Chapter 3 Gene Function. Transcription Prokaroyotes Eukaryotes Transcript processing Proteins Translation Genetic nomenclature
|
|
|
- Scarlett Craig
- 5 years ago
- Views:
Transcription

1 Chapter 3 Gene Function Transcription Prokaroyotes Eukaryotes Transcript processing Proteins Translation Genetic nomenclature

2 Transcription
3 RNA composition ATP, GTP, UTP, CTP are substrates for RNA polymerase. RNA is a ribonucleoside monophosphate polymer.
4 RNA types Informational: mrna, encode amino acids, ~ 1% of total cellular RNA Functional: trna, translation rrna, ribosome components; 5S, 5.8S, 18S, 28S; bulk snrna, splicing sirna, mirna inhibitory (regulatory) RNA other small stable RNAs, signal recognition particle, RNaseP ribozymes

5 Tandemly transcribed rrna genes (NO).
6 Transcription of a gene Promoter codogenic strand DNA 5' 3'...ApGpCpGpT......TpCpGpCpA... 3' 5' template strand mrna 5' pppapgpcpgpu... 3'

7 Both DNA strands can serve as template for transcription

8
9 RNA polymerisation The RNA strand grows from 5' to 3'.

10 Comparison of eukaryotic and procaryotic RNA polymerases.

11 RNA Polymerase Synthesizes RNA at the Transcription Bubble RNA polymerase is a complex enzyme with two grooves. The first groove holds a single strand of DNA, and the second groove holds the growing RNA. RNA polymerase travels down the DNA, adding ribonucleotides that complement each of the bases on the DNA template strand. Companion site for Biotechnology. by Clark Copyright 2009 by Academic Press. All rights reserved.
12 There are three eukaryotic nuclear RNA polymerases Polymerase Location Products I Nucleolus 28S, 18S, 5.8S rrna II Nucleoplasm mrnas, sirna, U1, U2, U4, U5 III Nucleoplasm 5S rrna, trnas, U3, U6 other small stable RNAs

13 Mitochondria and plastids have "prokaryotic" RNA polymerases


14 E. coli promoter consensus sequences

15 RNA polymerase in start position

16 Initiation of transcription in E. coli

17 The ternary elongation complex

18 Eukaryotic transcription
19 Structure and organization of a eukaryotic gene important sequence motifs in promotors
20 Transcription initiation in eukaryotes requires core transcription factors

21 Interaction with enhancers is required for efficient transcription initiation in eukaryotes

22 Enhancer and Insulator Sequences


23 RNA processing in eukaryotes The basic transcription mechanism in eukaryotoes is similar to prokaryotes Guanyltransferase adds 7-methylguanosin to 5' end

24 An endonuclease cuts approximately 20 nucleotides downstream of the polya addition signal.
25 PolyA polymerase synthezises the poly(a) tail (about 200bp) to yield the complete primary mrna (pre-mrna).
26 Eukaryotic genes contain exons and introns The ovalbumin gene is 7.7 kb long and encodes a mrna of 1.9 kb.




27 The ovalbumin gene is 7.7 kb long and encodes a mrna of 1.9 kb.
28 Splice consensus sequences

29 Conserved sequences in introns

30 Intron splicing in nuclear pre-mrna occurs within a large ribonucleoprotein complex, the splicosome.

31 Splicing mechanism Splicing is initiated by the nucleophilic attack of the 2' hydroxyl group of the branch site adenosine on the phosphodiester bond at the 5' splice site. Thereby the intron forms a lariat and releases the 5' exon. Splicing is completed with a second nucleophilic attack by the 3'- hydroxyl group of the 5' exon.
32 RNA splicing Group I introns can self-splice (ribozymes); in mitochondria of yeast, rrna genes of Tetrahymena, and plant organells Group II introns and splicocomal introns share the same splicing mechanism; in fungal mitochondria (self splicing), and in plant organells (not self splicing)
33 Three types of RNA splicing

34
 The black arrows indicate physical and/or functional")
35 An extensive network of coupling among gene expression machines Tom Maniatis, Robin Reed, Nature 416, (2002) The black arrows indicate physical and/or functional coupling
36 Gene expression factory model for coupling steps in gene expression. large RNAP subunit PIC, preinitiation complex TF, transcription factors CTD, carboxy-terminal domain CAP, capping factor SF, splicing factor pa, polyadenylation factor P, posphorylated CTD CTD consists of hepta repeats that are phosphosylated during initiation (serin 5). This is required for binding the capping apparatus. During elongation, serin 5 is dephosphorylated and serin 2 is phosphorylated, this is required for binding the splicing and poly(a) machineries. Genetic analysis: Deletion of CTD decreases mrna processing.
37 PIC, preinitiation complex TF, transcription factors CTD, carboxy-terminal domain CAP, capping factor SF, splicing factor pa, polyadenylation factor P, posphorylated CTD CTD is dephosphorylated during termination The mrna is released as a ribonucleoprotein
 UAP recruits ALY and NMD b) complex formation at exon/exon junction c) NMD remains bound in the cytoplasm d) if NMD is not")
38 Model for coupling splicing to mrna export and nonsensemediated decay (NMD). a) UAP recruits ALY and NMD b) complex formation at exon/exon junction c) NMD remains bound in the cytoplasm d) if NMD is not released by translation, degradation is triggered
39 mrna degradation pathways Reactions take place in the cytoplasm shortening the polya tail by 3' to 5' exonuclease removal of CAP degradation is completed by 5' to 3' exonuclease
40 Message The mrna of eukaryotes is processed by 5' capping 3' polyadenylation intron splicing
41 Message Prokaryotes Eukaryotes 1. All RNAs are syntesized by a single RNA polymerase 2. mrna is translated during transcription 3. Genes are colinear with mrna 4. mrna is often polycistronic 1. Three different RNA polymerases 2. mrna is processed before transport to the cytoplasm 3. Genes contain often introns 4. mrnas are monocistronic
42 Is splicing an advantage for eukaryotes? Exons often encode protein domains. 18 exons constitute the LDL receptor gene, containing six functional domains.
43 What is the evolutionary origin of splicing? Intron early vs. intron late hypothesis. (Walter Gilbert)
44 Alternativ splicing products e.g. adeno virus
45 Trans splicing intron 1 intron 2 pre-mrnas complementary intron sequences trans splice products normal products e.g. actin mrna C. elegans
46 Proteins Structure Genetic Code Translation

47 Protein structure

48

49 Protein structure α-helix

50 Protein structure: antiparallel β sheet
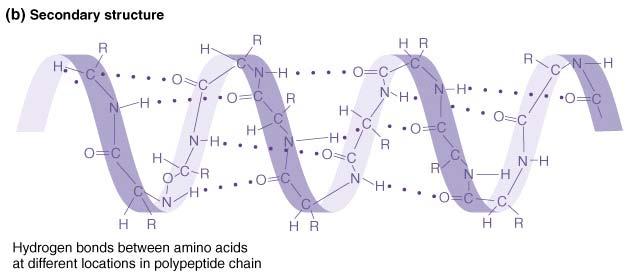
51


52 Protein structure

53 Translation

54 The genetic code
55 trna

56 FIGURE 2.16 Structure of trna Allows Wobble in the Third Position Transfer RNA recognizes the codons along mrna and presents the correct amino acid for each codon. The first position of the anticodon on trna matches the third position of the codon.

57

58 Ribosome

59 Decoding centre; Peptidyl transfer centre
60 Mechanismus of peptide synthesis The N3 of A2486 abstracts a proton from the NH2 group as the latter attacks the carbonyl carbon of the peptidyl-trna. (B) A protonated N3 stabilizes the tetrahedral carbon intermediate by hydrogen bonding to the oxyanion. (C) The proton is transferred from the N3 to the peptidyl trna 3' OH as the newly formed peptide deacylates.

61 Translation in Prokaryotes (A) (B) (C) Initiation of translation begins with the association of the small ribosome subunit with the Shine-Dalgarno sequence (S-D sequence) on the mrna. Next, the initiator trna that reads AUG is charged with fmet. The charged initiator trna associates with the small ribosome subunit and finds the start codon. Assembly is helped by initiation factors (IF1, IF2, and IF3) not shown. During elongation peptide bonds are formed between the amino acids at the A-site and the P-site. The movement of the ribosome along the mrna and addition of a new trna to the A-site are controlled by elongation factors (also not shown). The E-site binds exiting trna. Termination requires release factors. The various components dissociate. The completed protein folds into its proper three-dimensional shape.

62 Translation in Eukaryotes (A) (B) (C) (D) Assembly of the small subunit plus initiator Met-tRNA involves the binding of factors eif3 and eif2. The cap binding protein of eif4 attaches to the mrna before it joins the small subunit. The mrna binds to the small subunit via cap binding protein and the 40S initiation complex is assembled. Assembly of the large subunit requires factor eif5. After assembly, eif2 and eif3 depart.
63 Two more amino acids selenocysteine pyrrolysine COO - + H 3 N C H CH 2 SeH These are also called non-canonical amino acids
 to encode these amino acids.")
64 The genetic code for selenocysteine and pyrrolysine selenocysteine insertion element (SECIS); pyrrolysine insertion element (PYLIS). Codons that function usually as stop codons are used (in the context of a special structure of the mrna) to encode these amino acids. Nature (2004), 431, 257

65 Redefining the stop codon in mammalian messenger RNAs. Ribosomes move along mrna, deciphering the nucleotide sequence and making a protein according to the encoded amino-acid sequence. The nucleotide sequence UGA normally specifies that the ribosome should stop translation. But sometimes this stop codon can be redefined, so that the twenty-first amino acid selenocysteine is incorporated instead. This model shows how this might be done. a, A 'stem loop' structure in the downstream, untranslated part of the mrna binds to a protein called SBP2. SBP2 in turn binds to the eefsec protein, which itself has recruited the transfer RNA carrying selenocysteine. b, The selenocysteinebound trna is then delivered to the waiting UGA, for incorporation into the growing amino-acid string that constitutes the newly created protein.
, special signals in mrnas tag a subset of stop codons that are to have their meaning redefined.")
66 When stop means go. There are two ways in which the stop codon UAG could be redefined to specify the 22nd amino acid, pyrrolysine. In the first (top), special signals in mrnas tag a subset of stop codons that are to have their meaning redefined. In the second (bottom), a codon is redefined regardless of the mrna involved.
67 A short consideration of genetic nomenclature gene allele wild type wild type alleles mutation mutant alleles
68 Types of mutation loss of function mutation gain of function mutation
69 Types of mutation mutant site leaky mutation null mutation silent mutation
70 Position of mutant sites and functional consequences protein mutant site active site neutral null promoter null (or leaky, or neutral) leaky gain of function