Chapter 16 The Molecular Basis of Inheritance
|
|
|
- Melvin McLaughlin
- 5 years ago
- Views:
Transcription
1 Chapter 16 The Molecular Basis of Inheritance
2 Chromosomes and DNA Morgan s experiments with Drosophila were able to link hereditary factors to specific locations on chromosomes. The double-helical model of DNA was first proposed by James Watson and Francis Crick in We now know that the DNA component of chromosomes contains hereditary information.
3 Concept 16.1 DNA is the genetic material Experiments with DNA and viruses provided the first evidence that DNA is the genetic material in cells Watson and Crick developed the double helix model of DNA
4 Genetic Material After Morgan s research linked genes to chromosomes, the components of chromosomes were investigated to determine their role as genetic material. Initially, proteins were considered more likely than DNA, given that research had shown them to be varied and carry out specific functions.
5 Evidence from Bacterial Studies Frederick Griffith studied the Streptococcus pneumonia, in an attempt to develop a vaccine against pneumonia. Through his work, he discovered that a chemical component of dead cells was able to cause a heritable change in living cells. He called this transformation. Transformation a change in genotype and phenotype due to the assimilation of external DNA by a cell.
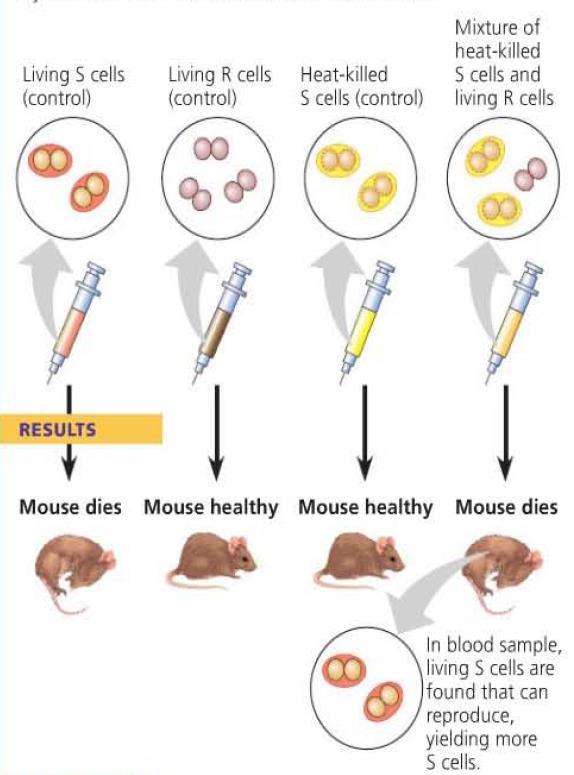
6
7 DNA, RNA, or Protein? From Griffith s work, the identity of the genetic material was still unknown. Oswald Avery carried out studies into DNA, RNA, and protein to determine which molecule is responsible for transfer of genetic information. For the studies, only one molecule was allowed to remain active in the dead cells. Of the three molecules, only active DNA resulted in transformation.
8 Evidence from Viral Studies Bacteriophages are viruses that can infect bacteria Alfred Hershey and Martha Chase conducted studies to determine that the genetic material in viruses is DNA and not proteins. Viruses were grown with either radioactive sulfur or radioactive phosphorus. The radioactive sulfur was incorporated into the viral protein, the radioactive phosphorus was incorporated into the viral DNA. Only viruses with radioactively labelled DNA were able to infect bacteria to show radioactivity inside the bacteria.
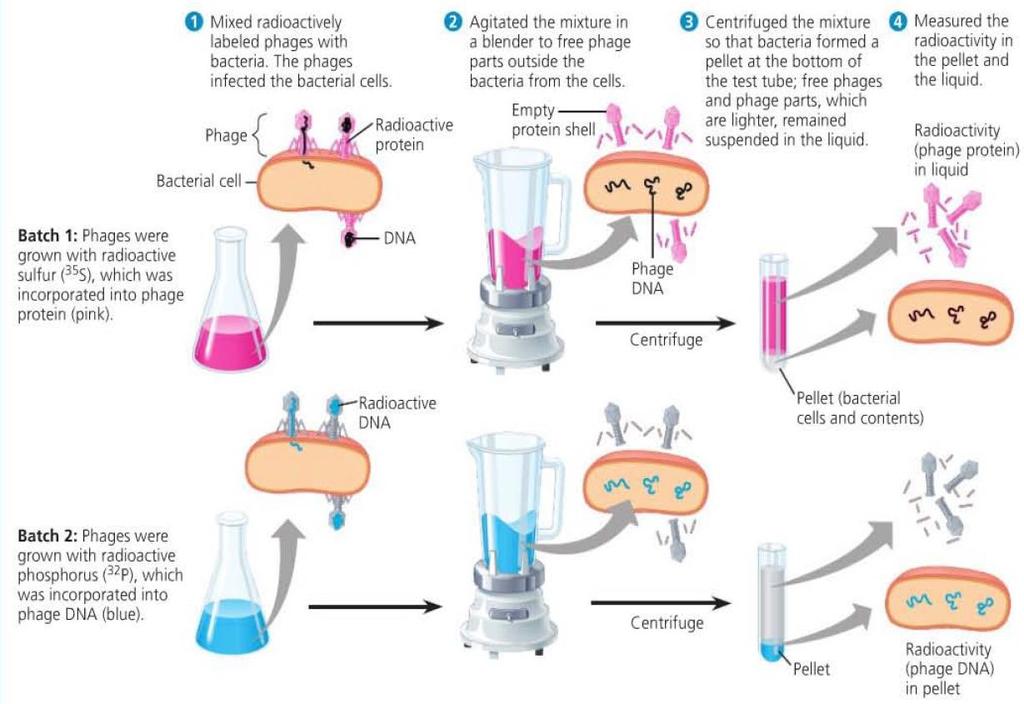
9
10 Chargaff s Rules Erwin Chargaff s research showed that the composition of DNA varied from species to species in terms of the ratio of the four nitrogen bases (adenine, guanine, cytosine, thymine). He also found that there was a pattern in the ratios within a species. The number of adenines was approximately equal to the number of thymines, and the number of cytosines was approximately equal to the number of guanines.
11 Structural Model of DNA Rosalind Franklin, while working with Maurice Wilkins, took an X-ray diffraction photograph of DNA. James Watson and Francis Crick helped to interpret the image to determine the three-dimensional structure of DNA. The image indicated that DNA had a helical structure, and the width and spacing of the molecule could also be determined. This helped to determine the double helix structure of DNA.
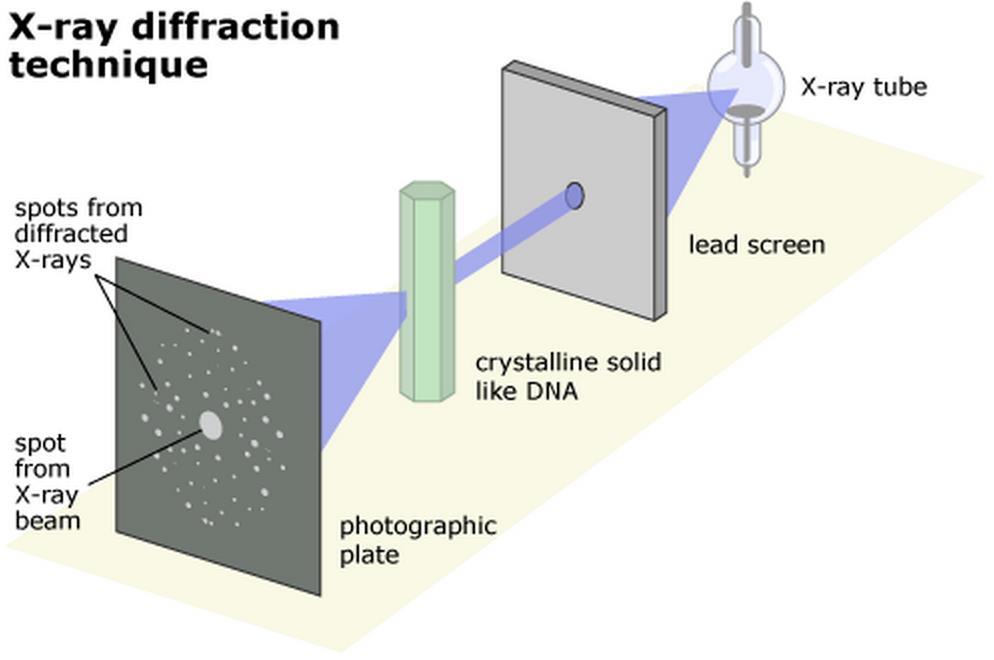
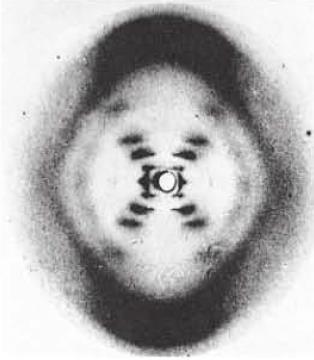
12

13 Base Pairing Based on the width of the DNA molecule, the pairings had to be purines with pyrimidines. Otherwise the width would be too wide or too narrow when compared with experimental data.
14 Base Pairing Based on hydrogen bonding, adenine forms two hydrogen bonds with thymine and cytosine forms three hydrogen bonds with guanine. This model is consistent with Chargaff s rules.
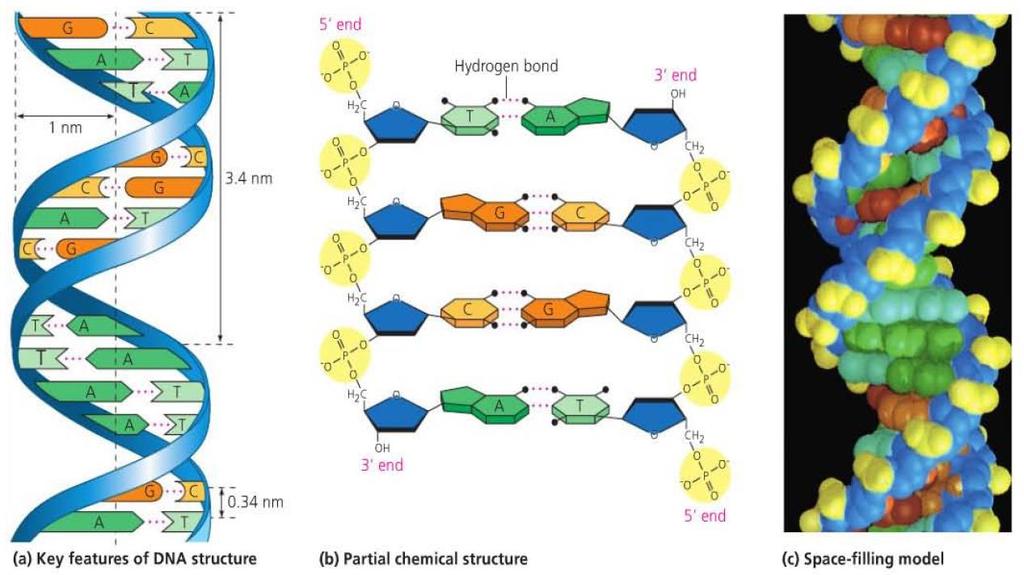
15
16 Concept 16.2 Many proteins work together in DNA replication and repair Matthew Meselson and Franklin Stahl showed that DNA replication is semi-conservative through their experiments with isotopes of nitrogen. Enzymes work together to proofread new DNA and correct errors
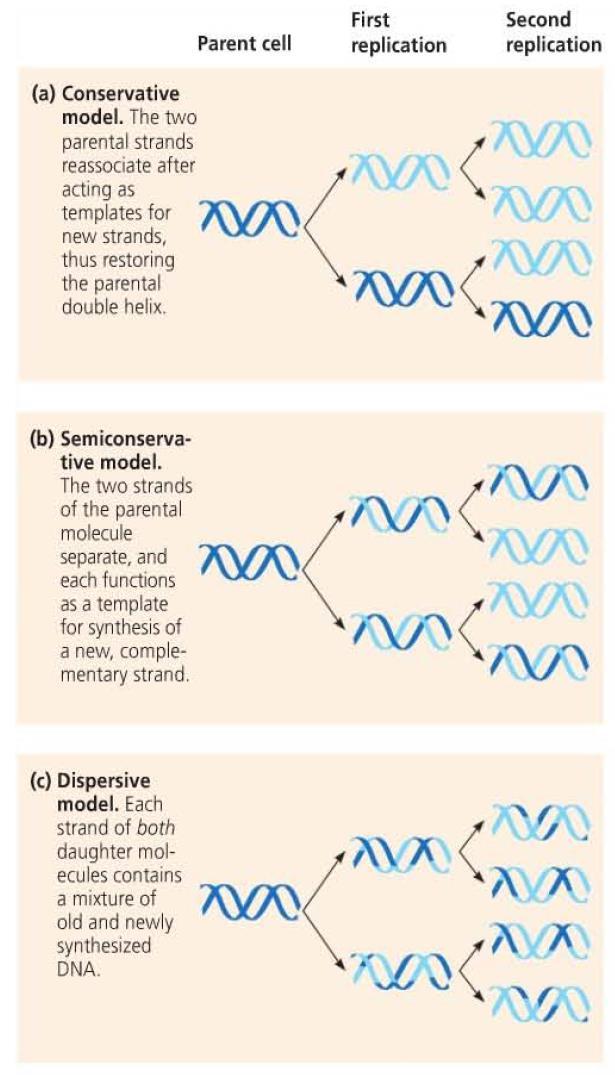
17 Base Pairing to a Template Strand Watson and Crick s model allowed for a possible copying mechanism for DNA However, this model of DNA replication was not yet tested and three models were considered. Conservative model Semiconservative model Dispersive model

18
19 Meselson and Stahl To determine the correct model of replication, Matthew Meselson and Franklin Stahl conducted an experiment involving a heavy isotope of nitrogen ( 15 N) Bacteria were cultured in medium containing heavy nitrogen before being transferred to medium containing lighter nitrogen ( 14 N) The extracted DNA was then centrifuged to separate the DNA by density. This method would allow for separation of DNA based on the incorporation of heavy vs. light nitrogen


20 Their experiment showed that there were two densities of DNA that corresponded with the semiconservative model of DNA replication
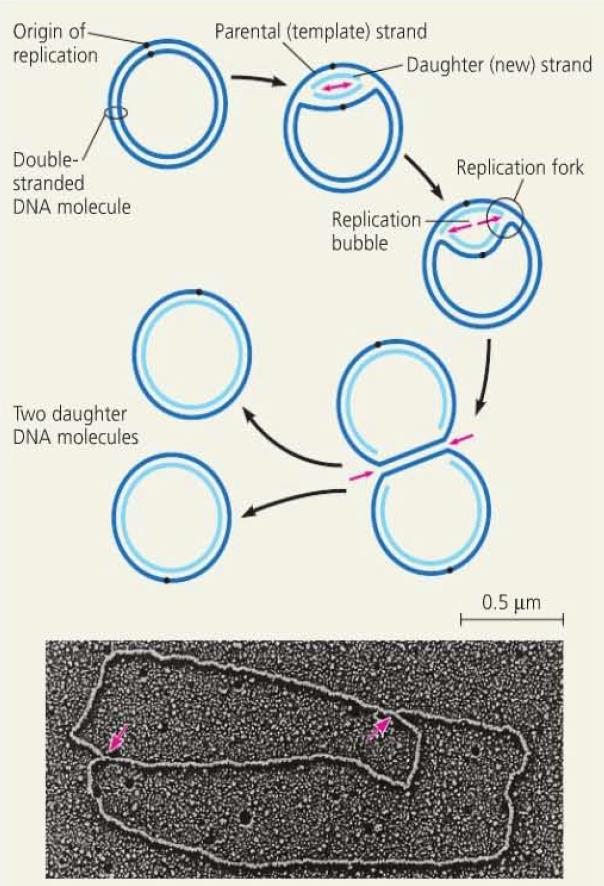
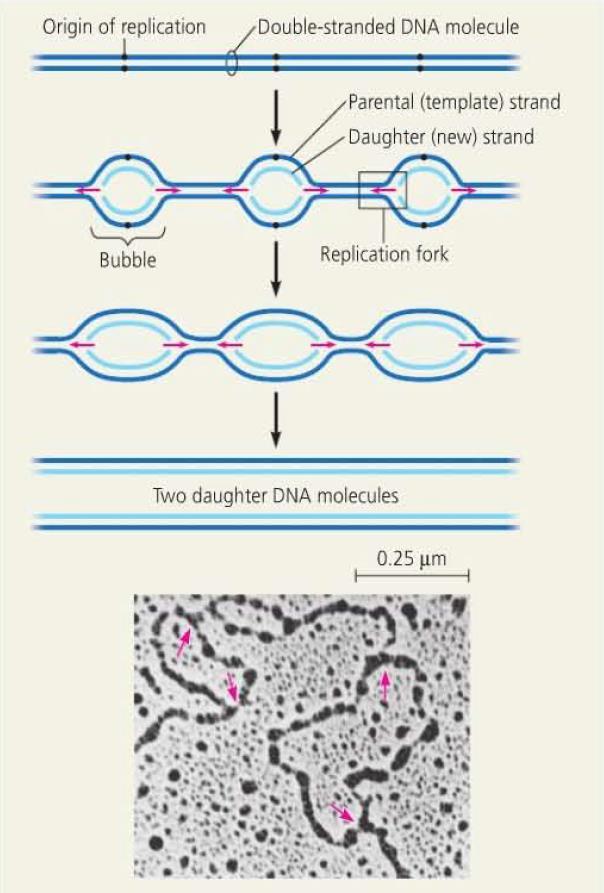
21 DNA Replication DNA replication begins at origins of replication. In prokaryotes, there is one origin of replication. In eukaryotes, there are many origins which allows for concurrent replication of different segments. The sequences of these origins are recognized by proteins which initiate replication by separating the strands, forming a replication bubble. Replication then proceeds in two directions, continuing until DNA replication is complete.
22
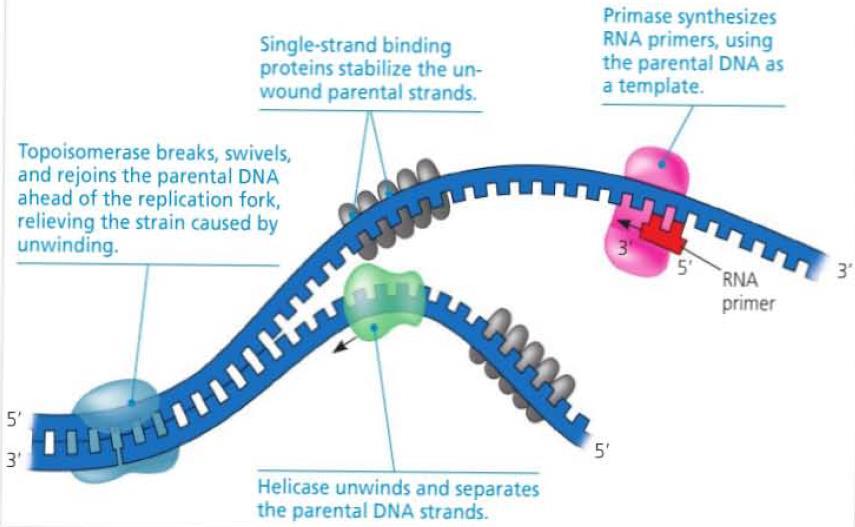
23 Replication Starter Enzymes The open bubble forms a replication fork at either end, where the DNA strands are unwound and separated by helicase enzymes. As the DNA is unwound and separated at the fork, it creates tension in front of the fork. This tension is eased by topoisomerase which breaks the DNA, allowing it to unwind, before rejoining. To stabilize the single stranded DNA that is exposed, single-strand binding protein binds to the separated strands.
24 Primers The enzyme DNA polymerase aides in the synthesis of DNA strands by adding nucleotides to a pre-existing chain. Since it cannot start synthesis without a pre-existing strand, a primer is needed. The enzyme primase synthesizes a short stretch of RNA, complementary to the exposed DNA template. This allows DNA polymerase to start synthesizing DNA from the 3 end of the primer.
25
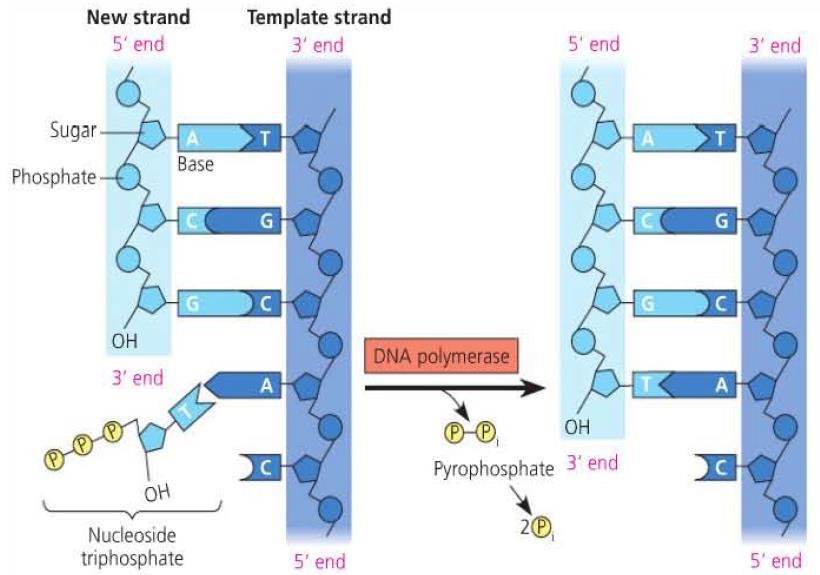
26 DNA Synthesis After primers have been formed, DNA polymerases can start extending the strand by adding nucleoside triphosphates (sugar, base, three phosphates) according to complementary base pairing with the template strand. Each addition releases two phosphate groups which provides energy for polymerization.
27
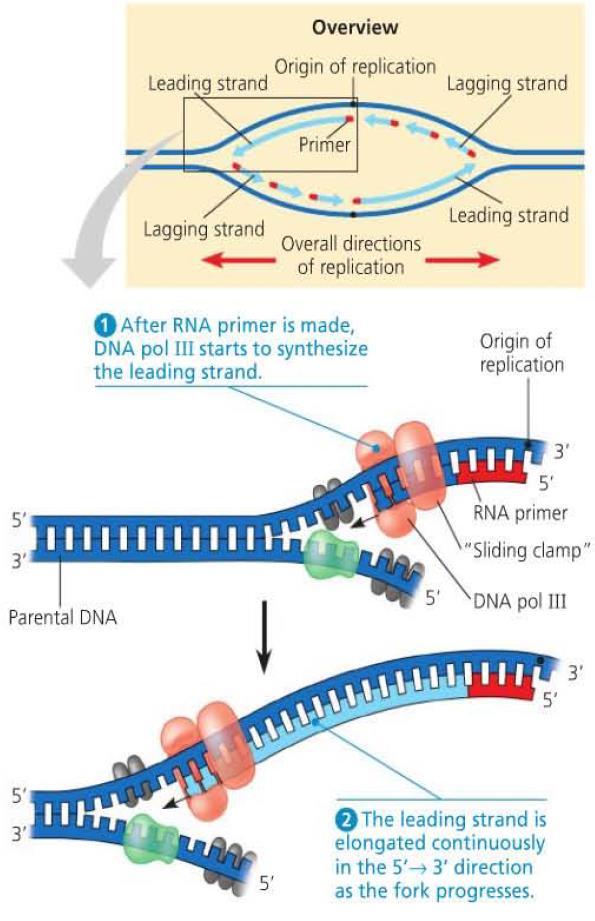
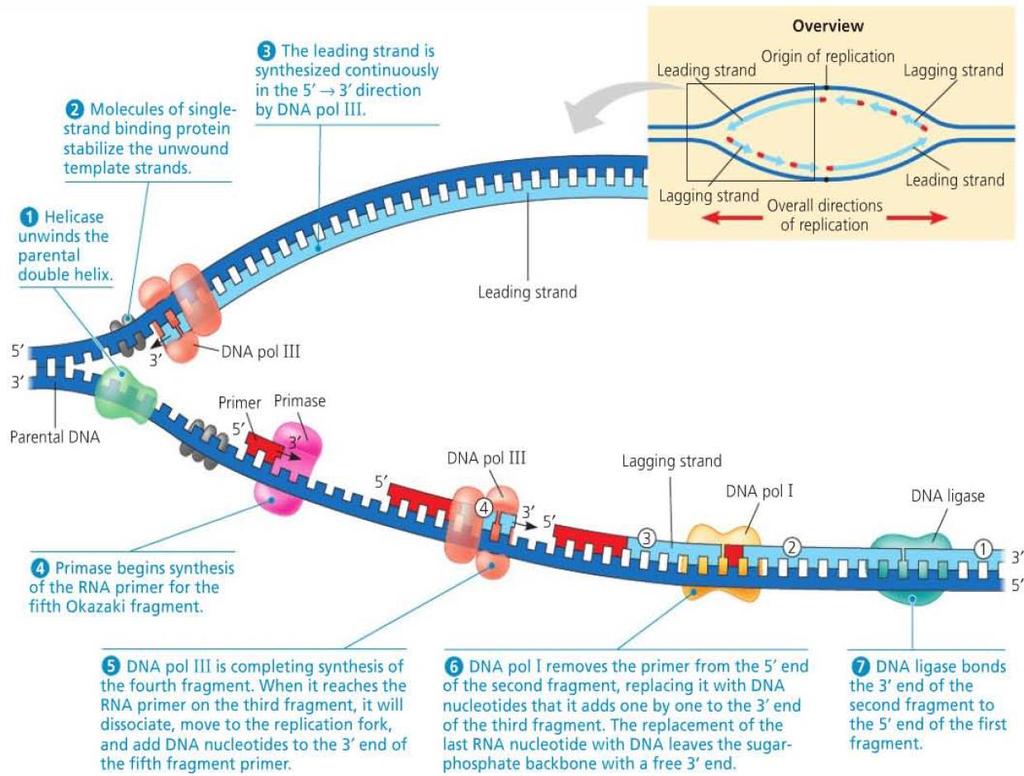
28 Antiparallel Elongation DNA polymerase only extends to the 3 end of a strand meaning that DNA is elongated in the 5 to 3 direction. This forms the leading strand of the replication fork in the 5 to 3 direction. To replicate the strand in the other direction, primers are continually added to allow DNA polymerase to extend. The resulting fragments form the lagging strand and are called Okazaki fragments. Another DNA polymerase then replaces the RNA primers with DNA segements by adding to the 3 ends. Separate fragments are then connected by DNA ligase.
29
30
31
32 DNA Replication Complex The enzymes involved in DNA replication form a DNA replication complex that works together to replicate the DNA. These complexes are anchored to the nuclear matrix. DNA is then moved through the numerous complexes to replicated the DNA.
33 DNA Replication Proteins and Their Functions Protein Helicase Function Unwinds parental double helix at replication forks Single-strand binding protein Topoisomerase Primase DNA pol III DNA pol I DNA ligase Binds to and stabilizes single-stranded DNA until it can be used as a template Relieves overwinding strain ahead of replication forks by breaking, swiveling, and rejoining DNA strands Synthesizes an RNA primer 5 end of leading strand and of each Okazaki fragment of lagging strand Using parental DNA as a template, synthesizes new DNA strand by covalently adding nucleotides to the 3 end of a pre-existing DNA strand or RNA primer Removes RNA nucleotides of primer from 5 end and replaces them with DNA nucleotides Joins 3 end of DNA that replaces primer to rest of leading strand and joins Okazaki fragments of lagging strand
34 Proofreading and DNA Repair Errors occur during DNA replication at a rate of one in nucleotides. Most of these errors are corrected during the process so that only approximately one error in 10 billion nucleotides exists. DNA polymerases proofread the new strands against the template during replication but mismatches still occur. Enzymes then perform mismatch repair such as nucleotide excision repair by the nuclease enzyme.

35 DNA damage can be caused by environmental factors such as ultraviolet rays. Exposure to ultraviolet rays can cause thymine dimers which interfere with DNA replication
36 Replicating the Ends of DNA Molecules Eukaryotic chromosomes are linear molecules. With the DNA replication complex, it is not possible to replicate or repair the end of the DNA molecule in the 3 to 5 direction. This results in a DNA molecule with uneven ends. This shortening of chromosomes occurs at the telomeres which are repeating sequences at the ends of DNA molecules. In germ cells, telomerase can lengthen the telomeres
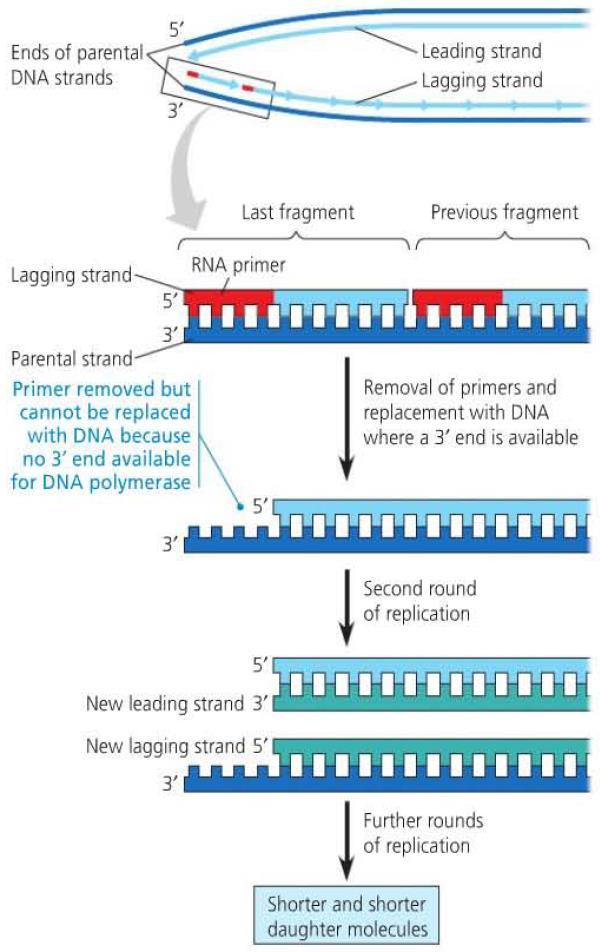
37
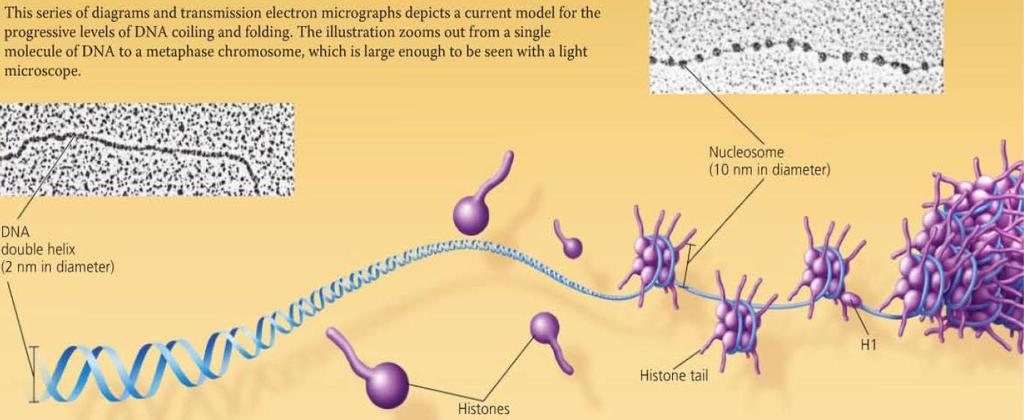
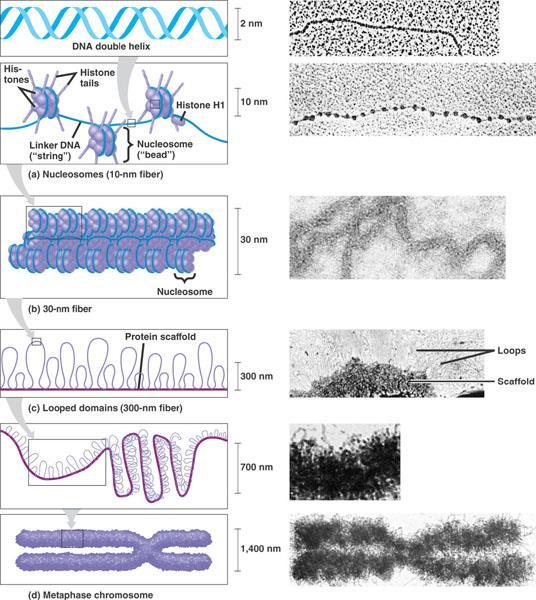
38 Concept 16.3 A chromosome consists of a DNA molecule packed together with proteins Bacterial chromosomes are usually circular DNA molecules associated with some proteins. Eukaryotic chromosomes are composed of DNA, proteins, and histones, organized to form tightly wound and packed chromatin.
39 Chromatin Packing in Eukaryotes DNA Double-helix structure composed of a sugar phosphate backbone and complementary nucleotide pairs. Histones Protein around which DNA is tightly bound. Nucleosomes basic unit of DNA packing consisting of DNA wound around a protein core composed of histones
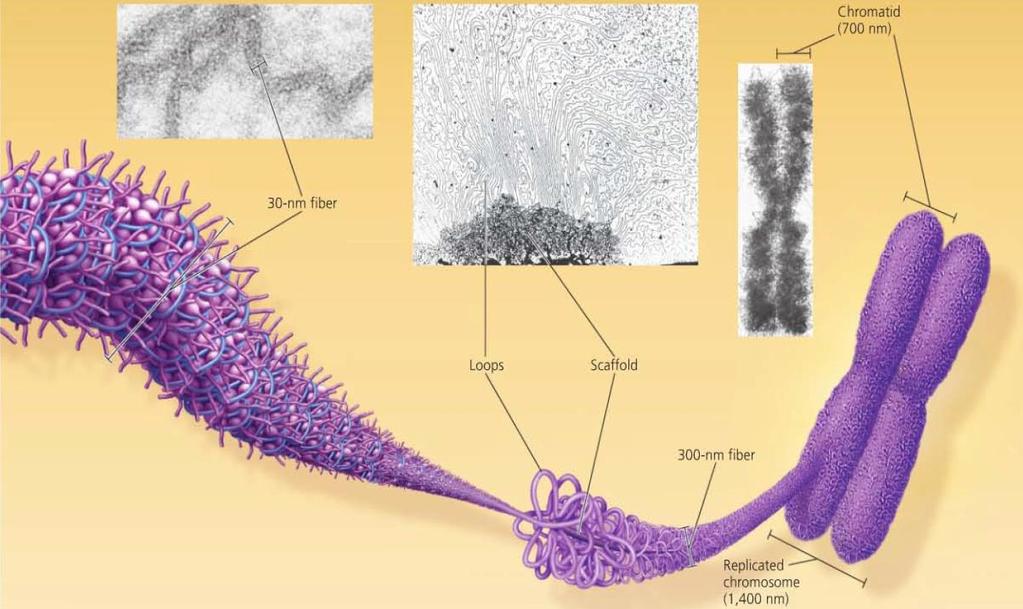
40 Chromatin Packing in Eukaryotes 30-nm Fibre Interactions between the histone tails cause the fiber to coil into a chromatin fibre that is 30-nm in thickness Looped domains Loops formed by the fibre, attached to a protein scaffold Metaphase chromosomes Condensed and replicated chromatin seen during metaphase
41
42
43 Chromatin Chromatin is not visible as individual stand until mitosis when the chromatin condenses to form visible chromosomes. The less compacted chromatin is know as euchromatin. Certain chromosomal regions are highly condensed even during interphase. This is called heterochromatin. The difference in compaction allows for certain regions to be more accessible than others. Modifications to different regions can control behaviour of the chromosome.