NAD kinases use substrate-assisted catalysis for specific recognition of NAD.
|
|
|
- Cory Singleton
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on August 8, 2007 as Manuscript M The latest version is at NAD kinases use substrate-assisted catalysis for specific recognition of NAD. Guillaume Poncet-Montange,*, Liliane Assairi, Stefan Arold, Sylvie Pochet, Gilles Labesse, Atelier de Bio- et Chimie Informatique Structurale, CNRS, UMR5048, Centre de Biochimie Structurale, F34090 Montpellier, France; INSERM, U554, F34090 Montpellier, France; Université Montpellier 1 et 2; IFR3, F34090 Montpellier, France, 29 rue de Navacelles Montpellier, France. INSERM, U759, Institut Curie, Orsay, France. Unité de Chimie Organique, Institut Pasteur, CNRS, URA 2128, Paris, France. Running title: Substrate-assisted catalysis of NAD kinases Address correspondance to: Gilles Labesse, Atelier de Bio- et Chimie Informatique Structurale, Centre de Biochimie Structurale, CNRS UMR INSERM U554 - Université Montpellier 1. Phone: ; Fax: labesse@cbs.cnrs.fr Here we describe the crystal structures of the NAD kinase (LmNADK1) from Listeria monocytogenes in complex with its substrate NAD, its product NADP or two synthesized NAD mimics. We identified one of the NAD mimics, di-adenosine diphosphate, as a new substrate for LmNADK1, while we showed that the closelyrelated compound, di-(5 -thio-adenosine) is a novel non-natural inhibitor for this enzyme. These structures suggest a mechanism involving substrate-assisted catalysis. Indeed, sequence/structure comparison and directed mutagenesis have previously shown that NAD kinases (NADKs) and the distantly related 6-phosphofructokinases (PFKs) share the same catalytically important GGDGT motif. However, in this study we have shown that these enzymes use the central aspartate of this motif differently. While this acidic residue chelates the catalytic Mg 2+ ion in PFKs, it activates the phosphoacceptor (NAD) in NADKs. Sequence/ structure comparisons suggest that the role of this aspartate would be conserved in NADKs and the related sphingosine and diacylglycerol kinases. With the emergence of antibiotic resistance worldwide, the search for new antibacterial compounds has become increasingly important (1). NADKs represent an attractive and novel drug target for antibiotic discovery. Indeed, NADKs (EC ) were recently shown to be essential for growth in many bacteria (2-4) including various human pathogens such as Mycobacterium tuberculosis (5), Staphylococcus aureus (6), Streptococcus pneumoniae (7), Salmonella enterica (8) and Pseudomonas aeruginosa (Jacobs, Urbach and Ausubel, personal communication). NADKs are ubiquitous enzymes involved in the last step of the biosynthesis of NADP catalyzing the transfer of a phosphate group on the substrate NAD to produce NADP in the presence of both ATP and Mg 2+. NADKs are also involved in the tight regulation of the NADH/NADPH ratio (8). Both NAD(P) and NAD(P)H are essential cofactors for a large number of enzymes involved in various metabolic pathways such as oxidoreductases. This activity of NADKs has been well known for decades, but their genes were cloned only recently (9), subsequently leading to rapid identification of NADK orthologs in many organisms from bacteria, archea, plants and human (Pfam PF01513). Bacterial and human enzymes show significant functional and sequence divergences and the NADK activity in human cells is low (10). However, despite recent advance in the characterization of NADKs, the reaction mechanism is not well understood, and some original features of NADKs remain unexplained: Firstly, during catalysis, the phosphorylation occurs specifically on 2'-hydroxyl of the adenosine moiety of NAD, and not on the neighboring 3 -hydroxyl. Secondly, the specificity for the catalytic dication is low (Mn 2+, Ca 2+, Zn 2+, Co 2+ and Fe 2+ appear equivalent to Mg 2+ ) (11) whereas it is generally high in other wellcharacterized kinases. Furthermore, the dication binding motif is yet unknown. Finally, most NADKs use as a phosphodonor various nucleoside triphosphates (9,12,13). Polyphosphates can also act as a donor but only for NADKs from Gram(+) bacteria and from mycobacteria (9). On the contrary, the human NADK counterpart preferentially uses ATP (7% activity with GTP) (10). The recent determination of the crystal structures of NADKs from Mycobacterium tuberculosis and Archaeoglobus fulgidus, unbound 11 1 Copyright 2007 by The American Society for Biochemistry and Molecular Biology, Inc.
2 or in the presence of NAD or NADP, allowed to more clearly locate the active site (14-16), but the precise reaction mechanism remained speculative (15). We now report crystal structures of NADK from the human pathogen Listeria monocytogenes (LmNADK1) 1, as a first representative of the enzymes from Gram(+) bacilli. The crystal structures of LmNADK1 ligand-free or in complex with its natural substrate or its product were solved at 2.3, 2.1 and 2.1 Å, respectively. Furthermore, we designed and synthesized two molecules mimicking NAD: di-adenosine diphosphate and di-(5'-thioadenosine). Additionally, we characterized two mutants whose function but not their structure was affected. Our analysis highlights important, and previously unnoticed, features of the NAD reaction mechanism: (i) We reveal the molecular basis for the 2 -hydroxyl specificity of NAD, (ii) we show that NADKs and the structurally related PFKs use the same catalytic GGDGT motif in a different manner and (iii) we suggest that the diphosphate group of the substrate plays the role of a dication chelator, revealing a mechanism involving substrate-assisted catalysis. EXPERIMENTAL PROCEDURES Chemicals, Bacterial strains, plasmids. The oligonucleotides were synthesized by MWG- Biotech. The restriction enzymes were purchased from Biolabs and Roche. Vent DNA polymerase, Tfu DNA polymerase was from Qbiogene. and T4 DNA ligase were purchased from Biolabs. Isopropyl-β-D-1-thiogalactopyranoside (IPTG) was from ICN Biomedical inc. Isocitrate dehydrogenase from porcine heart, sodium isocitrate, NAD, nucleosides triphosphates were from Sigma or Boehringer. The kits used for plasmid preparation were from either Sigma or Macherey-Nagel. Genomic DNA of Listeria monocytogenes strain EGD-e was a gift of P. Cossart (Institut Pasteur). The E. coli strains NM554 and BL21(DE3)/pDIA17 was purchased from Stratagen and Novagen. Cloning, site-directed mutagenesis and expression in E. coli. The NAD kinase 1 (EC ) coding sequence was amplified by PCR from the genomic DNA isolated from Listeria monocytogenes strain EGD-e, by using the Vent DNA polymerase, dntps and the following primers: 5'-Lm, 5 -GGAATTCCATATGAAATATATGATT ACTTCCAAAGGA-3 ' 3'-Lm, 5'-CGGCGCTCGAGTTAATCTTCAATAA ACGAATCGTGTAC-3' The amplified DNA was cloned into the expression vector pet22b (Novagen) at the NdeI and XhoI restriction sites (underlined) giving the following plasmid pla Site-directed mutagenesis was performed using the double PCR method by using the Tfu DNA polymerase the two above primers and the following mutagenic primers : D45N, 5 -AGACAGAAATGTTCCATTGCCACC AATAGAAAT-3 H223E, 5 -ATGCAAAATGCTCAGCTCGTCTAC AGAAATTTG-3 The final PCR products were purified and cloned into the pet22b vector at the NdeI and XhoI restriction sites. The mutated genes were sequenced and the corresponding plasmids were then used for transforming the E. coli strain BL21(DE3)/pDIA17 for protein expression. The transformants were grown at 37 C in 2YT medium in the presence of chloramphenicol and ampicillin at 30 µg/ml. When the absorbance reached at least 1.5 OD at 600 nm, the expression of the both recombinant proteins was then induced by the addition of 1 mm IPTG and the growth was continued for three more hours at 37 C. The cells were then pelleted by centrifugation and served as source for protein purification. For production of the selenomethionine derivative, selenomethionine was incorporated in place of methionine while the cells were grown in M9 minimum media supplemented with 20 mm glucose, 2 mm magnesium sulfate, ampicillin (30 µg/ml) and chloramphenicol (30 µg/ml) at 37 C. When the OD at 600 nm reached 0.5, the medium was supplemented with the amino acids lysine, threonine, phenylalanine at 100 mg/ml and leucine, isoleucine, valine and selenomethionine at 50 mg/ml. After 30 min, the expression was induced by addition of 0.5 mm IPTG for 16 hours at 37 C. Soluble protein was purified using cobalt-agarose affinity chromatography, followed by size exclusion chromatography. The protein was concentrated to 6 7 mg/ml in 25 mm sodium acetate, ph 5.2. The mutants were produced as described above for the wild-type enzyme and showed similar characteristics as monitored by SDS/PAGE, dynamic light scattering, and circular dichroïsm (data not shown). Enzymatic assay. The NAD kinase activity was determined by measuring the NADP produced by using isocitrate dehydrogenase as a coupling enzyme. Kinetic parameters for ATP were obtained by fitting data to the Michaelis-Menten equation. 22 2
3 Kinetics parameters for NAD were obtained by fitting data to the Hill equation. For determination of ADP produced, enzyme was incubated in 50 mm sodium phosphate ph 7.5, 100 mm NaCl, 10 mm MgCl2 with substrates or inhibitors at various concentrations for 30 min at 30 C. Cobalt resin was added to the sample for removal of the protein and centrifuged. The supernatant was tested for ADP production in 50 mm Tris-HCl ph 7.4, 50 mm KCl, 1 mm phosphoenolpyruvate, 0.2 mm NADH, 2 units each lactate dehydrogenase, pyruvate kinase and the decrease of absorbance was measured at 340 nm at 30 C using a spectrophotometer Eppendorf ECOM Phosphorylation of DTA was tested at rather high concentration of substrate (3mM) due to the low overall activity (no activity detected at 1 mm) of the wild-type and mutant enzymes. Crystallization. Initial crystallization conditions of LmNADK1 were found by using a hanging-dropbased sparse-matrix screening strategy. The best crystals were obtained by mixing 1 µl of the protein solution (concentration mg/ml) with an equal volume of crystallization buffer (0.3 M potassium chloride, 50 mm tri-sodium citrate dihydrate, ph 5.6, 15-20% w/v polyethylene glycol 400), equilibrated over 0.5 ml of the same buffer. Two independent crystal forms were grown in nearly identical conditions in the presence of 10 mm iodine (apo structure, space group P2 1 ) or in the presence (holo structure, space group I222) of 10 mm 5 -thioacetyl-adenosine (TAA, Fig. 1C). The complexes with bound NAD, NADP and NAD mimics were solved using crystals in the orthorhombic form. After growing them in presence of TAA, crystals were washed in the mother liquor and soaked with millimolar concentration of the desired ligand in a pre-equilibrated hanging-drop. Crystallographic Studies. X-ray diffraction data sets were collected from frozen single crystals at the European Synchrotron Radiation Facility (Grenoble, France, beamlines BM30A, ID14, and ID23) and processed with the programs MOSFLM, SCALA, and TRUNCATE from the CCP4 program suites (18) (Table 1 and Table 2). The structure was solved by using a multiple-wavelength anomalous diffraction experiment from a highly redundant 4 Å data set of the selenomethionine-labeled protein. Twenty-two of 24 Se sites were found by direct methods with the program SHAKE N'BAKE (19) and refined with the program SHARP (20). Electron density modification was performed with SOLOMON (20). A molecular replacement solution was obtained using the program PHASER (21) and the crystal structure PDB # 1U0R (14). Phases derived from MAD data and from molecular replacement were then combined using the program DMMULTI (22). Four-fold NCS was applyied in DMMULTI (22) and during the first steps of refinement in REFMAC5 as implemented in CCP4 (18). Iterative model rebuilding and refinement was performed first by using the program O (23) and then the program COOT (24) and the program REFMAC5, using a translation/libration/screw model (25) against the native data set at 2.4-Å resolution. In the final model, short segments of the protein (loops 69-TGHL-72, 110-GIGKK-114 and 129-SGGP-132) are not clearly visible in the electron density map. Similarly, the last residue and/or the affinity-tag (monomers A, C and D) or the last five histidines (monomer B), could not be modelled. Molecular replacement was performed to solve the first structure in orthorhombic form. Subsequent determinations were performed on isomorphous crystals. Refinement was performed using REFMAC5 using the same translation/libration/screw model. As in the case of the monoclinic form, some side chains and several short segments of the protein (mainly loops 69- TGHL-72, 110-GIGKK-114 and less frequently 129-SGGP-132) are not clearly visible in the electron density map. Similarly, the last residue and/or the affinity-tag could not be modelled in most protein-ligand complexes. Figures of ligands and corresponding electronic density were generated using PyMOL ( Chemical synthesis. Solvents were spectroscopic or HPLC grade. 1 H and 31 P NMR spectra were recorded on a Bruker Avance 400 spectrometer, operating at MHz and MHz, respectively. Preparative and analytical HPLC were carried out on a Perkin Elmer system (200 Pump) using a C18 reverse phase column (Kromasil, 5µ 100 Å) at a flow rate of 5.5 ml/min. or 1 ml/min and a linear gradient of CHCN in 20 mm triethylammonium acetate buffer at ph 7.5 over 20 min. Mass spectra were recorded at the mass spectroscopy laboratory (CNRS-ICSN, Gif-sur- Yvette). TAA was synthesized from commercially available 2',3'-isopropylidene-adenosine and thioacetic acid under Mitsunobu conditions as previously described (17). The acetonide group was then removed in aqueous formic acid to yield after purification by silica gel chromatography TAA in a total yield of 60%. 5'-thioadenosine (TA, Fig. 1C) 33 3
4 was obtained from TAA by hydrolysis using a mixture of MeOH/H2O saturated with ammonia under oxygen free conditions. The chemical structure of TA was confirmed by NMR and mass analysis, its purity (absence of disulfide formation) was checked by reverse phase HPLC analysis and compared with DTA as control. Di-adenosine diphosphate (Ap2A, Fig. 1B) was prepared by condensation of 5'-phosphomorpholidate of adenosine (26) and adenosine 5'-monophosphate as tributylammonium salt in anhydrous DMF. After purification by reverse phase HPLC, Ap2A was isolated as sodium salt in 50% yield. RESULTS Overall Structure and Topology Two independent crystal forms were grown in nearly identical conditions in the absence (apo structure, space group P2 1 ) or in the presence of an adenosine analogue (holo structure, space group I222) named 5 -thioacetyl-adenosine (hereafter TAA, Fig. 1C). First, we determined the apo structure of LmNADK1 in the P2 1 crystal form. Initial molecular replacement trials using 2.4 Å resolution data from native P2 1 crystals, and the structure of the mycobacterial NADK (14), which is only 24% identical in sequence to LmNADK1, yielded a solution with plausible crystal packing and score above noise level. However this model could not be easily refined. Structure determination was only successful once this molecular replacement solution were combined with low-resolution (4 Å) multiple-wavelength anomalous diffraction data from a selenomethionine derivative, and subjected to four-fold averaging. The resulting electron density allowed rebuilding and refinement of the LmNADK1 tetramer using native data to 2.4 Å resolution (Table 1). Then, the structure of the second crystal form (space group I222) was solved by molecular replacement using the native structure (Table 2). LmNADK1 is a homotetramer of approximate dimensions Å (Fig. 2). In the monoclinic crystal, one tetramer is observed in the asymmetric unit and corresponds to the biological tetramer. Only one monomer is present in the orthorhombic form and the biological tetramer was deduced from the crystallographic symmetries. The tetrameric arrangement is very similar to that described for the other NADKs, and corresponds to a dimer of dimers, stabilized by large interfaces (~3500 A²) between the C-terminal domains. The monomer structure of LmNADK1 is very similar to one of homologues from M. tuberculosis and A. fulgidus, showing root mean square deviations of 2.2 and 2.4 Å, respectively. Nevertheless, some structural features in the C- terminal domain appeared specific to the NADKs from Gram(+) bacilli, in particular a sequence motif 188-NnrvfR-193 in the vicinity of the bound nucleobases, corresponding to a small loop absent in other NADKs (Fig. S1). Within each monomer, the polypeptide chain is organized into an N-terminal and a C- terminal domain (Fig. 2). The C-terminal domain adopts an original topology composed of two β- sheets arranged in an antiparallel 12-stranded β- sandwich. It comprises most of the motifs strictly conserved among NADKs, including the motif 150- DG///sTPsGsTAY-163 involved in NAD recognition (27) (where '/' stands for any hydrophobic residue, lower/capital letters for partially/strictly conserved residues, respectively) (see supplemental data Fig. S1). The N-terminal domain is composed of 5 β- strands and 3 α-helices. In LmNADK1 and in its orthologs from other Gram(+) bacteria, the N- terminal domain is shorter than in other NADKs due to the loss of the second α-helix. This domain harbors the well-conserved motif GGDGT. Beside this motif, little conservation is observed in agreement with the low level of specificity for the phosphate donor (which can be polyphosphate or any nucleoside triphosphate). Binding of Natural Substrates to LmNADK1 To investigate the catalytic mechanism, we next tried to obtain crystal forms of LmNADK1 bound to NAD or NADP. Despite several attempts of soaking and co-crystallization with NAD or NADP, monoclinic P2 1 apo crystals failed to reveal electron density corresponding to these ligands. Soaking of NAD or NADP was finally successful with the second crystal form, I222. The ligand TAA was washed out, and successfully replaced by NAD or NADP. Since apo as well as NAD and NADP bound crystals yielded near 2 Å data sets (Table 2), the catalytic mechanism can be analyzed in more detail (Fig. S2 and S3). Despite the change in crystal forms, the overall structures of the apoenzyme and the bound states were highly similar, with only small reorientation of the two domains (Fig. 2), as well as of a few side chains in the substrate binding site (root mean square deviation of Å for isolated 44 4
5 N-terminal and C-terminal domains, versus Å for full-length chains). As in other NADKs, the dinucleosidediphosphate binding site of LmNADK1 was formed at the interface between the two domains. The conserved motif from the N-terminal domain faces those from the C-terminal domain at the edges of the interfacial groove. Two distinct binding pockets, named sites A and N, accommodate the adenosine and the nicotinamide-ribose, respectively (supplementary data Fig. S2 and S3). While the site A is composed of residues from only one monomer, the site N includes an aspartate (Asp-150' in LmNADK1) and an alanine (Ala-185') from a second monomer. In the site A (Fig. S2 and S3), the adenine ring is sandwiched between the side chain and the carboxylate of Ala-162 and part of the side chains of Asp-45 and Phe-74. Specific recognition of the adenine moiety involves mainly two residues: Asn- 122 and Thr-161. Their side chains are hydrogenbonded to the N6 amino group and the N7 and N3 nitrogens of the adenine. These two residues are well-conserved in most NADKs and participate in an intricate hydrogen-bond network involving the neighboring side chain of Tyr-75 and the Ser-158 carboxylate. In turn, the hydroxyl of Ser-158 is also hydrogen bonded to the hydroxyl of Tyr-75. On the contrary, few interactions are observed with the ribose moiety of adenosine beside the hydrogen bond between the Asp-45 and the 2 -hydroxyl of NAD. Disruption of this contact in LmNADK1- NADP complex is accompanied by a slight reorientation of the ribose (see below), while the 2 - phosphate group is hydrogen-bonded to the backbone of the second and third glycines of the motif 43-GGDGT-47. In the site N (Fig. S2 and S3), nicotinamide binding induced only a subtle rearrangement: the small rotation of the side chain of Tyr-163. The aromatic ring of the nicotinamide stacks onto the tyrosine ring. Both the hydroxyl of this tyrosine and the amide group of the nicotinamide are hydrogenbonded to a conserved Asp-150' from another subunit, as observed in other NADKs (15,16). The amide group of the nicotinamide forms additional hydrogen bonds with the mainchain carbonyl of Ala-185' and the side chain of Ser-166. Recognition of the ribose attached to the nicotinamide is ensured by three hydrogen bonds through its 2' and 3'- hydroxyl groups with the side chains of Asn-122 and Asp-123. This network of interactions ensures that the other natural dinucleoside diphosphate compound, FAD, will not enter the site N. While the recognition of the nucleobase moieties is made through various interactions, few contacts are observed between the protein and the two phosphate groups of the NAD molecule. The diphosphate link (group linking the nucleosides) is solvent-exposed and showed alternating conformations with significant mobility. Only one histidine (His-223) was hydrogen-bonded to one of the two phosphate groups (Fig. S2). The ribose of the adenosine moiety is also rather solvent-exposed. This raised the question of the basis for the specific 2' phosphorylation of NAD by NADKs. The role of the conserved GGDGT motif The conserved GGDGT motif of NADKs has been suggested to be catalytically important and to be involved in nucleotide binding (28) or in phosphate transfer (15). Previous sequence comparisons had revealed that the conserved motif GGDGT of NADKs corresponds to a consensus signature '/d/////ggdgs' which is well conserved in four kinase families: NAD kinases, sphingosine kinases, eukaryotic diacylglycerol kinases and phosphofructokinases (PFKs) (28). PFKs phosphorylate D-fructose 6-phosphate using ATP as a phosphodonor (29) (EC ; Pfam PF00365). From the crystal structure of PFKs, it was deduced that in the consensus sequence, glycines interact with the phosphates of the phospho-donor (ATP) and the second conserved aspartate is involved in chelation of the catalytic dication in order to activate the phospho-donor (29). Using our crystal structure, we performed structural comparison between the NADK-NAD and PFK-ATP complexes. It revealed that the two-domain organization of NADKs was reminiscent to those of PFKs. In both cases, the active site lies at the hinge separating the two domains. However, the C- terminal domains of NADKs and PFKs adopt two distinct architectures, whereas the N-terminal domains adopt a very similar fold (70 Cαs from their N-terminal domains superimpose with a root mean square deviation of 3.5 Å) (Fig. 3). Nevertheless, in PFKs and NADKs, the common GGDGT motif is similarly positioned at the twodomain interface, suggesting a similar role in catalysis. Surprisingly, in NADKs, the central aspartate is oriented in an opposite direction, incompatible with its role in PFKs (Fig. 3). Because this position is conserved in NADKs from M. tuberculosis and A. fulgidus, these data suggest that 55 5
6 even if NADKs and PFKs belong to the same superfamilly, their enzymatic mechanisms differ. The conformation of Asp-45 varies slightly between apo, NADP and NAD bound states of LmNADK1. In the unbound LmNADK1 structure or in the presence of the reaction product NADP, the Asp-45 of the GGDGT motif is hydrogen bonded to three backbone amide groups which belong to residues whose side chains are involved in adenine binding. In contrast, in LmNADK1-NAD, the side chain of Asp-45 becomes hydrogen bonded to 2'-hydroxyl group of NAD and two NADK backbone amide groups (residues 74-75). This rearrangement (Fig. S3) is accompanied by a concomitant change in the orientation of the ribose of the adenosine moiety (relative to its nucleobase). To investigate the role of Asp-45, we determined the crystal structure of the LmNADK1- D45N mutant, using the I222 TAA-bound crystal form. After washing and soaking with NAD, 2.2 Å data could be collected from these crystals (Table 2). The mutant structure showed only minor changes, and its active site was similar to that of the wild-type enzyme with the ligand present in the same conformation (see supplemental data Fig. S4). The asparagine adopted the same buried conformation as the aspartate, but did not form any hydrogen bond with NAD. The mutation Asp45Asn resulted in a tenfold decrease in activity (Table 3). Asp-45 appears clearly as a key residue for the catalytic activity of LmNADK1. This central and buried aspartate seems to have a structural role, but at the same time, it may also participate in the catalysis by attracting the hydrogen of the 2'-hydroxyl group. This role may be essential for the specificity of the catalyzed phosphorylation. Indeed, the neighbouring 3' hydroxyl is solvent exposed with no direct contact with the protein and it remains unreactive. In most known kinases, including PFKs, conserved and catalytic acidic residues activate Mg 2+ -ATP directly, priming it for the phosphate transfer. In contrast, our results show that in NADKs, the corresponding aspartate (Asp-45 in LmNADK1) can not fulfill this role. The absence of other conserved acidic residue in the vicinity of the substrate, suggested that a distinct reaction mechanism was involved. This raised the question of the presence of an activator of the phosphodonor, and of a chelation site for the required dication. Ligand recognition and reaction specificity Our results so far showed that the specific position of Asp-45 led to its interaction with the 2 - hydroxyl of NAD, rather than with the di-phosphate link. The absence of strong recognition between the diphosphate groups of NAD and NADKs let us to suggest the rational design of new ligands. Various linkers might be used as a replacement for the phosphate groups to bridge the two nucleobases of NAD. We synthesized 5'-thio-adenosine (TA, Fig. 1C) and its S-acetylated form (TAA, Fig. 1C) as precursors of NAD mimics. The structures of LmNADK1 in complex with each of these adenosine analogues (Table 2) showed few variations compared to the unbound structure and to those in complex with NAD or NADP. Surprisingly, these compounds bind in a dimeric form in the NAD binding site of LmNADK1 and occupy both sites A and N. In both complexes, the adenosine scaffold which occupied the site A is oriented like the adenosine moiety of NAD. The second adenosine perfectly mimics the nicotinamide and its ribose, and showed very similar interactions with the protein despite the change of ring nature (Fig. S5). Compared to NAD, only slight differences are observed in the relative orientation of the nucleobases and a small shift in the position of the water molecule bridging them. The crystal complexes showed that acetyl groups of TAAs point toward each other (distance S5'-S5': ~ 5 Å), while in the deprotected form (TA) a disulphide bridge (distance S5'-S5': 2.0 Å) covalently linked the compound into a dimer named di-(5 -thioadenosine) (DTA, Fig. 1C and Fig. S5). This latter might have formed in solution in absence of reductant. In conclusion, the dimerization of 5 -thioadenosine appeared to have mimicked a complete natural di-nucleotide substrate. Enzymology experiments confirmed the competitive inhibition by these two synthetic compounds with Ki values of 3.6 mm and 0.02 mm for TAA and DTA, respectively. These affinities are low, but similar to those observed for the natural ligands NAD (Km ~ 1 mm) and NADP (Ki ~ 0.04 mm). Intriguingly, the two novel inhibitors possessed a 2'-hydroxyl readily accessible for phosphorylation by LmNADK1. Furthermore, the ribose moieties occupy strictly identical positions in the NAD and DTA bound crystal structures (see Fig. 4A). However, no phosphotransfer in presence of ATP could be observed (data not shown). This prompted us to synthesize a closer mimic of both DTA and NAD: di-adenosine-diphosphate (Ap2A, Fig. 1B, Fig. 4B and Fig. S6) to investigate the role 66 6
7 of the phosphate groups in catalysis and the potential impact of the nucleobase substitution. The structure of Ap2A bound to LmNADK1 was also solved to a resolution of 2.2 Å (Table 2). It showed that the new ligand adopted a conformation highly similar to that of DTA, and perfectly mimicked NAD. Moreover, in the structure solved after crystal soaking with both Ap2A and Mg 2+ -ATP, a clear density corresponding to Ap2A bearing a 2' phosphate group was observed (2 -phospho-ap2a, Fig. 1B and Fig. S6), inferring that Ap2A could be used as a substrate by LmNADK1, in contrast to TAA and the closely related DTA. These results revealed that the presence of a correctly oriented 2'-hydroxyl in DTA is not sufficient for the catalysis to occur. Moreover, the presence of the diphosphate groups of the substrate seems to be important. Even though we failed to identify electron density for bound metal ions (Mg 2+ or Mn 2+ ), this suggested that the negatively charged phosphates might be involved in the dication chelation. To evaluate the putative substrate-assisted catalysis, a new directed mutation was performed. The histidine H223 is the only residue whose sidechain transiently interacts with the phosphate groups (see above). This histidine was mutated to glutamate in order to introduce a negatively charged residue. The glutamate was predicted to be able to chelate a dication. This mutant was twice less active than the wild-type (0.47 versus 1.1 U/mg) on the biologically-relevant substrate NAD. On the contrary, its activity toward DTA is increased twofold (from 0.13 to 0.24 U/mg). The crystal structure of the mutant H223E explained the low activity, because the glutamate sidechain folded back in a buried position. E223 forms hydrogen bonds with its own backbone nitrogen and the sidechain of the histidine 53 (data not shown) preventing a higher impact on catalysis. However, the observed change in the ratio of enzyme activity on NAD versus DTA for the wild-type and the mutant enzymes confirmed the proposed substrate-assisted mechanism. This mechanism would explain the low dication specificity and at the same time provide the molecular basis for specific productive recognition of dinucleotide-diphosphate substrates. DISCUSSION NADKs from Gram(+) bacteria represent a promising novel therapeutic target for a whole spectrum of human pathogens including bacilli and staphylococci. Here, we have determined the crystal structures of NADK from L. monocytogenes in the free state, and bound to NAD, NADP, and two nonnatural ligands. In contrast to the previously reported complexes of NADK-NADP and NADK-NADs from M. tuberculosis and A. fulgidus (15,16), all our structures were determined at near 2 Å resolution, allowing a more detailed analysis of the changes accompagnying the catalytic reaction. This is especially true for the aspartate of the conserved motif GGDGT. Our analysis revealed that the aspartate of the conserved GGDGT motif has a different catalytic role in PFKs and NADKs. In PFKs this conserved residue is involved in ATP activation, while in NADK it is involved in NAD activation. Indeed, based on these structures, in combination with site-directed mutagenesis and enzymology experiments, we propose that in NADKs, the conserved aspartate is involved in proton abstraction from the 2'-hydroxyl of NAD to activate the phospho-acceptor. This explains why NADK phosphorylates only the 2 -hydroxyl of adenosine, and not the functionally equivalent 3 - hydroxyl. In addition, our results infer that the diphosphate group of the substrate plays the role of a dication chelator, revealing a mechanism involving substrate-assisted catalysis (Fig. 5). Together, these two aspects of this mechanism would cooperatively ensure the specificity of the catalyzed reaction. Thus our results yield important insights into the specificity and mechanism of the catalytic action of NADKs. Nevertheless, this mechanism might be extended to the families of diacylglycerol/ sphingonsine kinases (30, 31). The first structure (PDB # 2JGR) of a member (YegS) of this protein family was recently solved (32). Structural comparison highlighted that the aspartate of the well conserved GGDGT motif adopts the same position (supplementary data, Fig. S7). In this structure, a pyrophosphate group interacts with residues from the GGDGT motif (mainly the last glycine and the threonine). It potentially mimics the phosphate donor. However, in both NADKs and the related lipid kinases, the precise orientation of the phosphate donor and/or the position of the catalytic dication remain presently unresolved. 77 7
8 As a first step toward designing new inhibitors for LmNADK1, we have described herein the structural and enzymatic characterization of two novel compounds mimicking NAD, namely Ap2A and DTA, the latter representing a novel synthetic inhibitor for NADK. Building tighter links between the two nucleobases would lead to more efficient inhibitors. 88 8
9 REFERENCES 1. Pucci, M. J. (2006) Biochem Pharmacol 71, Gerdes, S. Y., Scholle, M. D., D'Souza, M., Bernal, A., Baev, M. V., Farrell, M., Kurnasov, O. V., Daugherty, M. D., Mseeh, F., Polanuyer, B. M., et al. (2002) J Bacteriol 184, Kobayashi, K., Ehrlich, S. D., Albertini, A., Amati, G., Andersen, K. K., Arnaud, M., Asai, K., Ashikaga, S., Aymerich, S., Bessieres, P., Boland, F., et al. (2003) Proc Natl Acad Sci U S A 100, Suzuki, N., Okai, N., Nonaka, H., Tsuge, Y., Inui, M., and Yukawa, H. (2006) Appl Environ Microbiol 72, Sassetti, C. M., Boyd, D. H., and Rubin, E. J. (2003) Mol Microbiol 48, Thanassi, J. A., Hartman-Neumann, S. L., Dougherty, T. J., Dougherty, B. A., and Pucci, M. J. (2002) Nucleic Acids Res 30, Zalacain, M., Biswas, S., Ingraham, K. A., Ambrad, J., Bryant, A., Chalker, A. F., Iordanescu, S., Fan, J., Fan, F., Lunsford, R. D., et al. (2003) J Mol Microbiol Biotechnol 6, Grose, J. H., Joss, L., Velick, S. F., and Roth, J. R. (2006) Proc Natl Acad Sci U S A 103, Kawai, S., Mori, S., Mukai, T., Suzuki, S., Yamada, T., Hashimoto, W., and Murata, K. (2000) Biochem Biophys Res Commun 276, Lerner, F., Niere, M., Ludwig, A., and Ziegler, M. (2001) Biochem Biophys Res Commun 288, Schomburg, I., Chang, A., Ebeling, C., Gremse, M., Heldt, C., Huhn, G., and Schomburg, D. (2004) Nucleic Acids Res 32, D Garavaglia, S., Galizzi, A., and Rizzi, M. (2003) J Bacteriol 185, Kawai, S., Mori, S., Mukai, T., Hashimoto, W., and Murata, K. (2001) Eur J Biochem 268, Garavaglia, S., Raffaelli, N., Finaurini, L., Magni, G., and Rizzi, M. (2004) J Biol Chem 279, Liu, J., Lou, Y., Yokota, H., Adams, P. D., Kim, R., and Kim, S. H. (2005) J Mol Biol 354, Mori, S., Yamasaki, M., Maruyama, Y., Momma, K., Kawai, S., Hashimoto, W., Mikami, B., and Murata, K. (2005) Biochem Biophys Res Commun 327, Pignot, M., Pljevaljcic, G., and Weinhold. (2000) E. European Journal of Organic Chemistry, (1994) Acta Crystallogr D Biol Crystallogr 50, Weeks, C. M., and Miller, R. (1999) Acta Crystallogr D Biol Crystallogr 55, Bricogne, G., Vonrhein, C., Flensburg, C., Schiltz, M., and Paciorek, W. (2003) Acta Crystallogr D Biol Crystallogr 59, Cowtan, K. (1994) Newsletter on Protein Crystallography 31, McCoy, A. J., Storoni, L. C., and Read, R. J. (2004) Acta Crystallogr D Biol Crystallogr 60, Jones, T. A., Zou, J. Y., Cowan, S. W., and Kjeldgaard, M. (1991) Acta Crystallogr A 47, Emsley, P., and Cowtan, K. (2004) Acta Crystallogr D Biol Crystallogr 60, Winn, M. D., Isupov, M. N., and Murshudov, G. N. (2001) Acta Crystallogr D Biol Crystallogr 57, Moffatt, J. G. A. (1964) Canadian Journal of Chemistry 42, Raffaeli, N., Finaurini L, Mazzola F, Pucci L, Sorci L, Amici A, and Magni G (2004) Biochemistry. 43, Shirakihara, Y., and Evans, P. R. (1988) J Mol Biol 204, Labesse, G., Douguet, D., Assairi, L., and Gilles, A. M. (2002) Trends Biochem Sci 27, Taha, T. A., Hannun, Y. A., and Obeid, L. M. (2006) J Biochem Mol Biol 39, Topham, M. K. (2006) J Cell Biochem 97, Bakali, M.A., Dolores Herman, M., Johnson, K.A., Kelly, A., Wieslander, A., Hallberg, B.M. and Nordlund, P. (2007) J Biol Chem 282, Catherinot, V., and Labesse, G. (2004) Bioinformatics 20,
10 FOOTNOTES Acknowledgments: We thanks O. Barzu and A-M Gilles (Institut Pasteur, Paris), C. Royer, C. Roumestand, F. Hoh and C. Frauer (Centre de Biochimie Structurale, Montpellier) for helpful discussion and technical help. * Present address : Université Montpellier 2, Montpellier, F France; INSERM, U710, Montpellier, F France; EPHE, Paris, F France. 1 The abbreviations used are: Ap2A, diadenosine diphosphate; TA, 5 -thio-adenosine; TAA, 5'-thioacetyladenosine; DTA, di-(5 -thio-adenosine); LmNADK1, NADK1 from the Listeria monocytogenes. 2 Data deposition footnote: The refined models and structure factors have been deposited in the Research Collaboratory for Structural Biology ( under the following accession numbers: PDB # 2I1W (Native structure), PDB # 2I29 (LmNADK1-NAD complex), PDB # 2I2A (LmNADK1-NADP complex), PDB # 2I2B (LmNADK1-TAA complex), PDB # 2I2C (LmNADK1-DTA complex), PDB # 2I2D (LmNADK1- Ap2A complex), PDB # 2I2E (LmNADK1-2'p-Ap2A complex), PDB # 2I2F (D45N mutant of LmNADK1 in complex with NAD), PDB # 2Q5F (H223E mutant of LmNADK1 in complex with DTA). FIGURE LEGENDS Fig. 1: Schematic reaction and chemical structures of nucleotides describe in this study. A. Schematic reaction of the NADKs. B. Di-nucleotides linked by a di-phosphate group and representation of adenine and nicotinamide moiety. C. Di-nucleotide linked by a disulfide bridge (DTA) and its protected form (TAA). Fig. 2: Structural superposition of the tetrameric structure of LmNADK1 in absence (grey) and presence of NAD (black) using the C-terminal as an anchor. A monomer is zoomed out to highlight the position of the NAD at the domain hinge. Fig. 3: Stereo view of the superposed N-terminal domains of NADKs and PFKs. The N-termini of LmNADK1 and EcPFK were superposed using the program ViTO (33). The picture was generated by the program PYMOL ( Grey and black cartoons and wireframes represent EcPFK/ADP/FBP/Mg 2+ and LmNADK1/NAD, respectively. Fig. 4: A: Stereo view of the superposition of LmNADK1-NAD (black) and LmNADK1-DTA (grey) active site. B Stereo view of the superposition of LmNADK1-NADP (black) and LmNADK1-2 p-ap2a (grey) active site. Fig. 5: Schematic flow of the reaction of NAD phosphorylation. The substrate-assisted catalysis mechanism proposed for NADKs (see discussion in the text) implies chelation of the catalytic dication Mg 2+ by both the phosphate group of the donor and the disphosphate group of the subtrate. Protein domains are drawn as filled shapes with thick lines. The catalytic aspartate D45 is drawn as a thick Y symbol. The ligands observed in the various crystal structures are drawn as plain shapes while shaded forms correspond to the phosphate donor and the dication placed at their putative positions. Fig. S1: Sequence alignment of LmNADK1 with related NADKs. Structural alignment of various NADKs including the human NADK (its central catalytic domain). Enzymes names are from SWISSPROT but for the known crystal structures for which we use the PDB code. The motif of two binding sites, A and N, are highlighted by green triangles (phosphodonor) and purple stars (NAD), respectively. A loop specific of Grampositive bacteria is underline with blue circles. The figure was drawn using ESPRIT ( Fig. S2: Binding mode of NAD: Upper: Views of the 2Fo Fc electron density map in the active site with the NAD molecule omitted in the Fourier synthesis. Map is contoured at 1.5 σ. NAD are shown as a ball-and-stick model. Bottom: Schematic representation of the network of interactions between NAD and LmNADK1. Hydrogen bonds are shown as dashes in black labelled with interatomic distance. Important ring stacking are indicated between nicotinamide and tyrosine
11 Fig. S3: Binding mode of NADP: Views of the 2Fo Fc electron density map for NADP molecule drawn as described in Fig. S2. Fig. S4: NAD binding to D45N mutant. Structural comparison of the active site of the wild-type (green backbone) and D45N mutant (blue backbone) of LmNADK1 in complex with NAD. Fig. S5: Binding mode of DTA: Views of the 2Fo Fc electron density map for the DTA molecule drawn as described in Fig. S2. Fig. S6: Binding mode of 2'-phospho-Ap2A: Views of the 2Fo Fc electron density map for the 2'-phospho- Ap2A molecule drawn as described in Fig. S2. Fig. S7: Structural comparison of LmNADK1-NAD and YegS-pyrophosphate. Blue/Green cartoons represent LmNADK1 and YegS, respectively. Wheat cartoons represent the well conserved GGDGT motif and NAD molecule is represented in yellow. 11
12 TABLES TABLE 1: Data collection, phasing, and refinement statistics for apo LmNADK1. Data collection Native SeMet peak SeMet inflection SeMet remote Space group P2 1 P2 1 Cell dimension a, b, c (Å) 66.6, 119.0, , 119.3, 67.5 β ( ) No. molecules in a.u 4 4 Wavelength (Å) Resolution (Å) ( ) ( ) R merge (%) (43.1) 5.0 (8.4) 4.6 (9.2) 4.5 (7.2) I/σI (2.9) 18.9 (8.0) 18 (6.0) 20.1 (7.9) Completeness (%) (99.3) 96.6 (77.7) 94.1 (68.4) 97.7 (85.2) Redundancy (3.6) 3.4 (1.6) 2.9 (1.5) 3.4 (1.6) B-wilson Refinement Resolution (Å) No. Reflections 42,472 R work /R free (%) /26.6 No. Atoms Protein 8,194 Iodide 18 Water 362 B-factors (Å 2 ) Protein 38.9 Iodide 38.5 Water 39.8 R.m.s deviations 5 Bond lengths (Å) Bond angles ( ) Values in parentheses refer to the outermost resolution shell. 2 Rmerge = ΣhklΣi_ Ihkl,i Iaverage,hkl / ΣhklΣi_ Ihkl,i x Rwork = Σhkl Fobs Fcalc /Σhkl Fobs x Rfree.is calculated in the same way on a subset of reflections that are not used in the refinement (5 %). 5 Deviation from ideal values. 12
13 TABLE 2: Data collection and refinement statistics for LmNADK1-complexes. Data collection 1 Values in parentheses refer to the outermost resolution shell. Wild type D45N mutant TAA DTA NAD NADP Ap2A 2 -pappa NAD Space group I222 I222 I222 I222 I222 I222 I222 Cell dimension a, b, c (Å) 63.2, 74.6, , 75.3, , 73.5, , 77.5, , 75.9, , 75.7, , 76.4, No. molecules in a.u Wavelength (Å) Resolution (Å) ( ) ( ) ( ) ( ) ( ) ( ) ( ) Rmerge (%) (48.8) 6.8 (44.2) 7.1 (56.8) 7.7 (44.8) 5.4 (41.1) 4.8 (46.0) 7.1 (39.9) I/σI (2.9) 17.3 (1.9) 17.2 (2.0) 18.1 (4.2) 22.9 (3.5) 18.6 (1.9) 20.9 (4.1) Completeness (%) (100.0) 98.9 (93.5) 98.7 (91.3) 96.4 (98.0) 96.8 (81.2) 99.1 (94.5) 99.0 (94.0) Redundancy (5.7) 5.2 (2.9) 6.0 (4.4) 5.9 (5.8) 4.9 (4.0) 3.2 (2.1) 6.5 (5.2) B-wilson Refinement Resolution (Å) No. Reflections 15,878 22,941 15,633 15,714 13,383 13,914 21,902 Rwork/Rfree (%) / / / / / / /24.3 No, Protein Atoms 2,122 2,325 2,145 2,226 2,107 2,148 2,195 No, Waters molecules Ligands type TAA x2 DTA NAD NADP Ap2A 2 -pappa NAD B-factors (Å 2 ) Protein Ligands Water R.m.s deviations 5 Bond lengths (Å) Bond angles ( ) Rmerge = ΣhklΣi_ Ihkl,i Iaverage,hkl / ΣhklΣi_ Ihkl,i x Rwork = Σhkl Fobs Fcalc /Σhkl Fobs x Rfree.is calculated in the same way on a subset of reflections that are not used in the refinement (5 %). 5 Deviation from ideal values. 13
14 TABLE 3: Kinetic parameters of LmNADK1 and its variants D45N. Substrate Protein ATP (mm) V max b K m (mm) S 0.5 (mm) n H k cat (S -1 ) c k cat /K m (S -1 /mm) k cat / S 0.5 (S -1 /mm) ATP WT a ± ± ± D45N a ± ± ± NAD WT d ± ± ± ± ± ± ± ± D45N d ± ± ± ± ± ± ± ± The kinetic values are presented as means ± standard errors for three independent determinations a data for 10 various ATP concentrations at 3 various NAD concentrations for 3 enzymatic preparations (obtained by fitting the data to the Michaelis - Menten equation v=v m [ATP]/Km+[ATP]) b µmol NADP/mg protein /min c assuming that the molecular mass of the tetramer (with His tag ) is124kd d Kinetic parameters were obtained by fitting data to the Hill equation (v=v m [NAD] n /S n 0.5 +[NAD] n ) and determined for a fixed ATP concentration of 4 or 0.5 mm 14
15 Figure 1 15

16 Figure 2 16

17 Figure 3 17

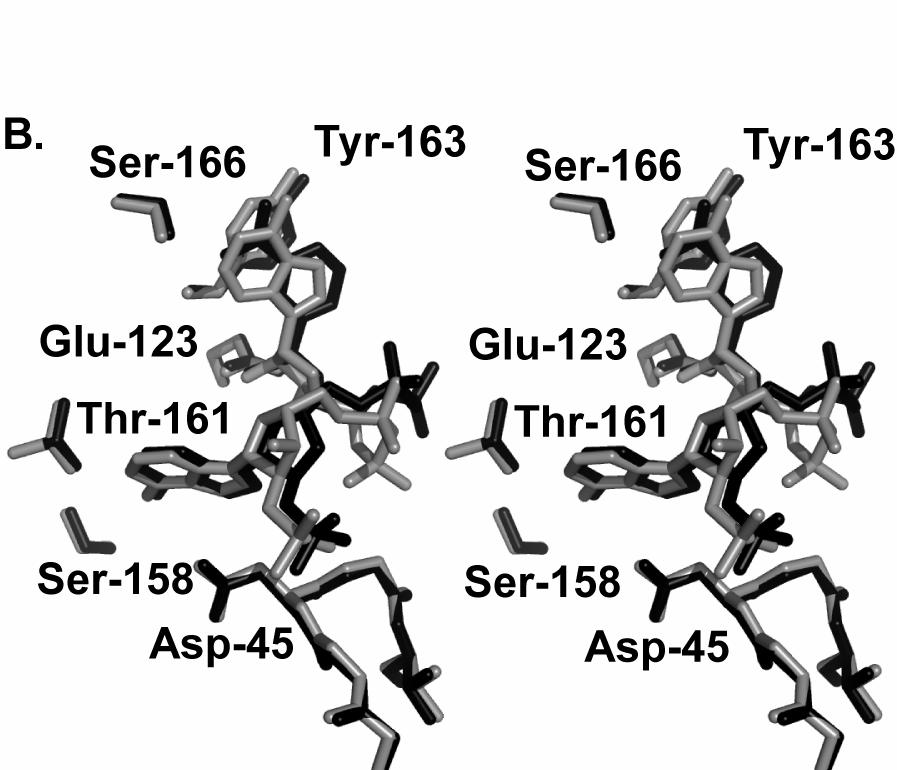
18 Figure 4 18

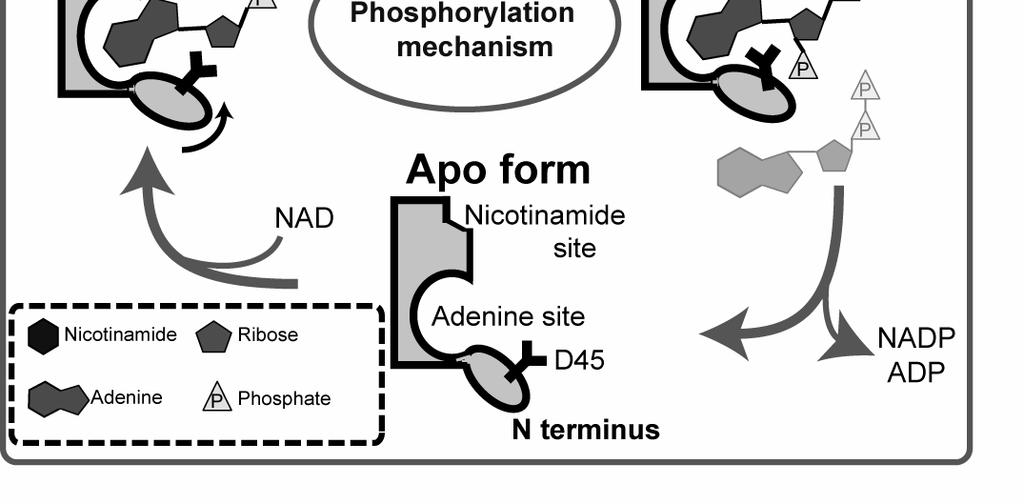
19 Figure 5 19
Supplementary Note 1. Enzymatic properties of the purified Syn BVR
 Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and
 Materials and Methods Immunoprecipitation and immunoblot analysis Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and immunoblot analyses with corresponding
Materials and Methods Immunoprecipitation and immunoblot analysis Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and immunoblot analyses with corresponding
Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid
 Amino acid") Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
6-Foot Mini Toober Activity
 Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction
 X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the
 Supplementary information, Data S1 Methods Clones and protein preparation Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the Saccharomyces cerevisiae genomic DNA by polymerase chain
Supplementary information, Data S1 Methods Clones and protein preparation Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the Saccharomyces cerevisiae genomic DNA by polymerase chain
Supplementary Information for. Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction
 Supplementary Information for Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction Takamasa Teramoto, Yukari Fujikawa, Yoshirou Kawaguchi, Katsuhisa
Supplementary Information for Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction Takamasa Teramoto, Yukari Fujikawa, Yoshirou Kawaguchi, Katsuhisa
SUPPLEMENTARY INFORMATION
 Molecular basis of RNA-dependent RNA polymerase II activity Elisabeth Lehmann, Florian Brueckner, and Patrick Cramer Gene Center Munich and Center for integrated Protein Science CiPS M, Department of Chemistry
Molecular basis of RNA-dependent RNA polymerase II activity Elisabeth Lehmann, Florian Brueckner, and Patrick Cramer Gene Center Munich and Center for integrated Protein Science CiPS M, Department of Chemistry
Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein
 Cell Wall Recycling Protein") Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
The Skap-hom Dimerization and PH Domains Comprise
 Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature06147 SUPPLEMENTARY INFORMATION Figure S1 The genomic and domain structure of Dscam. The Dscam gene comprises 24 exons, encoding a signal peptide (SP), 10 IgSF domains, 6 fibronectin
doi: 10.1038/nature06147 SUPPLEMENTARY INFORMATION Figure S1 The genomic and domain structure of Dscam. The Dscam gene comprises 24 exons, encoding a signal peptide (SP), 10 IgSF domains, 6 fibronectin
Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport
 Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport Ivan Laponogov 1,2, Dennis A. Veselkov 1, Isabelle M-T. Crevel
Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport Ivan Laponogov 1,2, Dennis A. Veselkov 1, Isabelle M-T. Crevel
Nature Structural & Molecular Biology: doi: /nsmb.3428
 Supplementary Figure 1 Biochemical characterization of the monou and oligou activity switch of TUT4(7). (a) Mouse TUT4 and human TUT7 were assayed for monou and Lin28-dependent oligou addition activities
Supplementary Figure 1 Biochemical characterization of the monou and oligou activity switch of TUT4(7). (a) Mouse TUT4 and human TUT7 were assayed for monou and Lin28-dependent oligou addition activities
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10963 Supplementary Table 1 Data collection, phasing and refinement statistics. Crystal Native Derivative-1 (OsO 4 ) Derivative-2 (Orange-Pt) Data collection Space group C2 C2 C2 Cell
doi:10.1038/nature10963 Supplementary Table 1 Data collection, phasing and refinement statistics. Crystal Native Derivative-1 (OsO 4 ) Derivative-2 (Orange-Pt) Data collection Space group C2 C2 C2 Cell
STRUCTURE, DYNAMICS AND INTERACTIONS OF PROTEINS BY NMR SPECTROSCOPY
 STRUCTURE, DYNAMICS AND INTERACTIONS OF PROTEINS BY NMR SPECTROSCOPY Constantin T. Craescu INSERM & Institut Curie - Recherche Orsay, France A SHORT INTRODUCTION TO PROTEIN STRUCTURE Chemical composition
STRUCTURE, DYNAMICS AND INTERACTIONS OF PROTEINS BY NMR SPECTROSCOPY Constantin T. Craescu INSERM & Institut Curie - Recherche Orsay, France A SHORT INTRODUCTION TO PROTEIN STRUCTURE Chemical composition
Clamping down on pathogenic bacteria how to shut down a key DNA polymerase complex
 Clamping down on pathogenic bacteria how to shut down a key DNA polymerase complex Bacterial DNA-replication machinery Pathogenic bacteria that are resistant to the current armoury of antibiotics are an
Clamping down on pathogenic bacteria how to shut down a key DNA polymerase complex Bacterial DNA-replication machinery Pathogenic bacteria that are resistant to the current armoury of antibiotics are an
Analysis of the AcrB binding domain of the membrane fusion protein AcrA. Introduction
 Analysis of the AcrB binding domain of the membrane fusion protein AcrA Introduction Antibiotic resistance in bacteria is due in large part to multidrug efflux complexes like the TolC-AcrAB complex in
Analysis of the AcrB binding domain of the membrane fusion protein AcrA Introduction Antibiotic resistance in bacteria is due in large part to multidrug efflux complexes like the TolC-AcrAB complex in
SUPPLEMENTARY INFORMATION
 This supplementary information is an extension of the letter with the same title and includes further discussion on the comparison of our designed Fe B Mb (computer model and crystal structure) with the
This supplementary information is an extension of the letter with the same title and includes further discussion on the comparison of our designed Fe B Mb (computer model and crystal structure) with the
Hmwk # 8 : DNA-Binding Proteins : Part II
 The purpose of this exercise is : Hmwk # 8 : DNA-Binding Proteins : Part II 1). to examine the case of a tandem head-to-tail homodimer binding to DNA 2). to view a Zn finger motif 3). to consider the case
The purpose of this exercise is : Hmwk # 8 : DNA-Binding Proteins : Part II 1). to examine the case of a tandem head-to-tail homodimer binding to DNA 2). to view a Zn finger motif 3). to consider the case
STRUCTURAL BIOLOGY. α/β structures Closed barrels Open twisted sheets Horseshoe folds
 STRUCTURAL BIOLOGY α/β structures Closed barrels Open twisted sheets Horseshoe folds The α/β domains Most frequent domain structures are α/β domains: A central parallel or mixed β sheet Surrounded by α
STRUCTURAL BIOLOGY α/β structures Closed barrels Open twisted sheets Horseshoe folds The α/β domains Most frequent domain structures are α/β domains: A central parallel or mixed β sheet Surrounded by α
Supplementary Information. Structural basis for duplex RNA recognition and cleavage by A.
 Supplementary Information Structural asis for duplex RNA recognition and cleavage y A. fulgidus C3PO Eneida arizotto 1, Edward D Lowe 1 & James S Parker 1 1 Department of Biochemistry University of Oxford
Supplementary Information Structural asis for duplex RNA recognition and cleavage y A. fulgidus C3PO Eneida arizotto 1, Edward D Lowe 1 & James S Parker 1 1 Department of Biochemistry University of Oxford
Molecular design principles underlying β-strand swapping. in the adhesive dimerization of cadherins
 Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
From mechanism to medicne
 From mechanism to medicne a look at proteins and drug design Chem 342 δ δ δ+ M 2009 δ+ δ+ δ M Drug Design - an Iterative Approach @ DSU Structural Analysis of Receptor Structural Analysis of Ligand-Receptor
From mechanism to medicne a look at proteins and drug design Chem 342 δ δ δ+ M 2009 δ+ δ+ δ M Drug Design - an Iterative Approach @ DSU Structural Analysis of Receptor Structural Analysis of Ligand-Receptor
Supplementary Fig. S1. SAMHD1c has a more potent dntpase activity than. SAMHD1c. Purified recombinant SAMHD1c and SAMHD1c proteins (with
 Supplementary Fig. S1. SAMHD1c has a more potent dntpase activity than SAMHD1c. Purified recombinant SAMHD1c and SAMHD1c proteins (with concentration of 800nM) were incubated with 1mM dgtp for the indicated
Supplementary Fig. S1. SAMHD1c has a more potent dntpase activity than SAMHD1c. Purified recombinant SAMHD1c and SAMHD1c proteins (with concentration of 800nM) were incubated with 1mM dgtp for the indicated
Solutions to 7.02 Quiz II 10/27/05
 Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
Genes to Proteins. Nucleic Acid Structure
 Genes to Proteins Pratt & Cornely Chapter 3 Nucleobase Nucleoside Nucleotide Nucleic acid Chromatin Chromosome Nucleic Acid Structure 1 Base Structure Purines and pyrimidines Aromatic Tautomers Nucleosides
Genes to Proteins Pratt & Cornely Chapter 3 Nucleobase Nucleoside Nucleotide Nucleic acid Chromatin Chromosome Nucleic Acid Structure 1 Base Structure Purines and pyrimidines Aromatic Tautomers Nucleosides
Bioinformatics. ONE Introduction to Biology. Sami Khuri Department of Computer Science San José State University Biology/CS 123A Fall 2012
 Bioinformatics ONE Introduction to Biology Sami Khuri Department of Computer Science San José State University Biology/CS 123A Fall 2012 Biology Review DNA RNA Proteins Central Dogma Transcription Translation
Bioinformatics ONE Introduction to Biology Sami Khuri Department of Computer Science San José State University Biology/CS 123A Fall 2012 Biology Review DNA RNA Proteins Central Dogma Transcription Translation
Dina Al-Tamimi. Faisal Nimri. Ma amoun Ahram. 1 P a g e
 1 Dina Al-Tamimi Faisal Nimri Ma amoun Ahram 1 P a g e **Difference between Molecular Biology and Genetics: Molecular Biology: is a fancy term of biochemistry. It is the science that deals with DNA, RNA
1 Dina Al-Tamimi Faisal Nimri Ma amoun Ahram 1 P a g e **Difference between Molecular Biology and Genetics: Molecular Biology: is a fancy term of biochemistry. It is the science that deals with DNA, RNA
Crystal Structure of a Self-Splicing Group I Intron with Both Exons
 Supplementary Data for: Crystal Structure of a Self-Splicing Group I Intron with Both Exons Peter L. Adams, Mary R. Stahley, Anne B. Kosek, Jimin Wang and Scott A. Strobel The supplementary material includes
Supplementary Data for: Crystal Structure of a Self-Splicing Group I Intron with Both Exons Peter L. Adams, Mary R. Stahley, Anne B. Kosek, Jimin Wang and Scott A. Strobel The supplementary material includes
Zool 3200: Cell Biology Exam 3 3/6/15
 Name: Trask Zool 3200: Cell Biology Exam 3 3/6/15 Answer each of the following questions in the space provided; circle the correct answer or answers for each multiple choice question and circle either
Name: Trask Zool 3200: Cell Biology Exam 3 3/6/15 Answer each of the following questions in the space provided; circle the correct answer or answers for each multiple choice question and circle either
of the Triphosphate of ATP
 A Small Aptamer with Strong and Specific Recognition of the Triphosphate of Peter L. Sazani, Rosa Larralde and Jack W. Szostak Howard Hughes Medical Institute, and Department of Molecular Biology, Massachusetts
A Small Aptamer with Strong and Specific Recognition of the Triphosphate of Peter L. Sazani, Rosa Larralde and Jack W. Szostak Howard Hughes Medical Institute, and Department of Molecular Biology, Massachusetts
Algorithms in Bioinformatics ONE Transcription Translation
 Algorithms in Bioinformatics ONE Transcription Translation Sami Khuri Department of Computer Science San José State University sami.khuri@sjsu.edu Biology Review DNA RNA Proteins Central Dogma Transcription
Algorithms in Bioinformatics ONE Transcription Translation Sami Khuri Department of Computer Science San José State University sami.khuri@sjsu.edu Biology Review DNA RNA Proteins Central Dogma Transcription
Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis
 Supplemental material to accompany Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis Jacob Bauer, a Gabriela Ondrovičová, a Lucie Najmanová, b Vladimír Pevala,
Supplemental material to accompany Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis Jacob Bauer, a Gabriela Ondrovičová, a Lucie Najmanová, b Vladimír Pevala,
Supporting Online Material. Av1 and Av2 were isolated and purified under anaerobic conditions according to
 Supporting Online Material Materials and Methods Av1 and Av2 were isolated and purified under anaerobic conditions according to published protocols (S1). Crystals of nf-, pcp- and adp-av2:av1 complexes
Supporting Online Material Materials and Methods Av1 and Av2 were isolated and purified under anaerobic conditions according to published protocols (S1). Crystals of nf-, pcp- and adp-av2:av1 complexes
HEK293T. Fig. 1 in the
 Supplementary Information Supplementary Figure 1 Zinc uptake assay of hzip4 and hzip4-δecd transiently expressed in HEK293T cells. The results of one representative e experiment are shown in Fig. 1 in
Supplementary Information Supplementary Figure 1 Zinc uptake assay of hzip4 and hzip4-δecd transiently expressed in HEK293T cells. The results of one representative e experiment are shown in Fig. 1 in
Speaker: Yu-Chen Ku Professor: Ching-Tsan Huang, Ph. D. Source: Biochemical Journal (2007) 402, Date: December 4th, 2007
 402, Date: December 4th, 2007") Directed evolution and structural analysis of N-carbamoyl-D-amino acid amidohydrolase provide insights into recombinant protein solubility in Escherichia coli Speaker: Yu-Chen Ku Professor: Ching-Tsan
Directed evolution and structural analysis of N-carbamoyl-D-amino acid amidohydrolase provide insights into recombinant protein solubility in Escherichia coli Speaker: Yu-Chen Ku Professor: Ching-Tsan
RNA Expression of the information in a gene generally involves production of an RNA molecule transcribed from a DNA template. RNA differs from DNA
 RNA Expression of the information in a gene generally involves production of an RNA molecule transcribed from a DNA template. RNA differs from DNA that it has a hydroxyl group at the 2 position of the
RNA Expression of the information in a gene generally involves production of an RNA molecule transcribed from a DNA template. RNA differs from DNA that it has a hydroxyl group at the 2 position of the
Chapter 14 Regulation of Transcription
 Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
Supplementary Information For. A genetically encoded tool for manipulation of NADP + /NADPH in living cells
 Supplementary Information For A genetically encoded tool for manipulation of NADP + /NADPH in living cells Valentin Cracan 1,2,3, Denis V. Titov 1,2,3, Hongying Shen 1,2,3, Zenon Grabarek 1* and Vamsi
Supplementary Information For A genetically encoded tool for manipulation of NADP + /NADPH in living cells Valentin Cracan 1,2,3, Denis V. Titov 1,2,3, Hongying Shen 1,2,3, Zenon Grabarek 1* and Vamsi
7 Synthesizing the ykkcd Mutant Toxin Sensor RNA in vitro
 7 Synthesizing the ykkcd Mutant Toxin Sensor RNA in vitro 7.1 Learning Objective In the quest toward understanding how the ykkcd toxin sensor recognizes the antibiotic tetracycline you thus far designed
7 Synthesizing the ykkcd Mutant Toxin Sensor RNA in vitro 7.1 Learning Objective In the quest toward understanding how the ykkcd toxin sensor recognizes the antibiotic tetracycline you thus far designed
 Problem: The GC base pairs are more stable than AT base pairs. Why? 5. Triple-stranded DNA was first observed in 1957. Scientists later discovered that the formation of triplestranded DNA involves a type
Problem: The GC base pairs are more stable than AT base pairs. Why? 5. Triple-stranded DNA was first observed in 1957. Scientists later discovered that the formation of triplestranded DNA involves a type
MBMB451A Section1 Fall 2008 KEY These questions may have more than one correct answer
 MBMB451A Section1 Fall 2008 KEY These questions may have more than one correct answer 1. In a double stranded molecule of DNA, the ratio of purines : pyrimidines is (a) variable (b) determined by the base
MBMB451A Section1 Fall 2008 KEY These questions may have more than one correct answer 1. In a double stranded molecule of DNA, the ratio of purines : pyrimidines is (a) variable (b) determined by the base
AP Biology Book Notes Chapter 3 v Nucleic acids Ø Polymers specialized for the storage transmission and use of genetic information Ø Two types DNA
 AP Biology Book Notes Chapter 3 v Nucleic acids Ø Polymers specialized for the storage transmission and use of genetic information Ø Two types DNA Encodes hereditary information Used to specify the amino
AP Biology Book Notes Chapter 3 v Nucleic acids Ø Polymers specialized for the storage transmission and use of genetic information Ø Two types DNA Encodes hereditary information Used to specify the amino
Drug DNA interaction. Modeling DNA ligand interaction of intercalating ligands
 Drug DNA interaction DNA as carrier of genetic information is a major target for drug interaction because of the ability to interfere with transcription (gene expression and protein synthesis) and DNA
Drug DNA interaction DNA as carrier of genetic information is a major target for drug interaction because of the ability to interfere with transcription (gene expression and protein synthesis) and DNA
Protocol S1: Supporting Information
 Protocol S1: Supporting Information Basis for the specificity of the kinase domain of Abl for peptide substrates The crystal structures reported in this work were obtained using two different ATP analog-peptide
Protocol S1: Supporting Information Basis for the specificity of the kinase domain of Abl for peptide substrates The crystal structures reported in this work were obtained using two different ATP analog-peptide
Structural Bioinformatics (C3210) DNA and RNA Structure
 DNA and RNA Structure") Structural Bioinformatics (C3210) DNA and RNA Structure Importance of DNA/RNA 3D Structure Nucleic acids are essential materials found in all living organisms. Their main function is to maintain and transmit
Structural Bioinformatics (C3210) DNA and RNA Structure Importance of DNA/RNA 3D Structure Nucleic acids are essential materials found in all living organisms. Their main function is to maintain and transmit
BC 367, Exam 2 November 13, Part I. Multiple Choice (3 pts each)- Please circle the single best answer.
- Please circle the single best answer.") Name BC 367, Exam 2 November 13, 2008 Part I. Multiple Choice (3 pts each)- Please circle the single best answer. 1. The enzyme pyruvate dehydrogenase catalyzes the following reaction. What kind of enzyme
Name BC 367, Exam 2 November 13, 2008 Part I. Multiple Choice (3 pts each)- Please circle the single best answer. 1. The enzyme pyruvate dehydrogenase catalyzes the following reaction. What kind of enzyme
The YTH domain (residues ) of human YTHDF2 (NP_ ) was subcloned
 of human YTHDF2 (NP_ ) was subcloned") Supplementary information, Data S1 Materials and Methods Protein Expression, Purification and Crystallization The YTH domain (residues 383-553) of human YTHDF2 (NP_057342.2) was subcloned into a modified
Supplementary information, Data S1 Materials and Methods Protein Expression, Purification and Crystallization The YTH domain (residues 383-553) of human YTHDF2 (NP_057342.2) was subcloned into a modified
Basic concepts of molecular biology
 Basic concepts of molecular biology Gabriella Trucco Email: gabriella.trucco@unimi.it Life The main actors in the chemistry of life are molecules called proteins nucleic acids Proteins: many different
Basic concepts of molecular biology Gabriella Trucco Email: gabriella.trucco@unimi.it Life The main actors in the chemistry of life are molecules called proteins nucleic acids Proteins: many different
has only one nucleotide, U20, between G19 and A21, while trna Glu CUC has two
 SPPLEMENTRY INFORMTION doi:1.138/nature9411 Supplementary Discussion The structural characteristics of trn Gln G in comparison to trn Glu 16 in trn Gln G is directed towards the G19 56 pair, or the outer
SPPLEMENTRY INFORMTION doi:1.138/nature9411 Supplementary Discussion The structural characteristics of trn Gln G in comparison to trn Glu 16 in trn Gln G is directed towards the G19 56 pair, or the outer
Supplementary Figure 1
 Supplementary Figure 1 2 Supplementary Figure 1: Sequence alignment of HsHSD17B8 and HsCBR4 of with KAR orthologs. The secondary structure elements as calculated by DSSP and residue numbers are displayed
Supplementary Figure 1 2 Supplementary Figure 1: Sequence alignment of HsHSD17B8 and HsCBR4 of with KAR orthologs. The secondary structure elements as calculated by DSSP and residue numbers are displayed
Peptide deformylase from superbacteria
 Peptide deformylase from superbacteria Antibiotics Most antibiotics were originally isolated from soil-derived actinomycetes between 1940s and 1960s (Golden era of antibiotic discovery) Natural product
Peptide deformylase from superbacteria Antibiotics Most antibiotics were originally isolated from soil-derived actinomycetes between 1940s and 1960s (Golden era of antibiotic discovery) Natural product
human Cdc45 Figure 1c. (c)
") 1 Details of the refined crystallographic model of human Cdc45 and comparison of its active-site region with that of bacterial RecJ. (a) Stereo view of a representative example of the final 2F o -F c electron
1 Details of the refined crystallographic model of human Cdc45 and comparison of its active-site region with that of bacterial RecJ. (a) Stereo view of a representative example of the final 2F o -F c electron
Supporting Information
 Supporting Information Slep et al. 10.1073/pnas.0801569105 SI Materials and Methods Mouse RGS16, residues 53 180, was subcloned into a modified pgex-2t vector (GE Healthcare) to yield a thrombin-cleavable
Supporting Information Slep et al. 10.1073/pnas.0801569105 SI Materials and Methods Mouse RGS16, residues 53 180, was subcloned into a modified pgex-2t vector (GE Healthcare) to yield a thrombin-cleavable
Nanobiotechnology. Place: IOP 1 st Meeting Room Time: 9:30-12:00. Reference: Review Papers. Grade: 50% midterm, 50% final.
 Nanobiotechnology Place: IOP 1 st Meeting Room Time: 9:30-12:00 Reference: Review Papers Grade: 50% midterm, 50% final Midterm: 5/15 History Atom Earth, Air, Water Fire SEM: 20-40 nm Silver 66.2% Gold
Nanobiotechnology Place: IOP 1 st Meeting Room Time: 9:30-12:00 Reference: Review Papers Grade: 50% midterm, 50% final Midterm: 5/15 History Atom Earth, Air, Water Fire SEM: 20-40 nm Silver 66.2% Gold
SUPPLEMENTAL MATERIAL BIOCHEMICAL AND STRUCTURAL STUDIES ON THE M. TUBERCULOSIS O 6 -METHYLGUANINE METHYLTRANSFERASE AND MUTATED VARIANTS
 SUPPLEMENTAL MATERIAL BIOCHEMICAL AND STRUCTURAL STUDIES ON THE M. TUBERCULOSIS O 6 -METHYLGUANINE METHYLTRANSFERASE AND MUTATED VARIANTS Riccardo Miggiano 1, Valentina Casazza 1, Silvia Garavaglia 1,
SUPPLEMENTAL MATERIAL BIOCHEMICAL AND STRUCTURAL STUDIES ON THE M. TUBERCULOSIS O 6 -METHYLGUANINE METHYLTRANSFERASE AND MUTATED VARIANTS Riccardo Miggiano 1, Valentina Casazza 1, Silvia Garavaglia 1,
Biochemistry Prof. S. Dasgupta Department of Chemistry. Indian Institute of Technology Kharagpur. Lecture - 16 Nucleic Acids - I
 Biochemistry Prof. S. Dasgupta Department of Chemistry. Indian Institute of Technology Kharagpur Lecture - 16 Nucleic Acids - I We start our discussion on Nucleic Acids and their components. Before we
Biochemistry Prof. S. Dasgupta Department of Chemistry. Indian Institute of Technology Kharagpur Lecture - 16 Nucleic Acids - I We start our discussion on Nucleic Acids and their components. Before we
Figure S2, related to Figure 1. Stereo images of the CarD/RNAP complex and. electrostatic potential surface representation of the CarD/RNAP interface
 Structure, Volume 21 Supplemental Information Structure of the Mtb CarD/RNAP -Lobes Complex Reveals the Molecular Basis of Interaction and Presents a Distinct DNA-Binding Domain for Mtb CarD Gulcin Gulten
Structure, Volume 21 Supplemental Information Structure of the Mtb CarD/RNAP -Lobes Complex Reveals the Molecular Basis of Interaction and Presents a Distinct DNA-Binding Domain for Mtb CarD Gulcin Gulten
Name. Student ID. Midterm 2, Biology 2020, Kropf 2004
 Midterm 2, Biology 2020, Kropf 2004 1 1. RNA vs DNA (5 pts) The table below compares DNA and RNA. Fill in the open boxes, being complete and specific Compare: DNA RNA Pyrimidines C,T C,U Purines 3-D structure
Midterm 2, Biology 2020, Kropf 2004 1 1. RNA vs DNA (5 pts) The table below compares DNA and RNA. Fill in the open boxes, being complete and specific Compare: DNA RNA Pyrimidines C,T C,U Purines 3-D structure
Nucleic Acids, Proteins, and Enzymes
 3 Nucleic Acids, Proteins, and Enzymes Chapter 3 Nucleic Acids, Proteins, and Enzymes Key Concepts 3.1 Nucleic Acids Are Informational Macromolecules 3.2 Proteins Are Polymers with Important Structural
3 Nucleic Acids, Proteins, and Enzymes Chapter 3 Nucleic Acids, Proteins, and Enzymes Key Concepts 3.1 Nucleic Acids Are Informational Macromolecules 3.2 Proteins Are Polymers with Important Structural
Supplementary Fig. 1. Initial electron density maps for the NOX-D20:mC5a complex obtained after SAD-phasing. (a) Initial experimental electron
 Initial experimental electron") Supplementary Fig. 1. Initial electron density maps for the NOX-D20:mC5a complex obtained after SAD-phasing. (a) Initial experimental electron density map obtained after SAD-phasing and density modification
Supplementary Fig. 1. Initial electron density maps for the NOX-D20:mC5a complex obtained after SAD-phasing. (a) Initial experimental electron density map obtained after SAD-phasing and density modification
SUPPLEMENTARY INFORMATION. Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly
 SUPPLEMENTARY INFORMATION Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly Dustin J. E. Huard, Kathleen M. Kane and F. Akif Tezcan* Department of Chemistry and Biochemistry,
SUPPLEMENTARY INFORMATION Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly Dustin J. E. Huard, Kathleen M. Kane and F. Akif Tezcan* Department of Chemistry and Biochemistry,
Protein Structure/Function Relationships
 Protein Structure/Function Relationships W. M. Grogan, Ph.D. OBJECTIVES 1. Describe and cite examples of fibrous and globular proteins. 2. Describe typical tertiary structural motifs found in proteins.
Protein Structure/Function Relationships W. M. Grogan, Ph.D. OBJECTIVES 1. Describe and cite examples of fibrous and globular proteins. 2. Describe typical tertiary structural motifs found in proteins.
Supplemental Information. OprG Harnesses the Dynamics of its Extracellular. Loops to Transport Small Amino Acids across
 Structure, Volume 23 Supplemental Information OprG Harnesses the Dynamics of its Extracellular Loops to Transport Small Amino Acids across the Outer Membrane of Pseudomonas aeruginosa Iga Kucharska, Patrick
Structure, Volume 23 Supplemental Information OprG Harnesses the Dynamics of its Extracellular Loops to Transport Small Amino Acids across the Outer Membrane of Pseudomonas aeruginosa Iga Kucharska, Patrick
Chapter 8 DNA Recognition in Prokaryotes by Helix-Turn-Helix Motifs
 Chapter 8 DNA Recognition in Prokaryotes by Helix-Turn-Helix Motifs 1. Helix-turn-helix proteins 2. Zinc finger proteins 3. Leucine zipper proteins 4. Beta-scaffold factors 5. Others λ-repressor AND CRO
Chapter 8 DNA Recognition in Prokaryotes by Helix-Turn-Helix Motifs 1. Helix-turn-helix proteins 2. Zinc finger proteins 3. Leucine zipper proteins 4. Beta-scaffold factors 5. Others λ-repressor AND CRO
His-Spin Protein Miniprep
 INSTRUCTIONS His-Spin Protein Miniprep Catalog No. P2001 (10 purifications) and P2002 (50 purifications). Highlights Fast 5 minute protocol to purify His-tagged proteins from cell-free extracts Screen
INSTRUCTIONS His-Spin Protein Miniprep Catalog No. P2001 (10 purifications) and P2002 (50 purifications). Highlights Fast 5 minute protocol to purify His-tagged proteins from cell-free extracts Screen
Chapter 3 Nucleic Acids, Proteins, and Enzymes
 3 Nucleic Acids, Proteins, and Enzymes Chapter 3 Nucleic Acids, Proteins, and Enzymes Key Concepts 3.1 Nucleic Acids Are Informational Macromolecules 3.2 Proteins Are Polymers with Important Structural
3 Nucleic Acids, Proteins, and Enzymes Chapter 3 Nucleic Acids, Proteins, and Enzymes Key Concepts 3.1 Nucleic Acids Are Informational Macromolecules 3.2 Proteins Are Polymers with Important Structural
Refold SK Protein Refolding Kit
 Refold SK Protein Refolding Kit 2.5mg SK v5. 2 Table of Contents Introduction...4 Kit Content...5 Instruction for Use...6 Troubleshooting Guide...9 Application Notes...16 Introduction Overexpression of
Refold SK Protein Refolding Kit 2.5mg SK v5. 2 Table of Contents Introduction...4 Kit Content...5 Instruction for Use...6 Troubleshooting Guide...9 Application Notes...16 Introduction Overexpression of
Biomolecules: nucleotides
 Biomolecules: nucleotides Nucleotides and nucleic acids Nucleic acids are linear hetero-polymers adapted to maintain and transmit the genetic information Depending on their sugar moiety they can be classified
Biomolecules: nucleotides Nucleotides and nucleic acids Nucleic acids are linear hetero-polymers adapted to maintain and transmit the genetic information Depending on their sugar moiety they can be classified
Technical University of Denmark. Written examination, 29 May 2012 Course name: Life Science. Course number: Aids allowed: Written material
 1 Technical University of Denmark Written examination, 29 May 2012 Course name: Life Science Course number: 27008 Aids allowed: Written material Exam duration: 4 hours Weighting: The exam set consists
1 Technical University of Denmark Written examination, 29 May 2012 Course name: Life Science Course number: 27008 Aids allowed: Written material Exam duration: 4 hours Weighting: The exam set consists
SPRIN Ketoreductase Kit
 SPRIN Ketoreductase Kit SPRIN Ketoreductase Kit Normal (12 preparations) Product code: SKKN Covalently immobilised preparations of different Ketoreductases on Acrylic Resin Kit Description: The kit contains
SPRIN Ketoreductase Kit SPRIN Ketoreductase Kit Normal (12 preparations) Product code: SKKN Covalently immobilised preparations of different Ketoreductases on Acrylic Resin Kit Description: The kit contains
Thermo Scientific MAbPac HIC Columns. Novel Hydrophobic Interaction HPLC Columns. Designed for Monoclonal Antibody Analysis
 Thermo Scientific MAbPac HIC Columns Novel Hydrophobic Interaction HPLC Columns Designed for Monoclonal Antibody Analysis Introduction The Thermo Scientific MAbPac HIC column family is designed for separations
Thermo Scientific MAbPac HIC Columns Novel Hydrophobic Interaction HPLC Columns Designed for Monoclonal Antibody Analysis Introduction The Thermo Scientific MAbPac HIC column family is designed for separations
Structure and Function of Nucleic Acids
 Structure and Function of Nucleic Acids E T Nyahangare Class Assignment 1. Write notes and outline the role of the following in protein biosynthesis a. DNA replication b. Transcription c. Genetic code
Structure and Function of Nucleic Acids E T Nyahangare Class Assignment 1. Write notes and outline the role of the following in protein biosynthesis a. DNA replication b. Transcription c. Genetic code
Kinetics Review. Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets)
") Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
Nucleic acid and protein Flow of genetic information
 Nucleic acid and protein Flow of genetic information References: Glick, BR and JJ Pasternak, 2003, Molecular Biotechnology: Principles and Applications of Recombinant DNA, ASM Press, Washington DC, pages.
Nucleic acid and protein Flow of genetic information References: Glick, BR and JJ Pasternak, 2003, Molecular Biotechnology: Principles and Applications of Recombinant DNA, ASM Press, Washington DC, pages.
Denis V. Kurek, Sergey A. Lopatin, *Vladimir E. Tikhonov, Valery P. Varlamov
 NEW AFFINITY SORBENTS FOR PURIFICATION OF RECOMBINANT PROTEINS WITH THE USE OF CHITIN-BINDING DOMAIN AS AN AFFINITY TAG Denis V. Kurek, Sergey A. Lopatin, *Vladimir E. Tikhonov, Valery P. Varlamov Centre
NEW AFFINITY SORBENTS FOR PURIFICATION OF RECOMBINANT PROTEINS WITH THE USE OF CHITIN-BINDING DOMAIN AS AN AFFINITY TAG Denis V. Kurek, Sergey A. Lopatin, *Vladimir E. Tikhonov, Valery P. Varlamov Centre
Structure formation and association of biomolecules. Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München
 Technische Universität München") Structure formation and association of biomolecules Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München Motivation Many biomolecules are chemically synthesized
Structure formation and association of biomolecules Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München Motivation Many biomolecules are chemically synthesized
Supplemental Online Material
 Supplemental Online Material Supplemental Figures Supplemental Figure 1. A) Crosslinking efficiencies of 5 -biotin tagged oligonucleotides screened with the FASTDXL method 1. Reactive and nonreactive cysteine
Supplemental Online Material Supplemental Figures Supplemental Figure 1. A) Crosslinking efficiencies of 5 -biotin tagged oligonucleotides screened with the FASTDXL method 1. Reactive and nonreactive cysteine
Nucleic Acids. Information specifying protein structure
 Nucleic Acids Nucleic acids represent the fourth major class of biomolecules (other major classes of biomolecules are proteins, carbohydrates, fats) Genome - the genetic information of an organism Information
Nucleic Acids Nucleic acids represent the fourth major class of biomolecules (other major classes of biomolecules are proteins, carbohydrates, fats) Genome - the genetic information of an organism Information
Enzymes That Bind Nucleotides
 Enzymes That Bind Nucleotides Nucleotides play a central role in cellular metabolism, in which they are the major currency of energy exchange. They channel the energy released during the catabolism of
Enzymes That Bind Nucleotides Nucleotides play a central role in cellular metabolism, in which they are the major currency of energy exchange. They channel the energy released during the catabolism of
Information specifying protein structure. Chapter 19 Nucleic Acids Nucleotides Are the Building Blocks of Nucleic Acids
 Chapter 19 Nucleic Acids Information specifying protein structure Nucleic acids represent the fourth major class of biomolecules (other major classes of biomolecules are proteins, carbohydrates, fats)
Chapter 19 Nucleic Acids Information specifying protein structure Nucleic acids represent the fourth major class of biomolecules (other major classes of biomolecules are proteins, carbohydrates, fats)
Antibody-Antigen recognition. Structural Biology Weekend Seminar Annegret Kramer
 Antibody-Antigen recognition Structural Biology Weekend Seminar 10.07.2005 Annegret Kramer Contents Function and structure of antibodies Features of antibody-antigen interfaces Examples of antibody-antigen
Antibody-Antigen recognition Structural Biology Weekend Seminar 10.07.2005 Annegret Kramer Contents Function and structure of antibodies Features of antibody-antigen interfaces Examples of antibody-antigen
What is necessary for life?
 Life What is necessary for life? Most life familiar to us: Eukaryotes FREE LIVING Or Parasites First appeared ~ 1.5-2 10 9 years ago Requirements: DNA, proteins, lipids, carbohydrates, complex structure,
Life What is necessary for life? Most life familiar to us: Eukaryotes FREE LIVING Or Parasites First appeared ~ 1.5-2 10 9 years ago Requirements: DNA, proteins, lipids, carbohydrates, complex structure,
Please sign below if you wish to have your grades posted by the last five digits of your SSN
 BIO 226R EXAM II (Sample) PRINT YOUR NAME SSN Please sign below if you wish to have your grades posted by the last five digits of your SSN Signature BIO 226R Exam II has 6 pages, and 27 questions. There
BIO 226R EXAM II (Sample) PRINT YOUR NAME SSN Please sign below if you wish to have your grades posted by the last five digits of your SSN Signature BIO 226R Exam II has 6 pages, and 27 questions. There
PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%)
") 1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%) DESCRIPTION Nickel NTA Magnetic Agarose Beads are products that allow rapid and easy small-scale purification of
1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%) DESCRIPTION Nickel NTA Magnetic Agarose Beads are products that allow rapid and easy small-scale purification of
Protocol for in vitro transcription
 Protocol for in vitro transcription Assemble the reaction at room temperature in the following order: Component 10xTranscription Buffer rntp T7 RNA Polymerase Mix grna PCR DEPC H 2 O volume 2μl 2μl 2μl
Protocol for in vitro transcription Assemble the reaction at room temperature in the following order: Component 10xTranscription Buffer rntp T7 RNA Polymerase Mix grna PCR DEPC H 2 O volume 2μl 2μl 2μl
 The replication of DNA Kornberg 1957 Meselson and Stahl 1958 Cairns 1963 Okazaki 1968 DNA Replication The driving force for DNA synthesis. The addition of a nucleotide to a growing polynucleotide
The replication of DNA Kornberg 1957 Meselson and Stahl 1958 Cairns 1963 Okazaki 1968 DNA Replication The driving force for DNA synthesis. The addition of a nucleotide to a growing polynucleotide
Basic concepts of molecular biology
 Basic concepts of molecular biology Gabriella Trucco Email: gabriella.trucco@unimi.it What is life made of? 1665: Robert Hooke discovered that organisms are composed of individual compartments called cells
Basic concepts of molecular biology Gabriella Trucco Email: gabriella.trucco@unimi.it What is life made of? 1665: Robert Hooke discovered that organisms are composed of individual compartments called cells
Nature Structural & Molecular Biology: doi: /nsmb.2548
 Supplementary Figure 1. Structure of GltPhout. (a) Stereo view of a slice through a single GltPhout protomer shown in stick representation along with 2Fo-Fc and anomalous difference electron maps. The
Supplementary Figure 1. Structure of GltPhout. (a) Stereo view of a slice through a single GltPhout protomer shown in stick representation along with 2Fo-Fc and anomalous difference electron maps. The
Supporting Information
 Title Structure of bacterial cellulose synthase subunit D Hu, Song-Qing; Gao, Yong-Gui; Tajima, Kenji; Sunagaw Author(s) Yoda, Takanori; Shimura, Daisuke; Satoh, Yasuharu; M CitationProceedings of the
Title Structure of bacterial cellulose synthase subunit D Hu, Song-Qing; Gao, Yong-Gui; Tajima, Kenji; Sunagaw Author(s) Yoda, Takanori; Shimura, Daisuke; Satoh, Yasuharu; M CitationProceedings of the
BACTERIAL PRODUCTION EXPRESSION METHOD OVERVIEW: PEF # GENE NAME EXPRESSION VECTOR MOLECULAR WEIGHT kda (full-length) 34.
 34.") BACTERIAL PRODUCTION PEF # GENE NAME EXPRESSION VECTOR MOLECULAR WEIGHT 2015-XXXX XXXX pet-32a 50.9 kda (full-length) 34.0 kda (cleaved) EXPRESSION METHOD OVERVIEW: Plasmid DNA was transformed into BL21
BACTERIAL PRODUCTION PEF # GENE NAME EXPRESSION VECTOR MOLECULAR WEIGHT 2015-XXXX XXXX pet-32a 50.9 kda (full-length) 34.0 kda (cleaved) EXPRESSION METHOD OVERVIEW: Plasmid DNA was transformed into BL21
Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination
 201 Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination Acknowlegements. The structure of the D-8-Ad:P450cam conjugate was determined by Anna-Maria A. Hays.
201 Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination Acknowlegements. The structure of the D-8-Ad:P450cam conjugate was determined by Anna-Maria A. Hays.
RNA does not adopt the classic B-DNA helix conformation when it forms a self-complementary double helix
 Reason: RNA has ribose sugar ring, with a hydroxyl group (OH) If RNA in B-from conformation there would be unfavorable steric contact between the hydroxyl group, base, and phosphate backbone. RNA structure
Reason: RNA has ribose sugar ring, with a hydroxyl group (OH) If RNA in B-from conformation there would be unfavorable steric contact between the hydroxyl group, base, and phosphate backbone. RNA structure
Protein analysis. Dr. Mamoun Ahram Summer semester, Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5
 Protein analysis Dr. Mamoun Ahram Summer semester, 2015-2016 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5 Bases of protein separation Proteins can be purified on the basis Solubility
Protein analysis Dr. Mamoun Ahram Summer semester, 2015-2016 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5 Bases of protein separation Proteins can be purified on the basis Solubility
D-3-Phosphoglycerate Dehydrogenase* Gregory A. Grant, Zhiqin Hu, and Xiao Lan Xu
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 2, Issue of January 12, pp. 1078 1083, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Specific Interactions
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 2, Issue of January 12, pp. 1078 1083, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. Specific Interactions
IB HL Biology Test: Topics 1 and 3
 October 26, 2011 IB HL Biology Test: Topics 1 and 3 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What conditions must be met for the t-test to be applied?
October 26, 2011 IB HL Biology Test: Topics 1 and 3 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What conditions must be met for the t-test to be applied?
University of Guelph 3570 Analytical Biochemistry Fall 2004 In-class Quiz #1 Thursday, October 14, 2004 Instructor: Prof. David Josephy Solutions
 Student name: I.D. number: p. 1 University of Guelph 570 Analytical Biochemistry Fall 004 In-class Quiz #1 Thursday, October 14, 004 Instructor: Prof. David Josephy Solutions Instructions: Please write
Student name: I.D. number: p. 1 University of Guelph 570 Analytical Biochemistry Fall 004 In-class Quiz #1 Thursday, October 14, 004 Instructor: Prof. David Josephy Solutions Instructions: Please write
3 Designing Primers for Site-Directed Mutagenesis
 3 Designing Primers for Site-Directed Mutagenesis 3.1 Learning Objectives During the next two labs you will learn the basics of site-directed mutagenesis: you will design primers for the mutants you designed
3 Designing Primers for Site-Directed Mutagenesis 3.1 Learning Objectives During the next two labs you will learn the basics of site-directed mutagenesis: you will design primers for the mutants you designed
Supplementary Information
 Supplementary Information Supplementary Figures Figure S1. Study of mgtl translation in vitro. (A) Detection of 5 LR RNA using wild-type and anti-sd (91-95) substituted templates in a transcription-translation
Supplementary Information Supplementary Figures Figure S1. Study of mgtl translation in vitro. (A) Detection of 5 LR RNA using wild-type and anti-sd (91-95) substituted templates in a transcription-translation