JBC Papers in Press. Published on November 28, 2000 as Manuscript M J.M. Sandra Healy, Margit Menges, John H. Doonan* and James A.H. Murray.
|
|
|
- Kerrie Cross
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on November 28, 2000 as Manuscript M The Arabidopsis D-type cyclins CycD2 and CycD3 both interact in vivo with the PSTAIRE cyclin-dependent kinase Cdc2a but are differentially controlled. J.M. Sandra Healy, Margit Menges, John H. Doonan* and James A.H. Murray. Institute of Biotechnology, University of Cambridge, Cambridge CB2 1QT, United Kingdom *Dept. of Cell Biology, John Innes Centre, Colney Lane, Norwich NR4 7UH Present address: Dipartimento di Genetica, IV Piano, Torre A, Università di Milano, Milan, Italy. To whom correspondence should be addressed: Institute of Biotechnology, University of Cambridge, Tennis Court Road, Cambridge CB2 1QT, United Kingdom. Tel.: Fax: Running Title: Characterization of Arabidopsis CycD2 and CycD3 proteins This work was financially supported by BBSRC grants G02552 and P09509, a BBSRC studentship to MM and Aventis CropScience. Copyright 2000 by The American Society for Biochemistry and Molecular Biology, Inc. 1
2 SUMMARY D-type cyclins (CycD) play key roles in linking the Arabidopsis cell cycle to extracellular and developmental signals, but little is known of their regulation at the post-transcriptional level or of their cyclin-dependent kinase (CDK) partners. Using new antisera to CycD2 and CycD3, we demonstrate that the CDK partner of these Arabidopsis cyclins is the PSTAIREcontaining CDK Cdc2a. Previous analysis has shown that transcript levels of CycD2 and CycD3 are regulated in response to sucrose levels, and that both their mrna levels and kinase activity are induced with different kinetics during G1 phase of cells re-entering the division cycle from quiescence (1). Here we analyse the protein levels and kinase activity of CycD2 and CycD3. We show that CycD3 protein and kinase activity parallel the abundance of its mrna, and that CycD3 protein is rapidly lost from cells in stationary phase or following sucrose removal. In contrast to both CycD3 and the regulation of its own mrna levels, CycD2 protein is present at constant levels. CycD2 kinase activity is regulated by sequestration of CycD2 protein in a form inaccessible to immunoprecipitation and probably not complexed to Cdc2a. 2
3 INTRODUCTION The cell cycle in eukaryotic organisms is primarily controlled by cyclin-dependent kinases (CDKs) in complexes with their activating and substrate-specifying partners, cyclins. The action of these protein complexes is perhaps best understood in yeast, in which a single CDK interacts with different cyclins during the G1 and G2/ mitotic phases of the cell cycle (2). In comparison to yeast, relatively little is known about the control of the plant cell cycle, but complementation of yeast strains lacking G1 cyclins has been used to identify the functionally equivalent proteins in plants (3, 4). These approaches identified plant D-type (CycD) cyclins, as less related to the yeast G1 cyclins than to mammalian and Drosophila D-type cyclins which function during G1 phase to control cell cycle commitment in response to growth and mitogenic signals (5, 6). CycD cyclins have low overall similarity with animal D-type cyclins (7), but share key features, including an L-x-C-x-E motif at the N-terminus which in both animal and plant D-type cyclins is the binding site for the retinoblastoma (Rb) protein (8-10). The interaction between animal D-type cyclins and Rb results in phosphorylation of Rb by the cyclin D-CDK complexes, and the resultant phosphorylated Rb is unable to bind (and therefore inhibit) E2F transcription factors involved in the G1/S transition and progression through S phase (reviewed 11, 12). The transcription of the mammalian D-type cyclins is dependent on stimulation by serum growth factors, and thus D-type cyclins act as mediators of external signals in the progression of the cell cycle (5, 6). Higher organisms express multiple CDKs, which bind distinct cyclins, and different CDKs are therefore characterized by sequence differences in their cyclin-interaction domain. In animals, mitotic cyclins (A- and B-type) bind CDKs that contain the conserved cyclinbinding sequence PSTAIRE, whereas D-type cyclins bind CDK4 and 6, which have the sequence PI/LSTV/IRE (reviewed 13). Plants express two main types of cell cycle regulating CDKs: those containing the PSTAIRE sequence (CDK-a or Cdc2a proteins), and a novel CDK class with the variant sequence PPTA/TLRE which shows cell cycle regulation 3
4 (reviewed 14, 15). Little is known of cyclin-cdk partnerships in plants, although an alfalfa A-type cyclin CycA2 has recently been shown to associate with the PSTAIRE-containing Cdc2a (16), as has a tobacco CycD3 (17). The isolation of plant Rb and E2F-like proteins and their interaction in vitro therefore suggests overall similarities between G1/S control in animals and plants (17-21, de Jager et al. submitted). Evidence that an in vitro assembled complex of tobacco CycD3 and the PSTAIRE CDK Cdc2a can phosphorylate a tobacco Rb-related protein also substantiates the broad parallels between plant and mammalian systems (17). In addition, mrna levels of the Arabidopsis cyclin D2 (CycD2) and cyclin D3 (CycD3) genes are controlled by external growth signals. In an Arabidopsis cell culture, CycD2 and CycD3 mrna levels are induced by sucrose and CycD3 mrna levels increase in response to the presence of the plant hormone cytokinin (22, 1). CycD1 expression is not detectable in this culture. Overexpression of the Arabidopsis CycD2 gene in tobacco has also been shown to increase overall plant growth rate (23), and high level expression of CycD3 results in extopic cell divisions and altered growth (22). Taken together these results suggests that plant CycD levels may serve to integrate such signals with commitment of cells to division, paralleling the role of D-type cyclins of animals. Despite the significance of CycD activity for cell cycle control, development and growth, almost nothing is known of the control of protein levels or kinase activity of CycD kinases in plants. This is particularly significant, since post-transcriptional and posttranslational controls are of considerable importance in determining CDK-cyclin activity (24). Moreover, the CDK partner of the Arabidopsis CycDs is unknown. Here we utilise antibodies that specifically detect CycD2 and CycD3 proteins of Arabidopsis to analyse CycD protein levels in an Arabidopsis cell culture and during plant growth, to identify the CDK partner of these cyclins, and to analyse the kinase activity and associations of the cyclin-cdk complexes in response to external growth signals. 4
5 MATERIALS AND METHODS Arabidopsis cell suspension and plant growth Suspension cultures of Arabidopsis thaliana ecotype Landsberg erecta (25, 26) were maintained as previously described (1). For cell cycle re-entry experiments, quiescent cells were prepared from cultures 7 days (7d) after previous subculture (7 d cultures); the cells were washed and sub-cultured into MS medium with hormones and all supplements except sucrose. After 24 hours (h), sucrose was added to the culture and time point samples taken as the cells re-entered the cell cycle (1, 22). Similarly for cell cycle exit experiments, 3 d (exponential culture growth phase) cells were harvested and replaced in MS medium containing hormones and all supplements except sucrose, and samples taken at specified times. Arabidopsis seedlings (Landsberg erecta) were grown in liquid medium as previously described (1). Plants were grown to flowering at 22 C in 16 h light in a growth room. Protein methods - Polyclonal rabbit antisera were raised against full length Arabidopsis CycD2 expressed with a 6x-histidine tag in Escherichia coli, and against Arabidopsis CycD3 C-terminal peptide MRGAEENEKKKPILHLPWAIVATP by the antibody facility at the Babraham Institute. Antisera against a common C-terminal peptide found in Arabidopsis Cdc2b and in its tobacco homologue CdkB1 has been described (Sorrell et al., submitted), and antisera against Arabidopsis Cdc2a was produced using the C-terminal peptide ARAALEHEYFKDLGGMP at the Babraham Institute. Procedures for protein extraction, SDS-polyacrylamide gel electrophoresis, western blot analysis, immunoprecipitations and histone H1 protein kinase assays have been described (1, 23). Antiserum was used at 1/1000 dilution and incubated with western blots overnight at room temperature. For competition of the antisera with CycD3 peptide or CycD2 protein, the appropriate antiserum was incubated 5
6 with CycD3 peptide (at a concentration of 1µM) or CycD2 protein (40 µg of E. coli expressed CycD2) for 1 h at 37ºC before probing the western blot. In Vitro Translation - CycD2 and CycD3 cdnas were cloned into pet23 (Novagen, Madison, WI) and proteins were produced using a T7 coupled transcription-translation rabbit reticulocyte lysate expression system with incorporation of [ 35 S]methionine (Promega, Madison, WI). 6
7 RESULTS Specific antisera which immunodetect and immunopreciptate Arabidopsis CycD2 and CycD3 in active kinase complexes For analysis of Arabidopsis CycD cyclin proteins, rabbit antibodies were generated against full length CycD2 protein and against a CycD3 C-terminal peptide. To verify that these antisera specifically detect the intended proteins, they were used to probe western blots of Arabidopsis whole cell extract (WCE) and in vitro translated (IVT) radiolabelled CycD2 and CycD3; the antibody probes were used both directly and in competition with the relevant purified CycD2 protein or CycD3 peptide (Figure 1A). The CycD2 antibody detected IVT CycD2 protein of approximately 46 kda and a protein of the same size in WCE. Neither were detected when the antibody was pre-incubated with purified CycD2 protein. Similarly the CycD3 antibody detected IVT CycD3 and a protein of the same size (approximately 60 KDa) from the WCE, and these bands were not detected after pre-incubation of the antibody with purified CycD3 peptide. No cross-reactivity was observed between the CycD2 and CycD3 antisera (not shown). These results were confirmed by immunoblots of plants overexpressing CycD2 or CycD3, in which a stronger signal was observed (not shown). The quantitative immunoprecipitation of CycD2 and CycD3 by these antibodies was demonstrated by the recovery of IVT CycD2 and CycD3 (Figure 1B). Native CycD2 and CycD3 were then immunopreciptitated from WCE of an exponential phase culture. The resultant immunoprecipitates were assayed for histone H1 kinase activity to verify the presence of active CycD-CDK complexes. Both CycD2 and CycD3 immunoprecipitates phosphorylate histone H1 (Figure 1C). The phosphorylation of two to four other proteins of kda present in WCE was also observed, and these may represent endogenous substrates that have co-immunopreciptitated with CycD2 and CycD3. In addition, a protein of 7
8 around 60 kda is phosphorylated in CycD3 immunoprecipitates, which may be a result of autophosphorylation of CycD3 protein (27, 28) Insert Figure 1 Arabidopsis CycD2 and CycD3 proteins interact with the PSTAIRE CDK Cdc2a In mammalian and Drosophila cells, D-type cyclins interact with the non-pstaire CDK4 and (in mammals) CDK6. However, no direct plant homologues of these CDKs have been identified, although plants contain two main groups of CDKs (14, 15). These are represented by the Arabidopsis PSTAIRE CDK Cdc2a and the PPTALRE CDK Cdc2b. To establish the CDK partners of Arabidopsis CycD2 and CycD3, the cyclins were immunoprecipitated from exponentially growing cells and eluants were analysed by protein gel blotting using antisera raised against Arabidopsis Cdc2a and Cdc2b (Figure 2). Cdc2a was detected in eluants from both CycD2 and CycD3 immunoprepitates, but Cdc2b did not co-immunoprecipitate with either cyclin. We conclude that Cdc2a is an in vivo partner of both CycD2 and CycD3 in Arabidopsis cells, but Cdc2b is not. Insert Figure 2 CycD2 and CycD3 proteins accumulate differentially during Arabidopsis development CycD proteins likely play key roles in activating cell division during development, so the presence of CycD2 and CycD3 was examined during Arabidopsis vegetative development (Figure 3A). CycD2 levels are lower in 4 d seedlings but are subsequently present at a constant level in whole seedling/plant extracts (5-17 d), whereas CycD3 is present at similar levels from 4 d to 14 d, but subsequently declines at 17 d. In contrast, the kinase partner Cdc2a is present at a constant level in these protein extracts. The distribution of the protein between roots and shoots was examined in 9 d seedlings. CycD2 is primarily expressed in the 8
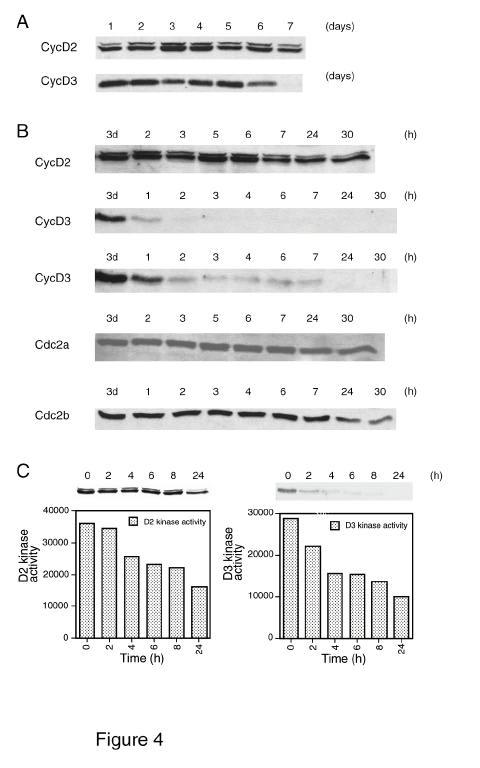
9 shoot although a weak signal was detected in root extract (Figure 3B), while CycD3 was detected less readily than D2 and primarily in the root. In flowering Arabidopsis plants, CycD2 was more abundant in leaves and stem and detectable at a lower level in flowers, but appeared to be absent from root extracts. In contrast CycD3 was present only in the root, where it was detected weakly. Insert Figure 3 CycD3 but not CycD2 protein level is higher in actively dividing cells Levels of CycD2 and CycD3 were examined throughout the growth cycle from samples taken on consecutive days after sub-culture. This culture reaches maximum cell density after 6 d (1). The level of CycD2 remained fairly constant throughout the growth cycle, compared to CycD3 which decreased dramatically after 6 d, corresponding to the onset of stationary phase (Figure 4A). CycD3 is therefore present only in cells from actively dividing cultures. Insert Figure 4 Differential abundance and kinase activity of CycD2 and CycD3 proteins after removal of growth stimulation signals Previous analysis has shown the dependence of CycD2 and CycD3 gene expression on sucrose availability (1). To measure the response of CycD2 and CycD3 protein levels and kinase activity to the removal of such growth stimulation signals, sucrose was removed from an exponentially growing (3 d) cell culture and samples taken at specific times (Figure 4B). CycD2 protein levels remained relatively constant, but CycD3 protein levels decreased rapidly with approximately 90% of CycD3 being degraded within 2 h of sucrose removal. Longer exposures of this blot show that low levels of CycD3 persist for at least 7 h, but by 24 h CycD3 is almost undetectable (Figure 4B). In contrast to CycD2 mrna, CycD2 protein 9
10 levels are constant following sucrose removal, whereas CycD3 protein rapidly declines in abundance. The abundance of Cdc2a and Cdc2b proteins after removal of sucrose remained constant for at least 24 h (Figure 4B). When the kinase activity of immunoprecipitations (IPs) was examined in a similar experiment, a somewhat different pattern was observed. In contrast to the constant abundance of CycD2 protein, CycD2-associated kinase activity started to decline within 4 h of sucrose removal, and by 24 h was reduced by more than 50%. CycD3 kinase activity decreased to 50% of its original level within 4 h and then declined more gradually up to 24 h. However, although CycD3 protein was almost undetectable after 24 h, there was still one third of the kinase activity observed in the exponentially growing cells. This may suggest that much of the early decline in CycD3 levels may be due to a reduction of cyclin not associated with active kinase complexes. Differential accumulation and activity of CycD2 and CycD3 in cells re-entering the division cycle Arabidopsis cells deprived of sucrose for 24 h resume division and show a relatively synchronous entry into S phase when sucrose is re-added (1, 22). CycD2 and CycD3 mrna levels decline after sucrose removal, and increase after re-addition. CycD2 mrna increases within 1 h of sucrose addition, whereas CycD3 mrna accumulates only after 4 h, corresponding to late G1 phase. To examine the abundance of CycD2 and CycD3 proteins and their associated kinase activity as cells re-enter the cell cycle, early stationary phase (7 d) cells were prepared by removing sucrose from the growth medium for 24 h and then the levels of D-type cyclins (Figure 5A) and kinase activity of their IPs (Figure 5B) were observed after sucrose was re-added. CycD2 was present at relatively high levels in stationary phase cells before (data not shown) and after sucrose removal. CycD3 was present at a low level before 10
11 (data not shown) sucrose removal, and absent after 24 h without sucrose, consistent with results observed after sucrose removal from an exponentially growing cell culture (Figure 4B). Figure 5A shows that in samples taken every hour after sucrose re-addition, CycD2 remains at a constant level. During this time cells progress through G1, reaching S phase after 6 h (1, 22). CycD3 protein was absent in early G1 cells, but started to accumulate rapidly at 4-5 h in late G1, co-incident with the increase in CycD3 mrna (1, 22). The abundance of Cdc2a, the CDK partner of CycD2 and CycD3, was constant during this experiment. The kinase activities of CycD2 and CycD3 were examined in a separate experiment (Figure 5B), and showed that despite the constant protein abundance of CycD2, its associated kinase activity is strongly regulated, being very low after sucrose removal, and increasing within 2 h of sucrose re-addition, as previously reported (1). CycD2 kinase activity continues to increase up to 24 h after sucrose re-addition, despite no change in protein abundance (Figure 5B). In contrast, CycD3 kinase activity starts to increase only after 4 h, and largely reflects mrna levels and protein abundance during G1/S phase, although a further increase in kinase activity is seen in cells 24 h after sucrose addition (Figure 5B). We conclude that CycD2 is subject to strong post-translational regulation. Insert Figure 5 CycD2 does not interact with Cdc2a in quiescent cells The experiments described above show that CycD2 protein abundance remains constant in actively dividing and non-dividing Arabidopsis suspension cells, although CycD2-associated kinase activity is only found in actively dividing cells. To analyse the nature of this posttranslational regulation, we first examined whether the CDK partner is present. Both CycD2 and Cdc2a were found in quiescent cells (prepared as for the cell cycle re-entry experiment) as well as exponentially growing cells, although Cdc2a levels were lower than in 11
12 exponentially growing cells (Figure 6A). However, the differences in the kinase activity of the CycD2-Cdc2a complex were marked between the two growth phases, and almost no activity was present in quiescent cells (Figure 6A). To investigate whether the lack of kinase activity in quiescent cells was due to the inability of CycD2 and Cdc2a to interact, CycD2 was immunoprecipitated from both quiescent and exponentially growing cells to identify if similar amounts of Cdc2a were coimmunoprecipitated. The amount of Cdc2a co-immunoprecipitated in quiescent cell extract was much lower (Figure 6B, c), suggesting that the interaction between Cdc2a and CycD2 was inhibited in the quiescent state. However, re-probing these blots with CycD2 antibody (Figure 6B, g) showed that the quantity of CycD2 protein immunoprecipitated from quiescent cell extract was much lower than from exponential cell extract even though CycD2 was equally abundant in quiescent cells (Figure 6A). Thus anti-cycd2 antiserum efficiently immunoprecipitates CycD2 from exponential cell extract, but not from quiescent cells; a result that was observed in several repeated experiments. From this experiment it is therefore not possible to determine whether CycD2 and Cdc2a are associated in quiescent cells, since CycD2 is not immunoprecipitated. To determine if this inefficient immunoprecipitation is a feature of quiescent cell extract or particular to CycD2, Cdc2a was immunoprecipitated directly from quiescent and exponentially growing cells, and the eluates were immunoblotted with the same antiserum (Figure 6C, c and d). Cdc2a was immunoprecipitated in proportion to its abundance (Figure 6A) from cells in both phases of growth, suggesting that the inefficient immunoprecipitation of CycD2 in quiescent cells was a behaviour specific to CycD2. The eluted Cdc2a immunoprecipitates were immunoblotted with anti-cycd2 antiserum (Figure 6C, a and b). CycD2 was efficiently co-immunoprecipitated by the anti-cdc2a antiserum from exponential cells (Figure 6C, a), but not from quiescent cells (Figure 6C, b). This demonstrates that the 12
13 majority of CycD2 present in quiescent cells is not associated with Cdc2a, nor as we show in Figure 6B is it accessible for immunoprecipitation by the CycD2 antiserum. We also noted that the small amount of CycD2 immunoprecipitated from quiescent cells by the CycD2 antiserum (Figure 6B, g) and the amount of CycD2 coimmunoprecipitated by the Cdc2a antiserum (Figure 6C, b), is a similar proportion of the equivalent immunoprecipitations from exponential cells (compare Figure 6B, g with Figure 6B, f and Figure 6C, b with Figure 6C, a). Moreover the ratio between the amount of Cdc2a co-immunoprecipitated by the CycD2 antiserum from quiescent and dividing cells (Figure 6B, c and Figure 6B, b), is similar to the ratio of the CycD2 directly immunoprecipitated from the same extracts (Figure 6B, g and f). Taken together, this suggests that the CycD2 that was immunoprecipitated was interacting with Cdc2a, and therefore that there is a correlation between the availability of CycD2 for immunoprecipitation and its ability to associate with Cdc2a. Insert Figure 6 13
14 DISCUSSION Plant D-type (CycD) cyclins play important roles in controlling the cell cycle in development and in response to external signals (1, 3, 4, 22, 23, 29). However, despite the importance of post-transcriptional mechanisms in regulating cyclin-cdk activity (24, 30), previous studies have either examined the consequences of CycD mis-expression (22, 23), the developmental regulation of their gene expression or the response of mrna levels to external signals (1, 3, 4, 22, 29). Here we show that the development of specific antisera to CycD2 and CycD3 of Arabidopsis allows the identification of the CDK partner of these cyclins and analysis of their protein abundance and kinase activity during development and during the response to sucrose removal and addition. We show that both CycD2 and CycD3 associate with the Arabidopsis PSTAIRE CDK Cdc2a, and that CycD2 and CycD3 show strikingly different modes of regulation at the protein level. Tobacco CycD3 has previously been shown to associate with a CDK of the PSTAIRE type in vitro and in BY-2 cell extract (17), and Arabidopsis CycD1 was identified in a yeast two-hybrid screen using Cdc2a as a bait (31). Moreover, Arabidopsis CycD2 expressed in tobacco forms functional kinase complexes with the PSTAIRE tobacco CDK Cdc2a (23). Here we present the first evidence that endogenous Arabidopsis CycD2 and CycD3 interact with and form functional kinase complexes with Cdc2a in vivo, whereas these cyclins do not interact with the non-pstaire CDK Cdc2b. These results are in contrast to mammalian and Drosophila D-type cyclins, which do not form functional complexes with PSTAIRE CDKs but rather associate with the non-pstaire-containing CDK4 and CDK6 (2, 32). Immunoprecipitates of CycD2 and CycD3 were found to exhibit in vitro protein kinase activity against added histone H1 as previously reported (1, 23) and against coimmunoprecipitated proteins from the Arabidopsis cell extract. The identity of these proteins 14
15 is currently unknown, but since CycD2 and CycD3 interact with maize Rb in vitro and in yeast two-hybrid assays (10), one candidate is the Arabidopsis homologue of the Rb protein. The suitability of histone H1 as a substrate for plant CycD kinases (1, 23) is further confirmed, highlighting a further difference with animal cyclin D-CDK4 complexes for which histone H1 is a poor substrate. Since Arabidopsis Rb was unpublished at the time of this work (33), we have been unable to confirm its suitability as an in vitro substrate of Arabidopsis CycD kinases, although this would be predicted from previous analysis in vitro (10, 17). Mammalian D-type cyclins are highly unstable proteins whose synthesis is linked to the presence of external growth signals (5, 6, 34-36). Previous analysis has shown that mrna levels of plant CycD respond to external signals of hormones and sucrose levels (1, 3, 22, 29, 37). Arabidopsis CycD2 and CycD3 mrna levels decline on sucrose removal and are induced on its re-addition, although the magnitude of the CycD3 response is twice that of CycD2. Here we examined the protein levels and kinase activity of CycD2 and CycD3, and found strikingly different results. When sucrose starved cells were induced to re-enter the cell cycle, CycD3 protein was detectable 4-5 h after re-addition of sucrose, at the same time as CycD3 mrna and kinase activity are induced (1). Since we found that Cdc2a is present throughout the sucrose starvation, this suggests that CycD3 kinase activity is regulated by CycD3 mrna abundance during this experiment. The importance of transcriptional regulation of CycD3 activity is supported by the strong phenotypes produced in vivo from its overexpression (22). When sucrose was removed from cells in a mid-exponential phase culture, the majority of CycD3 was no longer present after 1-2 h, suggesting that in this situation it is rapidly turned over. A residual level of CycD3 persists for several hours, and the reduction in kinase activity is less abrupt than the loss of CycD3 protein, suggesting that CycD3 not present in active kinase complexes is turned over more rapidly. In this regard it is interesting 15
16 to note the low amount of Cdc2a co-immunoprecipitated with CycD3 (Figure 2). We note that the response of CycD3 to sucrose removal appears to be an immediate and specific response to loss of this signal, since the levels of other cell cycle proteins remain constant for 24 h after sucrose removal, for example the kinases Cdc2a and Cdc2b (Figure 4B), even though Cdc2b abundance is cell cycle regulated and the protein is only present from S-M phases (14). We conclude that changes in CycD3 level are an immediate response to sucrose removal and not an indirect consequence of a cessation of cell division caused by declining intracellular carbohydrate levels. CycD2 regulation is strikingly different from CycD3 and D-type cyclins in other organisms. Despite the regulation of its mrna during sucrose removal and re-supply (1), CycD2 protein remains almost constant in abundance through sucrose starvation and re-entry into the cell cycle, although its kinase activity is strongly regulated, being activated more rapidly than that of CycD3. Further investigation showed that CycD2 present in quiescent or sucrose-starved cells is not associated with Cdc2a and is not accessible for immunoprecipitation. This suggests regulation of CycD2-associated kinase activity by a novel post-translational mechanism involving its sequestration in an inactive form. It is unknown whether the reappearance of CycD2-Cdc2a kinase activity within 2 h of sucrose re-addition is a result of release and activation of existing CycD2, or the rising levels of CycD2 activity result from de novo synthesis. The increase in CycD2 mrna shortly after sucrose re-addition may be consistent with the latter explanation. We conclude that the kinase activity of CycD2- containing complexes is not dependent on the level of CycD2 protein, in contrast to the situation observed with CycD3. Based on the data presented here, we present a model for activation of cell division in Arabidopsis cells (Table 1 and Figure 7). In quiescent cells, CycD2 protein is sequestered and presumably inactive as it is not associated with its CDK partner. CycD3 protein is absent. After stimulation of division, CycD2 mrna accumulates, and CycD2 protein associates in an 16
17 active form with Cdc2a, and kinase activity starts to accumulate within 2 h, in early G1 phase. The amount of CycD2 present in this active form is unknown. After 4 h, in late G1, CycD3 transcript levels increase sharply, accompanied by an increase in CycD3 protein levels and kinase activity. This is followed by entry into S phase after approximately 6 h. Insert Table 1 and Figure 7 The work presented here and elsewhere (reviewed 14) suggests that the overall similarities between plant and mammalian controls of the cell cycle are overlaid by complex and important differences in the control, interactions and targets of cell cycle regulators in plants. Acknowledgements We thank Bart den Boer, Marc de Block and Masami Sekine for useful discussions. 17
18 REFERENCES 1. Riou-Khamlichi, C., Menges, M., Healy, J. M., and Murray, J. A. H. (2000) Mol. Cell Biol. 20, Pines, J. (1995) Adv. Cancer Res. 66, Soni, R., Carmichael, J. P., Shah, Z. H., and Murray, J. A. H. (1995) Plant Cell 7, Dahl, M., Meskiene, I., Bögre, L., Ha, D. T. C., Swoboda, I., Hubmann, R., Hirt, H., and Heberle-Bors, E. (1995) Plant Cell 7, Sherr, C. J. (1993) Cell 73, Sherr, C. J. (1994) Cell 79, Renaudin, J.-P., Doonan, J. H., Freeman, D., Hashimoto, J., Hirt, H., Inzé, D., Jacobs, T., Kouchi, H., Rouzé, P., Sauter, M., Savouré, A., Sorrell, D. A., Sundaresan, V., and Murray, J. A. H. (1996) Plant Mol. Biol. 32, Ewen, M. E., Sluss, H. K., Sherr, C. J., Matsushime, H., Kato, J., and Livingston, D. M. (1993) Cell 73, Kato, J., Matsushime, H., Hiebert, S. W., Ewen, M. E., and Sherr, C. J. (1993) Genes Dev. 7, Huntley, R., Healy, S., Freeman, D., Lavender, P., de Jager, S., Greenwood, J., Makker, J., Walker, E., Jackman, M., Xie, Q., Bannister, A. J., Kouzarides, T., Gutierrez, C., Doonan, J. H., and Murray, J. A. H. (1998) Plant Mol. Biol. 37, Sherr, C. J. (1996) Science 274,
19 12. Dyson, N. (1998) Genes Dev. 12, Morgan, D. O. (1997) Annu. Rev. Cell Dev. Biol. 13, Mironov, V., De Veylder, L., Van Montagu, M., and Inzé, D. (1999) Plant Cell 11, Huntley, R. P., and Murray, J. A. H. (1999) Curr. Opin. Plant Biol. 2, Roudier, F., Fedorova, E., Gyorgyey, J., Feher, A., Brown, S., Kondorosi, A., and Kondorosi, E. (2000) Plant J. 23, Nakagami, H., Sekine, M., Murakami, H., and Shinmyo, A. (1999) Plant J. 18, Xie, Q., Sanz-Burgos, A. P., Hannon, G. J., and Gutiérrez, C. (1996) EMBO J. 15, Grafi, G., Burnett, R. J., Helentjaris, T., Larkins, B. A., DeCaprio, J. A., Sellers, W. R., and Kaelin, W. G. (1996) Proc. Natl. Acad. Sci. USA 93, Ramirez-Parra, E., Xie, Q., Boniotti, M. B., and Gutierrez, C. (1999) Nucleic Acids Res. 27, Sekine, M., Ito, M., Uemukai, K., Maeda, Y., Nakagami, H., and Shinmyo, A. (1999) FEBS Lett. 460, Riou-Khamlichi, C., Huntley, R., Jacqmard, A., and Murray, J. A. H. (1999) Science 283, Cockcroft, C. E., den Boer, B. G., Healy, J. M., and Murray, J. A. H. (2000) Nature 405,
20 24. Lew, D. J., and Kornbluth, S. (1996) Curr. Opin. Cell Biol. 8, May, M., and Leaver, C. (1993) Plant Physiol. 103, Fuerst, R. A. U. A., Soni, R., Murray, J. A. H., and Lindsey, K. (1996) Plant Physiol. 112, Diehl, J. A., Zindy, F., and Sherr, C. J. (1997) Genes Dev. 11, Diehl, J. A., and Sherr, C. J. (1997) Mol. Cell Biol. 17, Gaudin, V., Lunness, P. A., Fobert, P. R., Towers, M., Riou-Khamlichi, C., Murray, J. A. H., Coen, E., and Doonan, J. H. (2000) Plant Physiol. 122, Sherr, C. J., and Roberts, J. M. (1995) Genes Dev. 9, Helleboid, S., Hendriks, T., Bauw, G., Inze, D., Vasseur, J., and Hilbert, J. L. (2000) J. Exp. Bot. 51, Xiong, Y., Zhang, H., and Beach, D. (1992) Cell 71, Kong, L. J., Orozco, B. M., Roe, J. L., Nagar, S., Ou, S., Feiler, H. S., Durfee, T., Miller, A. B., Gruissem, W., Robertson, D., and Hanley-Bowdoin, L. (2000) EMBO J. 19, Matsushime, H., Ewen, M. E., Strom, D. K., Kato, J.-Y., Hanks, S. K., Roussel, M. F., and Sherr, C. J. (1992) Cell 71, Sewing, A., Bürger, C., Brüsselbach, S., Schalk, C., Lucibello, F. C., and Müller, R. (1993) J. Cell Sci. 104, Baldin, V., Lukas, J., Marcote, M. J., Pagano, M., and Draetta, G. (1993) Genes Dev. 7, 20
21 De Veylder, L., de Almeida Engler, J., Burssens, S., Manevski, A., Lescure, B., Van Montagu, M., Engler, G., and Inzé, D. (1999) Planta 208,
22 FIGURE LEGENDS Figure 1 Protein gel (western) blots of CycD2 and CycD3 demonstrating specificity and sensitivity of antibodies developed against CycD2 and CycD3. (A) Whole cell extracts and IVT products of CycD2 (left) and CycD3 (right) probed with CycD2 antiserum (left) and CycD3 antiserum (right). Left half of blots were probed with relevant antibody, right half probed with the antibody after pre-competition with purified protein/peptide (comp). The central lane containing IVT CycD2/ CycD3 protein was cut in half. CycD2 antibody was competed with purified CycD2 protein; CycD3 was competed with CycD3 C terminal peptide. (B) Immunoprecipitations of CycD2 and CycD3 IVT proteins. Left to right: IVT protein, immunoprecipitations (ip) of 1 µl, 2 µl and 4 µl of IVT protein. (C) Protein kinase assays of immunoprecipitates for the CycD2- (left) and CycD3- (right) CDK complexes using histone H1 as an added substrate; the left lane of each blot contains an immunoprecipitation using pre-immune sera (PI), the right lane contains an immunoprecipitation using immune sera specifc for each D-type cyclin protein (I). Dots indicate co-immunoprecipitated bands from WCE that are phosphorylated. Free standing arrow indicates band corresponding to potential autophosphorylation of CycD3. Figure 2 The PSTAIRE CDK Cdc2a co-immunoprecipitates with CycD2 and CycD3 from Arabidopsis cell extracts, but the non-pstaire Cdc2b does not. Protein gel (western) blots of immunoprecipitations from WCE using CycD2 and CycD3 antisera were probed with Cdc2a and Cdc2b antisera. WCE = 40 µg of whole cell extract; PI, immunoprecipitation with preimmune serum; I, immunoprecipitation of protein with immune serum, ip = immunoprepitation. 22
23 Figure 3 CycD2 and CycD3 protein levels during vegetative development and in mature tissues. (A) Levels of CycD2, CycD3 and Cdc2a in seedlings/plants of 4, 5, 7, 9, 14 and 17 days (d) grown in vitro. (B) Levels of CycD2 and CycD3 in shoots and roots of 9 d seedlings. (C) Presence of CycD2 and CycD3 in leaf, stem, root and flower of mature soil grown Arabidopsis. Figure 4 Levels of CycD proteins during the growth cycle of suspension cultured cells and in response to sucrose removal. (A) Protein gel (western) blots showing levels of CycD2 and CycD3 from the lag phase (days 1-2) through exponential growth (days 3-6) to stationary phase (day 7) (1). The upper band visible in some CycD2 blots is non-specific and not competed by preincubation of the CycD2 antiserum with purified CycD2 protein (data not shown). (B) Protein gel blots showing levels of CycD2, CycD3, Cdc2a and Cdc2b in response to removal of sucrose from medium of 3 d (exponential phase) suspension cultured cells. 3d; WCE before sucrose removal; further lanes are extracts at times (in hours; h) after replacing cells in fresh medium lacking sucrose. Short (above) and longer (below) exposures of CycD3 gel blots are shown to demonstrate continued presence of low levels of CycD3 for at least 7 h. (C) Protein kinase activity (measured in arbitrary units) of CycD2 and CycD3 immunoprecipitates against histone H1 at times indicated. Background phosphorylation observed using preimmune serum for immunoprecipitation has been subtracted from the data. Corresponding protein gel (western) blots for the extracts used for kinase assays are shown above the graphs. Figure 5 23
24 CycD2 and CycD3 levels and protein kinase activity during cell cycle re-entry. Stationary phase cells were resuspended in fresh medium lacking sucrose for 24 h, and sucrose was then added. Samples were taken for protein gel analysis at the time of sucrose addition (0) and at later times indicated in h. (A) Protein gel blots of CycD2, CycD3 and Cdc2a levels. (B) In a similar experiment, protein kinase levels (measured in arbitrary units) of CycD2 and CycD3 immunoprecipitates against histone H1 were determined from time of sucrose addition (0). Background obtained using pre-immune serum has been subtracted. Corresponding protein gel blots for extracts used for kinase assays are shown above graphs. Figure 6 CycD2 is present in stationary phase cells, but is not associated with Cdc2a and is inaccessible for immunoprecipitation. (A) Upper: protein gel blots of WCE from exponentially growing (E) and quiescent (Q) cells (prepared as for the cell cycle re-entry experiments) probed with antisera against CycD2 and Cdc2a. Lower: histone H1 kinase activity of CycD2 immunoprecipitates from exponentially growing and quiescent cells. PI, histone H1 kinase assays following immunoprecipitation with pre-immune sera; I, histone H1 kinase assays following immunoprecipitation with immune sera. (B) Immunoprecipitates from WCE of exponentially growing (E) and quiescent (Q) Arabidopsis cells using preimmune (PI) or immune CycD2 (I) antisera were subject to protein gel blotting and probed with Cdc2a or CycD2 antisera. CycD2 is efficiently immunoprecipitated only from exponentially growing cell extract. (C) Immunoprecipitates from WCE of exponentially growing (E) and quiescent (Q) cells using Cdc2a antiserum were blotted and probed with CycD2 and Cdc2a antisera. Note that CycD2 only co-immunoprecipitates efficiently from E cells, whereas Cdc2a is immunoprecipitated in equal proportions to its presence in E and Q cells (panel A). 24
25 Figure 7 Summary model of CycD2 and CycD3 protein associations with Cdc2a in an Arabidopsis cell culture during the transition from quiescent cells, through cell cycle re-entry to active division. See text and Table 1 for details. CycD2 is shown complexed with an unknown protein X in quiescent (non-dividing) cells; the rising CycD2-associated kinase activity in early G1 may result from the liberation of CycD2 from these complexes, alternatively CycD2 may be synthesized de novo (not illustrated). 25
26
27
28
29
30
31
32
33 Active division Quiescent cells Sucrose removal Cell cycle reentry Late G1 CycD2 mrna a +++ protein association CycD2-Cdc2a X-CycD2-X c X-CycD2-X c? c CycD2-Cdc2a Cdc2a Cdc2a kinase activity b ++ CycD3 mrna protein association CycD3-Cdc2a CycD3 d Cdc2a Cdc2a Cdc2a CycD3-Cdc2a kinase activity Table 1. Summary model of CycD2 and CycD3 mrna levels (1), protein abundance, association with Cdc2a and kinase activity levels in an Arabidopsis cell culture during the transition from exponential growth to stationary phase, and during re-entry into the cell cycle. See text for details. a ; increases within 1 h; b increases within 2 h of sucrose addition. c CycD2 is shown complexed in non-dividing cells with unknown protein X. Rising CycD2- associated kinase activity in early G1 may result from liberation of CycD2 from these complexes, or from de novo synthesized CycD2. d Note that CycD3 not complexed to Cdc2a may be present in exponentially growing cells, suggesting possible regulation at the level of complex assembly.
The Arabidopsis D-type Cyclins CycD2 and CycD3 Both Interact in Vivo with the PSTAIRE Cyclin-dependent Kinase Cdc2a but Are Differentially Controlled*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 10, Issue of March 9, pp. 7041 7047, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Arabidopsis D-type
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 10, Issue of March 9, pp. 7041 7047, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Arabidopsis D-type
Cell Cycle Regulation of Cyclin-Dependent Kinases in Tobacco Cultivar Bright Yellow-2 Cells 1
 Cell Cycle Regulation of Cyclin-Dependent Kinases in Tobacco Cultivar Bright Yellow-2 Cells 1 David A. Sorrell 2, Margit Menges, J.M. Sandra Healy 3, Yves Deveaux, Chinatsu Amano, Ya Su 4, Hirofumi Nakagami,
Cell Cycle Regulation of Cyclin-Dependent Kinases in Tobacco Cultivar Bright Yellow-2 Cells 1 David A. Sorrell 2, Margit Menges, J.M. Sandra Healy 3, Yves Deveaux, Chinatsu Amano, Ya Su 4, Hirofumi Nakagami,
Nature Structural & Molecular Biology: doi: /nsmb.1583
 Acetylation by GCN5 regulates CDC6 phosphorylation in the S-phase of the cell cycle Roberta Paolinelli 1,2, Ramiro Mendoza-Maldonado 2, Anna Cereseto 1 and Mauro Giacca 2 1 Molecular Biology Laboratory,
Acetylation by GCN5 regulates CDC6 phosphorylation in the S-phase of the cell cycle Roberta Paolinelli 1,2, Ramiro Mendoza-Maldonado 2, Anna Cereseto 1 and Mauro Giacca 2 1 Molecular Biology Laboratory,
Distinct Cyclin D Genes Show Mitotic Accumulation or Constant Levels of Transcripts in Tobacco Bright Yellow-2 Cells 1
 Plant Physiology, January 1999, Vol. 119, pp. 343 351, www.plantphysiol.org 1999 American Society of Plant Physiologists Distinct Cyclin D Genes Show Mitotic Accumulation or Constant Levels of Transcripts
Plant Physiology, January 1999, Vol. 119, pp. 343 351, www.plantphysiol.org 1999 American Society of Plant Physiologists Distinct Cyclin D Genes Show Mitotic Accumulation or Constant Levels of Transcripts
Phosphorylation of Retinoblastoma-Related Protein by the Cyclin D/Cyclin-Dependent Kinase Complex Is Activated at the G1/S-Phase Transition in Tobacco
 The Plant Cell, Vol. 14, 1847 1857, August 2002, www.plantcell.org 2002 American Society of Plant Biologists Phosphorylation of Retinoblastoma-Related Protein by the Cyclin D/Cyclin-Dependent Kinase Complex
The Plant Cell, Vol. 14, 1847 1857, August 2002, www.plantcell.org 2002 American Society of Plant Biologists Phosphorylation of Retinoblastoma-Related Protein by the Cyclin D/Cyclin-Dependent Kinase Complex
Supplementary data. sienigma. F-Enigma F-EnigmaSM. a-p53
 Supplementary data Supplemental Figure 1 A sienigma #2 sienigma sicontrol a-enigma - + ++ - - - - - - + ++ - - - - - - ++ B sienigma F-Enigma F-EnigmaSM a-flag HLK3 cells - - - + ++ + ++ - + - + + - -
Supplementary data Supplemental Figure 1 A sienigma #2 sienigma sicontrol a-enigma - + ++ - - - - - - + ++ - - - - - - ++ B sienigma F-Enigma F-EnigmaSM a-flag HLK3 cells - - - + ++ + ++ - + - + + - -
D-Type Cyclin Gene Expression
 REFERENCES CONTENT ALERTS Sugar Control of the Plant Cell Cycle: Differential Regulation of Arabidopsis D-Type Cyclin Gene Expression Catherine Riou-Khamlichi, Margit Menges, J. M. Sandra Healy and James
REFERENCES CONTENT ALERTS Sugar Control of the Plant Cell Cycle: Differential Regulation of Arabidopsis D-Type Cyclin Gene Expression Catherine Riou-Khamlichi, Margit Menges, J. M. Sandra Healy and James
Olga A. Koroleva, a Matthew Tomlinson, a Piyarat Parinyapong, b Lali Sakvarelidze, a David Leader, c,1 Peter Shaw, a and John H.
 This article is published in The Plant Cell Online, The Plant Cell Preview Section, which publishes manuscripts accepted for publication after they have been edited and the authors have corrected proofs,
This article is published in The Plant Cell Online, The Plant Cell Preview Section, which publishes manuscripts accepted for publication after they have been edited and the authors have corrected proofs,
Supplemental Online Material. The mouse embryonic fibroblast cell line #10 derived from β-arrestin1 -/- -β-arrestin2 -/-
 #1074683s 1 Supplemental Online Material Materials and Methods Cell lines and tissue culture The mouse embryonic fibroblast cell line #10 derived from β-arrestin1 -/- -β-arrestin2 -/- knock-out animals
#1074683s 1 Supplemental Online Material Materials and Methods Cell lines and tissue culture The mouse embryonic fibroblast cell line #10 derived from β-arrestin1 -/- -β-arrestin2 -/- knock-out animals
Supplemental Data. Wu et al. (2). Plant Cell..5/tpc RGLG Hormonal treatment H2O B RGLG µm ABA µm ACC µm GA Time (hours) µm µm MJ µm IA
. Plant Cell..5/tpc RGLG Hormonal treatment H2O B RGLG µm ABA µm ACC µm GA Time (hours) µm µm MJ µm IA") Supplemental Data. Wu et al. (2). Plant Cell..5/tpc..4. A B Supplemental Figure. Immunoblot analysis verifies the expression of the AD-PP2C and BD-RGLG proteins in the Y2H assay. Total proteins were extracted
Supplemental Data. Wu et al. (2). Plant Cell..5/tpc..4. A B Supplemental Figure. Immunoblot analysis verifies the expression of the AD-PP2C and BD-RGLG proteins in the Y2H assay. Total proteins were extracted
Tobacco RETINOBLASTOMA-RELATED protein is phosphorylated by different types of cyclin-dependent kinases during the cell cycle
 Plant Biotechnology 23, 467 473 (2006) Original Paper Tobacco RETINOBLASTOMA-RELATED protein is phosphorylated by different types of cyclin-dependent kinases during the cell cycle Kazue Kawamura 1, Ko
Plant Biotechnology 23, 467 473 (2006) Original Paper Tobacco RETINOBLASTOMA-RELATED protein is phosphorylated by different types of cyclin-dependent kinases during the cell cycle Kazue Kawamura 1, Ko
Supplemental Data. Cui et al. (2012). Plant Cell /tpc a b c d. Stem UBC32 ACTIN
. Plant Cell /tpc a b c d. Stem UBC32 ACTIN") A Root Stem Leaf Flower Silique Senescence leaf B a b c d UBC32 ACTIN C * Supplemental Figure 1. Expression Pattern and Protein Sequence of UBC32 Homologues in Yeast, Human, and Arabidopsis. (A) Expression
A Root Stem Leaf Flower Silique Senescence leaf B a b c d UBC32 ACTIN C * Supplemental Figure 1. Expression Pattern and Protein Sequence of UBC32 Homologues in Yeast, Human, and Arabidopsis. (A) Expression
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Thompson et al., http://www.jcb.org/cgi/content/full/jcb.200909067/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Modification-specific antibodies do not detect unmodified
Supplemental material Thompson et al., http://www.jcb.org/cgi/content/full/jcb.200909067/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Modification-specific antibodies do not detect unmodified
WesternMAX Alkaline Phosphatase Chemiluminescent Detection Kits
 WesternMAX Alkaline Phosphatase Chemiluminescent Detection Kits Code N221-KIT N220-KIT Description WesternMAX Chemiluminescent AP Kit, Anti-Mouse Includes: Alkaline Phosphatase (AP) Conjugated Anti-Mouse
WesternMAX Alkaline Phosphatase Chemiluminescent Detection Kits Code N221-KIT N220-KIT Description WesternMAX Chemiluminescent AP Kit, Anti-Mouse Includes: Alkaline Phosphatase (AP) Conjugated Anti-Mouse
Hossain_Supplemental Figure 1
 Hossain_Supplemental Figure 1 GFP-PACT GFP-PACT Motif I GFP-PACT Motif II A. MG132 (1µM) GFP Tubulin GFP-PACT Pericentrin GFP-PACT GFP-PACT Pericentrin Fig. S1. Expression and localization of Orc1 PACT
Hossain_Supplemental Figure 1 GFP-PACT GFP-PACT Motif I GFP-PACT Motif II A. MG132 (1µM) GFP Tubulin GFP-PACT Pericentrin GFP-PACT GFP-PACT Pericentrin Fig. S1. Expression and localization of Orc1 PACT
FIGURE LEGENDS, ONLINE SUPPLEMENT
 Online Supplement: Sierra O and Towler DA: Runx2 Transactivation Mediated by MINT, the Msx2- Interacting Nuclear Target, Requires Homeodomain Interacting Protein Kinase 3 ME-10-0029_v2 FIGURE LEGENDS,
Online Supplement: Sierra O and Towler DA: Runx2 Transactivation Mediated by MINT, the Msx2- Interacting Nuclear Target, Requires Homeodomain Interacting Protein Kinase 3 ME-10-0029_v2 FIGURE LEGENDS,
Supplementary methods Shoc2 In Vitro Ubiquitination Assay
 Supplementary methods Shoc2 In Vitro Ubiquitination Assay 35 S-labelled Shoc2 was prepared using a TNT quick Coupled transcription/ translation System (Promega) as recommended by manufacturer. For the
Supplementary methods Shoc2 In Vitro Ubiquitination Assay 35 S-labelled Shoc2 was prepared using a TNT quick Coupled transcription/ translation System (Promega) as recommended by manufacturer. For the
APA105Hu01 100µg Active Nerve Growth Factor (NGF) Organism Species: Homo sapiens (Human) Instruction manual
 Organism Species: Homo sapiens (Human) Instruction manual") APA105Hu01 100µg Active Nerve Growth Factor (NGF) Organism Species: Homo sapiens (Human) Instruction manual FOR RESEARCH USE ONLY NOT FOR USE IN CLINICAL DIAGNOSTIC PROCEDURES [ PROPERTIES ] 1th Edition
APA105Hu01 100µg Active Nerve Growth Factor (NGF) Organism Species: Homo sapiens (Human) Instruction manual FOR RESEARCH USE ONLY NOT FOR USE IN CLINICAL DIAGNOSTIC PROCEDURES [ PROPERTIES ] 1th Edition
Supplementary material for: Materials and Methods:
 Supplementary material for: Iron-responsive degradation of iron regulatory protein 1 does not require the Fe-S cluster: S.L. Clarke, et al. Materials and Methods: Fe-S Cluster Reconstitution: Cells treated
Supplementary material for: Iron-responsive degradation of iron regulatory protein 1 does not require the Fe-S cluster: S.L. Clarke, et al. Materials and Methods: Fe-S Cluster Reconstitution: Cells treated
7.06 Problem Set #3, 2006
 7.06 Problem Set #3, 2006 1. You are studying the EGF/Ras/MAPK pathway in cultured cells. When the pathway is activated, cells are signaled to proliferate. You generate various mutants described below.
7.06 Problem Set #3, 2006 1. You are studying the EGF/Ras/MAPK pathway in cultured cells. When the pathway is activated, cells are signaled to proliferate. You generate various mutants described below.
There are four phases: M = mitosis is when chromosomes become condensed and separate and the cell divides: visible changes. ~ 1 hr.
 The cell cycle in eukaryotes Jason Kahn, Biochem 465 Spring 2006 (Mostly from Alberts et al., Molecular Biology of the Cell, 2002). The book is available for free on line at http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4
The cell cycle in eukaryotes Jason Kahn, Biochem 465 Spring 2006 (Mostly from Alberts et al., Molecular Biology of the Cell, 2002). The book is available for free on line at http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/5/244/ra72/dc1 Supplementary Materials for An Interaction Between BZR1 and DELLAs Mediates Direct Signaling Crosstalk Between Brassinosteroids and Gibberellins
www.sciencesignaling.org/cgi/content/full/5/244/ra72/dc1 Supplementary Materials for An Interaction Between BZR1 and DELLAs Mediates Direct Signaling Crosstalk Between Brassinosteroids and Gibberellins
Supplemental Figure 1 HDA18 has an HDAC domain and therefore has concentration dependent and TSA inhibited histone deacetylase activity.
 Supplemental Figure 1 HDA18 has an HDAC domain and therefore has concentration dependent and TSA inhibited histone deacetylase activity. (A) Amino acid alignment of HDA5, HDA15 and HDA18. The blue line
Supplemental Figure 1 HDA18 has an HDAC domain and therefore has concentration dependent and TSA inhibited histone deacetylase activity. (A) Amino acid alignment of HDA5, HDA15 and HDA18. The blue line
Motivation From Protein to Gene
 MOLECULAR BIOLOGY 2003-4 Topic B Recombinant DNA -principles and tools Construct a library - what for, how Major techniques +principles Bioinformatics - in brief Chapter 7 (MCB) 1 Motivation From Protein
MOLECULAR BIOLOGY 2003-4 Topic B Recombinant DNA -principles and tools Construct a library - what for, how Major techniques +principles Bioinformatics - in brief Chapter 7 (MCB) 1 Motivation From Protein
Mechanisms of embryonic stem cell division and differentiation
 Mechanisms of embryonic stem cell division and differentiation Josephine Wbite.'.... Department of Molecular Biosciences (Biochemistry) The University of Adelaide Adelaide, South Australia Submitted for
Mechanisms of embryonic stem cell division and differentiation Josephine Wbite.'.... Department of Molecular Biosciences (Biochemistry) The University of Adelaide Adelaide, South Australia Submitted for
supplementary information
 DOI: 10.1038/ncb2116 Figure S1 CDK phosphorylation of EZH2 in cells. (a) Comparison of candidate CDK phosphorylation sites on EZH2 with known CDK substrates by multiple sequence alignments. (b) CDK1 and
DOI: 10.1038/ncb2116 Figure S1 CDK phosphorylation of EZH2 in cells. (a) Comparison of candidate CDK phosphorylation sites on EZH2 with known CDK substrates by multiple sequence alignments. (b) CDK1 and
Supplementary Figure S1. Immunodetection of full-length XA21 and the XA21 C-terminal cleavage product.
 Supplementary Information Supplementary Figure S1. Immunodetection of full-length XA21 and the XA21 C-terminal cleavage product. Total protein extracted from Kitaake wild type and rice plants carrying
Supplementary Information Supplementary Figure S1. Immunodetection of full-length XA21 and the XA21 C-terminal cleavage product. Total protein extracted from Kitaake wild type and rice plants carrying
Notes to accompany the slidecast on theory of SDS PAGE and Western blotting
 S317 Biological science: from genes to species Notes to accompany the slidecast on theory of SDS PAGE and Western blotting SDS PAGE SDS PAGE is a standard technique for determining the molecular size of
S317 Biological science: from genes to species Notes to accompany the slidecast on theory of SDS PAGE and Western blotting SDS PAGE SDS PAGE is a standard technique for determining the molecular size of
Supplemental Materials and Methods
 Supplemental Materials and Methods In situ hybridization In situ hybridization analysis of HFE2 and genin mrna in rat liver tissues was performed as previously described (1). Briefly, the digoxigenin-labeled
Supplemental Materials and Methods In situ hybridization In situ hybridization analysis of HFE2 and genin mrna in rat liver tissues was performed as previously described (1). Briefly, the digoxigenin-labeled
Single cell imaging of Bruton's Tyrosine Kinase using an irreversible inhibitor
 SUPPLEMENTARY INFORMATION Single cell imaging of Bruton's Tyrosine Kinase using an irreversible inhibitor Anna Turetsky 1,a, Eunha Kim 1,a, Rainer H. Kohler 1, Miles A. Miller 1, Ralph Weissleder 1,2,
SUPPLEMENTARY INFORMATION Single cell imaging of Bruton's Tyrosine Kinase using an irreversible inhibitor Anna Turetsky 1,a, Eunha Kim 1,a, Rainer H. Kohler 1, Miles A. Miller 1, Ralph Weissleder 1,2,
VERIFY Tagged Antigen. Validation Data
 VERIFY Tagged Antigen Validation Data Antibody Validation Figure 1. Over-expression cell lysate for human STAT3 (NM_139276) was used to test 3 commercial antibodies. Antibody A shows strong antigen binding.
VERIFY Tagged Antigen Validation Data Antibody Validation Figure 1. Over-expression cell lysate for human STAT3 (NM_139276) was used to test 3 commercial antibodies. Antibody A shows strong antigen binding.
Supplementary Figures 1-12
 Supplementary Figures 1-12 Supplementary Figure 1. The specificity of anti-abi1 antibody. Total Proteins extracted from the wild type seedlings or abi1-3 null mutant seedlings were used for immunoblotting
Supplementary Figures 1-12 Supplementary Figure 1. The specificity of anti-abi1 antibody. Total Proteins extracted from the wild type seedlings or abi1-3 null mutant seedlings were used for immunoblotting
Supplementary information
 Supplementary information Supplementary figures Figure S1 Level of mycdet1 protein in DET1 OE-1, OE-2 and OE-3 transgenic lines. Total protein extract from wild type Col0, det1-1 mutant and DET1 OE lines
Supplementary information Supplementary figures Figure S1 Level of mycdet1 protein in DET1 OE-1, OE-2 and OE-3 transgenic lines. Total protein extract from wild type Col0, det1-1 mutant and DET1 OE lines
ONE-HOUR Western TM Multiplex Kit II
 ONE-HOUR Western TM Multiplex Kit II Technical Manual No. 0256 Version 06192009 I Description... 1 II Kit Contents.. 2 III Related Products 2 IV Key Features. 2 V Storage... 2 VI ONE-HOUR Multiplex Western
ONE-HOUR Western TM Multiplex Kit II Technical Manual No. 0256 Version 06192009 I Description... 1 II Kit Contents.. 2 III Related Products 2 IV Key Features. 2 V Storage... 2 VI ONE-HOUR Multiplex Western
SUMOylated protein capture kit
 SUMOylated protein capture kit Cat no. A010-100 Capture and detect SUMOylated proteins High capacity, high specificity SUMO binding matrix Fast, convenient protein isolation using purification system provided
SUMOylated protein capture kit Cat no. A010-100 Capture and detect SUMOylated proteins High capacity, high specificity SUMO binding matrix Fast, convenient protein isolation using purification system provided
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary figures Supplementary Figure 1: Suv39h1, but not Suv39h2, promotes HP1α sumoylation in vivo. In vivo HP1α sumoylation assay. Top: experimental scheme. Middle: we
SUPPLEMENTARY INFORMATION Supplementary figures Supplementary Figure 1: Suv39h1, but not Suv39h2, promotes HP1α sumoylation in vivo. In vivo HP1α sumoylation assay. Top: experimental scheme. Middle: we
Discussion 42 The regulatory functions of chromatin such as transcription, replication and recombination occur at two levels (von Kries et a/., 1991).
.") DISCUSSION Discussion 42 The regulatory functions of chromatin such as transcription, replication and recombination occur at two levels (von Kries et a/., 1991). The first level involves the binding of
DISCUSSION Discussion 42 The regulatory functions of chromatin such as transcription, replication and recombination occur at two levels (von Kries et a/., 1991). The first level involves the binding of
SUPPLEMENTARY INFORMATION
 Ca 2+ /calmodulin Regulates Salicylic Acid-mediated Plant Immunity Liqun Du, Gul S. Ali, Kayla A. Simons, Jingguo Hou, Tianbao Yang, A.S.N. Reddy and B. W. Poovaiah * *To whom correspondence should be
Ca 2+ /calmodulin Regulates Salicylic Acid-mediated Plant Immunity Liqun Du, Gul S. Ali, Kayla A. Simons, Jingguo Hou, Tianbao Yang, A.S.N. Reddy and B. W. Poovaiah * *To whom correspondence should be
Takeshi Sekiguchi 1,2, Eishi Noguchi 2, Toshiro Hayashida 2, Torahiko Nakashima 2, Hideo Toyoshima 1, Takeharu Nishimoto 2 and Tony Hunter 1,*
 D-type cyclin expression is decreased and p21 and p27 CDK inhibitor expression is increased when tsbn462 CCG1/TAF II 250 mutant cells arrest in G1 at the restrictive temperature Takeshi Sekiguchi 1,2,
D-type cyclin expression is decreased and p21 and p27 CDK inhibitor expression is increased when tsbn462 CCG1/TAF II 250 mutant cells arrest in G1 at the restrictive temperature Takeshi Sekiguchi 1,2,
(phosphatase tensin) domain is shown in dark gray, the FH1 domain in black, and the
 domain is shown in dark gray, the FH1 domain in black, and the") Supplemental Figure 1. Predicted Domain Organization of the AFH14 Protein. (A) Schematic representation of the predicted domain organization of AFH14. The PTEN (phosphatase tensin) domain is shown in dark
Supplemental Figure 1. Predicted Domain Organization of the AFH14 Protein. (A) Schematic representation of the predicted domain organization of AFH14. The PTEN (phosphatase tensin) domain is shown in dark
S156AT168AY175A (AAA) were purified as GST-fusion proteins and incubated with GSTfused
 were purified as GST-fusion proteins and incubated with GSTfused") 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 Supplemental Materials Supplemental Figure S1 (a) Phenotype of the wild type and grik1-2 grik2-1 plants after 8 days in darkness.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 Supplemental Materials Supplemental Figure S1 (a) Phenotype of the wild type and grik1-2 grik2-1 plants after 8 days in darkness.
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Nakajima and Tanoue, http://www.jcb.org/cgi/content/full/jcb.201104118/dc1 Figure S1. DLD-1 cells exhibit the characteristic morphology
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Nakajima and Tanoue, http://www.jcb.org/cgi/content/full/jcb.201104118/dc1 Figure S1. DLD-1 cells exhibit the characteristic morphology
DIFFERENTIAL RESPONSE OF PCNA-ASSOCIATED CDK ACTIVITY TO PHYTOHORMONES DURING MAIZE GERMINATION
 Ms. 00 rec.//0 DIFFERENTIAL RESPONSE OF PCNA-ASSOCIATED CDK ACTIVITY TO PHYTOHORMONES DURING MAIZE GERMINATION María de la Paz Sánchez, Sunitha H. Gurusinghe, Kent J. Bradford and Jorge M. Vázquez-Ramos
Ms. 00 rec.//0 DIFFERENTIAL RESPONSE OF PCNA-ASSOCIATED CDK ACTIVITY TO PHYTOHORMONES DURING MAIZE GERMINATION María de la Paz Sánchez, Sunitha H. Gurusinghe, Kent J. Bradford and Jorge M. Vázquez-Ramos
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature10928 Materials and Methods 1. Plant material and growth conditions. All plant lines used were in Col-0 background unless otherwise specified. pif4-101 mutant
SUPPLEMENTARY INFORMATION doi:10.1038/nature10928 Materials and Methods 1. Plant material and growth conditions. All plant lines used were in Col-0 background unless otherwise specified. pif4-101 mutant
Transcriptional regulation of BRCA1 expression by a metabolic switch: Di, Fernandez, De Siervi, Longo, and Gardner. H3K4Me3
 ChIP H3K4Me3 enrichment.25.2.15.1.5 H3K4Me3 H3K4Me3 ctrl H3K4Me3 + E2 NS + E2 1. kb kb +82 kb Figure S1. Estrogen promotes entry of MCF-7 into the cell cycle but does not significantly change activation-associated
ChIP H3K4Me3 enrichment.25.2.15.1.5 H3K4Me3 H3K4Me3 ctrl H3K4Me3 + E2 NS + E2 1. kb kb +82 kb Figure S1. Estrogen promotes entry of MCF-7 into the cell cycle but does not significantly change activation-associated
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Dynamic Phosphorylation of HP1 Regulates Mitotic Progression in Human Cells Supplementary Figures Supplementary Figure 1. NDR1 interacts with HP1. (a) Immunoprecipitation using
SUPPLEMENTARY INFORMATION Dynamic Phosphorylation of HP1 Regulates Mitotic Progression in Human Cells Supplementary Figures Supplementary Figure 1. NDR1 interacts with HP1. (a) Immunoprecipitation using
At E17.5, the embryos were rinsed in phosphate-buffered saline (PBS) and immersed in
 and immersed in") Supplementary Materials and Methods Barrier function assays At E17.5, the embryos were rinsed in phosphate-buffered saline (PBS) and immersed in acidic X-gal mix (100 mm phosphate buffer at ph4.3, 3 mm
Supplementary Materials and Methods Barrier function assays At E17.5, the embryos were rinsed in phosphate-buffered saline (PBS) and immersed in acidic X-gal mix (100 mm phosphate buffer at ph4.3, 3 mm
Strep-Spin Protein Miniprep Kit Catalog No. P2004, P2005
 INSTRUCTION MANUAL Strep-Spin Protein Miniprep Kit Catalog No. P2004, P2005 Highlights Fast protocol to purify Strep-tagged proteins from cell-free extracts Screen your recombinant colonies directly for
INSTRUCTION MANUAL Strep-Spin Protein Miniprep Kit Catalog No. P2004, P2005 Highlights Fast protocol to purify Strep-tagged proteins from cell-free extracts Screen your recombinant colonies directly for
However, only a fraction of these genes are transcribed in an individual cell at any given time.
 All cells in an organism contain the same set of genes. However, only a fraction of these genes are transcribed in an individual cell at any given time. It is the pattern of gene expression that determines
All cells in an organism contain the same set of genes. However, only a fraction of these genes are transcribed in an individual cell at any given time. It is the pattern of gene expression that determines
As a control to demonstrate that we could isolate PCR products that result from ligating
 SUPPLEMENTAL INFORMATION (Mullen and Marzluff) RESULTS Oligo(dA) Clones and Oligo(dT) RT-PCR controls. As a control to demonstrate that we could isolate PCR products that result from ligating the authentic
SUPPLEMENTAL INFORMATION (Mullen and Marzluff) RESULTS Oligo(dA) Clones and Oligo(dT) RT-PCR controls. As a control to demonstrate that we could isolate PCR products that result from ligating the authentic
Supplementary Figure 1 Collision-induced dissociation (CID) mass spectra of peptides from PPK1, PPK2, PPK3 and PPK4 respectively.
 mass spectra of peptides from PPK1, PPK2, PPK3 and PPK4 respectively.") Supplementary Figure 1 lision-induced dissociation (CID) mass spectra of peptides from PPK1, PPK, PPK3 and PPK respectively. % of nuclei with signal / field a 5 c ppif3:gus pppk1:gus 0 35 30 5 0 15 10
Supplementary Figure 1 lision-induced dissociation (CID) mass spectra of peptides from PPK1, PPK, PPK3 and PPK respectively. % of nuclei with signal / field a 5 c ppif3:gus pppk1:gus 0 35 30 5 0 15 10
Supplementary Figure 1. jmj30-2 and jmj32-1 produce null mutants. (a) Schematic drawing of JMJ30 and JMJ32 genome structure showing regions amplified
 Schematic drawing of JMJ30 and JMJ32 genome structure showing regions amplified") Supplementary Figure 1. jmj30-2 and jmj32-1 produce null mutants. (a) Schematic drawing of JMJ30 and JMJ32 genome structure showing regions amplified by primers used for mrna expression analysis. Gray
Supplementary Figure 1. jmj30-2 and jmj32-1 produce null mutants. (a) Schematic drawing of JMJ30 and JMJ32 genome structure showing regions amplified by primers used for mrna expression analysis. Gray
SUPPLEMENTARY INFORMATION FILE
 SUPPLEMENTARY INFORMATION FILE Existence of a microrna pathway in anucleate platelets Patricia Landry, Isabelle Plante, Dominique L. Ouellet, Marjorie P. Perron, Guy Rousseau & Patrick Provost 1. SUPPLEMENTARY
SUPPLEMENTARY INFORMATION FILE Existence of a microrna pathway in anucleate platelets Patricia Landry, Isabelle Plante, Dominique L. Ouellet, Marjorie P. Perron, Guy Rousseau & Patrick Provost 1. SUPPLEMENTARY
ONE-HOUR Complete IP-Western Kit
 ONE-HOUR Complete IP-Western Kit Technical Manual No. 0218 Version 06192009 I Description.. 1 II Kit Contents.. 2 III Related Products 2 IV Key Features.. 2 V Storage.. 2 VI ONE-HOUR Western Protocol 3
ONE-HOUR Complete IP-Western Kit Technical Manual No. 0218 Version 06192009 I Description.. 1 II Kit Contents.. 2 III Related Products 2 IV Key Features.. 2 V Storage.. 2 VI ONE-HOUR Western Protocol 3
Western-GUARANTEED Antibody Service FAQ
 Western-GUARANTEED Antibody Service FAQ Content Q 1: When do I need a Western GUARANTEED Peptide Antibody Package?...2 Q 2: Can GenScript provide a Western blot guaranteed antibody?...2 Q 3: Does GenScript
Western-GUARANTEED Antibody Service FAQ Content Q 1: When do I need a Western GUARANTEED Peptide Antibody Package?...2 Q 2: Can GenScript provide a Western blot guaranteed antibody?...2 Q 3: Does GenScript
H3K36me3 polyclonal antibody
 H3K36me3 polyclonal antibody Cat. No. C15410192 Type: Polyclonal ChIP-grade/ChIP-seq grade Source: Rabbit Lot #: A1845P Size: 50 µg/32 µl Concentration: 1.6 μg/μl Specificity: Human, mouse, Arabidopsis,
H3K36me3 polyclonal antibody Cat. No. C15410192 Type: Polyclonal ChIP-grade/ChIP-seq grade Source: Rabbit Lot #: A1845P Size: 50 µg/32 µl Concentration: 1.6 μg/μl Specificity: Human, mouse, Arabidopsis,
Kinase Reaction and Alkylation Protocol
 Kinase Reaction and Alkylation Protocol Protocol for the treatment of substrates prior to detection by Thiophosphate Ester antibodies This product is for research use only and is not intended for diagnostic
Kinase Reaction and Alkylation Protocol Protocol for the treatment of substrates prior to detection by Thiophosphate Ester antibodies This product is for research use only and is not intended for diagnostic
Chapter 14 Regulation of Transcription
 Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
- PDI5. - Actin A 300-UTR5. PDI5 lines WT. Arabidopsis Chromosome 1. PDI5 (At1g21750) SALK_ SALK_ SALK_015253
 SALK_ SALK_ SALK_015253") Supplemental Data. Ondzighi et al. (2008). Arabidopsis Protein Disulfide Isomerase-5 Inhibits ysteine Proteases During Trafficking to Vacuoles Prior to Programmed ell Death of the dothelium in Developing
Supplemental Data. Ondzighi et al. (2008). Arabidopsis Protein Disulfide Isomerase-5 Inhibits ysteine Proteases During Trafficking to Vacuoles Prior to Programmed ell Death of the dothelium in Developing
Kawamata et al., Figure S1
 Kawamata et al., Figure S1 +Lysate Lysate Time (min) ss let-7 1 15 3 6 Duplex A Duplex B 1 15 3 6 1 15 3 6 ss let-7 Duplex A Duplex B Complex I Complex II Complex III Complex IV Complex V Free ighter exposure
Kawamata et al., Figure S1 +Lysate Lysate Time (min) ss let-7 1 15 3 6 Duplex A Duplex B 1 15 3 6 1 15 3 6 ss let-7 Duplex A Duplex B Complex I Complex II Complex III Complex IV Complex V Free ighter exposure
Cross Linking Immunoprecipitation
 301PR 03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Cross Linking Immunoprecipitation Utilizes Protein A/G Agarose& DSS for Antibody
301PR 03 G-Biosciences 1-800-628-7730 1-314-991-6034 technical@gbiosciences.com A Geno Technology, Inc. (USA) brand name Cross Linking Immunoprecipitation Utilizes Protein A/G Agarose& DSS for Antibody
Supplementary Information
 Supplementary Information Peroxiredoxin-2 and STAT3 form a redox relay for H 2 O 2 signaling Mirko C. Sobotta 1, Willy Liou 1, Sarah Stöcker 1, Deepti Talwar 1, Michael Oehler 1, Thomas Ruppert 2, Annette
Supplementary Information Peroxiredoxin-2 and STAT3 form a redox relay for H 2 O 2 signaling Mirko C. Sobotta 1, Willy Liou 1, Sarah Stöcker 1, Deepti Talwar 1, Michael Oehler 1, Thomas Ruppert 2, Annette
Supplemental Data. Benstein et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure 1. Purification of the heterologously expressed PGDH1, PGDH2 and PGDH3 enzymes by Ni-NTA affinity chromatography. Protein extracts (2 µl) of different fractions (lane 1 = total extract,
Supplemental Figure 1. Purification of the heterologously expressed PGDH1, PGDH2 and PGDH3 enzymes by Ni-NTA affinity chromatography. Protein extracts (2 µl) of different fractions (lane 1 = total extract,
Supplemental Data. Zhang et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") SDLTXgal SDLTH SDLTHA ADSV4 BD BDP53 BD BDP53 BD BDP53 ADAt1g1567(aa36358) ADAt1g844(aa9354) ADAt1g844(aa51354) ADAt1g844(full) Prey ADAt1g844(aa11354) ADAt3g5994(aa48418) BD BDPAL2 BD BDPAL2 BD BDPAL2
SDLTXgal SDLTH SDLTHA ADSV4 BD BDP53 BD BDP53 BD BDP53 ADAt1g1567(aa36358) ADAt1g844(aa9354) ADAt1g844(aa51354) ADAt1g844(full) Prey ADAt1g844(aa11354) ADAt3g5994(aa48418) BD BDPAL2 BD BDPAL2 BD BDPAL2
Cdc42 Activation Assay Kit
 A helping hand for your research Product Manual Configuration-specific Monoclonal Antibody Based Cdc42 Activation Assay Kit Catalog Number: 80701 20 assays 1 Table of Content Product Description 3 Assay
A helping hand for your research Product Manual Configuration-specific Monoclonal Antibody Based Cdc42 Activation Assay Kit Catalog Number: 80701 20 assays 1 Table of Content Product Description 3 Assay
all samples of a band with a molecular weight close to that expected for the endogenous!-
 SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure 1 : Specificity of anti-!-arrestin Abs and levels of!-arrestin in WHIM wt leukocytes. (A and B) HEK 293T cells were transiently transfected using the reagent
SUPPLEMENTAL FIGURE LEGENDS Supplemental Figure 1 : Specificity of anti-!-arrestin Abs and levels of!-arrestin in WHIM wt leukocytes. (A and B) HEK 293T cells were transiently transfected using the reagent
Strep-Spin Protein Miniprep Kit Catalog No. P2004 & P2005
 INSTRUCTION MANUAL Strep-Spin Protein Miniprep Kit Catalog No. P2004 & P2005 Highlights Fast & Simple: Purify Strep-tagged proteins from cell-free extracts using a spin-column in 7 minutes High-Quality:
INSTRUCTION MANUAL Strep-Spin Protein Miniprep Kit Catalog No. P2004 & P2005 Highlights Fast & Simple: Purify Strep-tagged proteins from cell-free extracts using a spin-column in 7 minutes High-Quality:
ENCODE DCC Antibody Validation Document
 ENCODE DCC Antibody Validation Document Date of Submission 09/12/12 Name: Trupti Kawli Email: trupti@stanford.edu Lab Snyder Antibody Name: SREBP1 (sc-8984) Target: SREBP1 Company/ Source: Santa Cruz Biotechnology
ENCODE DCC Antibody Validation Document Date of Submission 09/12/12 Name: Trupti Kawli Email: trupti@stanford.edu Lab Snyder Antibody Name: SREBP1 (sc-8984) Target: SREBP1 Company/ Source: Santa Cruz Biotechnology
Figure S1. USP-46 is expressed in several tissues including the nervous system
 Supplemental Figure legends Figure S1. USP-46 is expressed in several tissues including the nervous system Transgenic animals expressing a transcriptional reporter (P::GFP) were imaged using epifluorescence
Supplemental Figure legends Figure S1. USP-46 is expressed in several tissues including the nervous system Transgenic animals expressing a transcriptional reporter (P::GFP) were imaged using epifluorescence
Coleman et al., Supplementary Figure 1
 Coleman et al., Supplementary Figure 1 BrdU Merge G1 Early S Mid S Supplementary Figure 1. Sequential destruction of CRL4 Cdt2 targets during the G1/S transition. HCT116 cells were synchronized by sequential
Coleman et al., Supplementary Figure 1 BrdU Merge G1 Early S Mid S Supplementary Figure 1. Sequential destruction of CRL4 Cdt2 targets during the G1/S transition. HCT116 cells were synchronized by sequential
Technical tips Session 5
 Technical tips Session 5 Chromatine Immunoprecipitation (ChIP): This is a powerful in vivo method to quantitate interaction of proteins associated with specific regions of the genome. It involves the immunoprecipitation
Technical tips Session 5 Chromatine Immunoprecipitation (ChIP): This is a powerful in vivo method to quantitate interaction of proteins associated with specific regions of the genome. It involves the immunoprecipitation
ENCODE DCC Antibody Validation Document
 ENCODE DCC Antibody Validation Document Date of Submission 06/15/2012 Name: Trupti Kawli Email: trupti@stanford.edu Lab Snyder Antibody Name: CDP (sc6327) Target: CDP Company/ Source: Santa Cruz Catalog
ENCODE DCC Antibody Validation Document Date of Submission 06/15/2012 Name: Trupti Kawli Email: trupti@stanford.edu Lab Snyder Antibody Name: CDP (sc6327) Target: CDP Company/ Source: Santa Cruz Catalog
Construction of plant complementation vector and generation of transgenic plants
 MATERIAL S AND METHODS Plant materials and growth conditions Arabidopsis ecotype Columbia (Col0) was used for this study. SALK_072009, SALK_076309, and SALK_027645 were obtained from the Arabidopsis Biological
MATERIAL S AND METHODS Plant materials and growth conditions Arabidopsis ecotype Columbia (Col0) was used for this study. SALK_072009, SALK_076309, and SALK_027645 were obtained from the Arabidopsis Biological
Histone H3 Methylation Antibody Panel Pack I - Active Genes Base Catalog # C Component Size Shipping Temperature
 Histone H3 Methylation Antibody Panel Pack I - Active Genes Base Catalog # PACK CONTENTS Component Size Shipping Temperature Upon Receipt Checklist 3K4D Histone H3K4me2 (H3K4 Dimethyl) Polyclonal Antibody
Histone H3 Methylation Antibody Panel Pack I - Active Genes Base Catalog # PACK CONTENTS Component Size Shipping Temperature Upon Receipt Checklist 3K4D Histone H3K4me2 (H3K4 Dimethyl) Polyclonal Antibody
Strep-tag detection in Western blots
 Strep-tag detection in Western blots General protocol for the detection of Strep-tag fusion proteins Last date of revision April 2012 Version PR07-0010 www.strep-tag.com For research use only Important
Strep-tag detection in Western blots General protocol for the detection of Strep-tag fusion proteins Last date of revision April 2012 Version PR07-0010 www.strep-tag.com For research use only Important
This Technical Note describes these characterization studies in more detail.
 Technical Note Characterization of SOMAmer Reagents Binding Specificity in the SOMAscan 1.3k Assay Summary Slow Off-Rate Modified Aptamer (SOMAmer) reagents are identified via the SELEX process against
Technical Note Characterization of SOMAmer Reagents Binding Specificity in the SOMAscan 1.3k Assay Summary Slow Off-Rate Modified Aptamer (SOMAmer) reagents are identified via the SELEX process against
REGULATION OF CELL CYCLE SPECIFIC GENES THROUGH PLANT GROWTH AND DEVELOPMENT
 REGULATION OF CELL CYCLE SPECIFIC GENES THROUGH PLANT GROWTH AND DEVELOPMENT Ph.D. Theses Dr. Kovácsné Lendvai Ágnes MTA Biological Research Centre, Institute of Plant Biology Supervisors: Prof. Dudits
REGULATION OF CELL CYCLE SPECIFIC GENES THROUGH PLANT GROWTH AND DEVELOPMENT Ph.D. Theses Dr. Kovácsné Lendvai Ágnes MTA Biological Research Centre, Institute of Plant Biology Supervisors: Prof. Dudits
Herpes Simplex Virus Type 1 Infection Imposes a G 1 /S Block in Asynchronously Growing Cells and Prevents G 1 Entry in Quiescent Cells
 Virology 267, 335 349 (2000) doi:10.1006/viro.1999.0147, available online at http://www.idealibrary.com on Herpes Simplex Virus Type 1 Infection Imposes a G 1 /S Block in Asynchronously Growing Cells and
Virology 267, 335 349 (2000) doi:10.1006/viro.1999.0147, available online at http://www.idealibrary.com on Herpes Simplex Virus Type 1 Infection Imposes a G 1 /S Block in Asynchronously Growing Cells and
A Modified Digestion-Circularization PCR (DC-PCR) Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks
 Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks") A Modified Digestion-Circularization PCR (DC-PCR) Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks SARAH K. DICKERSON AND F. NINA PAPAVASILIOU Laboratory of Lymphocyte Biology, The
A Modified Digestion-Circularization PCR (DC-PCR) Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks SARAH K. DICKERSON AND F. NINA PAPAVASILIOU Laboratory of Lymphocyte Biology, The
Supplementary Fig S1 Nutlin-3a treatment does not affect cell cycle progression in the absence
 Supplementary Figure Legends Supplementary Fig S1 Nutlin-3a treatment does not affect cell cycle progression in the absence of p53 or p21. HCT116 cells which were null for either p53 (A) or p21 (B) were
Supplementary Figure Legends Supplementary Fig S1 Nutlin-3a treatment does not affect cell cycle progression in the absence of p53 or p21. HCT116 cells which were null for either p53 (A) or p21 (B) were
Document S1. Supplemental Experimental Procedures and Three Figures (see next page)
") Supplemental Data Document S1. Supplemental Experimental Procedures and Three Figures (see next page) Table S1. List of Candidate Genes Identified from the Screen. Candidate genes, corresponding dsrnas
Supplemental Data Document S1. Supplemental Experimental Procedures and Three Figures (see next page) Table S1. List of Candidate Genes Identified from the Screen. Candidate genes, corresponding dsrnas
Fig. S1. CrgA intracellular levels in M. tuberculosis. Ten and twenty micrograms of
 Supplementary data Fig. S1. CrgA intracellular levels in M. tuberculosis. Ten and twenty micrograms of cell free protein lysates from WT M. tuberculosis (Rv) together with various known concentrations
Supplementary data Fig. S1. CrgA intracellular levels in M. tuberculosis. Ten and twenty micrograms of cell free protein lysates from WT M. tuberculosis (Rv) together with various known concentrations
Histone H3K27 Methylation Antibody Panel Pack Base Catalog # C Component Size Shipping Temperature
 Histone H3K27 Methylation Antibody Panel Pack Base Catalog # PACK CONTENTS Component Size Shipping Temperature Upon Receipt Checklist 3K27M Histone H3K27me1 (H3K27 Monomethyl) Polyclonal Antibody 25 µg
Histone H3K27 Methylation Antibody Panel Pack Base Catalog # PACK CONTENTS Component Size Shipping Temperature Upon Receipt Checklist 3K27M Histone H3K27me1 (H3K27 Monomethyl) Polyclonal Antibody 25 µg
Custom Antibodies Services. GeneCust Europe. GeneCust Europe
 GeneCust Europe Laboratoire de Biotechnologie du Luxembourg S.A. 2 route de Remich L-5690 Ellange Luxembourg Tél. : +352 27620411 Fax : +352 27620412 Email : info@genecust.com Web : www.genecust.com Custom
GeneCust Europe Laboratoire de Biotechnologie du Luxembourg S.A. 2 route de Remich L-5690 Ellange Luxembourg Tél. : +352 27620411 Fax : +352 27620412 Email : info@genecust.com Web : www.genecust.com Custom
Arabidopsis KRPs have distinct inhibitory activity toward cyclin D2-associated kinases, including plant-specific B-type cyclin-dependent kinase
 FEBS Letters 580 (2006) 336 340 Arabidopsis KRPs have distinct inhibitory activity toward cyclin D2-associated kinases, including plant-specific B-type cyclin-dependent kinase Tomohiro Nakai a, Ko Kato
FEBS Letters 580 (2006) 336 340 Arabidopsis KRPs have distinct inhibitory activity toward cyclin D2-associated kinases, including plant-specific B-type cyclin-dependent kinase Tomohiro Nakai a, Ko Kato
Non-Organic-Based Isolation of Mammalian microrna using Norgen s microrna Purification Kit
 Application Note 13 RNA Sample Preparation Non-Organic-Based Isolation of Mammalian microrna using Norgen s microrna Purification Kit B. Lam, PhD 1, P. Roberts, MSc 1 Y. Haj-Ahmad, M.Sc., Ph.D 1,2 1 Norgen
Application Note 13 RNA Sample Preparation Non-Organic-Based Isolation of Mammalian microrna using Norgen s microrna Purification Kit B. Lam, PhD 1, P. Roberts, MSc 1 Y. Haj-Ahmad, M.Sc., Ph.D 1,2 1 Norgen
Lecture 22 Eukaryotic Genes and Genomes III
 Lecture 22 Eukaryotic Genes and Genomes III In the last three lectures we have thought a lot about analyzing a regulatory system in S. cerevisiae, namely Gal regulation that involved a hand full of genes.
Lecture 22 Eukaryotic Genes and Genomes III In the last three lectures we have thought a lot about analyzing a regulatory system in S. cerevisiae, namely Gal regulation that involved a hand full of genes.
Lecture 7: Affinity Chromatography-II
 Lecture 7: Affinity Chromatography-II We have studied basics of affinity purification during last lecture. The current lecture is continuation of last lecture and we will cover following: 1. Few specific
Lecture 7: Affinity Chromatography-II We have studied basics of affinity purification during last lecture. The current lecture is continuation of last lecture and we will cover following: 1. Few specific
Learning Objectives. Define RNA interference. Define basic terminology. Describe molecular mechanism. Define VSP and relevance
 Learning Objectives Define RNA interference Define basic terminology Describe molecular mechanism Define VSP and relevance Describe role of RNAi in antigenic variation A Nobel Way to Regulate Gene Expression
Learning Objectives Define RNA interference Define basic terminology Describe molecular mechanism Define VSP and relevance Describe role of RNAi in antigenic variation A Nobel Way to Regulate Gene Expression
Supporting Information
 Supporting Information Üstün and Börnke, 2015 Figure S1: XopJ does not display acetyltransferase activity. Autoacetylation activity in vitro. Acetylation reactions using MBP, MBP-XopJ, GST and GST-HopZ1a
Supporting Information Üstün and Börnke, 2015 Figure S1: XopJ does not display acetyltransferase activity. Autoacetylation activity in vitro. Acetylation reactions using MBP, MBP-XopJ, GST and GST-HopZ1a
Supplemental Data. Na Xu et al. (2016). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure 1. The weak fluorescence phenotype is not caused by the mutation in At3g60240. (A) A mutation mapped to the gene At3g60240. Map-based cloning strategy was used to map the mutated site
Supplemental Figure 1. The weak fluorescence phenotype is not caused by the mutation in At3g60240. (A) A mutation mapped to the gene At3g60240. Map-based cloning strategy was used to map the mutated site
Aminoacid change in chromophore. PIN3::PIN3-GFP GFP S65 (no change) (5.9) (Kneen et al., 1998)
 (5.9) (Kneen et al., 1998)") Supplemental Table. Table S1. Fluorophore characteristics of used fluorescent proteins, Related to Figure 2 and 3. Transgenic line Fluprescent marker Aminoacid change in chromophore pk(a) Ref. PIN3::PIN3-GFP
Supplemental Table. Table S1. Fluorophore characteristics of used fluorescent proteins, Related to Figure 2 and 3. Transgenic line Fluprescent marker Aminoacid change in chromophore pk(a) Ref. PIN3::PIN3-GFP
Supplemental Material Igreja and Izaurralde 1. CUP promotes deadenylation and inhibits decapping of mrna targets. Catia Igreja and Elisa Izaurralde
 Supplemental Material Igreja and Izaurralde 1 CUP promotes deadenylation and inhibits decapping of mrna targets Catia Igreja and Elisa Izaurralde Supplemental Materials and methods Functional assays and
Supplemental Material Igreja and Izaurralde 1 CUP promotes deadenylation and inhibits decapping of mrna targets Catia Igreja and Elisa Izaurralde Supplemental Materials and methods Functional assays and
Figure 1: TDP-43 is subject to lysine acetylation within the RNA-binding domain a) QBI-293 cells were transfected with TDP-43 in the presence or
 QBI-293 cells were transfected with TDP-43 in the presence or") Figure 1: TDP-43 is subject to lysine acetylation within the RNA-binding domain a) QBI-293 cells were transfected with TDP-43 in the presence or absence of the acetyltransferase CBP and acetylated TDP-43
Figure 1: TDP-43 is subject to lysine acetylation within the RNA-binding domain a) QBI-293 cells were transfected with TDP-43 in the presence or absence of the acetyltransferase CBP and acetylated TDP-43
This is the author's accepted version of the manuscript.
 This is the author's accepted version of the manuscript. The definitive version is published in Nature Communications Online Edition: 2015/4/16 (Japan time), doi:10.1038/ncomms7780. The final version published
This is the author's accepted version of the manuscript. The definitive version is published in Nature Communications Online Edition: 2015/4/16 (Japan time), doi:10.1038/ncomms7780. The final version published
Solutions for Your Research
 Solutions for Your Research Custom Antibody Services Polyclonal Monoclonal The service we offer is very complete starting from rabbits, mice and rats. The different formats provided by Primm depend on
Solutions for Your Research Custom Antibody Services Polyclonal Monoclonal The service we offer is very complete starting from rabbits, mice and rats. The different formats provided by Primm depend on
This Document Contains:
 This Document Contains: 1. In-Cell Western Protocol II. Cell Seeding and Stimulation Supplemental Protocol III. Complete Assay Example: Detailing the Seeding, Stimulation and Detection of the A431 Cellular
This Document Contains: 1. In-Cell Western Protocol II. Cell Seeding and Stimulation Supplemental Protocol III. Complete Assay Example: Detailing the Seeding, Stimulation and Detection of the A431 Cellular
Supplementary materials
 Supplementary materials Zidovudine Induces Down-Regulation of Mitochondrial Deoxynucleoside Kinases: Implications for Mitochondrial Toxicity of Antiviral Nucleoside Analogs Ren Sun, Staffan Eriksson, and
Supplementary materials Zidovudine Induces Down-Regulation of Mitochondrial Deoxynucleoside Kinases: Implications for Mitochondrial Toxicity of Antiviral Nucleoside Analogs Ren Sun, Staffan Eriksson, and
Histone H3 Methylation Antibody Panel Pack III - Active Genes Base Catalog # C Component Size Shipping Temperature
 Histone H3 Methylation Antibody Panel Pack III - Active Genes Base Catalog # PACK CONTENTS Component Size Shipping Temperature Upon Receipt Checklist 3R17A Histone H3R17 Dimethyl Asymmetric (H3R17me2a)
Histone H3 Methylation Antibody Panel Pack III - Active Genes Base Catalog # PACK CONTENTS Component Size Shipping Temperature Upon Receipt Checklist 3R17A Histone H3R17 Dimethyl Asymmetric (H3R17me2a)
Giardia RNAi Paper Discussion. Supplemental - VSG clonality. Variant-specific surface protein VSP9B10
 Giardia RNAi Paper Discussion Supplemental - VSG clonality VSP9B10 Green - VSP9B10 antibody Blue - DAPI Demonstration that cell line expresses a single VSP Variant-specific surface protein Outside Inside
Giardia RNAi Paper Discussion Supplemental - VSG clonality VSP9B10 Green - VSP9B10 antibody Blue - DAPI Demonstration that cell line expresses a single VSP Variant-specific surface protein Outside Inside