Conformation of the Mineralocorticoid Receptor N- terminal Domain: Evidence for Induced and Stable Structure
|
|
|
- Adele Edwards
- 5 years ago
- Views:
Transcription
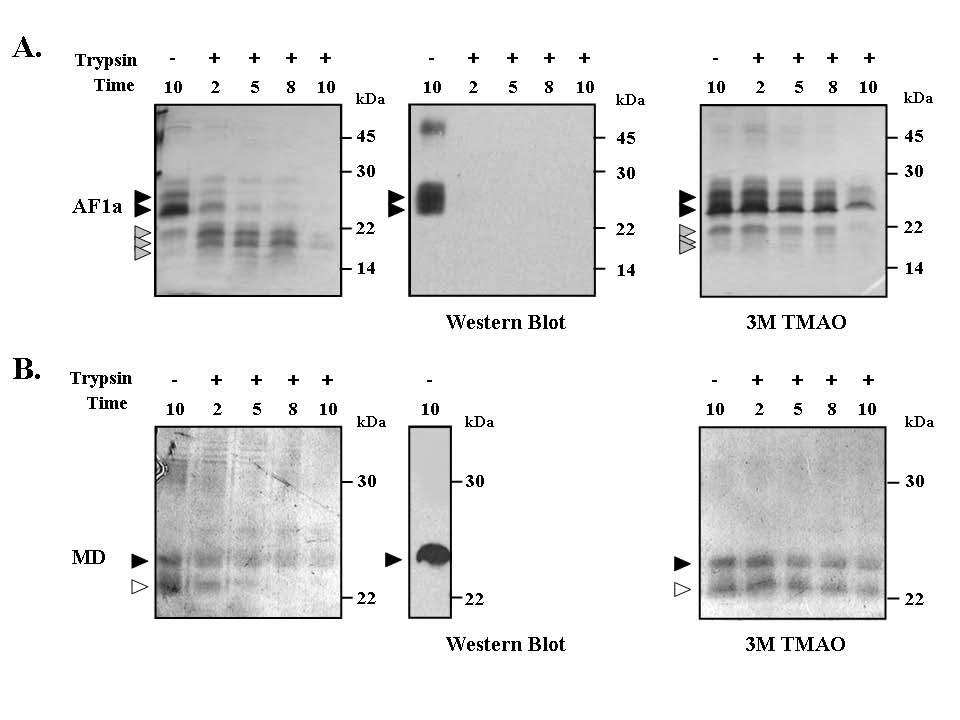
1 ME Conformation of the Mineralocorticoid Receptor N- terminal Domain: Evidence for Induced and Stable Structure Katharina Fischer 1, Sharon M. Kelly 2, Kate Watt 1, Nicholas C. Price 2 and Iain J. McEwan 1. 1 Institute of Medical Sciences, University of Aberdeen, Aberdeen AB25 2ZD, Scotland, UK. 2 Faculty of Biomedical and Life Sciences, University of Glasgow, Glasgow G12 8QQ, Scotland, UK Supplemental Figure Legends Supplemental Figure 1. Limited proteo lysis of MR-NTD transactivation domains. (A) Recombinant MR-AF1a was digested with 0.24 ng/µl trypsin for up to 10 minutes at 30 C in the absence (left panel) or presence (right panel) of 3 M TMAO. Experiments were performed in triplicate with 1 µm of protein, and representative data is shown. After digestion samples were resolved on 15% SDS polyacrylamide gel and subjected to silver staining. Two particularly stable fragments occurred at 20.5 and 16.9 kda after partial digest, in the absence of TMAO, which are marked by the grey filled arrows. These fragments were not detected using an anti-histidine tag antibody, suggesting that N-terminal sequences were lost (middle panel). From the size of these fragments and the theoretical cleavage pattern of the protein we can estimate that cleavage occurs around Arg 28 and/or 35. (B) Limited proteolysis of the MR-MD in the absence (left panel) or presence (right panel) of 3M TMAO. The polypeptide was digested with trypsin for the time periods indicated and the fragments resolved by SDS-PAGE and detected by silver staining. The open arrow head indicates a degradation product of MR-MD, which is not detected with an anti-histidine tag antibody (middle panel). In the presence of TMAO MR-MD, and the degradation product, were protected from digestion. The results are representative of two experiments. (C) MR-AF1b, stabilised in SDS, was digested at 1 µm with 0.24 ng/µl trypsin over time course indicated. Digested proteins were resolved on 15% polyacrylamide gels and
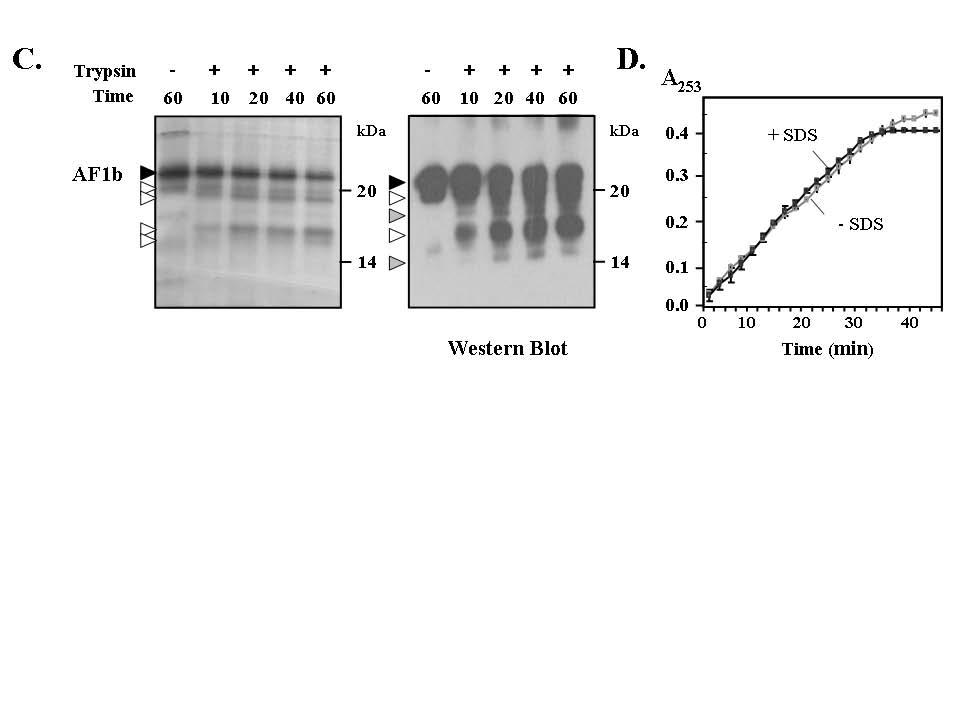
2 visualised by silver staining (left panel). Arrow heads indicate particularly stable fragments, at 17.1 kda fragment and 19.2 kda fragments; grey shading represents fragments more clearly detected on the Western blot. Some of these fragments (open arrow heads) were detected by Western blot analysis using an anti-histidine tag antibody, indicating that these fragments must retain N-terminal sequences and it was calculated that cleavage occurs around Arg 537 and/or Arg 540. Experiments were repeated three times. (D) The artificial trypsin substrate Nαbenzoyl-L-arginine ethyl ester was digested with trypsin in the absence or presence of 0.05% SDS and the reactions monitored at 253 nm over time. These two curves are virtually superimposable over the time course of the experiment indicating SDS at this concentration does not inhibit trypsin activity and the protease resistance of MR-AF1b is a property of the folded polypeptide. Supplemental Figure 2. Conformati on analysis of the MR-NTD. (A) Steadystate fluorescence emission spectra for MR-NTD in the absence (black line) or presence of either TMAO (black dotted line) or urea (grey line) after excitation (Ex.) at 278 nm to follow emission of two tryptophan (W) residues at the very amino- and carboxy terminal end (see Main Text). Data are representative for five independent experiments and readings are corrected for the buffer. Between µg/ml of protein were used for each reading. Respective λmax values are summarized in Table 1. (B) Purified MR- NTD protein (~ 0.2 mg/ml) was analysed by far UV CD spectroscopy. Experiments were done in the presence or absence of the structure stabilising agent TFE (25 and 50%). Results are representative for two independent experiments. (C) MR-NTD (50 pmoles) was subjected to partial proteolysis with trypsin (5 ng/μl)for the time points indicated in the absence or presence of TMAO (left panel). In the absence of TMAO two strong fragments are observed with molecular weights of around 25 kda (open arrow heads). Interestingly, these fragments are not detected using an anti-histidine tag antibody (right panel), which is consistent with them lacking N-terminal sequences. In the presence of TMAO intermediate fragments (grey arrow heads) and full-length protein are less susceptible to cleavage and there is a reduction in the levels of two fragments (open arrow heads) seen in the absence of osmolyte. Collectively this
3 data is consistent with folding of the MR-NTD in the presence of TMAO and adoption of a more protease resistant conformation. Supplemental Figure 3. Effect of point mutations of conserved residues in AF1b on transactivation activity of MR-NTD-DBD. Triple or single point mutations were introduced into pcmr-ntd-dbd-flag or pet-mr-af1b plasmids using Quickchange (Stratagene). The mutant or wild-type MR-NTD-DBD-flag constructs were transfected into COS-1 cells together with the (GRE) 2 -TATAluciferase reporter gene and transactivation activity measured. Transfections were done in triplicate and the mean ± standard deviation is shown. Above is shown a Western blot of the expressed proteins detected with an anti-flag antibody; the *, indicates a non-specific protein detected in blots for the MR-NTD-FLAG constructs. The % activity relative to the wild-type protein (100 %) is also shown. Mutations introduced into the N-terminal half (Figure 8A) impaired transactivation (i.e. M5 to M10). Supplemental Figure 4. Binding of TBP to MR-AF1b does not dramatically change secondary structure con tent in the complex. Far UV CD spectra were recorded for a complex of AF1b TBP, at a ratio of 1:1 with 0.1 mg/ml of each protein in the presence of 0.05% SDS (grey line). The spectrum resulting from the addition of the individual spectra for each protein is also shown (black line). Results are representative of two independent experiments. The experimental and calculated spectra are almost superimposable, which would suggest there is no net conformational change upon complex formation.
4
5
6

7

8
9