The interplay between cardiac progenitor cells and their microenvironment Mauretti, A.
|
|
|
- Noah Booker
- 6 years ago
- Views:
Transcription
1 The interplay between cardiac progenitor cells and their microenvironment Mauretti, A. Accepted/In press: 22/05/2018 Document Version Publisher s PDF, also known as Version of Record (includes final page, issue and volume numbers) Please check the document version of this publication: A submitted manuscript is the author's version of the article upon submission and before peer-review. There can be important differences between the submitted version and the official published version of record. People interested in the research are advised to contact the author for the final version of the publication, or visit the DOI to the publisher's website. The final author version and the galley proof are versions of the publication after peer review. The final published version features the final layout of the paper including the volume, issue and page numbers. Link to publication Citation for published version (APA): Mauretti, A. (2018). The interplay between cardiac progenitor cells and their microenvironment Eindhoven: Technische Universiteit Eindhoven General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. Users may download and print one copy of any publication from the public portal for the purpose of private study or research. You may not further distribute the material or use it for any profit-making activity or commercial gain You may freely distribute the URL identifying the publication in the public portal? Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Download date: 28. Apr. 2018
2 The interplay between cardiac progenitor cells and their microenvironment Arianna Mauretti
3 A catalogue record is available from the Eindhoven University of Technology Library ISBN: Copyright 2018 by A. Mauretti All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronically, mechanically, by print, photo print, recording or any other means without prior written permission by the author. Cover design by Arianna Mauretti and Aurora Mauretti Financial support by the Dutch Heart Association for the publication of this thesis is gratefully acknowledged. The work of this thesis was supported by the People Programme (Marie Curie Actions) of the European Union s Seventh Framework Programme [FP7/ /]; REA [grant agreement number ]. Printed by Ipskamp Printing, Enschede
4 The interplay between cardiac progenitor cells and their microenvironment PROEFSCHRIFT ter verkrijging van de graad van doctor aan de Technische Universiteit Eindhoven, op gezag van de rector magnificus prof.dr.ir. F.P.T. Baaijens, voor een commissie aangewezen door het College voor Promoties, in het openbaar te verdedigen op dinsdag 22 mei 2018 om uur door Arianna Mauretti geboren te Rome, Italië
5 Dit proefschrift is goedgekeurd door de promotoren en de samenstelling van de promotiecommissie is als volgt: Voorzitter: prof.dr. P.A.J. Hilbers 1 e promotor: prof.dr. C.V.C. Bouten 2 e promotor: prof.dr. C. Sahlgren copromotor: dr. N.A.M. Bax leden: prof.dr.ir. L. Brunsveld prof.dr. P.Y.W. Dankers prof.dr. K. Schenke-Layland (Eberhard-Karls-University Tübingen) adviseur: prof.dr. M.J.T.H. Goumans (Leiden University Medical Center) Het onderzoek dat in dit proefschrift wordt beschreven is uitgevoerd in overeenstemming met de TU/e Gedragscode Wetenschapsbeoefening.
6 A mamma e papà Cause baby there ain t no mountain high enough, ain t no valley low enough, ain t no river wide enough, to keep me from getting to you (Marvin Gaye and Tammi Terrell, 1967)
7
8 CONTENTS CONTENTS... VII CHAPTER INTRODUCTION...3 THE CARDIAC RESIDENT PROGENITOR CELL...6 THE CPC MICROENVIRONMENT...8 RATIONALE, AIM AND OUTLINE OF THE THESIS CHAPTER INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION CONCLUSIONS APPENDIX TO CHAPTER CHAPTER INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION APPENDIX TO CHAPTER CHAPTER INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION APPENDIX TO CHAPTER CHAPTER INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION CONCLUSIONS APPENDIX TO CHAPTER CHAPTER INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION AND OUTLOOK vii
9 APPENDIX TO CHAPTER CHAPTER INTRODUCTION MAIN FINDINGS CPC-MICROENVIRONMENT INTERPLAY IMPLICATIONS FOR ENGINEERED MICROENVIRONMENTS FUTURE PERSPECTIVES OF CARDIAC THERAPIES CONCLUSION BIBLIOGRAPHY SUMMARY CURRICULUM VITAE LIST OF PUBLICATIONS ACKNOWLEDGMENTS viii
10 CHAPTER 1 General introduction This chapter is based on: Mauretti A.*, Spaans S.*, Bax N.A.M.*, Sahlgren C., Bouten C.V.C., Cardiac progenitor cells and the interplay with their microenvironment, Stem Cells International (2017). *authors contributed equally
11 Abstract The extracellular microenvironment plays a crucial role in the behavior of stem and progenitor cells. In the heart, cardiac progenitor cells (CPCs) reside in specific niches, which are subject to drastic changes during cardiac disease or in response to injuries, such as a myocardial infarction. Currently, there is a lack of knowledge of niche composition in the heart, and especially of the interplay of CPCs with the niche components in health and disease. Insight into the complex interactions between the niche and CPCs can be used to promote the regenerative potential of these cells. In this chapter, the interplay of CPCs with the key elements of their niche is discussed, with a focus on the interactions between CPCs and supporting cells, extracellular matrix, mechanical stimuli and soluble factors. Finally, novel approaches are described to modulate the CPC niche, that may represent the next step in recreating an optimal CPC microenvironment and thereby improve the CPC regeneration capacity. 2
12 Introduction Cardiac tissue is composed of contractile and supportive cells, surrounded by extracellular matrix (ECM), intertwined with nervous and vascular networks. An ischemic event, such as a myocardial infarction (MI), not only induces cell death, but also affects the tissue structure and composition. This can eventually lead to loss of cardiac function due to changes in the key players of the cardiac microenvironment: 1) the stem/progenitor cells and supporting cells, 2) the extracellular matrix (ECM) proteins, 3) the mechanical environment of the cells, provided not only by the matrix, but also by e.g. the cyclic strain provided by the beating heart, and 4) the soluble components, such as oxygen and cytokines (Figure 1A). In this chapter we omit to describe the vascular components, innervation, and electrical conduction, as these are extensively reviewed elsewhere 1 3, although their derivatives, such as oxygen gradients and cyclic strain, are included. The myocardium shows limited self-renewal; nevertheless, the notion of the heart as a terminally differentiated organ, incapable of regenerating after injury, has been challenged in the last decade 4,5. There is ongoing debate on whether cardiac regeneration is to be attributed to de-differentiation and proliferation of cardiomyocytes 6,7 or to differentiation of cardiac stem or progenitor cells 8 10, which makes it difficult to identify the ideal therapeutic target. Nevertheless, the existence of resident cardiac progenitor cells (CPCs) in the heart and their relevance for cardiac regeneration have been demonstrated by several studies CPCs have emerged as a promising candidate for cardiac regeneration, due to their differentiation potential 10,14 and the ability to produce and remodel ECM proteins 15. Moreover, after acute MI the number of CPCs in the adult seems to increase, and differentiation into the cardiac lineages takes place 6. However, the post-mi microenvironment can affect CPC behavior: in chronic infarcts CPCs are characterized by decreased telomerase activity, leading to impaired cell division and cellular senescence, as well as increased CPC apoptosis 6. The traditional cell therapy approach to treat a MI entails isolation of CPCs, their expansion in vitro and transplantation into the infarcted area 16. Despite the immediate benefits on cardiac function, this treatment has shown limited improvement on the long term 17 19, mainly due to low cell survival and engraftment in the host tissue 20. In fact, a MI creates a hostile environment for the injected progenitor cells, due to the inflammatory response and tissue alterations, such as scar tissue formation, triggered by the cardiac injury, as extensively described elsewhere (Figure 1B). 3
 The simplified representation shows some of the key players of the healthy CPC niche: 1) cellular elements (CPCs and supporting cells: cardiomyocytes, endothelial cells, smooth muscle cells,")
13 Figure 1. The resident cardiac progenitor cell microenvironment. A) The simplified representation shows some of the key players of the healthy CPC niche: 1) cellular elements (CPCs and supporting cells: cardiomyocytes, endothelial cells, smooth muscle cells, stromal and immune cells), and cell-cell interactions such as signaling via Notch; 2) extracellular matrix (ECM), from which cells can receive mechanical signals via integrins; 3) mechanical stimuli, such as the cyclic strain provided by the beating heart; and 4) soluble components, such as cytokines, oxygen gradients and growth factors. B) Simplified representation of the infarcted heart, where the microenvironment is altered and the niche components modified: 1) cardiomyocyte death, infiltration of stromal cells (myofibroblasts) and immune cells; 2) excessive and disordered formation of ECM; 3) increased ECM stiffness and thus altered mechanical behavior; 4) increased secretion of growth factors and cytokines. In the adult, stem or progenitor cells reside in specific microenvironments, referred to as niches, that protect stem cells and regulate their fate and functions Stem cell niches are stored in specific anatomical compartments, located in tissue areas that are shielded from external damaging stimuli 23,26. In the adult mouse heart, putative progenitor cell niches have been identified in the atria, basemid region and apex 27,28. To date, however, the cardiac progenitor cell niche is still largely uncharacterized, and most studies have been performed on mouse models ; the human CPC niches are therefore largely unknown. For cell therapy, cells are isolated from their resident niche and expanded in an in vitro niche, prior to transplantation into the diseased niche of the infarcted heart tissue (Figure 2A). An alternative to cell therapy is to promote the regeneration provided by endogenous CPCs, for instance by promoting the migration of CPCs to the damaged cardiac area (and to the diseased niche ), as well as their proliferation and differentiation (Figure 2B). Another potential approach is to generate new, or engineered, microenvironments for the cells, in order to re-create optimal conditions to enhance their regenerative potential. Currently, there is a lack of knowledge of the composition of the resident, in vitro and diseased niches. Particularly, the interplay between CPCs and these niches (and the niche components) is largely unknown. 4

14 Understanding the influence of distinct elements of the microenvironment on CPC function is crucial, as it can provide information on how to elicit a regenerative response of CPCs by creating a regenerative niche. Therefore, here the key elements of the potential CPC microenvironments are highlighted, and the interplay between CPCs and the niche components is discussed. Improved knowledge on the CPC niches and the CPC-niche interactions will enhance our insight into CPC behavior and the influence of the niche on CPC regenerative capacity, which can ultimately help modulate the microenvironment to promote the regenerative potential of CPCs. In the last part of this chapter we provide an overview on recent advances in the field of engineered cardiac microenvironments, which can represent the next step in exploring and modulating the CPC niche and CPC behavior for cardiac repair. Figure 2. The CPC microenvironments or niches. For therapeutic application, CPCs can be isolated from their resident niche and A) cultured in an in vitro niche, prior to transplantation into the infarcted heart, or B) the local microenvironment can be modulated in order to recruit CPCs to the injured area. The aim of both these approaches is to regenerate the myocardium thanks to CPC proliferation and differentiation into cardiomyocytes. 5
15 The cardiac resident progenitor cell The presence of CPCs in the fetal and adult heart in mammals (including humans) has been extensively described (reviewed by 11 ). Since their discovery in , a number of cardiac progenitor cell population has been identified (based on cell behavior, membrane markers, or method of isolation): cardiac side population, c-kit +, Sca-1 +, Isl-1 +, cardiospheres (CSps) and cardiosphere-derived cells (CDCs). Although these cell populations present differences, they share some key characteristics and functions (an overview is provided in Table 1). CPCs reside throughout the heart in both embryonic and adult stage; they are self-renewing and multipotent, i.e. able to differentiate in at least three of the four cardiac cell types (cardiomyocytes, endothelial cells, smooth muscle cells and fibroblasts). Furthermore, some of the CPC populations were shown to be activated during cardiac injury, and to have regenerative potential, proven by the beneficial effects on cardiac function following transplantation of these cells into the diseased heart. The characteristics and regenerative potential of the CPC populations are summarized in Table 1 (adapted from 31 ). Some have suggested that all the described CPCs might represent the same cell population, and that the differences lie in the identification method or their differentiation stage 32. The work of this thesis is performed on human CPCs isolated using their reactivity for the murine membrane marker Sca-1 +. While these cells were originally defined as cardiomyocyte progenitor cells (CMPCs), due to the ongoing debate on the origin and identity of the several CPC populations, in this thesis we prefer to simply refer to them as CPCs. Below, the main characteristics and regenerative potential of this cell source are described. Sca-1 + cardiac progenitor cells Resident Sca-1 + cells are found in fetal and adult mouse and human hearts, in the atria, the intra-atrial septum, the myocardium and in the epicardium 10,12, Human Sca1 + cells harbor telomerase activity, which characterizes their proliferative potential and their ability to selfrenew 34,36,37. Among the several CPC populations, (murine) Sca-1 + cells have shown the highest correlation with cardiomyocytes 38. Culture-expanded Sca-1 + cells can be differentiated into cardiomyocytes in the presence of oxytocin or 5-azacytidine treatment 13,36,37 and their cardiac differentiation potential is enhanced by addition of transforming growth factor-beta (TGF-β) 36,37. Next to cardiomyocytes, Sca-1 + cells can differentiate into endothelial cells and smooth muscle cells, as observed both in vitro 10,35,39 and in vivo. Transplantation of isolated adult murine Sca-1 + CPCs induced revascularization and revealed differentiation into cardiomyocytes and endothelial cells in infarcted mouse heart 13,35,37. The differentiation versatility of Sca1 + CPCs is developmental stage- and subpopulation-dependent; whereas fetal cells are very suitable for cardiomyogenic and angiogenic development, adult cells prefer smooth muscle cell differentiation 40. Sca-1 + cells were demonstrated to be present in the hypertrophic human heart 12, and the number of resident cells after myocardial infarction was shown to expand 35. Transplantation of both fetal and adult Sca-1 + CPCs, either of murine or human origin, into the mouse injured 6
16 heart limits structural and functional deterioration, and thereby attenuates impairment of contractility. This regenerative potential is mediated both by differentiation of Sca-1 + cells and via paracrine mechanisms 35,37,40,41. Despite these functional benefits of Sca-1 + CPCs in vivo (pre-clinical testing), no clinical trials have yet been conducted. The Sca-1 marker has no homologue in the human genome, therefore the human epitope recognized by this antibody remains unknown. This has so far represented an obstacle to their clinical application, as it poses questions on the identity of these cells 42. A panel of antibodies has recently been published that were specifically raised against resident human Sca-1 + CPCs. These antibodies, such as mab C19, which targets the protein GRP78, recognize CPCs in human heart tissue. Isolated C19 + cells show CPC characteristics and differentiate into the same lineages as Sca-1 + CPCs. This discovery might represent a step forward for the application of these human CPCs in clinical trials 43. Table 1: Summary of identified cardiac progenitor cell populations (adapted from 31 ). Cell type Cardiac resident Selfrenewal Multipotent Activation after injury Improvement of cardiac function Embryonic Adult Side population c-kit Sca Isl1+ +? + +? + 44 Cardio- spheres ?? 7
17 The CPC microenvironment The regenerative potential of the heart is not only determined by the characteristics of CPCs, but also critically influenced by their microenvironment. In this paragraph, key components of the CPC niche (schematically represented in Figure 1A) will be described. Supporting cells In both healthy and diseased hearts, cells interact with each other directly via cell-cell contact or indirectly by the expression of paracrine factors (Figure 3). Interactions can be isotypic (same cell types) or heterotypic (cells of different phenotypes) and the crosstalk between different populations may affect the regenerative potential and cardiac function. These interaction mechanisms are complex and poorly understood. The role of the direct contact between CPCs and supporting cells in vivo is difficult to unravel. Therefore, most knowledge is derived from in vitro experiments and mainly focused on paracrine effects. In this paragraph the interactions of CPCs with supporting cells and their importance for cardiac repair are discussed. Cardiomyocytes In the first in vivo studies on the cardiac niche and putative supporting cells, connexins and cadherins were detected in cellular contacts between CPCs and cardiomyocytes as well as between CPCs and fibroblasts 27,45,46. However, these connections were not observed between CPCs and endothelial cells, and between CPCs and smooth muscle cells 27. Cardiomyocytes are able to transfer information to CPCs (and vice versa) through gap junctions. Co-culture of CPCs with cardiomyocytes promotes their expansion and results in beating CPC-derived cells, together with expression of cardiomyocyte-specific proteins and well organized sarcomeres 14,47 50, a process probably regulated by TGF-β 51,52 and indirectly via the Wnt/β-catenin signaling system 53. Therefore, coupling of CPCs with cardiomyocytes is critical to control the cardiac fate, and lack of appropriate interaction may hamper CPC differentiation 54. However, cardiomyocytes might not solely stimulate differentiation toward the cardiomyogenic lineage. In fact, under hypoxia, cardiomyocytes produce vascular endothelial growth factor (VEGF), which might induce endothelial differentiation of the CPCs 53. At the same time, CPCs can express growth factors and cytokines, which besides being necessary for their proliferation and senescence 55 are also important for cardiomyocyte proliferation, cell survival and prevention of hypertrophy 56. Endothelial cells Since CPCs are often found in the perivascular area, interaction with endothelial cells and smooth muscle cells is highly plausible, although direct cell-cell interactions have not been observed 27. It can be hypothesized that endothelial cell CPC interaction is regulated via Notch, since Notch receptors and ligands are predominantly expressed by the vascular endothelium 57. Notch signaling is crucial for cell fate decisions that underlie cardiomyogenic and vessel formation 57,58. Since Notch signaling is a highly conserved pathway that acts via cellcell contact, it will be discussed later on in more detail. Indirect interactions between 8
18 endothelial cells and CPCs, via the production of VEGF, might not only promote CPC migration 59 but also regulate CPC differentiation towards endothelial or smooth muscle cells 60,61. Immune cells Myocardial injury causes inflammation by activation of immune cells, which are involved in cardiac repair as well as scar tissue formation. Hence, crosstalk between CPCs and immune cells is likely to take place, although there are no proven interactions. Transplantation studies revealed that CPCs are able to dampen the immune response and thereby influence cardiac repair 62. Yet, the mechanisms underlying CPC modulation of the immune system are not completely clear. Macrophages Macrophages are a heterogeneous population of both protective and cytotoxic cells. They play a cardioprotective role by maintaining cardiac homeostasis via interactions with other cardiac cells 63. Macrophages are able to produce growth factors (e.g. IGF-1, VEGF, TGF-β), which stimulates CPC proliferation and induces differentiation towards both cardiomyocytes and endothelial cells 64,65. On the other hand, CPCs are able to polarize macrophages away from their pro-inflammatory phenotype, although the exact mechanism is unclear. It is not known toward which cell type the polarization acts, although it was proven not to be toward the antiinflammatory phenotype 66. Therefore, a cardioprotective effect seems to arise from the interaction between macrophages and CPCs. Natural killer cells Natural killer cells, a subset of the innate lymphoid cell compartment, are effectors of the innate immune system, which is essential in allogeneic transplantation. Their cytotoxic effects are mediated by exocytosis of granules that perforate the target cell to trigger apoptosis 67. Little is known about these cells and CPCs, but a recent study by Boukouaci et al revealed that CPCs are protected from killer cell cytotoxicity within an inflammatory context 68. On the other hand, CPCs are able to downregulate the toxicity of natural killer cells and bias cytokine secretion towards an anti-inflammatory state. Retention of CPCs is improved by this crosstalk with natural killer cells, and contributes to cardiac regeneration 68. Mast cells Mast cells are bone marrow-derived precursors, and although their number increases in the failing heart, their exact role in cardiac disease and regeneration is understudied. Moreover, mast cells express similar markers as CPCs 69. It is known that CPCs share distinct characteristics with mast cells, but not all CPCs are mast cells 69. Both mast cells and CPCs are located in the perivascular area, although cell contact is not reported. Paracrine effects can be assumed since mast cells produce several cytokines, growth factors and angiogenic factors that are all involved in cardiac repair 70. During mast cell degranulation TGF-β is released, which as described earlier is important for CPC differentiation 71. Stromal cells Fibroblasts Together with cardiomyocytes, fibroblasts were the first supporting cells of CPCs to be discovered 27,45. Like cardiomyocytes, fibroblasts are connected to CPCs via gap and adherens 9
19 junctions. Not only do fibroblasts maintain the supporting matrix of the CPC niche 53 (the importance of the ECM-cell interaction will be discussed later on in a dedicated paragraph), but they might also influence the differentiation potential of CPCs. It has recently been shown that fibroblast-conditioned medium can induce differentiation via the Wnt signaling pathway 72 and that fibroblasts produce angiogenic and anti-angiogenic factors 53. Cardiac fibroblasts originate mainly from the epicardium 73, therefore interactions between epicardium-derived cells (EPDCs) and CPCs need to be described. Epicardium-derived cells (EPDC) CPCs are often found in the subepicardial region, which mostly consists of EPDCs. EPDCs have a crucial modulatory role during cardiac development, and their activation after injury suggests a similar role in the adult heart 78. Due to these characteristics some research groups even suggest that the epicardium is a source of progenitor cells 79,80. The presence of CPCs near the epicardium and EPDCs suggest that important interactions occur between CPCs and EPDCs. Previous research showed that EPDCs stimulate the migration and proliferation of CPCs 77,81,82. Co-culture of CPCs with EPDCs revealed induction of metalloproteinases and their inhibitors, which affected infarct size 82. In fact, matrix remodeling is not only important to prevent cardiac dilation after injury, but it also plays a role in maintaining the supporting network of the CPC niche. Moreover, CPC-EPDC co-culture stimulated angiogenesis and consequently improved cardiac function. The interaction between CPCs and EPDCs is reciprocal and results in synergistic action, leading to improved cardiac function. This beneficial effect is at least partially explained by paracrine stimulation 82. Telocytes Another stromal cell type which is present in the subepicardial region is the telocyte, formally known as the Interstitial Cajal-like cell 83. Telocytes are in the close vicinity of CPCs, and stromal synapses and adherens junctions are formed between the cells both in vitro and in vivo 84,85. These adherens junctions not only control the retention of CPCs but might also be important for division and migration of CPCs 85. Therefore, it is assumed that telocytes provide guidance and nursing for CPCs to stimulate their activation, proliferation and differentiation leading to cardiac repair 84,86,87. Furthermore, telocytes produce growth factors (e.g. VEGF) 88 and macromolecular signals, such as micro RNAs 89, which might influence the differentiation potential of CPCs
, both via direct cell-cell signaling (such as the Notch")
20 Figure 3. Cell-cell interactions in the CPC niche. CPCs interact with each other and with supporting cells (cardiomyocytes, fibroblasts, endothelial cells, smooth muscle cells and immune cells), both via direct cell-cell signaling (such as the Notch pathway) and paracrine signaling. Cell-cell signaling via Notch in CPCs Cell-cell contact-dependent signaling is an essential component of the niche, and has an important influence on cellular behavior. Notch signaling is a fundamental and highly conserved pathway that acts via direct cell-cell contact, and has a key role in the heart. Notch regulates a number of cell functions, such as survival, proliferation and differentiation, as well as cardiac tissue development and homeostasis 90,91. In mammals, four Notch proteins have been identified (Notch1-4), which can bind to ligands of the Delta or Serrate/Jagged families expressed by neighboring cells, as extensively reviewed elsewhere 90,92 (Figure 3). Following cleavage by ɣ-secretase, Notch intracellular domain (NICD) is released and translocates to the nucleus, where it regulates the expression of target genes, such as members of the Hes and Hey families 90,92,93, as well as Nkx2.5 in cardiac cells 94. Notch signaling represents an essential element of the cardiac microenvironment. Firstly, Notch plays a crucial role in cardiomyogenesis, and Notch mutations have been linked to several congenital heart and heart valve defects 90,92,93. Active Notch signaling is needed for CPC 11
21 differentiation 95, whereas in cardiomyocytes Notch is activated during embryonic development 91,92, and inactivated during maturation 96 and after birth (reviewed by 91 ). Secondly, several studies demonstrated the re-activation of Notch in adult cardiomyocytes after MI, in small animal models 94 as well as in humans 97,98. Moreover, Notch plays a key role in cardiomyocyte survival 99,100 and cardiac repair after injury 91,101. In the adult mouse, about 60% of c-kit + CPCs express the Notch1 receptor, and signaling with surrounding cells, either CPCs or myocytes, is mediated by Jagged1 94. Notch signaling activation and its outcomes strongly depend on timing and dosage 102,103 ; it is needed for proliferation and expansion of the CPC pool 96, and is essential for CPC cardiomyogenic differentiation 94,95. The activation of Notch1 by Jagged1 in mouse c-kit + CPCs promotes the nuclear translocation of N1ICD, and enhances its co-localization with the cardiac transcription factor and Notch target gene, Nkx However, Notch becomes undetectable when differentiating CPCs lose their proliferative capacity, showing that its downregulation is needed for terminal differentiation 96. Moreover, overexpression of Notch1 in mouse c-kit + CPCs, leads to improved resistance to oxidative stress, and injection of these cells in mouse infarcted hearts resulted in an enhanced cardioprotective effect, as shown by smaller infarct length and area, functional improvement, and larger capillary density as compared to control cells (where endogenous Notch was activated via Jagged1) 58. In view of these findings, Notch has been proposed as a potential therapeutic target for treating myocardial disease. Hydrogels functionalized with a peptide mimic of Jagged1 were shown to activate Notch signaling in rat c-kit + CPCs, and injection of CPCs embedded in these hydrogels led to improved cardiac repair in a mouse MI model 104. Yet, very little is known about Notch in human CPCs. Recent studies, by our group and by others, have investigated Notch signaling in cardiac progenitors cultured as multicellular spheroids, or cardiospheres (if isolated from adult myocardial tissue). The spheroid model better mimics the in vivo cell-cell interactions as compared to cell monolayers, and therefore represents an interesting and promising model to study cell-cell signaling (see paragraph 4, Approaches to mimic the CPC microenvironment in vitro ; 105 ). The formation of cardiospheres from adult explant-derived cells increased the expression of Notch1 and Notch3 receptors 106. Moreover, multicellular spheroid formation was shown to activate Notch signaling in both fetal and adult CPCs, and more evidently so in combination with hypoxic culture, indicating the pivotal role of niche-like environmental conditions on a fundamental cellular pathway such as Notch signaling 107. Understanding the role and activity of this pathway in human endogenous CPCs will be important for the improvement of CPC-based cardiac regeneration therapies. Extracellular matrix Cardiac cells are surrounded by a highly organized and dynamic fibrous network, known as the extracellular matrix (ECM), which, together with cardiac cells, forms the cardiac tissue 108. The cardiac ECM is composed of different proteins, proteoglycans and glycosaminoglycans that 12
22 form a fibrillar mechanical support in which cells are embedded. The structural components of the ECM include collagen type I, III and V, as well as elastin that provides resilience to the cardiac tissue Furthermore, proteoglycans such as tenascin-c and decorin contribute to the cardiac tissue, and are crucial for the stability and integrity of the ECM 112,113. Next to structural components, the ECM is composed of non-structural elements that regulate important cellular functions, such as adhesion, proliferation and differentiation. These are primarily type IV collagen, laminins and fibronectin 109,114. Moreover, within the ECM network, different cell types secrete soluble macromolecules in the extracellular space, such as VEGF, TGF-β, and stromal cell-derived factor 1-α (SDF1-α), that regulate and stimulate important cellular processes Cardiac fibroblasts are primarily responsible for the production and remodeling of ECM 118, both under healthy and under pathological conditions. During and following a myocardial infarction, fibroblasts become activated and secrete an abundance of ECM components and ECM modifiers, such as MMPs 22, to compensate for the loss of cardiomyocytes. Eventually, this leads to excessive ECM formation and scarring, with adverse effects on the contractility of the cardiac tissue. Altogether, this fibrotic environment might not be ideal for the injected progenitor cells. For this reason, the effect of the ECM environment on progenitor cell survival and function should be investigated to improve the CPC contribution to cardiac regeneration. Below, recent literature on the interaction of CPCs with native and/or synthetic ECM in vitro and in vivo is summarized and discussed, with a focus on ECM properties such as stiffness, architecture, and composition. Integrins CPCs are linked to the ECM via focal adhesions (FAs), complexes made of transmembrane and intracellular proteins that localize at the cell-ecm adhesion sites. FAs directly link ECM components to intracellular actin junctions, but also to intermediate filaments and sarcomeres in cardiomyocytes 119 (Figure 4). Important components of the FA are integrins, transmembrane heterodimeric proteins which consist of a combination of α and β subunits. In mammals, 24 types of receptors can be formed and each combination has a specific binding affinity to a different ECM component. For CMs, the most occurring integrins are α1β1, α5β1 and α7β1, which bind specifically to collagen type I (COL), fibronectin (FN) and laminin (LN), respectively, with β1 being the prevalent β subunit 119. Furthermore, the protein expression of different types of integrin also changes from neonatal CMs, where the dominant subunit is α5, to adult CMs, where the α5 is replaced by α7 subunit 119,120. Similar changes in integrins expression can also be observed in response to pathological conditions 119. Human fetal CPCs subjected to cardiomyogenic differentiation protocol in vitro, showed unvaried expression but increased clustering of integrin β1, indicating FA maturation and improved mechanosensing with early cardiac differentiation (as shown in chapter 2 of this thesis) 121. These studies underline the importance of integrins in the heart, and suggest that the FA expression of CPCs and interactions with specific ECM components should also be studied to be able to guide CPCs towards specific lineages. 13

23 Figure 4: Schematic representation of integrin-mediated focal adhesions (FAs) binding the actin cytoskeleton. Integrin dimers composed of α and β subunits localize at the cell membrane, and bind to ECM fibers. An integrin signaling layer contains the integrin cytoplasmic tail and proteins such as FAK; a force transduction layer, where proteins like talin, vinculin and integrin-linked kinase (ILK) create a direct link between integrin and actin fibers, thereby allowing for force transmission between these two; finally, the actin regulatory layer includes proteins such as VASP, zyxin and α-actinin, that modulate and stabilize the actin cytoskeleton 122. Actin fibers can form a complex with myosin II to generate stress fibers (SFs), contractile units that allow the cell to exert forces on the ECM 123. CPC-ECM interactions ECM composition The versatility of interactions between CPCs and their direct surroundings generates the possibility to obtain highly regulated and regenerative cellular responses via intracellular signaling. It is currently known that the stiffness, composition and/or structure of the natural ECM have an effect on progenitor commitment in vitro 124. More importantly, the ECM can evoke different cellular responses depending on the cell type. This has been tested by culturing CPCs on different substrates in vitro and evaluating the cellular responses that can be associated with cardiac regeneration. The first studies were initiated by French et al., who studied c-kit + Sprague-Dawley rat CPC behavior, i.e. cardiomyogenic gene expression, cell survival and proliferation, cultured on decellularized porcine ventricular ECM (cecm) or standard collagen type I (COL), which more closely resembles the biochemical composition of a scar following a MI 125. Early cardiac genes for GATA-binding protein-4 (GATA-4), Nkx2.5, α- myosin heavy chain and troponin C and T were increased when CPCs were cultured for 2 days on cecm compared to COL. Moreover, fibroblast and endothelial/smooth muscle cell-specific genes decreased and remained constant, respectively, for CPCs cultured on cecm, as compared to COL. In a recent study, human Sca-1 + cells originating either from fetal heart (fcpcs) or adult hearts (acpcs) were encapsulated and cultured in three-dimensional (3D) hydrogels consisting of either cecm or COL 126. Similarly, gene expression of early cardiac markers, i.e. GATA-4, 14
24 Nkx2.5, myocyte enhancer factor 2c (Mef2c) and myosin light chain 2v (MLC2v), increased when fcpcs and acpcs were cultured in cecm compared to COL after 4 days. Furthermore, after 7 days these markers increased when fcpcs were cultured in cecm and remained constant for acpcs. One explanation for the minimal increase in cardiac markers after a longer period of time could be the development of an endogenous microenvironment by CPCs that decreases the early biochemical effect of cecm. Additionally, improved proliferation and survival was observed for both cecm-coated surfaces and for CPCs encapsulated in cecm hydrogels. These findings indicate the importance of the biochemical composition for the early maturation of fcpcs towards cardiac specific lineages. Nevertheless, more information is needed on the specific ECM composition to be able to understand which of the components generates the beneficial response of CPCs towards cardiac-derived ECM, considering the fact that CPCs reside in a specific, yet complex, niche that strongly determines their behavior 27. In a follow-up study by French et al., CPCs were cultured on cecm, COL-, FN- and LN-functionalized Bioflex plates and subjected to different cyclic strains 127. Proliferating cell nuclear antigen (PCNA) was used as a measure to determine the proliferation response of CPCs cultured for 30 hours on the different substrates. The highest number of PCNA positive cells was observed on substrates functionalized with fibronectin, demonstrating the benefit of a single ECM component compared to the whole complex cardiac ECM. Fibronectin has been shown to be crucial for the expansion of human CPCs during development and after a MI 128. Additionally, endogenous fibronectin production by human CPCs has been observed after 7 days in static culture 15. Moreover, the beneficial effect of fibronectin on the proliferation of CPCs seems to diminish at strain magnitudes of 10-15%. These findings suggest the importance of fibronectin on the initial proliferation response of CPCs, however the effect can be overruled by other microenvironmental components such as cyclic strain and/or stiffness. ECM stiffness The data described so far suggest that an ideal biochemical environment is not sufficient to completely obtain the desired regenerative response, but that mechanical and/or structural stimuli also contribute to a favorable response. To illustrate this, c-kit + human pediatric CPCs were cultured in neonatal or adult ECM derived from Sprague-Dawley rats and combined with fibrin to create 3D hybrid hydrogels with a range of Young s moduli, i.e. 2, 8, 14 and 32 kpa 129. By increasing the Young s modulus of the neonatal and adult ECM-fibrin hybrid hydrogel from 2 to 8 kpa, the gene expression of cardiac titin decreased, whereas it increased at higher moduli. These findings suggest that ECM stiffness has an effect on the gene expression of CPCs in terms of cardiac titin expression. Encapsulating CPCs in an environment with a stiffness that resembles the native mechanical properties would provide better conditions to study the development of CPCs into mature cardiomyocytes. However, this is complicated by the fact that stiffness values may differ between neonatal, fetal and adult heart, between healthy and diseased conditions, and also between species 130. To study the effect of ECM stiffness on cardiac stem/progenitor cell maturation, hydrogels with time-dependent and development-mimicking stiffnesses were developed based on thiolatedhyaluronic acid (HA) and crosslinked with poly(ethylene glycol) (PEG) diacrylate. By growing 15
25 pre-cardiac embryonic stem cells on HA-hydrogels with elastic moduli ranging from 1 to 10 kpa, a 60%-fold increase in myofibril orientation and a 3-fold increase in troponin T expression was observed as compared to cells grown on mechanically static polyacrylamide hydrogels 131. A recent example of modulating the ECM stiffness is shown by Choi et al., where a sol-to-gel transitional gelatin-peg-tyramine (GPT) hydrogel with tunable mechanical properties was developed 132. Hydrogels with elastic moduli of 1.8, 2.8, 5.8 and 8.1 kpa were created by varying the H2O2 concentration. Interestingly, CPCs isolated from 9 week-old Sprague Dawley rats showed inhibited F-actin organization and decreased proliferation in stiffer GPT hydrogels (elastic moduli of 5.8 and 8.1 kpa) compared to lower stiffnesses (1.8 and 2.8 kpa) 125. However, an enhanced expression level of early cardiac differentiation markers was observed in GPT hydrogels with higher elastic moduli. These results strongly suggest an inhibition of proliferation and enhancement of differentiation in cardiac stem/progenitor cells as a result of increasing the ECM stiffness. Mechanical loading: cyclic strain The adult human heart beats times per minute every day. Cells that reside in the myocardium are constantly subjected to this mechanical loading, which thus represents a significant component of the cardiac microenvironment that can influence the regenerative response of resident CPCs. However, while many studies have focused on the mechanoresponse of contractile cardiomyocytes, the effect of cyclic strain on CPCs has only been investigated by a few research groups. In chapter 2 of this thesis, we elucidate the mechanoresponse of human Sca-1 + CPCs. Cells were cultured on 2D substrates coated with collagen IV, which together with laminin represents the main component of the cardiomyocyte basement membrane. Whereas undifferentiated CPCs did not show a preferential orientation upon application of uniaxial cyclic strain, CPCs in the early stage of cardiomyogenic differentiation (predifferentiated), oriented perpendicularly to the main direction of the stretch (strain avoidance behavior) after 48 hrs 121. The different responses appear to be due to the development of the mechanosensing structures, such as focal adhesions (FAs) and actin stress fibers (the mechanosome), that we demonstrated to occur during the early phase of cardiac differentiation 121. In the study of French 127 mentioned above, cyclically strained rat c-kit + CPCs displayed a different orientation response when cultured on different ECM coatings. After 24 hours, rat CPCs displayed a strong strain avoidance response on fibronectin and collagen I. On the other hand, the strain avoidance response on decellularized porcine ventricular ECM (see above paragraph ECM composition ) was much weaker as compared to collagen I and fibronectin, whereas almost no strain avoidance was observed on laminin 127. Taken together, the reported studies suggest that CPCs on the natural cardiac ECM are less responsive to cyclic strain as compared to single ECM components (fibronectin, collagen I). Furthermore, the mechanoresponse of CPCs is weakened on certain ECM proteins (laminin, collagen IV). It is tempting to speculate that this behavior might be related to the affinity of different integrins for the ECM proteins. It would be interesting to investigate which integrins are expressed by the CPCs on the different substrates, and especially on the naturally-derived cardiac ECM, and 16
26 relate this expression pattern to the CPC mechanoresponse (as previously done in other cell types by 133,134 ). It should be noted that the above studies are limited by their 2D setup, which does not resemble the 3D physiologic environment. In a study by van Marion et al. 135, the effects of human Sca-1 + CPC engraftment in a collagen I/Matrigel hydrogels were investigated. Whereas CPCs showed a random orientation in stress-free hydrogels, in statically constrained hydrogels they aligned along the direction of the strain after 24 hrs. This effect was even more pronounced at day 9 of culture, suggesting that CPCs become readily mechanosensitive in 3D. Detailed investigation of the mechanosensing of (human) CPCs in 3D environments is needed in order to understand the response of these cells to the mechanical stimuli provided by the cardiac microenvironment. Soluble factors and oxygen tension After a myocardial infarction, cardiac cells are immediately exposed to hypoxia, due to the temporary lack of oxygen. Hypoxia has been shown to regulate the behavior of several stem and progenitor cells by dramatically influencing fundamental signaling pathways, such as Notch and Oct4, that determine self-renewal and multipotency In response to low oxygen tension, cells express hypoxia-inducible factors (HIFs), with HIF-1α being the key mediator of the cellular adaptive response to hypoxia 139. For example, HIF-1α is induced in the ischemic myocardium after MI 140. HIF-1 directly regulates the transcription of the chemokine stromalderived factor 1 (SDF-1) 141 and its receptors CXCR4 142, which plays an important role in the mobilization of progenitor cells Moreover, the upregulation of SDF-1 in ischemic tissues is directly proportional to the reduction of oxygen tension 141. The interaction between SDF-1 and CXCR4 plays a crucial role in the mobilization and migration of circulating progenitor cells in ischemic tissues 141,146,147. For cardiac regeneration, the response of resident progenitor cells to low oxygen tension is of great interest, due to the potential contribution of these cells to the cardiac regenerative mechanisms 148. In this respect, a number of studies on murine CPCs have been conducted. These cells express the SDF-1 receptors CXCR4 and CXCR7 149,150. In room air conditions (20% O2), CPCs show very limited expression of CXCR4; however, under (harsh) hypoxia (0.1% O2) expression of both the CXCR4 receptor and the chemokine SDF-1 is greatly enhanced 149. SDF-1 induces CPC migration in a time- and dose-dependent manner 149,150 ; this SDF-1-induced migration is however abolished by knockdown of CXCR4 or CXCR7 150, demonstrating the crucial role of the SDF-1/CXCR4 and SDF-1/CXCR7 axis for CPC motility. Pretreatment of murine CPCs with hypoxia results in increased migration toward SDF-1 in vitro, suppressed by cell transfection with CXCR4 shrna 149, once again indicating the key role of the SDF-1/CXCR4 interaction for CPC motility. Additionally, hypoxic pre-treatment results in improved recruitment of the CPCs to the ischemic myocardium in a mouse MI model 149, suggesting a potential therapeutic benefit was offered by this procedure. 17
.")
27 The response to hypoxia of human CPCs is less known. In a study by van Oorschot et al. 151, human Sca1 + CPCs displayed increased proliferation and motility when cultured under low oxygen tension (1% O2). The motility and migration of these cells were also enhanced by culture in 1% O2-conditioned media 151. Moreover, human Sca1 + CPCs displayed an increase in cell motility directly proportional to the reduction of oxygen tension, similarly to the SDF-1 induction observed in ischemic tissues by Ceradini et al In a hypoxia-gradient microfluidics chip, where high O2 tension was applied on one end (20% or 95%), and 1% O2 at the other end, an increasing number of human Sca1 + CPCs was detected after 24 hrs towards the condition of lowest oxygen tension (Figure 5). CPC displayed similar proliferation in all the areas of the chip, thereby suggesting that the higher amount of cells in the hypoxic area is indeed due to CPC migration. This suggests that, under hypoxic conditions, not only do human CPCs show improved motility, but also release chemoattractants and their receptors. However, the induction of SDF-1 and CXCR4/CXCR7 in human CPCs has not been investigated yet. Given the data here reported on murine CPCs and on other human progenitor cells, a mechanism similar to the SDF-1/CXCR4 axis might take place. Figure 5: CPCs migrate toward lower oxygen concentration in an oxygen gradient device. a) In a PDMS device (showed in the schematic representation) where 20% or 95% O 2 was applied at one end, and 1% O 2 at the other end, an increasing number of CPCs was observed at the lower oxygen side after 24 hrs. Representative images show the increased number of cells (nuclei stained with Hoechts 33342, blue). b) The quantification of cell number (normalized to the initial value after seeding) is reported as mean ± SD (n=4; *p<0.05; **p<0.01). Approaches to mimic the CPC microenvironment in vitro The CPC niche is complex and its importance for cardiac differentiation, maturation and contribution to repair is largely unknown. For a better understanding, engineering approaches to recapitulate the native cardiac microenvironment are required. To recreate the CPC niche in 18
28 vitro several key components are of importance. In this paragraph we will focus on current in vitro engineering approaches to mimic the cellular natural environment (Figure 6). As previously stated, the CPC microenvironment or niche should display key characteristics, such as optimal biochemical, physical and mechanical properties, to enhance the regenerative response of CPCs. This regenerative microenvironment should therefore stimulate either proliferation, differentiation, and/or organization, or elicit a beneficial effect on the paracrine signaling of CPCs. To date, little is known of the exact characteristics of this ideal niche and what is necessary to obtain optimal CPC contribution to cardiac regeneration. Multicellular spheroids are scaffold-free spherical cell aggregates that mimic in the most simplistic way the conditions of the niche As compared to 2D cell culture, spheroids provide improved cell-cell and cell-ecm interactions, as well as gradients of soluble factors, such as oxygen and nutrients Therefore, cell spheroids are used as a model to study cell behavior in a 3D environment that better resembles the in vivo conditions. At the same time, they could entail major advantages for clinical use over the injection of cells grown as a monolayer, especially in the treatment of cardiac disease (as extensively described by 105,158 ). However, an engineered microenvironment could provide more specific signals to CPCs, and its characteristics might be tuned to elicit a distinct response. Currently hydrogels, decellularized ECM, and synthetic matrices are used to create 3D cardiac environments that take into account cell-matrix interactions, as described above. However, though these matrices mimic ECM-like features, they do not always resemble the mechanical strength of the native tissue 159. Unfortunately, only a small amount of studies are performed with CPCs in matrices to create an engineered CPC-niche, although recently these are being increasingly explored 127, Cardiosphere-derived cells in both alginate 164 or biodegradable poly-(n-isoproplacrylamide) hydrogels showed cardiomyogenic differentiation and proliferation 165, and provided functional benefits 166. Nevertheless, these approaches neglect some of the key aspects of the cardiac microenvironment. By making use of biomaterials that can form well-defined and smart microenvironments, more knowledge can be extracted to be able to define the ideal CPC niche. Recently, scalable engineered and force-generating human myocardium was produced under well-defined conditions using embryonic stem cells, induced pluripotent stem cell-derived cardiomyocytes, and fibroblasts 167. Moreover, extensive evidence for cardiac molecular maturation and functional tissue formation was obtained using RNA sequencing techniques. According to Tiburcy et al., the most important responses that determine the degree of in vitro maturation of human cardiomyocytes are artificial electrical pacing 168, mechanical stimulation (uniaxial and cyclic load) 169, and co-cultures with fibroblast-like cells 170. Finally, they were able to develop a model that can be used to screen drugs, study heart repair or model heart disease, and to study the endogenous repair of CPCs. Another method to create well-defined microenvironments is by making use of micro- and nanoscale engineered biological systems on a chip 171. With these techniques different niche components, such as mechanical , electrical 175 or topographical cues 176, can be carefully modulated and stem cell responses can then be studied in more detail. For instance, Morez et al. have shown improved cardiomyocyte 19

29 differentiation from CPCs using silicone parallel microgrooves (10 μm wide and 3 μm deep) in vitro 177. Further in vitro research is needed to dissect the influence of different niche components on the behavior and regenerative potential of CPCs, in order to make the next step towards the successful (endogenous) cardiac repair by CPCs. In this thesis, the interplay between CPCs and their microenvironment is investigated, and the results discussed in view of the regenerative potential of CPCs. Figure 6: In vitro approaches to modulate the CPC niche. Strategies to optimize the regenerative potential of CPCs by modulating their microenvironment include (clockwise): engineering the ECM with synthetic or naturally-derived polymers with the right composition and physical properties; recreating a niche-like environment by growing cells as multicellular spheroids; applying electrical pacing and/or cyclic strain; co-and multi-culture of different cell types to optimize cell-cell interactions, with or without surrounding ECM; and modulating the cell recruiting potential held by gradients of oxygen and cytokines and growth factors. 20
30 Rationale, aim and outline of the thesis Cardiac progenitor cells have emerged as a potential cell source for myocardial repair and regeneration after cardiac damage. Despite their promising characteristics, and the reported activation of resident CPCs in the adult heart after acute myocardial infarction, their actual contribution to cardiac repair and regeneration is still mostly unknown. In addition, the beneficial outcomes of CPC transplantation into the infarcted heart are limited to the short term, and mostly due to paracrine effects. This is mainly due to the massive cell loss after injection, and to the reduced survival and differentiation of CPCs injected in the hostile environment of the damaged myocardium. To exploit the regenerative potential held by CPCs, and to improve current therapeutic strategies of CPC-mediated cardiac regeneration, a better understanding of the interactions of CPCs with their microenvironment (the CPC niche) is needed. Microenvironmental factors that can influence CPC behavior include: cyclic straining (due to cardiac beating), ECM, neighboring cells, and oxygen tension. The aim of this thesis is to explore and understand the impact of these environmental conditions on CPC function and differentiation, using 2D and 3D in-vitro model systems. In chapter 2, we investigated whether the response of CPCs to cyclic strain was influenced by the cell differentiation stage. The results show that upon early cardiomyogenic differentiation, CPCs develop their mechanosome, i.e. the assembly of molecular structures responsible for cell-ecm adhesions and signaling, as well as the ability to reorient perpendicular to the direction of cyclic stretching (strain avoidance). In chapter 3 the role of the mechanosome in CPC strain avoidance response was analyzed, by blocking the mechanosome components integrinβ1, integrin-linked kinase (ILK), or Rho-kinase (ROCK) in predifferentiated CPCs. The absence of strain avoidance following the mechanosome manipulation indicates the key role of the mechanosome in CPC reorientation response. Next, we studied the influence of cell density and cell-cell adhesions on the response of CPCs to cyclic strain (chapter 4). Undifferentiated CPCs displayed enhanced strain avoidance response in 2D with increasing cell density. N-cadherin mediated cell-cell adhesions, as well as ROCK activity, were shown to both contribute to the strain avoidance behavior of undifferentiated CPCs at high cell density. Next to the 2D cultures, we developed a fibrin-based 3D model system (chapter 5), to allow application of cyclic strain to undifferentiated CPCs in a 3D environment. Culture in the soft 3D fibrin hydrogel led to a shift of CPCs towards the vascular fate. Moreover, CPCs cyclically strained in 3D showed reorientation parallel to the strain direction (strain alignment), in contrast with the strain avoidance observed in 2D, and independently of SF development. The activity of YAP, one of the main sensors of the cell mechanical environment, was lower in 3D as compared to stiffer 2D substrates, indicating reduced cellular mechanosensing in the soft 3D matrix. These findings highlight the importance of studying CPC behavior in more physiologic environments. 21
31 The relevance of niche-like microenvironmental conditions was further studied in chapter 6, by culturing CPCs as spheroids on poly-lysine coated plates, under room air (20% O2) and physiologic (5% O2) oxygen conditions. Multicellular spheroids provide niche-like environments, with improved cell-cell and cell-ecm contacts as well as gradients of oxygen and nutrients. Increased expression and activation of Notch were observed in CPC spheroids, as compared to the cell monolayer, particularly under hypoxia. Finally, in chapter 7 the main results of this thesis are summarized and discussed in the context of mimicking an engineered microenvironment to investigate CPC behavior and enhance their regenerative potential. 22
32 CHAPTER 2 Cardiomyocyte progenitor cell mechanoresponse unrevealed: strain avoidance and mechanosome development This chapter is based on: Mauretti A., Bax N.A.M., van Marion M.H., Goumans M.J., Sahlgren C., Bouten C.V.C., Cardiomyocyte progenitor cell mechanoresponse unrevealed: strain avoidance and mechanosome development, Integrative Biology (2016).
33 Abstract For emerging cardiac regeneration strategies, it is essential to know if and how cardiac stem cells sense and respond to the mechanical stimuli provided by their environment in the beating heart. Here, we study the response to cyclic strain of undifferentiated and early differentiated (predifferentiated) human cardiac progenitor cells (CPCs), as well as the formation and activation of the cellular structures in direct contact with the extracellular matrix (ECM) and involved in mechanosensing, that we termed mechanosome. Once verified that the applied uniaxial cyclic strain (10%, 0.5Hz) did not alter the cardiac lineage commitment and differentiation state of CPCs, the cellular mechanoresponse to the applied strain was quantified by cellular orientation. While undifferentiated cells maintained a random orientation, predifferentiated CPCs exhibited a distinct reorientation perpendicular to the direction of applied cyclic strain (strain avoidance). The mechanosome development and the activation of the mechanotransduction pathways also occurred with early cardiac differentiation of the CPCs, regardless of the substrate or the applied cyclic strain. These results indicate that the mechanoresponse of CPCs depends on the presence of a developed mechanosome, which develops during early cardiomyogenic differentiation. Our findings provide a first understanding of mechanotransduction in human CPCs and as such can contribute to the improvement of cardiac regeneration strategies. 24
34 Introduction Cardiovascular disease is among the leading causes of mortality in the Western world. Cardiac regenerative therapies focus on the repair and restoration of cardiac function with the use of stem or progenitor cells. In the human heart, several cardiac progenitor cell (CPC) populations have been identified 178. Among these, Sca1+ cardiomyocyte progenitor cells (CPCs) have shown the ability to differentiate into the three cardiac lineages in vitro (cardiomyocytes, smooth muscle cells, and endothelial cells) 179,180. After transplantation in the injured heart, CPCs improve cardiac function and reduce scar formation 179. Our group has recently demonstrated that this can be attributed to the production and modulation of extracellular matrix (ECM) proteins 15 to obtain a structurally organized, healthy ECM. Therefore, CPCs represent a promising stem cell source for cardiac regenerative therapies. For successful regenerative outcomes the cells need to integrate into the cyclically strained myocardium. Yet, very little is known of cardiac stem cell mechanosensitivity and mechanoresponse. By revealing the mechanisms that trigger mechanosensitivity in CPCs, it may be possible to direct cell behavior towards the regeneration process. In response to external mechanical cues, several differentiated cell types, such as endothelial cells 181,182, fibroblasts 183,184, and smooth muscle cells 185, have shown the ability to re-orient perpendicularly to the direction of applied cyclic strains in 2D setups. This behavior is called strain avoidance, and it shows the tendency of cells to align in the direction of minimum deformation of the cell itself and its mechanosensing components, thereby allowing cells to reside in areas of high strains in the body While cell mechanoresponse in 3D is largely unknown, strain avoidance behavior is also exhibited in 2D by several stem cell types when subjected to cyclic strain, such as mesenchymal stem cells 189, bone marrow-derived progenitor cells 190, and adipose-derived stem cells 191. However, for CPCs this behavior is unknown. The process through which cells sense and respond to mechanical stimuli is called mechanotransduction, and the protein assemblies that mediate the signal transmission are known as focal adhesions (FAs). FAs are highly dynamic structures that form attachment sites of cells with the ECM. Normally, FAs localize at the end of actin stress fibers (SFs) 192,193 and thereby link the actin cytoskeleton with the ECM. Here, we refer to this cellular mechanosensing apparatus, which is in direct contact with the ECM, with the term mechanosome. The term mechanosome was first introduced in the beginning of this century by Pavalko et al. 194,195, to describe the load-bearing multi-protein complex in bone cells, which is composed of adhesion-associated and DNA-binding proteins. In the present study, the mechanosome is defined as the complex of cellular structures linked to the ECM responsible for cell mechanosensing and the mechanoresponse. The complex consists of SFs as well as integrin mediated adhesions, including vinculin, integrin-linked kinase (ILK) and focal adhesion kinase (FAK) 196. These adhesion complexes mediate bidirectional communication between cells and the ECM 197,198. Furthermore, integrins are also involved in the early formation of FAs, in which they form a stable complex with vinculin and talin. Vinculin is one of the best studied FA proteins, known for its important role in mechanotransduction. While its inactive form resides in the cytoplasm, vinculin acquires an extended conformation and localizes at the FAs upon 25
35 activation. 199 Another important player in mechanotransduction is ILK, which is fundamental for cardiomyocyte survival and force transmission, and for cardiac function 200,201. Upon integrin- ECM binding, FAK undergoes autophosphorylation at the Y397 tyrosine site (pfak), leading to phosphorylation of a number of cytoskeletal proteins that are responsible for the maturation of FAs and the SFs. However, the CPCs mechanosome and its key players are still unknown. In the present study, we investigated the mechanoresponse of L9TB CPCs to uniaxial cyclic strain, comparing undifferentiated cells with CPCs in their early differentiated state (referred to as predifferentiated CPCs). The applied cyclic strain caused strain avoidance of predifferentiated CPCs but not of undifferentiated cells. Furthermore, the mechanosome developed upon early cardiomyogenic differentiation. A different seeding substrate or applied cyclic strain did not induce mechanosome development in undifferentiated CPCs. Taken together, our results demonstrate that CPCs gain the ability to respond to cyclic strain upon early cardiac differentiation, guided by the maturation and activation of the mechanosome. These results provide insight in the mechanobiology of CPCs, and could help understand if and how these cells will react and adapt in the heart, which is an indispensable input for the development of new cardiac regenerative strategies. Materials and methods Culture of cardiac progenitor cells Human fetal CPCs were isolated and cultured as described previously 39,202. In this study two primary cell isolations were used, the L3 and L6. The third group of cells, the L9TB CPCs, was immortalized by lentiviral transduction of htert and BMI-1. All CPCs were cultured in SP++ growth medium consisting of M199 (Gibco)/EGM2 (3:1) supplemented with 10% (v/v) fetal bovine serum (FBS) (Greiner bio-one), 1% (v/v) non-essential amino acids (Gibco), and 1% (v/v) penicillin/streptomycin (Lonza) on 0.1% (w/v) gelatin (Sigma Aldrich)/PBS (Sigma) coated flasks. Differentiation of CPCs To induce differentiation, starting on ddiff 1 (Figure 1A) CPCs were treated with 5 μm 5- azacytidine (Sigma) for 72 hrs in differentiation medium (IFDIFF) consisting of IMDM (Iscove's Modified Dulbecco's Medium)/Ham-F12 (1:1) (Gibco) supplemented with 2% (v/v) horse serum, 10 4 M ascorbic acid (A-A, Sigma), 1% (v/v) non-essential amino acids, 1% (v/v) insulintransferrin-selenium (Lonza), and 1% (v/v) penicillin/streptomycin. After 72 hrs, the medium was changed, followed by transforming growth factor (TGF)-β1 (1 ng/ml; Sigma) stimulation. Every 3 days, the medium with TGF-β1 was refreshed (Figure 1A, ddiff 7, ddiff 10, ddiff 13), as described previously 202. The differentiation protocol was carried on for 14 days before strain application and verified by gene expression prior to exposure to dynamic strain. After 14 days of differentiation, CPCs reach an early differentiated state (predifferentiated CPCs), whereby cardiac markers such as cardiac troponin T and myocardin are upregulated, and the sarcomeric component α-actinin starts to be expressed, although sarcomeric organization cannot be observed at this point (Figure S1). 15 For the straining experiments, predifferentiated cells were seeded on Bioflex 6-well plates (Flexcell International Corporation, each well is made of a 26

36 silicone membrane, Figure 1B) at day0, and the differentiation protocol was continued up to day2. Predifferentiated CPCs were compared to undifferentiated CPCs (Figure 1A, ddiff 0). Figure 1: Timeline and setup of the experiment. A) Predifferentiated CPCs were differentiated for up to 16 days starting on d diff1, by switching to differentiation medium (IFDIFF), enriched with ascorbic acid (A- A) and TGFβ1 (in the second week, d diff7, d diff10). After 13 days of differentiation (d diff 13), predifferentiated and undifferentiated CPCs were seeded on Bioflex well plates and cultivated for 24 hrs. The next day (day0), uniaxial cyclic strain was applied for up to 48 hrs (day2). B-D) Mechanical loading was applied with a FlexCell device: the Bioflex plate (B) was placed on top of Teflon blocks oriented in the x direction, in order to apply strain in the y direction (0, blue arrow). The plate was placed on the FlexCell system (C) for straining. The FlexCell device creates vacuum below the plate (D, side view), and the cells seeded in the Bioflex plate are stretched in the direction perpendicular to the orientation of the post (D, 0, top view). 27
37 Cyclic straining Cyclic strain was applied using a FX-5000 Flexcell system (Flexcell International Corporation). Undifferentiated and predifferentiated L9TB CPCs were seeded on collagen IV-coated Bioflex plates. Cells were seeded at 75% confluency and left for 24 hrs to allow attachment to the membrane. Uniaxial cyclic strain was applied by stretching the membranes on rectangular posts oriented in the x direction (90, Figure 1 B, D). Strains of 10% (0.5 Hz, sine wave) were applied for up to 48 hrs (day 2) in the y direction (0, Figure 1 B, D). Samples were analyzed at day0 (before the onset of the straining protocol), after 24 hrs (day1), and 48 hrs (day2) of cyclic straining. Unstrained samples, cultured on identical Bioflex plates and under the same conditions, were used as controls. Quantification of cell alignment The orientation response of L9TB CPCs upon strain was determined from triplicates from three independent experiments. CPCs were incubated for 20 min with 1 μg/ml calcein AM (Sigma). Before visualization, the medium was refreshed. Only cells in the central part of the wells, where the strain field was homogenous according to previous in-house calibration, were considered. Cell orientation was quantified with Fiji, using the Directionality plug-in ( based on the Fourier transform (FFT) of each image by means of fitting a Gaussian to the FFT signal, measuring its peak position. For each group, 4 to 9 images were analyzed for each of the three independent experiments (n=16-23). Immunofluorescence labeling The CPCs that were cultured on cover glasses and Bioflex membranes were washed three times with Ham-F12 medium (Gibco) at 37 C, fixed in 3.7% (v/v) formaldehyde (Merck) for 15 min, washed three times with PBS, and permeabilized with 0.5% (v/v) Triton X-100 (Merck) in PBS for 10 min. The Bioflex membranes were cut out of the plates with a scalpel, using the center of the membrane for immunofluorescence staining. Non-specific antibody binding was blocked with 1% (v/v in PBS) horse serum incubation for 40 min. Cells were then incubated overnight at 4 C with primary antibodies in 10% (v/v in PBS) horse serum. Primary antibodies against FA proteins vinculin, integrin-linked kinase (ILK), and phosphorylated FAK (pfak) were used to determine FA development. The cells were rinsed with PBS four times for 5 min and incubated for 1.5 hrs with secondary antibodies, and TRITC-labeled phalloidin for visualization of F-actin, in PBS. All used antibodies and dyes are listed in Table 1. Nuclei were stained with DAPI for 5 min, after which cells were again washed four times with PBS and mounted on microscope glass slides with Mowiol. Except for primary antibody incubation, all incubation steps were performed at room temperature. Mechanosome development on silicone and glass substrates To observe the development of the mechanosome in undifferentiated and predifferentiated CPCs during cell spreading and on different substrates, cells were seeded on: a) collagen IVcoated Bioflex 6-well plates as purchased by the manufacturer, and b) 13 mm diameter glass coverslips, placed in 24-well culture plates and sterilized with 70% (v/v) ethanol, coated with 25 28
38 μg/ml collagen IV (Corning). Cells were seeded with a density of cells/cm 2, and fixed after 1 hr, 6 hrs and 24 hrs following plating for immunofluorescence staining. Total Protein Isolation and Isolation of Cytoskeletal and Soluble Protein Fractions Total protein extracts of L3, L6 and L9TB CPCs were obtained by lysing the cells in Ripa lysis buffer (Sigma-Aldrich) containing protease inhibitors (Sigma-Aldrich) (n=6). Protein concentrations were determined using a Bicinchronic Acid (BCA) protein assay (Pierce). Total protein extracts were stored at C until analysis. Soluble and cytoskeletal bound protein fractions of L3, L6, L9TB CPCs were isolated using the ProteoExtract cytoskeletal isolation and enrichment kit (Millipore) (ddiff1 n=6; ddiff 7, ddiff n=4; ddiff n=5). In short, after washing the cells with cold PBS, soluble proteins were isolated using cellular extraction buffer (CEB). After washing with cytoskeletal wash buffer (CWB), nuclear proteins were isolated after 10 min incubation with nuclear extraction buffer (NEB). Cells were washed again with CWB and cytoskeletal proteins were isolated after scraping in cytoskeletal solubilization buffer (CSB). All buffers contained a protease inhibitor cocktail and sodium orthovanadate, included in the kit. To further solubilize cytoskeletal protein extracts of 14 days cultured CPCs, lysates were sonicated 3 times for 10 s at an amplitude of 7 microns (Soniprep 150, Sanyo). Separate protein fractions are further referred to as soluble fraction (S), nuclear fraction (N) and cytoskeletal fraction (C). For comparison between fractions, equal volumes of the different fractions were used. To check for proper separation of the different fractions, samples were checked with Western blotting for GAPDH (only present in S and N) and vimentin (only present in C) (data not shown). Protein fractions were stored at 20 0 C until further analysis. Western Blotting The level of focal adhesion development was semi-quantitatively examined in total protein extracts and separated protein fractions (S and C) by Western blotting for β1-integrin and vinculin. Equivalents of 5 µg total protein or 12.5 µl of the separated protein fractions were size fractioned on a 10% polyacrylamide gel and blotted onto a Hybond PVDF membrane (Immobilon-P, Millipore). Non-specific binding sites were blocked with 5% milk powder (ELK) in 0.1 % PBS/Tween20. Membranes were probed with primary antibodies at 4 o C overnight, washed and subsequently incubated at room temperature with horse-radish peroxidase (HRP) labeled secondary antibodies for 1hr (for used antibodies and their dilutions see table 1). For total protein extracts, GAPDH was used as loading control. For visualization, membranes were incubated with enhanced chemiluminescence substrate (Thermo Scientific Pierce), and bands were detected using the Proxima C16 Phi+ (Isogen) imager. Bands were quantified by determining the intensity of the bands (INT*mm 2 with global background correction) using Quantity One image analysis software (Biorad, version 4.6.6). Statistical analysis Cell orientation data are presented as mean ± standard deviation (SD; n=16-23 from 3 independent experiments). Western blot data are shown as mean ± standard error of the mean 29
39 (SD; ddiff1: n=6, ddiff 7-14: n=4 from three independent experiments). Statistical analysis was performed using one-way ANOVA with Bonferoni post hoc test for multiple comparisons. Significance was assumed when p<0.05. Graphics and statistical analyses were performed with GraphPad Prism software (version 5.04). Table 1: List of used antibodies and dyes. Antigen Source Cat. No Isotype Label Species Dilutio n IF Dilution WB Vinculin (IF) SA V9131 IgG1 - Mouse 1:400 1:500 ILK SC sc IgG2b - Mouse 1:100 FAK (WB) BD bd IgG1 - Mouse 1:500 FAK (py397) (IF) BD IgG1 - Mouse 1:100 1:1000 β1-integrin SC sc IgG1 - Mouse 1:400 actin Ab ab-3280 IgG1 - Mouse 1:10000 GAPDH MI CS IgG1 - Mouse 1:5000 Mouse IgG1 MP A21121 IgG1 Alexa Fluor 488 Goat 1:300 Mouse IgG2b MP A21141 IgG2b Alexa Fluor 488 Goat 1:300 Mouse IgG + IgM Pi IgG + IgM HRP Rabbit 1:10000 Rabbit IgG IV A IgG (H+L) Alexa Fluor 555 Goat 1:300 Phalloidin SA P1951 TRITC Amanita phalloides 1:200 DAPI SA D :500 Abbreviations used in this table: SC, Santa Cruz; SA, Sigma Aldrich; BD, BD Biosciences; Ab, Abcam; MI, Millipore; MP, Molecular Probes; IG, Invitrogen; Pi, Pierce; HRP, horseradish peroxidase. 30
40 Results CPC strain avoidance is induced by differentiation The ability of CPCs to sense and respond to mechanical cues was analyzed by applying uniaxial cyclic strain for 48 hrs, and visualizing cell alignment with calcein staining. The early differentiation state was verified by gene expression and immunostainings of cardiac markers (Figure S1). The applied strain had no influence on the CPC cardiac lineage commitment and differentiation state, as demonstrated by the unchanged gene expression of cardiac markers following strain application (Figure S2). Before strain was applied (day0), both undifferentiated and predifferentiated cells were randomly oriented (Figure 2A, B). This random orientation was maintained throughout the experiment in the unstrained samples for both cell types (Figure 2C, D, G, H). After 24 hrs (day1), strained undifferentiated CPCs displayed random orientation, although some cells, especially in the areas characterized by higher cell density, showed orientation perpendicular to the strain direction (Figure 2 E, M). After 48 hrs of strain (day2), undifferentiated cells did not show a preferred orientation. However, in those areas where undifferentiated cells showed strain avoidance at day1, some reorientation at 90 was observed at day2 as well (Figure 2I, O). Strained predifferentiated CPCs displayed random orientation at day1, regardless of cell density (Figure 2F, M). In contrast, at day 2 strained predifferentiated CPCs showed a distinct strain avoidance response, and re-oriented perpendicularly to the direction of applied strain (Figure 2J, O). The amount of cells oriented at exactly 90 was used as representative value of orientation for all groups to perform statistical analysis (Figure 2P, Q). At day2, the fraction of oriented cells was significantly different between strained undifferentiated and strained predifferentiated CPCs (p<0.001, Figure 2P, Q). Moreover, a significant difference was observed between unstrained and strained predifferentiated CPCs (p<0.01, Figure 2P). 31
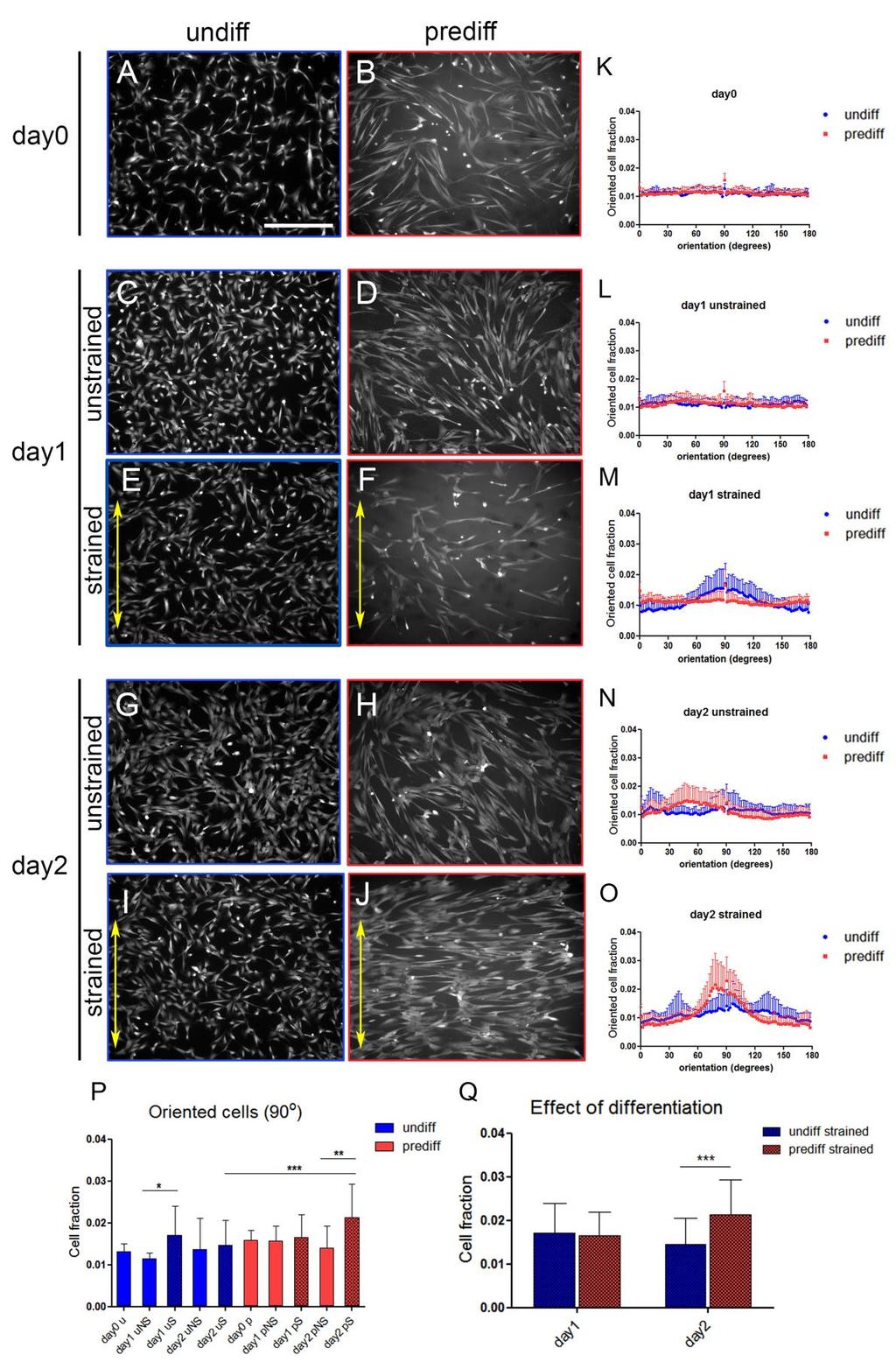
41 32
42 Figure 2: CPCs respond to cyclic strain upon differentiation. A-J) Representative fluorescent images (calcein staining) of undifferentiated (undiff) and predifferentiated (prediff) CPCs at day0, day1, and day2 of strain (strain direction is indicated by the arrow, 0 ). K-O) Frequency distributions display the cell orientation corresponding to each group.. While undifferentiated CPCs (blue) do not respond to the strain, predifferentiated cells (red) show strain avoidance response at day2. The fraction of cells oriented at 90 (P, Q) shows a clear strain avoidance response of predifferentiated CPCs at day2 (u=undifferentiated; p=predifferentiated; S=strained, patterned fill; NS=unstrained, solid fill). Scale bar indicates 400μm. Results are expressed as mean ± SD (n=16-23 from 3 biological replicates); *p<0.05; **p<0.01; ***p< Differentiation but not strain induces the mechanosome development in CPCs The mechanosome plays a crucial role in cellular mechanotransduction; it includes integrins, FA proteins, and actin SFs Upon application of forces to the cell, FA proteins at the cell-ecm adhesion sites, such as vinculin, ILK, and FAK, are activated, relocated to the FAs, and allow the cell to respond by modifying its actin cytoskeleton. 206 Here, we examined the protein distribution of vinculin, ILK, and FAK, and the development of SFs in undifferentiated and predifferentiated CPCs in response to cyclic strain. At day0 (Figure 3A-F), undifferentiated CPCs showed an undeveloped mechanosome. These cells displayed a mainly cytoplasmic expression of FA proteins, with few small protein clusters localized at the cell edges, typical of immature adhesions (Figure 3A, C, E). 193,207 Staining for F-actin showed the presence of cortical and cytoplasmic actin, while no SFs were visible (Figure 3A). Upon early cardiac differentiation (predifferentiated) CPCs exhibited a more developed mechanosome. At day0, the presence of mature FAs was demonstrated by the expression of all investigated FA proteins, distributed as clusters localized on the basal side of the cell and at the cell edges (Figure 3B, D, F). Cortical and cytoplasmic actin with a few SFs were visible (Figure 3B). After one (day1, Figure 3G-R) and two days (day2, Figure 3S-AD) of straining, undifferentiated CPCs still showed an immature mechanosome, with only small clusters of vinculin and absence of SFs (Figure 3G, M, S, Y), and cytoplasmic expression of ILK (Figure 3I, O, U, AA) and pfak (Figure 3K, Q, W, AC). Predifferentiated cells instead, exhibited FA clusters and well-developed SFs (Figure 3H, J, L, N, P, R, T, V, X, Z, AB, AD). In both cell types, slightly larger FAs were observed in unstrained samples (Figure 3G-L, S-X), suggesting the presence of more stable adhesions, whereas strained samples displayed thinner and more elongated FA assemblies (Figure 3M-R, Y-AD). Therefore, the applied strain does not seem to induce the development of the mechanosome. 33
 At day0 of straining, undifferentiated (undiff, A, C, E) CPCs display immature FAs (green) and absence of actin SFs (red), whereas predifferentiated (prediff, B, D, F) CPCs show a more developed")
43 Figure 3: The mechanosome development is enhanced in CPCs upon differentiation, and is not promoted by the applied strain. A-F) At day0 of straining, undifferentiated (undiff, A, C, E) CPCs display immature FAs (green) and absence of actin SFs (red), whereas predifferentiated (prediff, B, D, F) CPCs show a more developed mechanosome, characterized by large FA clusters and SFs. A) At day1 (G-R) and day2 (S-AD) of straining, undifferentiated CPCs (G, I, K, M, O, Q, S, U, W, Y, AA, AC) show small clusters of vinculin and cytoplasmic expression of ILK and pfak, indicating immature adhesions. Cortical and cytoskeletal actin, with absence of SFs are observed. In predifferentiated cells (H, J, L, N, P, R, T, V, X, Z, AB, AD), larger clusters of vinculin, ILK, and pfak are visible. Actin SFs are present in predifferentiated cells, with thicker fiber bundles in strained samples. In both cell types, unstrained cells display larger FA protein clusters, with more elongated assemblies in strained cells. Scale bar 100µm. The CPC mechanosome develops with differentiation regardless of the substrate characteristics To verify the differentiation-induced mechanosome development, we analyzed cell spreading and mechanosome development in undifferentiated and predifferentiated CPCs seeded on glass cover-slips. Since vinculin is one of the first adhesion proteins to be recruited to FAs 193,203, and actin SFs are responsible for cell contractility 123, we investigated the distribution of vinculin and the formation of stress fibers at 1hr, 6hrs, and 24hrs after seeding. It is well-known that some cell types only develop FA clusters and SFs when cultured on stiff substrates To verify whether the mechanosome maturation is influenced by the seeding substrate in CPCs, we analyzed the development of vinculin clusters and SFs in undifferentiated and predifferentiated CPCs on two collagen IV-coated surfaces: the silicone Bioflex membrane and glass. 34
44 During attachment to and spreading on both substrates cell morphology was remarkably different between undifferentiated and predifferentiated CPCs (Figure 4). Both on silicone and glass, undifferentiated CPCs showed a more elongated shape than predifferentiated cells, and displayed faster spreading and less extensive adhesions to the substrate. On the Bioflex membrane, undifferentiated cells showed an undeveloped mechanosome, with mostly perinuclear expression of vinculin (Figure 4A, E, I, M), and only cortical and cytoplasmic actin (Figure 4Q). In predifferentiated cells instead, vinculin was mainly localized around the nucleus and at the cell edges up to 6 hrs after plating (Figure 4B, F), while at 24 hrs vinculin clusters were visible especially at the cell edge (Figure 4J, O). On the silicon surface, SFs (Figure 4S) were formed in predifferentiated cells, as shown after 48 hrs of seeding in Figure 3 G, H. Undifferentiated CPCs did not develop the mechanosome even when seeded on glass coverslips (Figure 4B, F, J, N, R), whereas predifferentiated cells developed slightly larger vinculin clusters and more prominent stress fiber bundles on glass (Figure 4D, H, L, P, T), as compared to the silicone substrate. However, the absence of mechanosome in undifferentiated CPCs was evident on both substrates. Furthermore, CPCs displayed a similar mechanosome development on substrates with different composition (collagen I, collagen IV, fibronectin, and gelatin coating on glass, data not shown). This suggests that the development of the mechanosome is also not determined by the substrate composition (i.e. by specific integrin isotypes), but predominantly intrinsic to the cellular differentiation status. This shows that neither changes in the mechanical properties, nor substrate composition, can induce the maturation of FAs and SFs in undifferentiated CPCs. The differentiation-induced mechanosome development is an intrinsic property of these cells. 35
 Undifferentiated CPCs (undiff, A, C, E, G, I, K) show immature adhesions and weak expression of actin SFs; predifferentiated CPCs (prediff: B, D, F, H, J, L) present more pronounced expression")
45 Figure 4: Undifferentiated and predifferentiated CPCs show different cell spreading and mechanosome development after seeding. A-L) Undifferentiated CPCs (undiff, A, C, E, G, I, K) show immature adhesions and weak expression of actin SFs; predifferentiated CPCs (prediff: B, D, F, H, J, L) present more pronounced expression of vinculin and stress fiber formation. M-T) Both cell types show faster spreading on glass (N, P, R, T) than on the silicone Bioflex surface (M, O, Q, S), but the difference in the mechanosome development induced by early differentiation is maintained. Scale bar 100 μm. Mechanotransduction is activated during early cardiac differentiation To further verify the maturation of the mechanosome and the activation of the mechanotransduction pathways, we examined how different functional components of the mechanosome are activated during CPC differentiation. FA formation and maturation is initiated by the accumulation and clustering of integrins and FA proteins, such as vinculin and focal adhesion kinase (FAK). Vinculin is well known for its role in cell mechanotransduction; β1- integrin has high binding affinity for collagen IV, which is the main ECM component of the cardiac basement membrane. Thus, the expression levels and clustering of vinculin and β1- integrin, the cytoskeletal development, and the auto-phosphorylation of FAK were studied after 1, 7, 8, and 14 days from the start of the differentiation process. From immunofluorescent images it is evident that undifferentiated CPCs did not display neither vinculin clusters nor actin SFs, but only cytoplasmic expression of vinculin and actin was observed (ddiff 1, Figure 5A). On ddiff 7 and 8, a few SFs, mainly located close to the cell membrane, and small vinculin clusters could be observed (Figure 5B, C), whereas at ddiff 14 36
46 numerous prominent actin SFs throughout the whole cell were present, and large vinculin clusters co-localized with SFs (Figure 5D). Corresponding to our observations from immunofluorescent images, the results of Western blot analysis on total protein extracts show that the amount of vinculin increased from ddiff 1 onwards to approximately 2-fold on ddiff 14 (p<0.05, Figure 5E). The amount of β1-integrin first slightly increased on ddiff 7, followed by a small decrease on ddiff 8 and ddiff 14 (Figure 5F). FA maturation is mainly due to clustering of integrins and FA proteins. Therefore, soluble (S) and cytoskeletal bound (C) protein fractions were analyzed separately. Indeed, the C:S ratio of vinculin increased from 0.12±0.02 on ddiff 1 to 0.17±0.06 on ddiff 14 (Figure 5H), suggesting the incorporation of vinculin into FA complexes during the early phases of CPC differentiation. Although the C:S ratio of vinculin on ddiff 7 and 8 seem to be slightly higher compared to ddiff 14, the total amount of vinculin present in the cells was lower at these days, indicating that the formation of FA still increases towards ddiff 14. Moreover, integrin clustering at the cell membrane upon CPC differentiation was shown by an increase in the C:S ratio of β1-integrin from 0.83±0.50 on ddiff 1 to 2.50±1.21 on ddiff 14 (Figure 5 I). Surprisingly, pfak was only present in the S fractions, possibly because the binding of pfak to the FA complex is not strong enough and already loosens during the protein extraction and washing steps. Alternatively, the levels of pfak in the C-fraction were too low to detect with Western blot analysis. However, the level of pfak normalized to total FAK increased from 2.55±0.75 on ddiff 1 to 3.93±2.36on ddiff 14 (Figure 5J), indicative of the activation of the mechanotransduction pathway. Finally, the cytoskeletal development during CPC differentiation was further examined by determining the ratio of fibrillar actin (F-actin) to globular actin (G-actin), derived from the cytoskeletal and soluble protein fractions, respectively. The F- to G-actin ratio showed an increasing trend from 0.71±0.14 on ddiff 1 to 1.79±0.27 on ddiff 14 (p<0.001, Figure 5G), indicating a shift from globular to fibrillar actin upon cardiomyogenic differentiation of CPCs, and supporting our data from immunofluorescent stainings, showing the formation of actin fibers during the differentiation process. Altogether, these results suggest that the mechanosome of CPCs is activated during early cardiomyogenic differentiation. 37
 Immunofluorescent stainings of vinculin (green) and actin (red) of CPCs during induced cardiomyogenic differentiation.")
47 Figure 5: The CPC mechanosome components are activated during early differentiation. A-D) Immunofluorescent stainings of vinculin (green) and actin (red) of CPCs during induced cardiomyogenic differentiation. The mechanosome starts to form at d diff 8 (C) and is fully developed at d diff 14 (D). E-J) Western blots for mechanosome components from total protein extracts of CPCs during differentiation. The total amount of vinculin (E) increased with differentiation, while the levels of total β1-integrin stay roughly stable; cytoskeletal to soluble protein fractions (C:S) of vinculin (H) and β1-integrin (I) increased with differentiation of CPCs, as well as the ratio of fibrillar to globular actin (G, F-actin:G-actin); the amount of pfak over the amount of total FAK also increased during differentiation (J). Scale bar is 100 μm. Data are presented as mean fold change ± SD, (d diff1: n=6, d diff 7-14: n=4). *p<0.05, **p<0.01 and ***p< Discussion In this study, we investigated the mechanosensitivity of CPCs, and found a relationship between their differentiation state and their ability to respond to mechanical stimuli. Human CPCs are a promising cell source/target for cardiac regeneration strategies. Understanding the interaction of CPCs with their niche is therefore essential to identify target mechanisms for possible therapeutic strategies; in particular, mechanical stimuli can have a deep influence on cell behavior in the continuously strained heart. Here, we describe for the first time that the mechanosensitivity of CPCs is developed upon early cardiomyogenic differentiation, concomitantly with the maturation of the mechanosome. To investigate how CPCs respond to biomechanical stimuli, we determined the orientation of undifferentiated and predifferentiated cells upon applied uniaxial cyclic strain. We demonstrate that CPCs are able to respond with 38
48 strain avoidance to external mechanical stimuli only upon early differentiation. This is in agreement with numerous previous studies and models, describing strain avoidance as a behavior that allows cells to be minimally perturbed by the applied strain, and thus minimize the energy expenditure Strain avoidance after application of 10% strain at 1Hz has been reported in several other stem cells, e.g. adipose-derived stem cells 191, bone marrow mesenchymal stem cells, 189 and bone marrow-derived progenitor cells 190. These cells showed clear stress fiber development 172,190,191,211, although detailed information regarding the presence of the other components of the mechanosome in these other stem cells is lacking. In contrast to our study, French et al. described strain avoidance of rat c-kit + CPCs after 24 hrs of applied cyclic strain (5, 10 and 15%). They show that rat c-kit + CPCs give the strongest strain avoidance response when cultured on fibronectin and collagen I 127. However, they do not look at collagen IV, which is the coating substrate used in our study since it is the main component of the cardiomyocyte basement membrane. Moreover, the strain avoidance described by French and coworkers, that we only observed in predifferentiated cells, might depend on the different cell source and type used. Overall, this may suggest that the strain response of early cardiac progenitors is dependent on a delicate balance between environmental factors and the differentiation state of the cells. In cardiomyocytes, which are continuously exposed to mechanical cues, response to cyclic strain is dependent on the time of cultivation and on the straining protocol. In rat cardiomyocytes, 20% elongation at 0.5Hz applied 2 hrs 212 or 3 hrs 213 after seeding, led to alignment parallel to the strain direction. Cardiomyocytes aligned parallel to the direction of the stretch also when exposed to strain of 120% at 0.5 Hz applied 12 hrs after seeding, whereas when previously cultured for 24 hrs they oriented perpendicularly to the strain direction after one day of strain 214. However, cardiomyocytes failed to change their orientation when strained 72 hrs after seeding 213. Unfortunately, the different strain amplitudes and rates in these previous studies make it difficult to compare the mechanoresponse of cardiomyocytes to that of undifferentiated and predifferentiated CPCs. Additionally, it is important to mention that the presented results were obtained in a 2D setup, which is rather different from the in vivo environment. In this study, the different mechanoresponse of undifferentiated and predifferentiated CPCs is attributed to the development of the mechanosome, which is linked to the cardiac differentiation status of CPCs. We show that although undifferentiated CPCs express β1- integrin as well as FA proteins vinculin and (p)fak, undifferentiated cells do not form stable FA complexes. Upon differentiation, formation of focal adhesion complexes and cytoskeleton development is initiated as shown by clustering of β1-integrin and recruitment of vinculin at the FA sites, and a shift from globular to fibrillary actin. In line with our findings, the connection between differentiation and stress fiber development, has previously been described 215,216. Jacot et al. 217 suggested that pluripotent stem cells start to express and activate their mechanotransduction pathway during the process of cardiogenesis. Related to other cell types, a study by Mack et al. 215 showed that Rho kinase (ROCK), whose activation by RhoA is responsible for actin polymerization and stress fiber formation, has a crucial role in the regulation of differentiation markers in smooth muscle cell. Similarly, it has been shown that β1-integrin deficient cardiac precursors display impaired cardiac specification and formation of sarcomeric structures 218. These previous studies support our suggestion that differentiation is 39
49 responsible for the maturation of the mechanosome in CPCs. Furthermore, in undifferentiated CPCs the mechanosome development is neither induced by changed mechanical properties of the substrate, nor by applying strain. The more prominent mechanosome observed in predifferentiated CPCs seeded on glass as compared to silicone represents a behavior that has extensively been described by a number of papers, showing that a stiffer seeding substrate induces the formation of more static (larger) FAs and a more organized cytoskeleton Moreover, each seeding substrate might induce a different organization of the collagen coating, and therefore different distribution of the cellular adhesion sites 219. However, undifferentiated CPCs did not develop FAs and SFs on none of the used substrates, and the difference between the mechanosome in the two cell types was maintained both on silicone and on glass. This indicates that undifferentiated CPCs are not able to develop a mechanosome, regardless of the seeding substrate. We found that cyclic loading did not induce the mechanosome development in undifferentiated CPCs. Similarly, cyclic strain did not seem to enhance the maturation of FAs and SFs even in predifferentiated cells. Moreover, when CPCs were cultured statically on the Bioflex well-plate (unstrained cells), both undifferentiated and predifferentiated cells seemed to develop slightly larger, more static adhesions than strained samples, as demonstrated by clustering of vinculin (at day1 only), ILK and, to a minor extent, pfak. Although it is known that application of force typically results in FA strengthening in a seconds-to-minutes timescale (catch-bonds) , it appears that in cells that are cyclically strained, continuously and relatively quickly (0.5Hz), the formation of stable adhesions is impaired by the mechanical loading, as FAs undergo a faster assembly/disassembly process 206,223. Taken together, our data show that undifferentiated CPCs do not form stable FAs and lack a developed actin cytoskeleton, and have an impaired mechanosome. The mechanosome development in CPCs is intrinsically connected with differentiation, and cannot be induced neither by changing the substrate type or composition, nor by applying cyclic strain. With differentiation, CPCs start to develop SFs and consequently mature focal adhesions, along with the ability to respond to cyclic strain, indicating mechanosensing development. The presented results might suggest that resident undifferentiated CPCs are shielded from the external mechanical cues due to the lack of mechanosome. This might be a protective effect of the undifferentiated CPCs themselves. With an impaired mechanosensing apparatus these cells may remain in undifferentiated and quiescent state while continuously exposed to mechanical cues in the beating heart. Predifferentiated CPCs instead are more prone to align and integrate with the host cells and tissue, and might thus represent a more promising source for cardiac regeneration therapy. Conclusions In conclusion, our findings indicate that the cell mechanosensing apparatus in direct contact with the ECM, or mechanosome, is developed during cardiomyogenic differentiation in CPCs, regardless of the used substrate. Stress fiber development and focal adhesion maturation 40
50 enable predifferentiated cells to sense and respond to uniaxial cyclic strain, by aligning perpendicularly to the direction of the strain after two days. On the contrary, the lack of a developed mechanosome in undifferentiated CPCs shields them from external mechanical stimuli. Moreover, actin stress fiber formation and FA maturation cannot be induced in undifferentiated CPCs only by mechanical stimulation, but it requires (early) cardiomyogenic differentiation. Acknowledgements Dr. Sotiris Korossis from Hannover Medical School and Dr. Paola Aguiari from University of Padova are thankfully acknowledged for the fruitful discussions. This research was funded by the People Program (Marie Curie Actions) of the European Union s Seventh Framework Program FP7-People-2012-ITN TECAS, under grant agreement n Part of the research described here was supported by a grant of the Dutch Heart Foundation (DHF-2014T013 and DHF-2008T092). 41
, GATA4 (B), Mef2c (C), Myocardin (D), cardiac troponin T (E), and connexin 43 (F) in the three CPC isolations (L3, L6, and L9TB) during the first two weeks of cardiomyogenic differentiation, as")
51 Appendix to chapter 2 Figure S1: Predifferentiated CPCs are in an early differentiated state. A-F) Expression of cardiac transcription factors Nkx 2.5 (A), GATA4 (B), Mef2c (C), Myocardin (D), cardiac troponin T (E), and connexin 43 (F) in the three CPC isolations (L3, L6, and L9TB) during the first two weeks of cardiomyogenic differentiation, as determined by qpcr. Increased gene expression of all cardiac markers, and in particular of myocardin, troponint and connexin43 was observed (D-F). Data are presented as mean fold change ± SEM, n=2 per cell isolation per time point. *p<0.05; **p<0.01; ***p< G-N) Immunofluorescent images of the cardiac transcription factor Nkx 2.5 (G-J, red) and the sarcomeric component α-actinin (K-N, green) in L9TB CPCs during the first two weeks of differentiation. Both Nkx2.5 and α-actinin were expressed at day7, day8 and day14 (H-J), although no sarcomeric striations could be observed. However, α-actinin clusters close to the cell membrane were visible at day14 of differentiation (N), indicating early signs of sarcomeric organization. Nuclei are stained in blue (DAPI). Images are representative for all three cell isolations. Scale bar is 50μm. 42
52 Figure S2: Cyclic strain does not affect the cardiac lineage commitment of undifferentiated and predifferentiated CPCs. A-H) Gene expression of cardiac markers was not affected by the strain; at day1 upregulation of the cardiac transcription factor Nkx2.5 (C, n.s.) and of the cardiomyocyte marker ctnt (G, p<0.05) was observed in strained undifferentiated CPCs (n=3). This induction might be attributed to cells that spontaneously start to differentiate, especially in areas characterized by high cell density. These cells might therefore become sensitive to the applied strain, and change their alignment with the stretch. Moreover, due to the very low expression of troponint in undifferentiated CPCs, the large increase in the expression of this gene at day1 of strain can be seen as an on/off response, for which the spontaneously differentiated cells might partially be responsible. 43
 No significant differences are present among unstrained undifferentiated or predifferentiated CPCs, nor among B) strained undifferentiated or predifferentiated CPCs over time, indicating that time")
 and actin stress fibers (red) in undifferentiated cells (d diff1, A, E), and")
53 Figure S3: The mechanoresponse of CPCs is not induced over time or by the cyclic strain per se, unless cardiomyogenic differentiation occurs. A) No significant differences are present among unstrained undifferentiated or predifferentiated CPCs, nor among B) strained undifferentiated or predifferentiated CPCs over time, indicating that time or cyclic strain alone cannot induce mechanosensitivity in CPCs. N=16-23 from 3 independent experiments. Figure S4: The mechanosome is developed with differentiation in L3 and L6 CPCs. Immunofluorescent stainings of L3 and L6 CPCs during cardiomyogenic differentiation show absence of vinculin (green) and actin stress fibers (red) in undifferentiated cells (d diff1, A, E), and progressive maturation of the mechanosome at d diff7 (B, F), d diff8 (C, G) and d diff14 (D, H). Scale bar 50 μm. 44
54 CHAPTER 3 Selective inhibition of mechanosome components disrupts stress fiber integrity and mechanoresponse of predifferentiated cardiac progenitor cells
55 Abstract Cardiac progenitor cells (CPCs) have emerged as a promising cell source for cardiac regeneration strategies. CPCs reside in the human fetal and adult heart, and their interaction with the mechanically active cardiac environment may influence their regenerative potential. However, the mechanisms driving CPC mechanoresponse remain unclear. Upon early cardiomyogenic differentiation, CPCs develop their mechanosome, i.e. the mechanosensing apparatus in direct contact with the extracellular matrix, needed to respond to external mechanical stimuli. At the same time, early differentiated CPCs (predifferentiated) become able to reorient perpendicular to applied cyclic strain on 2D substrates (strain avoidance). Yet, the contribution of the mechanosome components in the CPC response to external mechanical stimuli is unknown. Here, we aimed to unravel the role of the mechanosome in the strain avoidance response of predifferentiated CPCs. Therefore, the mechanosome activity was manipulated in these cells by inhibiting the key mechanosome components: integrinβ1, integrin-linked kinase (ILK), and Rho-kinase (ROCK). Inhibition of the mechanosome components resulted in impaired cellular mechanoresponse to cyclic strain, shown by a weaker strain avoidance response. Furthermore, inhibition of the different mechanosome components affected the integrity of actin stress fibers, and led to a comparable reduction in the strain avoidance response of predifferentiated CPCs, regardless of the presence of vinculin clusters. The results indicate that, similarly to non-progenitor cells, the mechanosome and especially stress fibers play a crucial role in CPC mechanosensing and mechanoresponse. 46
56 Introduction Microenvironmental stimuli, such as mechanical and molecular signals coming from the extracellular matrix (ECM), can determine cell behavior and functions. In the cardiac tissue, cells are constantly subjected to the cyclic strain provided by the heart beating. Understanding the influence of such mechanical stimuli on CPCs is needed to improve current CPC-based approaches of heart repair and regeneration. Until recently, the heart was believed to be a terminally differentiated organ, and cardiomyocyte loss following cardiac injury to be irreversible. However, studies from the past couple of decades have revealed not only cardiomyocyte renewal (0.4 1% annual turnover rate 5 ), but also the presence of endogenous cardiac progenitor cell (CPC) pools 6,224. CPCs have shown promising characteristics for cardiac repair and regeneration, such as the ability to differentiate into all cardiac lineages and the capacity to produce and remodel ECM 15. Despite the beneficial effects on cardiac function upon CPC transplantation in the injured heart 17,180, long term effects are limited because of low integration, survival, and differentiation of the CPCs injected in the hostile environment of the infarcted myocardium 8,17. Cell-ECM transmission of mechanical signals occurs via the mechanosome, which is the complex of cellular structures in contact with the ECM that participate in cell mechanotransduction 225. The mechanosome consists of focal adhesion (FA) proteins, multiprotein complexes located at the cell-ecm interaction sites 196,226,227, and actomyosin stress fibers (SFs), which are the contractile units of the cell 123. Within the mechanosome, several functional layers can be identified: i) a signaling layer, where integrins initiate the transmission of mechanical cues; ii) a force transduction layer, where FA proteins such as vinculin and integrin-linked kinase (ILK) physically connect integrins and actin fibers; iii) and an actin regulatory layer, including proteins that regulate SF binding such as α-actinin and zyxin 122,196,228. The contractile elements of the mechanosome are SFs, myosin II-containing actin filaments whose assembly and contractility can be regulated through RhoA and its effector Rhokinase (ROCK) 123,229. In chapter 2, we describe that CPCs develop a mature mechanosome following early cardiomyogenic differentiation (predifferentiated CPCs) 121. In parallel to the mechanosome development, upon early differentiation CPCs acquire the ability to respond to applied cyclic stretching by changing their orientation. Whereas undifferentiated CPCs showed little to no reorientation in response to uniaxial cyclic strain, predifferentiated CPCs reoriented perpendicular to the stretching direction 121, a behavior known as strain avoidance 230,231. Here, the mechanosome activity of predifferentiated CPCs was manipulated, by inhibiting components from different functional layers (integrinβ1, ILK, ROCK). Manipulation of the different mechanosome components affected the integrity of SFs. All three inhibitors also led to a comparable reduction in the strain avoidance response of predifferentiated CPCs, regardless of the presence of vinculin clusters. The results indicate a crucial role for the mechanosome, and particularly SFs, in the mechanoresponse of human CPCs, and provide fundamental knowledge that can ultimately aid the improvement of CPC-based research and clinical application. 47
57 Materials and methods Culture of cardiac progenitor cells Human fetal Sca-1 + CPCs were isolated and immortalized by lentiviral transfection of htert abd BMI-1 as previously described 39,40. The immortalized cell line (L9TB CPCs) were cultured in SP++ growth medium consisting of M199 (Gibco)/EGM2 (3:1) supplemented with 10% (v/v) fetal bovine serum (FBS) (Greiner bio-one), 1% (v/v) non-essential amino acids (Gibco), and 1% (v/v) penicillin/streptomycin (Lonza) on 0.1% (w/v) gelatin (Sigma Aldrich)/PBS (Sigma) coated flasks. Early cardiomyogenic differentiation Early cardiomyogenic differentiation of CPCs was induced according to established protocols [Smits 2009]. Briefly, after reaching about 100% confluency (day-14), cells were treated with 5 μm 5 -azacytidine (5 -aza) (Sigma) for three consecutive days in differentiation medium (IFFDIFF), consisting of IMDM/Ham-F12 (1:1) (Gibco) with 2% (v/v) horse serum, 1% (v/v) P/S, 1% (v/v) NEAA, 10-4 M ascorbic acid (Sigma) and 1% (v/v) insulin-transferrin-selenium supplement (Lonza). After 72 hours, medium was changed followed by TGF-β1 (1 ng/ml) (Sigma) stimulation. IFFDIFF medium supplemented with TGF-β1 was changed every three days until seeding (Figure 1, day-1). The differentiation protocol was carried out in plastic culture flasks for 14 days before strain application, and verified by gene expression prior to strain exposure. Manipulation of mechanosome activity In order to study the role of the mechanosome in predifferentiated CPCs mechanoresponse, the activity of selected mechanosome components (integrinβ1, ILK, ROCK) from different functional layers was inhibited. Used inhibitors and their concentrations are listed in Table 1. Inhibitors were added to the cell culture medium at the moment of cell seeding (day-1). Since all inhibitors are dissolved in dimethyl sulfoxide (DMSO), same DMSO volumes were added to control samples. Cyclic stretching Cyclic strain was applied with a Flexcell FX-5000 TM Tension System (Flexcell International Corporation). At day-1, predifferentiated CPCs were seeded on collagen IV-coated flexible membranes of Bioflex plates (Flexcell International) with a density of 3x10 4 cells/cm 2. Predifferentiated CPCs were statically cultured for 24 hrs to allow attachment to the membrane, after which (day0) uniaxial cyclic strain (10%, 0.5Hz, sine wave) was applied by stretching the membranes on rectangular posts oriented along the 90 direction. Cells were cyclically stretched for up to 48 hrs (day 2) along the 0 direction. Cell orientation was analyzed at day2 (Figure 1). Static samples (unstrained) were cultured on Bioflex plates under the same conditions. Gene expression analysis Total RNA was isolated from one sample per group per experiment using the Qiagen RNAeasy isolation kit according to manufacturer s instructions (Qiagen). cdna was synthesized with
58 ng RNA per sample using M-MLV Reverse Transcriptase (Bio-Rad) and subjected to qpcr using iqtm Green Supermix and the CFX384 detection system (Bio-Rad). Primer sequences for qpcr and annealing temperatures are presented in Table 1. Immunofluorescence labeling and confocal imaging Samples were washed three times with Ham-F12 medium (Gibco) at 37 C, fixed in 3.7% (v/v) formaldehyde (Merck) for 15 min, washed three times with PBS, and permeabilized with 0.5% (v/v) Triton X-100 (Merck) in PBS for 5 min. The Bioflex membranes were cut out of the plates with a scalpel, using the center of the membrane for immunofluorescence staining. Nonspecific antibody binding was blocked with 10% (v/v in PBS) horse serum incubation for 45 min. Cells were then incubated with primary antibodies in 1% (v/v in PBS) horse serum for 1.5 hrs. Cells were rinsed with PBS four times for 5 min and incubated for 1 hr with secondary antibodies, and phalloidin for visualization of F-actin, in PBS. All used antibodies and dyes are listed in Table 2. Nuclei were stained with DAPI for 5 min, after which cells were again rinsed four times with PBS and mounted on microscopy glass coverslips with Mowiol. All incubation steps were performed at room temperature. Quantification of cell orientation CPCs were incubated for 20 min with 1 μg/ml calcein AM (Sigma). Only cells in the central part of the wells, where the strain field was homogenous according to previous in-house calibration, were considered. Cell orientation was quantified with Fiji, using the Directionality plug-in ( based on the Fourier transform (FFT) of each image by means of fitting a Gaussian to the FFT signal, measuring its peak position. For each group, 2 to 3 images were analyzed for each biological replicate (n=4-5). Statistical analysis All results are presented as mean ± SD. Statistical analysis was performed by Kruskal-Wallis test followed by Dunn s post-hoc test, or student s t-test where appropriate. Analysis of cell orientation distribution was performed with an in-house developed Matlab script, where each distribution is described by three parameters: peak location (mu), peak width (std) and distribution anisotropy (ani) 232. Significance was assumed for p<0.05; all statistical analyses were conducted with GraphPad Prism
59 Table 1: List of used pharmacological inhibitors. Protein Inhibitor Cat no. Final concentration Integrin β1 P5D2 (Abcam) ab μg/ml ILK Cpd-22 (Merck Millipore) μg/ml ROCK Y27632 (Sigma) Y μg/ml Table 2: List of primer sequences and annealing temperatures. Gene Primer sequences Ta Nkx 2.5 Myocardin Troponin T Notch2 Jagged1 HES 1 HEY 1 Fw: CCCCTGGATTTTGCATTCAC Rv: CGTGCGCAAGAACAAACG Fw: AGATGGATGAACTCCTGGAC Rv: AGT TGGACTTCGGGAAGATC Fw: CGACGAGAGGAGGAGGAGAAC Rv: CCGCTCTGTCTTCTGGATGTAAC Fw: GGCTATGAATGCCAGTGTGC Rv: GGATCGGGGTCACAGTTGTC Fw: AATGGC TACCGGTGTGTCTG Rv: CCCATGGTGATGCAAGGTCT Fw: AGTGAAGCACCT CCGGAAC Rv: CGTTCATGCACTCGCTGA Fw: TGGATCACCTGAAAATGCTG Rv: CGAAATCCCAAACTCCGATA 60 C 60 C 60 C 60 C 60 C 62 C 60 C Table 3: List of used antibodies and dyes. Antigen Source Cat. no Isotype Label Species Dilution Vinculin Sigma Aldrich V9131 IgG1 - Mouse 1:400 Phalloidin Sigma Aldrich P ATTO488 Amanita phalloides 1:200 Mouse IgG Miilipore A Alexa555 Goat 1:200 50
60 Results Early cardiomyogenic differentiation of CPCs Early cardiomyogenic differentiation of CPCs was carried out for 14 days (Figure 1A). Early differentiation (predifferentiated CPCs) was verified by gene expression analysis of cardiac markers. Undifferentiated CPCs showed expression of the cardiac transcription factor Nkx2.5, whereas low expression of the cardiomyocyte marker myocardin and no expression of troponin T (TnT) were observed (Table S1). Predifferentiated CPCs exhibited a minor decrease in Nkx2.5 expression, whereas increased RNA levels of myocardin and ctnt were detected (Figure 1B). Figure 1: Timeline of the experiment and early cardiomyogenic differentiation of predifferentiated CPCs. A) CPC cardiomyogenic differentiation protocol was carried out for 14 days, until the cells reached an early differentiated state (predifferentiated CPCs). At day-1, cells were seeded on Bioflex plates and let adhere for 24 hrs. At day0 uniaxial cyclic strain (10%, 0.5 Hz) was applied for 48 hrs (day2). B) The cell differentiation state was verified by gene expression analysis of cardiac markers. The modest decrease of Nkx2.5, increase of myocardin and troponint (TnT) indicate early cardiomyogenic CPC differentiation. Mean±SD (n=3 biological replicates); ****p< Mechanosome manipulation hinders the strain avoidance response of predifferentiated CPCs To elucidate the role of the mechanosome in the mechanoresponse of predifferentiated CPC, we evaluated the cell strain avoidance response while blocking proteins operating in different functional layers of the mechanosome complex. Integrins are transmembrane proteins in direct contact with the ECM, and thus represent the first sensors of external mechanical stimuli. ILK binds integrinβ1 and plays a crucial role as a structural FA protein, whereas ROCK is responsible for the development and activation of stress fibers. After 48 hrs of uniaxial cyclic strain (day2), control samples (DMSO) displayed a distinct strain avoidance response (Figure 2A), as shown by the cell orientation distribution peaked at 90 (B). The cell perpendicular reorientation was hampered by blocking either integrinβ1 (PD52; Figure 2C, D), ILK (Cpd-22; Figure 2E, F), or ROCK (Y27632; Figure 2G, H) (Figure S2). 51
 show a clear strain avoidance in response after 48 hrs of uniaxial")
61 Figure 2: Blocking integrinβ1, ILK, or ROCK reduces the strain avoidance response of predifferentiated CPCs. Predifferentiated CPCs (DMSO) show a clear strain avoidance in response after 48 hrs of uniaxial 52
62 cyclic strain (day2, A, B). Inhibition of the activity of integrinβ1 (PD52, C, D), ILK (Cpd-22, E, F), and ROCK (Y27632, G, H) hinders the strain avoidance response of predifferentiated CPCs (n=5). Scale bar 400 μm. Blocking different mechanosome components disturbs the mechanosome development with distinct dynamics To qualitatively determine the effect of the mechanosome manipulation on the development of the mechanosome itself, we conducted confocal imaging to visualize the distribution and clustering of vinculin, a FA protein well known for its role in cell mechanosensing, and the extent of SF development and bundling. At both day0 and day2, large vinculin cluster and thick SFs were observed in control predifferentiated CPCs (DMSO, Figure 3A-C). At day0, the presence of the inhibitors of integrinβ1 led to altered distribution of vinculin, which seemed to be more present in the cell cytoplasm and to form smaller clusters, mainly localized at the cell edges (Figure S3D, F). Conversely, integrinβ1 inhibition did not appear to affect the SF development (Figure S3E, F). Similarly, at day0, ILK inhibition left both vinculin and SFs intact (Figure S3G-I). On the contrary, the presence of ROCK inhibitor Y27632 led to smaller vinculin clusters and thinner SFs already at day0 (Figure S3J, K, L). The mechanosome manipulation exhibited much stronger effects at day2; at this time point, vinculin clustering was impaired by inhibition of integrinβ1 (Figure 3D) and ROCK (Figure 3J), whereas it was not affected by ILK inhibition (Figure 3G). At day2, actin SFs appeared thinner in all treated samples, as compared to the control (Figure 3E, H, K), indicating that inhibition of integrinβ1, ILK, and ROCK all led to disruption of SF integrity over time. 53
 show large vinculin clusters (A) and developed SFs (B).")
63 Figure 3: Inhibition of integrinβ1, ILK, and ROCK affects SF development in predifferentiated CPCs at day2. Predifferentiated CPCs after 72 hrs of culture (A-C) show large vinculin clusters (A) and developed SFs (B). Integrinβ1 blocking by PD52 (D-F) leads to increased cytoplasmic distribution and smaller clusters of vinculin (D), and thinner SFs (E). ILK inhibition by Cpd-22 (G-I) does not alter vinculin distribution (G), while SF integrity is affected (H). ROCK inhibition with Y27632 (J-L) results in smaller and less numerous vinculin clusters (J) and disrupted SFs (K). Scale bar 50μm. 54
64 Discussion Mechanical stimuli can have a profound influence on cellular behavior, particularly in a mechanically active environment such as the cardiac tissue. Here, we aimed at investigating the role of the mechanosome, i.e. the cell mechanosensing apparatus in contact with the ECM, in the mechanoresponse of CPCs. In a previous study, we showed that the mechanosome is developed upon early cardiomyogenic differentiation of CPCs, along with CPC strain avoidance response. We investigated the effects of manipulating the mechanosome of predifferentiated CPCs by inhibiting the activity of specific components operating in different functional layers of the mechanosome complex: integrinβ1, ILK, and ROCK. Inhibition of the investigated mechanosome components led to reduced strain avoidance response, as shown by the broader cellular orientation distribution as compared to control samples (Figure S1). This suggests that the activity of all three proteins, integrinβ1, ILK, and ROCK, is required for an efficient mechanoresponse of predifferentiated CPCs. In agreement with our data, others have reported impairment of cell mechanoresponse following inhibition of focal adhesion kinase (FAK) 233 or ROCK 234. The mechanosome manipulation did not affect the early differentiated state of predifferentiated CPCs, indicating that the hindered strain avoidance response is not caused by cell de-differentiation. Moreover, all three blocking agents eventually resulted in impaired SF development, although their effects followed distinct dynamics. As known, integrins are FA proteins in direct contact with the ECM, whose activation induces a conformational change that allows exposure of vinculin-binding sites 235,236. Our data indeed showed that inhibition of integrinβ1 activity resulted in impaired vinculin clustering already at day0. However, SF maturation was only affected over time (day2), which might be due to the indirect link of integrins to actin and the slow integrin turnover dynamics 237. Similarly, inhibition of ILK activity via Cpd-22 did not seem to cause changes in either vinculin distribution or SF development at day0. At day2 however, vinculin distribution was still unaffected, whereas thinner SFs were observed. Besides its fundamental role in cardiomyocyte survival, migration and force transmission 200,201, ILK clustering in CPCs only occurs upon early cardiomyogenic differentiation (predifferentiated CPCs), together with the development of SFs 121. On the contrary, small vinculin clusters can already be observed in undifferentiated cells 121, indicating that the translocation of ILK to FAs takes place at a later stage than that of vinculin. This likely explains the fact that vinculin localization is independent of ILK activity. The fastest and most evident effect on vinculin and SFs was provided by ROCK inhibitor Y27632, in agreement with previous reports 221,229,234. Activation of the Rho-ROCK pathway directly leads to SF bundling and contractility 123,238 and FA formation 192, thereby explaining the immediate effects of its inhibition on the mechanosome stability. An interesting parallel can be found between the strain avoidance impairment and the disruption of SFs integrity (Figure 4). Surprisingly, the absence of developed SFs after ILK inhibition hindered proper cellular reorientation regardless of the presence of stable vinculin clusters. This seems to suggest that the key player in CPC mechanoresponse is represented by 55
65 SF activation. The proper development of SFs is however dependent on the activity of (at least) integrinβ1, ILK and ROCK. It can be further speculated that the whole mechanosome is required for adequate mechanosensing and mechanoresponse of CPCs. In conclusion, our results shed light on the fundamental role of the mechanosome, and particularly SFs, for the mechanoresponse of human CPCs. Further research is needed to gain a deeper understanding of the relevance of the mechanosome composition and dynamics on CPC mechanosensing and mechanoresponse. Figure 4: Manipulation of the mechanosome activity disturbs the mechanosome development and mechanoresponse of predifferentiated CPCs. Schematic representation of the mechanosome of predifferentiated CPCs (control). Inhibition of the activity of different mechanosome components has distinct effects on the maturation of vinculin and stress fibers (SFs). However, all three inhibitors hindered the development of SF and the cellular strain avoidance response at day2. 56
66 Acknowledgements This research was funded by the People Program (Marie Curie Actions) of the European Union s Seventh Framework Program FP7-People-2012-ITN TECAS, under grant agreement n and the FP7-PEOPLE-2013-CIG Stressfate, under grant agreement n Dylan Mostert, MSc is kindly acknowledged for her help in designing and conducting the experiments. 57
67 Appendix to chapter 3 Figure S1: Mechanosome activity manipulation does not affect the differentiation state of predifferentiated CPCs. Scatter plots show the gene expression of cardiac transcription factor Nkx2.5, and cardiomyocyte markers myocardin and troponint in predifferentiated CPCs in response to inhibition of integrinβ1 (int b1), ILK, and ROCK. The early differentiated state of the cells does not seem to be affected by the inhibitors. Mean±SD (n=3-5). 58
68 Table S1: Cycle threshold values of analyzed samples and markers. sample GAPDH NKX 2.5 Myocardin ctnt DMSO day0 undifferentiated intb1 day0 undifferentiated 17,20 ± 1,12 21,91 ± 1,23 28,81± 4,04 35,52 ± 0,10 17,13 ± 1,32 21,93 ± 1,39 29,35 ± 4,85 36,22 ± 0,93 ILK day 0 undifferentiated 19,25 ± 2,53 24,13 ± 2,60 30,80 ± 2,14 34,24 ± 4,02 ROCK day0 undifferentiated DMSO day0 predifferentiated intb1 day0 predifferentiated 18,29 ± 2,04 23,15 ± 2,11 33,25 ± 0,59 34,90 ± 1,47 18,23 ± 0,45 23,69 ± 0,25 27,59 ± 1,25 28,24 ± 1,90 17,34 ± 0,63 23,04 ± 1,42 28,00 ± 1,35 29,57 ± 0,57 ILK day0 predifferentiated 17,31 ± 0,91 23,39 ± 1,70 28,09 ± 2,03 30,32 ± 0,33 ROCK day0 predifferentiated DMSO day2 predifferentiated intb1 day2 predifferentiated 17,53 ± 0,89 23,51 ± 1,67 29,09 ± 3,45 30,03 ± 0,19 18,02 ± 0,54 23,40 ± 0,40 27,09 ± 0,89 26,24 ± 1,90 17,57 ± 0,18 22,91 ± 0,33 28,14 ± 2,50 25,83 ± 1,20 ILK day2 predifferentiated 19,09 ± 2,60 22,68 ± 0,22 27,37 ± 1,67 25,01 ± 0,13 ROCK day2 predifferentiated 17,62 ± 0,11 23,09 ± 0,10 28,61 ± 3,29 26,57 ± 1,76 59
69 Figure S2: Mechanosome manipulation reduces the strain avoidance response of predifferentiated CPCs. Cyclically stretched predifferentiated CPCs show a narrower distribution of the cell orientation peak values (mu ; A, B, blue circles). Although no significant differences are observed, inhibition of the activity of integrinb1, ILK, and ROCK leads to broader localization of the peak (mu; A) and wider peak width (std; B), whereas the distribution anisotropy seems unchanged (ani; C). Mean; n=2-5 independent experiments. 60
 show large vinculin clusters (A) and developed SFs (B).")
70 Figure S3: Inhibition of integrinβ1, ILK, and ROCK differentially affects the mechanosome of predifferentiated CPCs at day0. Predifferentiated CPCs after 24 hrs of culture (A-C) show large vinculin clusters (A) and developed SFs (B). At day0, integrinβ1 blocking by PD52 (D-F) leads to increased cytoplasmic distribution of vinculin and reduced clustering (D), whereas the SFs remain unaltered (E). ILK inhibition by Cpd-22 (G-I) does not evidently alter the development of vinculin (G) or SFs (H). ROCK inhibition with Y27632 (J-L) results in smaller vinculin clusters (J) and thinner SFs (K). Scale bar 50μm. 61
71 62
72 CHAPTER 4 The role of cell density in the response of cardiac progenitor cells to cyclic strain This chapter is based on: Mauretti A., Bax N.A.M., Mostert D., Goumans M.J., Sahlgren C., Bouten C.V.C., The role of cell density in the response of cardiac progenitor cells to cyclic strain, submitted for publication.
73 Abstract The interactions of cardiac progenitor cells (CPCs) with the mechanically active cardiac environment are largely unknown. In order to improve current CPC-based cardiac therapies, there is need to understand if and how CPCs respond to environmental mechanical stimuli, such as the cyclic stretching provided by the beating heart. Cell-cell adhesions may be key players in this context, due to their recognized role in the transmission of mechanical signals between neighboring cells. Here, we show that increased cell density of human CPCs enhances the reorientation of CPCs perpendicular to applied uniaxial cyclic stretching (strain avoidance), and that both N-cadherin and Rho kinase (ROCK) activity are required for CPC strain avoidance behavior. Higher cell density did not affect CPC differentiation state, actin stress fiber and focal adhesion development, whereas it promoted N-cadherin clustering. Our findings provide new fundamental knowledge on the interactions of CPCs with their mechanical environment, and can ultimately contribute to optimize strategies of CPC-based cardiac regeneration. 64
74 Introduction In the mechanically active cardiac microenvironment, cardiac progenitor cells (CPCs) are continuously subjected to cyclic straining due to the heart beating. It is not well known how these cells sense and respond to such mechanical stimuli, while this is important in light of the potential role of CPCs in cardiac regeneration 15,39,40. Injection of CPCs in the injured myocardium generates beneficial effects, which are however limited to the short-term due to the massive cell loss, and the reduced survival and differentiation of engrafted cells 17,239. In order to optimize the clinical application of CPCs, there is the need to improve our understanding of their response to the cyclic stretching provided by the cardiac environment. Cell-cell adhesions play a pivotal role in cardiac tissue development, and can determine the cellular response to the pathological cardiac microenvironment 240,241. Therefore, here we investigated the role of cell density in the response of undifferentiated CPCs to cyclic straining. Cells receive mechanical stimuli from the ECM via integrin-mediated focal adhesions (FAs), and from neighboring cells via cadherin-mediated adhesions. Cadherin-based junctions are active cell mechanosensors 242. They allow force transmission and mechanotransduction , and can modulate stem cell mechanosensitivity to the substrate stiffness 246. N-cadherin is the main component of adherens junctions in the heart 247, and mediates mechanical coupling between adjacent cardiomyocytes 248,249. Yet, the role of cadherin-mediated junctions in the cellular response to cyclic stretching remains unknown. It has been recently shown that both cadherin- and integrin-based adhesions share a similar structure and organization 122,250. Both integrin- and cadherin-mediated adhesions are characterized by three functional layers: a signaling layer, in the proximity of the cell membrane; a force transduction layer, where proteins such as vinculin are localized and connect integrins/cadherins to actin fibers; and an actin regulatory layer, that modulates actin stress fibers 122. Stress fibers (SFs) are actomyosin bundles that provide contractility to the cell, and represent key players in cell mechanotransduction 123. An important role in SF regulation is played by the Rho Rho kinase (ROCK) pathway, that can directly promote SF assembly and its activation via phosphorylation of myosin light chain 123,229,251. Inhibition of ROCK via Y27632 was shown to reduce the development of SFs and FAs 229. Moreover, while cyclically strained cells typically reorient perpendicular to the stretching direction (strain avoidance response) 230,231, inhibition of ROCK led to cell reorientation parallel rather than perpendicular to the direction of the strain 234, indicating the crucial role of SF contractility in cell mechanosensing and mechanotransduction. In a previous study (chapter 2), we reported that human CPCs develop mature SF and FA complexes, (the mechanosome ), upon early cardiomyogenic differentiation 121. Furthermore, cyclically strained early differentiated CPCs displayed a more evident strain avoidance behavior than undifferentiated cells seeded at low cell density 121. Here, we show that a higher degree of cell-cell physical contact as a consequence of increased cell density leads to enhanced N-cadherin clustering and strain avoidance response of these cells, without affecting neither the cell phenotype, nor the mechanosome development. Inhibition of N-cadherin as well as that of ROCK in highly dense CPCs resulted in reduced strain 65
75 avoidance, suggesting that both cell-cell adhesions and ROCK activity are required for the reorientation response of human CPCs to cyclic stretching. Our findings provide new insights into the role of cell density in the interaction of CPCs with their mechanical environment, that can influence their regenerative potential. Materials and methods Culture of cardiac progenitor cells Human fetal Sca1 + cardiac progenitor cells (CPCs) were isolated and cultured as previously described 39,40. To create the L9TB cell line, CPCs were immortalized by lentiviral transfection of htert and BMI-1. Cells were cultured in SP++ growth medium consisting of M199 (Gibco)/EGM2 (3:1) supplemented with 10% (v/v) fetal bovine serum (FBS) (Greiner bio-one), 1% (v/v) nonessential amino acids (Gibco), and 1% (v/v) penicillin/streptomycin (Lonza) on 0.1% (w/v) gelatin (Sigma Aldrich)/PBS (Sigma) coated flasks. Cells were cultured with densities of 1x10 4 (dens1), 2.5x10 4 (dens2), and 5x10 4 (dens3) cells/cm 2 on collagen IV-coated (which is the main component of the cardiomyocyte basement membrane) Bioflex plates (Flexcell International Corporation), for 24 hrs (day0) or up to 72 hrs (day2). Gene expression analysis Total RNA was isolated from one sample per group using the Qiagen RNAeasy isolation kit according to manufacturer s instructions (Qiagen). cdna was synthesized with 250 ng RNA per sample using M-MLV Reverse Transcriptase (Bio-Rad) and subjected to qpcr using iqtm SYBR@ Green Supermix and the CFX384 detection system (Bio-Rad). Primer sequences for qpcr and annealing temperatures are presented in Table S1. Immunofluorescence labeling and confocal imaging Samples were washed three times with Ham-F12 medium (Gibco) at 37 C, fixed in 3.7% (v/v) formaldehyde (Merck) for 15 min, washed three times with PBS, and permeabilized with 0.5% (v/v) Triton X-100 (Merck) in PBS for 5 min. The Bioflex membranes were cut out of the plates with a scalpel, using the center of the membrane for immunofluorescence staining. Nonspecific antibody binding was blocked with 10% (v/v in PBS) horse serum incubation for 45 min. Cells were then incubated with primary antibodies in 1% (v/v in PBS) horse serum for 1.5 hrs. Cells were rinsed with PBS four times for 5 min and incubated for 1 hr with secondary antibodies, and phalloidin for visualization of F-actin, in PBS. All used antibodies and dyes are listed in Table 2. Nuclei were stained with DAPI for 5 min, after which cells were again rinsed four times with PBS and mounted on microscopy glass coverslips with Mowiol. All incubation steps were performed at room temperature. Total protein and protein fractions isolation To obtain total protein extracts, growth medium was removed, and cells were rinsed twice with F12-Ham medium. Cells were scraped off the well and centrifuged for 7 minutes at 1000 rpm at 4 C. The obtained cell pellet was lysed in RIPA buffer with addition of protease inhibitor cocktail 66
76 (1:100) and stored at -80 C. Protein fractions (cytosolic, membrane, nuclear, and cytoskeletal proteins) were isolated with the ProteoExtract Subcellular Proteome Extraction kit (Millipore), according to the manufacturer s instructions. Briefly, cells were rinsed twice with ice-cold wash buffer, scraped and centrifuged to obtain a cell pellet. Cytosolic proteins were extracted by incubation with extraction buffer I and 10 minute centrifugation at 4 C, after which the supernatant was collected, and the insoluble cell pellet was used for isolation of membrane and organelle proteins with extraction buffer II. The process was repeated, and nuclear proteins were extracted with extraction buffer III, with the addition of benzonase nuclease. Finally, by addition of extraction buffer IV the cytoskeletal proteins were extracted. All protein fractions were stored at -80 C until analysis. Western Blotting The level of SF development and N-cadherin expression and clustering was semi-quantitatively determined in total protein extracts and separated protein fractions (cytosolic, or soluble: S; membrane: MB) by Western blotting for actin and N-cadherin. Equivalents of 5 µg total protein were size fractioned on a 8% polyacrylamide gel and blotted onto a Hybond PVDF membrane (Immobilon-P, Millipore). Non-specific binding sites were blocked with 5% milk powder (ELK) in 0.1 % PBS/Tween20. Membranes were probed with primary antibodies (in 1% milk in 0.1% PBS/Tween) at 4 o C overnight, washed and subsequently incubated at room temperature with horse-radish peroxidase (HRP) labeled secondary antibodies for 1hr (for used antibodies and their dilutions see Table 1). For visualization, membranes were incubated with enhanced chemiluminescence substrate (Thermo Scientific Pierce), and bands were detected using the Proxima C16 Phi+ (Isogen) imager. Bands were quantified by determining the intensity of the bands (INT*mm 2 with global background correction) using ImageJ analysis software. Cyclic mechanical loading Cyclic strain was applied with a Flexcell FX-5000 system (Flexcell International Corporation). Prior to cyclic straining, CPCs were statically cultured for 24 hrs on collagen IV-coated Bioflex plates to allow attachment to the membrane. Starting at day0, uniaxial cyclic strain (10%, 0.5 Hz, sine wave) was applied for up to 48 hrs (day 2). Unstrained samples were cultured on Bioflex plates under the same conditions. The straining frequency of 0.5 Hz, lower that the physiological human cardiac beating frequency of 1 Hz, was chosen due to cell detachment from the substrate at higher frequencies. N-cadherin and ROCK inhibition To inhibit N-cadherin-mediated cell-cell adhesions and SF contractility, inhibitors of N-cadherin and of ROCK were used. N-cadherin inhibitor ADH-1 (1 μm, Hoelzel Diagnostika) was added to the culture medium 48 hrs after seeding (day1), since excessive exposure to the drug was shown to cause cell death. ROCK inhibitor Y27632 (10 μm, Sigma-Aldrich) was added at the moment of cell seeding, since the presence of this inhibitor was verified to not affect cell viability. 67
77 Quantification of cell orientation To quantify cell orientation, living CPCs were incubated for 20 min with 1 μg/ml calcein AM (Sigma) at day2 of cyclic stretching. Only cells in the central part of the wells, where the strain field was homogenous according to previous in-house calibration, were considered. Cell orientation was quantified with Fiji, using the Directionality plug-in ( based on the Fourier transform (FFT) of each image by means of fitting a Gaussian to the FFT signal, measuring its peak position. For each experimental condition, 3 to 10 images were analyzed (n=3-4 biological replicates). Statistical analysis All data are presented as mean ± standard deviation (SD). Statistical analysis was performed with Kruskal-Wallis test followed by a Dunn s post-hoc test for multiple comparisons, or with student s t-test where appropriate. Analysis of cell orientation distribution was performed with an in-house developed Matlab script, where each distribution is described by three parameters: peak location (mu), peak width (std) and distribution anisotropy (ani) 232. Significance was assumed for p<0.05; all statistical analyses were conducted with GraphPad Prism
78 Table 1: List of used antibodies and dyes. Antigen Source Cat. no Isotype Species Label Dilution vinculin Sigma V9131 IgG Mouse - 1:500 N-cadherin Sigma C3865 IgG Mouse - 1:200 phalloidin Sigma P Amanita phalloides ATTO 488 1:400 YAP Cell Signaling 4912S IgG Rabbit - 1:50 Mouse IgG Molecular Probes A21424 IgG (H+L) Goat Alexa Fluor 555 1:300 Rabbit IgG Jackson IgG (H+L) Donkey Alexa Fluor 647 1:300 DAPI Sigma-Aldrich D :500 Results Increased cell density enhances CPC strain avoidance response To assess if cell density affected the response of undifferentiated CPCs to mechanical stimuli, cell reorientation perpendicular to the main axis of the applied strain (strain avoidance) was determined. CPCs seeded at dens1 showed a very modest strain avoidance response at day2 ((1x10 4 cells/cm 2, Figure 1A, D), whereas at dens2 the perpendicular reorientation became more evident (2.5x10 4 cells/cm 2, Figure 1B, D). However, CPCs at dens3 exhibited a distinct strain avoidance response (5x10 4 cells/cm 2, Figure 1C, D). These results indicate that the strain avoidance behavior is strengthened in CPCs by a higher cell density (Figure S2). Unstrained cells displayed random orientation at all cell densities, as shown by the isotropic distribution (Figure 1E; Figure S1). 69
 display increasingly evident strain avoidance behavior (A-C), as shown by the orientation distribution of strained cells (D).")
79 Figure 1: Increased cell density strengthens CPC strain avoidance response. CPCs seeded at increasing cell densities (dens1, dens2, dens3) and subjected to uniaxial cyclic strain (10%, 0.5Hz) display increasingly evident strain avoidance behavior (A-C), as shown by the orientation distribution of strained cells (D). Unstrained samples exhibited isotropic orientation at all densities (E). Mean±SD (n=3 biological replicates). Scale bar 500 μm. Cell density does not affect CPC differentiation Because the onset of differentiation can lead to the development of CPC mechanosome and improved mechanosensing, we tested whether increasing the cell density would influence CPC fate decision. The expression of cardiac, vascular, and fibrotic markers in undifferentiated CPCs cultured at increasing seeding densities (dens1, dens2, dens3: Figure S1A-C) was analyzed. No differences were observed in the expression of cardiac markers of CPCs at dens2 and dens3 with respect to dens1, indicating that cardiomyogenic differentiation is not occurring up to day2. The only observed change was the decreased expression of Mef2c and myocardin at day0 (Figure S1D), though the difference was no longer significant at day2 (Figure S1E). Expression of endothelial gene CD31, smooth muscle marker αsma, and fibroblast marker CD90 remained unchanged at all cellular densities and time points (Figure S1D, E), indicating that CPCs are not differentiating toward the vascular or fibrotic lineages. 70
80 Increased cell density does not influence the mechanosome but it enhances N-cadherin assembly To determine the influence of cell density on the development of the mechanosome and cellcell adhesions, we performed qualitative immunofluorescent staining analyses for F-actin, the FA protein vinculin, and the adherens junction protein N-cadherin. The increased cell density did not seem to alter the development of the thin actin fibers exhibited by CPCs (day2, Figure 2A-C). Likewise, small vinculin clusters were observed at all cell densities, indicative of immature FAs (day2, Figure 2D-F), in contrast with the developed SFs and FAs previously observed in early differentiated CPCs (chapter 2 of this thesis) 121. Semi-quantitative analysis via Western Blotting confirmed similar levels of F-/G-actin at the different cell densities (Figure 2J). On the other hand, immunofluorescence analysis showed improved N-cadherin clustering at dens2 and dens3 with respect to dens1, with larger albeit less numerous N-cadherin clusters at dens3 (day2, Figure 2G-I). Total protein analysis revealed similar N-cadherin levels at dens1, dens2 and dens3, though with a rather high variability (Figure 2K). Further, we sought to estimate the degree of N-cadherin localization to the membrane by the amount of membranebound (MB) N-cadherin with respect to the soluble cytosolic N-cadherin content (S). Similar N- cadherin MB/S ratios were detected among the three cell densities, at both day0 and day2 (Figure 2L). Therefore, the increased cell density had no effect on the development of SFs and FAs. N-cadherin production and localization to the cell membrane were stable, whereas N- cadherin clustering was enhanced. 71
 and vinculin (D-F) show similar SF and vinculin development after 72 hrs of culture (day2) at dens1, dens2 and dens3.")
81 Figure 2: Cell density does not influence SF and FA development and N-cadherin production, but it enhances N-cadherin clustering. Immunofluorescent confocal images of F-actin (A-C) and vinculin (D-F) show similar SF and vinculin development after 72 hrs of culture (day2) at dens1, dens2 and dens3. Western blotting reveals comparable levels of F-/G-actin with increasing cell density both after 24 hrs (day0) and 72 hrs (day2) of culture (J). Confocal imaging of N-cadherin shows improved clustering associated with increased cell density (G-I), although Western blot analysis indicates similar MB/S N- cadherin levels at all cell densities (L). Mean±SD (n=2-3). Scale bar 50 μm. N-cadherin and ROCK both contribute to CPC strain avoidance N-cadherin mediates mechanical communication between cardiomyocytes. Therefore, we sought to verify whether N-cadherin played a role in the augmented strain avoidance of CPCs induced by increased cell density. N-cadherin adhesion was inhibited by addition of ADH-1 in CPCs cultured at the highest cell density (dens3: 5x10 4 cells/cm 2 ) and subjected to cyclic strain. CPCs treated with ADH-1 showed reduced, although not completely blocked, strain avoidance response at day2 (Figure 3B, D), as compared to untreated control samples (Figure 3A, D; Figure S3). Moreover, the strain avoidance of predifferentiated CPCs is dependent on SF development and contractility (chapter 3 of this thesis). Thus, we next analyzed the mechanoresponse of highly dense CPCs in the presence of the Rho-ROCK inhibitor Y27632, which disrupts SFs and reduces cell contractility. Treatment with Y27632 resulted in decreased, though not completely 72
.")
82 abolished, strain avoidance response of highly dense CPCs at day2 (Figure 3C, D; Figure S3). Unstrained cells showed isotropic orientation in all conditions (Figure 3E; Figure S3). To better understand the timing behind the impaired strain avoidance following N-cadherin inhibition, we compared the orientation of strained CPCs at day1, i.e. before adding the ADH-1 inhibitor, and 24 hrs after the inhibition (day2). First, we observed that highly dense CPCs responded to cyclic strain already after 24 hrs of cyclic stretching (day1; Figure 4A, F, G), though this response was enhanced at day2 (Figure 4B, F, G). However, addition of ADH1 at day1 prevented this increase in the fraction of reoriented cells at day2 (Figure 4A, C), resulting in overlapping orientation distributions at day1 and day2, i.e. before and after ADH-1 addition (Figure 4F, G). The same behavior was observed in two separate experiments (Figure 4F, G). Similarly, addition of ADH-1 to Y27632-treated CPCs (Y27+ADH-1, Figure 4E) at day1, prevented further reorientation of the cells at day2, and resulted in a proportion population of perpendicularly reoriented cells comparable to untreated CPCs at dens1 (Figure 1D). Consequently, the combined inhibition of N-cadherin and ROCK seemed to be more effective in hindering the strain avoidance response than the single inhibitors (Figure 4H), which could suggest a synergistic contribution of N-cadherin and ROCK activity in CPC strain avoidance. Figure 3: The strain avoidance of highly dense CPCs is reduced by inhibitors of N-cadherin and of ROCK. The strain avoidance displayed by CPCs at dens3 (A) is hindered by inhibitor of N-cadherin (ADH-1; B) and of ROCK (Y27632; C), as shown by the cell orientation distribution (D). Unstrained samples exhibit random orientation, as shown by the isotropic distribution (E). Mean±SD (n=3-4). Scale bar 500 μm. 73
83 74
84 Figure 4: N-cadherin inhibition at day1 prevents further strain avoidance of highly dense CPCs at day2. CPCs seeded with high cell density (dens3) show strain avoidance response after 24 hrs (day1; A, F, G), which becomes more evident after 48 hrs (day2; B, F, G) of uniaxial cyclic strain. However, when N- cadherin adhesions are inhibited at day1 (ADH-1), the same fraction of reoriented cells is observed at day1 and day2 (C, F, G). Also in the presence of Y27632, the amount of reoriented cells does not increase at day2 after ADH-1 addition at day1 (Y27+ADH-1; D, E H). Combined inhibition of ROCK and N- cadherin (Y27+ADH-1) reduces even further the strain avoidance response (H). Mean±SD (n=3-10 images per group from 1 technical replicate). Scale bar 500 μm. N-cadherin and ROCK inhibition differentially influence stress fiber development To evaluate the influence of N-cadherin and ROCK inhibition on the CPC mechanosome and cell-cell adhesions, we looked at the development of SFs, vinculin, and N-cadherin in highly dense CPCs (dens3) treated with ADH-1 or Y27632, as compared to untreated control samples (Figure 5A-C). Confocal imaging showed that the N-cadherin inhibitor ADH-1 induced SF thickening (Figure 5D), whereas it did not affect the expression and distribution of vinculin (Figure 5E). Moreover, ADH-1 did not influence the clustering of N-cadherin (Figure 5F). By contrast, the ROCK inhibitor Y27632 induced SF disruption and smaller vinculin clusters (Figure 5G, H), in agreement with previous observations 229,234. Additionally, N-cadherin clustering was decreased by the addition of Y27632 (Figure 5I), which might be explained by the dependence of cadherin ligation and clustering on cytoskeleton integrity and contractility 252,
 at day2 show thin actin SFs (A), and presence of vinculin (B) and N-cadherin clusters (C).")
85 Figure 5: Effect of N-cadherin and ROCK inhibition on the mechanosome and cell-cell adhesion development. Highly dense CPCs (dens3) at day2 show thin actin SFs (A), and presence of vinculin (B) and N-cadherin clusters (C). Inhibition of N-cadherin adhesions by ADH-1 leads to SF thickening (D), but does not affect the clustering of vinculin (E) or N-cadherin (F). Addition of the ROCK inhibitor Y27632 results in thinner SFs (G), and leads to reduced clustering of vinculin (H) and of N-cadherin (I). Scale bar 25 μm. 76
86 Discussion In this study we investigated the importance of cell density for the reorientation response of human undifferentiated CPCs to cyclic stretching. While considerable effort has been made to understand the role of cell-cell adhesions on collective cell behavior, such as collective migration, to the best of our knowledge this is the first study to investigate the effects of density and cadherin-mediated adhesions on cellular strain avoidance. The presented findings indicate that N-cadherin mediated adhesions, as well as ROCK activity, contribute to the reorientation response of CPCs to cyclic strain. Additionally, the combined inhibition of N- cadherin adhesions and ROCK seems to result in a stronger reduction of the strain avoidance as compared to the single inhibitors (Figure 6), leading to a degree of cellular reorientation comparable to the lowest cell density. This suggests that N-cadherin and ROCK play a synergistic role in the strain avoidance behavior of CPCs, and that blocking both of them can annihilate the effects of increased cell density on cellular reorientation. The cooperation between cell-cell adhesions and cell contractility is also confirmed by the fact that N-cadherin inhibition reduces the strain avoidance regardless of the SF thickening induced by ADH-1 (Figure 5). Our results are in agreement with the established link between cadherins and actin 242, , that provides mechanical communication between neighboring cells. Moreover, both cadherins and ROCK have been shown to play a role in cell mechanosensing and mechanotransduction 234,244. In particular, the cooperation between tensional and cohesive forces among neighboring cells, exerted via integrin- and cadherin- mediated adhesions, was demonstrated to be crucial for collective cell migration of epithelial and cancer cells 253, , suggesting that a similar mechanisms might occur for strain avoidance. Interestingly, addition of ADH-1 at day1 seemed to prevent further cell reorientation at day2 (Figure 4), which contributes to suggest that N-cadherin mediated communication between adjacent cells is required for CPC strain avoidance. Nevertheless, the observed effect of ADH-1 on the reorientation ability of CPCs could also be due to a reduced cell proliferation induced by the inhibitor (Figure 5), and the consequent loss of physical contact among neighboring cells. On the other hand, the improved strain avoidance at higher cell densities could also be provided by geometrical constraints imposed by adjacent cells, that is known to influence cellular motility 261,262. Besides, cell-cell adhesion via cadherins was shown to induce cell polarity 263, although this phenomenon is independent of ROCK activity 263, as opposed to the high density-induced strain avoidance of CPCs. Therefore, cell-cell contact induced polarity might not be the main/only mechanism underlying our observations. Cell density has been shown to play a role in stem cell differentiation and pluripotent stem cell generation 267. Thus, we analyzed the influence of increasing cell density on CPC lineage commitment, and verified that at the investigated timescales the enhanced cell density does not alter the undifferentiated state of the CPCs (Figure S1). This phenomenon has also been observed in mesenchymal stem cells, in which cell-cell contact has been shown to be irrelevant for the activation of transcription factors during osteogenic or adipogenic differentiation 268,269. Moreover, the similar development of SFs and FAs (whose maturation occurs upon cardiac differentiation of CPCs 121 at the different cell densities confirms that the 77
87 enhanced strain avoidance at higher densities is not due to enhanced mechanosome maturation and cell differentiation. Cell-cell adhesions play a key role in heart tissue development, where their maturation is crucial for the mechanical communication among cardiomyocytes and thereby for cardiac function 240. The presented results indicate that the response of undifferentiated CPCs to cyclic stretching is promoted by increased cell density and cell-cell adhesions mediated by N-cadherin. Moreover, cadherin-mediated adhesions and ROCK activity (and hence SF development and cell contractility) seem to be both required for CPC reorientation response to cyclic stretching. Our findings suggest that increased density of CPCs injected in the injured myocardium may help not only mechanical coupling of adjacent CPCs via cell-cell adhesions, but also mechanical integration into the host tissue, and can therefore potentially improve the outcome of CPCbased cardiac regenerative therapies. Acknowledgements This research was funded by the People Program (Marie Curie Actions) of the European Union s Seventh Framework Program FP7-People-2012-ITN TECAS, under grant agreement n Part of this study is supported by the Dutch Heart Foundation (project DHF-2014T013). Dr. Ir. Mark van Turnhout and Ir. Eline van Haaften are kindly thanked for their help with the orientation distribution analysis. 78
88 Appendix to chapter 4 Table S1: List of used primer sequences and annealing temperatures. Gene Primer sequence Ta Nkx2.5 Fw: CCCCTGGATTTTGCATTCAC Rv: CGTGCGCAAGAACAAACG 60 GATA4 Fw: TCCAGCAACTCCAGCAAC Rv: AGACATCGCACTGACTGAG 60 Mef2c Fw: TCGGGTCTTCCTTCATCAG Rv: GTTCATCCATAATCCTCGTAATC 60 Myocardin Fw: AGATGGATGAACTCCTGGAC Rv: AGTTGGACTTCGGGAAGATC 60 TroponinT (TnT) Fw: CGACGAGAGGAGGAGGAGAAC Rv: CCGCTCTGTCTTCTGGATGTAAC 60 Connexin43 (Cx43) Fw: TTTCTTCAAGGGCGTTAAGGATC Rv: AGGAGGAGACATAGGCGAGAG 60 CD31 (PECAM) Fw: AAGGAACAGGAGGGAGAGTATTA Rv: GTATTTTGCTTCTGGGGACACT 60 αsma Fw: CGTGTTGCCCCTGAAGAGCAT Rv: ACCGCCTGGATAGCCACATACA 60 CD90 (Thy-1) Fw: CTACTTATCCGCCTTCACTAGC Rv: AGCCACGAGGTGTTCTGAG 60 CDH2 (N-cadherin) Fw: AGGGGACCTTTTCCTCAAGA Rv: CAATGTCAATGGGGTTCTCC 60 Notch1 Fw: CCTGAAGAACGGGGCTAACA Rv: GATGTCCCGGTTGGCAAAGT 60 Notch2 Fw: GGCTATGAATGCCAGTGTGC Rv: GGATCGGGGTCACAGTTGTC 60 Notch3 Fw: GCCATGCTGATGTCAATGCT Rv: CAGCCCAGTGTAAGGCTGAT 60 Jagged1 Fw: AATGGCTACCGGTGTGTCTG Rv: CCCATGGTGATGCAAGGTCT 60 Hey1 Fw: CGAAATCCCAAACTCCGATA Rv: TGGATCACCTGAAAATGCTG 60 Hes1 Fw: AGTGAAGCACCTCCGGAAC Rv: CGTTCATGCACTCGCTGA 60 79
, dens2 (B), and dens3 (C) does not significantly influence cell fate decisions at 24 hrs (day0, D) or 72 hrs (day2, E) after seeding.")
89 Figure S1: Cell density does not affect CPC lineage commitment. CPC culture at increasing cell densities dens1 (A), dens2 (B), and dens3 (C) does not significantly influence cell fate decisions at 24 hrs (day0, D) or 72 hrs (day2, E) after seeding. Expression of cardiac-specific genes (Nkx2.5, Mef2c, myocardin, Cx43, TnT, N-cadherin (CDH2)), endothelial (CD31), smooth muscle (αsma), and fibroblast (CD90) markers is unchanged in dens2 and dens3 as compared to dens1, except for a decrease of Mef2c and myocardin at dens3 at day0 (D) that is no longer significant at day2 (E). Mean±SD (n=3), *p<0.05. Scale bar 500 μm. 80
, and their anisotropy (B, C).")
90 Figure S2: Increased cell density enhances the strain avoidance of undifferentiated CPCs. Cell orientation distributions were described by the location and width of their peak (A), and their anisotropy (B, C). Cyclically strained cells at increasing densities show the same peak location around 90, though the peak width is decreasing with higher density (ns, A). The distribution anisotropy increases with the cell density, particularly evident between dens1 and dens3 (B). The orientation distribution of strained CPCs are significantly more anisotropic than the distributions of unstrained cells (C), indicating an evident strain avoidance response. Mean±SD (n=3). *p<0.05; **p<
 at day2, with wider peak in the presence of ROCK inhibitor Y27632 and N-cadherin inhibitor ADH-1 (A).")
91 Figure S3: Inhibition of ROCK and N-cadherin hinders the strain avoidance of highly dense undifferentiated CPCs. Cyclically strained CPCs show reorientation around 90 (A) at day2, with wider peak in the presence of ROCK inhibitor Y27632 and N-cadherin inhibitor ADH-1 (A). Addition of either Y27632 or ADH-1 lead to smaller distribution anisotropy, which confirms the reduction in strain avoidance response of highly dense undifferentiated CPCs by the inhibitors (B). However, the strain avoidance is not completely abolished by inhibition of ROCK or N-cadherin, as showed by the significantly larger anisotropy of the orientation distribution of strained and unstrained CPCs, both in the absence (CTRL) and in the presence of the inhibitors (C). Mean±SD (n=3).*p<0.05; ***p<0.001; ****p<
92 CHAPTER 5 2D and 3D environments differentially direct cardiac progenitor cell fate and mechanoresponse This chapter is based on: Mauretti A., Bax N.A.M., Spaans S., Goumans M.J., Sahlgren C., Bouten C.V.C, Cardiac progenitor cell fate and mechanoresponse are differentially directed by 2D and 3D environments, submitted for publication.
93 Abstract Cardiac progenitor cells (CPCs) are multipotent cells that reside in the human heart, and represent a promising cell source for cardiac repair and regeneration. Mechanical cues from the cardiac microenvironment, such as extracellular matrix (ECM) stiffness and cyclic mechanical loading, can direct the behavior and function of cells inhabiting the heart, and thus may influence the regenerative potential of CPCs. However, the response of CPCs to mechanical stimuli in three-dimensional (3D) environments, more resembling of the in vivo physiological situation than two-dimensional (2D) substrates, is largely unknown. Here, we investigate the influence on human CPCs to culture in static and cyclically stretched 3D fibrin hydrogels. The soft 3D environment (E=0.95±0.30 kpa), promoted CPC differentiation toward the vascular rather than the cardiomyogenic fate, as compared to stiff 2D substrates (E=1.02±0.01 MPa). Moreover, CPCs cyclically strained in 3D fibrin hydrogels reoriented parallel to the strain direction, as opposed to the perpendicular reorientation shown on 2D substrates. The distinct cellular mechanoresponse to the 2D and 3D environments was corroborated by the differential activation of the YAP/TAZ pathway, one of the main pathways of cell mechanosensing and regulation of cell fate determination. Our findings provide novel insights into the role of environmental stiffness and dimensionality on the fate determination, mechanosensing and mechanoresponse of human CPCs. Moreover, the presented results highlight the relevance of engineering in vitro model systems that mimic the 3D physiological microenvironment for more accurate study of in vivo cell behavior. 84
94 Introduction Cardiac progenitor cells (CPCs) represent a valid cell source for cardiac regeneration, and have shown encouraging results in pre-clinical and clinical studies 17,239. Nevertheless, low cell engraftment, survival and differentiation limit the long-term benefits of CPC-based therapies. In order to improve current CPC-mediated cardiac regeneration therapies, there is the need to explore and understand how these cells interact with the mechanical cues offered by their microenvironment, such as ECM stiffness and the cyclic strain provided by the beating heart. The importance of studying cellular behavior in 3D, in vivo-like, environments is widely recognized, as different mechanisms and cellular responses can occur in 2D and 3D environments Yet, most of our understanding of cell mechanoresponse, including that of CPCs, is based on two-dimensional (2D) studies. As shown in chapter 2 of this thesis, in 2D human CPCs develop the ability to sense and respond to cyclic stretching upon early cardiomyogenic differentiation, along with the maturation of stress fibers and focal adhesions 121, or in the presence of high cell density (chapter 4). When exposed to uniaxial cyclic strain on 2D flexible substrates (Bioflex membranes), CPCs reoriented perpendicular to the stretching direction 121,127, a behavior observed in several cell types and known as strain avoidance 230,234,273. On the contrary, the cellular response of CPCs to cyclic strain in 3D environments is unknown, whereas experiments on other cell types, such as fibroblasts and smooth muscle cells, have been conducted. Contrasting results were reported, due to the diversity of the used cell types and model systems A key factor in cell mechanoresponse is represented by environmental stiffness. Tondon and Kaunas 278 showed that U2OS cells cyclically strained on 2D rubber substrates aligned perpendicular to the strain direction (strain avoidance), whereas the same cells cyclically stretched on soft collagen gels showed alignment parallel to the stretch direction (strain alignment) 278. The authors suggested that observed cell reorientation is caused by a different tensional state of the cells on a stiffer or softer surface, indicating the crucial role played by substrate stiffness in the cellular response to cyclic straining. Additionally, substrate stiffness plays a recognized role in the differentiation and function of stem and progenitor cells 211,279,280. Cells respond to substrate stiffness by developing stress fibers, actomyosin fiber bundles that represent the contractile elements of the cell 123,251. Stress fiber contractility is regulated by a number of kinases, such as myosin light chain kinase (MLCK) and Rho-associated kinase (ROCK) activated by RhoA 272,281. Another signaling route that has emerged as fundamental cellular mechanosensor is the Hippo pathway and its downstream effectors YAP/TAZ 282. In response to stiff seeding substrates, and downstream of RhoA-ROCK 283, dephosphorylation of YAP (and TAZ, which shows analogous behavior) leads to its activation and translocation to the cell nucleus. Nuclear YAP can then regulate gene expression and cell function, including proliferation, migration and differentiation 284,285. On the contrary, soft substrates determine the exclusion of YAP from the nucleus into the cytoplasm, following deactivation of the pathway 284,286. YAP activity was also shown to be regulated by substrate stiffness in CPCs 287. Interestingly, YAP activation on stiff substrates was shown to promote CPC 85
95 differentiation towards the cardiomyogenic phenotype, whereas YAP silencing induced endothelial differentiation 287. Here, we investigated the effect of the mechanical environment provided by static and cyclically stretched 3D fibrin hydrogels on human CPCs, as compared to 2D substrates, in terms of differentiation, cellular reorientation, and YAP activity. Human CPCs cultured in soft 3D fibrin hydrogels (E=0.95±0.30 kpa) showed downregulation of cardiac-specific genes and increase of endothelial and smooth muscle markers. Application of cyclic strain in 3D hydrogels did not affect CPC gene expression, and led to cellular reorientation parallel to the stretching direction (strain alignment), as opposed to the strain avoidance observed on 2D Bioflex membranes (chapter 2) 121 (E=1.02±0.01 MPa). Reduction of stress fiber contractility by inhibition of ROCK or MLCK did not modify the cellular response, suggesting that the observed CPC reorientation does not require contractile stress fibers. Lower nuclear-to-cytoplasmic YAP distribution was detected in CPCs cultured in the soft 3D hydrogels as compared to 2D Bioflex membranes, indicating decreased activity of the YAP/TAZ pathway in 3D. The reduced YAP activity confirmed the low cell mechanosensitivity in the fibrin 3D environment, and supports the shift towards the vascular lineage in 3D. The findings presented in this work provide novel insights into the interactions of CPCs with their mechanical environment, and highlight the relevance of mimicking 3D in vivo-like conditions in engineered in vitro microenvironments. Improved knowledge and understanding of the CPC response to environmental cues under physiologically relevant conditions can contribute to optimize and develop research and therapeutic strategies of CPC-based cardiac regeneration. Materials and methods Culture of cardiac progenitor cells Human fetal Sca-1 + cardiac progenitor cells (CPCs) were isolated and cultured as previously described 39,40, and L9TB CPCs immortalized cell line was produced by lentiviral transfection of htert and BMI-1. CPCs were cultured in SP++ growth medium consisting of M199 (Gibco)/EGM2 (3:1) supplemented with 10% (v/v) fetal bovine serum (FBS) (Greiner bio-one), 1% (v/v) nonessential amino acids (Gibco), and 1% (v/v) penicillin/streptomycin (Lonza) on 0.1% (w/v) gelatin (Sigma Aldrich)/PBS (Sigma) coated flasks. Preparation of 3D constructs Velcro strips (11 x 7 mm) were glued with Silastic MDX on the flexible membrane of untreated Bioflex plates (FlexCell International Corporation) in the shape of a cross, in order to obtain biaxially constrained hydrogels 160,274 (Figure 1A). The Velcro constructs were allowed to dry and sterilized with 70% ethanol for 1 hr, followed by UV illumination for 30 min. CPCs were seeded in fibrin gels at a concentration of cells/ml. Briefly, bovine fibrinogen (Sigma) was dissolved in culture medium (SP++) to a final concentration of 10 mg/ml, and sterile filtered through a 0.2 μm filter. Cells were mixed with thrombin (final concentration 5 IU/ml) and then 86
96 with the fibrinogen solution (500 μl/gel). The mix was quickly pipetted in between the Velcro cross-shaped constructs, and allowed to gelate at room temperature for 5 minutes before being incubated at 37 C. After 20 min, SP++ culture medium was added to the wells (4 ml/well). For inhibition of stress fiber contractility, ROCK inhibitor Y27632 (Sigma-Aldrich, 10 μm) and MLCK inhibitor ML7 (Sigma, 100 μm) were added to the culture medium during cell seeding (day0). Cyclic mechanical loading of 3D constructs Cyclic strain was applied with a Flexcell FX-5000 system (Flexcell International Corporation). CPC-seeded fibrin constructs (Figure 1A) were statically cultured for 2 hrs to allow for correct gelation of the fibrin before stretching. Uniaxial cyclic strain (10%, 0.5 Hz, sine wave) was applied for 48 hrs (day2) along the 0 direction (Figure 1B). Static control samples were cultured under the same conditions. Mechanical properties and cyclic strain validation The stiffness of the Bioflex membrane surface was determined with a Piuma nanoindenter (Optics11) by averaging the Young s modulus (E) of different points of the Bioflex membrane. For (cell-free) fibrin hydrogels, the shear s modulus G was measured with a Discovery HR-3 Rheometer (TA Instruments; Geometry used: 25-mm Peltier plate sand-blasted) (n=5). The Young s modulus E can then be calculated by: E = 2G(1 + ν) Where ν is the Poisson s ratio of the fibrin matrix, that was assumed equal to The efficacy of the 2D and 3D cyclic strain was validated by image analysis. First, cyclic stretching of the Velcro strips glued on the Bioflex membrane was analyzed. Videos of stretching cycles were converted into images with an in-house Matlab script, and the applied strain was measured by the displacement of the Velcro strips from one another with ImageJ. To validate the cyclic strain applied to the fibrin constructs, graphite powder was spread on top of the gels, and the displacement of the graphite pattern was analyzed via an in-house developed digital image correlation analysis (DIC) method (representative outcomes of the image analysis are shown in Figure 1C, D). Gene expression analysis Total RNA was isolated from one or two technical replicates per experimental condition (n=4 biological replicates). The constructs were rinsed 3 times with Ham-F12 medium at 37 C, cut out of the Velcro and snap-froze in liquid nitrogen. Frozen samples were then disrupted with RNAse-free pestles before using the Qiagen RNAeasy isolation kit according to manufacturer s instructions (Qiagen). cdna was synthesized with 100ng RNA per sample using M-MLV Reverse Transcriptase (Bio-Rad) and subjected to qpcr using iqtm SYBR@ Green Supermix and the CFX384 detection system (Bio-Rad). Primer sequences for qpcr and annealing temperatures are presented in Table 1. 87
97 Immunofluorescence staining and confocal imaging Samples were washed three times with Ham-F12 (Gibco) at 37 C, fixed in 3.7% (v/v) formaldehyde (Merck) for 30 min, washed three times with PBS, and permeabilized with 0.5% (v/v) Triton X-100 (Merck) in PBS for 15 min. For orientation analysis, samples were kept in between the Velcro strips, to keep track of the straining direction, and stained with phalloidin for 20 min at room temperature, for fluorescent labeling of F-actin fibers. Samples were visualized with a Leica TCS SP5X confocal microscope, by taking z-stacks of 100 μm thickness at different heights in the fibrin construct. For immunofluorescent staining of YAP and of the cell nuclei, constructs were cut out with a scalpel and placed in 48-well plates. Non-specific antibody binding was blocked with 10% (v/v in PBS) horse serum incubation for 45 min. The fibrin constructs were then incubated with primary antibodies in 1% (v/v in PBS) horse serum overnight at 4 C. The samples were rinsed with PBS four times for 5 min and incubated for 1 hr with secondary antibodies and DAPI for nuclei staining, diluted in PBS. The constructs were finally rinsed 4 times with PBS, and stored at 4 C in PBS until visualization. All used antibodies and dyes are listed in Table 2. All incubation steps, except primary antibody incubation, were performed at room temperature upon gentle shaking. Quantification of cell orientation and fluorescent signal Cell orientation was quantified from phalloidin-stained samples, by using the Directionality plug-in of Fiji ( The orientation analysis is performed based on the Fourier transform (FFT) of each image, by means of fitting a Gaussian to the FFT signal and measuring its peak position. Maximum projection images were obtained from 2 μm z-stacks of the central part of the constructs, for a total thickness of 100 μm analyzed per image. For each experimental condition, 4-9 images were analyzed from the center of 3D samples, for each independent experiment (n=3-4 biological replicates). The amount of nuclear-to-cytoplasmic YAP was determined from confocal images collected with identical acquisition parameters, by the ratio between the mean fluorescent signal in the cell nucleus and the whole cell, measured with ImageJ. The DAPI channel was used to determine the nuclear area. For each of the three biological replicates, at least 10 cells per group were analyzed (n=3). Statistical analysis All data are presented as mean ± standard deviation (SD). Statistical analysis was performed with Kruskal-Wallis test followed by a Dunn s post-hoc test for multiple comparisons or with student s t-test where appropriate. Analysis of cell orientation distributions was performed with an in-house developed Matlab script, where each distribution is described by three parameters: peak location (mu), peak width (std) and distribution anisotropy (ani) 232. Significance was assumed for p<0.05; statistical analysis was conducted with GraphPad Prism
98 Table1: List of used primer sequences and annealing temperatures. Gene Primer sequence T a Nkx2.5 Fw: CCCCTGGATTTTGCATTCAC Rv: CGTGCGCAAGAACAAACG 60 GATA4 Fw: TCCAGCAACTCCAGCAAC Rv: AGACATCGCACTGACTGAG 60 Mef2c Fw: TCGGGTCTTCCTTCATCAG Rv: GTTCATCCATAATCCTCGTAATC 60 Myocardin Fw: AGATGGATGAACTCCTGGAC Rv: AGTTGGACTTCGGGAAGATC 60 TroponinT Fw: CGACGAGAGGAGGAGGAGAAC Rv: CCGCTCTGTCTTCTGGATGTAAC 60 Connexin43 (Cx43) Fw: TTTCTTCAAGGGCGTTAAGGATC Rv: AGGAGGAGACATAGGCGAGAG 60 CD31 Fw: AAGGAACAGGAGGGAGAGTATTA Rv: GTATTTTGCTTCTGGGGACACT 60 asma Fw: CGTGTTGCCCCTGAAGAGCAT Rv: ACCGCCTGGATAGCCACATACA 60 CD90 Fw: CTACTTATCCGCCTTCACTAGC Rv: AGCCACGAGGTGTTCTGAG 60 CD105 Fw: GGCACCGTGGAACTCATC Rv: GTGTAGAAGTGGAGGAGGAAG 60 CDH2 Fw: AGGGGACCTTTTCCTCAAGA Rv: CAATGTCAATGGGGTTCTCC 60 Notch1 Fw: CCTGAAGAACGGGGCTAACA Rv: GATGTCCCGGTTGGCAAAGT 60 Notch2 Fw: GGCTATGAATGCCAGTGTGC Rv: GGATCGGGGTCACAGTTGTC 60 Notch3 Fw: GCCATGCTGATGTCAATGCT Rv: CAGCCCAGTGTAAGGCTGAT 60 Jagged1 Fw: AATGGCTACCGGTGTGTCTG Rv: CCCATGGTGATGCAAGGTCT 60 Hey1 Fw: CGAAATCCCAAACTCCGATA Rv: TGGATCACCTGAAAATGCTG 60 Hes1 Fw: AGTGAAGCACCTCCGGAAC Rv: CGTTCATGCACTCGCTGA 60 89
99 Table 2: List of used antibodies and dyes. Antigen Source Cat. no Isotype Species Label Dilution YAP Cell Signaling 4912S IgG Rabbit - 1:50 Phalloidin Sigma-Aldrich P Amanita phalloides ATTO 488 1:400 Rabbit IgG Molecular Probes A21428 IgG (H+L) Goat Alexa Fluor 555 1:200 DAPI Sigma-Aldrich D :500 Results Mechanical characterization and cyclic strain validation To determine the influence of different environments on CPCs, mechanical characterization of the 2D culture substrates (used to apply cyclic strain in chapter 2) and 3D fibrin hydrogel was conducted. Nanoindentation measurements provided a Young s modulus value of the Bioflex membrane of EBioflex=1.02±0.01 MPa. Rheology measurements revealed a shear s modulus of cell-free fibrin hydrogel (10 mg/ml) of G fibrin=0.38±0.12 kpa, corresponding to Efibrin=0.95±0.30 kpa. The Young s modulus of plastic tissue culture plates is known to be of approximately Eplastic 10 MPa 289. Therefore, fibrin is ~10 4 times softer than the tissue culture plastic, and ~10 3 times softer than the Bioflex membrane used for 2D straining experiments (Table 3). The fibrin hydrogel did not show strain stiffening behavior for strains lower than 10% (data not shown), in agreement with the literature 290. The efficacy of the 3D cyclic straining was determined by image analysis. The strain applied to the Bioflex membrane was measured to equal 9.9±2.3%. On top of the fibrin gels, cyclic strains between 6-8% were measured (as representatively shown in Figure 1B, C), indicating effective cyclic stretching of the 3D samples, though characterized by a lower amplitude than the 2D samples. 90
 [kpa] Assay Polystyrene culture plate (2D) ~1x10 4 289 Gene expression 2D Bioflex membrane (2D) ~1x10 3 Fibrin hydrogel (3D) ~1 Reorientation to cyclic strain 2D YAP")
100 Table 3: Stiffness values (Young s modulus) and performed assays for each of the substrates used in this chapter. Sample Stiffness (E) [kpa] Assay Polystyrene culture plate (2D) ~1x Gene expression 2D Bioflex membrane (2D) ~1x10 3 Fibrin hydrogel (3D) ~1 Reorientation to cyclic strain 2D YAP immunofluorescence Gene expression 3D Reorientation to cyclic strain 3D YAP immunofluorescence Figure 1: Mechanical characterization of 2D and 3D straining substrates. CPC-seeded, biaxially constrained, 3D fibrin hydrogel constructs were prepared (A), and uniaxial cyclic strain was applied along the 0 direction (B). Typical outcomes of the cyclic strain validation on 3D samples conducted by digital image correlation (DIC) analysis (C, D). 91
101 Culture in 3D soft hydrogels influences CPC fate decisions The lineage commitment of CPCs following culture in 3D fibrin hydrogels was evaluated by analyzing the expression of cardiac-specific and vascular markers. As compared to cells cultured on 2D substrates, after 48 hrs of culture in the 3D fibrin hydrogels CPCs showed decreased expression of the cardiac transcription factors Nkx2.5, Mef2c, GATA4, the cardiomyocyte gene myocardin, as compared to the cell monolayer (Figure 2A). At the same time, increased expression of the endothelial marker CD31 and of the smooth muscle marker αsma was detected, whereas the fibroblast marker CD90 was downregulated. Notch is a key signaling pathway in CPC fate determination, and plays a key role in cardiac development and repair after injury. CPC culture in the 3D fibrin matrix led to reduced expression of Notch components Notch1, Notch2, and Jagged1, while Notch3 increased (Figure 2B). Notch target gene Hes1 displayed decreased expression in 3D as compared to 2D samples (Figure 2C), similarly to the cardiac-specific Notch target Nkx2.5 (Figure 2A), whereas a modest increase in Hey1 expression (ns) was observed (Figure 2C). The gene expression of CPCs in cyclically strained 3D samples was unchanged with respect to static 3D samples, indicating that the applied cyclic strain does not influence CPC lineage commitment and Notch signaling (Figure 2). Our results indicate that culture of CPCs in 3D fibrin hydrogels determines a shift in cell lineage specification and influences the expression of components and downstream targets of Notch, a key molecular regulator of CPC function and cardiac development. 92
102 Figure 2: Culture in soft 3D fibrin hydrogels directs CPCs towards the vascular lineage specification. Culture of CPCs in 3D soft fibrin hydrogels leads to downregulation of cardiac-specific markers Nkx2.5, Mef2c, GATA4, and myocardin, as compared to 2D culture plastic substrates. 3D culture results in increased expression of the endothelial marker CD31 and of the smooth muscle marker αsma, and downregulation of the fibroblast marker CD90 (A). Notch receptors Notch1 and Notch2 and the ligand Jagged1 are downregulated in 3D samples, whereas the expression of Notch3 is increased (B). Expression of Notch target Hes1 is decreased in 3D samples, whereas a modest increase is observed in the expression of Hey1 (not significant) (C). No differences were observed between 3D strained and 3D static samples. Mean±SD; *p<0.05; **p<0.01. CPCs reorient parallel to the cyclic strain in 3D, independently of stress fiber development The mechanoresponse of CPCs subjected to uniaxial cyclic stretching was determined by their reorientation response. After 48 hrs of uniaxial cyclic straining in the 3D constructs (day2), CPCs reoriented parallel to the strain direction (strain alignment; Figure 3A, C), whereas static samples showed isotropic cellular orientation (Figure 3B, C; Figure S1). Stress fibers are actomyosin fiber bundles that allow cells to exert forces on the surrounding environment, and 93
 and of MLCK (ML7) to the culture medium of 3D")
103 actively respond to external mechanical stimuli. Therefore, we asked whether stress fiber contractility was responsible for the observed CPC mechanoresponse, and added inhibitors of Rho-ROCK (Y27632) and of MLCK (ML7) to the culture medium of 3D strained and unstrained samples. The cell orientation response was not affected by the inhibitors (Figure 3D, G, F; Figure S1), indicating that the CPC reorientation in 3D hydrogels is not caused by the presence of developed and contractile stress fibers. Addition of the inhibitors did not affect the random orientation of cells in static constructs (Figure 3E, H, I). Figure 3: Undifferentiated CPCs in 3D fibrin hydrogels reorient parallel to uniaxial cyclic strain, independently of stress fiber contractility. Undifferentiated CPCs in the center of 3D fibrin hydrogel reorient parallel to the direction of uniaxial cyclic strain (A, C). Reduction of stress fiber development and contractility by addition of ROCK inhibitor Y27632 (D) or MLCK inhibitor ML7 (G) does not change the parallel reorientation of cyclically strained CPCs (F). Static samples are isotropically oriented in all conditions (B, E, F, I). Mean±SD, n=3-4 independent experiments. Scale bar 100μm. YAP activity is lower in soft 3D hydrogels and is not affected by cyclic strain Hippo downstream effector YAP is increasingly recognized as one of the main cellular mechanosensors. The presence of YAP in the cell nucleus indicates activity of the YAP/TAZ 94
, with little signal coming from the cytoplasm as shown by the N/C YAP")
104 pathway. Therefore, we determined the ratio of nuclear/cytoplasmic (N/C) YAP in CPCs on 2D Bioflex membranes and in 3D fibrin hydrogels, as an indication of cell mechanosensing. CPCs cultured as cell monolayer on the Bioflex flexible membrane showed a predominantly nuclear distribution of YAP (Figure 4A), with little signal coming from the cytoplasm as shown by the N/C YAP ratio of 2.4±0.4 (Figure 4E), indicating activation of the YAP/TAZ pathway. Conversely, CPCs in 3D fibrin constructs exhibited presence of YAP in the cytoplasm as well as in the nucleus (Figure 4F), with a N/C ratio of 1.4±0.2 (Figure 4E). This indicates that although YAP is not completely inactivated in 3D samples, CPCs in 3D hydrogels display lower YAP activity, as compared to CPCs on 2D substrates (Figure 4E; p=0.024). YAP distribution in 3D samples was not affected by the application of cyclic strain (Figure 4J). Altogether these results indicate that CPC culture in the soft 3D fibrin environment leads to reduced activity of the YAP/TAZ pathway, with no major effect of the applied strain on cell mechanosensing. Figure 4: YAP activity is lower in CPCs cultured in soft 3D fibrin hydrogels, and is not influenced by the applied cyclic strain. Immunofluorescent confocal images show the expression of YAP (A, F), the cell nuclei (B, G), F-actin (C, H) in CPCs cultured for 2 days on 2D Bioflex flexible membranes (A-D) or in 3D fibrin hydrogels (F-I). YAP clearly translocases to the cell nuclei of cells on 2D Bioflex substrates (A), whereas it is evenly distributed in the cytoplasm and nuclei of CPCs in 3D fibrin hydrogels (F), as confirmed by the image quantification (E). The cyclic strain applied to 3D samples does not influence YAP distribution (J). Mean±SD; *p<0.05; scale bar 50μm. Discussion Extracellular mechanical signals can have a crucial influence on the regenerative potential of CPCs. Yet, the response of these cells to mechanical stimuli, such as stiffness and cyclic strain, in an in vivo-like 3D environment is largely unknown. Here, we examined the effects of static and cyclic culture of human CPCs in 3D fibrin hydrogels on cell fate decisions, mechanosensing and reorientation response, as compared to 2D substrates. 95
105 Culture in the softer 3D fibrin environment directed CPC lineage commitment toward a vascular rather than cardiomyogenic phenotype, and affected the expression of components and downstream targets of Notch, a key regulator of cell fate decisions in the developing and adult heart 92,93. The downregulation of Notch targets Nkx2.5 and Hes1, whose expression has been linked to cardiac differentiation 58,94, supports the observed decrease in cardiac markers. Moreover, the increased expression of Notch target Hey1 (p=0.07), observed in each independent experiment, supports the increase in vascular markers 291. In agreement with our results, Hey1 upregulation in CPCs cultured in 3D hydrogels with different stiffness was associated with induction of either vascular (E=1 kpa), or cardiac markers (E=2 kpa) 291. These findings confirm that environmental stiffness is a crucial factor in the fate decisions of CPCs, similarly to what was shown for other stem cell types 211. In particular, the stiffness of the fibrin constructs used in our work (~1 kpa) is much lower than the one of the native healthy cardiac tissue (~10-20 kpa) 292. Proper stiffness conditions are known to be crucial for the cardiomyogenic differentiation of cardiac progenitor cells 287,291,293 and induced pluripotent stem cells 294, as well as for cardiomyocyte maturation This highlights the relevance of mimicking physiological stiffness values when engineering in vitro model systems. The strain alignment of undifferentiated CPCs cyclically stretched in 3D hydrogels (Figure 3) is strikingly distinct from the strain avoidance previously observed on 2D substrates, that was more evident in early differentiated, more contractile cells (chapter 2) 121. Other groups have reported parallel cellular alignment in response to cyclic strain in 3D 276,298,299 ; however these studies are based on constructs with uniaxial constraints, which provide a static strain that can influence the cellular alignment 160. Perpendicular reorientation of fibroblasts in biaxially constrained, cyclically strained, collagen/matrigel constructs was reported by Foolen et al Due to the different size, ECM composition, and cell type of the used model systems, the comparison with our data is not straightforward. Particularly, fibroblasts are highly contractile cells, whereas CPCs have been shown to only develop mature stress fibers and focal adhesions with early differentiation 121. In fact, the strain alignment response of CPCs in 3D fibrin hydrogels is independent of developed stress fibers and cell contractility (Figure 3), which are instead required for the strain avoidance of fibroblasts in 3D 300. Recently, Tondon et al 278 have observed a strain avoidance behavior of cells cyclically stretched on 2D rubber substrates, and strain alignment on softer collagen gel layers. The authors proposed that cells on substrates with different mechanical properties actively reorient perpendicular or parallel to the applied strain in order to maintain an optimal intracellular tensional state 278. Similarly, we propose that CPCs in fibrin hydrogels, due to their intrinsically low contractility and to the low stiffness of the environment, align parallel to the stretching direction to increase their tensional state to an optimal level. Alternatively, the phenomenon can be explained by passive cellular organization due to the cell viscoelastic behavior. The reduced mechanosensing state of CPCs in the 3D fibrin constructs was confirmed by the YAP activity analysis (Figure 4). The reduced YAP activity of CPCs in soft 3D constructs (Figure 4) is in agreement with previous reports, showing YAP activation by increased substrate stiffness 284,286 downstream of the Rho-ROCK pathway 283. Further, the lower YAP activity supports our 96
106 hypothesis that the strain alignment in 3D, independent of stress fiber contractility, is caused by a low tensional state of these cells. Moreover, the decreased YAP activity in 3D can be responsible for the induction of vascular markers in the fibrin hydrogels. YAP has been shown to regulate cell fate decisions of CPCs in response to the substrate stiffness, and YAP knock-down in CPCs resulted in increase of endothelial genes and downregulation of cardiac markers 287. Additionally, the similar YAP activity in 3D static and cyclically strained samples (Figure 4) can be linked to the unchanged gene expression in static and cyclic 3D samples (Figure 2). Recent studies have shown a cell type-dependent link between the YAP and Notch pathways Our results suggest that both YAP and Notch activation may be relevant for CPC cardiomyogenic differentiation, in agreement with previous reports 58,94,287,291. More in-depth studies are needed to confirm our suggestion, particularly in view of the potential role of Notch in cardiac regeneration 58,91,156. The presented results suggest that culture of human CPCs in a soft 3D environment leads to a lower tensional state of the cells, associated to low Rho-ROCK and downstream YAP activity, which in turn directs cell fate decisions. CPC strain alignment in 3D hydrogels seems to be driven by the low intracellular tensional state, though the exact mechanism remains unclear. Nevertheless, our study lacks to decouple the effects of stiffness and dimensionality, which hinders the identification of a key factor directing the observed effects. To answer this intriguing question, future research with different experimental setups needs to be conducted. For instance, culturing CPCs in static and cyclic conditions on 2D and/or 3D substrates of varying stiffness would allow to isolate the effect of environmental stiffness and dimensionality on CPC mechanosensing and mechanoresponse. Conclusions The relevance of investigating cellular behavior in a 3D, in vivo-like, environment is widely acknowledged 270,304, as well as the crucial role of environmental stiffness in driving fundamental cellular processes such as survival and differentiation 211,279,280. Here, we showed that CPCs cultured in soft 3D fibrin hydrogels display a distinct change in fate determination, mechanosensing and mechanoresponse as compared to cells on 2D substrates. The reduced YAP activity in 3D hydrogels indicates that CPCs are characterized by a low mechanosensing and tensional state in this environment. Consequently, cells reorient parallel to the applied cyclic stretch, as opposed to the perpendicular reorientation observed on 2D substrates, where their tensional state is increased. Moreover, the reduced mechanosensing in 3D directs CPC fate toward the vascular rather than cardiomyogenic lineage. Our findings provide novel insight into the interactions of human CPCs with their mechanical environment, and highlight the importance of studying CPC behavior in a 3D, more physiologic, environment. These results provide relevant information that can be valuable for the development of an optimal engineered microenvironment. Acknowledgements 97
107 This research was funded by the People Program (Marie Curie Actions) of the European Union s Seventh Framework Program FP7-People-2012-ITN TECAS, under grant agreement n Part of this study was supported by the Dutch Heart Foundation for project DHF-2014T
, and their anisotropy (B, C).")
, as compared to untreated samples (CTRL).")
108 Appendix to chapter 5 Figure S1: CPC strain alignment response in 3D is independent of ROCK and MLCK. Cell orientation distributions were described by the location and width of their peak (A), and their anisotropy (B, C). Cyclically strained CPCs in 3D fibrin hydrogels in the presence of inhibitors of ROCK (Y27) or MLCK (ML7) show comparable peak location and width (A), as well as distribution anisotropy (B), as compared to untreated samples (CTRL). C) The distribution anisotropy of strained samples (S) is higher as compared to unstrained samples (NS), indicating a distinct strain alignment response in strained cells. Mean±SD (n=3). **p<
109 100
110 CHAPTER 6 Spheroid 3D culture enhances Notch signaling in cardiac progenitor cells This chapter is based on: Mauretti A.*, Rossi F.*, Bax N.A.M., Miano C., Miraldi F., Goumans M.J., Messina E., Giacomello A., Bouten C.V.C., Sahlgren C., Spheroid 3D culture enhances Notch signaling in cardiac progenitor cells, MRS Communications (2017). *authors contributed equally
111 Abstract Cardiac progenitor cells (CPCs) are promising candidates for cardiac regeneration, and the interaction between CPCs and their microenvironment can influence their regenerative response. Notch signaling plays a key role in cell fate decisions in the developing and adult heart. Here, we investigated the effect of 3D spheroid culture, as a model of the 3D microenvironment, on Notch in fetal and adult human CPCs, under room air (20%) and physiological (5%) oxygen tension. Notch signaling is enhanced in 3D spheroids; spheroid culture under 5% O2 further increases Notch signaling enhancement, and might ultimately improve the regenerative potential of CPCs. 102
112 Introduction Cardiac disease is one of the main causes of death worldwide. After injury, the heart muscle displays very limited regeneration, in which resident cardiac progenitor cells (CPCs) seem to play a role 148. Despite their acknowledged regenerative potential 17,180,305, very little is known about the actual role of CPCs, particularly of human origin, in cardiac repair and regeneration. Among the different CPC populations that have been identified, human cardiomyocyte progenitor cells (CPCs) represent a promising candidate for cardiac regeneration, due to their presence in fetal and adult heart, and their capacity to differentiate into the three cardiac lineages (cardiomyocytes, endothelial and smooth muscle cells) 40,305. Another interesting system to isolate and study human CPCs is represented by the cardiosphere model. Cardiospheres (CSps) are cardiac-derived multicellular spheroids, spontaneously formed by cells migrating from cardiac tissue explants and subsequently cultured on a poly-d-lysine-coated dish 163,306. CSps can serve as an interesting in vitro model of the cardiac stem cell niche 307. Compared to traditional 2D culture, multicellular spheroids provide a microenvironment that more closely mimics the in vivo physiology. These model systems recapitulate key features of the endogenous cell niche, such as improved cell-cell and cell-ecm contacts, as well as gradients of oxygen and nutrients between periphery and core of the spheroid 105,155,157,307. Improved knowledge on the mechanisms that regulate CPC behavior and functions, such as survival, proliferation, and differentiation, is needed. In particular, the interaction of CPCs with the niche in which they reside is of pivotal importance to ultimately direct their behavior towards cardiac regeneration. Notch is a cell-cell signaling pathway that regulates many cellular functions, such as survival, proliferation, and differentiation 94,96,308. In the heart, Notch is essential during development 93,309, and it is inactivated in cardiomyocytes after birth 96. However its reactivation after myocardial damage indicates that Notch plays a key role in cardiac repair 94,101. In CPCs, activation of the Notch pathway is known to induce cell proliferation 94, and is needed for the onset of cardiomyogenic differentiation, at the same time maintaining the cells in a proliferative state 94. Here we use cardiospheres, of both adult (CSps) and fetal origin (CPC-CSps), as a model to investigate the Notch pathway in CPCs under room air (20% O2) and tissue physiological (5% O2) oxygen tension (often referred to as hypoxia ). Our results show that a 3D in vivo-like microenvironment, as provided by the cardiosphere system, enhances Notch signaling in fetal and adult CPCs as compared to the respective cell monolayers. Moreover, the signaling enhancement provided by the 3D culture is further increased under hypoxic conditions. These findings provide fundamental knowledge on the interactions between CPCs and the conditions of the cardiac microenvironment, such as 3D culture and different oxygen tension conditions. 103
113 Materials and Methods CPCs were isolated from fetal human hearts by magnetic sorting using Sca-1-coupled beads. L9TB cell line was immortalized by lentiviral transduction of htert and BMI-1, and cultured in SP++ growth medium, according to published protocols 40. CPC-CSps were obtained by seeding L9TB CPCs on poly-d-lysine-coated 12-multiwell plates with a density of 1.6x10 4 cells/cm 2 and cultured in cardiosphere medium (CSM). Adult explant-derived cells (EDCs) were isolated from human surgical auricular biopsies, as previously described 163,306. Once 80% of confluency was reached, EDCs were collected and seeded on poly-d-lysine-coated 12-multiwell plates with a cell density of cells/cm 2 to form cardiospheres (CSps). CPC-CSps and CSps spontaneously formed after 5 days of culture in either room air (20% O2) or physiologic (5% O2) oxygen conditions, and collected at this time-point. CSps were seeded on a fibronectin-coated flask to obtain cardiosphere derived cells (CDCs). CPCs, CPC-CSps, EDCs, CSps and CDCs were grown in either room air (20% O2) or in tissue physiologic (5% O2) oxygen conditions (Table 1; for details on cell isolation methods and culture medium composition, see Appendix). Total RNA was isolated from one sample per group per experiment (n=4 for fetal CPCs; n=3-7 for adult CPCs) using the Qiagen RNAeasy isolation kit according to manufacturer s instructions (Qiagen). cdna was synthesized with 250 ng RNA per sample using M-MLV Reverse Transcriptase (Bio-Rad) and subjected to qpcr using iqtm SYBR@ Green Supermix and the CFX384 detection system (Bio-Rad). Primer sequences for qpcr and annealing temperatures are presented in Table 2. For immunofluorescence labelling, all samples were washed with PBS at 37 C, fixed in 3.7% (v/v) formaldehyde for 15 min, and rinsed four times with washing buffer (see Appendix). A blocking step in 1% horse serum was followed by staining with primary and secondary antibodies, and DAPI for nuclear staining. Used antibodies and dyes are listed in Table 3. Immunofluorescence images were acquired with a Leica TCS SP5X confocal microscope. All data are presented as mean ± standard deviation (SD). For qpcr data analysis, cyclethreshold (Ct) values were normalized to the reference gene (GAPDH) value, and signals of CPC- CSps, CSps, and CDCs were compared with those of the cell monolayer (CPCs and EDCs for fetal and adult cells, respectively) and presented as fold change (2 ΔΔCt ). A list of Ct values of fetal and adult cells is presented in Table S1 and Table S2, respectively. Statistical analysis was performed with Graphpad Prism Student s t-test or Kruskal-Wallis test with Dunn s posthoc for multiple comparisons were performed where appropriate. Significance was assumed for p<
114 Table 1: Schematic summary of all cell type and culture conditions used in this article. Abbreviation Origin Culture condition CPCs Monolayer (2D) 5% / 20% O2 Fetal CPC-CSps Spheroid (3D) 5% / 20% O2 EDCs Monolayer (2D) 5% / 20% O2 CSps Adult Spheroid (3D) 5% / 20% O2 CDCs Monolayer (2D) 5% / 20% O2 Table 2: List of sequences and annealing temperatures (Ta) of used primers (Fw: forward; Rv: reverse). Gene Primer sequence T a Notch 1 (N1) Notch 2 (N2) Notch 3 (N3) Jagged 1 (Jag1) Hey1 Fw: CCTGAAGAACGGGGCTAACA Rv: GATGTCCCGGTTGGCAAAGT Fw: GGCTATGAATGCCAGTGTGC Rv: GGATCGGGGTCACAGTTGTC Fw: GCCATGCTGATGTCAATGCT Rv: CAGCCCAGTGTAAGGCTGAT Fw: AATGGCTACCGGTGTGTCTG Rv: CCCATGGTGATGCAAGGTCT Fw: CGAAATCCCAAACTCCGATA Rv: TGGATCACCTGAAAATGCTG Table 3: List of used antibodies and dyes. Antigen Source Cat. no Isotype Species Label Dilution Notch1 (N1) Santa Cruz sc-6014 IgG Goat - 1:50 Notch 2 (N2) Santa Cruz sc-5545 IgG Rabbit - 1:50 Notch3 (N3) Santa Cruz sc-7424 IgG Goat - 1:50 Rabbit IgG Goat IgG DAPI Molecular Probes Molecular Probes Sigma- Aldrich A21428 IgG (H+L) Goat Alexa Fluor 555 1:200 A21432 IgG (H+L) Donkey Alexa Fluor 555 1:200 D :
, Notch2 (N2), and Notch3 (N3), Notch ligand Jagged1 (Jag1), and target gene Hey1, as compared to the respective cell monolayer from which the cardiospheres derive (CPCs and EDCs")
115 Results Notch signaling is enhanced in CPC spheroids The effect of 3D culture on Notch signaling in fetal (CPC-CSps) and adult (CSps) CPCs was evaluated by analyzing the gene expression of Notch receptors Notch1 (N1), Notch2 (N2), and Notch3 (N3), Notch ligand Jagged1 (Jag1), and target gene Hey1, as compared to the respective cell monolayer from which the cardiospheres derive (CPCs and EDCs for fetal and adult, respectively). Under 20% O2, CPC-CSps and CPC monolayer showed a similar expression of N1. However, CPC-CSps displayed increased expression of N2 and N3 (2.0- and 2.5-fold), and Jag1 (1.9-fold) (Figure 1A), and a 13.8-fold upregulation of the target gene Hey1 (Figure 1B). Adult CSps displayed upregulation of N1 and N3 (2.2- and 2.9-fold, respectively) as compared to both adult cell monolayers (explant-derived cells (EDCs) and cardiosphere-derived cells (CDCs)), whereas N2 and Jag1 showed similar gene expression levels in all samples, although a modest downregulation was shown in CDCs as compared to CSps (Figure 1C). Significant upregulation of Hey1 was observed in CSps as compared to both EDCs and CDCs (6.6-fold change; Figure 1D). Immunofluorescent staining revealed the presence of the N2 receptor in the cell nuclei in CPC- CSps as well as in the CPC monolayers, supporting the activation of Notch signaling suggested by the gene expression data (Figure 1E, F)). Similar protein localization was observed in adult CSps and cell monolayers (EDCs, CDCs; Figure S2). Staining for N1 and N3 showed localization at the cell membrane and around the cell nuclei, respectively (Figure S1). These results indicate that spheroid 3D culture favors the activation of Notch signaling in CPCs, more evidently in adult CSps than in fetal CPC-CSps. 106
116 Figure 1: Notch signaling is enhanced by 3D spheroid culture in human CPCs. (a-d) Gene expression of Notch components (N1, N2, N3, Jag1) and target gene (Hey1) in (a, b) fetal CPC-CSps (n=4) and (c, d) adult CSps (n=3-7) cultured under 20% oxygen. (e, f) Immunofluorescent staining for Notch2 (N2) in CPCs (e) and CPC-CSps (f) cultured at 20% oxygen. Nuclei are stained with DAPI. Scale bar 50μm. p <0.05, ** p <0.01; *** p < Notch signaling enhancement in CPC spheroids is increased by physiologic oxygen culture Oxygen tension is known to regulate many cell functions via the hypoxia and the Notch pathways, and is a key player in the stem cell niche 138,310,311. Therefore, we examined Notch signaling in fetal and adult CSps under physiological oxygen conditions (5% O2). CPC-CSps exhibited an increase in the RNA expression of all Notch components (Figure 2A), and a remarkable upregulation of the target gene Hey1 was observed (46.8-fold; Figure 2B). Similarly, adult CSps showed upregulation of N1 and N3 markers (2.3- and 2.2-fold, respectively) as compared to the monolayers (EDCs and CDCs), despite a slight downregulation of Jag1 (0.5-fold). No differences in the expression of N2 were observed between the three cell types (Figure 2C). Activation of the Notch downstream targets in adult CSps was demonstrated by Hey1 upregulation (22.7-fold; Figure 2D). Immunofluorescence staining showed the presence of N2 in the cell nuclei in CPC-CSps and in the respective cell monolayer (Figure 2E, F). To determine the effect of hypoxia on Notch signaling in each cell type (adult and fetal CPCs) and culture condition (monolayer, CSps), we evaluated the expression of Notch markers at 5% O2 as compared to 20% O2. In fetal CPCs and CPC-CSps, a general downregulation of all markers was observed (Figure S3A, B), except for a 2.1-fold upregulation of Hey1 in CPC-CSps at 5% O2. In adult cells (CDCs, CSps), no changes were observed at 5% oxygen, except for a modest increase in the expression of N1 in CDCs (1.5-fold) and of Jag1 in CSps (1.5-fold) (Figure S3C, D)). The presented results indicate that the enhanced expression of Notch components and target genes in fetal and adult 3D CSps is further improved by culture under physiological oxygen tension. 107
 Gene expression of Notch components (N1, N2, N3, Jag1) and target gene (Hey1) in (a, b) fetal CPC-CSps (n=4) and (c, d) adult CSps (n=3-7) cultured under 5% oxygen.")
117 Figure 2: Notch signaling enhancement by 3D spheroid culture is increased by combination with physiological oxygen conditions in human CPCs. (a-d) Gene expression of Notch components (N1, N2, N3, Jag1) and target gene (Hey1) in (a, b) fetal CPC-CSps (n=4) and (c, d) adult CSps (n=3-7) cultured under 5% oxygen. (e, f) Immunofluorescent staining for Notch2 (N2) in CPCs (e) and CPC-CSps (f) cultured at 5% oxygen. Nuclei are stained with DAPI. Scale bar 50μm. * p < 0.05, ** p < 0.01; *** p<
118 Discussion and outlook Multicellular spheroids, such as CSps, represent an in vitro model of the cardiac niche 153,307. The 3D environment and enhanced cell-cell and cell-ecm contacts offered by CSps can stimulate several molecular signaling and cellular functions. Notch is a highly conserved pathway that plays a key role in determining cell fate decisions in the heart. In CPCs, activation of Notch is needed to initiate cardiomyogenic differentiation 94 ; furthermore, transplantation of CPCs with increased Notch activity results in decreased infarct area and improved functions in an infarcted heart model 58. Previous studies have shown upregulation of Notch receptors 1 and 3 in CSps, as compared to the respective cell monolayer 312. Here, the upregulation of Notch receptors, and more importantly of the target gene Hey1 (13.8-fold and 6.6-fold in fetal and adult cells, respectively), suggests activation of Notch pathway in fetal and adult CPCs cultured as CSps. Moreover, in the presence of a physiological oxygen condition (5%), the 3D culture seemed to lead to an even stronger activation of Notch, as indicated by the enhanced upregulation of Hey1 (46.8-fold and 22.7-fold in fetal and adult CPCs, respectively). In CPC monolayers grown under 5% O2 the expression of Notch components and target genes is either decreased (fetal CPCs) or unchanged (adult CPCs) as compared to the 20% O2 culture (Figure S3). However, when CPCs are grown as CSps under 5% O2, the enhancement of Notch signaling due to CSp formation is even stronger than under 20% O2, suggesting that the 3D culture is needed for the increased activation of Notch signaling under physiological oxygen tension. In particular, the reported data suggest that, in both fetal and adult CPCs, N2 is the component that translocates to the cell nuclei in response to spheroid formation as a result of Notch signaling activation. However, unlike fetal CPCs where all Notch markers are upregulated by the 3D culture, in adult CSps the gene expression of N2 seems unvaried as compared to the cell monolayer, indicating that in adult CPCs the activation of Notch is not due to increased RNA levels. The observation might be explained by the different developmental stage of the two cell types, and a distinct response to environmental factors on Notch signaling in fetal and adult cells. Altogether, our findings suggest that in vivo-like conditions, as provided by the 3D spheroid system, particularly under physiologic oxygen tension, enhance Notch signaling in human CPCs, of both fetal and adult origin. Given the role of Notch in CPCs, this might be an important mechanism for several cell aspects of cardiac regeneration, such as maintenance of the CPC pool via enhanced survival and proliferation, maintenance of CPC multipotency and tissue homeostasis. However, it is tempting to speculate that a further increased activation of Notch (due to lower oxygen concentrations) might induce differentiation of CPCs into the cardiomyogenic lineage, resulting in improved extracellular matrix production and remodeling 15, mechanical integration 121, and might ultimately favor cardiac regeneration (Figure 3). Further studies are needed to better understand the influence of microenvironmental conditions on human CPCs, and disclose their full potential for cardiac regeneration. 109
119 Figure 3: Hypothesized putative mechanisms triggered by environmental conditions in CPCs. A) In the cell monolayer, Notch activity is very low; B, C) 3D spheroid culture provides improved cell-cell and cell- ECM contact and gradients of oxygen and nutrients 105,313, more similar to the in vivo microenvironment. This results in enhanced Notch activation and might thus lead to: B) CPC proliferation and maintenance of the stem cell pool and of niche homeostasis (20% O2); or (C) cardiac differentiation, increased ECM production and modulation, mechanical integration and ultimately to a cardioprotective effect and cardiac regeneration (5% O2). 110
120 Acknowledgements This research was funded by the People Program (Marie Curie Actions) of the European Union s Seventh Framework Program FP7-People-2012-ITN TECAS, under grant agreement n and the FP7-PEOPLE-2013-CIG Stressfate, under grant agreement n Dr. Mark van Turnhout (Eindhoven University of Technology) is thankfully acknowledged for his help with confocal image analysis. 111


121 Appendix to chapter 6 Figure S1: Notch1 and Notch3 expression in fetal and adult CPC monolayer and spheroids. (A-E) Notch1 (N1) is mainly localized on the cell membrane and cytoplasm; (F-J) Notch3 (N3) shows a perinuclear distribution in all samples. Nuclei are stained with DAPI. Scale bar 50 μm Figure S2: Notch2 (N2) expression and distribution in adult (A) EDCs, (B) CSps, (C) CDCs. Scale bar 50 μm. 112
122 Figure S3: Culture under physiological oxygen condition (5% O2) does not lead to upregulation of Notch gene expression in CPCs. A-D) Gene expression of Notch components (N1, N2, N3, Jag1) and target gene (Hey1) in (A, B) fetal CMPC-CSps (n=4) and (C, D) adult CSps (n=3-7) cultured under 5% oxygen as compared to 20% O 2. *p <0.05, **p<0.01; *** p <
123 114
124 CHAPTER 7 General discussion
125 Introduction The adult human heart has long been considered a terminally differentiated organ, incapable of regeneration. Today, we know that endogenous regenerative mechanisms occur in the heart following a myocardial infarction, although these are extremely limited, and not sufficient to recover cardiac function and structure. Besides, there is uncertainty on the main mechanisms driving this process. The observed endogenous cardiac regeneration could be ascribed to proliferation of cardiomyocytes, occurring at a rate of 0.4-1% annually in human adults 314, or to differentiation of progenitor cells residing in the cardiac tissue (CPCs) 315. Several clinical trials based on CPC transplantation reported beneficial outcomes on cardiac function and tissue structure, but this positive response is limited to the short term and mainly due to paracrine effects 17,239. A deeper understanding of the microenvironment (or niche) that surrounds CPCs, and of the interplay between CPCs and the niche components, is critical to achieve a long-term, optimal cardiac regeneration. Such knowledge can allow to pinpoint the characteristics of a regenerative microenvironment, to be recreated in vitro as an ideal model system, or to potentially modulate the in vivo microenvironment and overcome the current limitations of CPC-based cardiac therapies. The aim of this thesis was to explore and understand the interactions of human CPCs with some of the key components of the cardiac microenvironment: ECM, mechanical cues such as stiffness and cyclic strain, neighboring cells, and dimensionality of the environment. In this chapter, we discuss how the provided insights contribute to the current knowledge on the CPCniche interplay, to the improvement of in vitro engineered microenvironments. Finally, a perspective analysis on the future of cardiac regeneration is provided. Main findings Cells residing in the heart are constantly subjected to the cyclic strain provided by cardiac beating. The first study focused on the response of undifferentiated and predifferentiated CPCs to uniaxial cyclic strain on 2D substrates (chapter 2). We showed that following early cardiomyogenic differentiation, CPCs developed a mature mechanosome, along with the ability to reorient perpendicular to the strain direction (strain avoidance). These observations suggest that a developed and active mechanosome is required for CPC mechanosensing and mechanoresponse. This was verified in chapter 3, where inhibition of some of the key mechanosome components resulted in reduced strain avoidance and impaired mechanosome development of predifferentiated CPCs. It can be suggested that the lack of mature/active FAs and actin SFs in undifferentiated CPCs shields them from the harsh cardiac mechanical environment. Upon early cardiomyogenic differentiation, CPCs develop their mechanosome, which allows them to sense and respond to external mechanical cues. Consequently, injection of predifferentiated CPCs would seem to favor the mechanical integration of these cells into the cardiac tissue, and therefore might provide improved outcomes, due to better cell-ecm mechanical coupling. 116
126 Cells also receive mechanical stimuli from neighboring cells via cell-cell adhesions 122,244. Therefore, in chapter 4 we studied the effect of cell density on the response of undifferentiated CPCs to cyclic strain. We found that increased cell density enhances the strain avoidance of undifferentiated CPCs, without affecting their mechanosome and differentiation state. N- cadherin mediated cell-cell interactions thus emerge as a relevant factor of the CPCmicroenvironment interaction, that can act in synergy with FAs and SFs to provide improved mechanosensing and mechanoresponse. The importance of investigating cell behavior in an environment that mimics physiologic conditions has been widely recognized. In chapter 5 we analyzed the response to cyclic strain of undifferentiated CPCs using a 3D model system. As compared to CPCs cultured on 2D substrates, in 3D fibrin hydrogels CPCs showed a shift toward the vascular fate, lower YAP activity, and displayed a strain alignment response to cyclic strain. These profound differences highlight the importance of recreating physiologic-like conditions in vitro, and the relevance of the (3D) mechanical environment on CPC behavior. The same considerations are also valid when considering cell-cell signaling pathways. Lastly, in chapter 6, the cardiosphere model system was used to study the influence of a niche-like environment on Notch signaling in CPCs. Notch expression and activation was enhanced by CPC-cardiosphere formation, and this response was particularly evident under physiologic oxygen conditions (5% O2). This suggests that a niche-like environment can have a deep influence on a crucial cellular pathway such as Notch, which drives CPC proliferation and differentiation in the developing and adult heart. On one hand, the results obtained in 2D shed light on fundamental, and previously unknown, interaction mechanisms of CPCs with their microenvironment. On the other hand, studying cell behavior in an environment that recapitulates in vivo conditions can elicit different and more physiologic responses, and provide further input for the application of CPCs for cardiac regeneration. CPC-microenvironment interplay The interplay of stem and progenitor cells with their microenvironment (or niche) directs tissue homeostasis and regeneration by driving several cell functions, such as survival, proliferation, and differentiation. In this thesis I uncovered some of the fundamental interactions of human CPCs with their microenvironment. Obviously, the CPC niche is highly complex, as it is comprised of several components that can interact with each other. Moreover, CPCs can encounter a healthy (resident) or diseased niche, that provide different stimuli. Understanding the interplay of CPCs with such stimuli can be critical for enhancing the regenerative potential of these cells. Mechanical stimuli are crucial in the beating cardiac microenvironment, and can be transmitted to cells via cell-ecm or cell-cell adhesions 122. Furthermore, mechanical signals were shown to be particularly relevant for the behavior of stem and progenitor cells 279. At early developmental stages, cadherin-mediated cell-cell adhesions prevail. As development advances, cells differentiate, start producing ECM and forming cell-ecm adhesions 246. Consequently, the transmission of mechanical signals is mainly due to cadherin-mediated 117
127 adhesions during early development; upon development progression however, integrinmediated adhesions also acquire a relevant role in mechanosensing 279. We observed a similar behavior in CPCs, where the mechanosome is developed upon early cardiomyogenic differentiation and its activity is required for the reorientation response of predifferentiated CPCs (chapters 2 and 3). On the other hand, cell density-induced cell-cell adhesions emerged as an important player in the reorientation response of undifferentiated CPCs to mechanical stimuli (chapter 4). Investigating the role of cell-cell adhesions in the mechanoresponse of predifferentiated CPCs could provide relevant information to further understand the mechanisms that underlie CPC mechanosensing and mechanoresponse, and particularly their dependence on the cell differentiation state. Rho/ROCK activity (and hence SF development and contractility) was required for the reorientation response of both undifferentiated and predifferentiated CPCs, indicating Rho/ROCK activation as central to CPC mechanoresponse. Activation of the Rho/ROCK pathway is induced by a stiff environment, resulting in higher intracellular tension 278. In turn, Rho/ROCK activates YAP/TAZ 283, nuclear transcriptional coactivators that have emerged as key cell mechanosensors and regulators of cell mechanoresponse. Modulation of YAP/TAZ activity by substrate stiffness modification can direct CPC lineage commitment 316. Moreover, we showed in chapter 5 that culture of CPCs in a soft 3D environment also leads to reduced YAP activity, which can be linked to the vascular shift in cell fate, as well as to the strain alignment response to cyclic strain due to reduced mechanosensing. It is important to mention that in chapter 5 it was not possible, due to the experimental setup, to decouple the influence of stiffness and dimensionality (2D vs 3D). Thus, the main factor responsible for the observed cell response remains unknown. Nevertheless, the findings of chapter 5 underline that very different cellular responses can be obtained in 3D vs 2D. Mimicking physiologic conditions when engineering in vitro microenvironments is therefore of paramount importance to provide more reliable and relevant results. For instance, the reorientation of CPCs, either resident or injected in the infarcted myocardium, in response to the cyclic strain provided by the beating heart could be driven by different mechanisms from what observed in our 2D in vitro experiments. In fact, in the healthy cardiac tissue, cardiomyocytes are aligned parallel to the main stretching direction; therefore the reorientation response to cyclic strain observed in 3D might be more representative of the native situation, although other mechanisms may also occur in vivo. Multicellular spheroids, such as cardiospheres (CSps), represent 3D scaffold-free model systems that are believed to better resemble the stem/progenitor cell niche, as compared to 2D in vitro setups. CSps provide improved cell-cell and cell-ecm interactions, and physiologic-like gradients of oxygen, nutrients, and waste products 105. We showed that the activity of Notch signaling, a key regulator of heart development and CPC behavior, is enhanced by culture of CPCs as CSps, particularly when the culture was performed under physiologic oxygen conditions (5% O2). Increased Notch activity in CPC-CSps may favor CPC proliferation and multipotency, and hence maintenance of the CPC pool in the resident (healthy) niche. However, Notch is known to crosstalk with several other pathways, and to be influenced by mechanical factors such as matrix 118
128 stiffness 104. Therefore, pathological conditions, such as those encountered by cells in the infarcted cardiac microenvironment, might lead to different outcomes. For example, harsh hypoxia (oxygen tension <5%) might result in further enhancement of Notch activity, and thus in cardiomyogenic or vascular differentiation of the CPCs 58,94,104. Cardiac differentiation could result in improved mechanical integration of the cells in the host tissue via mechanosome development, and ECM production and remodeling 317. Vascular differentiation of CPCs can also provide beneficial effects, by contributing to the formation of new vessels in the injured cardiac tissue. It has to be noted that the cellular response to Notch activation is also highly sensitive to dose and timing. For instance, Notch activation is known to be required for the onset of CPC cardiac differentiation, but its activity needs to be reduced for terminal cardiomyocyte differentiation 58,94,96. Therefore, Notch represents an interesting therapeutic target for cardiac regeneration, to modulate CPC proliferation and differentiation, but also to promote cardiomyocyte proliferation 318. Nevertheless, timing and dosage of stimulation, for instance via Notch ligand Jagged1, must be accurately provided. Due to the relevance and complexity of the CPC-niche interactions, future research should focus on: a) engineering in vitro model systems that allow to reproduce the physiologic microenvironment; and b) using model systems that allow to dissect and manipulate the single niche components, to be able to specifically determine the role of each factor on CPC behavior. For instance, hydrogel biomaterials with tunable mechanical properties 319 might help investigate the role of stiffness in CPC behavior in a 3D environment. At the same time, interactions such as cell-cell adhesions could be studied on micropatterned substrates, which would allow for a more precise control of cell shape, cell-ecm, and cell-cell contact. Moreover, signaling interference by protein knock-down rather than pharmacological inhibitors would improve the specificity of inhibition, and increase the specificity of in vitro studies. Such improvements can aid in the engineering of in vitro microenvironments with specific features, to further understand the CPC-niche interactions and how to enhance their regenerative potential. Implications for engineered microenvironments The importance of engineering niche-like microenvironments has become increasingly evident for the study of cell behavior and for the potential implications on regeneration. From what has been shown and discussed in this thesis, several characteristics/components of the niche emerge as critical for CPC behavior and regenerative potential. In this paragraph, the implications for engineering in vitro microenvironments will be discussed. First, an engineered niche should provide a 3D microenvironment for the cells, as this is crucial to obtain a cellular response that resembles the in vivo behavior. Several options are available to recreate the cardiac 3D microenvironment, such as decellularized ECM, hydrogels, or synthetic biomaterials. Among the material characteristics, stiffness has emerged as a critical parameter for cell fate decisions and mechanoresponse. Therefore, engineered microenvironments need to mimic physiologic stiffness values to elicit a specific (healthy or 119
129 diseased) cellular response. 3D hydrogels with tunable stiffness can be helpful in mimicking healthy and diseased cardiac conditions. Another important factor that can influence CPC behavior is matrix composition. In our 2D studies we used collagen IV as a coating protein, because it is among the main components of the cardiac basement membrane, and together with laminins it has been identified in the CPC niche in mouse hearts 28. For 3D experiments, fibrin was used due its accessibility and mechanical properties, superior to e.g. collagen/matrigel hydrogels, that made it a good material of choice for our experiments. Moreover, fibrin is a suitable biomaterial for cardiac repair 320. However, the mechanical response of CPCs can vary according to the chemical composition of the substrate 127. A microenvironment where the cardiac ECM composition is reproduced 125 could elicit the expression of more specific integrins and other FA proteins, which might influence the CPC mechanical behavior. Interestingly, culture on a cardiac-derived ECM coating has been shown to promote the cardiac lineage commitment and proliferation of CPCs 126, indicating the relevant effects that the ECM composition can have on CPC cell fate as well. In chapter 4 we have described that cell density can deeply influence the response of CPCs to mechanical cyclic loading. In addition, although we did not observe an effect of cell density on CPC differentiation on a short time-scale (3 days), cell lineage commitment might be affected by the cell density over longer time-scales. Therefore, cell density emerged as another parameter that needs to be carefully considered in 3D engineered model systems. Although several microenvironmental components were considered in this thesis, I have not evaluated the effects of co-culturing CPCs with different cell types, that they encounter in the native healthy or diseased cardiac tissue. In the healthy resident niche, CPCs are surrounded by cardiomyocytes, with cell-cell adhesions mediated by connexins and cadherins 27,46. Although co-culture of these two cell types on 2D substrates has shown to improve the differentiation potential of CPCs 50 [Kubo 2008], it would be of interest to investigate whether the same effect occurs in a 3D niche-like environment. Observing the influence of CPC-cardiomyocyte coculture on the response of CPCs to cyclic strain might provide relevant information on the mechanoresponse of CPCs in vivo, such as on the alignment/reorientation of injected CPCs in the infarcted heart. Besides, the presence of different cell types in contact with each other can elicit a direct cell-cell signaling such as Notch. For example, Notch plays a crucial role on both CPCs and endothelial cells, and CPC pools are often located close to vessels. Engineered in vitro model systems that allow a controlled CPC-endothelial cell direct contact could be used to explore Notch signaling between these two cell types, and its possible role on CPC regenerative potential. In the myocardium, adjacent mature cardiomyocytes are electrically coupled via gap junction proteins, which allow their synchronized beating. Since the focus of this thesis was on CPCs, which are undifferentiated, non-beating cells, we did not include electrical stimulation among the investigated niche components. Nevertheless, electrical pacing becomes a very relevant cue when engineering an in vitro microenvironment that mimics the cardiac niche, and for the maturation of cardiomyocyte-based engineered tissues. For example, a recent study by Nunes 120
130 et al. 321 describes the development of a biowire platform, whereby electrical stimuli can improve the tissue maturation. Another feature of the cardiac microenvironment that was not considered in this thesis is the ECM topography. Substrate topography (in 2D) was demonstrated to influence the differentiation potential of CPCs 322, and to have an effect on YAP/TAZ activation in these cells 316. Several techniques allow to reproduce the topographical cues of the cardiac ECM in 3D, as extensively reviewed 323,324. Among those, bioprinting allows to prepare 3D tissues with strictly defined geometries, as described by Pati et al Of note, CPCs have been successfully cultured in a bioprinted porous alginate scaffold, which retained cell viability and did not affect their lineage specification 326. In both healthy and diseased cardiac microenvironments, CPCs are subjected to a number of soluble factors, either released from neighboring cells, or from the ECM upon its degradation. These factors can affect CPC differentiation, survival, and migration. Transforming growth factor β (TGF-β) stimulates CPC differentiation towards the cardiomyogenic lineage 39. Moreover, the migratory capacity of CPCs can be influenced by SDF1, the release of which is increased upon hypoxic conditions 149. Recently, SDF1 gradients have been associated with the distribution of CPCs in a rat MI model 327, suggesting a possible targeting mechanism to enhance CPC migration to the infarcted area after cardiac injury. A careful dosage of soluble factors, and gradients thereof, can be used to mimic the CPC microenvironment, and modulated to reproduce either healthy or diseased cardiac conditions. Future perspectives of cardiac therapies Cardiac regenerative therapies aim at recovering the heart functionality following an MI, through the generation of new cardiomyocytes and capillaries, reduction of the scar size and scar replacement by healthy ECM. Several approaches have been considered in the past couple of decades, including cell-based and cell-free therapies 239. Due to their cardiac origin and differentiation potential, CPCs seemed to be a perfect cell source to regenerate the heart. However, despite the encouraging results of the first clinical trials, the main issues involved with CPC transplantation, namely cell survival, engraftment and differentiation in the hostile post-mi environment, remain unsolved. The findings of this thesis shed light on the interplay between CPCs and their microenvironment, which can provide valuable information to engineer a regenerative niche, and understand how to drive CPCs towards regeneration in vivo. Nevertheless, our results show that the CPC-niche interplay is highly complex, and that the interactions with single niche components are difficult to isolate and modulate without affecting the others. Therefore, understanding the CPC-niche interplay can be extremely arduous without the help of additional tools. For example, mathematical modeling, supported by experimental data, can be instrumental to dissect the complex cardiac microenvironment and identifying the key mechanisms underlying specific behaviors. 121
131 Other limitations of the described regenerative approach can be associated with the chosen cell source. The focus of this thesis was on human fetal CPCs, because of their promising characteristics and regenerative potential; CPCs are able to differentiate into all the cardiac cell types, and hence generate beating cardiomyocytes as well as new vasculature, and can produce and remodel ECM. However, this cell type entails practical issues associated with their isolation, that involves an invasive surgical procedure. In addition, the use of fetal cells raises ethical concerns as well as the risk of immune rejection, that might be overcome by using autologous adult CPCs. Unfortunately, the scarcity of these cells in the adult human heart and the difficulties associated with their isolation represent an obstacle as well. In addition, fetal and adult CPCs have been shown to possess distinct differentiation potential 328 ; as compared to fetal cells, adult CPCs can form cardiomyocytes with improved electrophysiological characteristics, and favor smooth muscle over endothelial differentiation (upon angiogenic stimulation) 328. Fetal CPCs seem to be more suitable for developmental and in vitro studies; particularly, the immortalized cell line used in this thesis (L9TB CMPCs) is characterized by more stable behavior as compared to non-immortalized cells 329. Yet, the use of adult CPCs might provide different results in vitro and especially in vivo. Moreover, as suggested by the results of chapters 2 and 3, injection of predifferentiated (fetal) CPCs in the post-mi myocardial tissue might result in improved mechanical integration in the host cardiac tissue, due to presence of a mature mechanosome. However the in vitro differentiation protocol used in this thesis, which does not require co-culture with cardiomyocytes, involves treatment of the CPCs with 5- azacytidine that decreases cell proliferation by unmasking methylated genes, and is therefore not safe for clinical application of the CPCs. Several other cell types have been investigated in vitro and in vivo for cardiac repair and regeneration 239, although an ideal cell source has not been found yet. The recent discovery of induced pluripotent stem cells (ipscs) 330 opened the field to new and exciting possibilities. ipscs are pluripotent cells derived from stromal cell reprogramming, that hold an enormous potential for cardiac regeneration due to their high cardiomyogenic differentiation capacity. For engineered in vitro models, ipscs may have major advantages, such as the relative ease of access and the ability of differentiation in multiple cell types. ipscs might therefore represent a valuable alternative to more traditional cell sources, such as CPCs, for the in vitro study of cell behavior and of cardiac regenerative mechanisms. Moreover, transplantation of ipscs, or ipscderived cells, could entail great advantages for tissue regeneration, e.g. avoiding the risk of immune rejection and circumventing ethical issues. Nonetheless, transplantation of ipscs can induce the formation of teratoma 331, which of course questions the safety of this method. Despite the great promise held by ipscs, further research and careful characterization of these cells and of the safety of their vivo application are needed. Furthermore, ipsc injection still involves some of the issues associated with traditional stem cells, such as low survival and engraftment. Cell retention in the host tissue could be aided by delivering the cells with a biomaterial. This should be biodegradable and biocompatible, and preferably injectable to allow for non-invasive procedures. A number of biomaterials, either of natural or synthetic nature, has been 122
132 developed to promote stem/progenitor cell engraftment, that can be modified with adhesion peptides or proteins to improve cell attachment 332. In vitro models that closely mimic the cardiac niche can be of great help to study fundamental biological processes, and test new potential therapeutic approaches. However, they cannot predict the response of the whole organ/organism to therapeutic stimuli. Once again, mathematical models might help overcome this limitation, and circumvent the ethical issues involved with in vivo experiments. Nevertheless, pre-clinical studies are still necessary to understand the actual biological responses, and assess the safety of specific therapies. Despite the generation of new cardiomyocytes and capillaries following CPC injection, that might derive from cell differentiation, it seems that most of the beneficial effects of CPC therapies is provided by paracrine effects 333,334. CPCs release a number of cytokines and growth factors, as well as coding and non-coding RNAs contained in extracellular vesicles known as exosomes 333,335. Exosomes are small membrane vesicles ( nm) secreted by cells in response to physiological changes 333. They are produced by almost all cell types, and are involved in several cellular processes, such as long range intercellular communications, regulation of cell signaling and even immune modulation 333. While originally considered as mere cell waste, exosomes have recently been enormously reconsidered, in light of their role in (cardiac) tissue repair 336. In fact, delivery of exosomes from cardiosphere-derived cells (CDCs) was shown to reproduce most of the beneficial effects of CDC injection 333,336. Additionally, recent studies demonstrated the beneficial effects of delivering CDC exosomes in porcine MI models 337,338. Intramyocardial exosome injection resulted in attenuated cardiomyocyte apoptosis, increased angiogenesis, reduced scar size and impaired adverse remodeling, as well as enhanced left ventricular ejection fraction 337,338. Although the exosome content is diverse, most of the regenerative and cardioprotective effects seem to be provided by mirnas, such as mir210, mir , and mir146-a 339. Yet, whereas injection of the whole exosome produced the same positive effects as CDC delivery, the single mir146-a only partially reproduced these outcomes 339, indicating that other exosome components are required as well. Interestingly, ipscs-derived extracellular vesicles showed superior effects on cardiac function, angiogenesis and apoptosis, compared with ipscs 340. Thanks to these encouraging results, in addition to the proven safety and reduced immunogenicity of their delivery 337,340, exosomes appear as a valuable alternative to stem/progenitor cell therapy for cardiac repair. Nevertheless, further research is needed to deeply understand exosome biology and their effects on the different cells residing in the heart, to avoid undesired off-target outcomes. Moreover, the exosome cargo is highly dependent on the physiological state of the parent cell, and can be affected by disease 336,341. Therefore, a strict control over the conditions of exosome collection is required to prevent unwanted effects. Advantages and disadvantages can be listed for both cell-based and cell-free approaches. On one hand, cell delivery has the potential to regenerate the infarcted myocardium via 123
133 differentiation into cardiac cell types and scar tissue remodeling, although the issues associated with low cell engraftment and differentiation remain. On the other hand, exosomes have shown comparable or even improved benefits on cardiac function and angiogenesis, and allow to overcome cell-associated issues, such as ethics and safety. However exosomes might target the wrong cell type or deliver an undesirable cargo, and thus cause unexpected effects 341. Further research and (pre-)clinical studies will hopefully prove which is the best alternative to recover the heart functionality after an MI. Conclusion The aim of this thesis was to provide insights into the complex interplay between CPCs and their microenvironment, or niche. Our results show that the mechanoresponse of CPCs in 2D relies on the one hand on their mechanosome and on Rho/ROCK signaling, which are developed and activated upon early cardiomyogenic differentiation, and on the other hand on cell-cell adhesions induced by cell density. However, a more physiological 3D culture environment was shown to induce a shift in CPC fate and to change their response to cyclic stretching as compared to 2D environments. A 3D niche-like microenvironment can also alter a key cell-cell signaling pathway such as Notch, that regulates CPC fate decisions and cardiac development. These findings prove the importance of cell-matrix and cell-cell interactions in CPC mechanical behavior, and highlight that recapitulating the physiologic conditions is critical to engineer a regenerative in vitro niche, and determine the specific microenvironmental conditions required to elicit a regenerative response of CPCs. At the same time, due to the complexity of the CPC-niche interplay, understanding the key regulators of specific regenerative mechanisms requires computational models and in vitro model systems that allow for isolation and precise control of single CPC-microenvironment interactions. 124
134 BIBLIOGRAPHY 1. Ripplinger, C. M., Noujaim, S. F. & Linz, D. The nervous heart. Prog. Biophys. Mol. Biol. 120, (2016). 2. Ieda, M. Heart Development, Diseases, and Regeneration- New Approaches From Innervation, Fibroblasts, and Reprogramming. Circ. J. 80, (2016). 3. Smart, N. Prospects for improving neovascularization of the ischemic heart: Lessons from development. Microcirculation 24, (2017). 4. Graham, E. & Bergmann, O. Dating the Heart: Exploring Cardiomyocyte Renewal in Humans. Physiology (Bethesda). 32, (2017). 5. Bergmann, O. et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell (2015). doi: /j.cell Urbanek, K. et al. Myocardial regeneration by activation of multipotent cardiac stem cells in ischemic heart failure. Proc. Natl. Acad. Sci. U. S. A. 102, (2005). 7. Bloomekatz, J., Galvez-Santisteban, M. & Chi, N. C. Myocardial plasticity: cardiac development, regeneration and disease. Curr. Opin. Genet. Dev. 40, (2016). 8. van Berlo, J. H. & Molkentin, J. D. An emerging consensus on cardiac regeneration. Nat. Med. 20, (2014). 9. Liang, S. X. & Phillips, W. D. Migration of Resident Cardiac Stem Cells in Myocardial Infarction. Anat. Rec. 296, (2013). 10. Oh, H. et al. Cardiac progenitor cells from adult myocardium : Homing, differentiation, and fusion after infarction. 1 6 (2003). 11. Le, T. Y. L. & Chong, J. J. H. Cardiac progenitor cells for heart repair. Nat. Publ. Gr. 2, 1 4 (2016). 12. Urbanek, K. et al. Intense myocyte formation from cardiac stem cells in human cardiac hypertrophy. Proc. Natl. Acad. Sci. U. S. A. 100, (2003). 13. Matsuura, K. et al. Adult Cardiac Sca-1-positive Cells Differentiate into Beating Cardiomyocytes. J. Biol. Chem. 279, (2004). 14. Pfister, O. et al. CD31- but not CD31+ cardiac side population cells exhibit functional cardiomyogenic differentiation. Circ. Res. 97, (2005). 15. Bax, N. a M. et al. Matrix production and remodeling capacity of cardiomyocyte progenitor cells during in vitro differentiation. J. Mol. Cell. Cardiol. 53, (2012). 16. Boyle, A. J., Schulman, S. P., Hare, J. M. & Oettgen, P. Is stem cell therapy ready for patients? Stem Cell Therapy for Cardiac Repair. Ready for the Next Step. Circulation 114, (2006). 17. Zwetsloot, P. P. et al. Cardiac stem cell treatment in myocardial infarction: A systematic
135 126 review and meta-analysis of preclinical studies. Circ. Res. 118, (2016). 18. Menasche, P. Cardiac cell therapy: lessons from clinical trials. J. Mol. Cell. Cardiol. 50, (2011). 19. Malliaras, K. & Marban, E. Cardiac cell therapy: where we ve been, where we are, and where we should be headed. Br. Med. Bull. 98, (2011). 20. Vanhoutte, D., Schellings, M., Pinto, Y. & Heymans, S. Relevance of matrix metalloproteinases and their inhibitors after myocardial infarction: A temporal and spatial window. Cardiovasc. Res. 69, (2006). 21. Deddens, J. C. et al. Modeling the Human Scarred Heart In Vitro: Toward New Tissue Engineered Models. Adv. Healthc. Mater , (2016). 22. Dobaczewski, M., Gonzalez-Quesada, C. & Frangogiannis, N. G. The extracellular matrix as a modulator of the inflammatory and reparative response following myocardial infarction. J. Mol. Cell. Cardiol. 48, (2010). 23. Jones, D. L. & Wagers, A. J. No place like home: anatomy and function of the stem cell niche. Nat. Rev. Mol. Cell Biol. 9, (2008). 24. Moore, K. A., Lemischka, I. R., Intestinal, T. & Cell, S. Stem Cells and Their Niches. (1978). 25. Schofield, R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells 4, 7 25 (1978). 26. Morrison, S. J. & Spradling, A. C. Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell 132, (2008). 27. Urbanek, K. et al. Stem cell niches in the adult mouse heart. (2006). 28. Schenke-Layland, K. et al. Recapitulation of the embryonic cardiovascular progenitor cell niche. Biomaterials 32, (2011). 29. Leri, A., Rota, M., Hosoda, T., Goichberg, P. & Anversa, P. Cardiac stem cell niches. Stem Cell Res. 13, (2014). 30. Beltrami, A. P. et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 114, (2003). 31. Mauretti, A., Spaans, S., Bax, N. A. M., Sahlgren, C. & Bouten, C. V. C. Cardiac Progenitor Cells and the Interplay with Their Microenvironment. Stem Cells Int. 2017, (2017). 32. Ellison, G. M. et al. Cardiac stem and progenitor cell identification: different markers for the same cell? Front. Biosci. (Schol. Ed). 2, (2010). 33. Tallini, Y. N. et al. C-Kit Expression Identifies Cardiovascular Precursors in the Neonatal Heart. Proc Natl Acad Sci U S A 106, (2009). 34. Bolli, R. et al. Cardiac stem cells in patients with ischaemic cardiomyopathy (SCIPIO): initial results of a randomised phase 1 trial. Lancet (London, England) 378, (2011). 35. Chugh, A. R. et al. Administration of cardiac stem cells in patients with ischemic
136 cardiomyopathy: the SCIPIO trial: surgical aspects and interim analysis of myocardial function and viability by magnetic resonance. Circulation 126, S54-64 (2012). 36. van Vliet, P. et al. Progenitor cells isolated from the human heart: a potential cell source for regenerative therapy. Neth. Heart J. 16, (2008). 37. Wang, X. et al. The role of the sca-1+/cd31- cardiac progenitor cell population in postinfarction left ventricular remodeling. Stem Cells 24, (2006). 38. Dey, D. et al. Dissecting the molecular relationship among various cardiogenic progenitor cells. Circ. Res. 112, (2013). 39. Goumans, M. J. et al. TGF-β1 induces efficient differentiation of human cardiomyocyte progenitor cells into functional cardiomyocytes in vitro. Stem Cell Res. 1, (2008). 40. Smits, A. M. et al. Human cardiomyocyte progenitor cells differentiate into functional mature cardiomyocytes: an in vitro model for studying human cardiac physiology and pathophysiology. Nat. Protoc. 4, (2009). 41. van Vliet, P. et al. Hyperpolarization induces differentiation in human cardiomyocyte progenitor cells. Stem Cell Rev. 6, (2010). 42. van Vliet, P. et al. Foetal and adult cardiomyocyte progenitor cells have different developmental potential. J. Cell. Mol. Med. 14, (2010). 43. Matsuura, K. et al. Transplantation of cardiac progenitor cells ameliorates cardiac dysfunction after myocardial infarction in mice. J. Clin. Invest. 119, (2009). 44. Wang, L. et al. Transplantation of Isl1+ cardiac progenitor cells in small intestinal submucosa improves infarcted heart function. Stem Cell Res. Ther. 8, 230 (2017). 45. Bearzi, C. et al. Human cardiac stem cells. Proc. Natl. Acad. Sci. U. S. A. 104, (2007). 46. Bearzi, C. et al. Identification of a coronary vascular progenitor cell in the human heart. Proc. Natl. Acad. Sci. U. S. A. 106, (2009). 47. Messina, E. et al. Isolation and expansion of adult cardiac stem cells from human and murine heart. Circ. Res. 95, (2004). 48. Laugwitz, K.-L. et al. Postnatal isl1+ cardioblasts enter fully differentiated cardiomyocyte lineages. Nature 433, (2005). 49. Yamahara, K. et al. Heterogeneic nature of adult cardiac side population cells. Biochem. Biophys. Res. Commun. 371, (2008). 50. Kubo, H. et al. Increased cardiac myocyte progenitors in failing human hearts. Circulation 118, (2008). 51. Flanders, K. C., Holder, M. G. & Winokur, T. S. Autoinduction of mrna and protein expression for transforming growth factor-beta S in cultured cardiac cells. J Mol Cell Cardiol 27, (1995). 52. Behfar, A. et al. Stem cell differentiation requires a paracrine pathway in the heart. 127
137 128 FASEB J. 16, (2002). 53. Deb, A. Cell-cell interaction in the heart via Wnt/beta-catenin pathway after cardiac injury. Cardiovasc. Res. 102, (2014). 54. Hosoda, T. et al. Human cardiac stem cell differentiation is regulated by a mircrine mechanism. Circulation 123, (2011). 55. Gonzalez, A. et al. Activation of cardiac progenitor cells reverses the failing heart senescent phenotype and prolongs lifespan. Circ. Res. 102, (2008). 56. Samarel, A. M. Costameres, focal adhesions, and cardiomyocyte mechanotransduction. Am. J. Physiol. Heart Circ. Physiol. 289, H (2005). 57. High, F. A. et al. Endothelial expression of the Notch ligand Jagged1 is required for vascular smooth muscle development. Proc. Natl. Acad. Sci. 105, (2008). 58. Gude, N. et al. Notch activation enhances lineage commitment and protective signaling in cardiac progenitor cells. Basic Res. Cardiol. 110, (2015). 59. Urbich, C. et al. Soluble factors released by endothelial progenitor cells promote migration of endothelial cells and cardiac resident progenitor cells.[see comment]. J. Mol. Cell. Cardiol. 39, (2005). 60. Yoon, J., Choi, S. C., Park, C. Y., Shim, W. J. & Lim, D.-S. Cardiac side population cells exhibit endothelial differentiation potential. Exp. Mol. Med. 39, (2007). 61. Lushaj, E. B., Lozonschi, L., Barnes, M., Anstadt, E. & Kohmoto, T. Mitochondrial DNA deletion mutations in adult mouse cardiac side population cells. Mutat. Res. - Fundam. Mol. Mech. Mutagen. 734, (2012). 62. Zlatanova, I., Pinto, C. & Silvestre, J.-S. Immune Modulation of Cardiac Repair and Regeneration: The Art of Mending Broken Hearts. Front. Cardiovasc. Med. 3, 40 (2016). 63. Fujiu, K., Wang, J. & Nagai, R. Cardioprotective function of cardiac macrophages. Cardiovasc. Res. 102, (2014). 64. Hsieh, P. C. H., Davis, M. E., Gannon, J., MacGillivray, C. & Lee, R. T. Controlled delivery of PDGF-BB for myocardial protection using injectable self-assembling peptide nanofibers. J. Clin. Invest. 116, (2006). 65. Vannella, K. M. & Wynn, T. A. Mechanisms of Organ Injury and Repair by Macrophages. Annu. Rev. Physiol. 79, (2017). 66. de Couto, G. et al. Macrophages mediate cardioprotective cellular postconditioning in acute myocardial infarction. J. Clin. Invest. 125, (2015). 67. Campbell, K. S. & Hasegawa, J. Natural killer cell biology: an update and future directions. J. Allergy Clin. Immunol. 132, (2013). 68. Boukouaci, W. et al. Natural killer cell crosstalk with allogeneic human cardiac-derived stem/progenitor cells controls persistence. Cardiovasc. Res. 104, (2014). 69. Zhou, Y. et al. CD117-positive cells of the heart: progenitor cells or mast cells? J.
138 Histochem. Cytochem. 58, (2010). 70. Takeda, N. & Manabe, I. Cellular Interplay between Cardiomyocytes and Nonmyocytes in Cardiac Remodeling. Int. J. Inflam. 2011, (2011). 71. Balakumar, P. et al. Resident cardiac mast cells: are they the major culprit in the pathogenesis of cardiac hypertrophy? Basic Clin. Pharmacol. Toxicol. 102, 5 9 (2008). 72. Zhang, X. et al. Cardiomyocyte differentiation induced in cardiac progenitor cells by cardiac fibroblast-conditioned medium. Exp. Biol. Med. (Maywood). 239, (2014). 73. Moore-Morris, T. et al. Resident fibroblast lineages mediate pressure overload-induced cardiac fibrosis. J. Clin. Invest. 124, (2014). 74. Zhou, B. et al. Adult mouse epicardium modulates myocardial injury by secreting paracrine factors. J. Clin. Invest. 121, (2011). 75. Limana, F. et al. Myocardial infarction induces embryonic reprogramming of epicardial c- kit(+) cells: role of the pericardial fluid. J. Mol. Cell. Cardiol. 48, (2010). 76. Gittenberger-de Groot, A. C., Winter, E. M. & Poelmann, R. E. Epicardium-derived cells (EPDCs) in development, cardiac disease and repair of ischemia. J. Cell. Mol. Med. 14, (2010). 77. Lepilina, A. et al. A dynamic epicardial injury response supports progenitor cell activity during zebrafish heart regeneration. Cell 127, (2006). 78. Lie-Venema, H. et al. Origin, fate, and function of epicardium-derived cells (EPDCs) in normal and abnormal cardiac development. ScientificWorldJournal. 7, (2007). 79. Wessels, A. & Perez-Pomares, J. M. The epicardium and epicardially derived cells (EPDCs) as cardiac stem cells. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 276, (2004). 80. Bollini, S., Smart, N. & Riley, P. R. Resident cardiac progenitor cells: At the heart of regeneration. J. Mol. Cell. Cardiol. 50, (2011). 81. Chen, T. H. P. et al. Epicardial induction of fetal cardiomyocyte proliferation via a retinoic acid-inducible trophic factor. Dev. Biol. 250, (2002). 82. Winter, E. M. et al. A new direction for cardiac regeneration therapy: application of synergistically acting epicardium-derived cells and cardiomyocyte progenitor cells. Circ. Heart Fail. 2, (2009). 83. Popescu, L. M. & Faussone-Pellegrini, M.-S. TELOCYTES - a case of serendipity: the winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. Journal of cellular and molecular medicine 14, (2010). 84. Gherghiceanu, M. & Popescu, L. M. Cardiomyocyte precursors and telocytes in epicardial stem cell niche: Electron microscope images. J. Cell. Mol. Med. 14, (2010). 85. Popescu, L. M., Fertig, E. T. & Gherghiceanu, M. Reaching out: Junctions between cardiac telocytes and cardiac stem cells in culture. J. Cell. Mol. Med. 20, (2016). 129
139 86. Bei, Y., Wang, F., Yang, C. & Xiao, J. Telocytes in regenerative medicine. J. Cell. Mol. Med. 19, (2015). 87. Bani, D., Formigli, L., Gherghiceanu, M. & Faussone-Pellegrini, M. S. Telocytes as supporting cells for myocardial tissue organization in developing and adult heart. J. Cell. Mol. Med. 14, (2010). 88. Manole, C. G., Cismasiu, V., Gherghiceanu, M. & Popescu, L. M. Experimental acute myocardial infarction: telocytes involvement in neo-angiogenesis. J. Cell. Mol. Med. 15, (2011). 89. Cismasiu, V. B., Radu, E. & Popescu, L. M. mir-193 expression differentiates telocytes from other stromal cells. J. Cell. Mol. Med. 15, (2011). 90. Luxán, G., D Amato, G., MacGrogan, D. & de la Pompa, J. L. Endocardial Notch Signaling in Cardiac Development and Disease. Circ. Res. 118, e1 e18 (2016). 91. Gude, N. & Sussman, M. Notch signaling and cardiac repair. J. Mol. Cell. Cardiol. 52, (2012). 92. Niessen, K. & Karsan, A. Notch signaling in cardiac development. Circ. Res. 102, (2008). 93. De la Pompa, J. L. & Epstein, J. A. Coordinating Tissue Interactions: Notch Signaling in Cardiac Development and Disease. Dev. Cell 22, (2012). 94. Boni, A. et al. Notch1 regulates the fate of cardiac progenitor cells. Proc. Natl. Acad. Sci. U. S. A. 105, (2008). 95. Kwon, C. et al. A regulatory pathway involving Notch1/beta-catenin/Isl1 determines cardiac progenitor cell fate. Nat. Cell Biol. 11, (2009). 96. Collesi, C., Zentilin, L., Sinagra, G. & Giacca, M. Notch1 signaling stimulates proliferation of immature cardiomyocytes. J. Cell Biol. 183, (2008). 97. Øie, E. et al. Activation of Notch signaling in cardiomyocytes during post-infarction remodeling. Scand. Cardiovasc. J. 44, (2010). 98. Gude, N. A. et al. Activation of Notch-mediated protective signaling in the myocardium. Circ. Res. 102, (2008). 99. Kratsios, P. et al. Distinct roles for cell-autonomous notch signaling in cardiomyocytes of the embryonic and adult heart. Circ. Res. 106, (2010) Yu, B. & Song, B. Notch 1 Signalling Inhibits Cardiomyocyte Apoptosis in Ischaemic Postconditioning. Hear. Lung Circ. 23, (2014) Rizzo, P. et al. The role of notch in the cardiovascular system: potential adverse effects of investigational notch inhibitors. Front. Oncol. 4, 384 (2014) Mazzone, M. et al. Dose-dependent induction of distinct phenotypic responses to Notch pathway activation in mammary epithelial cells. Proc. Natl. Acad. Sci. U. S. A. 107, (2010). 130
140 103. Guentchev, M. & McKay, R. D. G. Notch controls proliferation and differentiation of stem cells in a dose-dependent manner. Eur. J. Neurosci. 23, (2006) Boopathy, A. V et al. The modulation of cardiac progenitor cell function by hydrogeldependent Notch1 activation. Biomaterials 35, (2014) Günter, J. et al. Microtissues in Cardiovascular Medicine: Regenerative Potential Based on a 3D Microenvironment. Stem Cells Int. 2016, (2016) Forte, E. et al. TGFβ-Dependent Epithelial-to-Mesenchymal Transition Is Required to Generate Cardiospheres from Human Adult Heart Biopsies. Stem Cells Dev. 21, (2012) Mauretti, A., Rossi, F., Bax, N. A. M., Miano, C. & Miraldi, F. Spheroid 3D culture and hypoxia activate Notch signaling in cardiac progenitor cells. (2017) Rossi, M. A. Connective tissue skeleton in the normal left ventricle and in hypertensive left ventricular hypertrophy and chronic chagasic myocarditis. Med Sci Monit 7: (2001) Jourdan-Lesaux, C., Zhang, J. & Lindsey, M. L. Extracellular matrix roles during cardiac repair. Life Sci. 87, (2010) Matsui, Y., Morimoto, J. & Uede, T. Role of matricellular proteins in cardiac tissue remodeling after myocardial infarction. World J. Biol. Chem. 1, (2010) Jugdutt, B. I. Ventricular remodeling after infarction and the extracellular collagen matrix: When is enough enough? Circulation 108, (2003) Engebretsen, K. V. T. et al. Decorin, lumican, and their GAG chain-synthesizing enzymes are regulated in myocardial remodeling and reverse remodeling in the mouse. J. Appl. Physiol. 114, (2013) Imanaka-Yoshida, K. Tenascin-C in Cardiovascular Tissue Remodeling. Circ. J. 76, (2012) Martin, G. R. & Timpl, R. Laminin and other Basement Membrane Components. Annu. Rev. Cell Biol. (1987). doi: /annurev.cb Corda, S., Samuel, J. L. & Rappaport, L. Extracellular matrix and growth factors during heart growth. Heart Fail. Rev. 5, (2000) Carmeliet, P. & Jain, R. K. Molecular mechanisms and clinical applications of angiogenesis. Nature 473, (2011) Bromage, D. I., Davidson, S. M. & Yellon, D. M. Stromal derived factor 1α: A chemokine that delivers a two-pronged defence of the myocardium. Pharmacol. Ther. 143, (2014) Porter, K. E. & Turner, N. a. Cardiac fibroblasts: At the heart of myocardial remodeling. Pharmacol. Ther. 123, (2009) Israeli-Rosenberg, S., Manso, A. M., Okada, H. & Ross, R. S. Integrins and integrinassociated proteins in the cardiac myocyte. Circ. Res. 114, (2014). 131
141 120. Merna, N. J. et al. Differential β3 Integrin Expression Regulates the Response of Human Lung and Cardiac Fibroblasts to Extracellular Matrix and Its Components. Tissue Eng. Part A 21, 1 11 (2015) Mauretti, A. et al. Cardiomyocyte progenitor cell mechanoresponse unrevealed: strain avoidance and mechanosome development. Integr. Biol. 8, (2016) Han, M. K. L. & de Rooij, J. Converging and Unique Mechanisms of Mechanotransduction at Adhesion Sites. Trends Cell Biol. xx, 1 12 (2016) Tojkander, S., Gateva, G. & Lappalainen, P. Actin stress fibers--assembly, dynamics and biological roles. J. Cell Sci. 125, (2012) Cosgrove, B. D. et al. Role of extracellular matrix signaling cues in modulating cell fate commitment for cardiovascular tissue engineering. Adv. Healthc. Mater. 3, (2016) French, K. M. et al. A naturally derived cardiac extracellular matrix enhances cardiac progenitor cell behavior in vitro. Acta Biomater. 8, (2012) Gaetani, R. et al. Cardiac derived extracellular matrix enhances cardiogenic properties of human cardiac progenitor cells. Cell Transplant. 25, (2015) French, K. M. et al. Fibronectin and cyclic strain improve cardiac progenitor cell regenerative potential in vitro. Stem Cells Int. (2016) Konstandin, M. H. et al. Fibronectin is essential for reparative cardiac progenitor cell response after myocardial infarction. Circ. Res. 113, (2013) Williams, C. et al. Cardiac extracellular matrix fibrin hybrid scaffolds with tunable properties for cardiovascular tissue engineering. Acta Biomater. 14, (2015) Tallawi, M., Rai, R., Boccaccini, A. R. & Aifantis, K. Effect of substrate mechanics on cardiomyocyte maturation and growth. Tissue Eng. Part B. Rev. 21, (2014) Young, J. L. & Engler, A. J. Hydrogels with time-dependent material properties enhance cardiomyocyte differentiation in vitro. Biomaterials 32, (2011) Choi, M. Y. et al. Engineered extracellular microenvironment with a tunable mechanical property for controlling cell behavior and cardiomyogenic fate of cardiac stem cells. Acta Biomater. 50, (2017) Hannafin, J. A., Attia, E. A., Henshaw, R., Warren, R. F. & Bhargava, M. M. Effect of cyclic strain and plating matrix on cell proliferation and integrin expression by ligament fibroblasts. J. Orthop. Res. 24, (2006) Balcioglu, H. E., van Hoorn, H., Donato, D. M., Schmidt, T. & Danen, E. H. J. The integrin expression profile modulates orientation and dynamics of force transmission at cellmatrix adhesions. J. Cell Sci. 128, (2015) van Marion, M. H. et al. Behavior of CMPCs in unidirectional constrained and stress-free 3D hydrogels. J. Mol. Cell. Cardiol. 87, (2015) Mohyeldin, A., Garzón-Muvdi, T. & Quiñones-Hinojosa, A. Oxygen in stem cell biology: a 132
142 critical component of the stem cell niche. Cell Stem Cell 7, (2010) Keith, B. & Simon, M. C. Hypoxia-inducible factors, stem cells, and cancer. Cell 129, (2007) Gustafsson, M. V. et al. Hypoxia requires Notch signaling to maintain the undifferentiated cell state. Dev. Cell 9, (2005) Adams, J. M. et al. HIF-1: a key mediator in hypoxia. Acta Physiol. Hung. 96, (2009) Jurgensen, J. S. et al. Persistent induction of HIF-1alpha and -2alpha in cardiomyocytes and stromal cells of ischemic myocardium. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 18, (2004) Ceradini, D. J. et al. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nat. Med. 10, (2004) Staller, P. et al. Chemokine receptor CXCR4 downregulated by von Hippel-Lindau tumour suppressor pvhl. Nature 425, (2003) Zernecke, A. et al. SDF-1alpha/CXCR4 axis is instrumental in neointimal hyperplasia and recruitment of smooth muscle progenitor cells. Circ. Res. 96, (2005) Jujo, K. et al. CXCR4 blockade augments bone marrow progenitor cell recruitment to the neovasculature and reduces mortality after myocardial infarction. Proc. Natl. Acad. Sci. U. S. A. 107, (2010) Zheng, H., Fu, G., Dai, T. & Huang, H. Migration of endothelial progenitor cells mediated by stromal cell-derived factor-1alpha/cxcr4 via PI3K/Akt/eNOS signal transduction pathway. J. Cardiovasc. Pharmacol. 50, (2007) Ghadge, S. K., Muhlstedt, S., Ozcelik, C. & Bader, M. SDF-1alpha as a therapeutic stem cell homing factor in myocardial infarction. Pharmacol. Ther. 129, (2011) Smart, N. & Riley, P. R. The stem cell movement. Circ. Res. 102, (2008) Emmert, M. Y. et al. Higher frequencies of BCRP+ cardiac resident cells in ischaemic human myocardium. Eur. Heart J. 34, (2013) Tang, Y. L. et al. Hypoxic preconditioning enhances the benefit of cardiac progenitor cell therapy for treatment of myocardial infarction by inducing CXCR4 expression. Circ. Res. 104, (2009) Chen, D. et al. Crosstalk between SDF-1/CXCR4 and SDF-1/CXCR7 in cardiac stem cell migration. Sci. Rep. 5, (2015) van Oorschot, A. a M., Smits, A. M., Pardali, E., Doevendans, P. a & Goumans, M.-J. Low oxygen tension positively influences cardiomyocyte progenitor cell function. J. Cell. Mol. Med. 15, (2011) Ceradini, D. J. & Gurtner, G. C. Homing to hypoxia: HIF-1 as a mediator of progenitor cell recruitment to injured tissue. Trends Cardiovasc. Med. 15, (2005). 133
143 153. Davis, D. R. et al. Validation of the Cardiosphere Method to Culture Cardiac Progenitor Cells from Myocardial Tissue. PLoS One 4, e7195 (2009) Li, T.-S. et al. Cardiospheres recapitulate a niche-like microenvironment rich in stemness and cell-matrix interactions, rationalizing their enhanced functional potency for myocardial repair. Stem Cells 28, (2010) Achilli, T.-M., Meyer, J. & Morgan, J. R. Advances in the formation, use and understanding of multi-cellular spheroids. Expert Opin. Biol. Ther. 12, (2012) Li, Y., Hiroi, Y. & Liao, J. K. Notch signaling as an important mediator of cardiac repair and regeneration after myocardial infarction. Trends Cardiovasc. Med. 20, (2010) Gattazzo, F., Urciuolo, A. & Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta - Gen. Subj. 1840, (2014) Cambria, E. et al. Cardiac regenerative medicine: The potential of a new generation of stem cells. Transfus. Med. Hemotherapy 43, (2016) Pok, S., Myers, J. D., Madihally, S. V & Jacot, J. G. A multilayered scaffold of a chitosan and gelatin hydrogel supported by a PCL core for cardiac tissue engineering. Acta Biomater. 9, (2013) van Marion, M. H. et al. Behavior of CMPCs in unidirectional constrained and stress-free 3D hydrogels. J. Mol. Cell. Cardiol. 87, (2015) Johnson, T. D. et al. Human versus porcine tissue sourcing for an injectable myocardial matrix hydrogel. Biomater. Sci. 29, (2014) Gaetani, R. et al. Cardiac tissue engineering using tissue printing technology and human cardiac progenitor cells. Biomaterials 33, (2012) Chimenti, I. et al. in Somatic Stem Cells: Methods and Protocols (ed. Singh, S. R.) (Humana Press, 2012). doi: / _ Liu, T. C. K., Ismail, S., Brennan, O., Hastings, C. & Duffy, G. P. Encapsulation of cardiac stem cells in superoxide dismutase-loaded alginate prevents doxorubicin-mediated toxicity. J. Tissue Eng. Regen. Med. 7, (2013) Leor, J., Amsalem, Y. & Cohen, S. Cells, scaffolds, and molecules for myocardial tissue engineering. Pharmacol. Ther. 105, (2005) Cheng, K. et al. Transplantation of platelet gel spiked with cardiosphere-derived cells boosts structural and functional benefits relative to gel transplantation alone in rats with myocardial infarction. Biomaterials 33, (2012) Tiburcy, M. et al. Defined Engineered Human Myocardium with Advanced Maturation for Applications in Heart Failure Modelling and Repair. Circulation CIRCULATIONAHA (2017). doi: /circulationaha Nunes, S. S. et al. Biowire: a platform for maturation of human pluripotent stem cellderived cardiomyocytes. Nat. Methods 10, (2013). 134
144 169. Tulloch, N. L. et al. Growth of engineered human myocardium with mechanical loading and vascular coculture. Circ. Res. 109, (2011) Zhang, D. et al. Tissue-engineered cardiac patch for advanced functional maturation of human ESC-derived cardiomyocytes. Biomaterials 34, (2013) Ghafar-Zadeh, E., Waldeisen, J. R. & Lee, L. P. Engineered approaches to the stem cell microenvironment for cardiac tissue regeneration. Lab Chip 11, (2011) Discher, D. E., Mooney, D. J. & Zandstra, P. W. Growth factors, matrices, and forces combine and control stem cells. Science 324, (2009) Discher, D. E., Janmey, P. & Wang, Y.-L. Tissue cells feel and respond to the stiffness of their substrate. Science 310, (2005) Throm Quinlan, A. M., Sierad, L. N., Capulli, A. K., Firstenberg, L. E. & Billiar, K. L. Combining dynamic stretch and tunable stiffness to probe cell mechanobiology in vitro. PLoS One 6, e23272 (2011) Tandon, N. et al. Electrical stimulation systems for cardiac tissue engineering. Nat. Protoc. 4, (2009) Murtuza, B., Nichol, J. W. & Khademhosseini, A. Micro- and Nanoscale Control of the Cardiac Stem Cell Niche for Tissue Fabrication. Tissue Eng. Part B 15, (2009) Morez, C. et al. Enhanced efficiency of genetic programming toward cardiomyocyte creation through topographical cues. Biomaterials 70, (2015) Leri, A., Kajstura, J. a N. & Anversa, P. Cardiac Stem Cells and Mechanisms of Myocardial Regeneration. Physiol Rev 86, (2005) Smits, A. M. et al. Human cardiomyocyte progenitor cell transplantation preserves longterm function of the infarcted mouse myocardium. Cardiovasc. Res. 83, (2009) van Vliet, P., Goumans, M.-J., Doevendans, P. a & Sluijter, J. P. G. Human cardiomyocyte progenitor cells: a short history of nearly everything. J. Cell. Mol. Med. 16, (2012) Barron, V. et al. The effect of physiological cyclic stretch on the cell morphology, cell orientation and protein expression of endothelial cells. J. Mater. Sci. Mater. Med. 18, (2007) Ngu, H. et al. Effect of focal adhesion proteins on endothelial cell adhesion, motility and orientation response to cyclic strain. Ann. Biomed. Eng. 38, (2010) Boccafoschi, F., Bosetti, M., Gatti, S. & Cannas, M. Dynamic fibroblast cultures: response to mechanical stretching. Cell Adh. Migr. 1, (2007) Jungbauer, S., Gao, H., Spatz, J. P. & Kemkemer, R. Two characteristic regimes in frequency-dependent dynamic reorientation of fibroblasts on cyclically stretched substrates. Biophys. J. 95, (2008) Standley, P. R., Cammarata, A., Nolan, B. P., Purgason, C. T. & Stanley, M. a. Cyclic stretch induces vascular smooth muscle cell alignment via NO signaling. Am. J. Physiol. Heart 135
145 136 Circ. Physiol. 283, H1907 H1914 (2002) Livne, A., Bouchbinder, E. & Geiger, B. Cell reorientation under cyclic stretching. Nat. Commun. 5, 3938 (2014) Tondon, A., Hsu, H. J. & Kaunas, R. Dependence of cyclic stretch-induced stress fiber reorientation on stretch waveform. J. Biomech. 45, (2012) Hsu, H.-J., Lee, C.-F. & Kaunas, R. A dynamic stochastic model of frequency-dependent stress fiber alignment induced by cyclic stretch. PLoS One 4, e4853 (2009) Park, J. S. et al. Differential effects of equiaxial and uniaxial strain on mesenchymal stem cells. Biotechnol. Bioeng. 88, (2004) Hamilton, D. W., Maul, T. M. & Vorp, D. a. Characterization of the response of bone marrow-derived progenitor cells to cyclic strain: implications for vascular tissueengineering applications. Tissue Eng. 10, (2004) Lee, W.-C. C., Maul, T. M., Vorp, D. a, Rubin, J. P. & Marra, K. G. Effects of uniaxial cyclic strain on adipose-derived stem cell morphology, proliferation, and differentiation. Biomech. Model. Mechanobiol. 6, (2007) Chrzanowska-Wodnicka, M. & Burridge, K. Rho-stimulated Contractility Drives the Contractility Formation of Stress Fibers and Focal Adhesions. J. Cell Biol. 133, (1996) Choi, C. K. et al. Actin and α-actinin orchestrate the assembly and maturation of nascent adhesions in a myosin II motor-independent manner. Nat. Cell Biol. 10, (2008) Pavalko, F. M. et al. A model for mechanotransduction in bone cells: The load-bearing mechanosomes. J. Cell. Biochem. 88, (2003) Bidwell, J. P. & Pavalko, F. M. The load-bearing mechanosome revisited. Clin. Rev. Bone Miner. Metab. 8, (2010) Kanchanawong, P. et al. Nanoscale architecture of integrin-based cell adhesions. Nature 468, (2010) Hynes, R. O. Integrins : Bidirectional, Allosteric Signaling Machines In their roles as major adhesion receptors, integrins. Cell 110, (2002) Schwartz, M. A. Integrins and Extracellular Matrix in Mechanotransduction. Cold Spring Harb. Perspect. Biol. 2, 1 13 (2010) Chen, H., Cohen, D. M., Choudhury, D. M., Kioka, N. & Craig, S. W. Spatial distribution and functional significance of activated vinculin in living cells. J. Cell Biol. 169, (2005) Bock-marquette, I., Saxena, A., White, M. D., Dimaio, J. M. & Srivastava, D. Thymosin b 4 activates integrin-linked kinase and promotes cardiac cell migration, survival and cardiac repair. Nature 432, (2004) Traister, A. et al. ILK induces cardiomyogenesis in the human heart. PLoS One 7, e37802
146 (2012) Smits, A. M. et al. Human cardiomyocyte progenitor cells differentiate into functional mature cardiomyocytes: an in vitro model for studying human cardiac physiology and pathophysiology. Nat. Protoc. 4, (2009) Carisey, A. & Ballestrem, C. Vinculin, an adapter protein in control of cell adhesion signalling. Eur. J. Cell Biol. 90, (2011) Carisey, A. et al. Vinculin regulates the recruitment and release of core focal adhesion proteins in a force-dependent manner. Curr. Biol. 23, (2013) Wickström, S. a, Lange, A., Montanez, E. & Fässler, R. The ILK/PINCH/parvin complex: the kinase is dead, long live the pseudokinase! EMBO J. 29, (2010) Hoffman, B. D., Grashoff, C. & Schwartz, M. a. Dynamic molecular processes mediate cellular mechanotransduction. Nature 475, (2011) Zaidel-Bar, R., Ballestrem, C., Kam, Z. & Geiger, B. Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. J. Cell Sci. 116, (2003) Discher, D. E., Janmey, P. & Wang, Y.-L. Tissue cells feel and respond to the stiffness of their substrate. Science 310, (2005) Pasapera, A. M., Schneider, I. C., Rericha, E., Schlaepfer, D. D. & Waterman, C. M. Myosin II activity regulates vinculin recruitment to focal adhesions through FAK-mediated paxillin phosphorylation. J. Cell Biol. 188, (2010) Huttenlocher, A. & Horwitz, A. R. 13. Integrins in Cell Migration. Cold Spring Harb. Perspect. Biol. 3, 1 16 (2011) Engler, A. J., Sen, S., Sweeney, H. L. & Discher, D. E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 126, (2006) Yamane, M. et al. Rac1 activity is required for cardiac myocyte alignment in response to mechanical stress. Biochem. Biophys. Res. Commun. 353, (2007) Matsuda, T. et al. N-cadherin-mediated cell adhesion determines the plasticity for cell alignment in response to mechanical stretch in cultured cardiomyocytes. Biochem. Biophys. Res. Commun. 326, (2005) Kada, K. et al. Orientation change of cardiocytes induced by cyclic stretch stimulation: time dependency and involvement of protein kinases. J. Mol. Cell. Cardiol. 31, (1999) Mack, C. P., Somlyo, A. V., Hautmann, M., Somlyo, A. P. & Owens, G. K. Smooth muscle differentiation marker gene expression is regulated by Rhoa-mediated actin polymerization. J. Biol. Chem. 276, (2001) Sandbo, N. et al. Delayed stress fiber formation mediates pulmonary myofibroblast differentiation in response to TGF-. AJP Lung Cell. Mol. Physiol. 301, L656 L666 (2011) Jacot, J. G., Martin, J. C. & Hunt, D. L. Mechanobiology of cardiomyocyte development. J. 137
147 138 Biomech. 43, 93 8 (2010) Fässler, R. et al. Differentiation and integrity of cardiac muscle cells are impaired in the absence of beta 1 integrin. J. Cell Sci. 109 ( Pt 1, (1996) Trappmann, B. et al. Extracellular-matrix tethering regulates stem-cell fate. Nat. Mater. 11, (2012) Galbraith, C. G., Yamada, K. M. & Sheetz, M. P. The relationship between force and focal complex development. J. Cell Biol. 159, (2002) Riveline, D. et al. Focal contacts as mechanosensors: Externally applied local mechanical force induces growth of focal contacts by an mdia1-dependent and ROCK-independent mechanism. J. Cell Biol. 153, (2001) Balaban, N. Q. et al. Force and focal adhesion assembly : a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 3, (2001) Chen, B., Kemkemer, R., Deibler, M., Spatz, J. & Gao, H. Cyclic Stretch Induces Cell Reorientation on Substrates by Destabilizing Catch Bonds in Focal Adhesions. PLoS One 7, (2012) Steinhauser, M. L. & Lee, R. T. Regeneration of the heart. EMBO Mol. Med. 3, (2011) Jaalouk, D. E. & Lammerding, J. Mechanotransduction gone awry. Nat. Rev. Mol. Cell Biol. 10, (2009) Burridge, K., Fath, K., Kelly, T., Nuckolls, G. & Turner, C. Focal adhesions: transmembrane junctions between the extracellular matrix and the cytoskeleton. Annu. Rev. Cell Biol. 4, (1988) Bershadsky, A. D. et al. Assembly and mechanosensory function of focal adhesions: Experiments and models. Eur. J. Cell Biol. 85, (2006) Horton, E. R., Astudillo, P., Humphries, M. J. & Humphries, J. D. Mechanosensitivity of integrin adhesion complexes: Role of the consensus adhesome. Exp. Cell Res. 343, 7 13 (2016) Katoh, K., Kano, Y. & Ookawara, S. Rho-kinase dependent organization of stress fibers and focal adhesions in cultured fibroblasts. Genes to Cells 12, (2007) Livne, A., Bouchbinder, E. & Geiger, B. Cell reorientation under cyclic stretching. Nat. Commun. 5, 3938 (2014) Tamiello, C., Buskermolen, A. B. C., Baaijens, F. P. T., Broers, J. L. V. & Bouten, C. V. C. Heading in the Right Direction: Understanding Cellular Orientation Responses to Complex Biophysical Environments. Cell. Mol. Bioeng. 9, (2015) Eaton, J. W. Free Your Numbers. Distribution (2011) Andersen, J. I., Pennisi, C. P., Fink, T. & Zachar, V. Focal Adhesion Kinase Activation Is Necessary for Stretch-Induced Alignment and Enhanced Differentiation of Myogenic Precursor Cells. Tissue Eng. Part A 0, ten.tea (2017).
148 234. Kaunas, R., Nguyen, P., Usami, S. & Chien, S. Cooperative effects of Rho and mechanical stretch on stress fiber organization. 102, (2005) Schiller, H. B. & Fässler, R. Mechanosensitivity and compositional dynamics of cell-matrix adhesions. EMBO Rep. 14, (2013) Sun, Z., Guo, S. S. & Fässler, R. Integrin-mediated mechanotransduction. J. Cell Biol. (2016). doi: /jcb Stutchbury, B., Atherton, P., Tsang, R., Wang, D.-Y. & Ballestrem, C. Distinct focal adhesion protein modules control different aspects of mechanotransduction. J. Cell Sci. 130, (2017) Amano, M. et al. Formation of actin stress fibers and focal adhesions enhanced by Rhokinase. Science 275, (1997) Cambria, E. et al. Translational cardiac stem cell therapy: advancing from first-generation to next-generation cell types. npj Regen. Med. 2, 17 (2017) McCain, M. L., Lee, H., Aratyn-Schaus, Y., Kléber, A. G. & Parker, K. K. Cooperative coupling of cell-matrix and cell cell adhesions in cardiac muscle. Proc. Natl. Acad. Sci. 109, 9881 LP-9886 (2012) Ackermann, M. A. et al. TGF-β1 affects cell-cell adhesion in the heart in an NCAM1-dependent mechanism. J. Mol. Cell. Cardiol. 112, (2018) Huveneers, S. & de Rooij, J. Mechanosensitive systems at the cadherin-f-actin interface. J. Cell Sci. 126, (2013) Borghi, N. et al. E-cadherin is under constitutive actomyosin-generated tension that is increased at cell cell contacts upon externally applied stretch. Proc. Natl. Acad. Sci. 109, LP (2012) Leckband, D. E., le Duc, Q., Wang, N. & de Rooij, J. Mechanotransduction at cadherinmediated adhesions. Curr. Opin. Cell Biol. 23, (2011) Ladoux, B. et al. Strength Dependence of Cadherin-Mediated Adhesions. Biophys. J. 98, (2010) Cosgrove, B. D. et al. N-cadherin adhesive interactions modulate matrix mechanosensing and fate commitment of mesenchymal stem cells. Nat. Mater. 1, 1 19 (2016) Noorman, M. et al. Cardiac cell-cell junctions in health and disease: Electrical versus mechanical coupling. J. Mol. Cell. Cardiol. 47, (2009) Peralta Soler 1994_N-Cadherin Involvement in Cardiac Myocyte Interaction and Myofibrillogenesis.pdf Kresh, J. Y. & Chopra, A. Intercellular and extracellular mechanotransduction in cardiac myocytes. Pflugers Arch. 462, (2011) Mui, K. L., Chen, C. S. & Assoian, R. K. The mechanical regulation of integrin cadherin crosstalk organizes cells, signaling and forces. J. Cell Sci. 129, 1093 LP-1100 (2016). 139
149 251. Pellegrin, S. & Mellor, H. Actin stress fibres. J. Cell Sci. 120, (2007) Ratheesh, A. & Yap, A. S. A bigger picture: classical cadherins and the dynamic actin cytoskeleton. Nat. Rev. Mol. Cell Biol. 13, 673 (2012) Ng, M. R., Besser, A., Danuser, G. & Brugge, J. S. Substrate stiffness regulates cadherindependent collective migration through myosin-ii contractility. J. Cell Biol. 199, (2012) Nelson, W. J. & Weis, W. I. 25 Years of Tension over Actin Binding to the Cadherin Cell Adhesion Complex: The Devil is in the Details. Trends Cell Biol. 26, (2016) Yap, A. S., Gomez, G. A. & Parton, R. G. Adherens Junctions Revisualized: Organizing Cadherins as Nanoassemblies. Dev. Cell 35, (2015) Yonemura, S. Cadherin-actin interactions at adherens junctions. Curr. Opin. Cell Biol. 23, (2011) Vedula, S. R. K. et al. Emerging modes of collective cell migration induced by geometrical constraints. Proc. Natl. Acad. Sci. 109, LP (2012) Tambe, D. T. et al. Collective cell guidance by cooperative intercellular forces. Nat. Mater. 10, 469 (2011) Labernadie, A. et al. A mechanically active heterotypic E-cadherin/N-cadherin adhesion enables fibroblasts to drive cancer cell invasion. Nat. Cell Biol. 19, 224 (2017) Trepat, X. et al. Physical forces during collective cell migration. Nat. Phys. 5, 426 (2009) Etienne-Manneville, S. Neighborly relations during collective migration. Curr. Opin. Cell Biol. 30, (2014) Basan, M., Elgeti, J., Hannezo, E., Rappel, W.-J. & Levine, H. Alignment of cellular motility forces with tissue flow as a mechanism for efficient wound healing. Proc. Natl. Acad. Sci. 110, 2452 LP-2459 (2013) Desai, R. A., Gao, L., Raghavan, S., Liu, W. F. & Chen, C. S. Cell polarity triggered by cellcell adhesion via E-cadherin. J. Cell Sci. 122, (2009) Singh, S. J., Turner, W., Glaser, D. E., McCloskey, K. E. & Filipp, F. V. Metabolic shift in density-dependent stem cell differentiation. Cell Commun. Signal. 15, 1 12 (2017) Tang, J., Peng, R. & Ding, J. The regulation of stem cell differentiation by cell-cell contact on micropatterned material surfaces. Biomaterials 31, (2010) Tsai, R. Y. & McKay, R. D. Cell contact regulates fate choice by cortical stem cells. J. Neurosci. 20, (2000) Chen, T. et al. E-cadherin-mediated cell-cell contact is critical for induced pluripotent stem cell generation. Stem Cells 28, (2010) Mao, A. S., Shin, J. W. & Mooney, D. J. Effects of substrate stiffness and cell-cell contact on mesenchymal stem cell differentiation. Biomaterials 98, (2016) Lu, H. et al. Effect of cell density on adipogenic differentiation of mesenchymal stem 140
150 cells. Biochem. Biophys. Res. Commun. 381, (2009) Baker, B. M. & Chen, C. S. Deconstructing the third dimension: how 3D culture microenvironments alter cellular cues. J. Cell Sci. 125, (2012) Doyle, A. D. & Yamada, K. M. Mechanosensing via cell-matrix adhesions in 3D microenvironments. Experimental Cell Research 343, (2016) Lee, S. & Kumar, S. Actomyosin stress fiber mechanosensing in 2D and 3D. F1000Research 5, 2261 (2016) Wang, H. B., Dembo, M., Hanks, S. K. & Wang, Y. Focal adhesion kinase is involved in mechanosensing during fibroblast migration. Proc. Natl. Acad. Sci. U. S. A. 98, (2001) de Jonge, N. et al. Matrix production and organization by endothelial colony forming cells in mechanically strained engineered tissue constructs. PLoS One 8, e73161 (2013) Foolen, J., Deshpande, V. S., Kanters, F. M. W. & Baaijens, F. P. T. The influence of matrix integrity on stress-fiber remodeling in 3D. Biomaterials 33, (2012) Bono, N. et al. Unraveling the role of mechanical stimulation on smooth muscle cells: A comparative study between 2D and 3D models. Biotechnol. Bioeng. 113, (2016) NAGAYAMA, K., MORISHIMA, N. & MATSUMOTO, T. Effects of Three-Dimensional Culture and Cyclic Stretch Stimulation on Expression of Contractile Proteins in Freshly Isolated Rat Aortic Smooth Muscle Cells. J. Biomech. Sci. Eng. 4, (2009) Tondon, A. & Kaunas, R. The direction of stretch-induced cell and stress fiber orientation depends on collagen matrix stress. PLoS One 9, (2014) Vining, K. H. & Mooney, D. J. Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 18, (2017) Lv, H. et al. Mechanism of regulation of stem cell differentiation by matrix stiffness. Stem Cell Res. Ther. 6, 103 (2015) Lee, C. F., Haase, C., Deguchi, S. & Kaunas, R. Cyclic stretch-induced stress fiber dynamics - Dependence on strain rate, Rho-kinase and MLCK. Biochem. Biophys. Res. Commun. 401, (2010) Dupont, S. et al. Role of YAP/TAZ in mechanotransduction. Nature 474, (2011) Nardone, G. et al. YAP regulates cell mechanics by controlling focal adhesion assembly. Nat. Commun. 8, (2017) Piccolo, S., Dupont, S. & Cordenonsi, M. The Biology of YAP/TAZ: Hippo Signaling and Beyond. Physiol. Rev. 94, (2014) Khalafalla, F. G. et al. P2Y 2 Nucleotide Receptor Prompts Human Cardiac Progenitor Cell Activation by Modulating Hippo Signaling. Circ. Res. 2610, CIRCRESAHA (2017). 141
151 286. Dupont, S. Role of YAP/TAZ in cell-matrix adhesion-mediated signalling and mechanotransduction. Exp. Cell Res. 343, (2016) Mosqueira, D. et al. Hippo pathway effectors control cardiac progenitor cell fate by acting as dynamic sensors of substrate mechanics and nanostructure. ACS Nano 8, (2014) Duong, H., Wu, B. & Tawil, B. Modulation of 3D Fibrin Matrix Stiffness by Intrinsic Fibrinogen Thrombin Compositions and by Extrinsic Cellular Activity. Tissue Eng. Part A 15, (2009) Skardal, A., Mack, D., Atala, A. & Soker, S. Substrate elasticity controls cell proliferation, surface marker expression and motile phenotype in amniotic fluid-derived stem cells. Journal of the mechanical behavior of biomedical materials 17, (2013) Piechocka, I. K. et al. Multi-scale strain-stiffening of semiflexible bundle networks. Soft Matter 12, (2016) Boopathy, A. V. et al. The modulation of cardiac progenitor cell function by hydrogeldependent Notch1 activation. Biomaterials 35, (2014) Berry, M. F. et al. Mesenchymal stem cell injection after myocardial infarction improves myocardial compliance. Am. J. Physiol. Heart Circ. Physiol. 290, H (2006) Qiu, Y. et al. A role for matrix stiffness in the regulation of cardiac side population cell function. Am. J. Physiol. Circ. Physiol. 308, H990 H997 (2015) Lee, S. et al. Contractile force generation by 3D hipsc-derived cardiac tissues is enhanced by rapid establishment of cellular interconnection in matrix with muscle-mimicking stiffness. Biomaterials 131, (2017) Tallawi, M., Rai, R., Boccaccini, A. R. & Aifantis, K. Effect of substrate mechanics on cardiomyocyte maturation and growth. Tissue Eng. Part B. Rev. 21, (2014) Young, J. L., Kretchmer, K., Ondeck, M. G., Zambon, A. C. & Engler, A. J. Mechanosensitive kinases regulate stiffness-induced cardiomyocyte maturation. Sci. Rep. 4, 6425 (2014) Jacot, J. G., McCulloch, A. D. & Omens, J. H. Substrate stiffness affects the functional maturation of neonatal rat ventricular myocytes. Biophys. J. 95, (2008) Tomei, A. A., Boschetti, F., Gervaso, F. & Swartz, M. A. 3D collagen cultures under welldefined dynamic strain: A novel strain device with a porous elastomeric support. Biotechnol. Bioeng. 103, (2009) Henshaw, D. R., Attia, E., Bhargava, M. & Hannafin, J. A. Canine ACL fibroblast integrin expression and cell alignment in response to cyclic tensile strain in three-dimensional collagen gels. J. Orthop. Res. 24, (2006) Foolen, J., Janssen-Van Den Broek, M. W. J. T. & Baaijens, F. P. T. Synergy between Rho signaling and matrix density in cyclic stretch-induced stress fiber organization. Acta Biomater. 10, (2014) Totaro, A. et al. YAP/TAZ link cell mechanics to Notch signalling to control epidermal stem cell fate. Nat. Commun. 8, (2017). 142
152 302. Manderfield, L. J. et al. Hippo signaling is required for Notch-dependent smooth muscle differentiation of neural crest. Development 142, 2962 LP-2971 (2015) Slemmons, K. K. et al. A Novel Notch YAP Circuit Drives Stemness and Tumorigenesis in Embryonal Rhabdomyosarcoma. Mol. Cancer Res. (2017) Caiazzo, M. et al. Defined three-dimensional microenvironments boost induction of pluripotency. Nat. Mater. 15, (2016) Bollini, S., Smart, N. & Riley, P. R. Resident cardiac progenitor cells: at the heart of regeneration. J. Mol. Cell. Cardiol. 50, (2011) Messina, E. et al. Isolation and expansion of adult cardiac stem cells from human and murine heart. Circ. Res. 95, (2004) Li, T.-S. et al. Cardiospheres Recapitulate a Niche-Like Microenvironment Rich in Stemness and Cell-Matrix Interactions, Rationalizing Their Enhanced Functional Potency for Myocardial Repair. Stem Cells 28, (2010) Luxán, G., D Amato, G., MacGrogan, D. & De La Pompa, J. L. Endocardial Notch Signaling in Cardiac Development and Disease. Circ. Res. 118, e1 e18 (2016) Nemir, M. & Pedrazzini, T. Functional role of Notch signaling in the developing and postnatal heart. J. Mol. Cell. Cardiol. 45, (2008) Main, H. et al. Interactions between Notch- and hypoxia-induced transcriptomes in embryonic stem cells. Exp. Cell Res. 316, (2010) Lendahl, U., Lee, K. L., Yang, H. & Poellinger, L. Generating specificity and diversity in the transcriptional response to hypoxia. Nat. Rev. Genet. 10, (2009) Forte, E. et al. TGFβ-dependent epithelial-to-mesenchymal transition is required to generate cardiospheres from human adult heart biopsies. Stem Cells Dev. 21, (2012) Di Costanzo, E. et al. A discrete in continuous mathematical model of cardiac progenitor cells formation and growth as spheroid clusters (Cardiospheres). Math. Med. Biol. A J. IMA dqw022-dqw022 (2017) Bergmann, O. et al. Evidence for cardiomyocyte renewal in humans. Science 324, (2009) Hsieh, P. C. H. et al. Evidence from a genetic fate-mapping study that stem cells refresh adult mammalian cardiomyocytes after injury. Nat. Med. 13, (2007) Mosqueira, D. et al. Hippo Pathway E ff ectors Control Cardiac Progenitor Cell Fate by Acting as Dynamic Sensors of Substrate Mechanics and Nanostructure (2014) Bax, N. A. M. et al. Matrix production and remodeling capacity of cardiomyocyte progenitor cells during in vitro differentiation. J. Mol. Cell. Cardiol. 53, (2012) Zhao, L. et al. Notch signaling regulates cardiomyocyte proliferation during zebrafish heart regeneration. Proc. Natl. Acad. Sci. U. S. A. 111, (2014). 143
153 319. Berger, A. J., Linsmeier, K. M., Kreeger, P. K. & Masters, K. S. Decoupling the effects of stiffness and fiber density on cellular behaviors via an interpenetrating network of gelatin-methacrylate and collagen. Biomaterials 141, (2017) Christman, K. L., Fok, H. H., Sievers, R. E., Fang, Q. & Lee, R. J. Fibrin Glue Alone and Skeletal Myoblasts in a Fibrin Scaffold Preserve Cardiac Function after Myocardial Infarction. Tissue Eng. 10, (2004) Nunes, S. S. et al. Biowire: a platform for maturation of human pluripotent stem cellderived cardiomyocytes. Nat. Methods 10, (2013) Forte, G. et al. Criticality of the biological and physical stimuli array inducing resident cardiac stem cell determination. Stem Cells 26, (2008) Atmanli, A. & Domian, I. J. Recreating the Cardiac Microenvironment in Pluripotent Stem Cell Models of Human Physiology and Disease. Trends Cell Biol. 27, (2017) Mathur, A., Ma, Z., Loskill, P., Jeeawoody, S. & Healy, K. E. In vitro cardiac tissue models: Current status and future prospects. Adv. Drug Deliv. Rev. 96, (2016) Pati, F. et al. Printing three-dimensional tissue analogues with decellularized extracellular matrix bioink. Nat. Commun. 5, 3935 (2014) Gaetani, R. et al. Cardiac tissue engineering using tissue printing technology and human cardiac progenitor cells. Biomaterials 33, (2012) Renko, O. et al. SDF1 gradient associates with the distribution of c-kit+ cardiac cells in the heart. Sci. Rep. 8, 1160 (2018) van Vliet, P. et al. Foetal and adult cardiomyocyte progenitor cells have different developmental potential. J. Cell. Mol. Med. 14, (2010) van Marion, M. H. The interplay between the extracellular environment and cardiomyocyte progenitor cells for material based cardiac repair. (2013) Takahashi, K. & Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 126, (2006) Aznar, J. & Tudela, J. Ten years since the discovery of ips cells: The current state of their clinical application. Rev. Clínica Española (English Ed. 217, (2017) Li, X., Tamama, K., Xie, X. & Guan, J. Improving Cell Engraftment in Cardiac Stem Cell Therapy. Stem Cells Int. 2016, (2016) Kishore, R. & Khan, M. More Than Tiny Sacks. Circ. Res. 118, (2016) Chimenti, I. et al. Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infarcted mice. Circ. Res. 106, (2010) Vrijsen, K. R. et al. Cardiomyocyte progenitor cell-derived exosomes stimulate migration of endothelial cells. J. Cell. Mol. Med. 14, (2010) Kishore, R. & Khan, M. Cardiac cell-derived exosomes: changing face of regenerative 144
154 biology. Eur. Heart J. 1 4 (2016). doi: /eurheartj/ehw Gallet, R. et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 38, (2017) Barile, L. et al. Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovasc. Res. 103, (2014) Ibrahim, A. G.-E., Cheng, K. & Marbán, E. Exosomes as Critical Agents of Cardiac Regeneration Triggered by Cell Therapy. Stem Cell Reports 2, (2014) Adamiak, M. et al. <span hwp:id= article-title-1 class= article-title >Induced Pluripotent Stem Cell (ipsc) Derived Extracellular Vesicles Are Safer and More Effective for Cardiac Repair Than ipscs</span><span hwp:id= article-title-41 class= sub-article-title >Novelty and. Circ. Res. 122, 296 LP-309 (2018) Sluijter, J. P. G. & Van Rooij, E. Exosomal microrna clusters are important for the therapeutic effect of cardiac progenitor cells. Circ. Res. 116, (2015). 145
155 146
156 SUMMARY Cardiac progenitor cells (CPCs), undifferentiated cells that reside in the human fetal and adult heart, have emerged as a promising cell source for myocardial repair and regeneration after cardiac damage. CPCs have the potential to differentiate into all of the cell types of the heart, and to produce and remodel extracellular matrix (ECM). Earlier studies have reported activation of resident CPCs in the adult heart following acute myocardial infarction. In addition, CPC therapy, i.e. CPC transplantation into the infarcted heart, has shown beneficial effects on cardiac function. Yet, the actual contribution of CPCs to cardiac repair and regeneration remains unclear, and the functional benefits are limited to the short-term. The latter issue is mainly due to massive cell loss and reduced survival and differentiation of the transplanted cells in the hostile environment of the damaged myocardium. To exploit the regenerative potential held by CPCs, and to improve current therapeutic strategies of CPC-mediated cardiac regeneration, a better understanding of the interactions of CPCs with their microenvironment (the CPC niche ) is needed. Microenvironmental factors that can influence CPC behavior include: cyclic cell straining (due to cardiac beating), the ECM, neighboring cells, and oxygen tension. The aim of this thesis was to systematically investigate and better understand the impact of these environmental conditions, either separate or combined, on CPC function and differentiation by using 2D and 3D in vitro model systems. First, we analyzed the response to uniaxial cyclic strain (10%, 0.5Hz) of undifferentiated CPCs and cells at an early stage of differentiation (predifferentiated CPCs) on flexible 2D substrates. When subjected to cyclic stretching, CPCs were able to reorient perpendicular to the main stretching direction (strain avoidance) only upon cardiomyogenic differentiation. In fact, after 48 hrs of uniaxial cyclic strain undifferentiated CPCs did not acquire a preferential orientation, whereas predifferentiated CPCs reoriented perpendicular to the strain direction. The results showed that CPCs develop a so-called mechanosome, i.e. the assembly of molecular structures responsible for cell-ecm adhesions and signaling, upon CPC differentiation. The role of the mechanosome in CPC strain avoidance was further investigated by blocking selected mechanosome components of predifferentiated CPCs (either integrinβ1, integrin-linked kinase (ILK), or Rho-kinase (ROCK)). The strain avoidance response was reduced by inhibition of each of these mechanosome components, indicating that the mechanosome integrity is required for CPC strain avoidance. In a second study, the response of CPCs to cyclic strain was shown to not only rely on cell-ecm adhesions, but also on cell density. With increasing cell density, undifferentiated CPCs displayed enhanced strain avoidance response in 2D, as well as increased N-cadherin clustering. Inhibition of N-cadherin-mediated cell-cell adhesions, as well as ROCK, resulted in reduced, though not completely absent, strain avoidance behavior of undifferentiated CPCs at high cell density. This suggests that both cell-cell adhesions and ROCK activity contribute to CPC mechanoresponse. 147
157 To better resemble the in vivo CPC environment, a 3D tissue model consisting of CPC-seeded fibrin gel was developed. Culture in the relatively soft fibrin environment (E=1 kpa) resulted in a shift in CPC fate toward the vascular lineage, as compared to culture of CPCs on 2D and stiffer substrates (E=1 MPa). Application of uniaxial cyclic strain (10%, 0.5Hz, 48 hrs) to the 3D fibrin model caused CPCs to orient in alignment with the strain direction, as opposed to the strain avoidance seen on 2D substrates. This strain alignment response was not affected by preventing stress fiber formation via inhibition of ROCK or myosin light chain kinase (MLCK), suggesting that the observed cell reorientation is independent of stress fiber development. Further experiments are required to conclude whether the observed differences are due to the environmental stiffness or dimensionality (cell configurations in 2D or 3D). In support of the notion that the substrate stiffness may influence the cellular response to cyclic strain, CPCs in 2D and 3D environments showed differential activation of YAP, one of the main sensors of the cell mechanical environment and regulator of cell fate determination. On the stiffer 2D substrate, YAP was distributed mainly in the cell nucleus, showing activation of the pathway. However, in the softer 3D environment of the fibrin hydrogel, YAP was distributed evenly in the cell nucleus and cytoplasm, indicating lower YAP activity and hence reduced cell mechanosensing. Altogether, these findings highlight the fundamental role of a more native 3D environment on CPC mechanobiology. The relevance of niche-like microenvironmental conditions was further demonstrated by culturing CPCs as spheroids on poly-lysine coated plates, under normoxic (20% O2) and hypoxic (5% O2) conditions. Multicellular spheroids are believed to provide niche-like environments, with improved cell-cell and cell-ecm contacts, as well as gradients of oxygen and nutrients. Increased expression and activation of Notch signaling were observed in CPC spheroids, as compared to CPCs grown as monolayer. Since Notch activity is known to regulate CPC proliferation and differentiation, these results suggest that a niche-like microenvironment promotes maintenance of the CPC pool and CPC multipotency. In conclusion, the results of this thesis showed that there is a clear and reciprocal interplay between CPCs and their microenvironment, mediated by cell-ecm and cell-cell adhesions. It was demonstrated that CPC differentiation state determines the ability to sense external mechanical stimuli. In turn, microenvironmental stimuli (mechanical and cellular signals) conversely influence CPC fate decisions. Furthermore, the presented results highlight the relevance of studying cell behavior in a 3D environment, as opposed to the traditional 2D in vitro culture. The improved knowledge on CPC-niche interactions provided by these findings can be used to re-evaluate and improve the current application of CPCs in cardiac therapy. 148
, she studied Materials Science at the University of Rome Tor Vergata.")
158 CURRICULUM VITAE Arianna Mauretti was born on December 15, 1986 in Rome, Italy. After completing her secondary education at Liceo Scientifico B. Croce in Rome (Italy), she studied Materials Science at the University of Rome Tor Vergata. She obtained her Master s degree in 2012, with a project on three-dimensional biomaterials for tissue engineering, performed at Technion Israel Institute of Technology in Haifa, Israel. In September 2013 she started her PhD in the group of Soft Tissue Mechanobiology and Engineering in the Department of Biomedical Engineering of Eindhoven University of Technology, investigating the interplay between human cardiac progenitor cells and their microenvironment. During her PhD, she carried on a collaboration with the Department of Molecular Medicine at University of Rome La Sapienza. The results of her research are presented in this thesis. 149
UNIT CELL PROCESSES UNDERLYING TISSUE ENGINEERING AND REGENERATIVE MEDICINE
 Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/20.441/HST522J UNIT CELL PROCESSES UNDERLYING TISSUE ENGINEERING AND REGENERATIVE
Massachusetts Institute of Technology Harvard Medical School Brigham and Women s Hospital VA Boston Healthcare System 2.79J/3.96J/20.441/HST522J UNIT CELL PROCESSES UNDERLYING TISSUE ENGINEERING AND REGENERATIVE
Contents. The Right Surface for Every Cell Extracellular Matrices and Biologically Coated Surfaces ECM Mimetic and Advanced Surfaces...
 Contents The Right Surface for Every Cell... 1 Extracellular Matrices and Biologically Coated Surfaces... 2 Corning Matrigel Matrix... 2 Corning BioCoat Cultureware... 3 ECM Mimetic and Advanced Surfaces...
Contents The Right Surface for Every Cell... 1 Extracellular Matrices and Biologically Coated Surfaces... 2 Corning Matrigel Matrix... 2 Corning BioCoat Cultureware... 3 ECM Mimetic and Advanced Surfaces...
Regenerative medicine approaches to the treatment of cardiovascular disease
 Regenerative medicine approaches to the treatment of cardiovascular disease Author: Noah Weisleder, Ph.D. Abstract: Ischemic heart disease related myocardial infarction (MI) remains the leading cause of
Regenerative medicine approaches to the treatment of cardiovascular disease Author: Noah Weisleder, Ph.D. Abstract: Ischemic heart disease related myocardial infarction (MI) remains the leading cause of
Lab Module 7: Cell Adhesion
 Lab Module 7: Cell Adhesion Tissues are made of cells and materials secreted by cells that occupy the spaces between the individual cells. This material outside of cells is called the Extracellular Matrix
Lab Module 7: Cell Adhesion Tissues are made of cells and materials secreted by cells that occupy the spaces between the individual cells. This material outside of cells is called the Extracellular Matrix
NEXT GENERATION ECM-BASED ALLOGRAFT TECHNOLOGY:
 NEXT GENERATION ECM-BASED ALLOGRAFT TECHNOLOGY: Potent biological scaffolds strategically control stem cell fate and function, allowing our allografts to harness the regenerative potential of patient s
NEXT GENERATION ECM-BASED ALLOGRAFT TECHNOLOGY: Potent biological scaffolds strategically control stem cell fate and function, allowing our allografts to harness the regenerative potential of patient s
At the conclusion of this lesson you should be able to:
 Learning Objectives At the conclusion of this lesson you should be able to: Understand the key terms and definitions regarding stem cells Differentiate between the adult and embryonic stem cells Differentiate
Learning Objectives At the conclusion of this lesson you should be able to: Understand the key terms and definitions regarding stem cells Differentiate between the adult and embryonic stem cells Differentiate
Bioreactors in tissue engineering
 Bioreactors in tissue engineering S. Swaminathan Director Centre for Nanotechnology & Advanced Biomaterials School of Chemical & Biotechnology SASTRA University Thanjavur 613 401 Tamil Nadu Joint Initiative
Bioreactors in tissue engineering S. Swaminathan Director Centre for Nanotechnology & Advanced Biomaterials School of Chemical & Biotechnology SASTRA University Thanjavur 613 401 Tamil Nadu Joint Initiative
Microenvironmental cues to influence endothelial colony forming cell fate : relevance for in situ vascular tissue engineering Fioretta, E.S.
 Microenvironmental cues to influence endothelial colony forming cell fate : relevance for in situ vascular tissue engineering Fioretta, E.S. DOI: 10.6100/IR762770 Published: 01/01/2014 Document Version
Microenvironmental cues to influence endothelial colony forming cell fate : relevance for in situ vascular tissue engineering Fioretta, E.S. DOI: 10.6100/IR762770 Published: 01/01/2014 Document Version
Exogenous and endogenous gene regulation for specific and efficient cancer gene therapy Gommans, Willemijn Maria
 University of Groningen Exogenous and endogenous gene regulation for specific and efficient cancer gene therapy Gommans, Willemijn Maria IMPORTANT NOTE: You are advised to consult the publisher's version
University of Groningen Exogenous and endogenous gene regulation for specific and efficient cancer gene therapy Gommans, Willemijn Maria IMPORTANT NOTE: You are advised to consult the publisher's version
1 Scaffolds. 2 Extracellular matrix
 This lecture describes the components of the extracellular matrix and their influence on the cell properties and functions. 1 Scaffolds One of the key components of the tissue engineering triad is the
This lecture describes the components of the extracellular matrix and their influence on the cell properties and functions. 1 Scaffolds One of the key components of the tissue engineering triad is the
Introduction to Cell- Biomaterial Engineering!
 Introduction to Cell- Biomaterial Engineering! Module 3, Lecture 1! 20.109 Spring 2010! Topics for Lecture 1!! Introduction to tissue engineerin! motivation"! basic principles + examples"! Introduction
Introduction to Cell- Biomaterial Engineering! Module 3, Lecture 1! 20.109 Spring 2010! Topics for Lecture 1!! Introduction to tissue engineerin! motivation"! basic principles + examples"! Introduction
A FUNCTIONAL GENOMICS STUDY OF EXTRACELLULAR PROTEASE PRODUCTION BY ASPERGILLUS NIGER
 A FUNCTIONAL GENOMICS STUDY OF EXTRACELLULAR PROTEASE PRODUCTION BY ASPERGILLUS NIGER Machtelt Braaksma A functional genomics study of extracellular protease production by Aspergillus niger Proefschrift
A FUNCTIONAL GENOMICS STUDY OF EXTRACELLULAR PROTEASE PRODUCTION BY ASPERGILLUS NIGER Machtelt Braaksma A functional genomics study of extracellular protease production by Aspergillus niger Proefschrift
Des cellules-souches dans le poumon : pourquoi faire?
 Des cellules-souches dans le poumon : pourquoi faire? Karl-Heinz Krause Dept. of Pathology and Immunology, Medical Faculty Dept. of Genetic and Laboratory Medicine, University Hospitals Geneva, Switzerland
Des cellules-souches dans le poumon : pourquoi faire? Karl-Heinz Krause Dept. of Pathology and Immunology, Medical Faculty Dept. of Genetic and Laboratory Medicine, University Hospitals Geneva, Switzerland
Fundamental properties of Stem Cells
 Stem cells Learning Goals: Define what a stem cell is and describe its general properties, using hematopoietic stem cells as an example. Describe to a non-scientist the current progress of human stem cell
Stem cells Learning Goals: Define what a stem cell is and describe its general properties, using hematopoietic stem cells as an example. Describe to a non-scientist the current progress of human stem cell
The Effects of Scaffold Rigidity on Retinal Pigment Epithelial Cells. Corina White Symposium on Biomaterials Science 24 October 2016
 The Effects of Scaffold Rigidity on Retinal Pigment Epithelial Cells Corina White Symposium on Biomaterials Science 24 October 2016 Background Physiology The retina is the light-responsive tissue layer
The Effects of Scaffold Rigidity on Retinal Pigment Epithelial Cells Corina White Symposium on Biomaterials Science 24 October 2016 Background Physiology The retina is the light-responsive tissue layer
Cell culture and drug treatment. Lineage - Sca-1+ CD31+ EPCs were cultured on
 Supplemental Material Detailed Methods Cell culture and drug treatment. Lineage - Sca-1+ CD31+ EPCs were cultured on 5µg/mL human fibronectin coated plates in DMEM supplemented with 10% FBS and penicillin/streptomycin
Supplemental Material Detailed Methods Cell culture and drug treatment. Lineage - Sca-1+ CD31+ EPCs were cultured on 5µg/mL human fibronectin coated plates in DMEM supplemented with 10% FBS and penicillin/streptomycin
7.06 Cell Biology EXAM #2 March 20, 2003
 7.06 Cell Biology EXAM #2 March 20, 2003 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #2 March 20, 2003 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
ACCEPTED MANUSCRIPT. In situ heart valve tissue engineering: Employing the innate immune response to do the
 Accepted Manuscript In situ heart valve tissue engineering: Employing the innate immune response to do the hard work F. Zafar, MD, D.L.S. Morales, MD PII: S0022-5223(18)30839-0 DOI: 10.1016/j.jtcvs.2018.03.059
Accepted Manuscript In situ heart valve tissue engineering: Employing the innate immune response to do the hard work F. Zafar, MD, D.L.S. Morales, MD PII: S0022-5223(18)30839-0 DOI: 10.1016/j.jtcvs.2018.03.059
ANAT Cell Biology Lecture 11 School of Medical Sciences The University of New South Wales. UNSW Copyright Notice
 ANAT3231 - Cell Biology Lecture 11 School of Medical Sciences The University of New South Wales The actin cytoskeleton Prof Peter Gunning Oncology Research Unit Room 502A Wallace Wurth Building Email:
ANAT3231 - Cell Biology Lecture 11 School of Medical Sciences The University of New South Wales The actin cytoskeleton Prof Peter Gunning Oncology Research Unit Room 502A Wallace Wurth Building Email:
Corning Microplates for Microscopy and High Content Imaging. Improve results with microplates for high resolution cell imaging
 Corning Microplates for Microscopy and High Content Imaging Improve results with microplates for high resolution cell imaging High Performance for Cell-based Assays Within the drug discovery process, high
Corning Microplates for Microscopy and High Content Imaging Improve results with microplates for high resolution cell imaging High Performance for Cell-based Assays Within the drug discovery process, high
Solution Key Problem Set
 Solution Key- 7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as
Solution Key- 7.013 Problem Set 5-2013 Question 1 During a summer hike you suddenly spot a huge grizzly bear. This emergency situation triggers a fight or flight response through a signaling pathway as
Cytomics in Action: Cytokine Network Cytometry
 Cytomics in Action: Cytokine Network Cytometry Jonni S. Moore, Ph.D. Director, Clinical and Research Flow Cytometry and PathBioResource Associate Professor of Pathology & Laboratory Medicine University
Cytomics in Action: Cytokine Network Cytometry Jonni S. Moore, Ph.D. Director, Clinical and Research Flow Cytometry and PathBioResource Associate Professor of Pathology & Laboratory Medicine University
Strategies for Assessment of Immunotoxicology in Preclinical Drug Development
 Strategies for Assessment of Immunotoxicology in Preclinical Drug Development Rebecca Brunette, PhD Scientist, Analytical Biology SNBL USA Preclinical Immunotoxicology The study of evaluating adverse effects
Strategies for Assessment of Immunotoxicology in Preclinical Drug Development Rebecca Brunette, PhD Scientist, Analytical Biology SNBL USA Preclinical Immunotoxicology The study of evaluating adverse effects
Neural Induction. Chapter One
 Neural Induction Chapter One Fertilization Development of the Nervous System Cleavage (Blastula, Gastrula) Neuronal Induction- Neuroblast Formation Cell Migration Mesodermal Induction Lateral Inhibition
Neural Induction Chapter One Fertilization Development of the Nervous System Cleavage (Blastula, Gastrula) Neuronal Induction- Neuroblast Formation Cell Migration Mesodermal Induction Lateral Inhibition
Citation for published version (APA): Lubbe, A. S. (2017). Molecular motors in new media [Groningen]: Rijksuniversiteit Groningen
![Citation for published version (APA): Lubbe, A. S. (2017). Molecular motors in new media [Groningen]: Rijksuniversiteit Groningen](/thumbs/73/69212531.jpg "Citation for published version (APA): Lubbe, A. S. (2017). Molecular motors in new media [Groningen]: Rijksuniversiteit Groningen") University of Groningen Molecular motors in new media Lubbe, Anouk Sophia IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check
University of Groningen Molecular motors in new media Lubbe, Anouk Sophia IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check
Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section B and ONE question from Section C.
 UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2013-2014 CELL BIOLOGY BIO-2B06 Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in
UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2013-2014 CELL BIOLOGY BIO-2B06 Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in
1. Introduction. 1 Page 1
 1. INTRODUCTION Osteoblast differentiation is the key component of the bone formation, growth and remodeling. During this incompletely understood process a subpopulation of multipotential mesenchymal stem
1. INTRODUCTION Osteoblast differentiation is the key component of the bone formation, growth and remodeling. During this incompletely understood process a subpopulation of multipotential mesenchymal stem
In Vitro Angiogenesis Assay Kit
 In Vitro Angiogenesis Assay Kit Catalog Number KA1323 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
In Vitro Angiogenesis Assay Kit Catalog Number KA1323 100 assays Version: 02 Intended for research use only www.abnova.com Table of Contents Introduction... 3 Background... 3 Principle of the Assay...
Towards definition of an ECM parts list: An advance on GO categories
 Towards definition of an ECM parts list: An advance on GO categories The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Towards definition of an ECM parts list: An advance on GO categories The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Characteristics. Capable of dividing and renewing themselves for long periods of time (proliferation and renewal)
") STEM CELLS (p 2-8) Overview The body is made up of about 200 different kinds of specialised cells, such as muscle cells, nerve cells, fat cells and skin cells. Cells with the capacity to give rise to the
STEM CELLS (p 2-8) Overview The body is made up of about 200 different kinds of specialised cells, such as muscle cells, nerve cells, fat cells and skin cells. Cells with the capacity to give rise to the
Adipose rabbit mesenchymal stem cells for the treatment of the chronic scar tissue of the vocal cords
 Adipose rabbit mesenchymal stem cells for the treatment of the chronic scar tissue of the vocal cords Dr. Vasiliki E Kalodimou, Head of Flow Cytometry-Research and Regenerative Medicine Department, IASO-Maternity
Adipose rabbit mesenchymal stem cells for the treatment of the chronic scar tissue of the vocal cords Dr. Vasiliki E Kalodimou, Head of Flow Cytometry-Research and Regenerative Medicine Department, IASO-Maternity
1.1 WHAT MAKES POLYMERS SO INTERESTING?
 1 INTRODUCTION Materials that can be applied in bionanotechnology and biomedicine are a subject of current research. Bio or multifunctional polymeric materials might help solving many of today s medical
1 INTRODUCTION Materials that can be applied in bionanotechnology and biomedicine are a subject of current research. Bio or multifunctional polymeric materials might help solving many of today s medical
Resource-aware business process management : analysis and support Nakatumba, J.
 Resource-aware business process management : analysis and support Nakatumba, J. DOI: 10.6100/IR760115 Published: 01/01/2013 Document Version Publisher s PDF, also known as Version of Record (includes final
Resource-aware business process management : analysis and support Nakatumba, J. DOI: 10.6100/IR760115 Published: 01/01/2013 Document Version Publisher s PDF, also known as Version of Record (includes final
1 Name. 1. (3 pts) What is apoptosis and how does it differ from necrosis? Which is more likely to trigger inflammation?
 What is apoptosis and how does it differ from necrosis? Which is more likely to trigger inflammation?") 1 Name MCB 150 Midterm Eam #1 (100 points total) Please write your full name on each page of the eam!! The eam consists of 17 questions (6 pages). Each has a different point count as indicated. Please
1 Name MCB 150 Midterm Eam #1 (100 points total) Please write your full name on each page of the eam!! The eam consists of 17 questions (6 pages). Each has a different point count as indicated. Please
PTH Effects Bone Healing and Vasculogenesis in Calvaria Bone Allograft Model
 PTH Effects Bone Healing and Vasculogenesis in Calvaria Bone Allograft Model Dmitriy Sheyn, PhD 1, Doron Cohn-Yakubovich, BSc 2, Ilan Kallai, MSc 2, Susan Su, MD 1, Xiaoyu Da, MSc 1, Gadi Pelled, DMD,
PTH Effects Bone Healing and Vasculogenesis in Calvaria Bone Allograft Model Dmitriy Sheyn, PhD 1, Doron Cohn-Yakubovich, BSc 2, Ilan Kallai, MSc 2, Susan Su, MD 1, Xiaoyu Da, MSc 1, Gadi Pelled, DMD,
Life Sciences. Novel ECM Mimetic Surfaces for Cell Expansion and Differentiation
 Life Sciences Novel ECM Mimetic Surfaces for Cell Expansion and Differentiation ASCB December 17, 2012 Marshall Kosovsky, Ph.D., Technical Support Manager Topics for Discussion Introduction Overview of
Life Sciences Novel ECM Mimetic Surfaces for Cell Expansion and Differentiation ASCB December 17, 2012 Marshall Kosovsky, Ph.D., Technical Support Manager Topics for Discussion Introduction Overview of
PRIME-XV Cell Therapy Products by
 PRIME-XV Cell Therapy Products by Product List 2017 TRINOVA BIOCHEM now distributes the PRIME-XV Cell Therapy line by Irvine Scientific in Germany, Austria and Switzerland. Irvine Scientific is a worldwide
PRIME-XV Cell Therapy Products by Product List 2017 TRINOVA BIOCHEM now distributes the PRIME-XV Cell Therapy line by Irvine Scientific in Germany, Austria and Switzerland. Irvine Scientific is a worldwide
Molecular Basis of Mechanotransduction in Endothelial Cells
 Molecular Basis of Mechanotransduction in Endothelial Cells Shu Chien, M.D., Ph.D. Departments of Bioengineering and Medicine, and The Whitaker Institute of Biomedical Engineering University of California
Molecular Basis of Mechanotransduction in Endothelial Cells Shu Chien, M.D., Ph.D. Departments of Bioengineering and Medicine, and The Whitaker Institute of Biomedical Engineering University of California
White Paper Authentic Recombinant Activin A and BMP- 4 Production Using HumanZyme s Proprietary Human Cell Line Expression Technology
 White Paper Authentic Recombinant Activin A and BMP- 4 Production Using HumanZyme s Proprietary Human Cell Line Expression Technology Mark Azam, Ph.D. VP - R&D and Manufacturing Patricia Ahrweiler, Ph.D.
White Paper Authentic Recombinant Activin A and BMP- 4 Production Using HumanZyme s Proprietary Human Cell Line Expression Technology Mark Azam, Ph.D. VP - R&D and Manufacturing Patricia Ahrweiler, Ph.D.
Tissue Engineered Medical Products
 WORKSHOP 8 Tissue Organizer: Jeremy J. Rawlinson PhD Speakers: Jack E. Lemons, PhD Lawrence J. Bonassar, PhD Mauro R. Alini, PhD Michael J. Yaszemski, MD, PhD Standards for Tissue J. Lemons, University
WORKSHOP 8 Tissue Organizer: Jeremy J. Rawlinson PhD Speakers: Jack E. Lemons, PhD Lawrence J. Bonassar, PhD Mauro R. Alini, PhD Michael J. Yaszemski, MD, PhD Standards for Tissue J. Lemons, University
Human Connective Tissue Growth Factor (CTGF) Elisa Kit
 Elisa Kit") Human Connective Tissue Growth Factor (CTGF) Elisa Kit Catalog No. CSB-E07875h (96 tests) This immunoassay kit allows for the in vitro quantitative determination of human CTGF concentrations in serum,
Human Connective Tissue Growth Factor (CTGF) Elisa Kit Catalog No. CSB-E07875h (96 tests) This immunoassay kit allows for the in vitro quantitative determination of human CTGF concentrations in serum,
leading the way in research & development
 leading the way in research & development people. passion. possibilities. ABBVIE 2 immunology AbbVie Immunology has a demonstrated record of success in identifying and developing both small molecule and
leading the way in research & development people. passion. possibilities. ABBVIE 2 immunology AbbVie Immunology has a demonstrated record of success in identifying and developing both small molecule and
INVESTIGATION OF MSC DIFFERENTIATION ON ELECTROSPUN NANOFIBROUS SCAFFOLDS
 With support of NSF Award no. EEC-0754741 INVESTIGATION OF MSC DIFFERENTIATION ON ELECTROSPUN NANOFIBROUS SCAFFOLDS NSF Summer Undergraduate Fellowship in Sensor Technologies Emily Wible (Bioengineering)
With support of NSF Award no. EEC-0754741 INVESTIGATION OF MSC DIFFERENTIATION ON ELECTROSPUN NANOFIBROUS SCAFFOLDS NSF Summer Undergraduate Fellowship in Sensor Technologies Emily Wible (Bioengineering)
ANAT 3231 Cell Biology Lecture 21 Stem Cells
 ANAT 3231 Cell Biology Lecture 21 Stem Cells Outline What are Stem Cells? Totipotency - Pluripotency - Multipotency What are different sources of Stem Cells? Embryonic vs Adult Pros and Cons for each type
ANAT 3231 Cell Biology Lecture 21 Stem Cells Outline What are Stem Cells? Totipotency - Pluripotency - Multipotency What are different sources of Stem Cells? Embryonic vs Adult Pros and Cons for each type
WOUND HEALING ON ARTIFICIAL EXTRACELLULAR MATRIX PROTEINS
 WOUND HEALING ON ARTIFICIAL EXTRACELLULAR MATRIX PROTEINS Thesis by Eileen Fong In Partial Fulfillment of the Requirements For the Degree of Doctor of Philosophy California Institute of Technology Pasadena,
WOUND HEALING ON ARTIFICIAL EXTRACELLULAR MATRIX PROTEINS Thesis by Eileen Fong In Partial Fulfillment of the Requirements For the Degree of Doctor of Philosophy California Institute of Technology Pasadena,
Facts and theories of organ regeneration in adults
 Harvard-MIT Division of Health Sciences and Technology HST.535: Principles and Practice of Tissue Engineering Instructor: I. V. Yannas Facts and theories of organ regeneration in adults I.V.Yannas, PhD
Harvard-MIT Division of Health Sciences and Technology HST.535: Principles and Practice of Tissue Engineering Instructor: I. V. Yannas Facts and theories of organ regeneration in adults I.V.Yannas, PhD
The Two-Hybrid System
 Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine The Two-Hybrid System Carolina Vollert & Peter Uetz Institut für Genetik Forschungszentrum Karlsruhe PO Box 3640 D-76021 Karlsruhe
Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine The Two-Hybrid System Carolina Vollert & Peter Uetz Institut für Genetik Forschungszentrum Karlsruhe PO Box 3640 D-76021 Karlsruhe
3D Cell Culture Product Intro. Bio-Byblos Biomedical
 3D Cell Culture Product Intro Bio-Byblos Biomedical Rundown Product Intro Cellusponge Series Go Matrix Applications Upcoming Products Degradable series Cellusponge CB series Cell Alignment plate Vivoalign
3D Cell Culture Product Intro Bio-Byblos Biomedical Rundown Product Intro Cellusponge Series Go Matrix Applications Upcoming Products Degradable series Cellusponge CB series Cell Alignment plate Vivoalign
Patient Handbook on Stem Cell Therapies
 Patient Handbook on Stem Cell Therapies WWW.ISSCR.ORG WWW.CLOSERLOOKATSTEMCELLS.ORG Patient Handbook on Stem Cell Therapies Introduction We have all heard about the extraordinary promise that stem cell
Patient Handbook on Stem Cell Therapies WWW.ISSCR.ORG WWW.CLOSERLOOKATSTEMCELLS.ORG Patient Handbook on Stem Cell Therapies Introduction We have all heard about the extraordinary promise that stem cell
Modeling Cardiac Hypertrophy: Endothelin-1 Induction with qrt-pcr Analysis
 icell Cardiomyocytes Application Protocol Modeling Cardiac Hypertrophy: Endothelin-1 Induction with qrt-pcr Analysis Introduction Cardiac hypertrophy is characterized by several different cellular changes,
icell Cardiomyocytes Application Protocol Modeling Cardiac Hypertrophy: Endothelin-1 Induction with qrt-pcr Analysis Introduction Cardiac hypertrophy is characterized by several different cellular changes,
SureSilencing sirna Array Technology Overview
 SureSilencing sirna Array Technology Overview Pathway-Focused sirna-based RNA Interference Topics to be Covered Who is SuperArray? Brief Introduction to RNA Interference Challenges Facing RNA Interference
SureSilencing sirna Array Technology Overview Pathway-Focused sirna-based RNA Interference Topics to be Covered Who is SuperArray? Brief Introduction to RNA Interference Challenges Facing RNA Interference
Supplemental Information Inventory
 Cell Stem Cell, Volume 6 Supplemental Information Distinct Hematopoietic Stem Cell Subtypes Are Differentially Regulated by TGF-β1 Grant A. Challen, Nathan C. Boles, Stuart M. Chambers, and Margaret A.
Cell Stem Cell, Volume 6 Supplemental Information Distinct Hematopoietic Stem Cell Subtypes Are Differentially Regulated by TGF-β1 Grant A. Challen, Nathan C. Boles, Stuart M. Chambers, and Margaret A.
Radius 24-Well Cell Migration Assay (Fibronectin Coated)
") Product Manual Radius 24-Well Cell Migration Assay (Fibronectin Coated) Catalog Number CBA-125-FN 24 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Cell migration is a highly
Product Manual Radius 24-Well Cell Migration Assay (Fibronectin Coated) Catalog Number CBA-125-FN 24 assays FOR RESEARCH USE ONLY Not for use in diagnostic procedures Introduction Cell migration is a highly
BD PureCoat ECM Mimetic Cultureware. Natural Performance. Scientifically Defined.
 BD PureCoat ECM Mimetic Cultureware Natural Performance. Scientifically Defined. The First Animal-Free Synthetic Surface To Mimic Natural ECM Attachment & Functionality. Next generation BD PureCoat ECM
BD PureCoat ECM Mimetic Cultureware Natural Performance. Scientifically Defined. The First Animal-Free Synthetic Surface To Mimic Natural ECM Attachment & Functionality. Next generation BD PureCoat ECM
The Road to Functional Bioanalysis: Development and Validation of a Cell-Based Assay for Neutralizing Anti-Drug Antibody Analysis
 The Road to Functional Bioanalysis: Development and Validation of a Cell-Based Assay for Neutralizing Anti-Drug Antibody Analysis Christelle Pythoud, PhD 2 nd EBF YSS, Barcelona Spain Celerion Switzerland
The Road to Functional Bioanalysis: Development and Validation of a Cell-Based Assay for Neutralizing Anti-Drug Antibody Analysis Christelle Pythoud, PhD 2 nd EBF YSS, Barcelona Spain Celerion Switzerland
Int. J. Mol. Sci. 2016, 17, 1259; doi: /ijms
 S1 of S5 Supplementary Materials: Fibroblast-Derived Extracellular Matrix Induces Chondrogenic Differentiation in Human Adipose-Derived Mesenchymal Stromal/Stem Cells in Vitro Kevin Dzobo, Taegyn Turnley,
S1 of S5 Supplementary Materials: Fibroblast-Derived Extracellular Matrix Induces Chondrogenic Differentiation in Human Adipose-Derived Mesenchymal Stromal/Stem Cells in Vitro Kevin Dzobo, Taegyn Turnley,
Development of non-substantially manipulated cell-based ATMPs 1 : flexibility introduced via the application of the risk-based approach
 3 July 2017 EMA/CAT/216556/2017 Inspections, Human Medicines, Pharmacovigilance and Committees Division Development of non-substantially manipulated cell-based ATMPs 1 : flexibility introduced via the
3 July 2017 EMA/CAT/216556/2017 Inspections, Human Medicines, Pharmacovigilance and Committees Division Development of non-substantially manipulated cell-based ATMPs 1 : flexibility introduced via the
Biomarkers, Early Prediction of Vaccine Efficacy and Safety
 Biomarkers, Early Prediction of Vaccine Efficacy and Safety Giuseppe Pantaleo, M.D. Professor of Medicine Head, Division of Immunology and Allergy Executive Director, Swiss Vaccine Research Institute Lausanne
Biomarkers, Early Prediction of Vaccine Efficacy and Safety Giuseppe Pantaleo, M.D. Professor of Medicine Head, Division of Immunology and Allergy Executive Director, Swiss Vaccine Research Institute Lausanne
Basic Antibody Structure. Multiple myeloma = cancerous plasma cells Monomer = 150,000. Chapter 4. Immunoglobulin Structure and Function
 Chapter 4. Immunoglobulin Structure and Function. Functional Regions. Types of chains. Constant & Variable regions 4. Glycoprotein * * * Heavy chain= 446 aa Light chain= 4aa Each heavy and light chain
Chapter 4. Immunoglobulin Structure and Function. Functional Regions. Types of chains. Constant & Variable regions 4. Glycoprotein * * * Heavy chain= 446 aa Light chain= 4aa Each heavy and light chain
Determining the Predictive Validity of 3D Models & Achieving Standardization for Preclinical Efficacy Testing
 Determining the Predictive Validity of 3D Models & Achieving Standardization for Preclinical Efficacy Testing August 28 2017 Jason Ekert, Head of Complex In Vitro Models Platform Technology and Sciences
Determining the Predictive Validity of 3D Models & Achieving Standardization for Preclinical Efficacy Testing August 28 2017 Jason Ekert, Head of Complex In Vitro Models Platform Technology and Sciences
Agilent Seahorse XF Live-Cell Metabolism Solutions for STEM CELL RESEARCH
 Agilent Seahorse XF Live-Cell Metabolism Solutions for STEM CELL RESEARCH MONITOR CELL FATE TRANSITIONS Improve Differentiation and Reprogramming Outcomes Adult Fibroblast Cells Reprogramming (ips Cells)
Agilent Seahorse XF Live-Cell Metabolism Solutions for STEM CELL RESEARCH MONITOR CELL FATE TRANSITIONS Improve Differentiation and Reprogramming Outcomes Adult Fibroblast Cells Reprogramming (ips Cells)
Quantification of Cell Migration and Invasion Using the IncuCyte Chemotaxis Assay
 APPLICATION NOTE IncuCyte ZOOM Live-Cell Imaging System Quantification of Cell Migration and Invasion Using the IncuCyte Chemotaxis Assay Lindy O Clair, Meagan Roddy, Maria Tikhonenko, Clare Syzbut, Nicola
APPLICATION NOTE IncuCyte ZOOM Live-Cell Imaging System Quantification of Cell Migration and Invasion Using the IncuCyte Chemotaxis Assay Lindy O Clair, Meagan Roddy, Maria Tikhonenko, Clare Syzbut, Nicola
NEURONAL CELL CULTURE MATRIX FOR BETTER MAINTENANCE AND SURVIVAL OF NEURONAL CELL CULTURES IN TISSUE CULTURE.
 NEURONAL CELL CULTURE MATRIX FOR BETTER MAINTENANCE AND SURVIVAL OF NEURONAL CELL CULTURES IN TISSUE CULTURE. D. R. Aguirre, N. DiMassa, Chrystal Johnson, H. Eran, R. Perez, C.V.R. Sharma, M.V.R. Sharma,
NEURONAL CELL CULTURE MATRIX FOR BETTER MAINTENANCE AND SURVIVAL OF NEURONAL CELL CULTURES IN TISSUE CULTURE. D. R. Aguirre, N. DiMassa, Chrystal Johnson, H. Eran, R. Perez, C.V.R. Sharma, M.V.R. Sharma,
Importance of inflammation reaction of scaffold for the application of regenerative medicine
 Review ArticleInflammation reaction of scaffold for regenerative medicine 178 Review Article Importance of inflammation reaction of scaffold for the application of regenerative medicine Gilson Khang Department
Review ArticleInflammation reaction of scaffold for regenerative medicine 178 Review Article Importance of inflammation reaction of scaffold for the application of regenerative medicine Gilson Khang Department
Xeno-Free Systems for hesc & hipsc. Facilitating the shift from Stem Cell Research to Clinical Applications
 Xeno-Free Systems for hesc & hipsc Facilitating the shift from Stem Cell Research to Clinical Applications NutriStem Defined, xeno-free (XF), serum-free media (SFM) specially formulated for growth and
Xeno-Free Systems for hesc & hipsc Facilitating the shift from Stem Cell Research to Clinical Applications NutriStem Defined, xeno-free (XF), serum-free media (SFM) specially formulated for growth and
Passive Immunization Trials to Inform Vaccine Design
 Passive Immunization Trials to Inform Vaccine Design Points for Consideration from deliberations held at the August 8, 2014 workshop Contents I. Introduction... 2 II. Types of trials... 2 1. Therapeutic
Passive Immunization Trials to Inform Vaccine Design Points for Consideration from deliberations held at the August 8, 2014 workshop Contents I. Introduction... 2 II. Types of trials... 2 1. Therapeutic
Stem Cell Principle -
 Effective Date: 31.10.2017 Doc ID: 20290214 Version: 1.0 Status: Approved Planned Effective Date: 31-Oct-2017 00:00 CET (Server Date) Stem Cell Principle - Rationale Research on human stem cells and their
Effective Date: 31.10.2017 Doc ID: 20290214 Version: 1.0 Status: Approved Planned Effective Date: 31-Oct-2017 00:00 CET (Server Date) Stem Cell Principle - Rationale Research on human stem cells and their
CHAPTER 7 CELLULAR BASIS OF ANTIBODY DIVERSITY: CLONAL SELECTION
 CHAPTER 7 CELLULAR BASIS OF ANTIBODY DIVERSITY: CLONAL SELECTION The specificity of humoral immune responses relies on the huge DIVERSITY of antigen combining sites present in antibodies, diversity which
CHAPTER 7 CELLULAR BASIS OF ANTIBODY DIVERSITY: CLONAL SELECTION The specificity of humoral immune responses relies on the huge DIVERSITY of antigen combining sites present in antibodies, diversity which
Interactions of cells with their environment; Engineering materials with biological recognition
 Interactions of cells with their environment; Engineering materials with biological recognition Last time: Today: Reading: Supplementary Reading: Polyelectrolyte hydrogel swelling thermodynamics Applications
Interactions of cells with their environment; Engineering materials with biological recognition Last time: Today: Reading: Supplementary Reading: Polyelectrolyte hydrogel swelling thermodynamics Applications
Embryonic development, epigenics and somatic cell nuclear transfer - The science and its social implications -
 Embryonic development, epigenics and somatic cell nuclear transfer - The science and its social implications - Moshe Yaniv Unité d Expression Génétique et Maladies, Institut Pasteur, Paris, France September
Embryonic development, epigenics and somatic cell nuclear transfer - The science and its social implications - Moshe Yaniv Unité d Expression Génétique et Maladies, Institut Pasteur, Paris, France September
Flow CAST : Testing Potency and Efficacy of Inhibitors of PI3K δ, PI3Kγ, BTK and SYK Activity
 Flow CAST : Testing Potency and Efficacy of Inhibitors of PI3K δ, PI3Kγ, BTK and SYK Activity Michele Romano, PhD Product Manager Flow CAST is for Research Use Only. Not for use in diagnostic procedures.
Flow CAST : Testing Potency and Efficacy of Inhibitors of PI3K δ, PI3Kγ, BTK and SYK Activity Michele Romano, PhD Product Manager Flow CAST is for Research Use Only. Not for use in diagnostic procedures.
Wnt16 smact merge VK/AB
 A WT Wnt6 smact merge VK/A KO ctrl IgG WT KO Wnt6 smact DAPI SUPPLEMENTAL FIGURE I: Wnt6 expression in MGP-deficient aortae. Immunostaining for Wnt6 and smooth muscle actin (smact) in aortae from 7 day
A WT Wnt6 smact merge VK/A KO ctrl IgG WT KO Wnt6 smact DAPI SUPPLEMENTAL FIGURE I: Wnt6 expression in MGP-deficient aortae. Immunostaining for Wnt6 and smooth muscle actin (smact) in aortae from 7 day
3D-Zellkulturgerüste für die biomedizinische Grundlagenforschung
 3D-Zellkulturgerüste für die biomedizinische Grundlagenforschung Zoologisches Institut, Fakultät für Chemie- und Biowissenschaften, Zell- und Neurobiologie KIT University of the State of Baden-Württemberg
3D-Zellkulturgerüste für die biomedizinische Grundlagenforschung Zoologisches Institut, Fakultät für Chemie- und Biowissenschaften, Zell- und Neurobiologie KIT University of the State of Baden-Württemberg
Learning Objectives. Define RNA interference. Define basic terminology. Describe molecular mechanism. Define VSP and relevance
 Learning Objectives Define RNA interference Define basic terminology Describe molecular mechanism Define VSP and relevance Describe role of RNAi in antigenic variation A Nobel Way to Regulate Gene Expression
Learning Objectives Define RNA interference Define basic terminology Describe molecular mechanism Define VSP and relevance Describe role of RNAi in antigenic variation A Nobel Way to Regulate Gene Expression
Chapter 3 Nucleic Acids, Proteins, and Enzymes
 3 Nucleic Acids, Proteins, and Enzymes Chapter 3 Nucleic Acids, Proteins, and Enzymes Key Concepts 3.1 Nucleic Acids Are Informational Macromolecules 3.2 Proteins Are Polymers with Important Structural
3 Nucleic Acids, Proteins, and Enzymes Chapter 3 Nucleic Acids, Proteins, and Enzymes Key Concepts 3.1 Nucleic Acids Are Informational Macromolecules 3.2 Proteins Are Polymers with Important Structural
ANAT 2341 Embryology Lecture 18 Stem Cells
 ANAT 2341 Embryology Lecture 18 Stem Cells 29 September 2010 Dr Antonio Lee Neuromuscular & Regenera
ANAT 2341 Embryology Lecture 18 Stem Cells 29 September 2010 Dr Antonio Lee Neuromuscular & Regenera
RoleofBiomaterialsinNeuralStemCellFate
 Global Journal of Science Frontier Research: G Bio-Tech & Genetics Volume 15 Issue 1 Version 1.0 Year 2015 Type: Double Blind Peer Reviewed International Research Journal Publisher: Global Journals Inc.
Global Journal of Science Frontier Research: G Bio-Tech & Genetics Volume 15 Issue 1 Version 1.0 Year 2015 Type: Double Blind Peer Reviewed International Research Journal Publisher: Global Journals Inc.
DNA Microarray Technology
 CHAPTER 1 DNA Microarray Technology All living organisms are composed of cells. As a functional unit, each cell can make copies of itself, and this process depends on a proper replication of the genetic
CHAPTER 1 DNA Microarray Technology All living organisms are composed of cells. As a functional unit, each cell can make copies of itself, and this process depends on a proper replication of the genetic
Introduction State the objectives of the work and provide adequate background, avoiding a detailed literature survey or a summary of the results.
 Format requirements for IJBCB articles For each article type, IJBCB has its own requirements regarding the article format. Please ensure you follow the exact guidelines (included below) for the specific
Format requirements for IJBCB articles For each article type, IJBCB has its own requirements regarding the article format. Please ensure you follow the exact guidelines (included below) for the specific
Lecture 23: Clinical and Biomedical Applications of Proteomics; Proteomics Industry
 Lecture 23: Clinical and Biomedical Applications of Proteomics; Proteomics Industry Clinical proteomics is the application of proteomic approach to the field of medicine. Proteome of an organism changes
Lecture 23: Clinical and Biomedical Applications of Proteomics; Proteomics Industry Clinical proteomics is the application of proteomic approach to the field of medicine. Proteome of an organism changes
Stem cells and motor neurone disease
 Stem cells and motor neurone disease F Stem cell research has fuelled hope of a treatment for a variety of conditions. This information sheet explains what these cells are and includes details of the current
Stem cells and motor neurone disease F Stem cell research has fuelled hope of a treatment for a variety of conditions. This information sheet explains what these cells are and includes details of the current
RayBio Human NF-κB p65 Transcription Factor Activity Assay Kit
 RayBio Human NF-κB p65 Transcription Factor Activity Assay Kit Catalog #: TFEH-p65 User Manual Mar 13, 2017 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,
RayBio Human NF-κB p65 Transcription Factor Activity Assay Kit Catalog #: TFEH-p65 User Manual Mar 13, 2017 3607 Parkway Lane, Suite 200 Norcross, GA 30092 Tel: 1-888-494-8555 (Toll Free) or 770-729-2992,
Charles Shuler, Ph.D. University of Southern California Los Angeles, California Approved for Public Release; Distribution Unlimited
 AD Award Number: DAMD17-01-1-0100 TITLE: Smad-Mediated Signaling During Prostate Growth and Development PRINCIPAL INVESTIGATOR: Charles Shuler, Ph.D. CONTRACTING ORGANIZATION: University of Southern California
AD Award Number: DAMD17-01-1-0100 TITLE: Smad-Mediated Signaling During Prostate Growth and Development PRINCIPAL INVESTIGATOR: Charles Shuler, Ph.D. CONTRACTING ORGANIZATION: University of Southern California
In vitro cultures of bone marrow stromal cells and progenitor B cells can accurately recapitulate the normal steps of B cell development.
 Regular Office Hours: Tuesdays 11-12 Extra office hours: Wed, Feb 7 12-1pm Thurs, Feb 8 11am-12 Fri, Feb 9 2-4pm I WILL NOT BE HOLDING OFFICE HOURS ON TUESDAY Feb 13!! Dina, Tim, and I encourage all confused
Regular Office Hours: Tuesdays 11-12 Extra office hours: Wed, Feb 7 12-1pm Thurs, Feb 8 11am-12 Fri, Feb 9 2-4pm I WILL NOT BE HOLDING OFFICE HOURS ON TUESDAY Feb 13!! Dina, Tim, and I encourage all confused
B cell development The stages of B cell development
 Regular Office Hours: Tuesdays 11-12 Extra office hours: Wed, Feb 7 12-1pm Thurs, Feb 8 11am-12 Fri, Feb 9 2-4pm I WILL NOT BE HOLDING OFFICE HOURS ON TUESDAY Feb 13!! Dina, Tim, and I encourage all confused
Regular Office Hours: Tuesdays 11-12 Extra office hours: Wed, Feb 7 12-1pm Thurs, Feb 8 11am-12 Fri, Feb 9 2-4pm I WILL NOT BE HOLDING OFFICE HOURS ON TUESDAY Feb 13!! Dina, Tim, and I encourage all confused
Genetics - Problem Drill 19: Dissection of Gene Function: Mutational Analysis of Model Organisms
 Genetics - Problem Drill 19: Dissection of Gene Function: Mutational Analysis of Model Organisms No. 1 of 10 1. The mouse gene knockout is based on. (A) Homologous recombination (B) Site-specific recombination
Genetics - Problem Drill 19: Dissection of Gene Function: Mutational Analysis of Model Organisms No. 1 of 10 1. The mouse gene knockout is based on. (A) Homologous recombination (B) Site-specific recombination
Functional Assessment and Clinical Outcome
 Tissue Engineering Functional Assessment and Clinical Outcome STEVEN A. GOLDSTEIN Orthopaedic Research Laboratories, Department of Orthopaedic Surgery, University of Michigan, Ann Arbor, Michigan 48109,
Tissue Engineering Functional Assessment and Clinical Outcome STEVEN A. GOLDSTEIN Orthopaedic Research Laboratories, Department of Orthopaedic Surgery, University of Michigan, Ann Arbor, Michigan 48109,
RegenACR. autologous cellular regeneration. RegenACR A-PRP treatment. collagen fibers. 1,2 texture and elasticity. 3 volume & strength.
 RegenACR autologous cellular regeneration RegenACR A-PRP treatment collagen fibers. 1,2 texture and elasticity. 3 volume & strength. 4,5 WHAT IS A-PRP Autologous Platelet Rich Plasma Plasma contains many
RegenACR autologous cellular regeneration RegenACR A-PRP treatment collagen fibers. 1,2 texture and elasticity. 3 volume & strength. 4,5 WHAT IS A-PRP Autologous Platelet Rich Plasma Plasma contains many
Application of RNA Interference to Anti-Doping. Matthew Fedoruk, Ph.D. Gene & Cell Doping Symposium 2013, Beijing, China
 Application of RNA Interference to Anti-Doping Matthew Fedoruk, Ph.D. Gene & Cell Doping Symposium 2013, Beijing, China From Plants to Worms to Humans: Discovery and Mechanism of RNAi Alnylam Pharmaceuticals
Application of RNA Interference to Anti-Doping Matthew Fedoruk, Ph.D. Gene & Cell Doping Symposium 2013, Beijing, China From Plants to Worms to Humans: Discovery and Mechanism of RNAi Alnylam Pharmaceuticals
Biological immune systems
 Immune Systems 1 Introduction 2 Biological immune systems Living organism must protect themselves from the attempt of other organisms to exploit their resources Some would-be exploiter (pathogen) is much
Immune Systems 1 Introduction 2 Biological immune systems Living organism must protect themselves from the attempt of other organisms to exploit their resources Some would-be exploiter (pathogen) is much
PCR Arrays. An Advanced Real-time PCR Technology to Empower Your Pathway Analysis
 PCR Arrays An Advanced Real-time PCR Technology to Empower Your Pathway Analysis 1 Table of Contents 1. Introduction to the PCR Arrays 2. How PCR Arrays Work 3. Performance Data from PCR Arrays 4. Research
PCR Arrays An Advanced Real-time PCR Technology to Empower Your Pathway Analysis 1 Table of Contents 1. Introduction to the PCR Arrays 2. How PCR Arrays Work 3. Performance Data from PCR Arrays 4. Research
3D In Vitro Living Systems for Biological Application
 3D In Vitro Living Systems for Biological Application Hossein Hosseinkhani Graduate Institute of Biomedical Engineering, National Taiwan University of Science and Technology (TAIWAN TECH), Taipei, Taiwan
3D In Vitro Living Systems for Biological Application Hossein Hosseinkhani Graduate Institute of Biomedical Engineering, National Taiwan University of Science and Technology (TAIWAN TECH), Taipei, Taiwan
Stem Cells and Regenerative Medicine
 Q 0 Stem Cells and Regenerative Medicine Technologies and capabilities to add value to your company Differentiation HA-PPS for cartilage Parallel culture optimisation Stem Cell Therapies Fetal EPCs and
Q 0 Stem Cells and Regenerative Medicine Technologies and capabilities to add value to your company Differentiation HA-PPS for cartilage Parallel culture optimisation Stem Cell Therapies Fetal EPCs and
Manipulation of a VEGF-Notch signaling circuit drives formation of functional vascular endothelial progenitors from human pluripotent stem cells
 Manipulation of a VEGF-Notch signaling circuit drives formation of functional vascular endothelial progenitors from human pluripotent stem cells The Harvard community has made this article openly available.
Manipulation of a VEGF-Notch signaling circuit drives formation of functional vascular endothelial progenitors from human pluripotent stem cells The Harvard community has made this article openly available.