order: Component 10xTranscription Buffer rntp T7 RNA Polymerase Mix grna PCR DEPC H 2 O volume 2μl 2μl 2μl
Protocol for in vitro transcription Assemble the reaction at room temperature in the following order: Component 10xTranscription Buffer rntp T7 RNA Polymerase Mix grna PCR DEPC H 2 O volume 2μl 2μl 2μl
AFFINITY HIS-TAG PURIFICATION
 DESCRIPTION Resins are products that allow batch or column purifications. This product is supplied as a suspension in 50% aqueous suspension containing 30 vol % ethanol. INSTRUCTIONS The resins are adapted
DESCRIPTION Resins are products that allow batch or column purifications. This product is supplied as a suspension in 50% aqueous suspension containing 30 vol % ethanol. INSTRUCTIONS The resins are adapted
has only one nucleotide, U20, between G19 and A21, while trna Glu CUC has two
 SPPLEMENTRY INFORMTION doi:1.138/nature9411 Supplementary Discussion The structural characteristics of trn Gln G in comparison to trn Glu 16 in trn Gln G is directed towards the G19 56 pair, or the outer
SPPLEMENTRY INFORMTION doi:1.138/nature9411 Supplementary Discussion The structural characteristics of trn Gln G in comparison to trn Glu 16 in trn Gln G is directed towards the G19 56 pair, or the outer
6-Foot Mini Toober Activity
 Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
OPPF-UK Standard Protocols: Insect Cell Purification
 OPPF-UK Standard Protocols: Insect Cell Purification Last Updated 6 th October 2016 Joanne Nettleship joanne@strubi.ox.ac.uk OPPF-UK SOP: Insect Cell Purification Table of Contents Suggested Schedule...
OPPF-UK Standard Protocols: Insect Cell Purification Last Updated 6 th October 2016 Joanne Nettleship joanne@strubi.ox.ac.uk OPPF-UK SOP: Insect Cell Purification Table of Contents Suggested Schedule...
SUPPLEMENTAL INFORMATION. Center Hamburg-Eppendorf, Center for Molecular Neurobiology, ZMNH, Hamburg, Germany
 SUPPLEMENTAL INFORMATION Title: Inter-motif communication induces hierarchical Ca 2+ -filling of Caldendrin Authors: Uday Kiran 1, Phanindranath Regur 1, Michael R. Kreutz 3,4*, Yogendra Sharma 1,2*, Asima
SUPPLEMENTAL INFORMATION Title: Inter-motif communication induces hierarchical Ca 2+ -filling of Caldendrin Authors: Uday Kiran 1, Phanindranath Regur 1, Michael R. Kreutz 3,4*, Yogendra Sharma 1,2*, Asima
Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis
 Supplemental material to accompany Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis Jacob Bauer, a Gabriela Ondrovičová, a Lucie Najmanová, b Vladimír Pevala,
Supplemental material to accompany Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis Jacob Bauer, a Gabriela Ondrovičová, a Lucie Najmanová, b Vladimír Pevala,
Structure/function relationship in DNA-binding proteins
 PHRM 836 September 22, 2015 Structure/function relationship in DNA-binding proteins Devlin Chapter 8.8-9 u General description of transcription factors (TFs) u Sequence-specific interactions between DNA
PHRM 836 September 22, 2015 Structure/function relationship in DNA-binding proteins Devlin Chapter 8.8-9 u General description of transcription factors (TFs) u Sequence-specific interactions between DNA
Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and
 and") Supplementary Tables Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and their side chain contacting residues in the second chain (B) a Interface Res. in Contacting
Supplementary Tables Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and their side chain contacting residues in the second chain (B) a Interface Res. in Contacting
Conformational changes in IgE contribute to its. uniquely slow dissociation rate from receptor FcεRI
 Conformational changes in IgE contribute to its uniquely slow dissociation rate from receptor FcεRI M.D. Holdom, A.M. Davies, J.E. Nettleship, S.C. Bagby, B. Dhaliwal, E. Girardi, J. Hunt, H.J. Gould,
Conformational changes in IgE contribute to its uniquely slow dissociation rate from receptor FcεRI M.D. Holdom, A.M. Davies, J.E. Nettleship, S.C. Bagby, B. Dhaliwal, E. Girardi, J. Hunt, H.J. Gould,