8.1 Why Do Cells Divide?
|
|
|
- August Farmer
- 5 years ago
- Views:
Transcription
1 8.1 Why Do Cells Divide? Cells reproduce by cell division. One cell gives rise to two or more cells, called daughter cells. Each daughter cell receives a complete set of heredity information identical to the information in the parent cell and about half of the cytoplasm. Cell division transmits hereditary information to each daughter cell. The hereditary information in each cell is deoxyribonucleic acid (DNA). DNA is contained in chromosomes. A molecule of DNA consists of smaller subunits called nucleotides.
2 8.1 Nucleotide Structure A nucleotide consists of a phosphate, a sugar (deoxyribose), and one of four bases. Adenine (A) Thymine (T) Guanine (G) Cytosine (C). The nucleotides are held together by hydrogen bonding between the bases in two strands forming a double helix.

3 8.1 Why Do Cells Divide? The structure of DNA phosphate nucleotide base sugar A C T G A G C C C G A T A C G A T T T A (a) A single strand of DNA (b) The double helix Fig. 8-1
4 8.1 Why Do Cells Divide? Genes, segments DNA, are the units of inheritance. Each gene spells out the instructions for making one or more proteins. When a cell divides, it first replicates its DNA, and a copy is transferred into each daughter cell.
5 8.1 Why Do Cells Divide? Cell division is required for growth and development. Cell division in which the daughter cells are genetically identical to the parent cell is called mitotic cell division. After cell division, the daughter cells may grow and divide again, or may differentiate, becoming specialized for specific functions. The repeating pattern of division, growth, and differentiation followed again by division is called the cell cycle.
6 8.1 Why Do Cells Divide? Most multicellular organisms have three categories of cells. Stem cells: retain the ability to divide and can differentiate into a variety of cell types Other cells capable of dividing: typically differentiate only into one or two different cell types (progenitor cells) Permanently differentiated cells: differentiated cells that can never divide again
7 8.1 Why Do Cells Divide? Cell division is required for sexual and asexual reproduction. Sexual reproduction in eukaryotic organisms occurs when offspring are produced by the fusion of gametes (sperm and eggs) from two adults. Gametes are produced by meiotic cell division, which results in daughter cells with exactly half of the genetic information of their parent cells. Fertilization of an egg by a sperm results in the restoration of the full complement of hereditary information in the offspring.
8 8.1 Why Do Cells Divide? Reproduction in which offspring are formed from a single parent, without having a sperm fertilize an egg, is called asexual reproduction. Asexual reproduction produces offspring that are genetically identical to the parent. Examples of asexual reproduction occur in bacteria, single-celled eukaryotic organisms, multicellular organisms such as Hydra, and many trees, plants, and fungi.

9 8.2 What Occurs During The Prokaryotic Cell Cycle? The prokaryotic cell cycle consists of a long period of growth, during which the cell duplicates its DNA. cell division by binary fission cell growth and DNA replication The prokaryotic cell cycle
10 8.2 What Occurs During the Prokaryotic Cell Cycle? Cell division in prokaryotes occurs by binary fission, which means splitting in two. The prokaryotic chromosome is attached at one point to the plasma membrane of the cell.




11 8.2 The Prokaryotic Cell Cycle? cell wall attachment site plasma membrane The circular DNA double helix is attached to the plasma membrane at one point. circular DNA The DNA replicates and the two DNA double helices attach to the plasma membrane at nearby points. New plasma membrane is added between the attachment points, pushing them further apart. The plasma membrane grows inward at the middle of the cell. Fig. 8-3b(1)
 The parent cell divides into two daughter cells. Fig.")
12 8.2 The Prokaryotic Cell Cycle The prokaryotic cell cycle (continued) The parent cell divides into two daughter cells. Fig. 8-3b(5)
13 8.3 How Is The DNA In Eukaryotic Cells Organized? Unlike prokaryotic chromosomes, eukaryotic chromosomes are separated from the cytoplasm by a membrane-bound nucleus. Eukaryotic cells always have multiple chromosomes. Eukaryotic chromosomes contain more DNA than prokaryotic chromosomes. The eukaryotic chromosome consists of DNA bound to protein.
14 8.3 How Is The DNA In Eukaryotic Cells Organized? Duplicated chromosomes separate during cell division. Prior to cell division, the DNA within each chromosome is replicated. The duplicated chromosomes then consist of two DNA double helixes and associated proteins that are attached to each other at the centromere. Each of the duplicated chromosomes attached at the centromere is called a sister chromatid. During mitotic cell division, the sister chromatids separate and each becomes a separate chromosome that is delivered to one of the two resulting daughter cells.
 (a) A replicated chromosome consists of two sister chromatids")
15 8.3 How Is the DNA In Eukaryotic Cells Organized? Eukaryotic chromosomes during cell division centromere genes duplicated sister chromosome chromatids (2 DNA double helices) (a) A replicated chromosome consists of two sister chromatids independent daughter chromosomes, each with one identical DNA double helix (b) Sister chromatids separate during cell division Fig. 8-5
16 8.3 How Is The DNA In Eukaryotic Cells Organized? Chromosomes with the same genes are called homologous chromosomes, or homologues. Cells with pairs of homologous chromosomes are called diploid. Homologous chromosomes are usually not identical. The same genes on homologous chromosomes may be different due to mutations, changes in the sequence of nucleotides in the DNA.
17 8.3 How Is The DNA In Eukaryotic Cells Organized? Not all cells have paired chromosomes. The ovaries and testes undergo a special kind of cell division, called meiotic cell division, to produce gametes (eggs and sperm). Gametes contain only one member of each pair of autosomes, plus one of the two sex chromosomes. Cells with half the number of each type of chromosome are called haploid cells. Fusion of two haploid cells at fertilization produces a diploid cell with the full complement of chromosomes.
18 8.3 How Is The DNA In Eukaryotic Cells Organized? The number of different types of chromosomes in a species is called the haploid number and is designated n. In humans, n = 23. Diploid cells contain 2n chromosomes. Humans body cells contain 2n = 46 (2 x 23) chromosomes.
19 8.4 What Occurs During The Eukaryotic Cell Cycle? The eukaryotic cell cycle is divided into two major phases: interphase and cell division. During interphase, the cell acquires nutrients from its environment, grows, and duplicates its chromosomes. During cell division, one copy of each chromosome and half of the cytoplasm are parceled out into each of two daughter cells.

20 8.4 What Occurs During The Eukaryotic Cell Cycle? The eukaryotic cell cycle prophase metaphase anaphase mitotic cell division cytokinesis telophase and cell growth and differentiation cell growth interphase synthesis of DNA; chromosomes are duplicated Fig. 8-7
21 8.4 What Occurs During The Eukaryotic Cell Cycle? There are two types of division in eukarytic cells: mitotic cell division and meiotic cell division. Mitotic cell division may be thought of as ordinary cell division, such as occurs during development from a fertilized egg, during asexual reproduction, and in skin, liver, and the digestive tract every day. Meiotic cell division is a specialized type of cell division required for sexual reproduction.
22 8.4 What Occurs During The Eukaryotic Cell Cycle? Mitotic cell division Mitotic cell division consists of nuclear division (called mitosis) followed by cytoplasmic division (called cytokinesis) and the formation of two daughter cells.
23 8.4 What Occurs During The Eukaryotic Cell Cycle? Meiotic cell division Is a prerequisite for sexual reproduction in all eukaryotic organisms. Meiotic cell division involves a specialized nuclear division called meiosis. It involves two rounds of cytokinesis, producing four daughter cells that can become gametes.
24 8.4 The Eukaryotic Cell Cycle The life cycle of eukaryotic organisms include both mitotic and meiotic cell division. mitotic cell division, differentiation, and growth mitotic cell division, differentiation, and growth baby adults mitotic cell division, differentiation, and growth haploid diploid embryo fertilized egg egg fusion of gametes meiotic cell division in ovaries sperm meiotic cell division in testes Flash Fig. 8-8
25 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? Mitosis is divided into four phases. Prophase Metaphase Anaphase Telophase Flash

26 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? Interphase, prophase, and metaphase nuclear envelope chromatin nucleolus condensing chromosomes spindle pole spindle microtubules centriole pairs beginning of spindle formation spindle pole kinetochore (a) Late Interphase (b) Early Prophase (c) Late Prophase The (d) Metaphase Duplicated chromosomes are in the relaxed uncondensed state; duplicated centrioles remain clustered. Chromosomes condense and shorten; spindle microtubules begin to form between separating centriole pairs. nucleolus disappears; the nuclear envelope breaks down; spindle microtubules attach to the kinetochore of each sister chromatid. Kinetochores interact; spindle microtubules line up the chromosomes at the cell s equator. Fig. 8-9a d
 Anaphase Sister (f) Telophase One set of (g) Cytokinesis (h)")
27 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? Anaphase, telophase, cytokinesis, and interphase unattached spindle microtubules chromosomes extending nuclear envelope re-forming (e) Anaphase Sister (f) Telophase One set of (g) Cytokinesis (h) Interphase of chromatids separate chromosomes reaches The cell divides in daughter cells Spindles and move to opposite each pole and relaxes two; each daughter disappear, intact nuclear poles of the cell; spindle into the extended state; cell receives one envelopes form, microtubules that are nuclear envelopes start nucleus and about chromosomes extend not attached to the to form around each set; half of the cytoplasm. completely, and the chromosomes push the spindle microtubles nucleolus reappears. poles apart. begin to disappear. Fig. 8-9e h
28 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? Three major events happen in prophase: The duplicated chromosomes condense. The spindle microtubules form and attach to the kinetochore of the chromatids. The chromosomes migrate with the spindle poles to opposite sides of the nucleus. Fig. 8-9b c

29 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? During metaphase, the chromosomes line up along the equator of the cell. At this phase, the spindle apparatus lines up the sister chromatids at the equator, with one kinetochore facing each cell pole. Fig. 8-9d
30 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? During anaphase, sister chromatids separate and move to opposite poles of the cell. Sister chromatids separate, becoming independent daughter chromosomes. The kinetochores pull the chromosomes poleward along the spindle microtubules.

31 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? During telophase, nuclear envelopes form around both groups of chromosomes. Telophase begins when the chromosomes reach the poles. The spindle microtubules disintegrate and the nuclear envelop forms around each group of chromosomes.

32 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? During Telophase cytokinesis occurs splitting the cytoplasm between the two daughter cells ending mitosis and beginning Interphase. Microfilaments form a ring around the cell s equator. The microfilament ring contracts, pinching in the cell s waist. The waist completely pinches off, forming two daughter cells (a) Microfilaments contract, pinching the cell in two (b) Scanning electron micrograph of cytokinesis. Fig. 8-10

33 8.5 How Does Mitotic Cell Division Produce Genetically Identical Daughter Cells? Cytokinesis in plant cells has an additional step. Fig. 8-11
34 8.6 How Does Meiotic Cell Division Produce Haploid Cells? Meiosis is the production of haploid cells with unpaired chromosomes derived from diploid parent cells with paired chromosomes. Meiosis includes two nuclear divisions, known as meiosis I and meiosis II. In meiosis I, homologous chromosomes pair up, but sister chromatids remain connected to each other. In meiosis II, chromosomes behave as they do in mitosis sister chromatids separate and are pulled to opposite poles of the cell. Flash
 Prophase I (b) Metaphase I (c) Anaphase I (d) Telophase I Duplicated chromosomes condense.")
35 8.6 How Does Meiotic Cell Division Produce Haploid Cells? paired homologous chromosomes recombined chromatids chiasma spindle microtubule kinetochores (a) Prophase I (b) Metaphase I (c) Anaphase I (d) Telophase I Duplicated chromosomes condense. Homologous chromosomes pair up and chiasmata occur as chromatids of homologues exchange parts by crossing over. The nuclear envelope disintegrates, and spindle microtubules form. Paired homologous chromosomes line up along the equator of the cell. One homologue of each pair faces each pole of the cell and attaches to the spindle microtubules via the kinetochore (blue). Homologues separate, one member of each pair going to each pole of the cell. Sister chromatids do not separate. Spindle microtubules disappear. Two clusters of chromosomes have formed, each containing one member of each pair of homologues. The daughter nuclei are therefore haploid. Cytokinesis commonly occurs at this stage. There is little or no interphase between meiosis I and meiosis II. Fig. 8-12a d

36 8.6 How Does Meiotic Cell Division Produce Haploid Cells? (e) Prophase II (f) Metaphase II (g) Anaphase II (h) Telophase II (i) If the chromosomes have relaxed after telophase I, they recondense. Spindle microtubules re-form and attach to the sister chromatids. The chromosomes line up along the equator, with sister chromatids of each chromosome attached to spindle microtubules that lead to opposite poles. The chromatids separate into independent daughter chromosomes, one former chromatid moving toward each pole. The chromosomes finish moving to opposite poles. Nuclear envelopes re-form, and the chromosomes become extended again (not shown here). Four haploid cells Cytokinesis results in four haploid cells, each containing one member of each pair of homologous chromosomes (shown here in the condensed state). Fig. 8-12e i

37 8.6 How Does Meiotic Cell Division Produce Haploid Cells? Flash
38 8.7 How Do Meiotic Cell Division And Sexual Reproduction Produce Genetic Variability? Ways to produce genetic variability from meiotic cell division and sexual reproduction Shuffling of homologues Crossing over Fusion of gametes

39 8.7 Meiotic Cell Division And Sexual Reproduction Produce Genetic Variability Shuffling of homologues creates novel combinations of chromosomes during meiosis producing genetic variability. (a) The four possible chromosome arrangements at metaphase of meiosis I (b) The eight possible sets of chromosomes after meiosis I Fig. 8-13
 parts of chromosomes that have been exchanged between")
40 8.7 Meiotic Cell Division And Sexual Reproduction Produce Genetic Variability Crossing over creates chromosomes with novel combinations of genetic material. sister chromatids of one duplicated homologue pair of homologous duplicated chromosomes chiasmata (sites of crossing over) parts of chromosomes that have been exchanged between homologues Fig. 8-14
41 8.7 How Do Meiotic Cell Division And Sexual Reproduction Produce Genetic Variability? Fusion of gametes creates genetically variable offspring. Because every egg and sperm are genetically unique, and it is random as to which sperm fertilizes which egg, every fertilized egg is also genetically unique.
42 9.1 What Is The Physical Basis Of Inheritance? Inheritance occurs when genes are transmitted from parent to offspring. The units of inheritance are genes. Genes are segments of DNA at specific locations on chromosomes. A gene s physical location on a chromosome is called its locus. Each member of a pair of homologous chromosomes carries the same genes, located at the same loci. Different versions of a gene at a given locus are called alleles.
43 9.1 What Is The Physical Basis Of Inheritance? The relationship among genes, alleles, and chromosomes gene loci a pair of homologous chromosomes Both chromosomes carry the same allele of the gene at this locus; the organism is homozygous at this locus This locus contains another gene for which the organism is homozygous Each chromosome carries a different allele of this gene, so the organism is heterozygous at this locus the chromosome from the male parent the chromosome from the female parent Fig. 9-1
44 9.1 What Is The Physical Basis Of Inheritance? Mutations are the source of alleles. Differences in alleles at a given locus are due to mutations at that gene. If a mutation occurs in gametes, it can be passed on from parent to offspring. An organism s two alleles may be the same or different. A diploid organism has pairs of homologous chromosomes with two copies of each gene at a given locus.
45 9.1 What Is The Physical Basis Of Inheritance? An organism s two alleles may be the same or different (continued). If both homologous chromosomes have the same allele at a locus, the organism is said to be homozygous. If two homologous chromosomes have different alleles at a locus, the organism is heterozygous at that locus. The gametes of a homozygous individual are all the same at a particular locus, while gametes of a heterozygous individual would contain half one allele and half the other allele.
46 9.3 How Are Single Traits Inherited? True-breeding traits of organisms, such as purple flower color, are always inherited by all of their offspring that result from selffertilization. In one experiment, Mendel cross-fertilized white-flowered plants with purple-flowered plants. When he grew the resulting seeds, he found all the first-generation offspring, or the F 1 generation, produced purple flowers. What happened to the white color?
47 9.3 How Are Single Traits Inherited? The F 2 generation Next, Mendel allowed the F 1 flowers to selffertilize, collected the seeds, and grew the second generation, called the F 2 generation. Flowers in the F 2 generation were threefourths purple and one-fourth white, in a ratio of 3 purple to 1 white. This showed that the gene for white flowers was hidden in the F 1 generation, but appeared again in the F 2 generation.
 self-fertilize")
48 9.3 How Are Single Traits Inherited? Cross of F 1 plants with purple flowers Firstgeneration offspring (F 1 ) self-fertilize Secondgeneration offspring (F 2 ) 3/4 purple 1/4 white Fig. 9-5
49 9.3 How Are Single Traits Inherited? All the white-flowered plants in the F 2 generation only produced additional whiteflowered plants. Purple-flowered plants were of two types: About ⅔ were true-breeding for purple, while ⅔ produced both purple- and white-flowered offspring (ratio 3 purple/1 white). Therefore, the F 2 generation included ¼ truebreeding purple plants, ½ hybrid purple, and ¼ true-breeding white plants.
50 9.3 How Are Single Traits Inherited? This allows us to develop a five-part hypothesis to explain the inheritance of single traits. 1. Each trait is determined by pairs of distinct physical units called genes. There are two alleles for each gene, one on each homologous chromosome. 2. When two different alleles are present in an organism, the dominant allele may mask the expression of the recessive allele; but the recessive allele is still present. 3. The two alleles of a gene segregate (separate) from one another during meiosis (Mendel s law of segregation).
51 9.3 How Are Single Traits Inherited? 4. Which allele ends up in any given gamete is determined by chance. 5. True-breeding (homozygous) organisms have two copies of the same allele for a given gene; hybrid (heterozygous) organisms have two different alleles for a given gene.
 Gametes produced by a homozygous parent")
52 9.3 How Are Single Traits Inherited? The distribution of alleles in gametes homozygous parent gametes A A A A (a) Gametes produced by a homozygous parent heterozygous parent gametes A a A a (b) Gametes produced by a heterozygous parent Fig. 9-6
53 9.3 How Are Single Traits Inherited? Mendel s hypothesis was that two plants may look alike (phenotype) but have a different allele composition (genotype). Purple pea plants had PP or Pp genotypes, but their phenotype (purple color) was the same. The F 2 generation could be described as having three genotypes (¼ PP, ½ Pp, and ¼ pp) and two phenotypes (¾ purple and ¼ white).

54 9.3 How Are Single Traits Inherited? The Punnett square method Pp self-fertilize 1 2 P eggs 1 2 p 1 2 P sperm 1 4 PP 1 4 Pp 1 2 p pp 4 pp Fig. 9-8
55 9.3 How Are Single Traits Inherited? Mendel predicted the outcome of cross-fertilizing Pp plants with homozygous recessive plants (pp) there should be equal numbers of Pp (purple) and pp (white) offspring. all sperm if PP P PP or Pp sperm unknown p eggs all Pp pollen if Pp sperm P p pp all eggs 1 2 p Pp p eggs 1 2 pp Fig. 9-9
56 9.4 How Are Multiple Traits Inherited? Mendel next crossed pea plants that differed in two traits, such as seed color (yellow or green) and seed shape (smooth or wrinkled). He knew from previous crosses that smooth and yellow were both dominant traits in peas. His first cross was a true-breeding plant with smooth, yellow seeds (SSYY) to a truebreeding plant with wrinkled, green seeds (ssyy).
57 9.4 How Are Multiple Traits Inherited? All the offspring of this cross (F 1 generation) were SsYy and had smooth, yellow seeds (both dominant traits). F 1 plants were allowed to self-fertilize and produced F 2 offspring in the phenotypic ratio 9:3:3:1.
58 9.4 How Are Multiple Traits Inherited? Mendel concluded that multiple traits are inherited independently. Mendel realized that these results could be explained if the genes for seed color and seed shape were inherited independently. The independent inheritance of two or more distinct traits is called the law of independent assortment. Multiple traits are inherited independently because the alleles of one gene are distributed to gametes independently of the alleles of other genes.

59 9.4 How Are Multiple Traits Inherited? Predicting genotypes and phenotypes SsYy self-fertilize eggs 1 4 SY 1 4 Sy 1 4 sy 1 4 sy 1 4 SY 1 16 SSYY 1 16 SSYy 1 16 SsYY 1 16 SsYy sperm Sy sy SSyY ssyy SSyy ssyy SsyY ssyy Ssyy ssyy 1 4 sy 1 16 ssyy 1 16 ssyy 1 16 ssyy 1 16 ssyy Fig. 9-11

60 9.4 How Are Multiple Traits Inherited? Independent assortment of alleles S Y chromosomes replicate s y pairs of alleles on homologous chromosomes in diploid cells S s Y y replicated homologues pair during metaphase of meiosis I, orienting like this or like this S s y Y meiosis I S Y s y S y s Y S Y s y S y s Y meiosis II S S s s S S s s Y Y y y y y Y Y SY sy Sy sy independent assortment produces four equally likely allele combinations during meiosis Fig. 9-12
61 9.5 How Are Genes Located on the Same Chromosome Inherited? Genetic linkage is the inheritance of genes as a group because they are on the same chromosome. Genes that are located on the same chromosome are inherited together, rather than sorted independently. In peas, the gene for flower color and the gene for pollen shape occur on the same chromosome and are inherited together. Because the two genes are located on the same chromosomes, they tend to end up in gametes together, and are then expressed in the plants.
62 9.5 How Are Genes Located on the Same Chromosome Inherited? Crossing over can create new combinations of linked alleles. Genes on the same chromosome do not always stay together. During prophase I of meiosis, homologous chromosomes sometimes exchange parts in the process, called crossing over. Crossing over produces a new allele combination on both homologous chromosomes. Therefore, the chromosomes of each haploid daughter cell receives different combinations of alleles from those of the parent cell.

63 9.6 How Is Sex Determined? Offspring sex is determined sex chromosomes. In mammals, females have two X chromosomes and males have an X chromosome and a Y chromosome. Y chromosomes are much smaller than the X chromosomes. female parent X 1 X 2 eggs X 1 X 2 X 1 X m X 2 X m male parent X m X m Y sperm female offspring X 1 Y X 2 Y Y male offspring
64 9.7 How Are Sex-Linked Genes Inherited? Genes that are found on one sex chromosome but not on the other are called sex-linked. Because females have two X chromosomes, they can be either homozygous or heterozygous for genes on the X chromosome. Males only have one X chromosome, and therefore express all the alleles they have on their X chromosome.
65 9.8 Do Mendelian Rules Of Inheritance Apply To All Traits? When a heterozygous phenotype is intermediate between the two homozygous phenotypes, the pattern of inheritance is called incomplete dominance. Human hair texture is influenced by a gene with two incompletely dominant alleles, C 1 and C 2. A person with two copies of the C 1 allele has curly hair; two copies of the C 2 allele produces straight hair; heterozygotes with C 1 C 2 genotype have wavy hair.
, ½ wavy C 1 C 2 eggs (C 1 C 2 ), and ¼ straight (C 2 C 2")
66 9.8 Do Mendelian Rules Of Inheritance Apply To All Traits? mother Two wavy-haired people could have the following children: ¼ curly (C 1 C 1 ), ½ wavy C 1 C 2 eggs (C 1 C 2 ), and ¼ straight (C 2 C 2 ). father C 1 sperm C 1 C 2 C 1 C 1 C 1 C 2 C 1 C 2 C 2 C 1 C 2 C 2 C 2 Fig. 9-17
67 9.8 Do Mendelian Rules Of Inheritance Apply To All Traits? A single gene may have multiple alleles. A single individual can have only two alleles for any gene, one on each homologous chromosomes. However, within all the members of a species there could be dozens of alleles for every gene.

68 9.8 Do Mendelian Rules Of Inheritance Apply To All Traits?
69 9.8 Do Mendelian Rules Of Inheritance Apply To All Traits? A single trait may be influenced by several genes. Many physical traits are governed not by single genes, but by interactions among two or more genes, a phenomenon called polygenic inheritance. The more genes that contribute to a single trait, the greater the number of phenotypes and the finer the distinctions among them.
70 9.10 How Are Single-Gene Disorders Inherited? Some human genetic disorders are caused by recessive alleles. Many genes encode information to synthesize enzymes or structural proteins in cells, and a defective allele in such a gene may cause damaged or inactive protein. In some cases, a defective gene may be masked when one normal allele is also present and makes enough functional protein.
71 9.10 How Are Single-Gene Disorders Inherited? Some human genetic disorders are caused by dominant alleles. Many genetic diseases are caused by dominant alleles, in which a single defective allele is enough to cause the disorder. For dominant diseases to be inherited, at least one parent must suffer from the disease but live long enough to have children. Some diseases, like Huntington disease, do not appear until after the affected person has reproduced.
72 10.1 What Is The Structure Of DNA? Individual traits of an organism are transmitted from parent to offspring in discrete units of DNA called genes. Genes are located on chromosomes found within the nucleus of cells. What makes all organisms different from each other is the arrangement and molecular composition of its genes.
73 10.1 What Is The Structure Of DNA? DNA is composed of four different subunits, called nucleotides. Each nucleotide has three parts: A phosphate group Deoxyribose, a 5 Carbon sugar One of four different nitrogen-containing bases Thymine Cytosine Adenine Guanine
74 10.1 What Is The Structure Of DNA? A DNA molecule contains two nucleotide strands. A DNA molecule consists of two DNA strands of linked nucleotides. Within each strand, the phosphate group of one nucleotide binds to the sugar group of the next nucleotide. The sugar-phosphate bonding produces a sugar-phosphate backbone to the DNA molecule.
 sugar free sugar (a) Hydrogen bonds hold")
75 10.1 What Is The Structure Of DNA? The Watson-Crick model of DNA structure nucleotide nucleotide free phosphate phosphate base (cytosine) sugar free sugar (a) Hydrogen bonds hold complementary base pairs (b) Two DNA strands form (c) Four turns of a together in DNA a double helix DNA double helix Fig. 10-2
76 10.1 What Is The Structure Of DNA? Nucleotide rungs only result in specific pair combinations. Adenine only pairs with Thymine. Guanine only pairs with Cytosine. This A T and G C coupling is called complementary base pairing.
77 10.3 How Is DNA Copied? Cells reproduce themselves by making two daughter cells from each parental cell, each with a complete copy of all the parental cell s genetic information. During cell reproduction, the parental cell synthesizes two exact copies of its DNA through a process called DNA replication. One copy goes into each daughter cell.
78 10.3 How Is DNA Copied? DNA replication produces two DNA double helices, each with one original strand and one new strand. DNA replication requires three ingredients: The parental DNA strands Free nucleotides that were synthesized in the cytoplasm and then imported to the nucleus Enzymes that unwind the parental DNA double helix and synthesize the new DNA strands
79 10.3 How Is DNA Copied? DNA replication produces two DNA double helices, each with one original strand and one new strand (continued). The first step involves enzymes called DNA helicases, which pull apart the parental DNA double helix. Next, enzymes called DNA polymerases move along each separated parental DNA strand, matching each base on the strand with free nucleotides. DNA replication keeps, or conserves, one parental DNA strand and produces one new daughter strand (semiconservative replication).

80 10.3 How Is DNA Copied? The basic features of DNA replication 1 Parental DNA double helix 2 The parental DNA is unwound 3 New DNA strands are synthesized with bases complementary to the parental strands free nucleotides 4 Each new double helix is composed of one parental strand (blue) and one new strand (red) Fig. 10-3
81 10.4 What Are The Mechanisms Of DNA Replication? DNA helicase separates the parental DNA strands by breaking the hydrogen bonds between complementary bases. This activity separates the two strands and forms a replication bubble where the parental strands are no longer paired. Replication then proceeds. There is a replication fork on each end of the bubble, where replication is taking place and the original DNA strand is unzipping. The unzipping and replication continues in both directions until the new strands are completely formed.
 DNA helicase replication forks")
82 10.4 What Are The Mechanisms Of DNA Replication? The mechanism of DNA replication, step (2) DNA helicase replication forks DNA helicase Fig. 10-5(2)
83 10.4 What Are The Mechanisms Of DNA Replication? DNA polymerase synthesizes new DNA strands. At the replication forks, DNA polymerase recognizes unpaired nucleotide bases in the parental strand and matches them up with free nucleotides. It then links up the phosphate of the incoming nucleotide with the sugar of the previously added nucleotide, thereby contributing to the growing molecule backbone.
84 10.4 What Are The Mechanisms Of DNA Replication? DNA helicase and DNA polymerase work together to copy each strand of separated parental DNA. Polymerase # 1 lands on one strand of DNA and follows behind the helicase toward the free phosphate end of the DNA, making a continuous new DNA strand. DNA polymerase # 2 on the other parental strand moves away from the helicase and makes only part of the new DNA strand.
85 10.4 What Are The Mechanisms Of DNA Replication? As the helicase continues to unwind more of the double helix, additional DNA polymerase (# 3, # 4, etc.) must land on this strand to synthesize more pieces of DNA. Therefore, DNA synthesis on the second parental strand is discontinuous.
 DNA polymerase #1 continues along the parental DNA strand")
86 10.4 What Are The Mechanisms Of DNA Replication? The mechanism of DNA replication, step (4) DNA polymerase #1 continues along the parental DNA strand DNA polymerase #2 leaves DNA polymerase #3 Fig. 10-5(4)
87 10.4 What Are The Mechanisms Of DNA Replication? Multiple DNA polymerases make many pieces of DNA of varying lengths that need to be tied together to form a single continuous DNA polymer. DNA ligase joins together the separate segments of DNA.
 DNA polymerase #3 leaves DNA polymerase #4 DNA ligase")
88 10.4 What Are The Mechanisms Of DNA Replication? The mechanism of DNA replication, step (5) DNA polymerase #3 leaves DNA polymerase #4 DNA ligase joins the daughter DNA strands together Fig. 10-5(5)
89 10.4 What Are The Mechanisms Of DNA Replication? Proofreading produces almost error-free replication of DNA. DNA polymerase is almost 100% perfect in matching free nucleotides with those on the original parental strands. Once in every 10,000 base pairs, there is an error in replication. Some types of DNA polymerase recognize errors when they are made and correct them. This keeps the total errors in a complete DNA molecule to one mistake in every billion base pairs. Mistakes that remain in the DNA nucleotide sequence are called mutations.
90 Types of Mutations Point mutation - individual nucleotide in the DNA sequence is changed Insertion mutation - one or more nucleotide pairs are inserted into the DNA double helix Deletion mutation - one or more nucleotide pairs are removed from the double helix Inversion - piece of DNA is cut out of a chromosome, turned around, and re-inserted into the gap Translocation - chunk of DNA (often very large) is removed from one chromosome and attached to another
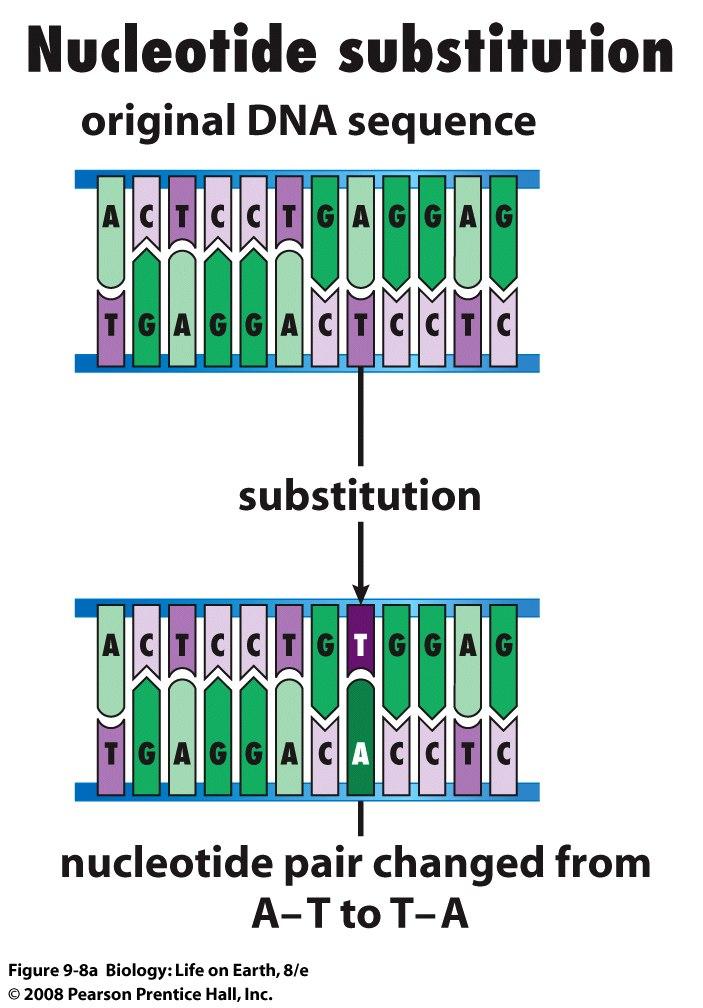
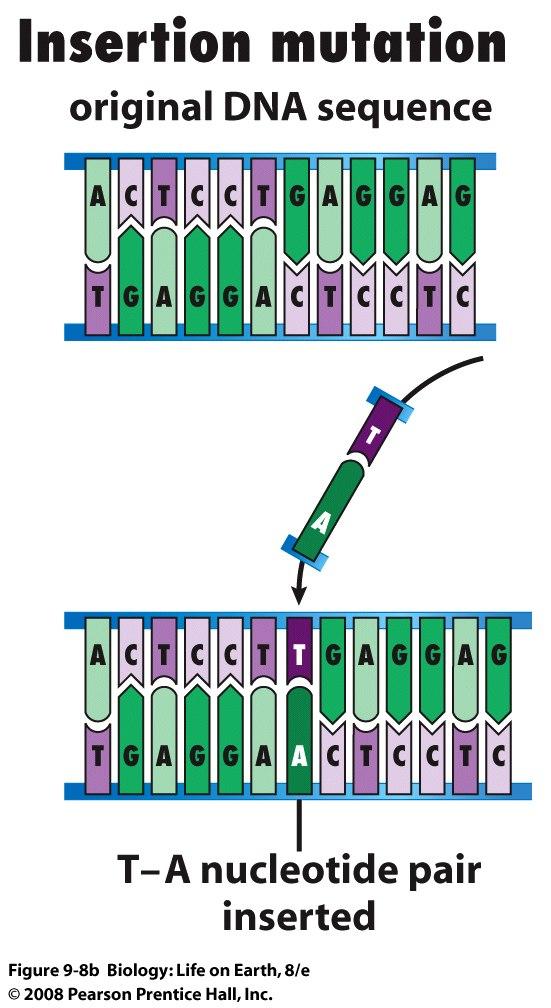
91
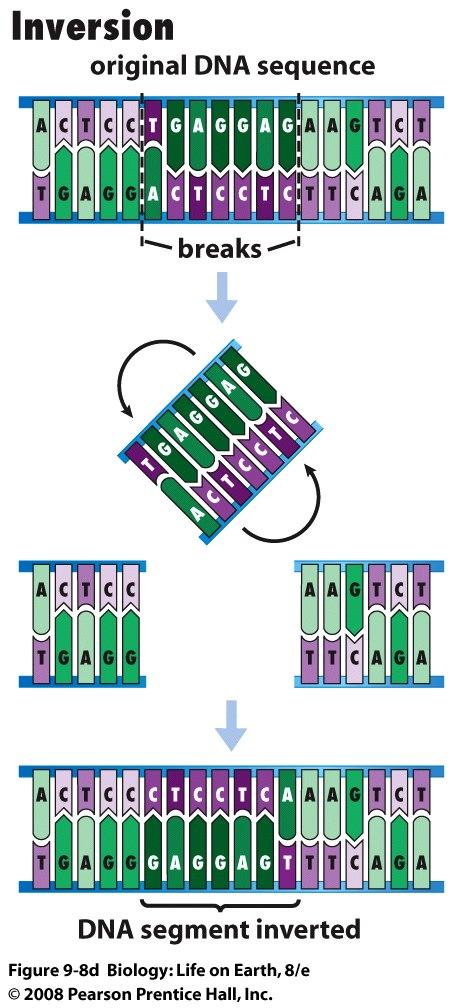
92
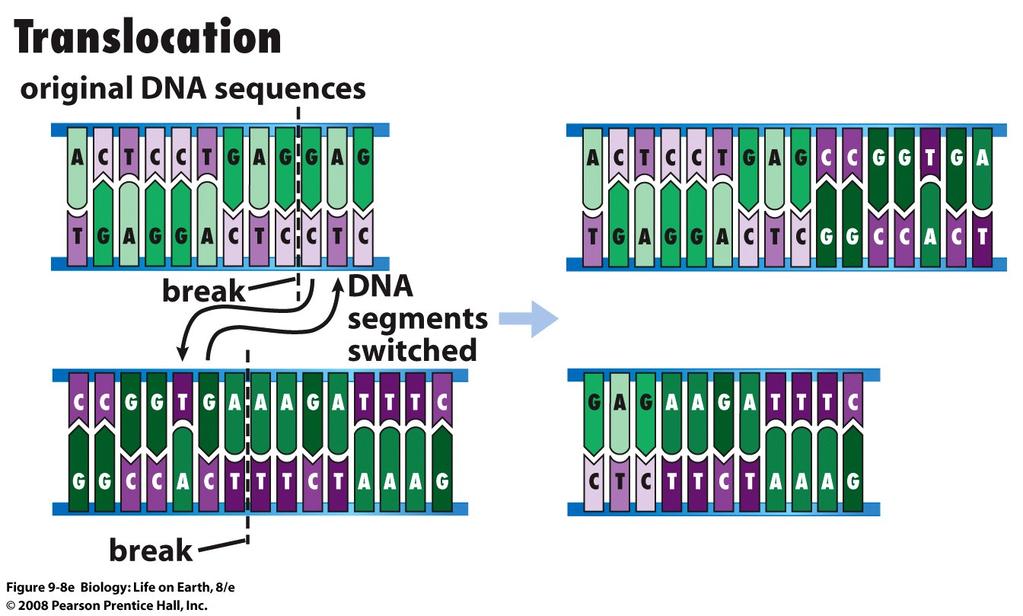
93
94 11.1 How Is The Information In DNA Used In A Cell? Most genes contain information for the synthesis of a single protein. A gene is a stretch of DNA encoding the instructions for the synthesis of a single protein. Proteins form cellular structures and the enzymes that catalyze cellular chemical reactions.

95 11.1 How Is The Information In DNA Used In A Cell?
 Transcription messenger RNA DNA (nucleus) (cytoplasm) Transcription of the gene produces an mrna with a nucleotide sequence complementary to one of the DNA strands ribosome Translation of")
96 11.1 How Is The Information In DNA Used In A Cell? Protein synthesis occurs in two steps, called transcription and translation. gene (a) Transcription messenger RNA DNA (nucleus) (cytoplasm) Transcription of the gene produces an mrna with a nucleotide sequence complementary to one of the DNA strands ribosome Translation of the mrna produces a protein molecule with an amino acid sequence determined by the nucleotide sequence in the mrna (b) Translation protein
97 11.1 How Is The Information In DNA Used In A Cell? Transcription: the information contained in the DNA of a specific gene is copied into one of three types of RNA Messenger RNA (mrna) Transfer RNA (trna) Ribosomal RNA (rrna) In eukaryotic cells, transcription occurs in the nucleus. Flash
98 11.1 How Is The Information In DNA Used In A Cell? Translation: ribosomes convert the base sequence in mrna to the amino acid sequence of a protein In eukaryotic cells, translation occurs in the cytoplasm. Flash
99 11.2 What Are The Functions Of RNA? Messenger RNA carries the code for a protein from the nucleus to the cytoplasm. All RNA is produced by transcription from DNA, but only mrna carries the code for amino acid sequence of a protein. mrna is synthesized in the nucleus and enters the cytoplasm through nuclear envelope pores. In the cytoplasm, mrna binds to ribosomes, which synthesize a protein specified by the mrna base sequence; DNA remains in the nucleus.
100 11.2 What Are The Functions Of RNA? Ribosomal RNA and proteins form ribosomes. Each ribosome consists of two subunits one small and one large. The small subunit has binding sites for mrna, a start trna, and other proteins that cooperate to read mrna to start protein synthesis. The large subunit has two binding sites for trna molecules, and one catalytic site where peptide bonds join amino acids together into a protein. During protein synthesis, the two subunits come together, clasping an mrna molecule between them.
101 11.2 What Are The Functions Of RNA? Transfer RNA molecules carry amino acids to the ribosomes. Each cell synthesizes many different kinds of transfer RNA, one or more for each amino acid. Twenty different kinds of enzymes in the cytoplasm, one for each amino acid, recognize the rrna and attach the correct amino acid. These loaded trna molecules deliver their amino acids to the ribosome, where they are incorporated into the growing protein chain.
102 11.3 What Is The Genetic Code? The genetic code translates the sequence of bases in nucleic acids into the sequence of amino acids in proteins. A sequence of three bases codes for an amino acid; the triplet is called a codon. There are 64 possible combinations of codons, which is more than enough to code for the 20 amino acids in proteins.

103 11.3 What Is The Genetic Code?
104 11.3 What Is The Genetic Code? How does a cell recognize where codons start and stop, and where the code for an entire proteins starts and stops? Most codons specify a specific amino acid in a protein sequence, but others are punctuation marks that indicate the end of one protein sequence and the start of another. All proteins begin with the start codon AUG (methionine), and all end with UAG, UAA, or UGA, called stop codons. Almost all amino acids are coded for by more than one codon (e.g., six codons code for leucine).
105 11.4 How Is The Information In A Gene Transcribed Into RNA? Transcription copies the genetic information of DNA into RNA in the nucleus of eukaryotic cells. Transcription is made up of three different processes: Initiation: the promotor region at the beginning of a gene starts transcription Elongation: the main body of a gene is where the RNA strand is elongated Termination: the termination signal at end of a gene is where RNA synthesis stops
106 11.4 How Is The Information In A Gene Transcribed Into RNA? Transcription begins when RNA polymerase binds to the promotor of a gene. RNA polymerase catalyzes the transcription of DNA to RNA. RNA polymerase first finds the promoter region (a non-transcribed sequence of DNA bases) that marks the start of a gene, and then binds to it, opening up the DNA as it does. Transcription of the gene begins after the promoter is bound to RNA polymerase.
107 11.4 How Is The Information In A Gene Transcribed Into RNA? Elongation generates a growing strand of RNA. RNA polymerase adds complementary bases to those in the DNA template strand, to make a growing RNA strand that has uracil rather than thymine complementary to adenine. The two strands of DNA re-form the original double helix. One end of the growing RNA strand drifts away from the DNA molecule, while the other remains attached to the DNA template strand by the RNA polymerase.
108 11.4 How Is The Information In A Gene Transcribed Into RNA? Transcription stops when RNA polymerase reaches the termination signal. RNA polymerase continues along the DNA template strand until it comes to the termination signal (a specific sequence of DNA bases). At the termination signal, RNA polymerase drops off the DNA and releases the completed RNA molecule. The enzyme is ready to bind to another promoter, to start the process over.
109 11.4 How Is The Information In A Gene Transcribed Into RNA? Transcription is selective. Some genes are transcribed in all cells because they encode essential proteins, like the electron transport chain of mitochondria. Other genes are transcribed only in specific types of cells. Proteins bind to control regions near gene promotors and block or enhance the binding of RNA polymerase. By this means, the amount of a specific protein encoded by a specific gene in a cell can be controlled.
110 11.5 How Is The Information In Messenger RNA Translated Into Protein? mrna, with a specific base sequence, is used during translation to direct the synthesis of a protein with the amino acid sequence encoded by the mrna. Decoding the base sequence of mrna is the job of trna and ribosomes in the cytoplasm. The ability of trna to deliver the correct amino acid to the ribosomes depends on base pairing between each codon of mrna and a set of three complementary bases in trna, called the anticodon.
111 11.5 How Is The Information In Messenger RNA Translated Into Protein? Like transcription, translation has three steps: Initiation of protein synthesis Elongation of the protein chain Termination of translation

112 11.5 How Is The Information In Messenger RNA Translated Into Protein? Initiation Initiation: initiation complex met U A C amino acid small ribosomal subunit methionine trna A trna with an attached methionine amino acid binds to a small ribosomal subunit, forming an initiation complex. mrna trna met U A C anticodon G C A U G G U U C A start codon The initiation complex binds to an mrna molecule. The methionine (met) trna anticodon (UAC) base-pairs with the start codon (AUG) of the mrna. catalytic site first trna binding site second trna binding site met U A C G C A U G G U U C A large ribosomal subunit The large ribosomal subunit binds to the small subunit. The methionine trna binds to the first trna site on the large subunit. Fig. 11-5(1,2,3)

113 11.5 How Is The Information In Messenger RNA Translated Into Protein? Elongation Elongation: met val catalytic site met val peptide bond initiator trna detaches met val U A C C A A U A C C A A C A A G C A U G G U U C A G C A U G G U U C A G C A U G G U U C A U A G The second codon of mrna (GUU) base-pairs with the anticodon (CAA) of a second trna carrying the amino acid valine (val). This trna binds to the second trna site on the large subunit. The catalytic site on the large subunit catalyzes the formation of a peptide bond linking the amino acids methionine and valine. The two amino acids are now attached to the trna in the second binding site. ribosome moves one codon to the right The empty trna is released and the ribosome moves down the mrna, one codon to the right. The trna that is attached to the two amino acids is now in the first trna binding site and the second trna binding site is empty. Fig. 11-5(4,5,6)
 met val his met val his C A A G U A C A A G U A G C A U G G U U C A U A G G C A U G G U U C A U A G The third codon of mrna (CAU) base-pairs with the anticodon (GUA) of a trna")
114 11.5 How Is The Information In Messenger RNA Translated Into Protein? Elongation (continued) met val his met val his C A A G U A C A A G U A G C A U G G U U C A U A G G C A U G G U U C A U A G The third codon of mrna (CAU) base-pairs with the anticodon (GUA) of a trna carrying the amino acid histidine (his). This trna enters the second trna binding site on the large subunit. The catalytic site forms a peptide bond between valine and histidine, leaving the peptide attached to the trna in the second binding site. The trna in the first site leaves, and the ribosome moves one codon over on the mrna. Fig. 11-5(7,8)

115 11.5 How Is The Information In Messenger RNA Translated Into Protein? Termination Termination: met val completed peptide his arg arg ile stop codon C G A A U C U A G U A A This process repeats until a stop codon is reached; the mrna and the completed peptide are released from the ribosome, and the subunits separate. Fig. 11-5(9)
116 11.5 How Is The Information In Messenger RNA Translated Into Protein? Summing up: transcription and translation With a few exceptions, each gene codes for a single protein. Transcription of a protein-coding gene produces an mrna that is complementary to the template strand of the DNA for the gene. Enzymes in the cytoplasm attach the appropriate amino acid to each trna. The mrna moves from the nucleus to the cytoplasm. trnas carry their attached amino acids to the ribosome.
117 11.5 How Is The Information In Messenger RNA Translated Into Protein? Summing up: transcription and translation (continued) At the ribosome, the bases in trna anticodons bind to the complementary bases in mrna codons. The amino acids attached to the trnas line up in the sequence specified by the codons. The ribosome joins the amino acids together with peptide bonds to form a protein. When a stop codon is reached, the finished protein is released from the ribosome.
 DNA complementary DNA strand template DNA strand gene A T G G G A G T T T A C C C T C A A codons etc. etc. (b) mrna A U G G G A G U U etc. anticodons (c) trna U A C C C U C A A etc.")
118 11.5 How Is The Information In Messenger RNA Translated Into Protein? Complementary base pairing is critical to decoding genetic information. (a) DNA complementary DNA strand template DNA strand gene A T G G G A G T T T A C C C T C A A codons etc. etc. (b) mrna A U G G G A G U U etc. anticodons (c) trna U A C C C U C A A etc. amino acids (d) protein methionine glycine valine etc. Fig. 11-6
119 11.6 How Do Mutations Affect Gene Function? Mutations are the raw material for evolution. Mutations are the ultimate source of all genetic differences among individuals. Without mutations, individuals would share the same DNA sequence. Most mutations are harmful; some improve the individual s ability to survive and reproduce. The mutation may be passed from generation to generation and become more common over time. This process is known as natural selection, and is the major cause of evolutionary change.
120 11.7 Are All Genes Expressed? All of the genes in the human genome are present in each body cell, but individual cells express only a small fraction of them. The particular set of genes that is expressed depends on the type of cell and the needs of the organism. This regulation of gene expression is crucial for proper functioning of individual cells and entire organisms.
121 11.7 Are All Genes Expressed? Gene expression differs from cell to cell and over time. The set of genes that are expressed depends on the function of a particular cell. Hair cells synthesize the protein keratin, while muscle cells make the proteins actin and myosin but do not make keratin. A human male does not express a casein gene, the protein in human milk, but will pass on the gene for casein synthesis to his daughter, who will express it if she bears children.
122 11.7 Are All Genes Expressed? Environmental cues influence gene expression. Changes in an organism s environment help determine which genes are transcribed. Longer spring days stimulate the sex organs of birds to enlarge and produce sex hormones. These hormones cause the birds to produce eggs and sperm, to mate, and to build nests. All these changes result directly or indirectly from alterations in gene expression.
123 11.8 How Is Gene Expression Regulated? A cell may regulate gene expression in many different ways. It may alter the rate of transcription of mrna. It may affect how long a given mrna molecule lasts before being broken down. It may affect how fast the mrna is translated into protein. It may affect how long the protein lasts, or how fast a protein enzyme catalyzes a reaction.
124 11.8 How Is Gene Expression Regulated? Regulatory proteins that bind to promoters alter the transcription of genes. Many steroid hormones act in this way. In birds, estrogen enters cells of the female reproductive system and binds to a receptor protein during the breeding season. The estrogen receptor combination then binds to the DNA in a region near the promotor of an albumen gene.
125 11.8 How Is Gene Expression Regulated? Regulatory proteins that bind to promoters alter the transcription of genes (continued). This attachment makes it easier for RNA polymerase to bind to the promotor and to transcribe large amounts of albumen mrna, which is translated into the albumin protein needed to make eggs.
126 11.8 How Is Gene Expression Regulated? Some regions of chromosomes are condensed and not normally transcribed. Certain parts of eukaryotic chromosomes are in a highly condensed, compact state in which most of the DNA is inaccessible to RNA polymerase. Some of these tightly condensed regions may contain genes that are not currently being transcribed, but when those genes are needed, the portion of the chromosome containing those genes becomes decondensed so that transcription can occur.
127 11.8 How Is Gene Expression Regulated? Entire chromosomes may be inactivated and not transcribed. In some cases, almost an entire chromosome may be condensed, making it largely inaccessible to RNA polymerase. In human females, one of their two X chromosomes may become inactivated by a special coating of RNA called Xist, which condenses the chromosome and prevents gene transcription.