DNA and the Molecular Structure of Chromosomes
|
|
|
- Alvin Watts
- 5 years ago
- Views:
Transcription
1 Chapter 8. DNA and the Molecular Structure of Chromosomes 1. Function of the Genetic Material 2. The Structures of DNA and RNA 3. Chemical Composition of Eukaryotic Chromosomes 4. Three Levels of DNA Packaging in Eukaryotic Chromosomes 5. Eukaryotic genomes: Repeated DNA sequences 1

2 Color-enhanced transmission of electron micrograph of the DNA from bacteriophage lambda 2
3 1. Function of the Genetic Material The genetic material must replicate, control the growth and development of the organism and allow the organism to adapt to changes in the environment. The genetic material has three essential functions: replication = genotypic function; gene expression = phenotypic function; and mutation = evolutionary function. Chromosomes are composed of nucleic acid, e.g., DNA and RNA, and proteins. 3
4 Proof that DNA mediate transformation 4
5 Avery, Macleod and McCarty s proof that the transforming principle is DNA 5

6 Proof that DNA carries the Genetic information in Bacteriophage T2 31P 32P 32S 35S 6
 is RNA, not protein. TMV contains no DNA.")
7 Proof that RNA stores the genetic information in Some Viruses Demonstration that the genetic material of tobacco mosaic virus (TMV) is RNA, not protein. TMV contains no DNA. 7
8 Viroids, Heritable infectious Naked RNA molecules Some infectious agents that cause diseases in plants, and others that are thought to cause diseases in animals, contain small circular molecules of RNA. However, unlike RNA viruses such as TMV, these RNA molecules are not packaged in protein coats. viroids meaning viruslike. The potato spindle tuber viroid (PSTV) was one of the first viroids studied; it causes the tubers of infected potatoes to be long and pointed like spindles. PSTV is a circular RNA molecule consisting of 359 nucleotides; 8
9 PSTV has a rod-shaped structure because of basepairing within the molecule. The pathogenic effects of viroids are thought to result from their ability to alter normal patterns of gene expression. 9
10 Prions, Heritable infectious proteins Other transmissible infectious agents contain no nucleic acid of any kind, just protein. These proteinaceous agents were named prions (derived from protein and infectious) in 1982 by Stanley Prusiner Prions are altered forms of normal cellular proteins in mammals. They are responsible for a group of fatal neurodegenerative diseases Prion diseases are spread by the consumption of meat containing the infective proteins. 10

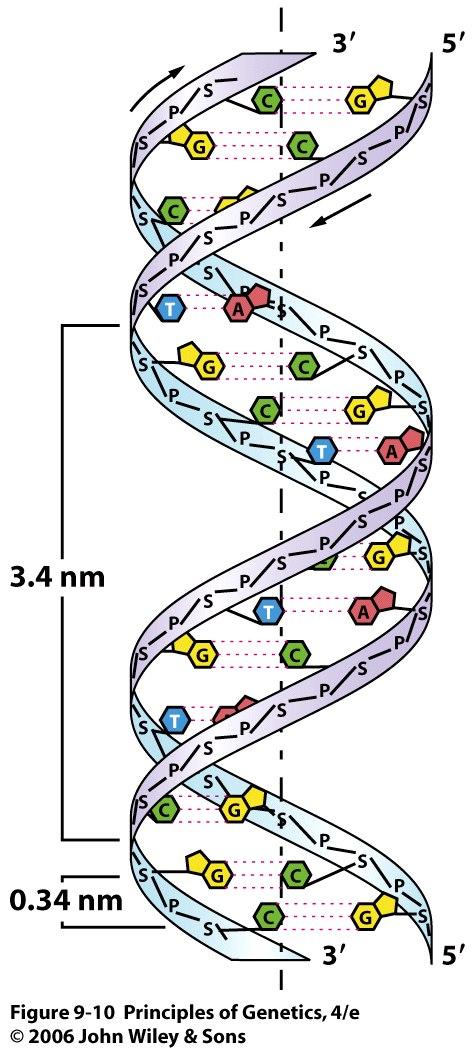
11 2. The Structures of DNA and RNA 11
12 12
13 13

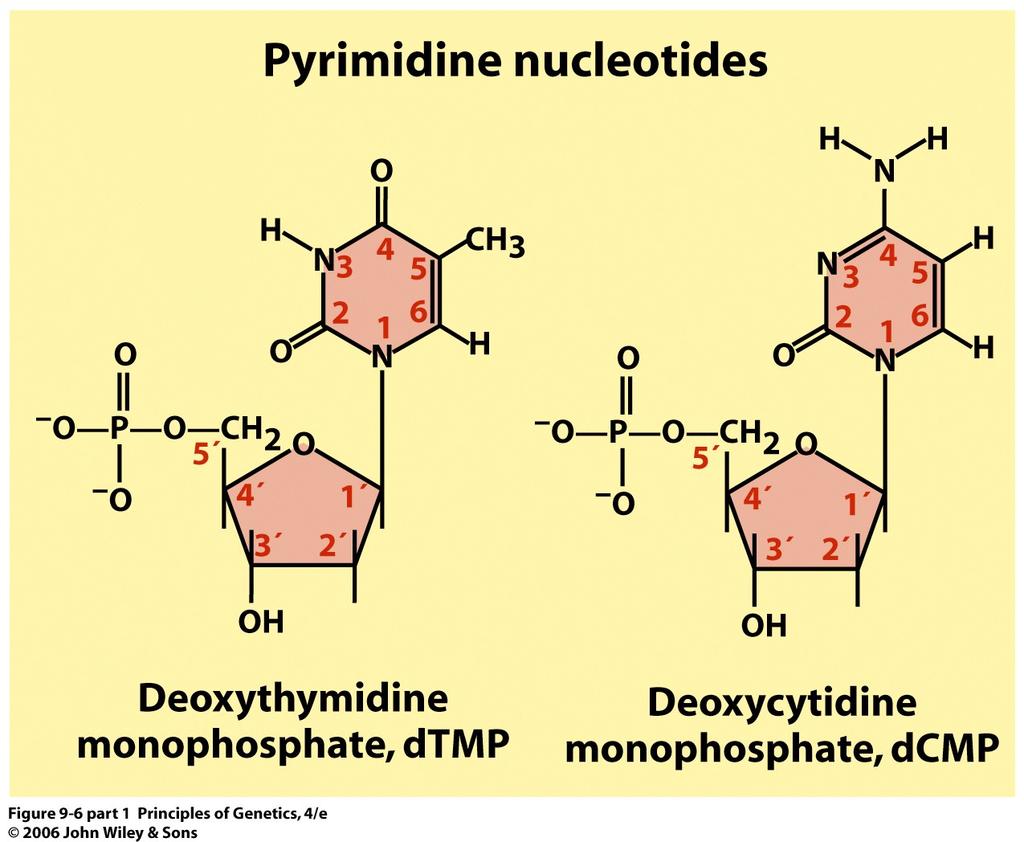
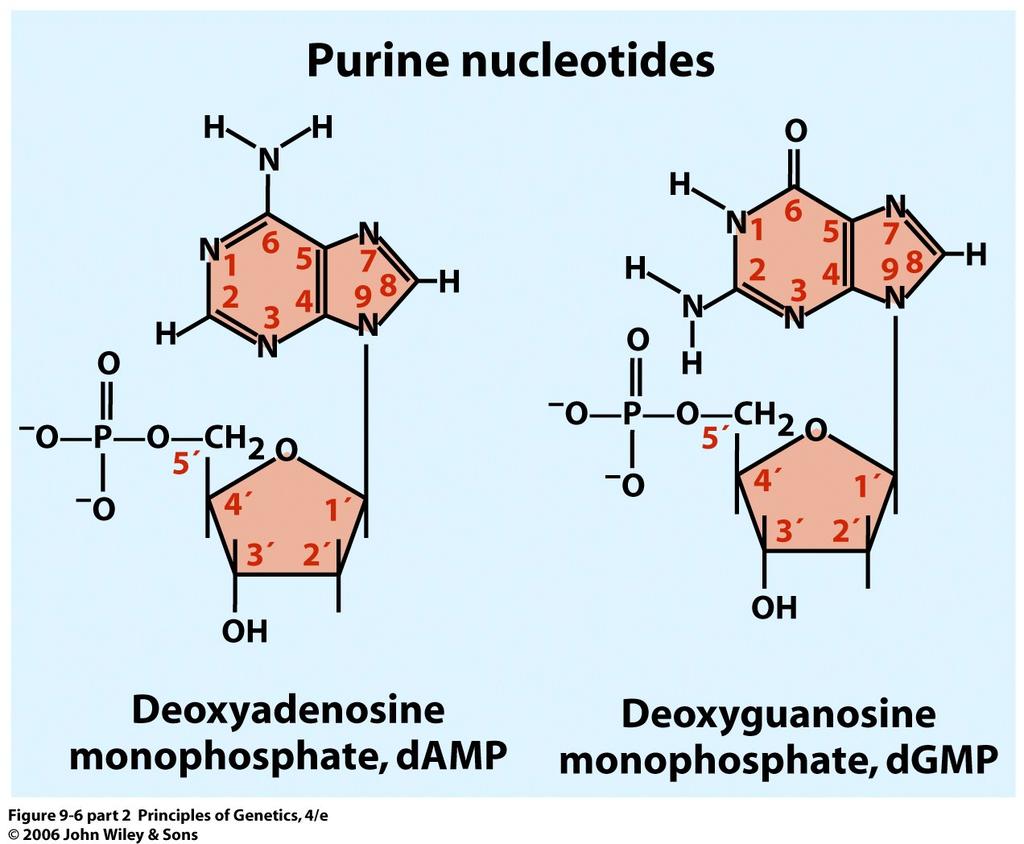
14 Formation of a polynucleotide chain by joining nucleotides with phosphodiester linkages. Note that the polynucleotides has a 5 to 3 chemical polarity (top to bottom) 14
15 15

16 James D. Watson and Francis H. C. Crick 16

17 Photograph of the x-ray diffraction pattern obtained with DNA. 17
18 18
19 complementarity The relationship between the two strands of a double helix of DNA. Thymine in one strand pairs with adenine in the other strand, and cytosine in one strand pairs with guanine in the other strand. 19

20 The sugar-phosphate backbones of the two complementary strands are antiparallel 20
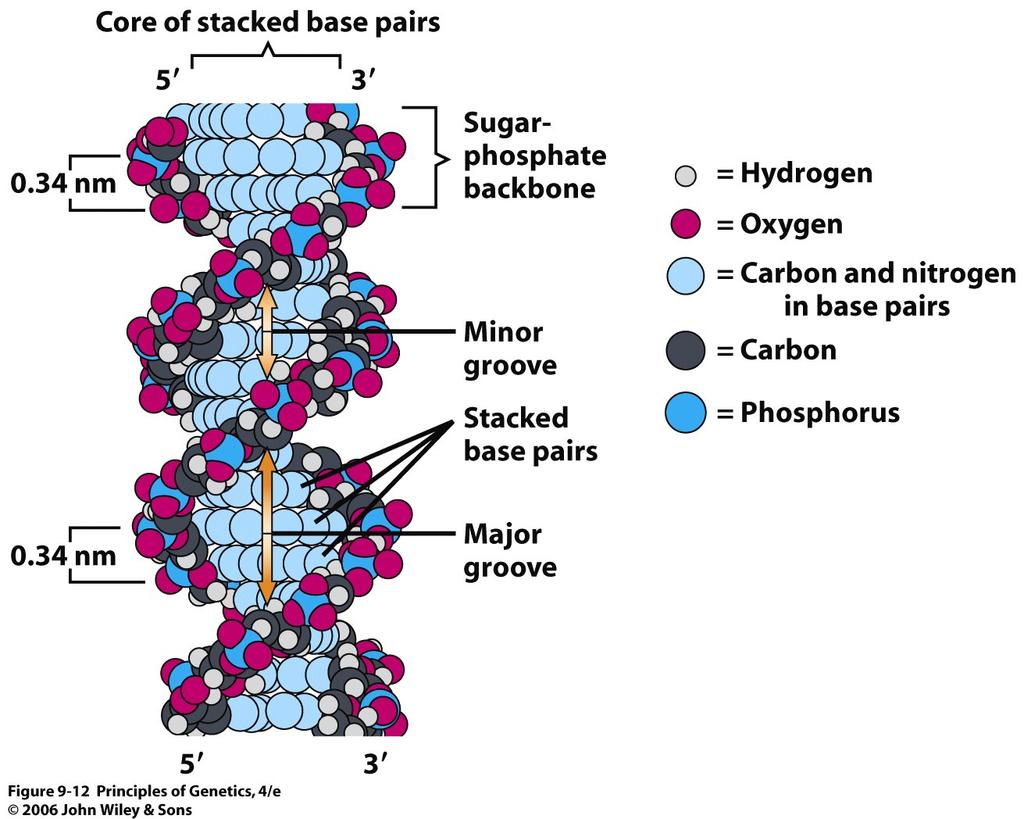
21 21

22 B-DNA B-DNA is the conformation that DNA takes under physiological conditions (in aqueous solutions containing low concentrations of salts). A-DNA A right-handed DNA double helix that has 11 base pairs per turn. DNA exists in this form when partially dehydrated. DNA-RNA, RNA-RNA duplexes in vivo. Z-DNA A left-handed double helix that forms in G:C-rich DNA molecules. The Z refers to the zig-zagged paths of the sugar-phosphate backbones in this form of DNA. 22
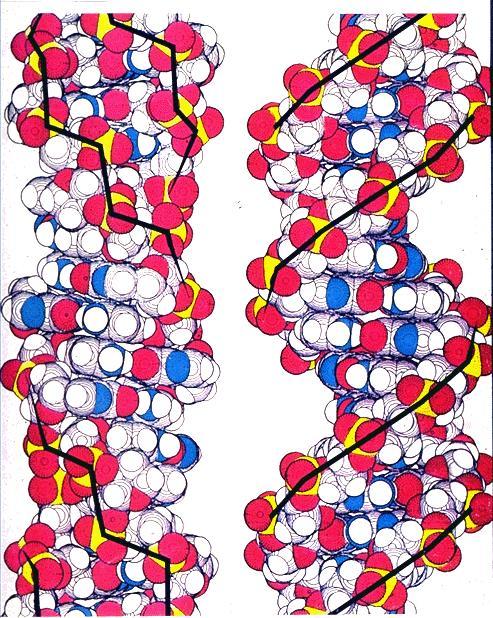
23 23

24 DNA Structure: Negative Supercoils In Vivo 24
25 Supercoiling occurs only in DNA molecules with fixed ends, ends that are not free to rotate. Obviously, the ends of the circular DNA molecules present in most prokaryotic chromosomes and in the chromosomes of eukaryotic organelles such as mitochondria are fixed. The large linear DNA molecules present in eukaryotic chromosomes are also fixed by their attachment at intervals and at the ends to non-dna components of the chromosomes. 25
26 If we rotate the free end in the same direction as the DNA double helix is wound (right-handed), a positive supercoil (overwound DNA) will be produced. If we rotate the free end in the opposite direction (left-handed), a negative supercoil (underwound DNA) will result. 26
27 3. Chemical Composition of Eukaryotic Chromosomes chromatin The complex of DNA and proteins in eukaryotic chromosomes; originally named because of the readiness with which it stains with certain dyes. When chromatin is isolated from interphase nuclei, the individual chromosomes are not recognizable. Chemical analysis of isolated chromatin shows that it consists primarily of DNA and proteins with lesser amounts of RNA 27
28 The proteins are of two major classes: (1) basic (positively charged at neutral ph) proteins called histones and (2) a heterogeneous, largely acidic (negatively charged at neutral ph) group of proteins collectively referred to as nonhistone chromosomal proteins. 28
29 The histones of all plants and animals consist of five classes of proteins. These five major histone types, called H1, H2a, H2b, H3, and H4, are present in almost all cell types. A few exceptions exist, most notably some sperm, where the histones are replaced by another class of small basic proteins called protamines. The five histone types are present in molar ratios of approximately 1 H1: 2 H2a: 2 H2b: 2 H3: 2 H4. Four of the five types of histones are specifically complexed with DNA to produce the basic structural subunits of chromatin, small (approximately 11 nm in diameter by 6 nm high) ellipsoidal beads called nucleosomes. 29
30 The histones are basic because they contain 20 to 30 percent arginine and lysine, two positively charged amino acids. The exposed NH 3 + groups of arginine and lysine allow histones to act as polycations. The positively charged side groups on histones are important in their interaction with DNA, which is polyanionic because of the negatively charged phosphate groups. 30
31 Structures of the amino acids arginine and lysine (at ph 7), which together account for 20 to 30 percent of the amino acids in histones 31
32 The nonhistone protein fraction of chromatin consists of a large number of heterogeneous proteins. The composition of the nonhistone chromosomal protein fraction varies widely among different cell types of the same organism. Thus, the nonhistone chromosomal proteins probably do not play central roles in the packaging of DNA into chromosomes. Instead, they are likely candidates for roles in regulating the expression of specific genes or sets of genes. 32

33 Separation of the chromosome-size DNA molecules of the yeast Saccharomyces cerevisiae by pulsed-field agarose gel electrophoresis. The large DNA molecules present in all 16 of the yeast chromosomes can be resolved by this procedure. 33
34 4. Three Levels of DNA Packaging in Eukaryotic Chromosomes The largest chromosome in the human genome contains about 85 mm (85,000 μm, or nm) of DNA that is believed to exist as one giant molecule. This DNA molecule somehow gets packaged into a metaphase structure that is about 0.5 μm in diameter and about 10 μm in length a condensation of almost fold in length from the naked DNA molecule to the metaphase chromosome. How? 34
35 When isolated chromatin is examined by electron microscopy, it is found to consist of a series of ellipsoidal beads (about 11 nm in diameter and 6 nm high) joined by thin threads. 35

36 Electron micrograph of rat liver chromatin showing the beads-on-a-string nucleosome substructure 36
37 Further evidence for a regular, periodic packaging of DNA has come from studies on the digestion of chromatin with various nucleases. Partial digestion of chromatin with these nucleases yielded fragments of DNA in a set of discrete sizes that were integral multiples of the smallest size fragment. These results are nicely explained if chromatin has a repeating structure, supposedly the bead seen by electron microscopy, within which the DNA is packaged in a nuclease-resistant form. This bead or chromatin subunit is called the nucleosome. 37
38 After partial digestion of the DNA in chromatin with an endonuclease (an enzyme that cleaves DNA internally), DNA approximately 200 nucleotide pairs in length is associated with each nucleosome (produced by a cleavage in each linker region). After extensive nuclease digestion, a 146-nucleotide-pairlong segment of DNA remains present in each nucleosome. This nuclease-resistant structure is called the nucleosome core. 38
39 from species to species and from cell to cell 39
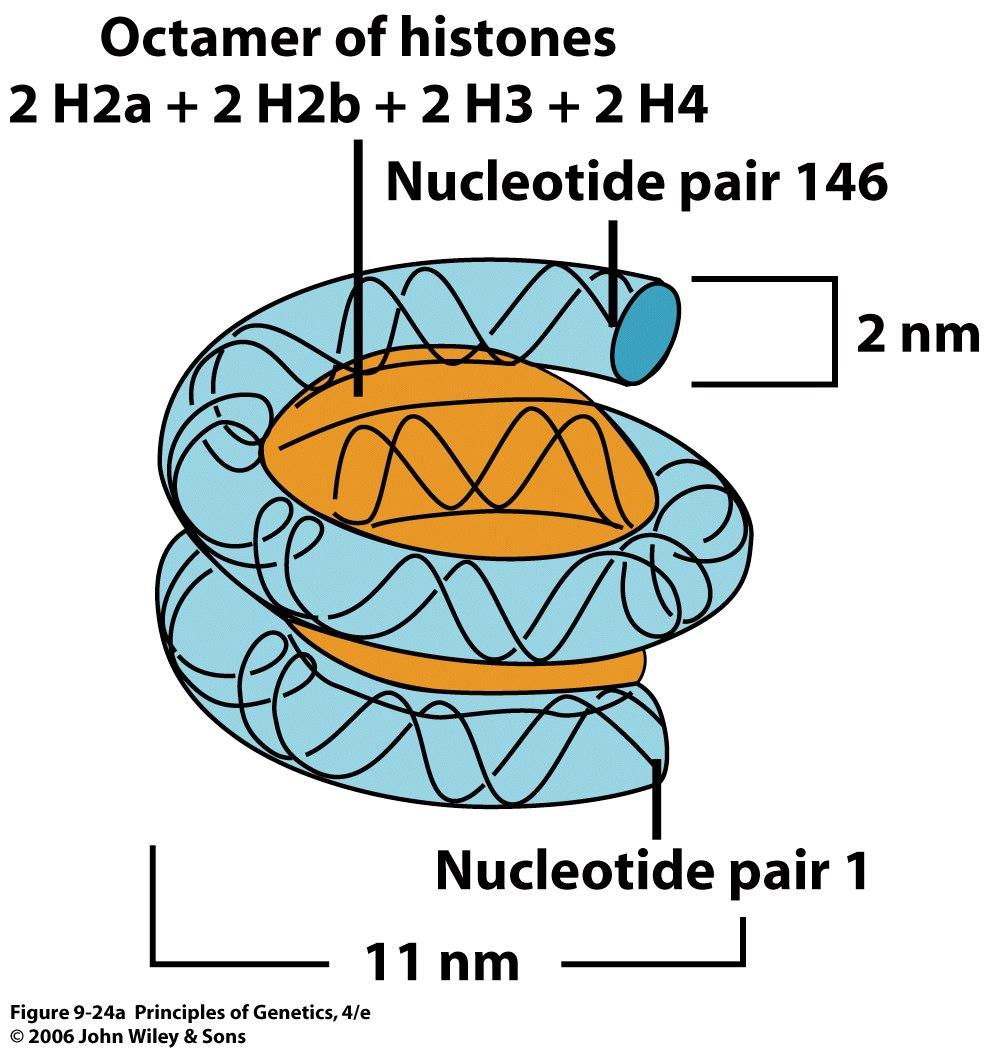
40 40

41 Complete nucleosome contains two full turns of DNA super helix 41
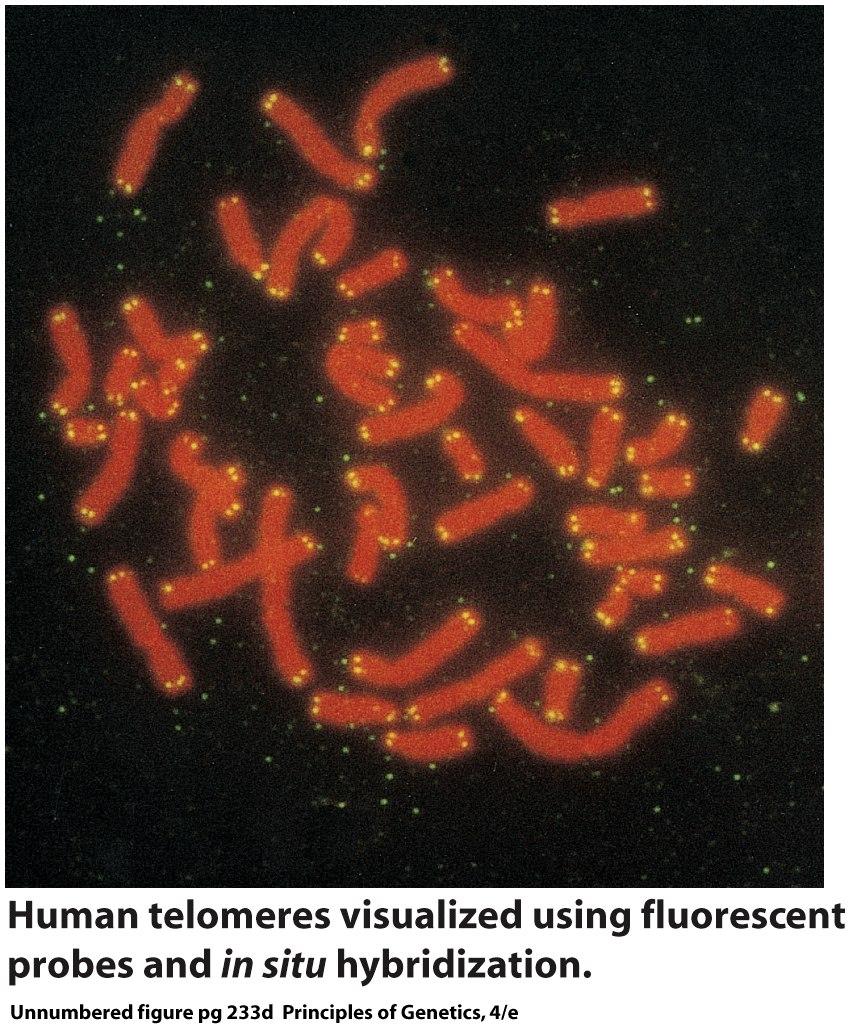
42 The basic structural component of eukaryotic chromatin is the nucleosome. But are the structures of all nucleosomes the same? What role(s), if any, does nucleosome structure play in gene expression and the regulation of gene expression? The structure of nucleosomes in transcriptionally active regions of chromatin is known to differ from that of nucleosomes in transcriptionally inactive regions. But what are the details of this structure function relationship? 42
43 The tails of some of the histone molecules protrude from the nucleosome and are accessible to enzymes that add and remove chemical groups such as methyl ( CH 3 ) and acetyl groups. The addition of these groups can change the level of expression of genes packaged in nucleosomes containing the modified histones (Chapter 21). 43

44 chromatin fibers A basic organizational unit of eukaryotic chromosomes that consists of DNA and associated proteins assembled into strands of average diameter 30 nm. 44

45 Diagram of the solenoid model of the 30-nm chromatin fiber. Histone H1 appears to stabilize the 10- nm nucleosome fiber and contribute to the formation of the 30-nm chromatin fiber. 45

46 Electron micrographs of isolated metaphase chromosomes from which the histones have been removed reveal a scaffold, or central core, which is surrounded by a huge pool or halo of DNA Scaffold: The central core structure of condensed chromosomes. The scaffold is composed of nonhistone chromosomal proteins. 46
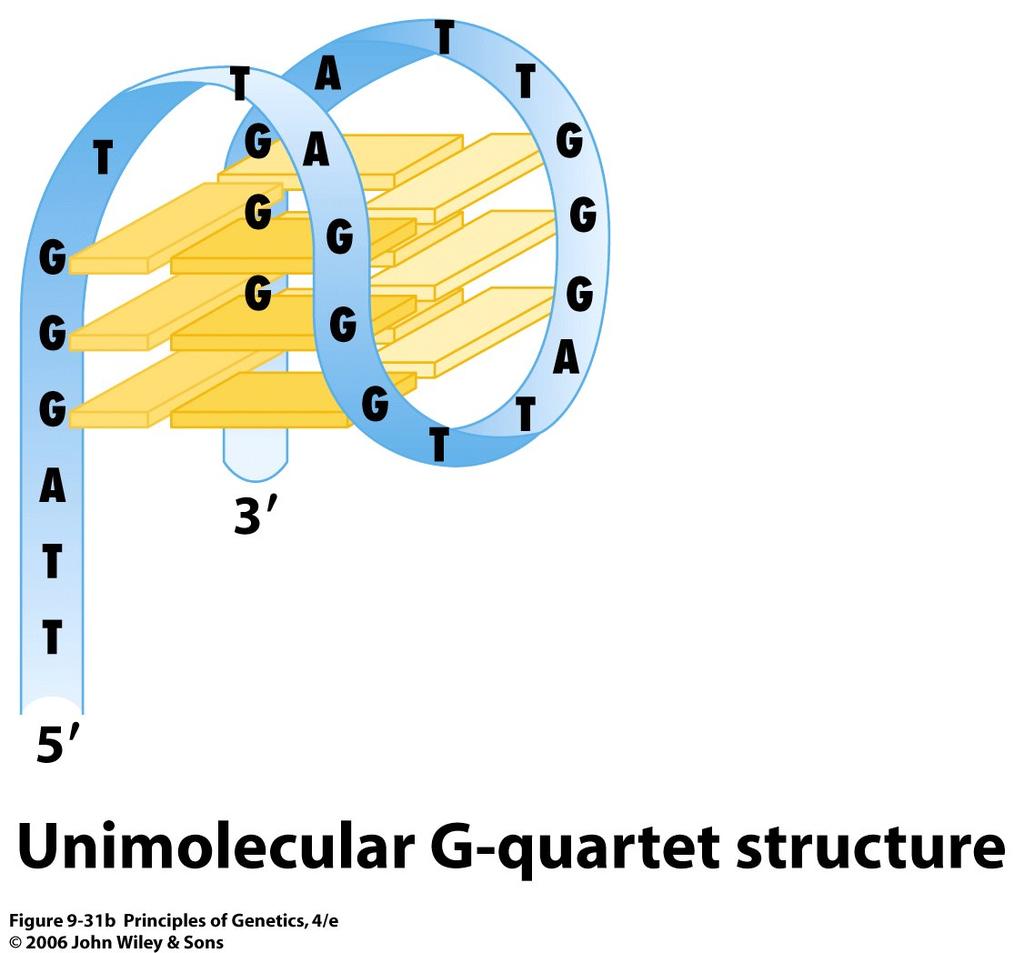
47 Centromeres The spindle fibers, which attach to centromeres, are responsible for the separation of homologous chromosomes during anaphase I of meiosis and progeny chromosomes (derived from chromatids) during anaphase II of meiosis and anaphase of mitosis. 47
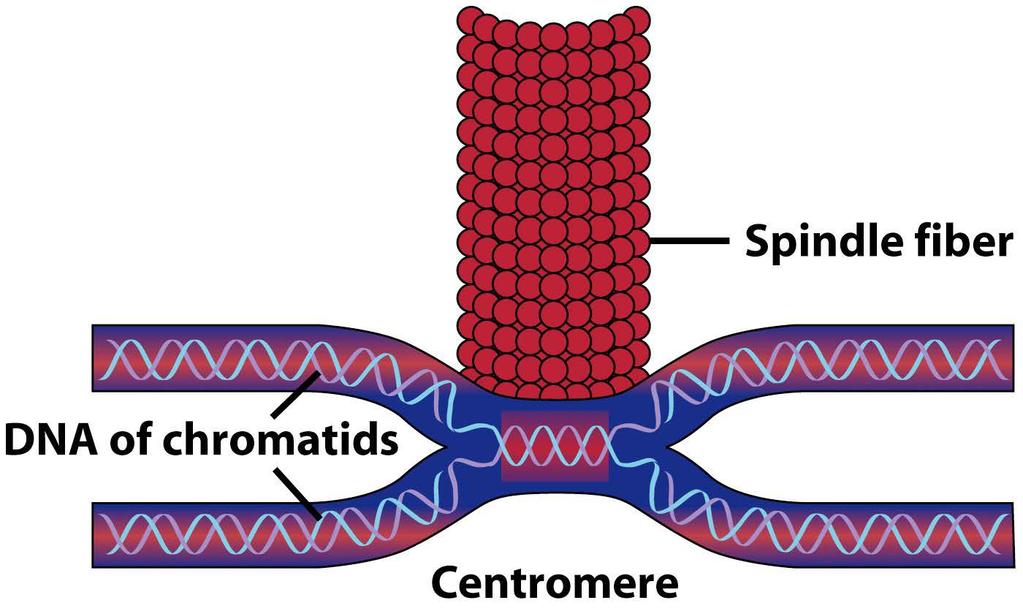
48 48
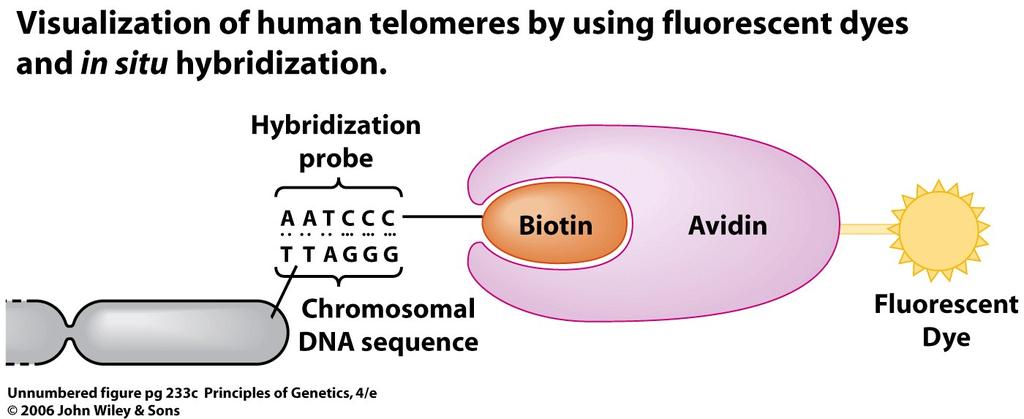
49 Diagram of the conserved structure of the centromeres in Saccharomyces cerevisiae (top) and the sequence of the centromeres (CEN region) of chromosome 3 of this species (bottom). 49
50 Telomeres telomeres (from the Greek terms telos and meros, meaning end and part ), or ends of eukaryotic chromosomes, have unique properties. The repeat sequence in humans and other vertebrates is TTAGGG, that of the protozoan Tetrahymena thermophila is TTGGGG, and that of the plant Arabidopsis thaliana is TTTAGGG. In most species, additional repetitive DNA sequences are present adjacent to telomeres; these are referred to as telomere-associated sequences. 50
51 In vertebrates, the TTAGGG repeat is highly conserved; it has been identified in more than 100 species, including mammals, birds, reptiles, amphibians, and fishes. The number of copies of this basic repeat unit in telomeres varies from species to species, from chromosome to chromosome within a species, and even on the same chromosome in different cell types. In normal (noncancerous) human somatic cells, telomeres usually contain 500 to 3000 TTAGGG repeats and gradually shorten with age. In contrast, the telomeres of germ-line cells and cancer cells do not shorten with age 51
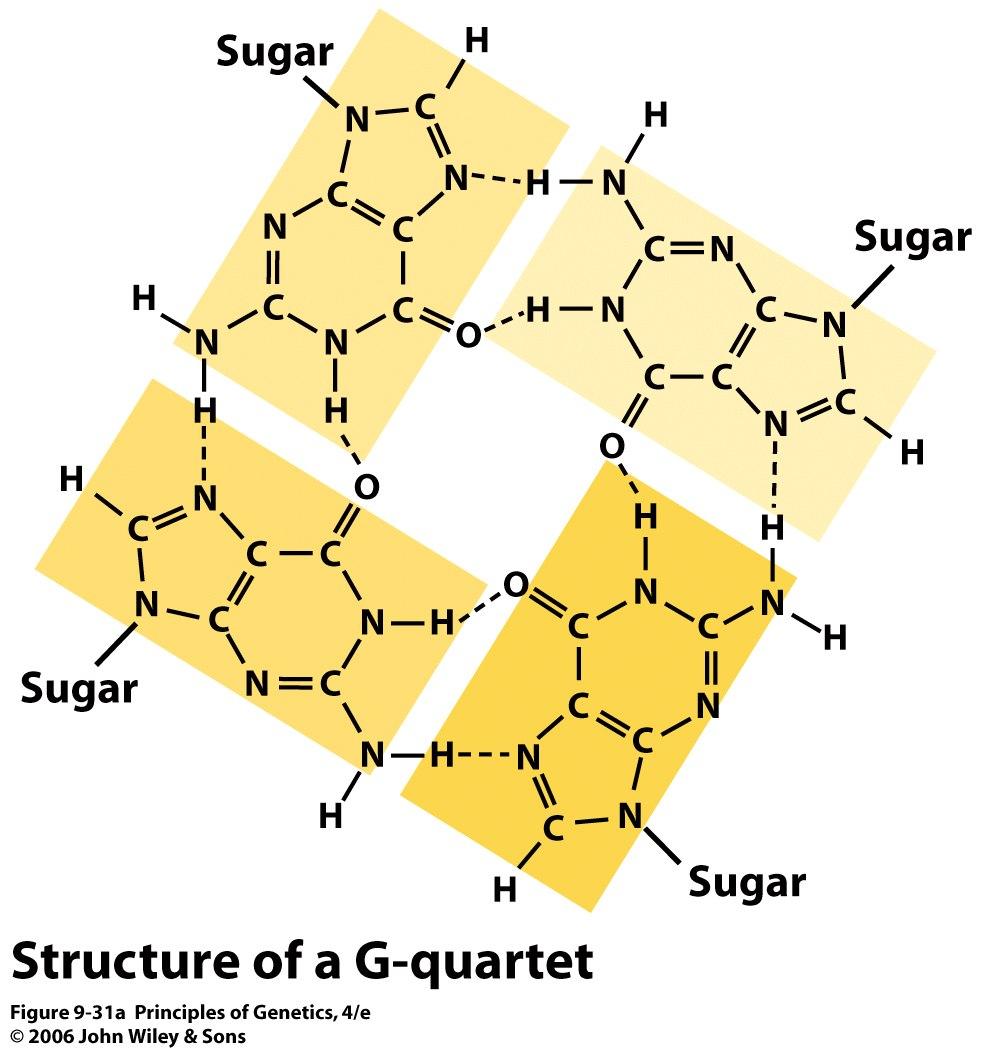
52 Most telomeres terminate with a G-rich singlestranded region of the DNA strand with the 3 end (a so-called 3 overhang). These overhangs are short (12 to 16 bases) in ciliates ( 섬모충 ) such as Tetrahymena, but they are quite long (125 to 275 bases) in humans. The guanine-rich repeat sequences of telomeres have the ability to form numerous hydrogen-bonded structures distinct from those produced by Watson Crick base-pairing in DNA. For example, four guanine residues can form a Gquartet by a special type of hydrogen bonding (Hoogstein base-pairing) 52
53 53
54 54
55 G quartets or very similar structures play an important role in stabilizing the ends of linear chromosomes. In humans, a protein called TRF-2 (Telomere Repeatbinding Factor 2) has been shown to prevent end-toend fusion of chromosomes. 55
 coats the DNA single strand displaced by the invading 3 - terminus of the")
56 TRF-1 and TRF-2 are telomere repeat-binding factors 1 and 2; both are complexes containing several proteins. Protein POT-1 (Protection Of Telomeres protein 1) coats the DNA single strand displaced by the invading 3 - terminus of the telomeric DNA. 56
57 In situ hybridization 57
58 58
59 59
.")
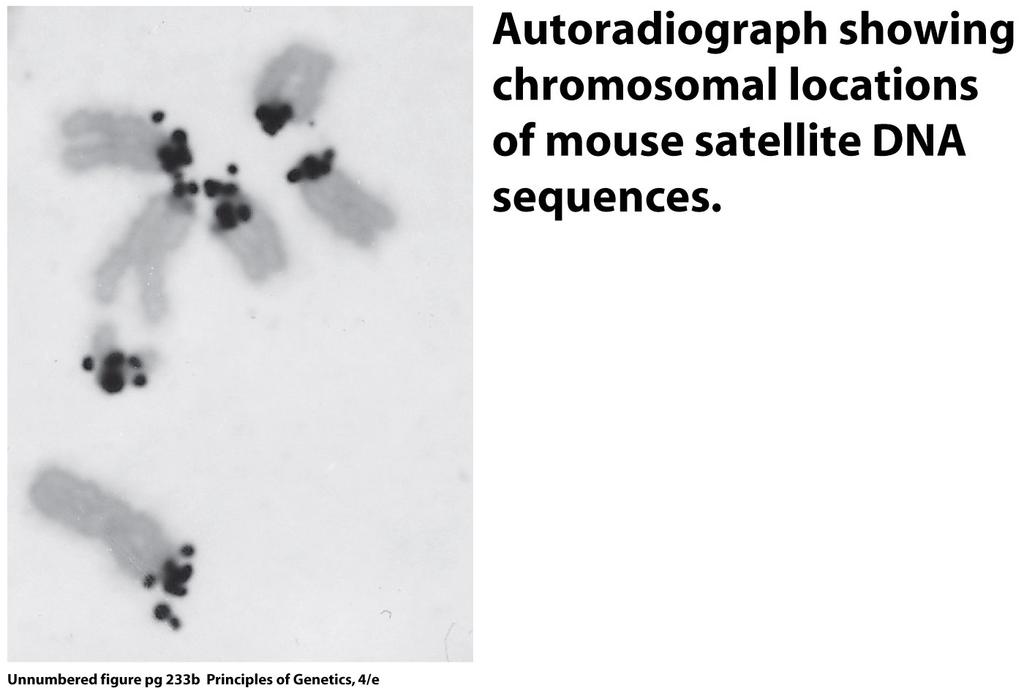
60 The location of alpha satellite DNA sequences (yellow) in the centromeres of human chromosomes (red). See the Focus on In Situ Hybridization. 60
61 61
62 5. Eukaryotic Genomes: Repeated DNA Sequences Many of the DNA sequences in eukaryotic genomes are present in multiple copies sometimes as many as a million. satellite DNAs A component of the genome that can be isolated from the rest of the DNA by density gradient centrifugation. Usually, it consists of short, highly repetitious sequences. 62
63 Detection of Repeated Sequences Denaturation Loss of native configuration of a macromolecule, usually accompanied by loss of biological activity. Denatured proteins often unfold their polypeptide chains and express changed properties of solubility. When DNA molecules in aqueous solution are heated to near 100 C, these bonds are broken and the complementary strands of DNA separate. Renaturation The restoration of a molecule to its native form. In nucleic acid biochemistry, this term usually refers to the formation of a double-stranded helix from complementary single-stranded molecules. 63
64 Surprisingly, some of the DNA sequences in the complex genomes of eukaryotes renature very rapidly. These sequences are repeated in the genome many times, sometimes a million times or more. A sequence that is repeated in the genome will be present at a higher concentration than a single-copy sequence during renaturation, thus increasing its rate of renaturation. Indeed, the rate of DNA renaturation is directly proportional to copy number the higher the copy number, the faster the rate and the less time required for renaturation. 64