Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1. Linkage preference of USP30 and USP domain architecture.
|
|
|
- Jacob Walters
- 6 years ago
- Views:
Transcription
1 Supplementary Figure 1 Linkage preference of USP30 and USP domain architecture. a, Catalytic efficiencies determined from Coomassie-stained gel-based kinetics for all eight diub linkages (see Supplementary Data Set 1 for raw data). Error bars represent standard deviations of the mean, obtained from five (Lys6), three (Lys11, Lys48) or two (Lys27, Lys33, Lys63, Met1) time courses. b, Time course of tetraubiquitin cleavage for Lys6, Lys11, Lys48, Lys63 and Met1-linked chains, resolved by SDS-PAGE and visualised by silver staining. The gel of Lys6-tetraubiquitin from Fig. 1c is included for comparison. It shows at the earliest time points that tri- and monoubiquitin are generated, consistent with exo-dub activity (Hospenthal, M.K et al., Nat Struct Mol Biol 20, , 2013). c, Annotation of the USP domain of USP7 (PDB 1NBF, Hu, M. et al., Cell 111, , 2002) according to the box system with insertion points indicated (Ye, Y. et al., Mol Biosyst 5, , 2009). d, USP domain nomenclature indicating thumb, finger and palm subdomains (Hu, M. et al., Cell 111, , 2002). e, Plot of average sequence conservation of USP30 (rolling window average of 9 residues. 10: high sequence conservation, 0: low sequence conservation, obtained from ConSurf, and deuterium incorporation obtained from HDX experiments on USP30 c2 (compare Fig. 1d), suggesting flexibility of regions with low sequence conservation.

2 Supplementary Figure 2 USP30 construct optimization through insertion deletions. a, Sequence alignment of the C-terminal end of box 4 of USP30 with other USPs that have been crystallised. Of note, the corresponding residues of Phe348, Met350 and Ile353 are typically hydrophilic residues that are solvent exposed on the domain backside in other USP structures, and were mutated in USP30 to increase protein solubility (see Supplementary Table 1). b, Coomassie-stained gel of indicated USP30 proteins used for crystallography. c, Melting temperatures of apo and covalent Ub-PA complexes of indicated USP30 constructs, indicating similar melting temperatures for constructs with or without the box 4-5 insertion and with or without mutation of the three hydrophobic residues shown in a, suggesting that the protein is similarly stable once purified. In contrast, additional deletion of the box 2-3 insertion significantly increased protein stability. Crystallised proteins are indicated with an arrow. Experiments were performed in triplicate and error bars represent standard deviation of the mean. d, Representative anisotropy traces for indicated USP30 proteins in the ubiquitin-kg-tamra cleavage assay. e, Catalytic efficiencies of indicated constructs derived from ubiquitin-kg-tamra cleavage. Experiments were performed in triplicate and error bars represent standard error. f, Lys6- diubiquitin cleavage assay for indicated constructs.
. a, Overall view of the asymmetric unit for USP30 c13 ~Ub-PA. b, Overall view of the asymmetric unit for USP30 c8 ~Ub-PA.")
3 Supplementary Figure 3 Electron density and SEC-MALS experiment. Electron density maps in this Figure correspond to weighted 2 F O F C electron density contoured at 1 (0.8 for the 3.6 Å USP30 c8 ~Ub-PA structure). a, Overall view of the asymmetric unit for USP30 c13 ~Ub-PA. b, Overall view of the asymmetric unit for USP30 c8 ~Ub-PA. c, Close-up view of the covalently modified catalytic triad residues of USP30 c13 ~Ub-PA (top) and USP30 c8 ~Ub-PA (bottom). d, Size-exclusion chromatography multi-angle light scattering (SEC-MALS) experiment on complex formation of USP30 c13i and Lys6-diubiquitin. While a clear shift is apparent from the elution profile, the observed complex mass is smaller than expected, indicating that the complex is not fully gel-filtration stable at the concentrations tested (80 M sample concentration, 1.2-fold molar excess of Lys6-diubiquitin). e, Overall view of the asymmetric unit for USP30 c13i + Lys6-diUb. f, Close-up view of the isopeptide bond in the USP30 active site from two different perspectives.

4 Supplementary Figure 4 HDX-MS analysis of ubiquitin binding to USP30. a, Deuterium incorporation measured by HDX-MS on USP30 c8 mapped onto the USP30 molecule from USP30 c13i -Lys6-diUb. Data for residues not part of the model are shown as horizontal bars. b, Deuterium incorporation measured by HDX-MS on USP30 c8 ~Ub-PA mapped as in a. c, Subtraction of data from HDX-MS runs on USP30 c8 in the presence and absence of Ub-PA mapped onto the model. The deuterium incorporation in the palm and thumb domain is virtually unchanged by ubiquitin binding. There is a significant reduction in deuterium uptake in the fingers domain, suggesting rigidification through ubiquitin binding. The box 2-3 loop insertion (residues ) has regions of high deuterium uptake (>45%) in both the apo and the Ub-PA-bound state, consistent with this part not being required for ubiquitin binding and with no density being observable for this part in the 3.6 Å structure of USP30 c8 ~Ub-PA.
5 Supplementary Figure 5 Binding of the distal ubiquitin moiety to USP30 and characterization of USP30 active site mutations. a, Sequence alignment of crystallised USP proteins centring on Leu328 and Phe453 of USP30, showing differences in the coordination of the distal ubiquitin s C-terminal tail compared to other USPs (compare Fig. 2a). b,c, Fluorescence polarisation binding experiment of USP30 c8 C77A (one loop deleted) and USP30 c2 C77A (no loop deleted) resulting in similarly poor binding of ubiquitin-kg-tamra as compared to USP30 c13i (Fig. 2b), indicating that loop deletions do not impact on the distal ubiquitin binding site of USP30. Experiments were performed in triplicate and error bars represent standard deviation from the mean. d, Fluorescence polarisation experiment of ubiquitin-kg-tamra binding to inactive USP21 C221A, as a positive control for b, c and Fig. 2b. Experiments were performed in triplicate and error bars represent standard deviation from the mean. e, Triplicate-averaged anisotropy traces for indicated USP30 proteins with active site residues mutated in the ubiquitin-kg-tamra cleavage assay. f, Observed rate constants were obtained from fitting anisotropy-time courses plotted over enzyme concentrations. The slopes correspond to catalytic efficiencies (k cat /K M ) and are plotted in Fig. 2d.
 binding experiment with FlAsH-tagged Lys6-diubiquitin and indicated USP30 proteins, showing identical affinities compared to USP30 c13i (Fig.")
6 Supplementary Figure 6 Selective recognition and cleavage of Lys6-diUb by USP30. a, Fluorescence polarisation (FP) binding experiment with FlAsH-tagged Lys6-diubiquitin and indicated USP30 proteins, showing identical affinities compared to USP30 c13i (Fig. 3a), indicating that the deleted loops do not affect binding to Lys6-diubiquitin. b, Control FP binding experiment of FlAsH-tagged diubiquitin probes used in Fig. 3a with inactive USP21 C221A, indicating that all probes can bind USP domains in principle. Mean ± standard error from three independent measurements. c, Isothermal titration calorimetry (ITC) data for asymmetric Lys6-diUb* (distal Ub: K6R, K48R; proximal Ub: K48R, Gly76) binding to USP30 c13i, confirming the affinities obtained from FP binding assays. d, Superposition of free ubiquitin (PDB 1UBQ, Vijay-Kumar, S. et al., J Mol Biol 194, , 1987), the proximal Ub of Lys6-diubiquitin (PDB 2XK5, Virdee, S. et al., Nat Chem Biol 6, , 2010) and the proximal Ub in the USP30 c13i -Lys6-diUb structure. Close-up on the 1-2 loops showing how binding to USP30 induces a distinct conformation in the proximal Ub moiety. e, Sequence alignment of structurally characterised USP domains, centred on Glu159. f, Surface representation of USP30 c13i -Lys6-diUb and close-up view of the recognition site for the isopeptide linkage. The distal Ub C-terminus and the isopeptide bond are fully engulfed by the USP domain, with Glu159 of the thumb subdomain contacting the palm subdomain and coordinating the Gly76 amide backbone. g, Specificity analysis for USP30 S477D (compare Fig. 1b). h, Diubiquitin cleavage assay with indicated substrates and USP30 Ser477 mutants at identical concentrations.
, revealing a significantly better distal as compared to proximal binding interface: larger interface area (1652 Å 2 vs.")
7 Supplementary Figure 7 Analysis of Lys6-diUb upon binding to USP30. a, Comparison of proximal versus distal ubiquitin binding sites in USP30. The USP30 c13i -Lys6-diUb complex was characterised by PISA ( revealing a significantly better distal as compared to proximal binding interface: larger interface area (1652 Å 2 vs. 535 Å 2 ), larger numbers of hydrogen bonds (39 vs. 8) and salt bridges (10 vs. 2) and lower solvation free energy ( 6.4 vs. 2.0 kcal/mol), suggesting that the distal ubiquitin may bind USP30 first. b, Cartoon representation of Lys6-diubiquitin (PDB 2XK5). Lys6-diubiquitin adopts a compact conformation, with Ile36, Leu8, Leu71 of the distal ubiquitin contacting the Ile44-patch of the proximal ubiquitin (Hospenthal, M.K et al., Nat Struct Mol Biol 20, , 2013). c, Superposition of a and b on the distal Ub moiety. USP30 binds the distal ubiquitin s Ile36 patch via Trp330, which requires Lys6-diubiquitin to open for the initial binding event. d,e,g, Superposition centred on the proximal ubiquitin, analogously to a,b,c. The proximal ubiquitin binding site of USP30 recognises the ubiquitin Phe4 patch, which is exposed in Lys6-diubiquitin. This binding site, hence, could recognise compact Lys6-diubiquitin and enable it to open subsequently.
8 Supplementary Figure 8 Characterization USP30 mutations of the proximal ubiquitin binding site. a, Anisotropy traces for indicated USP30 proteins with proximal binding site residues mutated in the ubiquitin-kg-tamra cleavage assay. b, Observed rate constants from a plotted over enzyme concentration to derive catalytic efficiencies. c, Catalytic efficiencies of indicated USP30 proteins derived from the ubiquitin-kg-tamra cleavage assay. Replacement of His445 and Trp475 leads to a reduction of catalytic activity on ubiquitin-kg-tamra, presumably due to their close proximity to the catalytic residues. Mean ± standard error are shown, derived from three independent experiments. d, Diubiquitin cleavage assay with indicated substrates and USP30 mutants, titrated to similar activity levels. H445N and D479Q retain the preference for Lys6-diUb, whereas H445S and H445Q show a cleavage pattern similar to H445E and W475K (see Fig. 3f).

9 Supplementary Figure 9 CYLD recognizes Lys63-diubiquitin by a distinctly different interface. a, Open-book representation of the CYLD(C596A)-Lys63-diubiquitin binding interface, compare Fig. 3d. b, Side-on view of the proximal ubiquitin binding site in CYLD. Specific recognition of the proximal Ub is achieved by extensive contacts made by CYLD s unique insertion, which is contacted by ubiquitin Glu64, Thr14, Thr12 and Phe4, as well as coordination of Met1, Glu16 and Glu18 by the USP domain. While Phe4 packs against the aliphatic part of the Arg824 side chain, polar contacts and salt bridges overall dominate this interface. Hence, while similar ubiquitin surfaces (Phe4 patch) are involved, the distinct orientations of the proximal ubiquitin in USP30 and CYLD dictate different linkage preferences, facilitated by unique structural elements.
 formed by a cleavage in the flexible part of the box 2-3 insertion. As for USP30 c13 (Fig.")
10 Supplementary Figure 10 Parkin-mediated USP30 ubiquitination. a, Left, ubiquitination of USP30 c2 by phosphorylated Parkin, visualised by western blotting with an anti-usp30 antibody. The antibody also detects a prominent proteolytic fragment of USP30 (marked with *) formed by a cleavage in the flexible part of the box 2-3 insertion. As for USP30 c13 (Fig. 5a) ubiquitination is enhanced for inactive USP30, suggesting that USP30 can auto-deubiquitinate in trans. Mass spectrometry of these samples identified several (Lys289, Lys235, Lys310) ubiquitination sites. See Supplementary Table 2 for mass spectrometry results. Right, the same samples blotted with an anti-ubiquitin antibody, revealing extensive polyubiquitin assembly by activated Parkin. This contrasts the USP30 blot which does not show evidence for USP30 polyubiquitination. b, Gel shown in Fig. 5c. Anion-exchange chromatography allowed partial separation of F4R-ubiquinated and unmodified USP30 with coeluting E1 and Parkin species. c, Superposition of USP30 c13 ~Ub-PA and USP30 c13i -Lys6-diUb, and close-up views on (1) the coordination of Phe4 of the distal ubiquitin moiety by the fingers domain, explaining the rigidification observed in HDX-MS through ubiquitin binding in this region as well as the inability of USP30 to process Ub F4R, (2) the coordination of Lys6 of the distal ubiquitin moiety by the side chain of Glu227 and the carbonyl backbone of Phe273, consistent with the observed exo activity in Lys6-tetraubiquitin cleavage experiments (Fig. 1b).
11 Supplementary Figure 11 PINK1-mediated monophosphorylation of Lys6-diubiquitin on the distal Ub moiety. a, Asymmetric Lys6-diubiquitin was monophosphorylated by PhPINK1, and following PhPINK1 heat-inactivation cleaved by USP21. Deconvoluted mass spectra are shown. The major phosphorylated species corresponds to the distal Ub moiety. The small peak at 8,618 Da may contain both unphosphorylated distal Ub as well as phosphorylated proximal Ub, which could not be separated due to limited resolution of the mass spectrometer used. b, Close-up view on the distal Ub Ser65 environment of USP30 (see Fig. 6d for superposition in full). c, Deconvoluted mass spectra of the ubiquitin-kg-tamra and the Ser65-phosphorylated phosphoub-kg- TAMRA reagents. Oxidation products (+ 1 oxygen, +16 Da), present also in the untreated sample, are indicated by + [O]. d,e Triplicate averaged anisotropy time traces and observed rate constants for USP30-mediated cleavage of ubiquitin- and phosphoub-kg-tamra (see Fig. 6e for catalytic efficiencies).
12 Supplementary Figure 12 Characterization data on differently phosphorylated Lys6-diUb substrates. a, Sample characterisation data for assay shown in Fig. 6f. USP30 c13i was ubiquitinated by Parkin, the sample then split and one half treated with PINK1. Analysis of the ubiquitin smear indicates the presence of Ser65-phosphoubiquitin only in the PINK1-treated sample. USP30 c13i ~Ub is not recognised by either the ubiquitin or the pser65-ubiquitin antibody. b, Asymmetric Lys6-diUb chains with different phosphorylation states were obtained through assembly from phosphorylated monoub species. Deconvoluted intact protein mass spectra for reagent characterisation are shown for the chains and the respective monoub species obtained after treatment with USP21 (compare Supplementary Fig. 11a).

13 Supplementary Figure 13 Regulation through USP30 overexpression of Lys6-linked polyubiquitin chains on MOM proteins unaffected by USP30 knockdown. a, Immunoblotting for CISD1 in eluates of Lys6-linked polyubiquitin chain pull-downs from HeLa Flp-In T-REx cells, doxycyclineinducibly expressing wild-type Parkin. Cells treated as indicated, all samples were treated with USP21 to deplete other polyubiquitin linkages. CISD1 forms a constitutive dimer, explaining the pull-down of monoubiquitinated CISD1 protein forms. b, Experiment as in a with USP30 knock down. Immunoblotting for VDAC1, MIRO1 and CISD1. *, non-specific bands. c, Quantification from b with n = 3 independent experiments. Mean ± s.e.m. Wilcoxon-Mann-Whitney test. n.s., non-significant. d, Experiment as in a with overexpression of indicated USP30 variants via transient transfection. e, Quantification from d. Mean ± s.e.m. One-Way ANOVA (F = 48) with Dunnett s correction, n = 3 independent experiments. ***, P <

14 Supplementary Figure 14 Model of localized mitophagy. USP30 displays normal levels of activity in the healthy part of the mitochondrial network. In the zone around mitochondrial damage, however, USP30 activity would be reduced due to the activity of PINK1, the presence of phosphoubiquitin and the impaired catalytic of USP30 for phosphoubiquitin substrates.
15 Supplementary Table 1: USP30 constructs used in this study. Construct Boundaries Mutations Box 4/5 deletion Box 2/3 deletion Solubility Used for wt - - good wt - - good wt ( )SGS - poor wt ( )SNA - poor wt ( ) - poor F348Q, M350N, I353Q ( )SNA - moderate F348D, M350S, I353E ( )SNA - good F348D, M350D, I353E ( )SNA - good 3.6 Å ~Ub-PA structure F348G, M350N, I353E ( )SNA - good F348D, M350S, I353E ( )SNA ( ) good F348D, M350S, I353E ( )SNA ( ) good F348D, M350S, I353E ( )SNA ( ) good F348D, M350S, I353E ( )SNA ( )GSGS good 13i C77A, F348D, M350S, I353E ( )SNA ( )GSGS moderate 2.3 Å ~Ub-PA structure 2.8 Å structure of USP30 / Lys6- diub complex
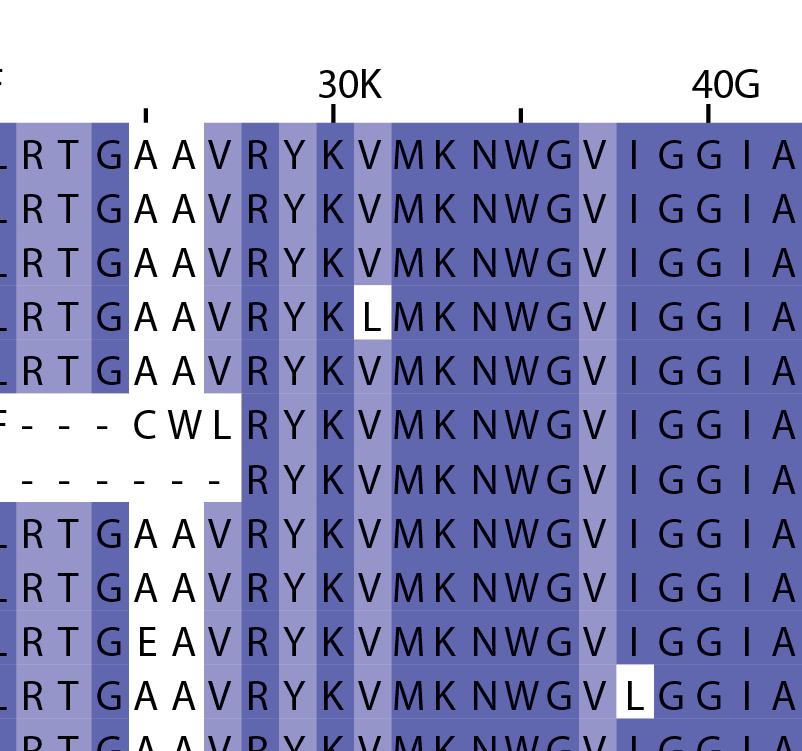


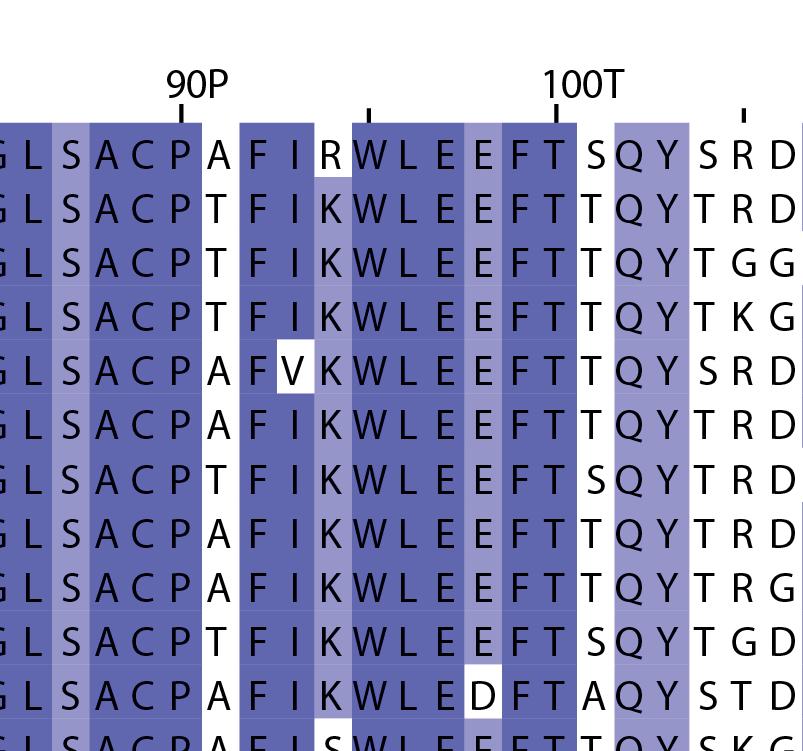
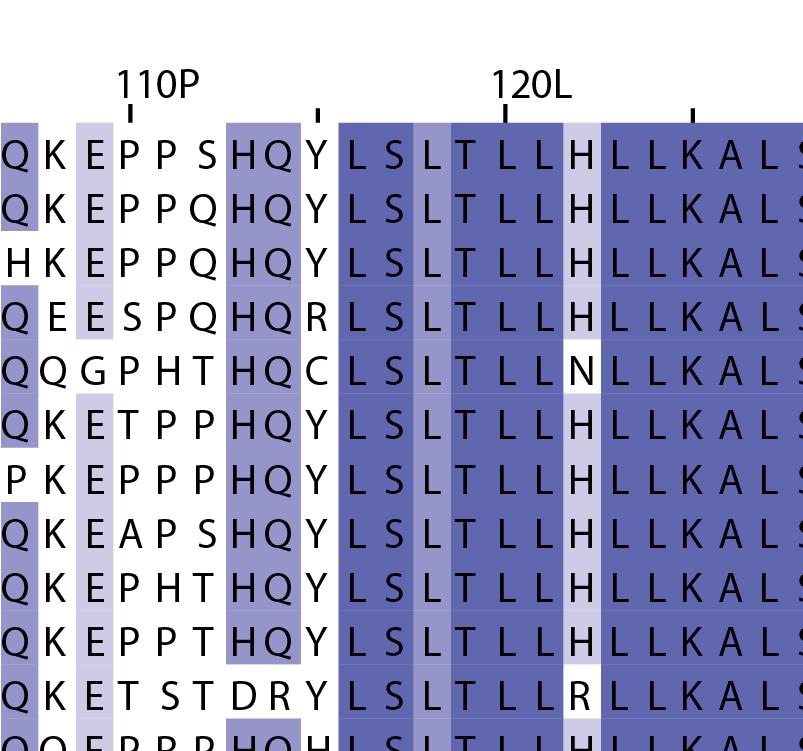











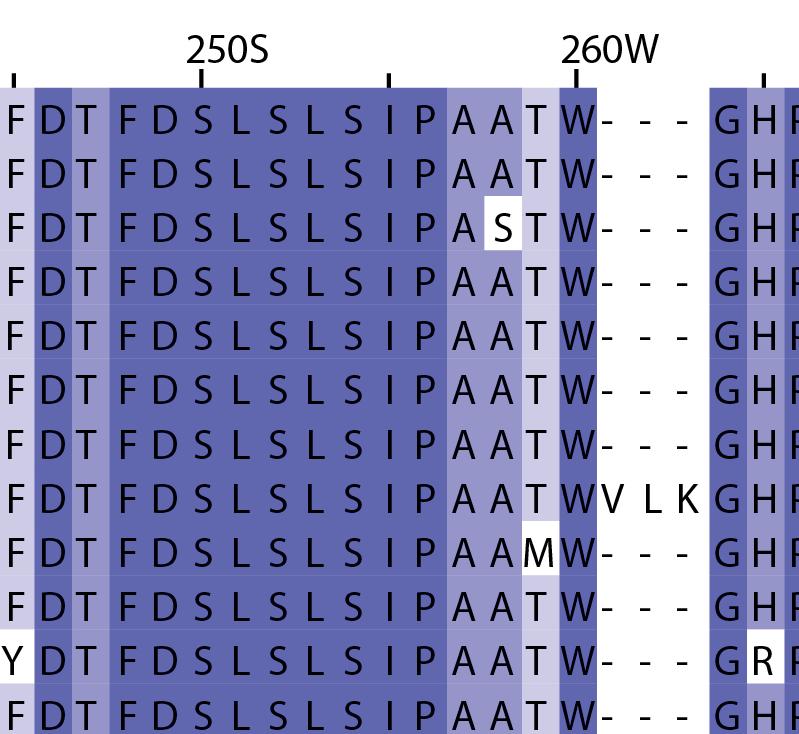







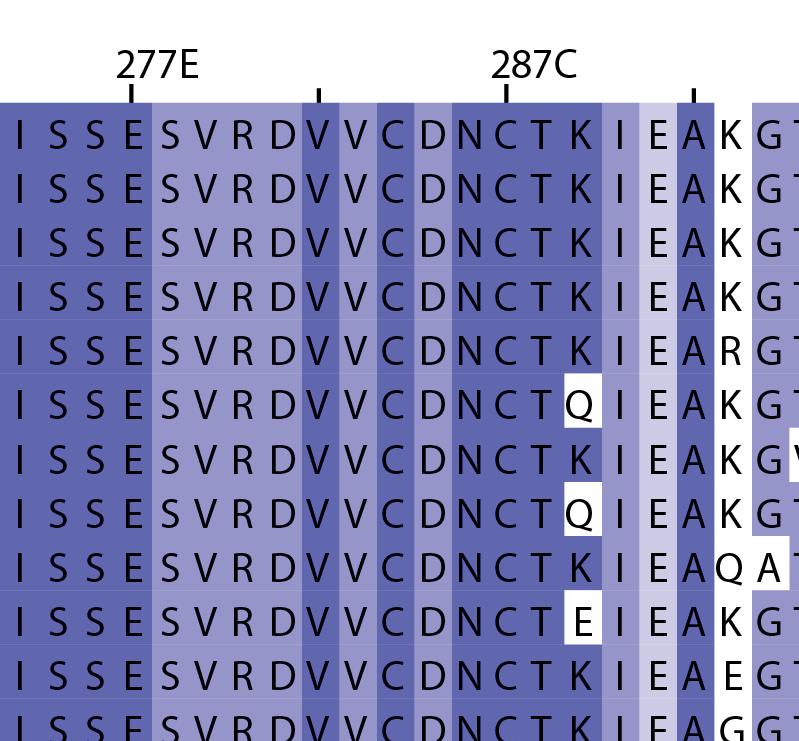


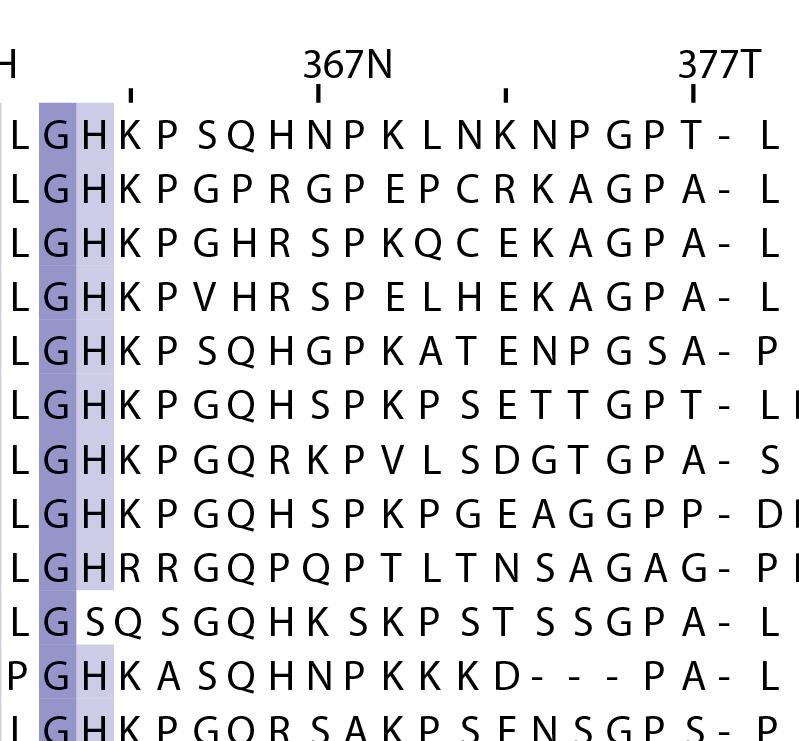
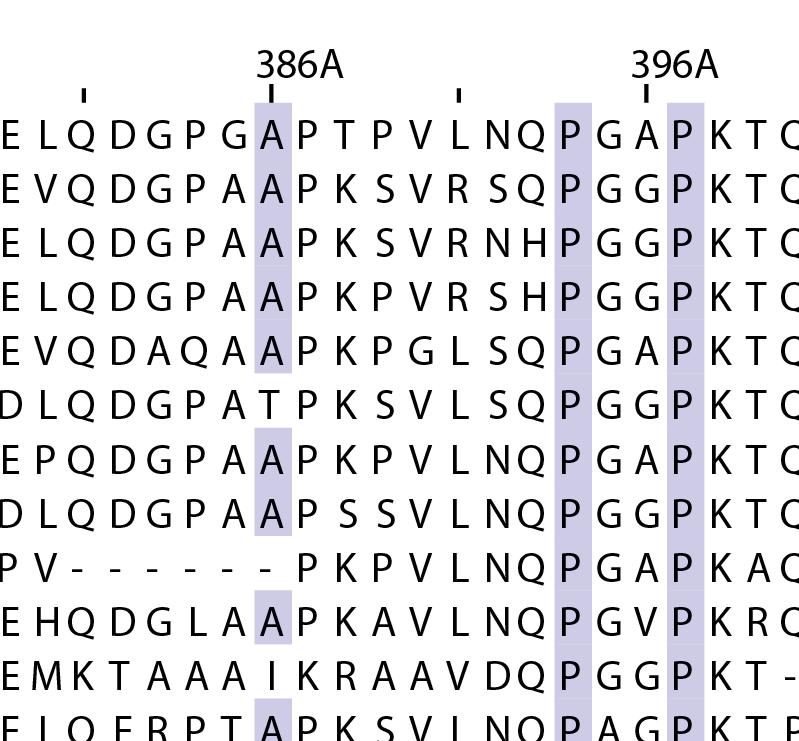
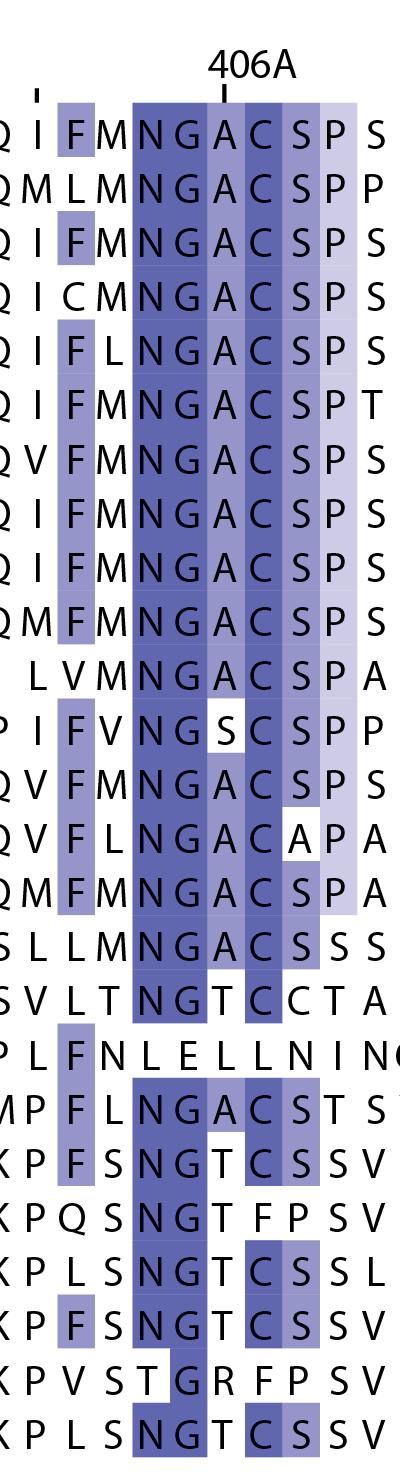


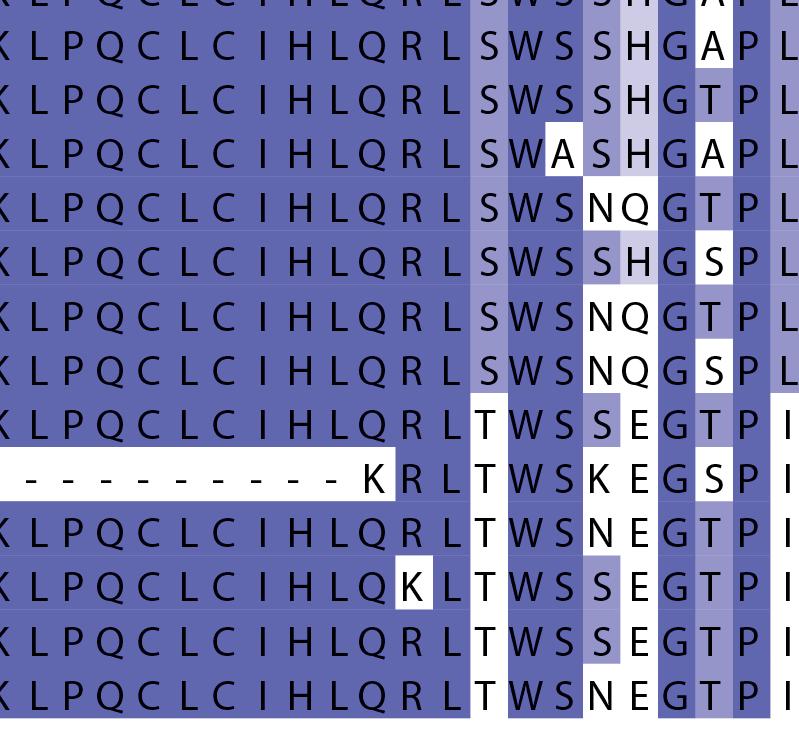

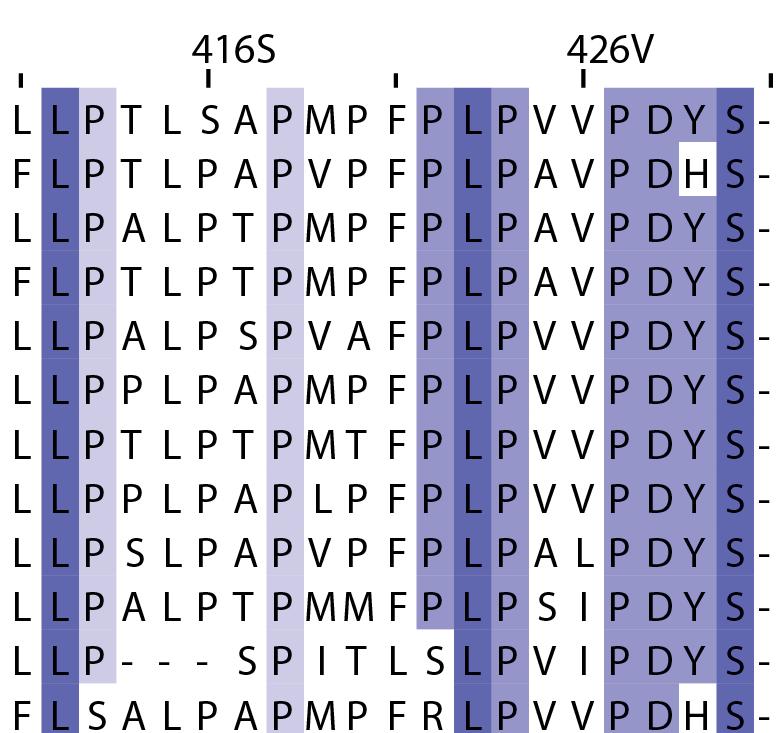
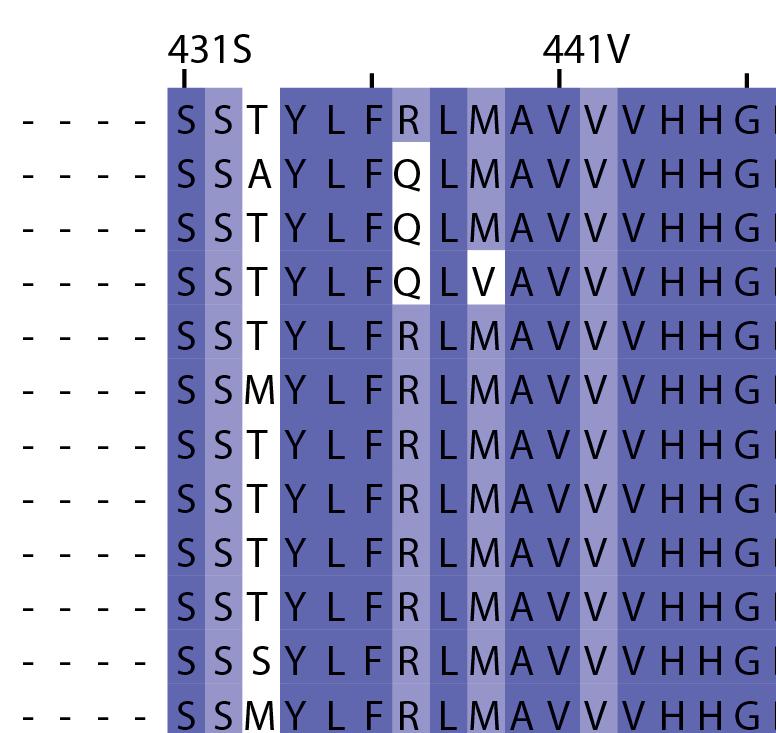









16 Supplementary Note 1: Sequence alignment of USP30 orthologs. The mitochondrial intermembrane part, the transmembrane helix, the polybasic signal anchor sequence and the USP boxes are indicated. Residues are coloured according to sequence identity. Select residues are marked with stars according to the legend. Of note, several USP30 orthologs have a C-terminal truncation similar to the construct used for structure determination. Strikingly, a subset of USP30 sequences in the lower part of the alignment shows hydrophilic residues (Ser, Arg) at the positions of the hydrophobic residues that were mutated along with the box 4-5 insertion deletion, suggesting that hydrophobic contacts between the insertion sequence and the USP domain at this position are not universally conserved. mitochondrial intermembrane part transmembrane helix anchor USP box 1 USP box 2 box 2/3 insertion deletion ( ) USP box 2 USP box 3 box 4/5 insertion deletion ( ) USP box 4 USP box 5 USP box 6 catalytic residues residues in interface for proximal Ub moiety key residues responsible for weakened distal Ub binding hydrophobic residues mutated for box 4/5 insertion deletion
Supplementatry Fig 1. Domain structure, biophysical characterisation and electron microscopy of a TD. (a) XTACC3/Maskin and XMAP215/chTOG domain
 XTACC3/Maskin and XMAP215/chTOG domain") Supplementatry Fig 1. Domain structure, biophysical characterisation and electron microscopy of a TD. (a) XTACC3/Maskin and XMAP215/chTOG domain architecture. Various C-terminal fragments were cloned and
Supplementatry Fig 1. Domain structure, biophysical characterisation and electron microscopy of a TD. (a) XTACC3/Maskin and XMAP215/chTOG domain architecture. Various C-terminal fragments were cloned and
Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and
 and") Supplementary Tables Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and their side chain contacting residues in the second chain (B) a Interface Res. in Contacting
Supplementary Tables Supplementary Table 1: List of CH3 domain interface residues in the first chain (A) and their side chain contacting residues in the second chain (B) a Interface Res. in Contacting
Solutions to 7.02 Quiz II 10/27/05
 Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
Nature Structural & Molecular Biology: doi: /nsmb.3018
 Supplementary Figure 1 Validation of genetic complementation assay in Bmal1 / Per2 Luc fibroblasts. (a) Only Bmal1, not Bmal2, rescues circadian rhythms from cells. Cells expressing various Bmal constructs
Supplementary Figure 1 Validation of genetic complementation assay in Bmal1 / Per2 Luc fibroblasts. (a) Only Bmal1, not Bmal2, rescues circadian rhythms from cells. Cells expressing various Bmal constructs
Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid
 Amino acid") Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
Supplementary Figures Suppl. Figure 1: RCC1 sequence and sequence alignments. (a) Amino acid sequence of Drosophila RCC1. Same colors are for Figure 1 with sequence of β-wedge that interacts with Ran in
Supplementary Figure S1 Purification of deubiquitinases HEK293 cells were transfected with the indicated DUB-expressing plasmids.
 Supplementary Figure S1 Purification of deubiquitinases HEK293 cells were transfected with the indicated DUB-expressing plasmids. The cells were harvested 72 h after transfection. FLAG-tagged deubiquitinases
Supplementary Figure S1 Purification of deubiquitinases HEK293 cells were transfected with the indicated DUB-expressing plasmids. The cells were harvested 72 h after transfection. FLAG-tagged deubiquitinases
HEK293T. Fig. 1 in the
 Supplementary Information Supplementary Figure 1 Zinc uptake assay of hzip4 and hzip4-δecd transiently expressed in HEK293T cells. The results of one representative e experiment are shown in Fig. 1 in
Supplementary Information Supplementary Figure 1 Zinc uptake assay of hzip4 and hzip4-δecd transiently expressed in HEK293T cells. The results of one representative e experiment are shown in Fig. 1 in
SUPPLEMENTARY INFORMATION Figures. Supplementary Figure 1 a. Page 1 of 30. Nature Chemical Biology: doi: /nchembio.2528
 SUPPLEMENTARY INFORMATION Figures Supplementary Figure 1 a b c Page 1 of 0 11 Supplementary Figure 1: Biochemical characterisation and binding validation of the reversible USP inhibitor 1. a, Biochemical
SUPPLEMENTARY INFORMATION Figures Supplementary Figure 1 a b c Page 1 of 0 11 Supplementary Figure 1: Biochemical characterisation and binding validation of the reversible USP inhibitor 1. a, Biochemical
Protein analysis. Dr. Mamoun Ahram Summer semester, Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5
 Protein analysis Dr. Mamoun Ahram Summer semester, 2015-2016 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5 Bases of protein separation Proteins can be purified on the basis Solubility
Protein analysis Dr. Mamoun Ahram Summer semester, 2015-2016 Resources This lecture Campbell and Farrell s Biochemistry, Chapters 5 Bases of protein separation Proteins can be purified on the basis Solubility
Supplementary Information
 Supplementary Information Peroxiredoxin-2 and STAT3 form a redox relay for H 2 O 2 signaling Mirko C. Sobotta 1, Willy Liou 1, Sarah Stöcker 1, Deepti Talwar 1, Michael Oehler 1, Thomas Ruppert 2, Annette
Supplementary Information Peroxiredoxin-2 and STAT3 form a redox relay for H 2 O 2 signaling Mirko C. Sobotta 1, Willy Liou 1, Sarah Stöcker 1, Deepti Talwar 1, Michael Oehler 1, Thomas Ruppert 2, Annette
Figure 1: TDP-43 is subject to lysine acetylation within the RNA-binding domain a) QBI-293 cells were transfected with TDP-43 in the presence or
 QBI-293 cells were transfected with TDP-43 in the presence or") Figure 1: TDP-43 is subject to lysine acetylation within the RNA-binding domain a) QBI-293 cells were transfected with TDP-43 in the presence or absence of the acetyltransferase CBP and acetylated TDP-43
Figure 1: TDP-43 is subject to lysine acetylation within the RNA-binding domain a) QBI-293 cells were transfected with TDP-43 in the presence or absence of the acetyltransferase CBP and acetylated TDP-43
SUPPLEMENTAL FIGURE LEGENDS. Figure S1: Homology alignment of DDR2 amino acid sequence. Shown are
 SUPPLEMENTAL FIGURE LEGENDS Figure S1: Homology alignment of DDR2 amino acid sequence. Shown are the amino acid sequences of human DDR2, mouse DDR2 and the closest homologs in zebrafish and C. Elegans.
SUPPLEMENTAL FIGURE LEGENDS Figure S1: Homology alignment of DDR2 amino acid sequence. Shown are the amino acid sequences of human DDR2, mouse DDR2 and the closest homologs in zebrafish and C. Elegans.
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Contents: Supplementary Figure 1. Additional structural and binding data for designed tuim peptides. Supplementary Figure 2. Subcellular localization patterns of designed tuim
SUPPLEMENTARY INFORMATION Contents: Supplementary Figure 1. Additional structural and binding data for designed tuim peptides. Supplementary Figure 2. Subcellular localization patterns of designed tuim
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Domain architecture and conformational states of the decapping complex, as revealed by structural studies. (a) Domain organization of Schizosaccharomyces pombe (Sp) and Saccharomyces
Supplementary Figure 1 Domain architecture and conformational states of the decapping complex, as revealed by structural studies. (a) Domain organization of Schizosaccharomyces pombe (Sp) and Saccharomyces
466 Asn (N) to Ala (A) Generate beta dimer Interface
 to Ala (A) Generate beta dimer Interface") Table S1: Amino acid changes to the HexA α-subunit to convert the dimer interface from α to β and to introduce the putative GM2A binding surface from β- onto the α- subunit Residue position (α-numbering)
Table S1: Amino acid changes to the HexA α-subunit to convert the dimer interface from α to β and to introduce the putative GM2A binding surface from β- onto the α- subunit Residue position (α-numbering)
Ali Yaghi. Tamara Wahbeh. Mamoun Ahram
 28 Ali Yaghi Tamara Wahbeh Mamoun Ahram This sheet is a continuation of protein purification methods. Isoelectric focusing Separation of proteins based on Isoelectric points(charge),and it is a horizontal
28 Ali Yaghi Tamara Wahbeh Mamoun Ahram This sheet is a continuation of protein purification methods. Isoelectric focusing Separation of proteins based on Isoelectric points(charge),and it is a horizontal
human Cdc45 Figure 1c. (c)
") 1 Details of the refined crystallographic model of human Cdc45 and comparison of its active-site region with that of bacterial RecJ. (a) Stereo view of a representative example of the final 2F o -F c electron
1 Details of the refined crystallographic model of human Cdc45 and comparison of its active-site region with that of bacterial RecJ. (a) Stereo view of a representative example of the final 2F o -F c electron
Nature Structural & Molecular Biology: doi: /nsmb.1583
 Acetylation by GCN5 regulates CDC6 phosphorylation in the S-phase of the cell cycle Roberta Paolinelli 1,2, Ramiro Mendoza-Maldonado 2, Anna Cereseto 1 and Mauro Giacca 2 1 Molecular Biology Laboratory,
Acetylation by GCN5 regulates CDC6 phosphorylation in the S-phase of the cell cycle Roberta Paolinelli 1,2, Ramiro Mendoza-Maldonado 2, Anna Cereseto 1 and Mauro Giacca 2 1 Molecular Biology Laboratory,
Supplementary Note 1. Enzymatic properties of the purified Syn BVR
 Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 29 September 2005
 Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 9 September 005 Focus concept Purification of a novel seed storage protein allows sequence analysis and
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Last modified 9 September 005 Focus concept Purification of a novel seed storage protein allows sequence analysis and
1 24 C63 C β- β-
 M40 Signal leaved RS1 Domain 59 110 142 Discoidin Domain 223 1 24 63 219 224 + - β- β- M M e e O O H H His 6 -Tag 250 150 100 75 50 37 * 25 20 Supplementary Figure 1 Purification of wild-type retinoschisin.
M40 Signal leaved RS1 Domain 59 110 142 Discoidin Domain 223 1 24 63 219 224 + - β- β- M M e e O O H H His 6 -Tag 250 150 100 75 50 37 * 25 20 Supplementary Figure 1 Purification of wild-type retinoschisin.
The molecular basis of lysine 48 ubiquitin chain synthesis by Ube2K
 Supplementary Information The molecular basis of lysine 48 ubiquitin chain synthesis by Adam J. Middleton, Catherine L. Day* Department of Biochemistry, Otago School of Medical Sciences, University of
Supplementary Information The molecular basis of lysine 48 ubiquitin chain synthesis by Adam J. Middleton, Catherine L. Day* Department of Biochemistry, Otago School of Medical Sciences, University of
Supplementary Online Material. Structural mimicry in transcription regulation of human RNA polymerase II by the. DNA helicase RECQL5
 Supplementary Online Material Structural mimicry in transcription regulation of human RNA polymerase II by the DNA helicase RECQL5 Susanne A. Kassube, Martin Jinek, Jie Fang, Susan Tsutakawa and Eva Nogales
Supplementary Online Material Structural mimicry in transcription regulation of human RNA polymerase II by the DNA helicase RECQL5 Susanne A. Kassube, Martin Jinek, Jie Fang, Susan Tsutakawa and Eva Nogales
Molecular design principles underlying β-strand swapping. in the adhesive dimerization of cadherins
 Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
Supplementary information for: Molecular design principles underlying β-strand swapping in the adhesive dimerization of cadherins Jeremie Vendome 1,2,3,5, Shoshana Posy 1,2,3,5,6, Xiangshu Jin, 1,3 Fabiana
Supplementary Figure 1 Sequence alignment of representative CbbQ sequences.
 Walker A Pore loop 1 Walker B Supplementary Figure 1 Sequence alignment of representative CbbQ sequences. Amino acid sequences of CbbQ1 and CbbQ2 proteins from selected chemoautotrophic bacteria were aligned
Walker A Pore loop 1 Walker B Supplementary Figure 1 Sequence alignment of representative CbbQ sequences. Amino acid sequences of CbbQ1 and CbbQ2 proteins from selected chemoautotrophic bacteria were aligned
Conformation of the Mineralocorticoid Receptor N- terminal Domain: Evidence for Induced and Stable Structure
 ME-10-0005 Conformation of the Mineralocorticoid Receptor N- terminal Domain: Evidence for Induced and Stable Structure Katharina Fischer 1, Sharon M. Kelly 2, Kate Watt 1, Nicholas C. Price 2 and Iain
ME-10-0005 Conformation of the Mineralocorticoid Receptor N- terminal Domain: Evidence for Induced and Stable Structure Katharina Fischer 1, Sharon M. Kelly 2, Kate Watt 1, Nicholas C. Price 2 and Iain
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Nakajima and Tanoue, http://www.jcb.org/cgi/content/full/jcb.201104118/dc1 Figure S1. DLD-1 cells exhibit the characteristic morphology
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Nakajima and Tanoue, http://www.jcb.org/cgi/content/full/jcb.201104118/dc1 Figure S1. DLD-1 cells exhibit the characteristic morphology
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction
 X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
X-ray structures of fructosyl peptide oxidases revealing residues responsible for gating oxygen access in the oxidative half reaction Tomohisa Shimasaki 1, Hiromi Yoshida 2, Shigehiro Kamitori 2 & Koji
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Structure and mechanism of a canonical poly(adp-ribose) glycohydrolase Mark S. Dunstan 1$, Eva Barkauskaite 2$, Pierre Lafite 3, Claire E. Knezevic 4, Amy Brassington 1, Marijan
SUPPLEMENTARY INFORMATION Structure and mechanism of a canonical poly(adp-ribose) glycohydrolase Mark S. Dunstan 1$, Eva Barkauskaite 2$, Pierre Lafite 3, Claire E. Knezevic 4, Amy Brassington 1, Marijan
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Dynamic Phosphorylation of HP1 Regulates Mitotic Progression in Human Cells Supplementary Figures Supplementary Figure 1. NDR1 interacts with HP1. (a) Immunoprecipitation using
SUPPLEMENTARY INFORMATION Dynamic Phosphorylation of HP1 Regulates Mitotic Progression in Human Cells Supplementary Figures Supplementary Figure 1. NDR1 interacts with HP1. (a) Immunoprecipitation using
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor
 Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Focus concept Purification of a novel seed storage protein allows sequence analysis and determination of the protein
Case 7 A Storage Protein From Seeds of Brassica nigra is a Serine Protease Inhibitor Focus concept Purification of a novel seed storage protein allows sequence analysis and determination of the protein
6-Foot Mini Toober Activity
 Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
Big Idea The interaction between the substrate and enzyme is highly specific. Even a slight change in shape of either the substrate or the enzyme may alter the efficient and selective ability of the enzyme
Kinetics Review. Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets)
") Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Multiple sequence alignments of four Swi2/Snf2 subfamily proteins, ScChd1, SsoRad54 and the RNA helicase Vasa. The sequence alignments of the Swi2/Snf2 subfamily proteins, ScChd1
Supplementary Figure 1 Multiple sequence alignments of four Swi2/Snf2 subfamily proteins, ScChd1, SsoRad54 and the RNA helicase Vasa. The sequence alignments of the Swi2/Snf2 subfamily proteins, ScChd1
Isolation of the recombinant middle and head + middle modules.
 Supplementary Figure 1 Isolation of the recombinant middle and head + middle modules. (a) Scheme illustrating the multi-step purification protocol for the reconstituted middle module. Extract from infected
Supplementary Figure 1 Isolation of the recombinant middle and head + middle modules. (a) Scheme illustrating the multi-step purification protocol for the reconstituted middle module. Extract from infected
Supplementary Figure 1. Electron microscopy of gb-698glyco/1g2 Fab complex. a)
") Supplementary Figure 1. Electron microscopy of gb-698glyco/1g2 Fab complex. a) Representative images of 2D class averages of gb-698glyc bound to 1G2 Fab. Top views of the complex were underrepresented
Supplementary Figure 1. Electron microscopy of gb-698glyco/1g2 Fab complex. a) Representative images of 2D class averages of gb-698glyc bound to 1G2 Fab. Top views of the complex were underrepresented
Figure S1. USP-46 is expressed in several tissues including the nervous system
 Supplemental Figure legends Figure S1. USP-46 is expressed in several tissues including the nervous system Transgenic animals expressing a transcriptional reporter (P::GFP) were imaged using epifluorescence
Supplemental Figure legends Figure S1. USP-46 is expressed in several tissues including the nervous system Transgenic animals expressing a transcriptional reporter (P::GFP) were imaged using epifluorescence
Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1.
 GR24 (μm) 0 20 0 20 GST-ShHTL7 anti-gst His-MAX2 His-COI1 PVDF staining Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1. Pull-down assays using GST-ShHTL7
GR24 (μm) 0 20 0 20 GST-ShHTL7 anti-gst His-MAX2 His-COI1 PVDF staining Supplementary information, Figure S1A ShHTL7 interacted with MAX2 but not another F-box protein COI1. Pull-down assays using GST-ShHTL7
SUPPLEMENTARY INFORMATION
 Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
Supplementary Table 1. Crystallographic statistics CRM1-SNUPN complex Space group P6 4 22 a=b=250.4, c=190.4 Data collection statistics: CRM1-selenomethionine SNUPN MAD data Peak Inflection Remote Native
SUPPLEMENTARY INFORMATION. Supplementary Figures 1-8
 SUPPLEMENTARY INFORMATION Supplementary Figures 1-8 Supplementary Figure 1. TFAM residues contacting the DNA minor groove (A) TFAM contacts on nonspecific DNA. Leu58, Ile81, Asn163, Pro178, and Leu182
SUPPLEMENTARY INFORMATION Supplementary Figures 1-8 Supplementary Figure 1. TFAM residues contacting the DNA minor groove (A) TFAM contacts on nonspecific DNA. Leu58, Ile81, Asn163, Pro178, and Leu182
Supplementary Fig. 1 Identification of Nedd4 as an IRS-2-associated protein in camp-treated FRTL-5 cells.
 Supplementary Fig. 1 Supplementary Fig. 1 Identification of Nedd4 as an IRS-2-associated protein in camp-treated FRTL-5 cells. (a) FRTL-5 cells were treated with 1 mm dibutyryl camp for 24 h, and the lysates
Supplementary Fig. 1 Supplementary Fig. 1 Identification of Nedd4 as an IRS-2-associated protein in camp-treated FRTL-5 cells. (a) FRTL-5 cells were treated with 1 mm dibutyryl camp for 24 h, and the lysates
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature06147 SUPPLEMENTARY INFORMATION Figure S1 The genomic and domain structure of Dscam. The Dscam gene comprises 24 exons, encoding a signal peptide (SP), 10 IgSF domains, 6 fibronectin
doi: 10.1038/nature06147 SUPPLEMENTARY INFORMATION Figure S1 The genomic and domain structure of Dscam. The Dscam gene comprises 24 exons, encoding a signal peptide (SP), 10 IgSF domains, 6 fibronectin
SUPPLEMENTARY INFORMATION. Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly
 SUPPLEMENTARY INFORMATION Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly Dustin J. E. Huard, Kathleen M. Kane and F. Akif Tezcan* Department of Chemistry and Biochemistry,
SUPPLEMENTARY INFORMATION Reengineering Protein Interfaces Yields Copper-Inducible Ferritin Cage Assembly Dustin J. E. Huard, Kathleen M. Kane and F. Akif Tezcan* Department of Chemistry and Biochemistry,
This is the author's accepted version of the manuscript.
 This is the author's accepted version of the manuscript. The definitive version is published in Nature Communications Online Edition: 2015/4/16 (Japan time), doi:10.1038/ncomms7780. The final version published
This is the author's accepted version of the manuscript. The definitive version is published in Nature Communications Online Edition: 2015/4/16 (Japan time), doi:10.1038/ncomms7780. The final version published
Cleavage of tau by asparagine endopeptidase mediates the neurofibrillary pathology in
 Supplementary information Cleavage of tau by asparagine endopeptidase mediates the neurofibrillary pathology in Alzheimer s disease Zhentao Zhang, Mingke Song, Xia Liu, Seong Su Kang, Il-Sun Kwon, Duc
Supplementary information Cleavage of tau by asparagine endopeptidase mediates the neurofibrillary pathology in Alzheimer s disease Zhentao Zhang, Mingke Song, Xia Liu, Seong Su Kang, Il-Sun Kwon, Duc
Stargazin regulates AMPA receptor trafficking through adaptor protein. complexes during long term depression
 Supplementary Information Stargazin regulates AMPA receptor trafficking through adaptor protein complexes during long term depression Shinji Matsuda, Wataru Kakegawa, Timotheus Budisantoso, Toshihiro Nomura,
Supplementary Information Stargazin regulates AMPA receptor trafficking through adaptor protein complexes during long term depression Shinji Matsuda, Wataru Kakegawa, Timotheus Budisantoso, Toshihiro Nomura,
Coleman et al., Supplementary Figure 1
 Coleman et al., Supplementary Figure 1 BrdU Merge G1 Early S Mid S Supplementary Figure 1. Sequential destruction of CRL4 Cdt2 targets during the G1/S transition. HCT116 cells were synchronized by sequential
Coleman et al., Supplementary Figure 1 BrdU Merge G1 Early S Mid S Supplementary Figure 1. Sequential destruction of CRL4 Cdt2 targets during the G1/S transition. HCT116 cells were synchronized by sequential
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11070 Supplementary Figure 1 Purification of FLAG-tagged proteins. a, Purification of FLAG-RNF12 by FLAG-affinity from nuclear extracts of wild-type (WT) and two FLAG- RNF12 transgenic
doi:10.1038/nature11070 Supplementary Figure 1 Purification of FLAG-tagged proteins. a, Purification of FLAG-RNF12 by FLAG-affinity from nuclear extracts of wild-type (WT) and two FLAG- RNF12 transgenic
Electronic Supplementary Information. Simultaneously sensitive detection of multiple DNA glycosylases from lung
 Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 Electronic Supplementary Information Simultaneously sensitive detection of multiple DNA
Electronic Supplementary Material (ESI) for Chemical Science. This journal is The Royal Society of Chemistry 2017 Electronic Supplementary Information Simultaneously sensitive detection of multiple DNA
JCB. Supplemental material THE JOURNAL OF CELL BIOLOGY. Hong et al.,
 Supplemental material JCB Hong et al., http://www.jcb.org/cgi/content/full/jcb.201412127/dc1 THE JOURNAL OF CELL BIOLOGY Figure S1. Analysis of purified proteins by SDS-PAGE and pull-down assays. (A) Coomassie-stained
Supplemental material JCB Hong et al., http://www.jcb.org/cgi/content/full/jcb.201412127/dc1 THE JOURNAL OF CELL BIOLOGY Figure S1. Analysis of purified proteins by SDS-PAGE and pull-down assays. (A) Coomassie-stained
Supplementary Fig. 1
 a FL (1-2266) NL (1-1190) CL (1191-2266) HA-ICE1: - HA-ICE1: - - - FLAG-ICE2: + + + + FLAG-ELL: + + + + + + IP: anti-ha FLAG-ICE2 HA-ICE1-FL HA-ICE1-NL HA-ICE1-CL FLAG-ICE2 b IP: anti-ha FL (1-2266) NL
a FL (1-2266) NL (1-1190) CL (1191-2266) HA-ICE1: - HA-ICE1: - - - FLAG-ICE2: + + + + FLAG-ELL: + + + + + + IP: anti-ha FLAG-ICE2 HA-ICE1-FL HA-ICE1-NL HA-ICE1-CL FLAG-ICE2 b IP: anti-ha FL (1-2266) NL
Aminoacid change in chromophore. PIN3::PIN3-GFP GFP S65 (no change) (5.9) (Kneen et al., 1998)
 (5.9) (Kneen et al., 1998)") Supplemental Table. Table S1. Fluorophore characteristics of used fluorescent proteins, Related to Figure 2 and 3. Transgenic line Fluprescent marker Aminoacid change in chromophore pk(a) Ref. PIN3::PIN3-GFP
Supplemental Table. Table S1. Fluorophore characteristics of used fluorescent proteins, Related to Figure 2 and 3. Transgenic line Fluprescent marker Aminoacid change in chromophore pk(a) Ref. PIN3::PIN3-GFP
supplementary information
 DOI: 10.1038/ncb2116 Figure S1 CDK phosphorylation of EZH2 in cells. (a) Comparison of candidate CDK phosphorylation sites on EZH2 with known CDK substrates by multiple sequence alignments. (b) CDK1 and
DOI: 10.1038/ncb2116 Figure S1 CDK phosphorylation of EZH2 in cells. (a) Comparison of candidate CDK phosphorylation sites on EZH2 with known CDK substrates by multiple sequence alignments. (b) CDK1 and
Purification: Step 1. Lecture 11 Protein and Peptide Chemistry. Cells: Break them open! Crude Extract
 Purification: Step 1 Lecture 11 Protein and Peptide Chemistry Cells: Break them open! Crude Extract Total contents of cell Margaret A. Daugherty Fall 2003 Big Problem: Crude extract is not the natural
Purification: Step 1 Lecture 11 Protein and Peptide Chemistry Cells: Break them open! Crude Extract Total contents of cell Margaret A. Daugherty Fall 2003 Big Problem: Crude extract is not the natural
Purification: Step 1. Protein and Peptide Chemistry. Lecture 11. Big Problem: Crude extract is not the natural environment. Cells: Break them open!
 Lecture 11 Protein and Peptide Chemistry Margaret A. Daugherty Fall 2003 Purification: Step 1 Cells: Break them open! Crude Extract Total contents of cell Big Problem: Crude extract is not the natural
Lecture 11 Protein and Peptide Chemistry Margaret A. Daugherty Fall 2003 Purification: Step 1 Cells: Break them open! Crude Extract Total contents of cell Big Problem: Crude extract is not the natural
Supplementary Figure S1. Binding of HSA mutants to hfcrn. (a) The levels of titrated amounts of HSA
 The levels of titrated amounts of HSA") Supplementary Figure S1. Binding of HSA mutants to hfcrn. (a) The levels of titrated amounts of HSA variants (5.0-0.002 μg/ml) directly coated in the wells at ph 6.0 were controlled using a horseradish
Supplementary Figure S1. Binding of HSA mutants to hfcrn. (a) The levels of titrated amounts of HSA variants (5.0-0.002 μg/ml) directly coated in the wells at ph 6.0 were controlled using a horseradish
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary figures Supplementary Figure 1: Suv39h1, but not Suv39h2, promotes HP1α sumoylation in vivo. In vivo HP1α sumoylation assay. Top: experimental scheme. Middle: we
SUPPLEMENTARY INFORMATION Supplementary figures Supplementary Figure 1: Suv39h1, but not Suv39h2, promotes HP1α sumoylation in vivo. In vivo HP1α sumoylation assay. Top: experimental scheme. Middle: we
T H E J O U R N A L O F C E L L B I O L O G Y
 T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Han et al., http://www.jcb.org/cgi/content/full/jcb.201311007/dc1 Figure S1. SIVA1 interacts with PCNA. (A) HEK293T cells were transiently
T H E J O U R N A L O F C E L L B I O L O G Y Supplemental material Han et al., http://www.jcb.org/cgi/content/full/jcb.201311007/dc1 Figure S1. SIVA1 interacts with PCNA. (A) HEK293T cells were transiently
Supplementary Fig. S1. SAMHD1c has a more potent dntpase activity than. SAMHD1c. Purified recombinant SAMHD1c and SAMHD1c proteins (with
 Supplementary Fig. S1. SAMHD1c has a more potent dntpase activity than SAMHD1c. Purified recombinant SAMHD1c and SAMHD1c proteins (with concentration of 800nM) were incubated with 1mM dgtp for the indicated
Supplementary Fig. S1. SAMHD1c has a more potent dntpase activity than SAMHD1c. Purified recombinant SAMHD1c and SAMHD1c proteins (with concentration of 800nM) were incubated with 1mM dgtp for the indicated
SUPPLEMENTAL MATERIAL BIOCHEMICAL AND STRUCTURAL STUDIES ON THE M. TUBERCULOSIS O 6 -METHYLGUANINE METHYLTRANSFERASE AND MUTATED VARIANTS
 SUPPLEMENTAL MATERIAL BIOCHEMICAL AND STRUCTURAL STUDIES ON THE M. TUBERCULOSIS O 6 -METHYLGUANINE METHYLTRANSFERASE AND MUTATED VARIANTS Riccardo Miggiano 1, Valentina Casazza 1, Silvia Garavaglia 1,
SUPPLEMENTAL MATERIAL BIOCHEMICAL AND STRUCTURAL STUDIES ON THE M. TUBERCULOSIS O 6 -METHYLGUANINE METHYLTRANSFERASE AND MUTATED VARIANTS Riccardo Miggiano 1, Valentina Casazza 1, Silvia Garavaglia 1,
The Skap-hom Dimerization and PH Domains Comprise
 Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
Molecular Cell, Volume 32 Supplemental Data The Skap-hom Dimerization and PH Domains Comprise a 3 -Phosphoinositide-Gated Molecular Switch Kenneth D. Swanson, Yong Tang, Derek F. Ceccarelli, Florence Poy,
Supplementary Figure 1 Collision-induced dissociation (CID) mass spectra of peptides from PPK1, PPK2, PPK3 and PPK4 respectively.
 mass spectra of peptides from PPK1, PPK2, PPK3 and PPK4 respectively.") Supplementary Figure 1 lision-induced dissociation (CID) mass spectra of peptides from PPK1, PPK, PPK3 and PPK respectively. % of nuclei with signal / field a 5 c ppif3:gus pppk1:gus 0 35 30 5 0 15 10
Supplementary Figure 1 lision-induced dissociation (CID) mass spectra of peptides from PPK1, PPK, PPK3 and PPK respectively. % of nuclei with signal / field a 5 c ppif3:gus pppk1:gus 0 35 30 5 0 15 10
Extracting Pure Proteins from Cells
 Extracting Pure Proteins from Cells 0 Purification techniques focus mainly on size & charge 0 The first step is homogenization (grinding, Potter Elvejhem homogenizer, sonication, freezing and thawing,
Extracting Pure Proteins from Cells 0 Purification techniques focus mainly on size & charge 0 The first step is homogenization (grinding, Potter Elvejhem homogenizer, sonication, freezing and thawing,
ARBRE-P4EU Consensus Protein Quality Guidelines for Biophysical and Biochemical Studies Minimal information to provide
 ARBRE-P4EU Consensus Protein Quality Guidelines for Biophysical and Biochemical Studies Minimal information to provide Protein name and full primary structure, by providing a NCBI (or UniProt) accession
ARBRE-P4EU Consensus Protein Quality Guidelines for Biophysical and Biochemical Studies Minimal information to provide Protein name and full primary structure, by providing a NCBI (or UniProt) accession
Nature Structural & Molecular Biology: doi: /nsmb.3428
 Supplementary Figure 1 Biochemical characterization of the monou and oligou activity switch of TUT4(7). (a) Mouse TUT4 and human TUT7 were assayed for monou and Lin28-dependent oligou addition activities
Supplementary Figure 1 Biochemical characterization of the monou and oligou activity switch of TUT4(7). (a) Mouse TUT4 and human TUT7 were assayed for monou and Lin28-dependent oligou addition activities
Chapter 5: Proteins: Primary Structure
 Instant download and all chapters Test Bank Fundamentals of Biochemistry Life at the Molecular Level 4th Edition Donald Voet https://testbanklab.com/download/test-bank-fundamentals-biochemistry-life-molecular-level-
Instant download and all chapters Test Bank Fundamentals of Biochemistry Life at the Molecular Level 4th Edition Donald Voet https://testbanklab.com/download/test-bank-fundamentals-biochemistry-life-molecular-level-
Fig. S1. CrgA intracellular levels in M. tuberculosis. Ten and twenty micrograms of
 Supplementary data Fig. S1. CrgA intracellular levels in M. tuberculosis. Ten and twenty micrograms of cell free protein lysates from WT M. tuberculosis (Rv) together with various known concentrations
Supplementary data Fig. S1. CrgA intracellular levels in M. tuberculosis. Ten and twenty micrograms of cell free protein lysates from WT M. tuberculosis (Rv) together with various known concentrations
BC 367, Exam 2 November 13, Part I. Multiple Choice (3 pts each)- Please circle the single best answer.
- Please circle the single best answer.") Name BC 367, Exam 2 November 13, 2008 Part I. Multiple Choice (3 pts each)- Please circle the single best answer. 1. The enzyme pyruvate dehydrogenase catalyzes the following reaction. What kind of enzyme
Name BC 367, Exam 2 November 13, 2008 Part I. Multiple Choice (3 pts each)- Please circle the single best answer. 1. The enzyme pyruvate dehydrogenase catalyzes the following reaction. What kind of enzyme
Supplementary Figure 1 a
 3 min PMA 45 min PMA AnnexinV-FITC Supplementary Figure 1 5 min PMA 15 min PMA a 9 min PMA 12 min PMA 5 min FGF7 15 min FGF7 3 min FGF7 6 min FGF7 9 min FGF7 12 min FGF7 5 min control 3 min control 6 min
3 min PMA 45 min PMA AnnexinV-FITC Supplementary Figure 1 5 min PMA 15 min PMA a 9 min PMA 12 min PMA 5 min FGF7 15 min FGF7 3 min FGF7 6 min FGF7 9 min FGF7 12 min FGF7 5 min control 3 min control 6 min
Supplementary Information
 Supplementary Information Supplemental Figure 1. VVD-III purifies in a reduced state. (a) The cell pellet of VVD-III (VVD 36 C108A:M135I:M165I) is green compared to VVD-I (wild type VVD 36) due to the
Supplementary Information Supplemental Figure 1. VVD-III purifies in a reduced state. (a) The cell pellet of VVD-III (VVD 36 C108A:M135I:M165I) is green compared to VVD-I (wild type VVD 36) due to the
SUPPLEMENTARY INFORMATION
 doi: 10.1038/nature06721 SUPPLEMENTARY INFORMATION. Supplemental Figure Legends Supplemental Figure 1 The distribution of hatx-1[82q] in Cos7 cells. Cos7 cells are co-transfected with hatx-1[82q]-gfp (green)
doi: 10.1038/nature06721 SUPPLEMENTARY INFORMATION. Supplemental Figure Legends Supplemental Figure 1 The distribution of hatx-1[82q] in Cos7 cells. Cos7 cells are co-transfected with hatx-1[82q]-gfp (green)
Supplementary Information for. Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction
 Supplementary Information for Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction Takamasa Teramoto, Yukari Fujikawa, Yoshirou Kawaguchi, Katsuhisa
Supplementary Information for Structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction Takamasa Teramoto, Yukari Fujikawa, Yoshirou Kawaguchi, Katsuhisa
Supplementary methods Shoc2 In Vitro Ubiquitination Assay
 Supplementary methods Shoc2 In Vitro Ubiquitination Assay 35 S-labelled Shoc2 was prepared using a TNT quick Coupled transcription/ translation System (Promega) as recommended by manufacturer. For the
Supplementary methods Shoc2 In Vitro Ubiquitination Assay 35 S-labelled Shoc2 was prepared using a TNT quick Coupled transcription/ translation System (Promega) as recommended by manufacturer. For the
Supplementary Figure 1 PZA inhibits root hair formation as well as cell elongation in the maturation zone of eto1-2 roots. (A) The PI staining of the
 The PI staining of the") Supplementary Figure 1 PZA inhibits root hair formation as well as cell elongation in the maturation zone of eto1-2 roots. (A) The PI staining of the roots of three-day-old etiolated seedlings of Col-0
Supplementary Figure 1 PZA inhibits root hair formation as well as cell elongation in the maturation zone of eto1-2 roots. (A) The PI staining of the roots of three-day-old etiolated seedlings of Col-0
Introduction to Protein Purification
 Introduction to Protein Purification 1 Day 1) Introduction to Protein Purification. Input for Purification Protocol Development - Guidelines for Protein Purification Day 2) Sample Preparation before Chromatography
Introduction to Protein Purification 1 Day 1) Introduction to Protein Purification. Input for Purification Protocol Development - Guidelines for Protein Purification Day 2) Sample Preparation before Chromatography
Hossain_Supplemental Figure 1
 Hossain_Supplemental Figure 1 GFP-PACT GFP-PACT Motif I GFP-PACT Motif II A. MG132 (1µM) GFP Tubulin GFP-PACT Pericentrin GFP-PACT GFP-PACT Pericentrin Fig. S1. Expression and localization of Orc1 PACT
Hossain_Supplemental Figure 1 GFP-PACT GFP-PACT Motif I GFP-PACT Motif II A. MG132 (1µM) GFP Tubulin GFP-PACT Pericentrin GFP-PACT GFP-PACT Pericentrin Fig. S1. Expression and localization of Orc1 PACT
SUPPLEMENTAL MATERIAL. Supplemental Methods:
 SUPPLEMENTAL MATERIAL Supplemental Methods: Immunoprecipitation- As we described but with some modifications [22]. As part of another ongoing project, lysate from human umbilical vein endothelial cells
SUPPLEMENTAL MATERIAL Supplemental Methods: Immunoprecipitation- As we described but with some modifications [22]. As part of another ongoing project, lysate from human umbilical vein endothelial cells
Supporting Information Contents
 Supporting Information Choy Theng Loh, Kiyoshi Ozawa, Kellie L. Tuck, Nicholas Barlow, Thomas Huber, Gottfried Otting, and Bim Graham Lanthanide tags for site-specific ligation to an unnatural amino acid
Supporting Information Choy Theng Loh, Kiyoshi Ozawa, Kellie L. Tuck, Nicholas Barlow, Thomas Huber, Gottfried Otting, and Bim Graham Lanthanide tags for site-specific ligation to an unnatural amino acid
Programme Good morning and summary of last week Levels of Protein Structure - I Levels of Protein Structure - II
 Programme 8.00-8.10 Good morning and summary of last week 8.10-8.30 Levels of Protein Structure - I 8.30-9.00 Levels of Protein Structure - II 9.00-9.15 Break 9.15-11.15 Exercise: Building a protein model
Programme 8.00-8.10 Good morning and summary of last week 8.10-8.30 Levels of Protein Structure - I 8.30-9.00 Levels of Protein Structure - II 9.00-9.15 Break 9.15-11.15 Exercise: Building a protein model
Supplementary Figure 1. Nature Structural & Molecular Biology: doi: /nsmb.3494
 Supplementary Figure 1 Pol structure-function analysis (a) Inactivating polymerase and helicase mutations do not alter the stability of Pol. Flag epitopes were introduced using CRISPR/Cas9 gene targeting
Supplementary Figure 1 Pol structure-function analysis (a) Inactivating polymerase and helicase mutations do not alter the stability of Pol. Flag epitopes were introduced using CRISPR/Cas9 gene targeting
Supplementary Figure 1.
 Supplementary Figure 1. Assessment of quaternary structure of soluble RSV F proteins. Soluble variants of F proteins from A2 and B1 RSV strains were expressed in HEK293 cells. The cell culture supernatants
Supplementary Figure 1. Assessment of quaternary structure of soluble RSV F proteins. Soluble variants of F proteins from A2 and B1 RSV strains were expressed in HEK293 cells. The cell culture supernatants
Expanded View Figures
 EMO reports rystal structure of Mis18 Yippee-like domain Lakxmi Subramanian et al Expanded View Figures Figure EV1. Structural characterization of the N-terminal Yippee-like globular domain of spmis18.
EMO reports rystal structure of Mis18 Yippee-like domain Lakxmi Subramanian et al Expanded View Figures Figure EV1. Structural characterization of the N-terminal Yippee-like globular domain of spmis18.
Prediction of Protein-Protein Binding Sites and Epitope Mapping. Copyright 2017 Chemical Computing Group ULC All Rights Reserved.
 Prediction of Protein-Protein Binding Sites and Epitope Mapping Epitope Mapping Antibodies interact with antigens at epitopes Epitope is collection residues on antigen Continuous (sequence) or non-continuous
Prediction of Protein-Protein Binding Sites and Epitope Mapping Epitope Mapping Antibodies interact with antigens at epitopes Epitope is collection residues on antigen Continuous (sequence) or non-continuous
pt7ht vector and over-expressed in E. coli as inclusion bodies. Cells were lysed in 6 M
 Supplementary Methods MIG6 production, purification, inhibition, and kinase assays MIG6 segment 1 (30mer, residues 334 364) peptide was synthesized using standard solid-phase peptide synthesis as described
Supplementary Methods MIG6 production, purification, inhibition, and kinase assays MIG6 segment 1 (30mer, residues 334 364) peptide was synthesized using standard solid-phase peptide synthesis as described
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature10324 Fig. S1: Two dimensional IEF/SDS-PAGE/Western blot analysis of RBC lysate crosslinked with 4 mm BS 3. The blot was probed with αsyn antibody C20. Fig. S2: SDS-PAGE/silver stain
doi:10.1038/nature10324 Fig. S1: Two dimensional IEF/SDS-PAGE/Western blot analysis of RBC lysate crosslinked with 4 mm BS 3. The blot was probed with αsyn antibody C20. Fig. S2: SDS-PAGE/silver stain
Supplementary information
 Supplementary information The E3 ligase RNF8 regulates KU80 removal and NHEJ repair Lin Feng 1, Junjie Chen 1 1 Department of Experimental Radiation Oncology, The University of Texas M. D. Anderson Cancer
Supplementary information The E3 ligase RNF8 regulates KU80 removal and NHEJ repair Lin Feng 1, Junjie Chen 1 1 Department of Experimental Radiation Oncology, The University of Texas M. D. Anderson Cancer
Supplementary Results Supplementary Table 1. P1 and P2 enrichment scores for wild-type subtiligase.
 Supplementary Results Supplementary Table 1. P1 and P2 enrichment scores for wild-type subtiligase. Supplementary Table 2. Masses of subtiligase mutants measured by LC-MS. Supplementary Table 2 (cont d).
Supplementary Results Supplementary Table 1. P1 and P2 enrichment scores for wild-type subtiligase. Supplementary Table 2. Masses of subtiligase mutants measured by LC-MS. Supplementary Table 2 (cont d).
Engineering splicing factors with designed specificities
 nature methods Engineering splicing factors with designed specificities Yang Wang, Cheom-Gil Cheong, Traci M Tanaka Hall & Zefeng Wang Supplementary figures and text: Supplementary Figure 1 Supplementary
nature methods Engineering splicing factors with designed specificities Yang Wang, Cheom-Gil Cheong, Traci M Tanaka Hall & Zefeng Wang Supplementary figures and text: Supplementary Figure 1 Supplementary
11/22/13. Proteomics, functional genomics, and systems biology. Biosciences 741: Genomics Fall, 2013 Week 11
 Proteomics, functional genomics, and systems biology Biosciences 741: Genomics Fall, 2013 Week 11 1 Figure 6.1 The future of genomics Functional Genomics The field of functional genomics represents the
Proteomics, functional genomics, and systems biology Biosciences 741: Genomics Fall, 2013 Week 11 1 Figure 6.1 The future of genomics Functional Genomics The field of functional genomics represents the
T H E J O U R N A L O F C E L L B I O L O G Y
 Supplemental material Thompson et al., http://www.jcb.org/cgi/content/full/jcb.200909067/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Modification-specific antibodies do not detect unmodified
Supplemental material Thompson et al., http://www.jcb.org/cgi/content/full/jcb.200909067/dc1 T H E J O U R N A L O F C E L L B I O L O G Y Figure S1. Modification-specific antibodies do not detect unmodified
Homology Modelling. Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen
 Homology Modelling Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen Why are Protein Structures so Interesting? They provide a detailed picture of interesting biological features,
Homology Modelling Thomas Holberg Blicher NNF Center for Protein Research University of Copenhagen Why are Protein Structures so Interesting? They provide a detailed picture of interesting biological features,
Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein
 Cell Wall Recycling Protein") Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
Paper Presentation PLoS ONE 2011 Structure and Function of the First Full-Length Murein Peptide Ligase (Mpl) Cell Wall Recycling Protein Debanu Das, Mireille Herve, Julie Feuerhelm, etc. and Dominique
Supplemental Materials and Methods
 Supplemental Materials and Methods Co-immunoprecipitation (Co-IP) assay Cells were lysed with NETN buffer (20 mm Tris-HCl, ph 8.0, 0 mm NaCl, 1 mm EDT, 0.5% Nonidet P-40) containing 50 mm β-glycerophosphate,
Supplemental Materials and Methods Co-immunoprecipitation (Co-IP) assay Cells were lysed with NETN buffer (20 mm Tris-HCl, ph 8.0, 0 mm NaCl, 1 mm EDT, 0.5% Nonidet P-40) containing 50 mm β-glycerophosphate,
Supplementary Fig. 1. Schematic structure of TRAIP and RAP80. The prey line below TRAIP indicates bait and the two lines above RAP80 highlight the
 Supplementary Fig. 1. Schematic structure of TRAIP and RAP80. The prey line below TRAIP indicates bait and the two lines above RAP80 highlight the prey clones identified in the yeast two hybrid screen.
Supplementary Fig. 1. Schematic structure of TRAIP and RAP80. The prey line below TRAIP indicates bait and the two lines above RAP80 highlight the prey clones identified in the yeast two hybrid screen.
Notes to accompany the slidecast on theory of SDS PAGE and Western blotting
 S317 Biological science: from genes to species Notes to accompany the slidecast on theory of SDS PAGE and Western blotting SDS PAGE SDS PAGE is a standard technique for determining the molecular size of
S317 Biological science: from genes to species Notes to accompany the slidecast on theory of SDS PAGE and Western blotting SDS PAGE SDS PAGE is a standard technique for determining the molecular size of
Supplementary Figure 1 Two distinct conformational states of the HNH domain in crystal structures. a
 Supplementary Figure 1 Two distinct conformational states of the HNH domain in crystal structures. a HNH-state 1 in PDB 4OO8, in which the distance from the C atom of the HNH catalytic residue 840 to the
Supplementary Figure 1 Two distinct conformational states of the HNH domain in crystal structures. a HNH-state 1 in PDB 4OO8, in which the distance from the C atom of the HNH catalytic residue 840 to the
42 fl organelles = 34.5 fl (1) 3.5X X 0.93 = 78,000 (2)
 3.5X X 0.93 = 78,000 (2)") SUPPLEMENTAL DATA Supplementary Experimental Procedures Fluorescence Microscopy - A Zeiss Axiovert 200M microscope equipped with a Zeiss 100x Plan- Apochromat (1.40 NA) DIC objective and Hamamatsu Orca
SUPPLEMENTAL DATA Supplementary Experimental Procedures Fluorescence Microscopy - A Zeiss Axiovert 200M microscope equipped with a Zeiss 100x Plan- Apochromat (1.40 NA) DIC objective and Hamamatsu Orca
Lecture 5: 8/31. CHAPTER 5 Techniques in Protein Biochemistry
 Lecture 5: 8/31 CHAPTER 5 Techniques in Protein Biochemistry Chapter 5 Outline The proteome is the entire set of proteins expressed and modified by a cell under a particular set of biochemical conditions.
Lecture 5: 8/31 CHAPTER 5 Techniques in Protein Biochemistry Chapter 5 Outline The proteome is the entire set of proteins expressed and modified by a cell under a particular set of biochemical conditions.
Supplementary Figure 1. α-synuclein is truncated in PD and LBD brains. Nature Structural & Molecular Biology: doi: /nsmb.
 Supplementary Figure 1 α-synuclein is truncated in PD and LBD brains. (a) Specificity of anti-n103 antibody. Anti-N103 antibody was coated on an ELISA plate and different concentrations of full-length
Supplementary Figure 1 α-synuclein is truncated in PD and LBD brains. (a) Specificity of anti-n103 antibody. Anti-N103 antibody was coated on an ELISA plate and different concentrations of full-length
Ch Biophysical Chemistry
 Ch 247.53. Biophysical Chemistry Nina Rosario L. Rojas 2012-2013 sem 1 Review of Protein Structure Why structure? Primary, secondary, tertiary structure Disulfide bonds scheme 2 STRUCTURE- REGULAR STRUCTURE
Ch 247.53. Biophysical Chemistry Nina Rosario L. Rojas 2012-2013 sem 1 Review of Protein Structure Why structure? Primary, secondary, tertiary structure Disulfide bonds scheme 2 STRUCTURE- REGULAR STRUCTURE