Supporting Online Material. Av1 and Av2 were isolated and purified under anaerobic conditions according to
|
|
|
- Gabriel Booth
- 6 years ago
- Views:
Transcription
1 Supporting Online Material Materials and Methods Av1 and Av2 were isolated and purified under anaerobic conditions according to published protocols (S1). Crystals of nf-, pcp- and adp-av2:av1 complexes were prepared in sitting drops by vapor diffusion in an anaerobic chamber at room temperature. All solutions and chemicals used for crystallization were thoroughly deaerated using vacuum/ar-fill cycles. Precipitating solutions for the nf-complex contained 16-18% (w/v) PEG 10000, and those for pcp- and adp- complexes contained 18-22% PEG 8000 (w/v), in addition to 0-50 mm NaCl, 100 mm Tris buffer (ph 8.5), and 10 mm dithionite. The protein solutions were prepared by mixing appropriate amounts of Av1 and Av2 stock solutions ([Av1] stock = ~20 mg/ml, [Av2] stock = ~40 mg/ml), both of which were in 100 mm Tris (ph 7.75), 200 mm NaCl and 5 mm dithionite. Best crystals for nf-, pcp- and adp-complexes were obtained using 1:1.2 to 1:1.5 Av1:Av2 ratios (v/v), corresponding to 5 to 6-fold molar excess of Av2 over per active site of Av1. The protein solutions for pcp- and adp-complexes also contained 10 mm MgCl 2 (added from a 1 M stock), and 10 mm AMPPCP or ADP, whose 100 mm stock solutions were prepared in 100 mm Tris buffer (ph 8.5) immediately prior to setting up the crystallization trays. The crystallization drops contained 2 ml of the precipitating solution and 2 ml of the protein mixture, and the reservoir contained 250 ml of the precipitating solution. Brown, rod-shaped crystals appeared after 2-3 days and reached full growth after 1-2 weeks. The crystals for nf- and pcp-complexes were orthorhombic (P ), and those for the adp-complex were triclinic (P1) (see Table S1 for unit cell dimensions). 17
2 For data collection, suitable crystals were exchanged into 25% PEG 400 over five steps in one hour for cryoprotection and flash-frozen in liquid nitrogen. X-ray diffraction data for the nf-complex were collected at SSRL (BL 9-1) and those for pcp- and adp-complexes were acquired at ALS (BL 8.2.1) on ADSC CCD detectors using x-rays of wavelengths Å, Å, and Å, respectively. Data collection and refinement statistics are summarized in Table S1. Data sets were processed using DENZO-SCALEPACK (S2) for nf- and adpcomplexes and MOSFLM (S3)-SCALA (S4) for the pcp-complex. The initial phases were obtained by molecular replacement using MOLREP (S4) with Av1 (PDB code: 1M1N) as a model. Initial electron density maps clearly revealed the positions of the iron-sulfur clusters and most secondary structure elements of Av2 molecules, allowing their manual placement using native Av2 (PDB code: 1G5P) and ADP AlF - 4 complexed Av2 (PDB code: 1M34) as models. After an initial round of rigid-body refinement with CNS (S5) and manual rebuilding with MAIN (S6), simulated annealing using Ramachandran potentials and a very low X-ray weight was performed to regularize geometry. Ramachandran potentials were utilized during refinement of the adp-av2:av1 structure. In the absence of such potentials, the refinement was less well-behaved as evidenced by high free R-factors (>33%). Simulated-annealing, positional and thermal refinement with CNS, along with manual rebuilding and water placement with XFIT (S7) and MAIN (S6) produced the final models. In case of the adp-complex, eight-fold cyclic NCS-averaging of the individual g-subunits of the four Av2 molecules in the asymmetric unit (using MAIN), in combination with 4-fold averaging of the individual Av1 ab-dimers produced higher quality maps in the regions where the four Av2 molecules were structurally similar. The 18
3 averaged maps were used along with the non-averaged maps as a guide for manual rebuilding. Bulk solvent correction and anisotropic B-factor scaling were applied to the reflection data. NCS restraints (300 kcal mol -1 Å -2 ) were applied to four equivalent Av1 ab-dimers and kept throughout the refinement. The stereochemistry of the final models was calculated using PROCHECK (S8). Final refinement statistics are indicated in Table S1. A listing of all residues included in the final model is provided in Table S3, with an evaluation of the quality of electron density for backbone and sidechain atoms. Figures were prepared using PYMOL (S9), MOLSCRIPT (S10) and RASTER3D (S11). The coordinates of the nf-av2:av1, pcp-av2:av1 and adp-av2:av1 complexes will be deposited in the Protein Data Bank ( for release upon publication. S1. D. Wolle, C. H. Kim, D. Dean, J. B. Howard, J. Biol. Chem. 267, 3667 (1992). S2. Z. Otwinowski, W. Minor, in Macromolecular Crystallography, Part A. (1997), vol. 276, pp S3. A. G. W. Leslie, MOSFLM v6.2.0, Cambridge, U.K. (2003) S4. CCP4, Acta Crystallogr. Sect. D Biol. Crystallogr. 50 (1994). S5. A. T. Brünger et al., Acta Crystallogr. D 54, 905 (1998). S6. D. Turk, in Proceedings from the 1996 Meeting of the International Union of Crystallography Macromolecular Computing School P. E. Bourne, K. Watenpaugh, Eds. (1996). S7. D. E. McRee, J. Mol. Graphics 10, 44 (1992). S8. R. A. Laskowski, M. W. Macarthur, D. S. Moss, J. M. Thornton, J. Appl. Crystallogr. 26, 283 (1993). S9. W. L. DeLano, The PYMOL Molecular Graphics System ( (2003). S10. P. J. Kraulis, J. Appl. Cryst. 24, 946 (1991). S11. E. A. Merritt, M. E. P. Murphy, Acta Crystallogr. D 50, 869 (1994). 19
4 Table S1. Data collection and refinement statistics. nf-av2:av1 pcp-av2:av1 adp-av2:av1 Data Collection Space group P P P1 Cell dimensions a, b, c (Å) a, b, g ( ) 170.9, 75.9, , 90, , 120.9, , 90, , 141.4, , 79.4, 76.6 Resolution (Å) No. of total observations 578, , ,772 Redundancy R symm (%) a,b 14.0 (53.6) 12.3 (37.8) 7.5 (24.2) I/s(I) a 8.7 (2.5) 12.0 (3.4) 8.8 (2.7) Completeness (%) a 95.7 (89.5) 89.2 (85.7) 88.2 (72.0) Refinement Resolution (Å) R (%) a,c R free (%) a,d R.m.s. deviations from ideal values bond lengths (Å) bond angles ( ) Ramachandran statistics most favored (%) additionally allowed (%) generously allowed (%) disallowed (%)
5 a Values in parentheses correspond to the highest resolution shell. b R symm = S hkl,i (S i ÁI hkl,i - I hkl ÒÁ/S hkl,i I hkl Ò, where I hkl,i is the scaled intensity of the i th individual measurement of the reflection with indices hkl, and I hkl Ò is the mean intensity of that reflection. c R= SÁÁF obs Á-ÁF calc ÁÁ/SÁF obs Á for all reflections (no s cutoff). d Free R calculated against 5% (nf), 7% (pcp) and 10% (adp) of the reflections randomly removed. e Ramachandran potentials were applied throughout the refinement for the adp:av2-av1 complex. 21
6 Table S2 Residues included in the final structural models for the nf-av2:av1, pcp- Av2:Av1 and adp-av2:av1 complexes. Residues with observable electron density for backbone atoms are included in the structural models; residues with poorly defined sidechain electron density are also included, but with the occupancy set to zero. Chains ABCD and IJKL correspond to the subunits of crystallographically distinct Av1 tetramers, while EF, GH, MN and OP are the subunits of crystallographically distinct Av2 dimers. nf-av1:av2 Chain Backbone defined A , 332 B C D E F pcp-av1:av2 Chain Backbone defined A B C D , 258 Residues with poor sidechain density Residues with poor sidechain density E 1-50, , 74, 84, 92, , 127, 154, 159, 166, , , 194, 202, 211, , 233, 235, 262 F 1-50, , 80, 111, , 155, 163, , 233, 235, 257, 261 G 1-51, , 69, 74, 77, 116, 159, 186, 215, 221, , 230, 233, 235, 245 H , 54, 77, 118, 129, 155,
7 adp-av1:av2 Chain Backbone defined Residues with poor sidechain density A , 30, 50-51, 101, 129, 168 B , 171, , 211, 303, 400, 417 C , 30, 43, 68, 70, 288 D , 171 E 2-88, , , , , , F 1-62, , , 32, 39, 41, 46, 56, 58-60, 63, 72-74, 77, 84, 87, 100, 102, 109, 116, , 130, 135, 137, , 143, 154, 166, 171, , 179, 183, 185, , 191, 206, 211, , 219, 223, , 233, , , , , 270 8, 32, 46, 48-49, 52, 58, 60, 67-68, 70, 72-73, 77, 84, 92, 107, 110, 116, 127, 129, 137, , 143, 146, 166, , 187, 190, 194, 199, 202, 219, 221, , , 245, 251, 261 G 1-89, 93-94, , 15, 28, 31-32, 41, 46, 52, 54-56, 59, 68, 71, 75-77, 84, 100, , 108, 116, 118, 127, 135, 140, 155, 179, 182, , , , 202, 213, , 229, 235, , 249, , 262 H 2-7, 10-41, 81-88, , , , , , , , , I J K , 129, 133 L M 4-49, 52-69, 71-76, 78-88, 91-95, , , , , N 4-22, 33-36, 40-44, 81-86, , , , 150, , , , , 10, 15, 18, 20-21, 23, 26, 28-29, 31-32, 39, 41, 84, 87, 92, 100, , 121, 127, 137, 140, , 146, , , 163, 166, 169, 178, 182, , , 202, 210, , 241, , 250, 255, 257, 261, 263, 266, 269 4, 10, 15, 21, 29, 31-32, 39, 41, 43, 46-47, 52, 54-55, 60, 66-68, 71-72, 74-76, 84, 92, 100, 111, 116, 119, 135, 146, 151, , , 166, 170, 178, 182, , 199, , 207, 210, , 221, 224, 235, 241, 245, , , , 269, 271 4, 8, 10, 18, 20, 22, 33-36, 41, 84, 100, , , , 121, , 137, , , 156, , 166, 170, , , 199, , , 213, 216, , 221, , , 243, , , , , ,
8 Chain Backbone defined , , , Residues with poor sidechain density O 2-114, , 22, 26, 29, 31-33, 40, 46, 52, 57, 67-68, 74, 76-77, 84, 100, 108, 111, , , 141, 146, , , 182, 187, 191, 194, , 213, 217, 219, 221, , , 238, 241, , 249, , 262 P 6-20, 22-31, 36-39, 54-61, 80-82, , , , , , , , , , 15, 31, 58-60, 80, 92, , 115, 117, 123, 130, , , 154, 163, 165, 170, 194, 196, , 205, 208, 210, , , 229, 233, 235, , 243, 246, , 261, , 269,
9 Table S3 Pairwise comparisons of different Fe-protein conformations native Av2 Av2 + ADP alf- Av2 Leug127 Leug127+ ATP structure native Av2 (1G5P) Av2 + ADP (1FP6) alf-av2 (1M34) Leu-g127 Av2 (1G20) Leu-g127 Av2 + ATP (1G21) nf-av2 (chains EF) nf- Av2 pcp-av2 adp-av2 pcp-av2 (chains EF) pcp-av2 (chains GH) adp1-av2 (chains EF) adp2-av2 (chains GH) adp3-av2 (chains MN) adp4-av2 (chains OP)
10 Table S3 Root mean square deviations (in Å) between pairs of superimposed Fe-protein structures. The first five structures and their PDB IDs (in parentheses) have been previously deposited and serve as reference structures. When multiple Fe-protein dimers were present in the asymmetric unit of a reference structure, the first set of subunits was used. The remaining seven structures have been determined in the course of this work; the chain labels for the specified dimer are in parentheses. The Ca's of 236 residues (6-49, , , and ) present in each subunit of all available Fe-protein models were used for the comparison. The first line in each entry corresponds to the superposition of both subunits in each dimer; for example, subunits AB of one dimer are superimposed onto EF of the other (but subunits AB are not superimposed onto subunits FE). The second line corresponds to the superposition of individual subunits, calculated as the average of the A-E, A-F, B-E and B-F comparisons. The entries outlined in dark boxes designate the closest comparisons between the reference structures and the complexes determined in this work; specifically, the nf-av2 structure most closely resembles native Av2; the pcp-av2 structure most closely resembles the Leug127+ATP structure; three of the adp-av2 dimers (adp1, adp2 and adp4) most closely resemble the ADP bound state of Av2 in the absence of MoFe-protein, while the adp3 most closely resembles native Av2. In all cases, the best superpositions exhibit root mean square deviations for both the dimer and individual subunit comparisons < 1 Å. 26
11 Table S4 Intermolecular polar interactions in the nitrogenase complexes. Contacts between oxygen and/or nitrogen atoms in the indicated Fe-protein - MoFe-protein complexes were identified with the CCP4 program CONTACT (S4) using a cutoff of 3.4 Å. "mc" indicates that the contact involves the main chain atoms of a specified residue; otherwise, the contacts are mediated by side chain groups. g 1 - and g 2 - designate the Fe-protein subunits in a given dimer that interact most closely with the a- and b-subunits of the MoFe-protein, respectively. "*" designates poor electron density for the designated side chain so that reliable placement could not be established; these residues are included to identify the interaction region. Due to disordering of the Fe-protein subunits, side chain density was weak for the adp3-av2 and adp4- Av2 molecules. Contacts in the alf-av2:av1 structure (PDB ID 1M34) are included for comparison to the pcp-av2:av1 complex. 27
12 Table S4 cont'd pcp-av2:av1 intermolecular contacts Av1 residue mc Av2 residue mc Lys a51 * Gly g 1-65 mc Glu a120 Arg g Glu a120 Thr g Ile a123 mc Cys g 2-97 mc Val a124 mc Cys g 2-97 mc Gly a157 mc Arg g Ile a159 mc Gly g mc Gly a160 mc Arg g Asp a161 mc Arg g Glu a184 Arg g Glu b120 Arg g Glu b120 Thr g Asp b121 Gly g 1-65 mc Val b124 mc Cys g 1-97 mc Glu b156 mc Arg g Ile b158 mc Gly g mc Asn b163 Glu g Asn b167 Ser g mc Asn b168 Lys g mc alf-av2:av1 (PDB ID 1M34) intermolecular contacts Av1 residue mc Av2 residue mc Lys a51 Gly g 1-65 mc Glu a120 Arg g Glu a120 Thr g Ile a123 mc Cys g 2-97 mc Val a124 mc Cys g 2-97 mc Asp a128 Lys g Gly a157 mc Arg g Ile a159 mc Gly g mc Gly a160 mc Arg g Asp a161 mc Arg g Ser a165 Ser g Lys a168 Glu g Arg a182 Arg g Glu a184 Arg g Glu b120 Arg g Glu b120 Thr g Ala b123 mc Cys g 1-97 mc Val b124 mc Cys g 1-97 mc Glu b156 Arg g Ile b158 mc Gly g mc Gly b159 mc Arg g Asp b160 mc Arg g Asp b161 mc Arg g Asn b163 Glu g Asn b167 Glu g Asn b167 Ser g mc Asn b168 Lys g mc Lys b171 Asn g mc Lys b400 Asp g
13 Table S4 cont'd nf-av2:av1 intermolecular contacts Av1 residue mc Av2 residue mc Glu a120 Asn g Lys a121 Ala g 2-61 mc Lys a121 Ala g 2-64 mc Lys a121 Thr g 2-66 mc Gly a160 mc Arg g Glu b156 mc Arg g Val b157 mc Arg g * Ile b158 mc Arg g * Asp b161 Tyr g Asn b163 Lys g mc Asn b163 Asn g Lys b211 Ala g mc Lys b211 Gly g mc Lys b211 Arg g Asn b399 Glu g * Lys b400 Glu g 2-68 mc Lys b400 Leu g 2-70 mc Lys b400 Glu g Arg b401 Glu g mc adp-av2:av1 intermolecular contacts Av1 residue mc Av2 residue mc adp1 (chains ABEF) Lys a50 * Glu g 1-68 * Val a124 mc Arg g Phe a125 mc Arg g Gly a157 mc Arg g * Ile a159 mc Arg g Asp a162 Arg g * Asp b121 Gly g 1-65 mc Asp b121 Val g 1-67 mc adp2 (chains CDGH) Lys a50 Glu g 1-68 * Lys a168 Asn g mc Val b157 mc Arg g * adp3 * (chains IJMN) Ile a159 mc Arg g Gly a160 mc Arg g adp4 * (chains KLOP) Lys a26 Asp g 1-69 Lys a30 Asp g 1-69 Lys a50 Glu g 1-68 Lys a50 Glu g Leu a158 mc Arg g Asp a162 Lys g Ser a165 Lys g Lys a168 Asn g Glu b120 mc Arg g
14 Table S5. Parameters describing Av2 Av1 docking interactions for the individual adpconformers. adp1-av2, adp2-av2, adp3-av2 and adp4-av2 designate specific Fe-protein dimers in the adp-av2:av1 structure with Av2 chain identifiers EF, GH, MN and OP, that are associated with Av1 subunits AB, CD, IJ and KL, respectively. complex Buried surface area (Å 2 ) g-100 helixhelix angle f ( ) [4Fe:4S] P-cluster distance (Å) adp1-av adp2-av adp3 -Av adp4 -Av
15 Supplementary Figure Legends Fig. S1. Docking geometries and protein-protein interaction surfaces in (A) adp1- Av2:Av1, (B) adp2- Av2:Av1, and (C) adp3- Av2:Av1. Fig. S2. Crosslinking region in nf-av2:av1, illustrating the H-bonding network involving Lysb400, Glu-g112, Glu-g68 and Glu-g70. The 2F o F c map (purple) is contoured at 1s. 31
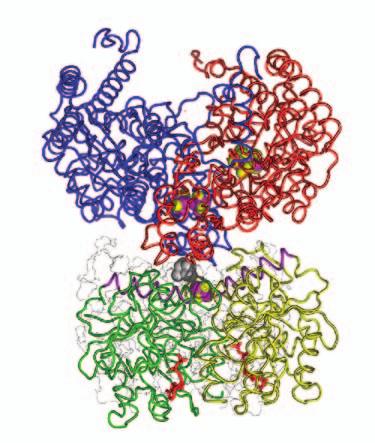
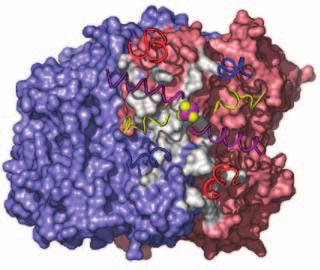
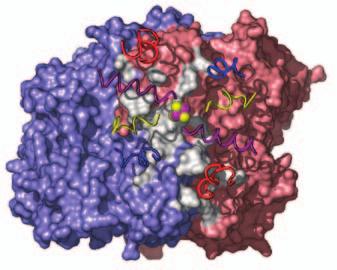
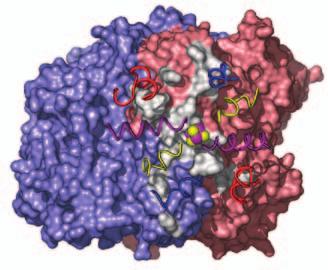
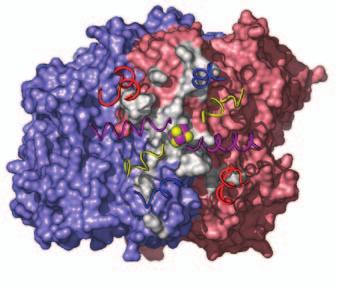
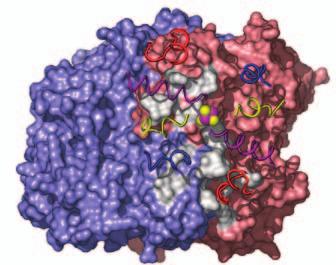
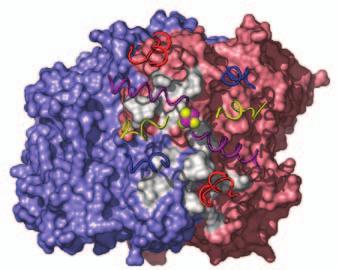
16 A adp1-av2:av1 B adp2-av2:av1 C adp4-av2:av1 Fig. S1
 -E68(Av2) Fig.")
17 -K400(Av1) -E112(Av2) -V70(Av2) -E68(Av2) Fig. S2
Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination
 201 Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination Acknowlegements. The structure of the D-8-Ad:P450cam conjugate was determined by Anna-Maria A. Hays.
201 Appendix B Dansyl probe syntheses and characterization and D-8-Ad:P450cam structure determination Acknowlegements. The structure of the D-8-Ad:P450cam conjugate was determined by Anna-Maria A. Hays.
SUPPLEMENTARY INFORMATION
 Structure of a tyrosyl-trna synthetase splicing factor bound to a group I intron RNA Paul J. Paukstelis 1, Jui-Hui Chen 2, Elaine Chase 2, Alan M. Lambowitz 1,*, and Barbara L. Golden 2,*,. 1 Institute
Structure of a tyrosyl-trna synthetase splicing factor bound to a group I intron RNA Paul J. Paukstelis 1, Jui-Hui Chen 2, Elaine Chase 2, Alan M. Lambowitz 1,*, and Barbara L. Golden 2,*,. 1 Institute
Protein expression and purification from Hi5 insect cells is as described (1, 2).
.") Materials & methods Protein expression and purification Protein expression and purification from Hi5 insect cells is as described (1, 2). The purified complex exhibited a 2:2:2 stoichiometry as determined
Materials & methods Protein expression and purification Protein expression and purification from Hi5 insect cells is as described (1, 2). The purified complex exhibited a 2:2:2 stoichiometry as determined
Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid
 Amino acid") Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction
 X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
Supporting Information
 Supporting Information Slep et al. 10.1073/pnas.0801569105 SI Materials and Methods Mouse RGS16, residues 53 180, was subcloned into a modified pgex-2t vector (GE Healthcare) to yield a thrombin-cleavable
Supporting Information Slep et al. 10.1073/pnas.0801569105 SI Materials and Methods Mouse RGS16, residues 53 180, was subcloned into a modified pgex-2t vector (GE Healthcare) to yield a thrombin-cleavable
Supporting Information
 Supporting Information Nishimura et al. 10.1073/pnas.1003553107 SI Text SI Materials and Methods. Plasmid construction. The cdnas of human Gα q were amplified and subcloned into pcmv5. Mutants of Gα q
Supporting Information Nishimura et al. 10.1073/pnas.1003553107 SI Text SI Materials and Methods. Plasmid construction. The cdnas of human Gα q were amplified and subcloned into pcmv5. Mutants of Gα q
Virtual bond representation
 Today s subjects: Virtual bond representation Coordination number Contact maps Sidechain packing: is it an instrumental way of selecting and consolidating a fold? ASA of proteins Interatomic distances
Today s subjects: Virtual bond representation Coordination number Contact maps Sidechain packing: is it an instrumental way of selecting and consolidating a fold? ASA of proteins Interatomic distances
Dr. R. Sankar, BSE 631 (2018)
") Pauling, Corey and Branson Diffraction of DNA http://www.nature.com/scitable/topicpage/dna-is-a-structure-that-encodes-biological-6493050 In short, stereochemistry is important in determining which helices
Pauling, Corey and Branson Diffraction of DNA http://www.nature.com/scitable/topicpage/dna-is-a-structure-that-encodes-biological-6493050 In short, stereochemistry is important in determining which helices
absorption spectra were measured on a Hewlett Packard 8452A diode array spectrophotometer.
 S1 Experimental General: P450cam was expressed and purified as previously described. 1 Steady state UV-visible absorption spectra were measured on a Hewlett Packard 8452A diode array spectrophotometer.
S1 Experimental General: P450cam was expressed and purified as previously described. 1 Steady state UV-visible absorption spectra were measured on a Hewlett Packard 8452A diode array spectrophotometer.
SUPPLEMENTARY INFORMATION
 This supplementary information is an extension of the letter with the same title and includes further discussion on the comparison of our designed Fe B Mb (computer model and crystal structure) with the
This supplementary information is an extension of the letter with the same title and includes further discussion on the comparison of our designed Fe B Mb (computer model and crystal structure) with the
SUPPLEMENTARY INFORMATION
 Molecular basis of RNA-dependent RNA polymerase II activity Elisabeth Lehmann, Florian Brueckner, and Patrick Cramer Gene Center Munich and Center for integrated Protein Science CiPS M, Department of Chemistry
Molecular basis of RNA-dependent RNA polymerase II activity Elisabeth Lehmann, Florian Brueckner, and Patrick Cramer Gene Center Munich and Center for integrated Protein Science CiPS M, Department of Chemistry
Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and
 and") Supplementary Tables Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and their side chain contacting residues in the second chain (B) a Interface Res. in Contacting
Supplementary Tables Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and their side chain contacting residues in the second chain (B) a Interface Res. in Contacting
Structure of the C-cadherin ectodomain and implications for the mechanism of cell adhesion
 Structure of the C-cadherin ectodomain and implications for the mechanism of cell adhesion Titus J. Boggon, John Murray, Sophie Chappuis-Flament, Ellen Wong, Barry M. Gumbiner, and Lawrence Shapiro Ref.
Structure of the C-cadherin ectodomain and implications for the mechanism of cell adhesion Titus J. Boggon, John Murray, Sophie Chappuis-Flament, Ellen Wong, Barry M. Gumbiner, and Lawrence Shapiro Ref.
Table S1. Crystallographic Data and Refinement Statistics, Related to Experimental Procedures hsddb1-drddb2 CPD
 Data collection Table S1. Crystallographic Data and Refinement Statistics, Related to Experimental Procedures hsddb1-drddb2 CPD #1 hsddb1-drddb2 CPD #2 Values in parentheses are for the highest resolution
Data collection Table S1. Crystallographic Data and Refinement Statistics, Related to Experimental Procedures hsddb1-drddb2 CPD #1 hsddb1-drddb2 CPD #2 Values in parentheses are for the highest resolution
6-Foot Mini Toober Activity
 Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
Supplemental Information. The structural basis of R Spondin recognition by LGR5 and RNF43
 Supplemental Information The structural basis of R Spondin recognition by LGR5 and RNF43 Po Han Chen 1, Xiaoyan Chen 1, Deyu Fang 2, Xiaolin He 1* 1 Department of Molecular Pharmacology and Biological
Supplemental Information The structural basis of R Spondin recognition by LGR5 and RNF43 Po Han Chen 1, Xiaoyan Chen 1, Deyu Fang 2, Xiaolin He 1* 1 Department of Molecular Pharmacology and Biological
Figure S1 Alpha-carbon backbones of components from FSH-FSHRHB complex. a, Stereo view of FSHRHB (red) with every 10 th residue marked.
 with every 10 th residue marked.") a 230 80 80 130 110 230 110 180 60 180 130 60 210 200160 40 210 200 160 40 90 20 90 20 250 150 150 190 100 250 50 30 100 220 140 220 190 140 50 30 240 240 120 70 120 70 170 170 b 70 70 60 60 20 20 50 50
a 230 80 80 130 110 230 110 180 60 180 130 60 210 200160 40 210 200 160 40 90 20 90 20 250 150 150 190 100 250 50 30 100 220 140 220 190 140 50 30 240 240 120 70 120 70 170 170 b 70 70 60 60 20 20 50 50
Computational Methods for Protein Structure Prediction
 Computational Methods for Protein Structure Prediction Ying Xu 2017/12/6 1 Outline introduction to protein structures the problem of protein structure prediction why it is possible to predict protein structures
Computational Methods for Protein Structure Prediction Ying Xu 2017/12/6 1 Outline introduction to protein structures the problem of protein structure prediction why it is possible to predict protein structures
Supplementary Figure 1. Electron microscopy of gb-698glyco/1g2 Fab complex. a)
") Supplementary Figure 1. Electron microscopy of gb-698glyco/1g2 Fab complex. a) Representative images of 2D class averages of gb-698glyc bound to 1G2 Fab. Top views of the complex were underrepresented
Supplementary Figure 1. Electron microscopy of gb-698glyco/1g2 Fab complex. a) Representative images of 2D class averages of gb-698glyc bound to 1G2 Fab. Top views of the complex were underrepresented
Diversity in DNA recognition by p53 revealed by crystal structures with Hoogsteen base pairs
 SUPPLEMENTARY INFORMATION Diversity in DNA recognition by p53 revealed by crystal structures with Hoogsteen base pairs Malka Kitayner 1, 3, Haim Rozenberg 1, 3, Remo Rohs 2, 3, Oded Suad 1, Dov Rabinovich
SUPPLEMENTARY INFORMATION Diversity in DNA recognition by p53 revealed by crystal structures with Hoogsteen base pairs Malka Kitayner 1, 3, Haim Rozenberg 1, 3, Remo Rohs 2, 3, Oded Suad 1, Dov Rabinovich
Structural bioinformatics
 Structural bioinformatics Why structures? The representation of the molecules in 3D is more informative New properties of the molecules are revealed, which can not be detected by sequences Eran Eyal Plant
Structural bioinformatics Why structures? The representation of the molecules in 3D is more informative New properties of the molecules are revealed, which can not be detected by sequences Eran Eyal Plant
Protein NMR II. Lecture 5
 Protein NMR II Lecture 5 Standard and NMR chemical shifts in proteins Residue N A A B O Ala 123.8 4.35 52.5 19.0 177.1 ys 118.8 4.65 58.8 28.6 174.8 Asp 120.4 4.76 54.1 40.8 177.2 Glu 120.2 4.29 56.7 29.7
Protein NMR II Lecture 5 Standard and NMR chemical shifts in proteins Residue N A A B O Ala 123.8 4.35 52.5 19.0 177.1 ys 118.8 4.65 58.8 28.6 174.8 Asp 120.4 4.76 54.1 40.8 177.2 Glu 120.2 4.29 56.7 29.7
A Brief Introduction to Structural Biology and Protein Crystallography
 A Brief Introduction to Structural Biology and Protein Crystallography structural biology of H2O http://courses.cm.utexas.edu/jrobertus/ch339k/overheads-1/water-structure.jpg Protein polymers fold up into
A Brief Introduction to Structural Biology and Protein Crystallography structural biology of H2O http://courses.cm.utexas.edu/jrobertus/ch339k/overheads-1/water-structure.jpg Protein polymers fold up into
doi: /nature09408 Figure S1
 doi:10.1038/nature09408 A Figure S1 www.nature.com/nature 1 RESEARCH SUPPLEMENTARY INFORMATION Figure S1 Primary sequence, and structure of NorM VC. A. Amino acid sequence alignment of NorM VC with selected
doi:10.1038/nature09408 A Figure S1 www.nature.com/nature 1 RESEARCH SUPPLEMENTARY INFORMATION Figure S1 Primary sequence, and structure of NorM VC. A. Amino acid sequence alignment of NorM VC with selected
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1133488/dc1 Supporting Online Material for An Inward-Facing Conformation of a Putative Metal-Chelate Type ABC Transporter H. W. Pinkett, A. T. Lee, P. Lum, K. P. Locher,
www.sciencemag.org/cgi/content/full/1133488/dc1 Supporting Online Material for An Inward-Facing Conformation of a Putative Metal-Chelate Type ABC Transporter H. W. Pinkett, A. T. Lee, P. Lum, K. P. Locher,
Supporting Information Contents
 Supporting Information Choy Theng Loh, Kiyoshi Ozawa, Kellie L. Tuck, Nicholas Barlow, Thomas Huber, Gottfried Otting, and Bim Graham Lanthanide tags for site-specific ligation to an unnatural amino acid
Supporting Information Choy Theng Loh, Kiyoshi Ozawa, Kellie L. Tuck, Nicholas Barlow, Thomas Huber, Gottfried Otting, and Bim Graham Lanthanide tags for site-specific ligation to an unnatural amino acid
Description of Changes and Corrections for PDB File Format Version 4.0. Provisional Document April 12, 2011
 Description of Changes and Corrections for PDB File Format Version 4.0 Provisional Document April 12, 2011 The wwpdb has reviewed the PDB archive and created a new set of corrected files that will be released
Description of Changes and Corrections for PDB File Format Version 4.0 Provisional Document April 12, 2011 The wwpdb has reviewed the PDB archive and created a new set of corrected files that will be released
In silico measurements of twist and bend. moduli for beta solenoid protein self-
 In silico measurements of twist and bend moduli for beta solenoid protein self- assembly units Leonard P. Heinz, Krishnakumar M. Ravikumar, and Daniel L. Cox Department of Physics and Institute for Complex
In silico measurements of twist and bend moduli for beta solenoid protein self- assembly units Leonard P. Heinz, Krishnakumar M. Ravikumar, and Daniel L. Cox Department of Physics and Institute for Complex
Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the
 Supplementary information, Data S1 Methods Clones and protein preparation Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the Saccharomyces cerevisiae genomic DNA by polymerase chain
Supplementary information, Data S1 Methods Clones and protein preparation Six genes, Lsm1, Lsm2, Lsm3, Lsm5, Lsm6, and Lsm7, were amplified from the Saccharomyces cerevisiae genomic DNA by polymerase chain
Structure formation and association of biomolecules. Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München
 Technische Universität München") Structure formation and association of biomolecules Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München Motivation Many biomolecules are chemically synthesized
Structure formation and association of biomolecules Prof. Dr. Martin Zacharias Lehrstuhl für Molekulardynamik (T38) Technische Universität München Motivation Many biomolecules are chemically synthesized
2 Results and Discussion
 ZHAO Ying et al: Crystallization of Nitrogenase MoFe Protein (NifB Av1) from a nifb Mutated Strain UW45 of Azotobacter vinelandii 821 In order to prevent the NifB Av1 from a damage resulted possibly from
ZHAO Ying et al: Crystallization of Nitrogenase MoFe Protein (NifB Av1) from a nifb Mutated Strain UW45 of Azotobacter vinelandii 821 In order to prevent the NifB Av1 from a damage resulted possibly from
Chapter 8. One-Dimensional Structural Properties of Proteins in the Coarse-Grained CABS Model. Sebastian Kmiecik and Andrzej Kolinski.
 Chapter 8 One-Dimensional Structural Properties of Proteins in the Coarse-Grained CABS Model Abstract Despite the significant increase in computational power, molecular modeling of protein structure using
Chapter 8 One-Dimensional Structural Properties of Proteins in the Coarse-Grained CABS Model Abstract Despite the significant increase in computational power, molecular modeling of protein structure using
The Skap-hom Dimerization and PH Domains Comprise
 Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
Supporting Information
 Supporting Information Xiao et al. 10.1073/pnas.1309211110 SI Methods Protein Expression and Purification. Bacmid was generated and recombinant baculovirus amplified using standard procedures (Bac-to-Bac;
Supporting Information Xiao et al. 10.1073/pnas.1309211110 SI Methods Protein Expression and Purification. Bacmid was generated and recombinant baculovirus amplified using standard procedures (Bac-to-Bac;
Programme Good morning and summary of last week Levels of Protein Structure - I Levels of Protein Structure - II
 Programme 8.00-8.10 Good morning and summary of last week 8.10-8.30 Levels of Protein Structure - I 8.30-9.00 Levels of Protein Structure - II 9.00-9.15 Break 9.15-11.15 Exercise: Building a protein model
Programme 8.00-8.10 Good morning and summary of last week 8.10-8.30 Levels of Protein Structure - I 8.30-9.00 Levels of Protein Structure - II 9.00-9.15 Break 9.15-11.15 Exercise: Building a protein model
The YTH domain (residues ) of human YTHDF2 (NP_ ) was subcloned
 of human YTHDF2 (NP_ ) was subcloned") Supplementary information, Data S1 Materials and Methods Protein Expression, Purification and Crystallization The YTH domain (residues 383-553) of human YTHDF2 (NP_057342.2) was subcloned into a modified
Supplementary information, Data S1 Materials and Methods Protein Expression, Purification and Crystallization The YTH domain (residues 383-553) of human YTHDF2 (NP_057342.2) was subcloned into a modified
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
Nature Structural & Molecular Biology: doi: /nsmb.1969
 Supplementary Methods Structure determination All the diffraction data sets were collected on BL-41XU (using ADSC Quantum 315 HE CCD detector) at SPring8 (Harima, Japan) or on BL5A (using ADSC Quantum
Supplementary Methods Structure determination All the diffraction data sets were collected on BL-41XU (using ADSC Quantum 315 HE CCD detector) at SPring8 (Harima, Japan) or on BL5A (using ADSC Quantum
Steps in solving a structure. Diffraction experiment. Obtaining well-diffracting crystals. Three dimensional crystals
 Protein structure from X-ray diffraction Diffraction images: ciprocal space Protein, chemical structure: IALEFGPSLKMNE Conformation, 3D-structure: CRYST1 221.200 73.600 80.900 90.00 90.00 90.00 P 21 21
Protein structure from X-ray diffraction Diffraction images: ciprocal space Protein, chemical structure: IALEFGPSLKMNE Conformation, 3D-structure: CRYST1 221.200 73.600 80.900 90.00 90.00 90.00 P 21 21
Acceleration of protein folding by four orders of magnitude through a single amino acid substitution
 Acceleration of protein folding by four orders of magnitude through a single amino acid substitution Daniel J. A. Roderer 1, Martin A. Schärer 1, Marina Rubini 2 * and Rudi Glockshuber 1 AUTHOR ADDRESS
Acceleration of protein folding by four orders of magnitude through a single amino acid substitution Daniel J. A. Roderer 1, Martin A. Schärer 1, Marina Rubini 2 * and Rudi Glockshuber 1 AUTHOR ADDRESS
Zwitterion Chromatography ZIC
 Zwitterion Chromatography ZIC A novel technique, with unique selectivity, suitable for preparative scale separations? PhD Einar Pontén What is Zwitterion Chromatography? Our definition: Liquid chromatography
Zwitterion Chromatography ZIC A novel technique, with unique selectivity, suitable for preparative scale separations? PhD Einar Pontén What is Zwitterion Chromatography? Our definition: Liquid chromatography
Protein 3D Structure Prediction
 Protein 3D Structure Prediction Michael Tress CNIO ?? MREYKLVVLGSGGVGKSALTVQFVQGIFVDE YDPTIEDSYRKQVEVDCQQCMLEILDTAGTE QFTAMRDLYMKNGQGFALVYSITAQSTFNDL QDLREQILRVKDTEDVPMILVGNKCDLEDER VVGKEQGQNLARQWCNCAFLESSAKSKINVN
Protein 3D Structure Prediction Michael Tress CNIO ?? MREYKLVVLGSGGVGKSALTVQFVQGIFVDE YDPTIEDSYRKQVEVDCQQCMLEILDTAGTE QFTAMRDLYMKNGQGFALVYSITAQSTFNDL QDLREQILRVKDTEDVPMILVGNKCDLEDER VVGKEQGQNLARQWCNCAFLESSAKSKINVN
Supplementary Data for Monti, et al.
 Supplementary Data for Monti, et al. Supplementary Figure S1 Legend to Supplementary Figure S1 Tumor spectrum associated with germline p53 alleles (restricted to the 7 most frequent tissue targets). Structural
Supplementary Data for Monti, et al. Supplementary Figure S1 Legend to Supplementary Figure S1 Tumor spectrum associated with germline p53 alleles (restricted to the 7 most frequent tissue targets). Structural
466 Asn (N) to Ala (A) Generate beta dimer Interface
 to Ala (A) Generate beta dimer Interface") Table S1: Amino acid changes to the HexA α-subunit to convert the dimer interface from α to β and to introduce the putative GM2A binding surface from β- onto the α- subunit Residue position (α-numbering)
Table S1: Amino acid changes to the HexA α-subunit to convert the dimer interface from α to β and to introduce the putative GM2A binding surface from β- onto the α- subunit Residue position (α-numbering)
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature06147 SUPPLEMENTARY INFORMATION Figure S1 The genomic and domain structure of Dscam. The Dscam gene comprises 24 exons, encoding a signal peptide (SP), 10 IgSF domains, 6 fibronectin
doi: 10.1038/nature06147 SUPPLEMENTARY INFORMATION Figure S1 The genomic and domain structure of Dscam. The Dscam gene comprises 24 exons, encoding a signal peptide (SP), 10 IgSF domains, 6 fibronectin
Amino Acid Sequences and Evolutionary Relationships
 Amino Acid Sequences and Evolutionary Relationships One technique used to determine evolutionary relationships is to study the biochemical similarity of organisms. Though molds, aardvarks, and humans appear
Amino Acid Sequences and Evolutionary Relationships One technique used to determine evolutionary relationships is to study the biochemical similarity of organisms. Though molds, aardvarks, and humans appear
Comparative Modeling Part 1. Jaroslaw Pillardy Computational Biology Service Unit Cornell Theory Center
 Comparative Modeling Part 1 Jaroslaw Pillardy Computational Biology Service Unit Cornell Theory Center Function is the most important feature of a protein Function is related to structure Structure is
Comparative Modeling Part 1 Jaroslaw Pillardy Computational Biology Service Unit Cornell Theory Center Function is the most important feature of a protein Function is related to structure Structure is
Supplementary Fig. 1. Initial electron density maps for the NOX-D20:mC5a complex obtained after SAD-phasing. (a) Initial experimental electron
 Initial experimental electron") Supplementary Fig. 1. Initial electron density maps for the NOX-D20:mC5a complex obtained after SAD-phasing. (a) Initial experimental electron density map obtained after SAD-phasing and density modification
Supplementary Fig. 1. Initial electron density maps for the NOX-D20:mC5a complex obtained after SAD-phasing. (a) Initial experimental electron density map obtained after SAD-phasing and density modification
Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and
 Materials and Methods Immunoprecipitation and immunoblot analysis Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and immunoblot analyses with corresponding
Materials and Methods Immunoprecipitation and immunoblot analysis Proteins were extracted from cultured cells using a modified buffer, and immunoprecipitation and immunoblot analyses with corresponding
Solutions to Problem Set 1
 MIT Department of Biology 7.014 Introductory Biology, Spring 004 Question 1 Solutions to 7.014 Problem Set 1 a) Describe the conditions of the atmosphere on prebiotic earth and how these conditions differ
MIT Department of Biology 7.014 Introductory Biology, Spring 004 Question 1 Solutions to 7.014 Problem Set 1 a) Describe the conditions of the atmosphere on prebiotic earth and how these conditions differ
NUCLEOTIDE DEPENDENT CONFORMATIONAL CHANGES. IN THE NITROGENASE Fe PROTEIN. Sanchayita Sen
 NUCLEOTIDE DEPENDENT CONFORMATIONAL CHANGES IN THE NITROGENASE Fe PROTEIN by Sanchayita Sen A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in
NUCLEOTIDE DEPENDENT CONFORMATIONAL CHANGES IN THE NITROGENASE Fe PROTEIN by Sanchayita Sen A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in
Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis
 Supplemental material to accompany Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis Jacob Bauer, a Gabriela Ondrovičová, a Lucie Najmanová, b Vladimír Pevala,
Supplemental material to accompany Structure and Possible Mechanism of the CcbJ Methyltransferase from Streptomyces caelestis Jacob Bauer, a Gabriela Ondrovičová, a Lucie Najmanová, b Vladimír Pevala,
03-511/711 Computational Genomics and Molecular Biology, Fall
 03-511/711 Computational Genomics and Molecular Biology, Fall 2011 1 Problem Set 0 Due Tuesday, September 6th This homework is intended to be a self-administered placement quiz, to help you (and me) determine
03-511/711 Computational Genomics and Molecular Biology, Fall 2011 1 Problem Set 0 Due Tuesday, September 6th This homework is intended to be a self-administered placement quiz, to help you (and me) determine
Solving Structure Based Design Problems using Discovery Studio 1.7 Building a Flexible Docking Protocol
 Solving Structure Based Design Problems using Discovery Studio 1.7 Building a Flexible Docking Protocol C. M. (Venkat) Venkatachalam Fellow, Life Sciences Dipesh Risal Marketing, Life Sciences Overview
Solving Structure Based Design Problems using Discovery Studio 1.7 Building a Flexible Docking Protocol C. M. (Venkat) Venkatachalam Fellow, Life Sciences Dipesh Risal Marketing, Life Sciences Overview
Packing of Secondary Structures
 7.88 Lecture Notes - 5 7.24/7.88J/5.48J The Protein Folding and Human Disease Packing of Secondary Structures Packing of Helices against sheets Packing of sheets against sheets Parallel Orthogonal Table:
7.88 Lecture Notes - 5 7.24/7.88J/5.48J The Protein Folding and Human Disease Packing of Secondary Structures Packing of Helices against sheets Packing of sheets against sheets Parallel Orthogonal Table:
research papers 784 Wang et al. C-phycocyanin Acta Cryst. (2001). D57, 784± Introduction
. D57, 784± Introduction") Acta Crystallographica Section D Biological Crystallography ISSN 0907-4449 Structure of C-phycocyanin from Spirulina platensis at 2.2 AÊ resolution: a novel monoclinic crystal form for phycobiliproteins
Acta Crystallographica Section D Biological Crystallography ISSN 0907-4449 Structure of C-phycocyanin from Spirulina platensis at 2.2 AÊ resolution: a novel monoclinic crystal form for phycobiliproteins
Ali Yaghi. Tamara Wahbeh. Mamoun Ahram
 28 Ali Yaghi Tamara Wahbeh Mamoun Ahram This sheet is a continuation of protein purification methods. Isoelectric focusing Separation of proteins based on Isoelectric points(charge),and it is a horizontal
28 Ali Yaghi Tamara Wahbeh Mamoun Ahram This sheet is a continuation of protein purification methods. Isoelectric focusing Separation of proteins based on Isoelectric points(charge),and it is a horizontal
Homology Modelling. Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen
 Homology Modelling Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen Why are Protein Structures so Interesting? They provide a detailed picture of interesting biological features,
Homology Modelling Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen Why are Protein Structures so Interesting? They provide a detailed picture of interesting biological features,
Homology Modelling. Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen
 Homology Modelling Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen Why are Protein Structures so Interesting? They provide a detailed picture of interesting biological features,
Homology Modelling Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen Why are Protein Structures so Interesting? They provide a detailed picture of interesting biological features,
The effect of metal distribution on the luminescence properties of mixedlanthanide metal-organic frameworks
 Electronic Supplementary Material (ESI) for Dalton Transactions. This journal is The Royal Society of Chemistry 2018 The effect of metal distribution on the luminescence properties of mixedlanthanide metal-organic
Electronic Supplementary Material (ESI) for Dalton Transactions. This journal is The Royal Society of Chemistry 2018 The effect of metal distribution on the luminescence properties of mixedlanthanide metal-organic
Amino Acids and Proteins
 Various Functions of Proteins SB203 Amino Acids and Proteins Jirundon Yuvaniyama, Ph.D. Department of Biochemistry Faculty of Science Mahidol University Enzymes Transport proteins utrient and storage proteins
Various Functions of Proteins SB203 Amino Acids and Proteins Jirundon Yuvaniyama, Ph.D. Department of Biochemistry Faculty of Science Mahidol University Enzymes Transport proteins utrient and storage proteins
Fundamentals of Protein Structure
 Outline Fundamentals of Protein Structure Yu (Julie) Chen and Thomas Funkhouser Princeton University CS597A, Fall 2005 Protein structure Primary Secondary Tertiary Quaternary Forces and factors Levels
Outline Fundamentals of Protein Structure Yu (Julie) Chen and Thomas Funkhouser Princeton University CS597A, Fall 2005 Protein structure Primary Secondary Tertiary Quaternary Forces and factors Levels
research papers 1. Introduction
 Acta Crystallographica Section D Biological Crystallography ISSN 0907-4449 NADP + and NAD + binding to the dual coenzyme specific enzyme Leuconostoc mesenteroides glucose 6-phosphate dehydrogenase: different
Acta Crystallographica Section D Biological Crystallography ISSN 0907-4449 NADP + and NAD + binding to the dual coenzyme specific enzyme Leuconostoc mesenteroides glucose 6-phosphate dehydrogenase: different
Amino Acid Sequences and Evolutionary Relationships. How do similarities in amino acid sequences of various species provide evidence for evolution?
 Amino Acid Sequences and Evolutionary Relationships Name: How do similarities in amino acid sequences of various species provide evidence for evolution? An important technique used in determining evolutionary
Amino Acid Sequences and Evolutionary Relationships Name: How do similarities in amino acid sequences of various species provide evidence for evolution? An important technique used in determining evolutionary
Amino Acid Sequences and Evolutionary Relationships
 Amino Acid Sequences and Evolutionary Relationships Pre-Lab Discussion Homologous structures -- those structures believed to have a common origin but not necessarily a common function -- provide some of
Amino Acid Sequences and Evolutionary Relationships Pre-Lab Discussion Homologous structures -- those structures believed to have a common origin but not necessarily a common function -- provide some of
Crystal Structures of the Oxidized and Reduced Forms of UDP-galactose 4-Epimerase Isolated from Escherichia coli,
 Biochemistry 1996, 35, 2557-2566 2557 Crystal Structures of the Oxidized and Reduced Forms of UDP-galactose 4-Epimerase Isolated from Escherichia coli, James B. Thoden, Perry A. Frey, and Hazel M. Holden*
Biochemistry 1996, 35, 2557-2566 2557 Crystal Structures of the Oxidized and Reduced Forms of UDP-galactose 4-Epimerase Isolated from Escherichia coli, James B. Thoden, Perry A. Frey, and Hazel M. Holden*
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature10258 Supplementary Figure 1 Reconstitution of the CENP-A nucleosome with recombinant human histones H2A, H2B, H4, and CENP-A. a, Purified recombinant human
SUPPLEMENTARY INFORMATION doi:10.1038/nature10258 Supplementary Figure 1 Reconstitution of the CENP-A nucleosome with recombinant human histones H2A, H2B, H4, and CENP-A. a, Purified recombinant human
Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1.
 GR24 (μm) 0 20 0 20 GST-ShHTL7 anti-gst His-MAX2 His-COI1 PVDF staining Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1. Pull-down assays using GST-ShHTL7
GR24 (μm) 0 20 0 20 GST-ShHTL7 anti-gst His-MAX2 His-COI1 PVDF staining Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1. Pull-down assays using GST-ShHTL7
Improving RNA Crystallographic Models Using Rosetta. Fang-Chieh Chou (Das Lab) For RosettaCon 2011
 For RosettaCon 2011") Improving RNA Crystallographic Models Using Rosetta Fang-Chieh Chou (Das Lab) For RosettaCon 211 The number RNA crystal structures increased explosively after the first Ribosome crystal structure is solved
Improving RNA Crystallographic Models Using Rosetta Fang-Chieh Chou (Das Lab) For RosettaCon 211 The number RNA crystal structures increased explosively after the first Ribosome crystal structure is solved
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 29 September 2005
 Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 9 September 005 Focus concept Purification of a novel seed storage protein allows sequence analysis and
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 9 September 005 Focus concept Purification of a novel seed storage protein allows sequence analysis and
DNA/Protein Binding, Molecular Docking and in Vitro Anti-cancer Activity of some Thioether-Dipyrrinato Complexes
 DNA/Protein Binding, Molecular Docking and in Vitro Anti-cancer Activity of some Thioether-Dipyrrinato Complexes Rakesh Kumar Gupta, Gunjan Sharma, ξ Rampal Pandey, Amit Kumar, Biplob Koch, ξ Pei- Zhou
DNA/Protein Binding, Molecular Docking and in Vitro Anti-cancer Activity of some Thioether-Dipyrrinato Complexes Rakesh Kumar Gupta, Gunjan Sharma, ξ Rampal Pandey, Amit Kumar, Biplob Koch, ξ Pei- Zhou
SUPPLEMENTARY INFORMATION. Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly
 SUPPLEMENTARY INFORMATION Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly Dustin J. E. Huard, Kathleen M. Kane and F. Akif Tezcan* Department of Chemistry and Biochemistry,
SUPPLEMENTARY INFORMATION Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly Dustin J. E. Huard, Kathleen M. Kane and F. Akif Tezcan* Department of Chemistry and Biochemistry,
Basic concepts of molecular biology
 Basic concepts of molecular biology Gabriella Trucco Email: gabriella.trucco@unimi.it Life The main actors in the chemistry of life are molecules called proteins nucleic acids Proteins: many different
Basic concepts of molecular biology Gabriella Trucco Email: gabriella.trucco@unimi.it Life The main actors in the chemistry of life are molecules called proteins nucleic acids Proteins: many different
Key crystallographic concepts: Theory of diffraction. (Crystallography y without tears, Part 1)
") Protein Crystallography (3) Key crystallographic concepts: Theory of diffraction. (Crystallography y without tears, Part 1) Cele Abad-Zapatero University of Illinois at Chicago Center for Pharmaceutical
Protein Crystallography (3) Key crystallographic concepts: Theory of diffraction. (Crystallography y without tears, Part 1) Cele Abad-Zapatero University of Illinois at Chicago Center for Pharmaceutical
The study of protein secondary structure and stability at equilibrium ABSTRACT
 The study of protein secondary structure and stability at equilibrium Michelle Planicka Dept. of Physics, North Georgia College and State University, Dahlonega, GA REU, Dept. of Physics, University of
The study of protein secondary structure and stability at equilibrium Michelle Planicka Dept. of Physics, North Georgia College and State University, Dahlonega, GA REU, Dept. of Physics, University of
Molecular design principles underlying β-strand swapping. in the adhesive dimerization of cadherins
 Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
Supplementary Figure 1
 Supplementary Figure 1 2 Supplementary Figure 1: Sequence alignment of HsHSD17B8 and HsCBR4 of with KAR orthologs. The secondary structure elements as calculated by DSSP and residue numbers are displayed
Supplementary Figure 1 2 Supplementary Figure 1: Sequence alignment of HsHSD17B8 and HsCBR4 of with KAR orthologs. The secondary structure elements as calculated by DSSP and residue numbers are displayed
Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport
 Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport Ivan Laponogov 1,2, Dennis A. Veselkov 1, Isabelle M-T. Crevel
Supplementary materials for Structure of an open clamp type II topoisomerase-dna complex provides a mechanism for DNA capture and transport Ivan Laponogov 1,2, Dennis A. Veselkov 1, Isabelle M-T. Crevel
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor
 Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Focus concept Purification of a novel seed storage protein allows sequence analysis and determination of the protein
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Focus concept Purification of a novel seed storage protein allows sequence analysis and determination of the protein
IB HL Biology Test: Topics 1 and 3
 October 26, 2011 IB HL Biology Test: Topics 1 and 3 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What conditions must be met for the t-test to be applied?
October 26, 2011 IB HL Biology Test: Topics 1 and 3 Multiple Choice Identify the choice that best completes the statement or answers the question. 1. What conditions must be met for the t-test to be applied?
Supplementary Information For. A genetically encoded tool for manipulation of NADP + /NADPH in living cells
 Supplementary Information For A genetically encoded tool for manipulation of NADP + /NADPH in living cells Valentin Cracan 1,2,3, Denis V. Titov 1,2,3, Hongying Shen 1,2,3, Zenon Grabarek 1* and Vamsi
Supplementary Information For A genetically encoded tool for manipulation of NADP + /NADPH in living cells Valentin Cracan 1,2,3, Denis V. Titov 1,2,3, Hongying Shen 1,2,3, Zenon Grabarek 1* and Vamsi
Supplementary Materials. Figure S1 Chemical structures of cisplatin and carboplatin.
 Supplementary Materials Figure S1 Chemical structures of cisplatin and carboplatin. Figure S2 (a) RMS difference plot between the RT and 100K structures for the carboplatin_dmso case after 13 months of
Supplementary Materials Figure S1 Chemical structures of cisplatin and carboplatin. Figure S2 (a) RMS difference plot between the RT and 100K structures for the carboplatin_dmso case after 13 months of
Main-Chain Conformational Tendencies of Amino Acids
 PROTEINS: Structure, Function, and Bioinformatics 60:679 689 (2005) Main-Chain Conformational Tendencies of Amino Acids Robert J. Anderson, 1,2 Zhiping Weng, 2 Robert K. Campbell, 1 and Xuliang Jiang 1
PROTEINS: Structure, Function, and Bioinformatics 60:679 689 (2005) Main-Chain Conformational Tendencies of Amino Acids Robert J. Anderson, 1,2 Zhiping Weng, 2 Robert K. Campbell, 1 and Xuliang Jiang 1
Protein Structure Prediction by Constraint Logic Programming
 MPRI C2-19 Protein Structure Prediction by Constraint Logic Programming François Fages, Constraint Programming Group, INRIA Rocquencourt mailto:francois.fages@inria.fr http://contraintes.inria.fr/ Molecules
MPRI C2-19 Protein Structure Prediction by Constraint Logic Programming François Fages, Constraint Programming Group, INRIA Rocquencourt mailto:francois.fages@inria.fr http://contraintes.inria.fr/ Molecules
Dynamic Features of camp-dependent Protein Kinase Revealed by Apoenzyme Crystal Structure
 doi:10.1016/s0022-2836(02)01446-8 J. Mol. Biol. (2003) 327, 159 171 Dynamic Features of camp-dependent Protein Kinase Revealed by Apoenzyme Crystal Structure Pearl Akamine 1, Madhusudan 1, Jian Wu 1, Nguyen-Huu
doi:10.1016/s0022-2836(02)01446-8 J. Mol. Biol. (2003) 327, 159 171 Dynamic Features of camp-dependent Protein Kinase Revealed by Apoenzyme Crystal Structure Pearl Akamine 1, Madhusudan 1, Jian Wu 1, Nguyen-Huu
CFSSP: Chou and Fasman Secondary Structure Prediction server
 Wide Spectrum, Vol. 1, No. 9, (2013) pp 15-19 CFSSP: Chou and Fasman Secondary Structure Prediction server T. Ashok Kumar Department of Bioinformatics, Noorul Islam College of Arts and Science, Kumaracoil
Wide Spectrum, Vol. 1, No. 9, (2013) pp 15-19 CFSSP: Chou and Fasman Secondary Structure Prediction server T. Ashok Kumar Department of Bioinformatics, Noorul Islam College of Arts and Science, Kumaracoil
1/4/18 NUCLEIC ACIDS. Nucleic Acids. Nucleic Acids. ECS129 Instructor: Patrice Koehl
 NUCLEIC ACIDS ECS129 Instructor: Patrice Koehl Nucleic Acids Nucleotides DNA Structure RNA Synthesis Function Secondary structure Tertiary interactions Wobble hypothesis DNA RNA Replication Transcription
NUCLEIC ACIDS ECS129 Instructor: Patrice Koehl Nucleic Acids Nucleotides DNA Structure RNA Synthesis Function Secondary structure Tertiary interactions Wobble hypothesis DNA RNA Replication Transcription
NUCLEIC ACIDS. ECS129 Instructor: Patrice Koehl
 NUCLEIC ACIDS ECS129 Instructor: Patrice Koehl Nucleic Acids Nucleotides DNA Structure RNA Synthesis Function Secondary structure Tertiary interactions Wobble hypothesis DNA RNA Replication Transcription
NUCLEIC ACIDS ECS129 Instructor: Patrice Koehl Nucleic Acids Nucleotides DNA Structure RNA Synthesis Function Secondary structure Tertiary interactions Wobble hypothesis DNA RNA Replication Transcription
SUPPLEMENTARY INFORMATION
 Electronic Supplementary Material (ESI) for Dalton Transactions. This journal is The Royal Society of Chemistry 2014 SUPPLEMENTARY INFORMATION NO-binding in {Ru(NO) 2 } 8 -type [Ru(NO) 2 (PR 3 ) 2 X]BF
Electronic Supplementary Material (ESI) for Dalton Transactions. This journal is The Royal Society of Chemistry 2014 SUPPLEMENTARY INFORMATION NO-binding in {Ru(NO) 2 } 8 -type [Ru(NO) 2 (PR 3 ) 2 X]BF
CHAPTER 7 CRYSTALLIZATION AND CRYSTAL STRUCTURE STUDIES OF PGA
 CHAPTER 7 CRYSTALLIZATION AND CRYSTAL STRUCTURE STUDIES OF PGA 7.. Introduction This chapter describes the crystallization and structural studies attempted on two penicillin acylases KcPGA and AfPGA. Because
CHAPTER 7 CRYSTALLIZATION AND CRYSTAL STRUCTURE STUDIES OF PGA 7.. Introduction This chapter describes the crystallization and structural studies attempted on two penicillin acylases KcPGA and AfPGA. Because
Folding simulation: self-organization of 4-helix bundle protein. yellow = helical turns
 Folding simulation: self-organization of 4-helix bundle protein yellow = helical turns Protein structure Protein: heteropolymer chain made of amino acid residues R + H 3 N - C - COO - H φ ψ Chain of amino
Folding simulation: self-organization of 4-helix bundle protein yellow = helical turns Protein structure Protein: heteropolymer chain made of amino acid residues R + H 3 N - C - COO - H φ ψ Chain of amino
Supporting Information
 Supporting Information Peroxidase vs. Peroxygenase Activity: Substrate Substituent Effects as Modulators of Enzyme Function in the Multifunctional Catalytic Globin Dehaloperoxidase Ashlyn H. McGuire, Leiah
Supporting Information Peroxidase vs. Peroxygenase Activity: Substrate Substituent Effects as Modulators of Enzyme Function in the Multifunctional Catalytic Globin Dehaloperoxidase Ashlyn H. McGuire, Leiah
Bio5325 Fall Crystal Vocabulary
 Crystals and Crystallization Bio5325 Fall 2007 Crystal Vocabulary Mosaicity (mosaic spread) Protein crystals are imperfect, consisting of a mosaic of domains that are slightly misaligned. As a result,
Crystals and Crystallization Bio5325 Fall 2007 Crystal Vocabulary Mosaicity (mosaic spread) Protein crystals are imperfect, consisting of a mosaic of domains that are slightly misaligned. As a result,
Figure S1
 Supplementary Figure 1 The distribution of chlorophyll containing complexes eluted from DEAE-cellulose column in a sucrose gradient tube Six pigment-containing bands were resolved and identified as: B1,
Supplementary Figure 1 The distribution of chlorophyll containing complexes eluted from DEAE-cellulose column in a sucrose gradient tube Six pigment-containing bands were resolved and identified as: B1,
The Cys 2. His 2. Research Article 451
 Research Article 451 High-resolution structures of variant Zif268 DNA complexes: implications for understanding zinc finger DNA recognition Monicia Elrod-Erickson 1, Timothy E Benson 1 and Carl O Pabo
Research Article 451 High-resolution structures of variant Zif268 DNA complexes: implications for understanding zinc finger DNA recognition Monicia Elrod-Erickson 1, Timothy E Benson 1 and Carl O Pabo
A Designed 3D Porous Hydrogen-Bonding Network Based on a Metal-Organic Polyhedron
 Supporting Information A Designed 3D Porous Hydrogen-Bonding Network Based on a Metal-Organic Polyhedron Wei Wei,*, Wanlong Li, Xingzhu Wang, Jieya He Department of Chemistry, Capital Normal University,
Supporting Information A Designed 3D Porous Hydrogen-Bonding Network Based on a Metal-Organic Polyhedron Wei Wei,*, Wanlong Li, Xingzhu Wang, Jieya He Department of Chemistry, Capital Normal University,
Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein
 Cell Wall Recycling Protein") Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
Hmwk # 8 : DNA-Binding Proteins : Part II
 The purpose of this exercise is : Hmwk # 8 : DNA-Binding Proteins : Part II 1). to examine the case of a tandem head-to-tail homodimer binding to DNA 2). to view a Zn finger motif 3). to consider the case
The purpose of this exercise is : Hmwk # 8 : DNA-Binding Proteins : Part II 1). to examine the case of a tandem head-to-tail homodimer binding to DNA 2). to view a Zn finger motif 3). to consider the case
Protocol S1: Supporting Information
 Protocol S1: Supporting Information Basis for the specificity of the kinase domain of Abl for peptide substrates The crystal structures reported in this work were obtained using two different ATP analog-peptide
Protocol S1: Supporting Information Basis for the specificity of the kinase domain of Abl for peptide substrates The crystal structures reported in this work were obtained using two different ATP analog-peptide