Replication of DNA and Chromosomes
|
|
|
- Suzan Potter
- 5 years ago
- Views:
Transcription
1 Chapter 9. Replication of DNA and Chromosomes 1. Semiconservative Replication 2. DNA Polymerases and DNA Synthesis In Vitro 3. The Complex Replication Apparatus 4. Unique Aspects of Eukaryotic Chromosome Replication 1

2 Seven pairs of identical twins: No instructions regarding how to pose. 2
3 People often explain nearly identical phenotypes of monozygotic twins like Merry and Sherry by stating that they contain the same genes. Of course, that is not true. To be accurate, the statement should be that identical twins contain progeny replicas of the same parental genes. A human life emerges from a single cell, a tiny sphere about 0.1 mm in diameter. 65 trillion (65,000,000,000,000) cells. 3

4 1. Semiconservative Replication Each of the parental strands is conserved and serves as a template for the synthesis of a new complementary strand; that is, the base sequence in each progeny strand is determined by the hydrogen-bonding potentials of the bases in the parental strand. 4

5 The chromosomal DNA of the broad bean, Vicia faba, replicates by a semiconservative mechanism. 5
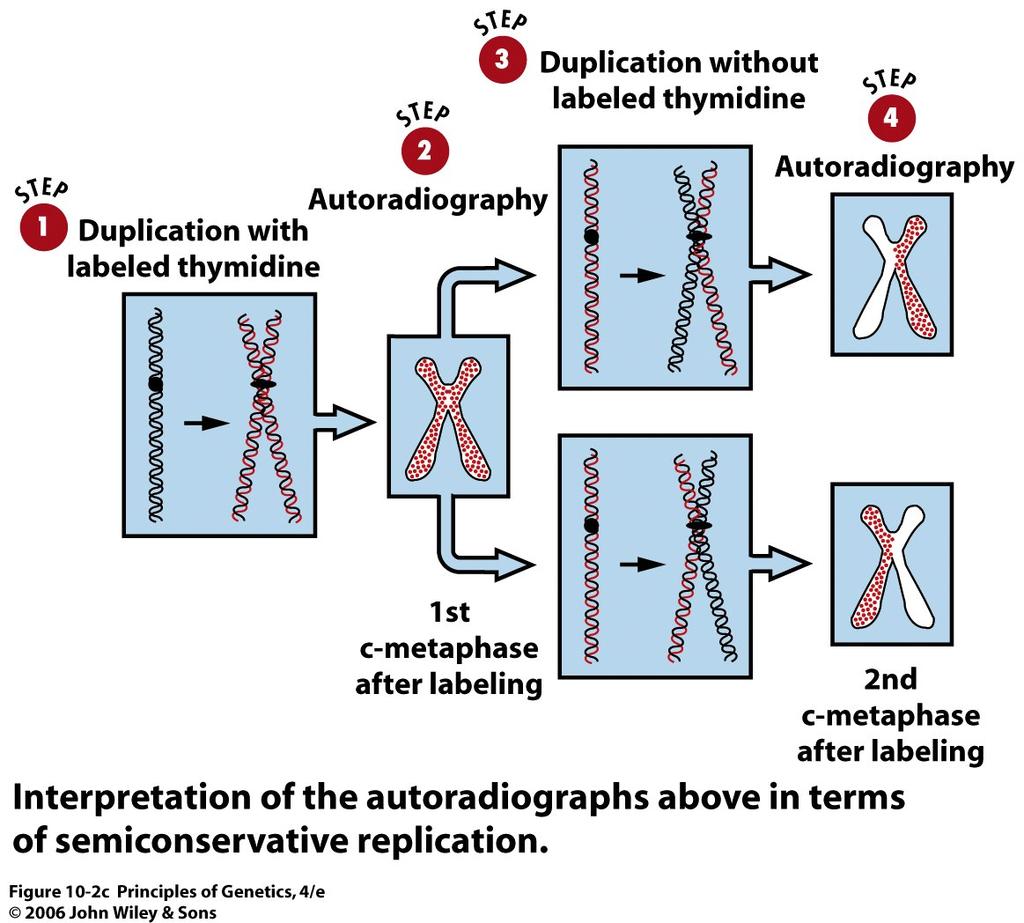
6 6

7 θ-shaped replicating chromosome 7
8 8
9 Unique Origins of Replication Cairns' results established the existence of a site of initiation or origin of replication on the circular chromosome of E. coli but provided no hint as to whether the origin was a unique site or occurred at randomly located sites in a population of replicating chromosomes. The single origin of replication, called oric, in the E. coli chromosome has been characterized in considerable detail. oric is 245 nucleotide pairs long and contains two different conserved repeat sequences 9
10 One 13-bp sequence is present as three tandem repeats. These three repeats are rich in A:T base pairs, facilitating the formation of a localized region of strand separation referred to as the replication bubble. 10
11 Another conserved component of oric is a 9-bp sequence that is repeated four times and is interspersed with other sequences. These four sequences are binding sites for a protein that plays a key role in the formation of the replication bubble. 11
12 In the yeast Saccharomyces cerevisiae, segments of chromosomal DNA that allow a fragment of circularized DNA to replicate as an independent unit (autonomously), that is, as an extrachromosomal self-replicating unit, have been identified and characterized. These sequences are called ARS (for Autonomously Replicating Sequences) elements. where Pu is either of the two purines and Py is either of the two pyrimidines 12
13 The core origin of replication in simian virus 40 (SV40). The core origin is the minimal sequence that can initiate replication in vivo. Palindromes. They are often the binding sites for proteins that interact with DNA, such as the large T antigen of SV40, which is the first protein to bind to the core origin during the initiation of SV40 replication. 13
14 Bidirectional Replication The phage λ chromosome is somewhat unusual in that it has a single-stranded region, 12 nucleotides long, at the 5 end of each complementary strand These single-stranded ends, called cohesive or sticky ends, are complementary to each other. The cohesive ends of a lambda chromosome can thus base-pair to form a hydrogen-bonded circular structure. One of the first events to occur after a lambda chromosome is injected into a host cell is its conversion to a covalently closed circular molecule. 14
15 15
16 The feature of the lambda chromosome that facilitated the demonstration of bidirectional replication is its differentiation into regions containing high concentrations of adenine and thymine (A:T-rich regions) and regions with large amounts of guanine and cytosine (G:C-rich regions). 16
17 17
18 18
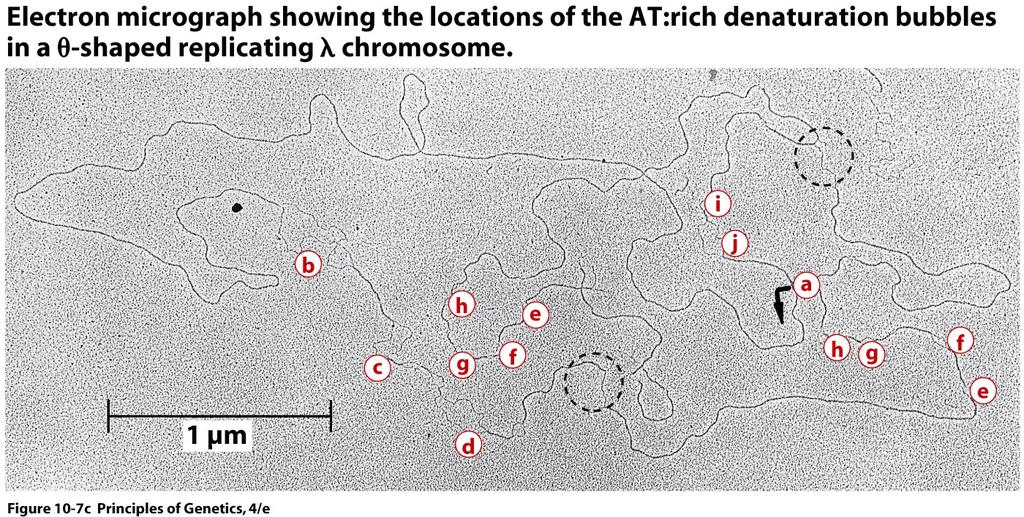
19 19
20 20
21 21

22 Electron micrographs of replicating bacteriophage T7 chromosomes. The phage T7 chromosome, unlike the E. coli and phage λ chromosomes, replicates as a linear structure. 22

23 Replication proceeds bidirectionally from the origin until the fork moving in a leftward direction reaches the left end of the molecule, yielding a Y-shaped structure such as that 23 shown in (b).
24 2. DNA Polymerases and DNA Synthesis In Vitro In vitro, in vivo DNA polymerase I The in vitro synthesis of DNA was first accomplished by Arthur Kornberg and his coworkers in Kornberg, who received a Nobel Prize in 1959 for this work, isolated an enzyme from E. coli that catalyzes the covalent addition of nucleotides to preexisting DNA chains. 24
25 The enzyme requires the 5 -triphosphates of each of the four deoxyribonucleosides deoxyadenosine triphosphate (datp), deoxythymidine triphosphate (dttp), deoxyguanosine triphosphate (dgtp), and deoxycytidine triphosphate (dctp) and is active only in the presence of Mg 2+ ions and preexisting DNA. 25
26 The primer DNA provides a terminus with a free 3 -OH to which nucleotides are added during DNA synthesis. The template DNA provides the nucleotide sequence that specifies the complementary sequence of the nascent DNA chain. 26
27 27
28 28
29 DNA polymerase I has two other enzymatic activities, both exonuclease activities. A nuclease is an enzyme that degrades nucleic acids. An exonuclease degrades nucleic acids starting at one or both ends, whereas an endonuclease cleaves nucleic acids at internal sites. DNA polymerase I contains both 5 3 exonuclease activity, which cuts back DNA strands starting at 5 termini, and 3 5 exonuclease activity, which cleaves off mononucleotides from the 3 termini of DNA strands. 29
30 The 5 3 exonuclease activity of DNA polymerase I usually excises small oligomers containing up to 10 nucleotides. Thus, DNA polymerase I has three different enzymatic activities: (1) a 5 3 polymerase activity, (2) a 5 3 exonuclease activity, and (3) a 3 5 exonuclease activity. 30
31 31
32 32
33 33
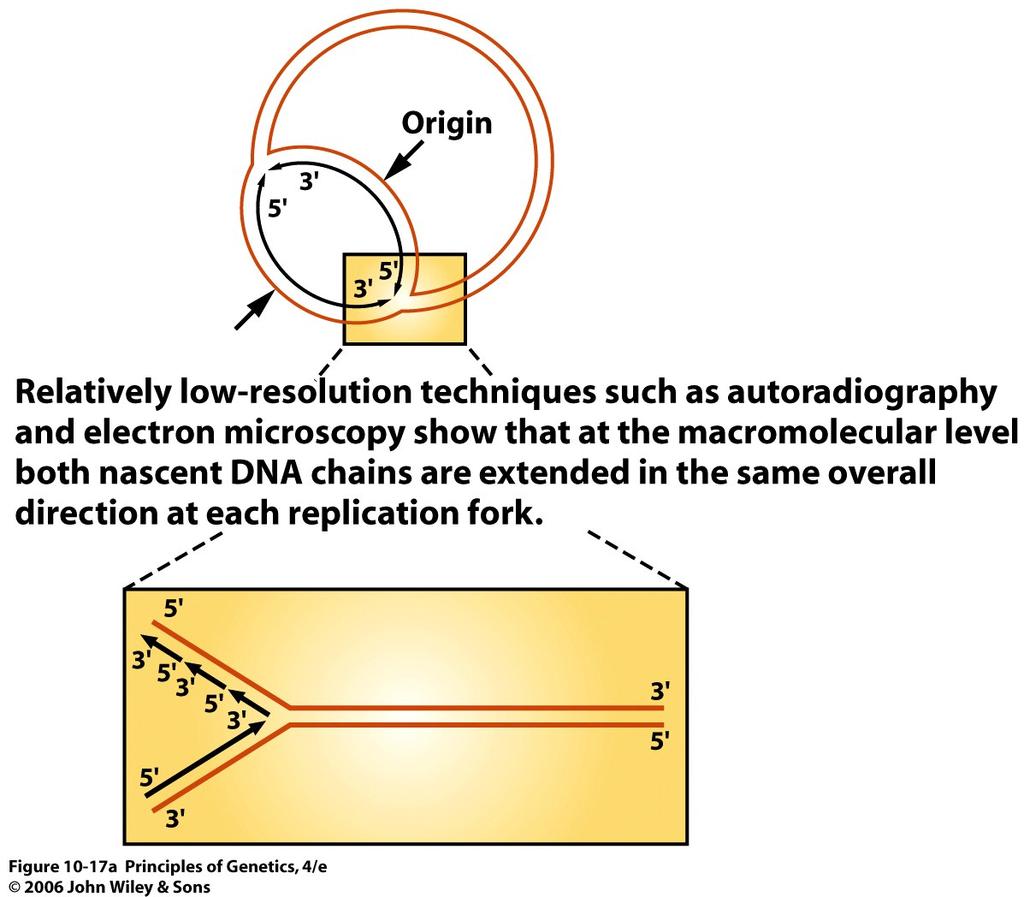
34 DNA polymerase I is not the true DNA replicase in E. coli. It does not catalyze the semiconservative replication of the E. coli chromosome; that function is performed by another enzyme. Nevertheless, DNA polymerase I does perform important functions in the E. coli cell, including playing a key role in chromosome replication and a central role in repairing damaged DNA. 34
35 Multiple DNA Polymerases There are at least four other DNA polymerases, DNA polymerase II, DNA polymerase III, DNA polymerase IV, and DNA polymerase V, in E. coli. Like DNA polymerase I, DNA polymerase II is a DNA repair enzyme; but it represents a small proportion of the polymerase activity in an E. coli cell. DNA polymerase II is a single polypeptide with 5 3 polymerase and 3 5 exonuclease activities. However, it has no 5 3 exonuclease activity. 35
36 In contrast to DNA polymerases I and II, DNA polymerase III is a complex enzyme composed of many different subunits. Like DNA polymerase II, DNA polymerase III has 5 3 polymerase and 3 5 exonuclease activities; however, it has a 5 3 exonuclease that is active only on single-stranded DNA. The more recently characterized DNA polymerases IV and V, along with polymerase II, play important roles in the replication of damaged DNA, with the polymerase involved depending on the type of damage. 36
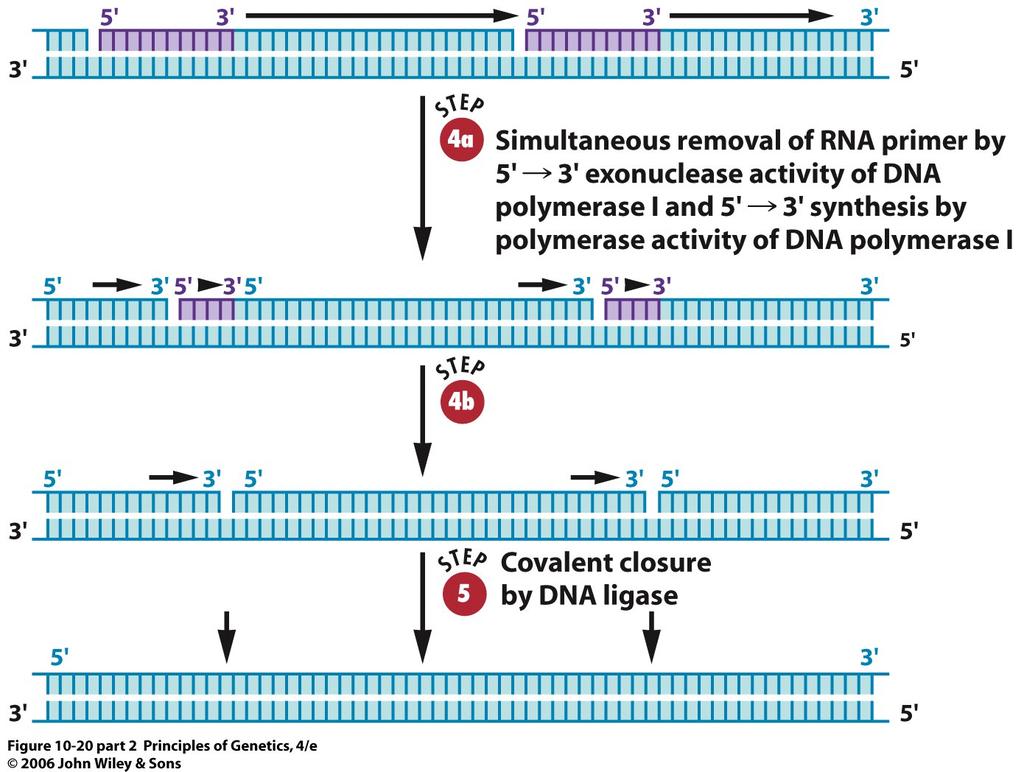
37 DNA Polymerase III: The Replicase in Escherichia coli DNA polymerase III is the true DNA replicase responsible for the semiconservative replication. polc, now renamed dnae. This mutant strain produced active DNA polymerase III when grown at 25 C, but totally inactive polymerase III when grown at 43 C. When dnae mutant cells growing at 25 C were shifted to 43 C, DNA replication stopped, indicating that the product of the dnae gene is required for DNA synthesis. 37
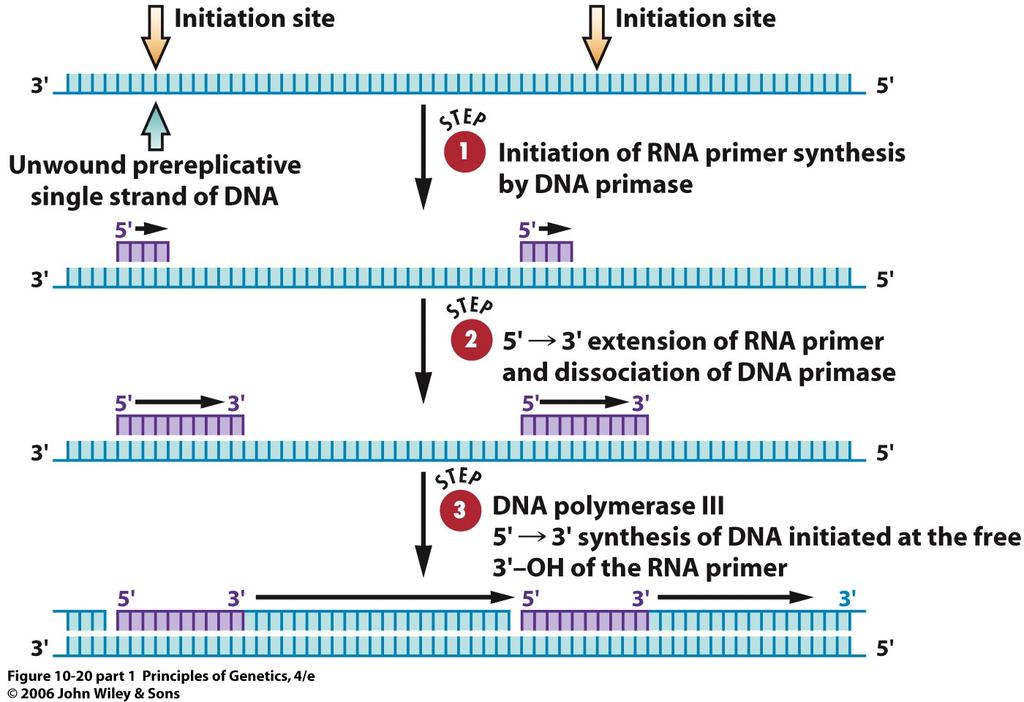
38 Proofreading Activities of DNA Polymerases The fidelity of DNA duplication is amazing with only about one error present in every billion base pairs shortly after synthesis. This high fidelity is necessary to keep the mutation load at a tolerable level, especially in large genomes such as those of mammals, which contain nucleotide pairs. One error per 10,000 to 100,000 incorporated nucleotides. Without the high fidelity of DNA replication, the monozygotic twins discussed at the beginning of this chapter would be less similar in phenotype. 38
39 Living organisms have solved the potential problem of insufficient fidelity during DNA replication by evolving a mechanism for proofreading the nascent DNA chain as it is being synthesized. The proofreading process involves scanning the termini of nascent DNA chains for errors and correcting them. This process is carried out by the 3 5 exonuclease activities of DNA polymerases. When a template-primer DNA has a terminal mismatch (an unpaired or incorrectly paired base or sequence of bases at the 3 end of the primer), the 3 5 exonuclease activity of the DNA polymerase clips off the unpaired base or bases 39
40 40
41 3. The Complex Replication Apparatus -DNA replication is a complex process, requiring the concerted action of a large number of proteins. -Continuous synthesis of One Strand; Discontinuous Synthesis of the Other Strand 41
42 42

43 Leading strand-continuous Lagging strand 43
44 Covalent Closure of Nicks in DNA by DNA Ligase DNA ligase catalyzes the covalent closure of nicks in DNA. The energy required to form the ester linkage is provided by either adenosine triphosphate (ATP) or nicotinamide-adenine dinucleotide (NAD), depending on the species. 44
45 Initiation of DNA Chains with RNA Primers All known DNA polymerases have absolute requirements for a free 3 -OH on a DNA primer strand and an appropriate DNA template strand for activity. No known DNA polymerase can initiate the synthesis of a new strand of DNA. Thus, some special mechanism must exist to initiate or prime new DNA chains. Whereas the continuous synthesis of the leading strand requires the priming function only at the origin of replication, a priming event is required to initiate each Okazaki fragment during the discontinuous synthesis of the lagging strand. 45

46 Subsequent research has shown that each new DNA chain is initiated by a short RNA primer synthesized by DNA primase 46
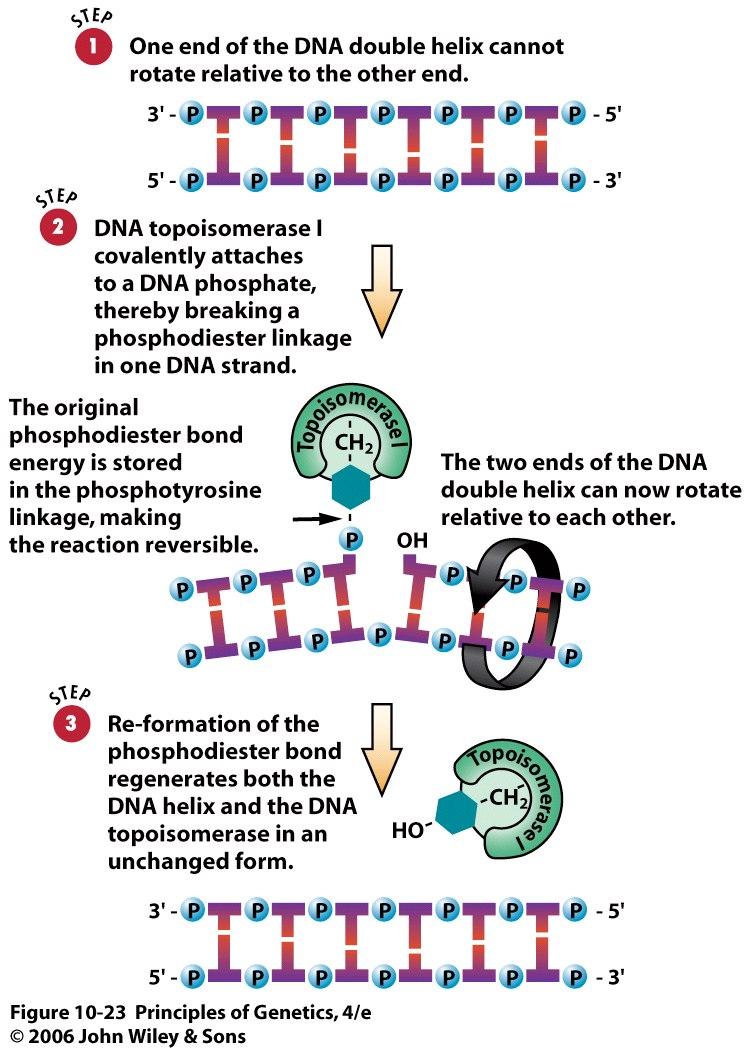
47 The RNA primers subsequently are excised and replaced with DNA chains. This step is accomplished by DNA polymerase I in E. coli. Recall that of the five DNA polymerases in E. coli, only DNA polymerase I possesses a 5 3 exonuclease that acts on double-stranded DNA. The 5 3 exonuclease activity of DNA polymerase I excises the RNA primer, and, at the same time, the 5 3 polymerase activity of the enzyme replaces the RNA with a DNA chain by using the adjacent Okazaki fragment with its free 3 -OH as a primer. 47
48 48
49 49
50 Unwinding DNA with Helicases, DNA-Binding Proteins, and Topoisomerases 50
51 51
52 Replicate 30,000 nucleotides per minute. 52
53 53
54 The topoisomerases are of two types: (1) DNA topoisomerase I enzymes produce temporary single-strand breaks or nicks in DNA, and (2) DNA topoisomerase II enzymes produce transient double-strand breaks in DNA. An important result of this difference is that topoisomerase I activities remove supercoils from DNA one at a time, whereas topoisomerase II enzymes remove and introduce supercoils two at a time. 54
55 55
56 56
57 The Replication Apparatus: Prepriming Proteins, Primosomes, and Replisomes Primosome: A protein replication complex that catalyzes the initiation of Okazaki fragments during discontinuous synthesis. It contains DNA primase and DNA helicase activities. Replisome: The complete replication apparatus present at a replication fork that carries out the semiconservative replication of DNA. 57
58 Rolling-Circle Replication Another important type of DNA replication called rolling-circle replication. Rolling-circle replication is used (1) by many viruses to duplicate their genomes, (2) in bacteria to transfer DNA from donor cells to recipient cells during one type of genetic exchange, and (3) in amphibians to amplify extrachromosomal DNAs carrying clusters of ribosomal RNA genes during oogenesis 58
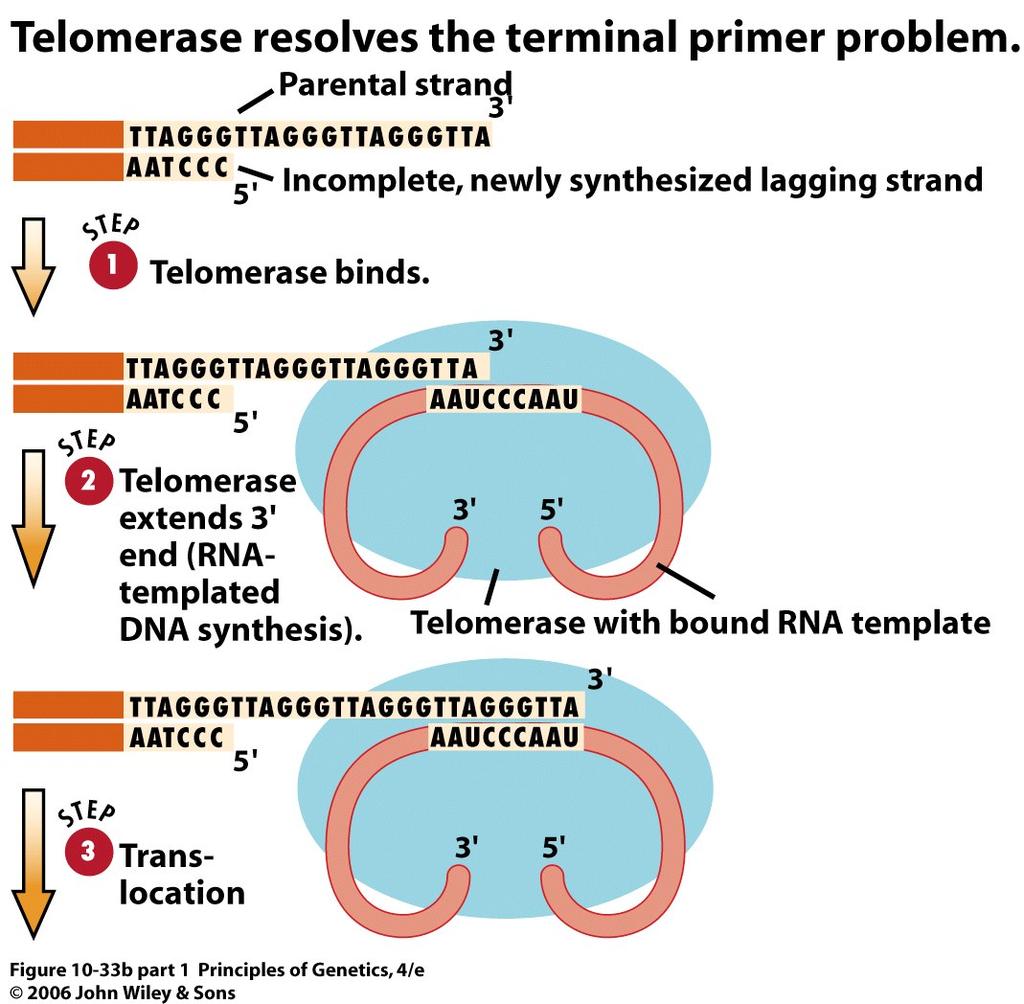
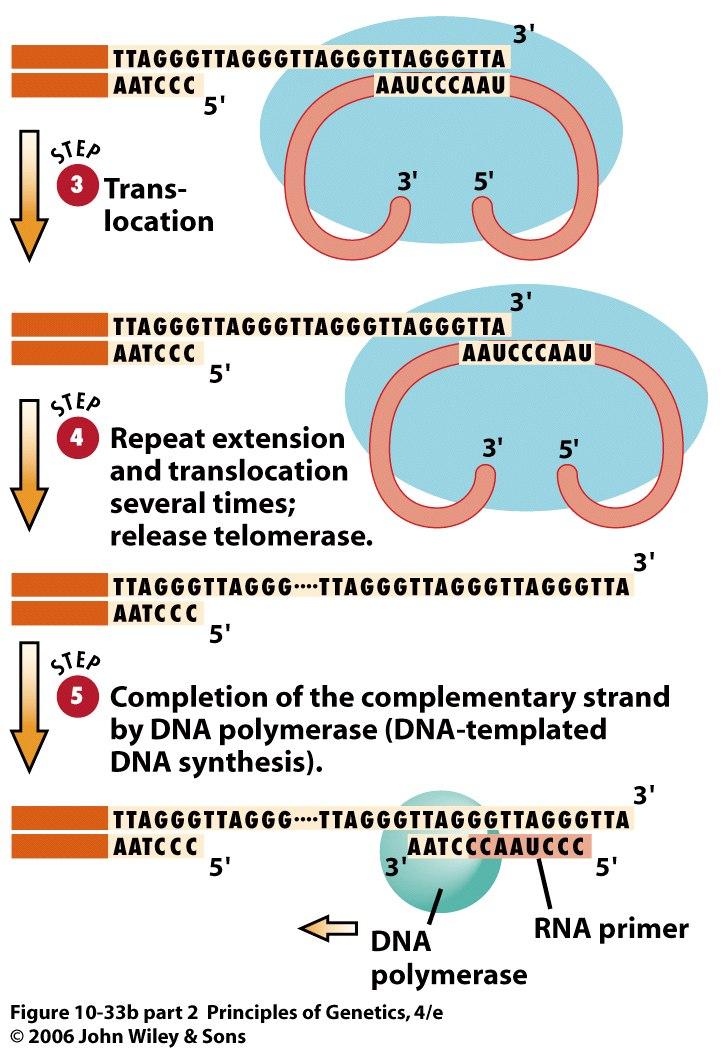
59 59
60 60
61 4. Unique Aspects of Eukaryotic Chromosome Replication Although the main features of DNA replication are the same in all organisms, some processes occur only in eukaryotes. A segment of DNA whose replication is under the control of one origin and two termini is called a replicon. In prokaryotes, the entire chromosome is usually one replicon. 61
62 The giant DNA molecules in the largest chromosomes of Drosophila melanogaster contain about nucleotide pairs. The rate of DNA replication in Drosophila is about 2600 nucleotide pairs per minute at 25 C. A single replication fork would therefore take about 17.5 days to replicate one of these giant DNA molecules. With two replication forks moving bidirectionally from a central origin, such a DNA molecule could be replicated in just over 8.5 days. Given that the chromosomes of Drosophila embryos replicate within 3 to 4 minutes require 7000 replication forks 62
63 Evidence for bidirectional replication of the multiple replicons in the giant DNA molecules of eukaryotes. 63
64 64
 are present in the segment of the DNA molecule shown.")
65 Electron micrograph of a DNA molecule in D. melanogaster showing multiple sites of replication. Four eye-shaped replication structures (labeled A-D) are present in the segment of the DNA molecule shown. The arrows show the positions of replication forks. 65
66 Telomerase: Replication of Chromosome Termini DNA polymerases cannot replicate the terminal DNA segment of the lagging strand of a linear chromosome. At the end of the DNA molecule being replicated discontinuously, there would be no DNA strand to provide a free 3 -OH (primer) for polymerization of deoxyribonucleotides after the RNA primer of the terminal Okazaki fragment has been excised 66
67 67
68 68
69 69
70 Telomere Length and Aging in Humans Unlike germ-line cells, most human somatic cells lack telomerase activity. When human somatic cells are grown in culture, they divide only a limited number of times (usually only 20 to 70 cell generations) before senescence and death occur. Cells with longer telomeres survive longer go through more cell divisions than cells with shorter telomeres. 70
71 Further evidence of a relationship between telomere length and aging in humans has come from studies of individuals with disorders called progerias, inherited diseases characterized by premature aging. In the most severe form of progeria, Hutchinson Gilford syndrome, senescence wrinkles, baldness, and other symptoms of aging begins immediately after birth, and death usually occurs in the teens. 71