Proceedings of the meeting at. Murcia (Spain) 14 th 18 th May 2006
|
|
|
- Brianna McCormick
- 6 years ago
- Views:
Transcription
 14 th 18 th May 2006 Editors: Cristina Castañé and Juan Antonio Sanchez IOBC wprs Bulletin Bulletin OILB srop Vol.")
1 IOBC / WPRS Working Group Integrated Control in Protected Crops, Mediterranean Climate OILB / SROP Groupe de Travail Lutte Intégrée en Cultures Protegées, Climat Méditerranéen Proceedings of the meeting at Murcia (Spain) 14 th 18 th May 2006 Editors: Cristina Castañé and Juan Antonio Sanchez IOBC wprs Bulletin Bulletin OILB srop Vol. 29 (4) 2006
2 The IOBC/WPRS Bulletin is published by the International Organization for Biological and Integrated Control of Noxious Animals and Plants, West Palearctic Regional Section (IOBC/WPRS) Le Bulletin OILB/SROP est publié par l organisation Internationale de Lutte Biologique et Intégrée contre les Animaux et les Plantes Nuisibles, section Régionale Ouest Paléarctique (OILB/SROP) Copyright: IOBC/WPRS 2006 The Publication Commission: Dr. Horst Bathon Federal Biological Research Center for Agriculture and Forestry (BBA) Institute for Biological Control Heinrichstrasse 243 D Darmstadt (Germany) Tel Fax h.bathon@bba.de Prof. Dr. Luc Tirry University of Gent Laboratory of Agrozoology Department of Crop Protection Coupure Links 653 B-9000 Gent (Belgium) Tel , Fax luc.tirry@ugent.be Address General Secretariat IOBC/WPRS: INRA Centre de Recherches de Dijon Laboratoire de Recherches sur la Flore Pathogène dans le Sol 17, Rue Sully, BV 1540 F Dijon Cedex France ISBN web: ii



3 The organization of this meeting was supported by the following firms and institutions: IOBC Victoria 2002 Organizing Committee CICYT Agencia Regional de Ciencia y Tecnología Región de Murcia iii
4 iv
5 Preface This bulletin contains the contributions to the meeting of the IOBC working group "Integrated Control in Protected Crops, Mediterranean Climate" held in Murcia (Spain), from 14 th to 18 th of May More than 200 authors from the West Palaearctic Regional Section, and also from other regions, produced a total of sixty full articles and seven abstracts that constitute this volume. I would like to thank all for their enthusiasm in preparing their contributions and for their collaboration in the success of the meeting. Topics include major pests and diseases problems in greenhouse crops and their possible biological solutions. Integration of diverse methodologies in IPM systems and how are faced in the different countries is one of the issues raised. Among greenhouse pests, whiteflies are still one of the most important problems addressed by a number of contributors. Other arthropod pest problems, as mites, thrips, leafminers, aphids and Lepidoptera, are also presented. General predators, represented mainly by mirids and anthocorids, have an important role in the biological control of greenhouse pests and this is reflected in several contributions. Fungal and virus diseases together with soilborne diseases of plants had also a considerable number of articles. The compatibility of chemical insecticides with beneficials and the evaluation of natural pesticides are also presented. I would like to thank to all the public and private sponsors that contribute to finance this meeting. Very special thanks are given to the local organizing committee, integrated by Juan- Antonio Sanchez, Alfredo Lacasa (Institute for Development of Agrarian and Food Research of Murcia, IMIDA), Josefina Contreras and Pablo Bielza (Polytechnic University of Cartagena) who made possible this meeting. Cristina Castañé Convenor of the Working Group March, 2006 v
6 vi
7 Contents Preface... v Contents... vii IPM Systems Combining pathogens and predators of insects in biological control. D. Gillespie, J. Brodeur. C. Cloutier, M. Goettel, P. Jaramillo, R. Labbe, R. Roitberg, C. Thompson (Alma), S. VanLaerhoven... 3 Integrated pest and disease management for greenhouse-grown vegetable crops in Cyprus N. Ioannou, N. A. Seraphides... 9 Status of biological and integrated pest control in CA&ES research greenhouses at UC Davis: Successes, challenges, and costs G. Pearson, R. Starnes, T. Costamagna, R. Lane, S. Silva, A. Almedhi, A. Rumsey, M. Parrella A biological pest control story J. A. Sanchez, A. Lacasa Implementing a Geographical Information System (GIS) for pepper greenhouse natural enemy and pest management in Southeast Spain F. Cánovas, A. Lacasa, J. A. Sanchez Mise en place d'une protection biologique intégree préventive contre les ravageurs des cultures de rosiers sous serre dans le sud de la France J. Pizzol, C. Poncet, S. Hector, M. Ziegler Reduction of pesticide use on plastic house cucumber in Varamin and Shahriar region of Tehran Province of Iran through Participatory IPPM. (Abstract) A. Nasrollahi, M. S. Moghaddam Mechanical distribution of beneficials: laboratory tests G. Tropea Garzia, L. Zappalà, G. Siscaro, G. Blandini, S. Failla, G. Manetto Whiteflies Bumble bees: An effective delivery system for microbial control agents for arthropod pest and disease management L. Shipp, J P. Kapongo, P. Kevan, J. Sutton, B. Broadbent Current situation of Bemisia tabaci in protected vegetables crops in the South of France Y. Trottin-Caudal, C. Chabrière, C. Fournier, J. M. Leyre, L. Schoen vii
8 Natural parasitism of Bemisia tabaci and Trialeurodes vaporariorum in an horticultural area of Sardinia, Italy M. Nannini. L. Manca, M. Giorgini Alternative whitefly biocontrol options for greenhouse tomatoes M. Nannini, F. Foddi, G. Murgia, R. Pisci, F. Sanna, S. Spanedda Releases of Eretmocerus mundus and Macrolophus caliginosus for controlling Bemisia tabaci on spring and autumn greenhouse tomato crops R. Gabarra, R. Zapata, C. Castañe, J. Riudavets, J. Arnó Amblyseius swirskii (Acari: Phytoseiidae) as a biological control agent of the tobacco whitefly Bemisia tabaci (Horn.: Aleyrodidae) in protected sweet pepper crops in Southern Spain J. Calvo, P. Fernández, K. Bolckmans, J. E. Belda Efficacy of the Beauveria bassiana strain ATCC (Naturalis ) against whitef1ies on protected crops F. Mayoral, M. Benuzzi, E. Ladurner Efficacy of the technical grade product and commercial formulation based on Paecilomyces fumosoroseus for controlling whitefly under laboratory conditions A. Padilla-Cubas, S. Amador, E. Hernández-Suárez, A. Carnero, J. M. Lara, C. Fernández Evaluation of the effect of two potassium based products on Bemisia tabaci in greenhouse tomatoes L. Schoen Imidacloprid as soil application against whitefly Bemisia tabaci in greenhouse cucumber. (Abstract) V. Baniameri, A. Sheikhi Laboratory screening of pesticides against Lecanoideus floccissimus. (Abstract) E. Hernández-Suárez, C. Ramos-Cordero, A. Carnero Does the PGPR Bacillus subtilis induce plant resistance to whiteflies and Phythium in greenhouse tomato? (Abstract) A. Hanafi, K. Fellah Mites Crop loss assessment of Pediculaster fletchmani (Acari: Pygmephoridae) on button mushrooms K. Kheradmand, K. Kamali, Y. Fathipour, E. M. Goltapeh A. R. Nemati Biology of the two spotted spider mite, Tetranychus turkestani (Acari: Tetranychidae) on four common varieties of eggplant in Iran E. Soleimannejadian, A. Nemati, P. Shishehbor, K. Kamali, V. Baniameri viii
9 Efficacy of Phytoseiulus persimilis versus Neoseiulus californicus for controlling spider mites on greenhouse sweet pepper P. Weintraub, S. Kleitman, N. Shapira, Y. Argov, E. Palevsky Identification of pollens for small scale mass rearing of Neoseiulus californicus and a novel method for quality control Y. Argov, M. Berkeley, S. Domeratzky, E. Melamed, P. Weintraub, E. Palevsky Are spider mites aware of the presence of thrips? B. Ahmadi, S. H. Goldansaz, A. Ashouri Effect of different types of prey on fecundity of the phytoseiid mite Typhlodromips swirskii, a potential biological control agent for horticultural greenhouse pests in the Canary Islands. (Abstract) E. Hernández-Suárez, M. C. Velásquez, M. I. Paz-González, F. J. González, A. Carnero, F. J. Ferragut Feeding rate of Stethorus gilvifrons on Tetranychus urticae in three greenhouse cucumber cultivars with different resistance levels N. Kheradpir,.J. Khalghani, H. Ostovan, M R. Rezapanah Thrips, Leafminers, Aphids and Lepidoptera Pests Development of traps for monitoring thrips (Thysanoptera: Thripidae) C C Chu, M A. Ciomperlik, M. Richards, T. J. Henneberry Preliminary field trials with the synthetic sexual aggregation pheromone of Frankliniella occidentalis on protected pepper and tomato crops in South-east Spain M. Gómez, F. García, R. GreatRex, M. Lorca, A. Serna The addition of potassium silicate to the fertilizer mix to suppress Liriomyza leafminers attacking chrysanthemums M. P. Parrella, T. Costamagna Natural parasitism of Liriomyza sativae (Diptera: Agromyzidae) on cucumber under field and greenhouse conditions Y. Fathipour, M. Haghani, A. A. Talebi, V. Baniameri, A. A. Zamani First data on the population dynamics of aphidophagous syrphids in Mediterranean pepper greenhouses A. Pineda, M a Á. Marcos-Garciá Evaluation of two aphid parasitoids as candidates for biocontrol of aphid pests in protected cultivation in Brazil. V. H. P. Bueno, M. V. Sampaio, J. C. van Lenteren, B. F. De Conti, R. J. Silva, S. M. M. Rodrigues, A. B. Carnevale ix
10 Host stage preference by Aphidius colemani and Aphidius matricariae (Hymenoptera: Aphidiidae) as parasitoids of Aphis gosoypii (Hemiptera: Aphididae) on greenhouse cucumber A. A. Talebi, A. A. Zamani, Y. Fathipour, V Baniameri, K. Kheradmand and M. Haghani The number of Cotesia plutellae needed to suppress the population growth of diamondback moth, Plutella xylostella, in a greenhouse S. Urano, J. Abe, M. Matsumura, K. Nagasaka, J. Takabayasi Functional response of Chelonus oculator (Hym.: Braconidae) in greenhouse conditions: influence of the parasitoid release ratio. (Abstract) M. García-Martin, M. Gámez-Cámara, A. Torres-Ruiz, T. Cabello Generalist Predators When does alternative food promote biological pest control? M. W. Sabelis, P. C.J. Van Rijn Feeding behavior of the rove beetle, Atheta coriaria Kraatz (Coleoptera: Staphylinidae) under laboratory conditions E. M. Birken, R. A. Cloyd The predatory bug Orius niger: its biology and potential for controlling Thrips tabaci in Iran V Baniameri, E. Soleyman-nejadian, J. Mohaghegh Predatory capacity and longevity of adults of Orius thyestes (Hemiptera, Anthocoridae) on different prey E. C. Pedroso, V. H. P. Bueno, R. J. Silva, A. R. Carvalho, A. J. F. Diniz, M. P. F. Silva, L. M. Carvalho Disinfection of oviposition substrate with sodium hypochlorite: effects on some biological traits of Orius thyestes A. J. F. Diniz, V. H. P. Bueno, A. R. Carvalho, E. C. Pedroso, R. J. Silva, L. M. Carvalho Intraguild predation between Orius insidiosus and Aphidius colemani, and biological control of Aphis gossypii L. S. R. Pierre, V. H. P. Bueno, M. V. Sampaio, J. C. van Lenteren, B. F. De Conti, M. P. F. Da Silva, L. C. P. Silveira Differentiation of Macrolophus pygmaeus (Rambur 1839) and Macrolophus melanotoma (Costa 1853) (Heteroptera: Miridae) based on molecular data J. I. Martinez-Cascales, J. L. Cenis, J. A. Sanchez Reproductive traits of the generalist predator Maerolophus caliginosus C. Castañé, 0. Alomar, J. Riudavets, C. Gemeno x
11 Rearing predatory bugs using the brine shrimp Artemia sp. as alternative prey food. J. Riudavets, J. Arnó, C. Castañé Investigation on the efficiency of a predator: a strategy beyond satiation D. Perdikis, P. Labropoulos, D. Maselou, A. Fantinou Zoophytophagy of the plantbug Nesidiocoris tenuis in tomato crops in southeast Spain J. A. Sanchez, M del Pino-Pérez, M. M. Davó, J. I. Martinez-Cascales, A. Lacasa Characterization of damage to tomato plants produced by the zoophytophagous predator Nesidiocoris tenuis J. Arnó, C. Castañé, J. Riudavets, R. Gabarra Susceptibility to virus infection of candidate plants used to enhance predatory dicyphine (Heteroptera: Miridae) I. Morales, J. A. Sanchez, O. Alomar, A. Lacasa, A. Fereres Plant Diseases Biological Control of Fusarium oxysporum fsp. melonis racc 1.2 by cross protection H. Chikh-Rouhou, J. M. Álvarez, R. González-Torres Evaluation of susceptibility of varieties of Gerbera jamesonii to Fusarium wilt A. Minuto, D. Bertetti, M. L. Gullino, A. Garibaldi Efficacy of Ampelomyces quisqualis isolate M-10 (AQ 10 ) against powdery mildews (Erysiphaceae) on protected crops M. Benuzzi, E. Ladurner, F. Mayoral Quantification of powdery mildew removal by the mycophagous beetle Psyllobora vigintimaculata (Coleoptera: Coccinellidae) A. M. Sutherland, M. P. Parrella UV interfering nets reduce TYLCD incidence and progress in tomato crops: influence of host genotype L. Velasco, D. Janssen, M. Catalá, J. Costa Iris yellow spot virus (IYSV): a new disease in Spain C. Córdoba, L. Martínez-Priego, R. Muñoz, C. Jordá Identification and determination of transmission ability of thrips species as vectors of two tospovirus, tomato spotted wilt virus (TSWV) and impatiens necrotic spot virus (INSV), on ornamental plants in Iran. (Abstract) T. Ghotbi, V. Baniameri Soil-borne Diseases Effect of soil disinfestation on fungi in greenhouses planted with sweet peppers M. Á. Martínez, A. Lacasa, M. M. Guerrero, C. Ros, M. C. Martínez, P. Bielza, J. Tello xi
12 Biofumigation as a technique for the control of pathogenic viruses and bacteria M. J. Zanón, J. C. Vilaseca, M.I. Font, C. Jordá Biofumigation vs. biofumigation plus solarization to control Meloidogyne incognita in sweet pepper M. M. Guerrero, C. Ros, M. A. Martínez, M. C. Martínez, A. Bello, A. Lacasa Integrated management of Meloidogyne resistance in sweet pepper in greenhouses C. Ros, M. M. Guerrero, M. A. Martíinez, A. Lacasa, A. Bello Effect of organic amendments of Ricinus communis and Azadirachta indica on root-knot nematodes Meloidogyne javanica infecting tomatoes in Morocco Z. Ferji, E. H. Mayad, T. Laghdaf, E. M. Cherif Natural Pesticides and Side Effects on Beneficials Effects of natural insecticides on Frankliniella occidentalis and Orius spp J. Contreras, V. Quinto, J. Abellán, E. Fernández, C. Grávalos, L. Moros, P. Bielza Insecticidal activity of essential oil from Vitex pseudo-negundo against Brevicoryne brassicae S. Moharramipour, B. Z. Sahaf Evaluation de l'effet nematicide de l'extrait methanolique de quelques plantes médicinales au Maroc E. H. Mayad, Z Ferji, L. M I. Hassani Side effects of pesticides on Orius insidiosus (Hemiptera: Anthocoridae) G. A. Carvalho, V. H. P. Bueno, A. P. Moura, L. C. D. Rocha, F. Z. V. Torres Side effects of pesticides on Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) G. A. Carvalho, A. P. Moura, V. H. P. Bueno Compatibility of Hyposoter didymator, an endoparasitoid of Spodoptera littoralis, with several insecticides used on horticultural crops J. J. Morales, P. Medina, E. Viñuela xii
13 IPM Systems
14 2
15 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp. 3-8 Combining pathogens and predators of insects in biological control Dave Gillespie 1, Jacques Brodeur 2, Conrad Cloutier 3, Mark Goettel 4, Patricia Jaramillo 5, Roselyne Labbe 6, Bernie Roitberg 5, Colleen Thompson (Alma) 5, Sherah VanLaerhoven 7 1 Agriculture and Agri-Food Canada, Research Centre, POB 1000, Agassiz, British Columbia, 2 Centre de Recherche en Horticulture, Université Laval, Québec, Québec, 3 Département de Biologie, Université Laval, Québec, Québec, 4 Agriculture and Agri-Food Canada Research Centre, POB 3000, Lethbridge, Alberta, 5 Department of Biology, Simon Fraser University, 8888 University Drive, Burnaby, British Columbia, 6 Institut de Recherche en Biologie Végétale, Département des Sciences Biologiques, Université de Montréal, Montréal, Québec, 7 Department of Biology, University of Windsor, 401 Sunset Ave, Windsor, Ontario, Canada. Abstract: When multiple natural enemies are used for biological control, the contribution of each to the combined mortality of the target is of concern. Using a functional response model, we show that, for the special case of a predator foraging after application of a microbial insecticide, competition should reduce the efficiency of the predator and result in a combined prey mortality that is less than additive over short intervals. We summarize results from three recent studies with microbial insecticides in combination with predators. Over time scales in excess of one generation, numerical response of the predator can reduce the importance of competition in these studies. Keywords: functional response, interference competition, Verticillium lecanii, Beauveria bassiana, Paecilomyces fumosoroseus, Encarsia formosa, Dicyphus hesperus, Aphidoletes aphidimyza Introduction In Canada, microbial insecticides based on entomopathogenic fungi have not yet been registered for use in greenhouses. As a consequence, biological control and IPM programs have developed that do not rely on these agents. Insecticide-based interventions have been increasing in greenhouse IPM (Gillespie, 2002). Many of these insecticides kill natural enemies and ultimately cause more damage to the crop. The need for insecticide approaches that can reduce pest populations without disrupting natural enemies of target or non-target pests is clear. Microbial insecticides based on entomopathogenic fungi might provide a biological control-based approach to the problem. There has been much discussion in the biological control literature in recent years regarding the combined effects of natural enemies on prey/pest populations (Brodeur et al. 2002, Ferguson & Stiling 1996, Rosenheim et al. 1995, Polis et al. 1989) and extensive literature on intraguild predation has developed as a result. Mutual predation and interference among natural enemy species used in biological control can cause pest mortality to be less than expected (e.g Fergusen & Stiling 1996). If the combined mortality is the sum of mortality from each acting alone, the enemies act in an additive fashion. If the sum of mortality is greater than additive, the enemies act synergistically, and if it is less, the enemies act in a non-additive fashion. The combined mortality from two natural enemies foraging for the same prey over different densities of prey could be described as a combined functional response (Soluk 1993). Fungi that are used in microbial insecticides tend, as a general rule, to be weak 3
16 pathogens. Applied as microbial insecticides, their interactions with the target pest are better described by a dose-response rather than a functional response. Application of a certain density of infectious units will result in a level of mortality that is relatively independent of pest density. The way in which entomopathogens interact with other natural enemies in biological control may be of interest for several reasons. In particular, the application of a microbial insecticide instantaneously changes the foraging impact of extant predators by killing, but not removing, part of the prey population. How this affects the predator population, and the efficacy of biological control is an important consideration (Hokkanen & Hajek 2003, and chapters therein). A theoretical perspective How should mortalities from an arthropod natural enemy and an entomopathogen combine? We can use Holling s disc equation (Holling, 1959) as a simple model of functional response in a predator to develop some predictions. a* N * T * P Na = (1) 1+ a* Th * N In the Holling Disc equation Na, the number of prey attacked, is a function of the attack rate (a), number of prey available (N), time (T), number of predators (P), and handling time (T h ). The number of prey attacked increases with attack rate and number of prey available, and asymptotes to a maximum value as handling time approaches total time available (Figure 1). 50 Prey Attacked Prey Available Figure 1. The relationship between prey available and prey attacked, derived using Holling's disc equation (a = 0.1; T = 10, P = 1 and Th = 0.5) For entomopathogens, the number of infected hosts, N i can simply be described by Ni = FN (2) i.e. a constant proportion of prey are killed regardless of host population size for a given dose, F. In an environment where a pathogen and an arthropod natural enemy are applied together the functional response of the latter must be adjusted to account for the presence of cadavers 4
17 resulting from the pathogen. Thus: a * NH * T * P Na = a + ( a * Th * NH) + ( a * Thc * N (3) i) where N H is the number of healthy prey (N-N i ), T hc is the handling time for cadavers. Even if T hc =0, and all other parameters are unchanged in the presence of the pathogen, the mortalities will not be perfectly additive. The combined effect is: a * NH * T * P Na = FN + (4) a + ( a * Th * NH) + ( a * Thc * Ni) This is illustrated for a pathogen mortality of 50% (Figure 2). This relationship only holds where there is sequential action, with the fungus attacking first and the predator second. Functional response with competition Mortality from a pathogen Functional response sum Hypothetical functional response with no competition A. Th cadavers = B. Th cadavers = 0 Prey Attacked Prey Attacked Prey Available Prey Available Figure 2. Functional responses based on a predator competing with a pathogen that causes 50% mortality in the prey (Equation 3 and 4). Equation parameters are as in Figure 1 except: A. Handling time for cadavers = handling time for prey = 0.5); B. Handling time for cadavers = 0 and handling time for prey = 0.5). In Fig 3A, the functional response of the predator is based on Equation 3, with handling time for cadavers identical to handling time for prey (solid line). The combined functional response (Dash-dot-dot line) is less than fully additive (dotted line). In Figure 3B, the handling time for cadavers is set to zero, which illustrates a condition where the predator discriminates perfectly between infected and healthy prey (solid line). The combined functional response approaches additive over the complete range of prey density, but is still less than the hypothetical fully additive functional response as a result of competition. In a simple biological control setting, such as in greenhouse agriculture, the phenomenon that we postulate should be a universal problem for predators. The number of prey attacked should be reduced by encounters with already-dead prey, and unproductive time (from a biological control perspective), should be spent handling these prey. Most studies that have examined the interactions between natural enemies postulate that mortalities should be fully additive, and that partial additivity suggests interference competition between the natural 5
18 enemies. We would argue that short-term interactions in a pathogen-predator biological control system should always result in partially additive mortality due to the obvious competition effects. Results from three microbial insecticide studies In order to illustrate the problems with additivity and the importance of interpreting results of pathogen-predator interactions, we summarize three recent studies. The first examined interactions between the pathogen Beauveria bassiana (Bals.-Criv.) Vuill. formulated as Botanigard, and the arthropod natural enemies Encarsia formosa (Gahan) and Dicyphus hesperus (Knight) simultaneously attacking the greenhouse whitefly, Trialeurodes vaporariorum (Westwood) on tomato (Labbe et al. in press, Labbé 2005). A second study examined the interaction between Paecilomyces fumosoroseus (Wize) Brown & Smith, formulated as PFR-97, and D. hesperus, simultaneously attacking greenhouse whitefly (Alma, 2005). The third study is currently investigating the interactions between Verticillium lecanii (Zimm.) Viégas, formulated as Mycotal, and Aphidoletes aphidimyza (Rondani) simultaneously attacking green peach aphid, Myzus persicae (Sulzer). We particularly focus on the ability of the predator to discriminate between infected and healthy prey, the potential for infection of the predator by the fungal disease, and the outcomes of experiments where the organisms interacted with the target prey. Beauveria bassiana D. hesperus fed on infected nymphs approximately 85% less frequently than healthy nymphs (Labbé 2005). Assuming that the actual time spent feeding on infected nymphs was similar to that spent feeding on healthy nymphs, the overall handling time for cadavers was less than that for healthy prey. It was possible to infect adults and nymphs of D. hesperus with B. bassiana. In a greenhouse trial, greenhouse whitefly populations were established on tomato plants in compartments with E. formosa and D. hesperus as biological control agents. Half of the compartments were treated three times with B. bassiana at 7 9 x10 7 conidia/ml, and mortality of whitefly due to the three natural enemies was measured over 8 weeks. Whitefly numbers were significantly lower in the presence of B. bassiana. There were no effects of B. bassiana on numbers of D. hesperus (all stages) or E. formosa (adults) found on the crop, although numbers of whitefly nymphs killed by D. hesperus were lower and percent parasitism by E. formosa was higher, in the presence of the fungus (Table 1). Table 1. Percent of nymphs of greenhouse whitefly that were killed by one of three natural enemies in the absence or presence of B. bassiana (Labbé et al. in press). Percent of whitefly nymphs Without B. bassiana With B. bassiana Killed by E. formosa Killed by D. hesperus Killed by B. bassiana E. formosa + B. bassiana 5% 6% 0.25% 0 % 7.5% 2.5% 10.5% 1% Mortality caused by the two arthropod natural enemies combined was very similar in compartments with and without B. bassiana, although partitioned differently. Applications of B. bassiana were almost additive with mortality from the two arthropod natural enemies. The mortality from B. bassiana was relatively low but there was a significant reduction in 6
19 mortality of whitefly in compartments relative to those with the arthropod natural enemies alone. Table 2. Numbers of whitefly in tomato leaf samples after 6 weeks of exposure to different natural enemy treatments (Alma, 2005). Whitefly nymphs Natural enemy treatment Control D. hesperus P. fumosoroseus Combined Paecilomyces fumosoroseus. The predator, D. hesperus, discriminated between uninfected prey and prey infected with P. fumosoroseus (Alma, 2005). Adult D. hesperus became infected with P. fumosoroseus when exposed to the disease on treated leaves, but not when treated by direct application of conidial solutions to the thorax. When foraging on leaves that were treated with P. fumosoroseus, adults of D. hesperus consumed fewer nymphs than on control leaves. Adults that died during this trial were excluded from the analysis, suggesting that either D. hesperus changed its foraging behaviour on treated leaves or that the fungal disease caused sub-lethal effects on the health of D. hesperus. The fact that adults can become infected when exposed to treated leaves suggests the latter. A 6 week trial was conducted, with three sequential applications of P. fumosoroseus at 18 x 10 7, 1 x 10 7, and 1 x 10 7 blastospores/ml. Both natural enemies produced significant mortality by the end of the trial, but the mortality was partially additive (Table 2). If mortalities had been additive, the combined treatment should have contained 15 whitefly nymphs per sample. Verticillium lecanii The predator, A. aphidimyza did not discriminate between infected and uninfected aphids (Jaramillo, unpublished data). Third instar larvae that were treated with conidial solutions of V. lecanii, then allowed to pupate, did not show any differences in emergence of adults, compared to untreated larvae. In a 30 day cage trial on pepper plants with green peach aphids (factorial design : +/- A. aphidimyza and +/- V. lecanii) aphid populations in the treatments with A. aphidimyza went extinct after 30 days. At 20 days, there were clearly additive effects of the application of the two natural enemies (Table 3). Under an assumption of additivity, the combined effects of the two natural enemies should have left 1085 aphids on plants with both natural enemies applied, clearly within the range of variation of the actual mean. Table 3. Numbers of aphids on pepper plants after 20 days of exposure to different natural enemy treatments (Jaramillo, unpublished data). Natural enemy treatment Control A. aphidimyza V. lecanii Both Numbers of 3343± ± ± ± aphids per plant Summary and Conclusions The predator D. hesperus appeared to reject prey that were infected with fungi. Where this 7
20 predator was used alone, mortality was partially additive, but the predator provided a significant increment of mortality in the presence of the fungal agent. In combination with E. formosa, mortality from D. hesperus was reduced in the presence of B. bassiana; however, an increase in the mortality due to E. formosa compensated for this. Overall mortality in this system of two natural enemies approached fully additive. In experiments with A. aphidimyza and V. lecanii, combined mortality agreed with the fully additive hypothesis. Why then have we questioned the additive hypothesis as the null expectation when natural enemies interact? In two of the systems we present, competition and sublethal effects clearly affected the efficacy of the predator, D. hesperus, but mortality from A. aphidimyza and V. lecanii was additive. Competition effects should result in partial additivity when an arthropod natural enemy and a pathogen affect the same prey. If this does not happen, understanding the mechanisms involved may help to improve biological control. Although A. aphidimyza was oblivious to infected prey, thus spending time killing already dead prey, it combined with V. lecanii in an additive fashion. Why should this occur? Does the numerical response of the predator compensate for the competition? Can the predator complete development on prey that are infected with the fungus? Are aphids more likely to drop from plants in the presence of fungus-infected conspecifics? The point is that science is not truly about answers it is about asking the right questions. References Alma, C. 2005: Intraguild interactions between two natural enemies of Trialeurodes vaporariorum (Homoptera: Aleyrodidae), the predator, Dicyphus hesperus (Heteroptera: Miridae), and the entomopathogenic fungus, Paecilomyces fumosoroseus Apopka-97 (Deuteromycotina: Hyphomycetes) (PFR-97 ). M.Sc. Thesis. Department of Biological Sciences, Simon Fraser University, Burnaby, British Columbia, Canada. Brodeur, J. Cloutier, C & Gillespie, D. 2002: Higher-order predators in greenhouse systems. IOBC/WPRS Bull. 24(1): Gillespie, D.R. 2002: Biological and integrated control in vegetables in British Columbia: The challenge of success. IOBC/WPRS Bull. 24(1): Hokkanen, H. & Hajak, A. (Eds.) 2003: Environmental Impacts of Microbial Insecticides: Needs and Methods for Risk Assessment. Kluwer Academic Publishers. Dordrecht, The Netherlands. Holling, C.S. 1959: Some characteristics of simple types of predation and parasitism. Can. Entomol. 91: Labbé, R. 2005: Intraguild interactions of the greenhouse whitefly natural enemies, predator Dicyphus hesperus, pathogen Beauveria bassiana and parasitoid Encarsia formosa. M.Sc. Thesis, Université Laval. Faculté des Sciences de l Agriculture et de l Alimentation. Québec, Québec, Canada. Labbé, RM., Cloutier, C & Brodeur, J. 2006: Prey selection by Dicyphus hesperus of infected or parasitized greenhouse whitefly. Biol. Control Sci. Technol. 16 (in press). Polis G.A., Myers C.A., & Holt R.D. 1989: The ecology and evolution of intraguild predation: potential competitors that eat each other. Annu. Rev. Ecol. Syst. 20: Rosenheim J.A., Kaya H.K., Ehler L.E., Marois J.J., & Jaffee B.A. 1995: Intraguild predation among biological control agents: theory and evidence. Biological control: theory and applications in pest management 5: Soluk, D.A. 1993: Multiple predator effects: predicting combined functional responses of stream fish and invertebrate predators. Ecology 74:
21 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Integrated pest and disease management for greenhouse-grown vegetable crops in Cyprus Nicolaos Ioannou, Nicos A. Seraphides Agricultural Research Institute, 1516 Nicosia, Cyprus Abstract: This study was carried out for three consecutive years at Zygí Experimental Station in Cyprus, with main objective to develop effective IPM programs for tomato, cucumber and pepper under greenhouse conditions. The study was carried out in two heated plastic greenhouses, one undergoing conventional pest management (CPM), based mainly on preventive chemical treatments applied on schedule, and the other under an IPM program, based primarily on the following elements: insect-proof construction of the greenhouse and mass trapping for managing insect pests and insectborne viruses, weekly monitoring of plants for pests and diseases, soil solarization for control of soilborne pathogens and pests and automatic mechanical regulation of the greenhouse environment for control of air-borne fungal and bacterial diseases. The main pests encountered in both greenhouses were the whitefly Bemicia tabaci, aphids and thrips. In the IPM greenhouse, insect populations were significantly lower compared to the CPM greenhouse. Soil-borne diseases were effectively controlled under both regimes. Similarly, no major virus or bacterial problems were encountered in either greenhouse. The most important fungal diseases were downey mildew on cucumber, and botrytis, sclerotinia and powdery mildew on all three crops. With the exception of powdery mildew, all airborne fungal diseases were effectively controlled in the IPM greenhouse, enabling a reduction of pesticide applications by about 50%, compared to the CPM greenhouse. Key words: IPM, tomato, cucumber, pepper Introduction Greenhouse vegetable production is an important agricultural sector in Cyprus, with a significant contribution to the Agricultural Gross Domestic Product of the island. Although greenhouses cover only 10% of the area in use for vegetable production, more than 20% of the total value of vegetable production is achieved in this small area. In the Mediterranean, the mild climate enables pests to develop year-round and pest pressure is, therefore, very high (Albajes et al., 1999). In the early years of vegetable greenhouse production, soil-borne disease problems were relatively simple, but they increased in importance as intensive cultivation continued (van Alebeek & van Lenteren, 1990). To cope with the above problems, farmers are obliged to make heavy use of pesticides with all negative consequences on human health, the environment and the sustainability of the plant protection practices themselves. In order to alleviate problems associated with heavy pesticide use in the greenhouse vegetable industry of Cyprus, the Agricultural Research Institute initiated the present research program aiming to develop, evaluate and introduce IPM programs for tomato, cucumber and pepper under greenhouse conditions. Materials and methods This study was conducted for three consecutive years in two old-technology plastic greenhouses, with similar crop history, located at Zygi Experimental Station (long.32 E, lat. 9
22 35 N) in Cyprus. One greenhouse was subjected to conventional pest management (CPM) practices and the other to an IPM program. The two greenhouses were planted to various tomato, cucumber and pepper varieties and for each crop there were four replicated plots with 150 plants each (i.e. a total of 600 plants/crop). CPM Greenhouse Conventional methods applied for control of pests and diseases included: a) soil fumigation with methyl bromide to control soil-borne pathogens and pests, b) preventive pesticide sprays applied on schedule and c) elementary regulation of greenhouse environment through a central heating system (circulation of hot water) and by manual regulation of the side openings (opened during the day and closed during the night). The fruit setting hormone β- naphthoxy acetic acid (ORTOMONE) was applied at 15-day intervals for pollination. IPM Greenhouse Integrated pest management practices included: a) a double-door entrance hall to control unwished insects access, b) fine-mesh screens on all openings to exclude insect pests, c) mass trapping of insect pests with 36 yellow sticky traps 25x30x0.3 cm, uniformly distributed in the greenhouse, d) limited application of biological control by releasing the parasitic wasp Diglyphus isaea to control the leafminer Liriomyza sp., e) soil solarization to control soilborne pathogens and pests, and f) advanced regulation of the greenhouse environment using a system identical to that of the CPM greenhouse, supplemented by an automatic RH control system, consisting of 4 computerized exhaust fans for better ventilation. The application of pesticides was based on pest and disease monitoring. For whiteflies the following action thresholds were used (Greer, 2000): a) for young plants, 0.5 whiteflies per trap per day and b) for mature plants, 2 whiteflies per trap per day. For disease control, infected plant parts were removed in order to reduce inoculum pressure. If necessary, the infected and neighbouring plants were sprayed with fungicides. Overall spays were applied only whenever the above measures failed. Bumblebees were used for pollination purposes. Records Environmental parameters (T and RH), both within and outside the two greenhouses, were continuously monitored through a computerized monitoring system. Six yellow sticky traps 25x30x0.3 cm (hanged cm higher from the top of the plants) and 12 plants for each crop (3 per plot) were used for monitoring the pest population densities on a weekly basis. Disease records were also taken at weekly intervals. In the case of bbotrytis and sclerotinia, disease incidence records were taken by counting separately the number of shoot/leaf infections and the number of infected fruits. The intensity of the disease (disease index) was calculated as the logarithm of the cumulative number of infections counted per 100 plants during the growing season. In the case of downey and powdery mildew, disease incidence was determined by counting the number of infected leaves per 100 plants. In addition, disease severity was rated on a scale from 1 to 10, based on the proportion of leaf surface affected by the disease. Disease intensity (disease index) was calculated as the logarithm of the product obtained by multiplying disease incidence by disease severity, using the following formula: D.I. = Log [(N 1 x 1) + (N 2 x2) +.. (N 10 x10)] where N 1, N 2.. N 10 the number of leaves (per 100 plants) with disease severity rating of 1, 2,
23 Results and Discussion ο 20 ΟΦΠ IPM ΣΦΠ CPM Out :00 01:00 13:00 01:00 13:00 01:00 13:00 01:00 13:00 01:00 13:00 01:00 Figure1. Ambient temperature variations inside and outside the IPM and CPM greenhouses RH% IPM RH% CPM RH% Out :00 01:00 13:00 01:00 13:00 01:00 13:00 01:00 13:00 01:00 13:00 01:00 Figure 2. Variations of relative humidity (RH) inside and outside the IPM and CPM greenhouses IPM CPM CM DEC JAN FEB MAR APR MAY JUNE Figure 3. Number of insect pests counted on plants inside the IPM and CPM greenhouses. 11
24 2 1,5 A) A) Sclerotinia Sclerotinia (shoot (shoot infection) infection) IPM CPM 1 0,5 0 2,5 2 B) Sclerotinia (fruit infection) IPM IPM CM CPM 1,5 1 0,5 0 2,5 2 C) Botrytis (leaf & shoot infection) IPM CM CPM 1,5 1 0, ,5 D) Botrytis (fruit infection) IPM CPM CM 2 1,5 1 0,5 0 YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 Tomato Cucumber Pepper Figure 4. Intensity (disease index) of Sclerotinia (A, B) and Botrytis (C, D) on greenhousegrown tomato, cucumber and pepper under conventional (CPM) or integrated pest management (IPM) practices. 12
25 4 3,5 3 A) Late Blight / Downy Mildew IPM CPM CM 2,5 2 1,5 1 0, ,5 4 3,5 3 2,5 2 1,5 1 0,5 B) Powdery Mildew IPM CPM CM 0 YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 Tomato Cucumber Pepper Figure 5. Disease index for Late Blight or Downey Mildew (A) and Powdery Mildew (B) on greenhouse-grown tomato, cucumber and pepper under conventional (CPM) or integrated pest management (IPM) practices IPM CPM CM YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 Tomato Cucumber Pepper Figure 6. Number of pesticide applications in greenhouse-grown tomato, cucumber and pepper under conventional (CPM) or integrated pest management (IPM) practices. The heating system was effective since the minimum night temperature was kept at around 15º C in both greenhouses (Figure 1). During the night, when greenhouses were closed, the relative humidity (RH) inside the CPM greenhouse approached 100% (saturation), while inside the IPM greenhouse was around 85% (Figure 2). 13
26 The main pests encountered in both greenhouses were the whitefly Bemicia tabaci, aphids and thrips. In the IPM greenhouse, insect populations were significantly lower compared to the CPM greenhouse (Figure 3). Soil-borne diseases were effectively controlled under both regimes. Similarly, no major virus or bacterial problems were encountered in either greenhouse. The most important fungal diseases were downey mildew on cucumber, and botrytis, sclerotinia and powdery mildew on all three crops (Figures 4 and 5). With the exception of powdery mildew, all air-borne fungal diseases were effectively controlled in the IPM greenhouse, enabling a reduction of pesticide applications by about 50%, compared to the CPM greenhouse (Figure 6). Powdery mildew was apparently favoured by lower relative humidity in the IPM greenhouse and may necessitate the installation of electric sulphur applicators. The marketable yield of all three crops was not affected by pest management practices (Figure 7) IPM CM CPM YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 YR 1 YR 2 YR 3 Tomato Cucumber Pepper Figure 7. Marketable yield of greenhouse-grown tomato, cucumber and pepper under conventional (CPM) or integrated pest management (IPM) practices. The results obtained in this study are immediately applicable in the agricultural practice of Cyprus, where over 90% of greenhouses are still of old technology, like the ones used in the present study. In recent years, however, the new greenhouses being built feature modern technology, with automated environmental control and hydroponic growing systems. Since this system is completely different from the one used in the present study, a new series of experiments were initiated with modern-technology greenhouses to develop and evaluate IPM programs under these conditions. References Albajes, R., Gullino, M.L., Lenteren, J.C. van & Elad, Y. eds.1999: Integrated Pest and Disease Management in Greenhouse Crops. Kluwer Publishers, Dordrecht. Greer, L, Integrated Pest Management for Greenhouse Crops. ATTRA, Florida p.12. van Alebeek, F.A.N. & van Lenteren, J.C. 1990: Integrated pest management for vegetables grown under protected cultivation in the Near East. FAO, part I and II. 14
27 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Status of biological and integrated pest control in CA&ES research greenhouses at UC Davis: Successes, challenges, and costs Garry Pearson, Robert Starnes, Thomas Costamagna, Ron Lane, Steve Silva, Ali Almedhi, Andy Rumsey, Michael Parrella College of Agricultural and Environmental Sciences, University of California, Davis, CA 95616, USA Abstract: UC Davis is the major land grant research and teaching university within the ten campus University of California system. With almost 1500 faculty, 31,000 undergraduate and 5,000 graduate students, requests for greenhouse space on the campus are varied and complicated. We recently centralized all greenhouse space (approximately 150,000 sq. ft.) with the intent of increasing availability of space in addition to standardized overall greenhouse maintenance, including pest control. A major objective has been to reduce pesticide use and increase the use of biological control. Greenhouse pests originate primarily from outdoors or are introduced through specialized research projects. Past methods of pest control have been the use of organophosphates, carbamates or other broad spectrum pesticides. The UC Davis Greenhouse Operations Staff has embarked on an ambitious program to use biological control in several key research and teaching greenhouse crops. Key words: Tomato, rice, tobacco, beans grapes, floral crops, aphids, thrips, mealy bug, whiteflies, natural enemies, and biological control agents Introduction The University of California at Davis is one of the largest plant research and teaching universities in the United States. Our departments of Plant Sciences, Viticulture and Enology, Land, Air and Water Resources, Nematology, Plant Pathology, Entomology, and Biological and Agricultural Engineering contain more than 150 faculty conducting research in the various areas of the plant sciences. The campus is located within the central region of California (about 80 miles east of San Francisco) where there is a Mediterranean climate. Summers are in excess of 35 C and winters can get down to -2 C. The research and teaching greenhouses total more than 150,000 sq. ft. We recently centralized all greenhouse space (approximate with the intent of increasing availability of space to faculty in addition to standardizing overall greenhouse maintenance, including pest control. There are 5 greenhouse complexes on the campus, each with its own superintendent, and these are spread amongst campus buildings and/or adjacent to research production land. Greenhouses were divided into different levels, based on the degree of environmental control (i.e., Level 1 with virtually no control to Level 5 with complete computer control). The charge to the greenhouse staff was to manage this space such that the wide variety of faculty needs for plant research and teaching could be satisfied. Greenhouses varied considerably in size (400 to 1800 sq. ft.) with more than 20 different crops grown. In addition, some faculty involved in breeding programs would keep their plant material for years, while those involved in floriculture bedding plant research would need the plant material for only a few weeks. A major focus has been to reduce the use of broad spectrum pesticides and to utilize more biological control across the myriad of crops, pests, research and teaching demands, and different levels of greenhouse space on the campus. Producing high quality plants for faculty research and teaching is the top priority for 15
28 the greenhouse managers. Most of the research done in greenhouses is used as a model for commercial field or greenhouse production and higher quality plant materials means the researchers can better extrapolate their results to the field. In addition, consistently high quality plants reinforce the quality of the scientific outcome. Material and methods Biological control in research greenhouse tomatoes, beans, and rice The main pests of these crops are Trialeurodes vaporariorim and occasionally Bemisia tabaci. Introduction of E. formosa & E. eremicus began with each cropping cycle and continued at weekly intervals until completion of the crop. These weekly releases, along with scouting techniques, good greenhouse management practices and spot treatments (using chemistry soft on beneficials) were successful in maintaining T. vaporariorim at low levels. Seasonal changes (movement of plant material and dropping temperatures) saw the reduction of the effectiveness of E. formosa & E. eremicus. Low populations of these pests were controlled by using 6 wide by 1500 foot rolls of yellow sticky tape and this was changed monthly. Another common pest was the western flower thrips, Frankliniealla occidentalis. Weekly releases of Amblyseius cumumeris when thrips first appeared in the greenhouse were made and this was continued weekly until the end of the crop cycle. Monitoring of this pest was done with yellow sticky cards, daily scouting and banker plants (see below). Biological control in research floral crops The UC Davis Greenhouse Staff grows and maintains a wide range of floral crops for research and teaching. The main pests of these crops are T. vaporariorim, F. occidentalis, Myzus persicae, Planococcus citri, Pseudococcos longispinus, P. maritimus and Tetranychus urticae. Introduction of E. formosa & E. eremicus, A cucumeris, Aphidius.colemani, Cryptolaemus montrouzieri, Leptomatix. dactylopii occurred in both small research greenhouses and multi-user greenhouses at the beginning of the crop cycles and continued until completion. Banker plants Introduction of banker plants as a technique for control of greenhouse pests was made in both small and large greenhouses. Fifty banker plants were established on a continued basis using Tagetes erecta (Dwarf Marigold var. Inka II) and Petunia hybrida (trailing Petunia var. Ramblin). The banker plants were spaced at 1/250 sq. ft. in the greenhouse. Hand watering and commercial fertilizers were used to maintain vigorous plants. Banker plants were infested with selected pests in a dedicated greenhouse and as the plants grew and pests developed, natural enemies were added. Banker plants were then introduced into greenhouses where the natural enemies could spread and provide control of pests on the targeted crops. Costs, labor and supplies. All costs associated with the introduction of biological control were maintained and documented. Greenhouse consolidation occurred a little over one year ago, and we are in the process of summarizing these data for the first year of operation. A portion of the technical staff time was allocated for the weekly ordering, monitoring, scouting, and distribution of biological control agents. Greenhouse managers in individual areas were responsible for communication to the technical staff regarding specific needs of the biological agents. Following up on the success of these biological control efforts and the possible intervention with additional pest control strategies was the responsibility of the individual manager. Initial training was done on life cycles of pests and biological control agents in addition to release techniques. This was achieved with the help of Plant Sciences, Inc. (Watsonville, CA), the 16
29 distributor for Koppert Inc. in California. Managers meet weekly to discuss overall greenhouse management in addition to the biological control program. See our website for more information. Results and discussion Successes Whiteflies, thrips, aphids and mites were easily controlled in the tomatoes, rice, beans and tobacco using the prescribed biological agents. Good greenhouse practices in addition to regular monitoring by greenhouse staff was critical in keeping these pests under control. Challenges The volatilization of sulfur for control of powdery mildew and bud mites on a variety of crops (e.g., grapes) grown in the greenhouses is an impediment to biological control. The greenhouse staff are working with university researchers to investigate other promising control measures that can reduce or eliminate the need for sulfur burning. The long tailed and grape mealy bugs proved difficult to control with the available biological control agents. Good greenhouse management practices helps to keep these pests under control, but we had to consistently rely on the use of insecticides (many of which are not compatible with natural enemies) for control. Acknowledgements We acknowledge the support of Plant Sciences, Inc, Koppert Biological Systems ( Goldsmith Seeds ( and Peaceful Valley Farm Supply ( We acknowledge the CA&ES Greenhouse Managers (Ron Lane, Steve Silva, Andy Rumsey and Ali Almehdi). This team has provided feedback and expertise in using biological agents. 17
30 18
31 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp A biological pest control story Juan Antonio Sanchez, Alfredo Lacasa Department of Biotechnology and Crop Protection, Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (IMIDA), C/Mayor s/n, La Alberca, Murcia, Spain. juana.sanchez23@carm.es Abstract: The use of biological pest control and IPM has quickly spread in pepper greenhouses in Campo de Cartagena (Spain) in the last ten years. IPM is currently applied in 90% (1,600 ha) of the greenhouse surface. The development of biological pest control in Campo de Cartagena is especially relevant for the whole Mediterranean area because it was possible despite of the high risk of pest outbreaks and the high incidence of insect-borne diseases (Frankliniella occidentalis-tomato spotted wilt virus, TSWV). The first trials to investigate the potential of native Orius spp. for controlling F. occidentalis in Campo de Cartagena were carried out in 1994 in experimental greenhouses at the IMIDA research station. In the following years, the use of natural enemies was optimised and the economic viability of the program was tested. Further assays were carried out to study the epidemiology of TSWV and to establish cultural practices to reduce the incidence of this virus. The adoption of IPM strategies greatly reduced the use of pesticides. The incidence of TSWV experienced a great reduction in the years following the establishment of biological pest control: the incidence of the virus in IPM greenhouses in 2002 and 2003 was 1.2% and 0.87%, respectively. Key words: biological pest control, IPM, pepper, Frankliniella occidentalis, Orius spp., Eretmocerus mundus, Neoseiulus californicus, TSWV. Introduction The successful development of biological pest control programs in greenhouse peppers in Campo de Cartagena has no precedent in the Mediterranean area. There are several aspects that make the development of biological control in this area especially interesting: (1) Biological pest control was established in spite of the high risk of pest outbreaks and the high incidence of insect-borne diseases (Frankliniella occidentalis (Pergande)-Tomato Spotted Wilt Virus, TSWV); (2) The adoption of new biocontrol strategies spread very quickly; in fewer than ten years almost the entire surface of protected pepper crops (about 1,600 ha) switched from chemical to pest control using natural enemies; (3) Some of the natural enemies (e.g. Neoseiulus (Amblyseius) californicus (McGregor) and Eretmocerus mundus Mercet) used to control key vegetable crop pests throughout the world were first collected for mass rearing and assayed for pest control on a large scale in this area. Campo de Cartagena is a flat area located along the coastal strip of the Murcia and Alicante regions. In this area, peppers grown under plastic greenhouses coexist with other vegetable crops (artichokes, lettuce, broccoli and celery), cucurbits (melon and watermelon) and citrus (lemon and orange). The highly heterogeneous landscape and the relatively open structures of the plastic greenhouses facilitate a high degree of interaction with other crops. Open fields and wild plants serve as pest and disease reservoirs during pepper crop-free periods and vice versa. However, the surrounding environment is also a great source of natural enemies that immigrate and establish in crops when the use of pesticides is reduced. Many stories could be told about the change in pest control strategies in pepper greenhouses in Campo de Cartagena. Here we are telling the story of a research group involved in 19
32 establishing the scientific bases of biological pest control for greenhouses grown peppers. Evolution of Biological Pest Control in Greenhouse pepper crops in Campo de Cartagena Before the introduction of western flower thrips (F. occidentalis) in about the mid-1980s, Thrips tabaci Lindeman was a secondary pest for pepper crops in southern Spain. The introduction of this exotic thrips occasioned a great disruption in the existing chemical pest control programs and truncated the first attempt to develop integrated pest management (IPM) programs. Neoseiulus (Amblyseius) cucumeris (Oudemans), which had started to be successfully used against T. tabaci, following experiences in northern Europe, completely failed to control F. occidentalis. Orius insidiosus (Say) was later tested with the same unsatisfactory results. Western flower thrips was a more serious pest than T. tabaci because of the greater amount of damage produced when it fed on leaves and fruits and, particularly, because of its great ability and efficacy to transmit Tomato spotted wilt virus (TSWV). We cannot be sure whether TSWV was present in the Iberian Peninsula before the appearance of F. occidentalis but, it is quite evident that TSWV started to spread soon after the introduction of western flower thrips (Lacasa, 1990). The peculiarities of the Mediterranean region made it necessary to develop ad hoc programs for greenhouse peppers based on native natural enemies. The aim of the first works was to catalogue the main predators and parasitoids of F. occidentalis. The anthocorids Orius laevigatus (Fieber) and Orius albidipennis (Reuter) were among the most common and abundant thrips predators (Lacasa et al., 1996). They occurred in many wild plants and spontaneously colonized pepper crops when pesticide applications were reduced, normally at the end of the growing season (Lacasa et al., 1996). Predatory thrips (Aeolothrips intermedius Bagnall and Aeolothrips tenuicornis Bagnall), mites [N. californicus, Neoseiulus barkeri (Hughes) and Euseius stipulatus (Athias-Henriot)], mirids [Deraeocoris punctulatus (Fallén)] and Eulophids (Ceranisus menes Walker) were other minor groups that helped to regulate F. occidentalis populations in crops and surrounding habitats. The first trials to investigate the potential of native Orius spp. in controlling F. occidentalis in Campo de Cartagena were carried out in 1994 in experimental greenhouses at the IMIDA research farm (Sanchez et al., 1995). The main conclusions drawn from these assays were: (1) O. laevigatus quickly established in the crop and reduced F. occidentalis population in a short time; (2) O. albidipennis spontaneously colonized the crop by the beginning of the summer and reached high densities; and (3) N. californicus immigrated to the crop and controlled Tetranychus urticae Koch. These preliminary assays formed the basis for the subsequent development of the pepper IPM program in Campo de Cartagena (Sanchez et al., 1995). Other assays were carried out in the following years in experimental and commercial greenhouses to optimise the use of natural enemies and to assess the economic viability of the program. The first assays in commercial greenhouses were carried out in two commercial greenhouses during the 1995/96 growing season (Sanchez et al., 1997a). O. laevigatus, O. albidipennis and N. cucumeris were used against F. occidentalis. N. cucumeris was released at the beginning of the season and during the winter months, when low temperatures made the establishment of O. laevigatus difficult. Preliminary trials showed that F. occidentalis could be controlled using natural enemies with efficacy levels similar to those obtained by chemical methods (Sanchez et al., 1997a). Further works characterized the biological parameters of O. laevigatus and O. albidipennis and established release rates in relation to F. occidentalis population dynamics (Sanchez & Lacasa, 2002). However, by the 20
33 mid-1990s the high incidence of TSWV still remained an obstacle to the development of IPM programs Surface (ha) Year Surface (%) Figure 1. Evolution of the area of pepper greenhouses under IPM in Campo de Cartagena. The study of TSWV epidemiology and the implementation of cultural practices to reduce the incidence of the virus were addressed in subsequent years. The incidence of TSWV depends on the immigration of viruliferous thrips and it spreads in the crop from primary foci. Thus, physical barriers that reduced thrips immigration and cultural practices aiming to reduce primary foci were expected to diminish TSWV incidence. Lacasa et al. (1994) had already proved that thrips-excluding meshes in lateral vents of greenhouses delayed thrips immigration and reduced the incidence of TWSV. In subsequent assays, Alcázar et al. (2000) concluded that, once TSWV was present in the crop, the virus mainly spread from primary foci. Removing infected plants greatly reduced the final incidence of the virus (Sanchez et al., 1999, Sanchez et al., 2000). The adoption of appropriate cultural practices and the use of resistant varieties, which were available from around 2000, considerably reduced the incidence of TSWV in pepper greenhouses. In the following years, the area of pepper greenhouses under IPM in the Campo de Cartagena region started to rapidly increase (Figure 1). The increase in pesticide applications to control F. occidentalis and the reduction of active ingredients following new European Union norms, encouraged growers to consider natural enemies as an alternative to pesticides for pest control. In less than ten years, almost the entire area switched from chemical to biological pest control programs (Figure 1). When we speak of biological pest control or IPM, we are referring to programs in which pest control is primarily based on predators and parasitoids, with little or no use of pesticides. A few species such as Macrosiphum euphorbiae (Thomas), Aulacorthum solani (Kaltenbach), Ostrinia nubilalis (Hübner) or Nezara viridula (L.) which were kept under control when chemical control was applied, became secondary pests after the generalised application of biological control. Bemisia tabaci Gennadius turned out to be a major pest for both chemical and IPM greenhouses by the end of the 1990s. The first attempts at controlling B. tabaci with the parasitoid Eretmocerus eremicus Rose & Zolnerowich did not provide satisfactory results (Fernández et al., 2003). The native parasitoid E. mundus frequently 21
34 immigrated to crops providing a good B. tabaci. The mass rearing and commercialisation of E. mundus made inoculative releases possible. In most cases this aphelinid provides a satisfactory control for B. tabaci (Calvo et al., 2002, Urbaneja et al., 2003, Fernández et al., 2003). Natural enemy and pest management in IPM pepper greenhouses is based on periodic sampling that supports decision-making. Sampling protocols used in research were highly time-consuming and inappropriate for commercial greenhouses. Binomial or presenceabsence methods were therefore developed for several pests and natural enemies (e.g. F. occidentalis, Orius spp., Amblyseius spp., B. tabaci, aphids) in order to reduce the cost of sampling without losing precision and accuracy in the estimates (Sanchez et al., 1997b, Guirao et al., 2004). The extension of the IPM program to commercial greenhouses took place towards the end of the 1990s. The regional agriculture agency (Consejería de Agricultura, Agua y Medio Ambiente de la Región de Murcia) took part in a project with the local growers association FECOAM (Federación de Cooperativas Agrarias de la Región de Murcia) to help transferring the IPM program from an experimental to a commercial scale. The aim of this collaboration was to train technicians and growers and to help solving problems that arose when applying IPM on a larger scale. Natural enemy producers also played a major role in training and implementing IPM in Campo de Cartagena. Many companies got involved at the sight of a good prospect for a natural enemy market that, with time, was going to be the biggest in the Mediterranean area. Outcome of the Biological and Integrated pest control programs One of the major outcomes of IPM was a great reduction in the use of pesticides in pepper greenhouses in Campo de Cartagena. In a survey involving conventional greenhouses that was carried out in 1998 and 1999, we found that, on average, crops were sprayed 27 times in a growing season using 2.5 active ingredients in each application (Sanchez et al. 2000); 33 of these active ingredients were against western flower thrips, 18 against B. tabaci, 4 against caterpillars (Spodoptera exigua (Hübner) and O. nubilalis), 0.5 against T. urticae and 5 were used to control fungal diseases [Botrytis cinerea Pers. and powdery mildew, Leveillula taurica (Lév.)]. In greenhouses under IPM, most of the sprays were used against caterpillars (8.5 with Bacillus thuringiensis Berliner) and to control fungal diseases. Reducing the number of spray applications in IPM has many advantages relating to the busy spray calendar of the conventional chemical control. (1) It reduces health risks for growers and consumers. Growers are much happier to release natural enemies than to handle chemical products. Releasing natural enemies is much safer and does not require all the safety measures associated with the use of chemical products. (2) It reduces the negative environmental impact on surrounding ecosystems. (3) Returning times are greatly reduced and this gives growers more freedom and confidence in harvesting and commercialising their produce. (4) Production in IPM greenhouses is generally higher than in greenhouses under chemical control: (i) pepper plants grow better when the intensity of such treatments is reduced; (ii) in parral type greenhouses, chemicals are normally applied from lateral openings using a spray cannon; in most crops, a gradient in plant growth can be observed from the lateral to the middle of the greenhouse due to the impact of applications at high pressure: the production of plants from the first three or four lateral rows is always very low; (iii) the growing season has extended by a few months since biological pest control is applied due to the lower incidence of TSWV. The incidence of TSWV experienced a great reduction in pepper greenhouses in the years following the establishment of biological pest control in Campo de Cartagena. In the mid- 22
35 1990s, many pepper crops were removed by the beginning of July and the incidence of TSWV in many greenhouses was close to a hundred percent at the end of the growing season. This situation had completely changed by the beginning of In a survey carried out in 2002, involving 96 randomly selected IPM greenhouses and 75 using chemical methods, the incidence of TSWV was 1.2% and 1.1%, respectively. Most of the pepper varieties used (92.8% and 98.7% for IPM and chemical greenhouses, respectively), were TSWV sensitive (e.g. Herminio, Spirit, Fiesta and Velez); TSWV-resistant varieties (e.g. Requena) only represented 7.2% and 1.3% for IPM and chemical greenhouses, respectively. In 2003, in a survey carried out in 174 IPM pepper greenhouses, the incidence of TSWV was 0.87% and the percentage of TSWV-resistant varieties was 10%. Although it is difficult about the factors responsible for the reduction in the incidence of TSWV in pepper crops, there are good reasons to believe that the change in pest control strategies may have played an important role. It is quite unlikely that resistant varieties were entirely responsible for the reduction in the incidence of the virus, among other reasons, because the area devoted to these varieties had been much smaller than that devoted to susceptible ones. The reduction of the population of viruliferous thrips at a global level might have been one of the main factors contributing to the fall in the impact of TSWV. In greenhouses under chemical control, western flower thrips populations reach high densities at the end of the growing season once pesticide applications stop. At the end of the growing season many plants may be infected with TSWV and young larvae feeding on these plants may produce abundant generations of viruliferous thrips. Adults may immigrate and infect susceptible crops as well as wild plants, which may serve as virus inoculum for the subsequent growing season. In IPM greenhouses high Orius spp. populations keep thrips density low at the end of the growing season. Thus, the immigration of infected thrips to potential hosts and the contribution to global TSWV inoculum is expected to be much lower for pepper crops under IPM than for those under chemical control. Future development Crops under biological pest control are dynamic systems and new problems will tend to arise that will make it necessary to adjust the program. One of the advantages of IPM is that periodic sampling provides information about the state and the evolution of the system. This information is used for immediate decision making, but may also be used to establish management strategies at broader geographical and temporal levels. Over the last few years we have been involved in integrating all of these data within a Geographical Information System (GIS) (Sanchez et al. 2005). This GIS may be a very useful tool for establishing management strategies for controlling pests and natural enemies in line with the local problems and the environmental conditions of each area. Acknowledgments We would like to thank all the colleagues who made it possible to carry out the experiments to establish the scientific bases of the IPM program in the Campo de Cartagena: Josefina Contreras, Federico García, Mercedes Lorca, Laura Gutiérrez, Mª Carmen Martínez, Jeronimo Torres, Antonio Alcázar, Pedro Fernández, Marta Miguel and Francisco Torró. References Alcázar, A, Sanchez, J.A., Lacasa, A., Gutiérrez, L., Fernández, P. & Oncina, M. 2000: Epidemiología del virus del bronceado del tomate (TSWV) en cultivos de pimiento en invernadero. Relación entre la incidencia de la virosis y las poblaciones de Frankliniella 23
36 occidentalis. X Congr. Soc. Española Fitopatol. Valencia 3-6 october Calvo, J., León, P., Giménez, A., Stansly P. & Urbaneja, A. 2002: Control biológico de Bemisia tabaci (Hom.: Aleyrodidae) en cultivo de pimiento en el Campo de Cartagena mediante sueltas de Eretmocerus mundus y E. eremicus (Hym.: Aphelinidae). Terralia 30: Fernández P., Miguel M., Lacasa, A. & Sanchez, J. A. 2003: Eretmocerus mundus and Eretmocerus eremicus (Hymenoptera: Aphelinidae) for Bemisia tabaci (Homoptera: Aleyrodidae) control on greenhouse pepper in the southeast of Spain. 3 rd International Bemisia Workshop, Barcelona March. Guirao, P., Fernández, P., Miguel, M. & Lacasa, A. 2004: Distribución espacial y muestreo de Bemisia tabaci en pimiento. Phytoma España 164: Lacasa, A. 1990: Un trienio de Frankliniella occidentalis (Pergande) en España. Evolución temporal y espacial de una plaga importada. Cuadernos Phytoma España, 6: Lacasa, A., Contreras, J., Sanchez, J.A., Lorca, M. & García, F. 1996: Ecology and natural enemies of Frankliniella occidentalis (Pergande 1895) in the southeast Spain. Folia Entomologica Hungarica. 57: Lacasa, A., Contreras, J., Torres, J., González, A., Martínez, M.C., García, F. & Hernández, A. 1994: Utilización de mallas en el control de Frankliniella occidentalis y el virus del bronceado del tomate en el pimiento en invernadero. Bol. San. Veg. Plagas 20: Sanchez, J.A., Contreras, J., Lacasa, A. & Lorca, M. 1995: Datos preliminares sobre la utilización de Orius laevigatus (Fieber) en el control de Frankliniella occidentalis (Pergande) en pimiento en invernadero. Phytoma España 68: Sanchez, J.A., García, F., Lacasa, A., Gutiérrez, L., Oncina, M., Contreras, J., Gómez, J. 1997a: Response of the Antocorids Orius laevigatus and Orius albidipennis and the Phytoseiid Amblyseius cucumeris for the control of Frankliniella occidentalis in commercial crops of sweet peppers in plastic houses in Murcia (Spain). IOBC/WPRS Bull. 20(4): Sanchez, J.A., Lacasa, a., Gutiérrez, L. & Contreras, J. 1997b: Distribution pattern and binomial samplig for Frankliniella occidentalis and Orius spp. in sweet pepper crops. IOBC/WPRS Bull. 20(4): Sanchez, J.A., Lacasa, A., Gutiérrez, l., Torres, J., González, A. & Alcázar, A. 1999: Efecto del arranque de las plantas con síntomas de Tomato Spotted Wilt Virus (TSWV) sobre la incidencia de la enfermedad y la producción en cultivos de pimiento en invernadero. Actas de Horticultura 26: Sanchez, J.A., Alcázar, A. Llamas, A. Lacasa, A. & Bielza P. 2000: Integrated pest management strategies in sweet pepper plastic houses in the Southeast of Spain. IOBC/WPRS Bull. 23(1): Sanchez, J.A. & Lacasa, A. 2002: Modelling population dynamics of Orius laevigatus and Orius albidipennis (Hemiptera: Anthocoridae) to optimize their use as Biological control agents of Frankliniella occidentalis. Bull. Entomol. Research 92: Sanchez, J.A., Cánovas, F. & Lacasa, A. 2005: GIS. Un sistema de información geográfica (SIG) para el manejo de enemigos naturales, plagas y enfermedades en los cultivos de pimiento del Campo de Cartagena (Murcia). Urbaneja, A., Stansly, P., Calvo, J., Beltrán, D., Lara L. & Van der Blom, J. 2003: Eretmocerus mundus: Control Biológico de Bemisia tabaci. Phytoma 144:
37 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Implementing a Geographical Information System (GIS) for pepper greenhouse natural enemy and pest management in Southeast Spain Fernando Cánovas, Alfredo Lacasa, Juan Antonio Sanchez Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (IMIDA), Department of Biotechnology and Crop Protection, c/ Mayor, s/n, La Alberca, Murcia, Spain. juana.sanchez23@carm.es Abstract: Biological pest control is applied in 90% of protected pepper crops in Campo de Cartagena (Southeast Spain). The quick transition from chemical to biological pest control methods was motivated in part by problems in controlling western flower thrips, Frankliniella occidentalis, using pesticides. Decision-making in greenhouses under biological pest control is based on periodical sampling that generates a great amount of information which can be stored and analysed using Geographical Information Systems (GIS). The aim of this work was to show the application of GIS in optimising management strategies in greenhouse pepper crops; Orius laevigatus release was analysed in relation to F. occidentalis and Orius spp. spatio-temporal population dynamics. Thrips and Orius spp. population abundance data were gathered weekly for greenhouses in a 64 square km area. Greenhouses were located using Global Positioning System (GPS) and satellite digital images. Inverse distance weighted (IDW) interpolation was performed to estimate F. occidentalis and Orius spp. incidence at non-sampling points. F. occidentalis and Orius spp. showed an uneven geographical abundance distribution through time. High F. occidentalis incidence (percentage of occupied flowers) was in correspondence with areas of late O. laevigatus establishment. O. laevigatus was realeased during April, May and June in areas were it was already successfully established. GIS may be used to optimise O. laevigatus releases according to population dynamics. Key words: Frankliniella occidentalis, Orius spp., integrated pest management, geographic information systems, GIS, spatial interpolation. Introduction Biological pest control and integrated pest management (IPM) strategies have been almost generally adopted in greenhouse pepper crops in Campo de Cartagena (Southeast Spain). In less than ten years 90% of protected pepper crops (about 1600 ha) switched from chemical to IPM and Biological pest control (Sanchez & Lacasa, 2006). The quick transition from chemical to IPM programs was due among other reasons to: (1) increasing difficulties to satisfactorily control western flower thrips [Frankliniella occidentalis (Pergande)] using pesticides and the high Tomato spotted wilt virus (TSWV) incidence; (2) restrictions in pesticide application following the European Union normatives. The change in pest control strategies brought in a new decision-making concept based on periodical sampling (Sanchez et al., 1997). F. occidentalis is the key pest and its populations are carefully monitored almost during the entire growing season. Orius laevigatus (Fieber) is a key predator released for F. occidentalis control according to thrips and Orius spp. population dynamics (Sanchez & Lacasa, 2002). Although only O. laevigatus is used in augmentative releases, Orius albidipennis (Reuter) spontaneously colonize the crops and it usually represents the most abundant species during summer months (Sanchez et al., 1997). This weekly sampling scheme generates a big amount of poorly utilised information as it is uniquely used for immediate decision-making purposes. The proper storage and analysis of 25
38 this information may revert in optimising IPM practices. Geographic Information Systems (GIS) may be used to store and analyse this information spatially (Liebhold et al., 1993). The aim of this work was to show how GIS can be used as a management tool to optimise IPM practices. In this work we analyse the spatio-temporal evolution of F. occidentalis and Orius spp. We also discuss O. laevigatus release strategies in relation to thrips and anthocorid population dynamics. Material and methods Area of study Campo the Cartagena is a flat area located along the coastal strip of Murcia and Alicante regions. The study area was circumscribed to a 8.5x7.5 km cell located in the northern part of Campo de Cartagena (Figure 2). Table 1. Number of greenhouses used in the analyses and number of greenhouses where Orius laevigatus was released during the first two weeks of each month. Month Jan Feb Mar Apr May Jun Week 1 st 2 nd 1 st 2 nd 1 st 2 nd 1 st 2 nd 1 st 2 nd 1 st Orius Thrips Orius releases Population dynamic information Orius laevigatus releases and F. occidentalis and Orius spp. abundance data (percentage of occupied flowers) data were gathered weekly by IPM technicians. Thrips and Orius spp. abundance for each greenhouse was given as the average of two weeks in each month. The average of all greenhouses was used to calculate the population dynamics of F. occidentalis and Orius spp. We used Orius spp. when describing population dynamics because O. albidipennis usually immigrates and establishes in pepper crops. Greenhouses were located using global positioning system (GPS) and satellite digital images (Figure 2). The number of greenhouses used in the analyses and O. laevigatus releases are specified in Table 1. Spatial interpolation Inverse distance weighted (IDW) interpolation was performed to estimate F. occidentalis and Orius spp. incidence at non-sampling points (100x100 m grid) using ArcView GIS (ESRI )- Spatial Analyst extension. All analysis were done using the R statistical software (R Development Core Team, 2004). Results and discussion Temporal evolution The percentage of occupied flowers by F. occidentalis progressively increased from January to mid May and slowly decreased thereafter (Figure 1). O. laevigatus population increased at the same rate as F. occidentalis until mid March and at a much higher rate in the following weeks; at the end of June almost 80% of the flowers were occupied with Orius spp. Most O. laevigatus 26
39 releases were done between January and the end of March (Figure 1), and only a quarter of the releases (28%) were carried out between April and mid June. Orius spp. and F. occidentalis population dynamics in Campo de Cartagena during 2005 was very characteristic and similar to what was observed in previous years (Sanchez et al., 1997). Western flower thrips is usually present in most greenhouses at the beginning of the growing season but its populations remain low during winter months due to low temperatures (Sanchez et al., 2000). In most greenhouses population outbreaks occur between mid April and the beginning of June (Sanchez et al., 2002). Figure 2. Population dynamics of F. occidentalis and Orius spp. in pepper greenhouses during The arrows indicate the moment of releases and the upper numbers the percentage of releases in relation to the total in the growing season. Spatio-temporal dynamics The spatio-temporal analysis showed an uneven geographical distribution of F. occidentalis and Orius spp. abundance over time (Figure 2). The abundance of F. occidentalis during January was null through the entire working area. At the beginning of February F. occidentalis was already present in the northern area, but its population remained stable until March. In April F. occidentalis spread through all the territory and hotspots were registered during May and June. The overall thrips incidence was higher for the northern than for the southern part (Figure 2). The first O. laevigatus establishment spots were observed in February. At the beginning of March, O. laevigatus was already established through a big area in the lower part of the map (Figure 2). In April the bug was present in all the territory but its abundance was higher in places with early establishment. The higher O. laevigatus flower occupancy (76-100%) was reached during June (Figure 2). 27
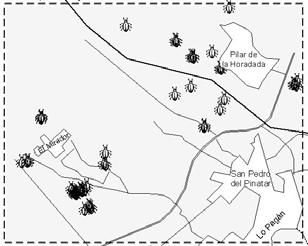
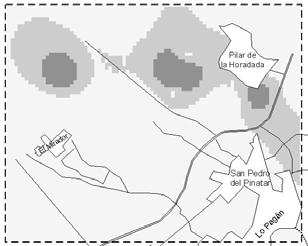
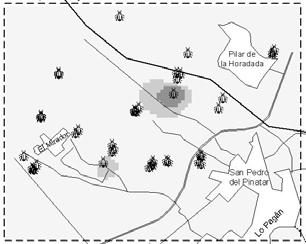
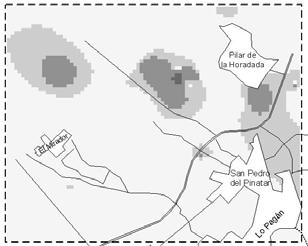
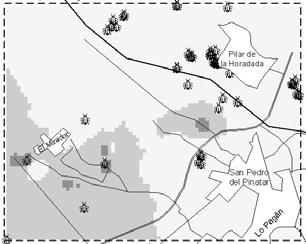
40 F.occidentalis January & Location map Orius spp. - January F.occidentalis - February Orius spp. - February F.occidentalis March Orius spp. - March Figure 2. 28


.")



41 F.occidentalis April Orius spp. - April F.occidentalis May Orius spp. - May F.occidentalis June Orius spp. - June Figure 2 (continued). Interpolated surfaces of Orius spp. and F. occidentalis abundance: average of the percentage of occupied flowers in the first two weeks of each month. The dashed square in the very first map shows the study area; dots represents all greenhouses used in spatial interpolations. 29
42 O. laevigatus release strategies Although O. laevigatus was abundantly released from January to May, timing was different from one area to the other (Figure 2). Early releases in January were effectuated in greenhouses through the entire territory. In February the intensity of releases was higher in the middle and lower part, while in March, it was more intense at the upper right corner. Most of the releases during April, May and June were done in areas where Orius spp. abundance was already high (Figure 2). The failure in early Orius laevigatus establishment and the absence of releases in thrips hotspots during February may have greatly determined the higher thrips incidence in greenhouses at the upper area (Figure 2). Releases in these areas should be optimised in order to accomplish a more efficient thrips control. The lack of establishment in some of the northern areas might as well be a consequence of bad crop practices interfering with the activity of Orius spp. During April, May and June most O. laevigatus was released in areas where it was already successfully established. Avoid releasing O. laevigatus in areas where it were already established would significantly reduce the cost of IPM programs. Acknowledgements We thank Modesto del Pino and Mª del Mar Davó for technical support. Pedro Fernández helped in positioning greenhouses. We thank Paco Torró and Antonio Oliver from SURINVER growers association for providing the data for the analyses. This work was funded by a joint project of IMIDA, FECOAM and Consejería de Agricultura y Agua de la Región de Murcia. References Liebhold, A.M., Rossi, R.E. & Kemp, W.P. 1993: Geostatistics and geographic information systems in applied insect ecology. Annu. Rev. Entomol. 38: R Development Core Team. 2004: R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Sanchez, J.A., Lacasa, A., Gutiérrez, L. & Contreras, J. 1997: Distribution pattern and binomial sampling for Frankliniella occidentalis and Orius spp. in sweet pepper crops. IOBC/WPRS Bull. 20(4): Sanchez, J.A., García, F., Lacasa, A., Gutiérrez, L., Oncina, M., Contreras, J., Gómez, J. 1997: Response of the antocorids Orius laevigatus and Orius albidipennis and the Phytoseiid Amblyseius cucumeris for the control of Frankliniella occidentalis in commercial crops of sweet peppers in plastic houses in Murcia (Spain). IOBC/WPRS Bull. 20(4): Sanchez, J.A., Alcázar, A, Llamas, A., Lacasa, A. & Bielza, P. 2000: Integrated pest management strategies in sweet pepper plastic houses in the southeast of Spain. IOBC/WPRS Bull. 23(1): Sanchez, J.A. & Lacasa, A. 2002: Modelling population dynamics of Orius laevigatus and Orius albidipennis (Hemiptera: Antochoridae) to optimize their use as Biological control agents of Frankliniella occidentalis (Thysanoptera: Thripidae). Bull. Entomol. Res. 92: Sanchez, J.A. & Lacasa, J.A. 2006: A biological pest control story. IOBC/WPRS Bull. In this volume. 30
43 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Mise en place d une protection biologique intégrée préventive contre les ravageurs des cultures de rosiers sous serre dans le sud de la France Jeannine Pizzol, Christine Poncet, Sandy Hector, Michel Ziegler INRA - URIH - 400, route des Chappes - BP Sophia-Antipolis Cedex - France - pizzol@antibes.inra.fr Résumé: Le thrips, Frankliniella occidentalis et l acarien, Tetranychus urticae sont les principaux ravageurs des cultures de rosiers sous serre, tout particulièrement en climat méditerranéen. Pendant l été 2005, une stratégie biologique préventive fondée sur une succession raisonnée de lâchers d auxiliaires a été testée sur rosiers en serre. Cette stratégie a permis d éviter tout traitement chimique contre les ravageurs sur cette culture, de début juin à décembre 2005, soit plus de six mois incluant la période de risque maximal. Ces résultats nécessitent d être confirmés dans différentes situations de production et la méthodologie employée peut être optimisée du point de vue du rapport coût/efficacité. Néanmoins, le niveau de protection déjà obtenu laisse entrevoir de réelles possibilités pour la mise en place d une telle stratégie sur le rosier en serre. Mots-clés: Protection intégrée, lâchers préventifs, Frankliniella occidentalis, Tetranychus urticae, Neoseiulus cucumeris, Neoseiulus californicus, Phytoseiulus persimilis, serre, rosier Introduction Bien que la protection intégrée sous serre ait démarré dans les années 1980 en France (Ramakers & Rabasse, 1995), la lutte chimique reste encore très dominante sur les cultures ornementales. Les deux principales raisons sont : une moindre pression des consommateurs puisque les produits ne sont pas destinés à être consommés et des seuils de tolérance beaucoup plus faibles dans la mesure où sont vendues des fleurs, des tiges et des feuilles et pas uniquement le fruit du végétal. Les risques pour la santé des travailleurs sous serre sont aujourd hui pris en compte par la profession et constituent le principal moteur d une orientation vers la protection intégrée. En France, environ 50 hectares de cultures ornementales sous serres sont actuellement conduits en Protection Intégrée et les surfaces ainsi protégées ont presque triplé de 1998 à 2001 (Astredhor, 2004). Les cultures ornementales sous serres sont sensibles à de nombreuses maladies et ravageurs (Poncet et al., 2003), en particulier aux thrips et aux acariens phytophages. Le thrips Frankliniella occidentalis Pergande, originaire de l ouest des Etats-Unis est l un des plus importants ravageurs sous serre en Europe (Van Lenteren & Loomans, 1998). Introduit en France depuis 1986 (Bournier & Bournier, 1987), il occasionne de nombreux dégâts sur cultures ornementales et maraîchères. Sur rosier, ce ravageur est à l origine de dégâts sur jeunes pousses (points jaunes et déformations), mais c est sur les fleurs que les dégâts sont les plus importants (Brun et al. 2004). En effet, les piqûres et les pontes entraînent des nécroses sur les pétales et des cicatrices sur les sépales, dépréciant ainsi les fleurs touchées qui deviennent impossibles à commercialiser. L autre ravageur majeur est l acarien Tetranychus urticae McGregor qui entraîne des décolorations ou jaunissements le long des nervures (Bertaux et al., 2003). Il est surtout présent sur la face inférieure des feuilles. Lors d attaques importantes, les fleurs et les jeunes pousses sont aussi touchées jusqu à la formation d un entoilage. 31
44 Pour lutter contre ces ravageurs, des acariens prédateurs, Neoseiulus (=Amblyseius) cucumeris Oudemans (prédateur de larves de thrips) et Neoseiulus (=Amblyseius) californicus McGregor (prédateur d acariens) sont préconisés. Lors de fortes attaques de T. urticae, un autre acarien prédateur Phytoseiulus persimilis Athias-Henriot peut être utilisé en traitement curatif (Pralavorio, 1976; Brun et al., 2005; Olivier et al., 2005). Ces acariens prédateurs sont distribués, soit en sachets (mini-élevages de 1000 N. cucumeris qui permettent chacun une diffusion progressive de 6000 à 7000 acariens), soit en vrac. Sur la base de lâchers préventifs d auxiliaires, nous avons pu protéger la culture de rosier sous serre insect-proof sans traitement chimique contre les ravageurs pendant l été et l automne Cet article présente les premiers résultats de cette stratégie de lutte et propose les étapes de son amélioration. Matériel et méthodes L étude a été réalisée dans une serre de 576m 2 comportant trois chapelles de 9,6 mètres sur 20 mètres, située sur le site de l INRA à Sophia-Antipolis (France Alpes Maritimes). Il n y a pas de séparation entre les trois chapelles contiguës. La serre de type Multiclair 9600 est couverte d un film Triclair-Inc 45, 200 microns. Elle est orientée Nord-Sud avec des ouvrants de type papillon en faîtière. La serre est équipée de filets «anti-bemisia» (Ref : 3306 : dimension des mailles 400 x 770 microns). Chaque chapelle comprend cinq rangs doubles de rosiers et sur chaque rang trois blocs, chaque bloc correspondant à un cultivar : cv. Magnum (Meilland), cv. Milva (Tantau), cv. Suela (Tantau). Les rosiers plantés le 6 mai 2003 sont cultivés hors sol, sur laine de roche de type Grodan, et conduits en coupe continue, avec un «poumon» (partie végétative maintenue à la base des plants). Dans chaque chapelle, les trois cultivars sont répartis en alternance (Suela, Magnum, Milva pour les rangs 1 à 5 ; Milva, Suela, Magnum pour les rangs 6 à 9 ; Magnum, Milva, Suela pour les rangs 10 à 15). Le climat et la fertirrigation sont gérés à l optimum par un ordinateur de serre (chauffage de l air par un système à circulation d eau chaude dans des tubes de type Alcoa, déshumidification par chauffage/aération, chauffage du substrat basse température, brumisation de type Coolnet). Des observations hebdomadaires ont été effectuées sur toute la période considérée. Pour le thrips, nous avons réalisé des frappages de roses à raison de 10 roses sur chaque rang soit 150 roses, ce qui nous a permis de suivre la dynamique de l infestation de la serre. Les paramètres suivants ont été notés : stade de la rose, présence de dégâts et nombre de thrips. Pour les autres ravageurs, des prélèvements hebdomadaires de 180 folioles ont été effectués de façon uniforme dans la serre (4 folioles par variété et par rang). Ces folioles ont été observées à la loupe binoculaire afin de détecter la présence des différents ravageurs et de les quantifier. Ces observations nous ont servi pour décider des traitements biologiques curatifs (ou chimiques s il y avait lieu) à réaliser. En effet, il a été décidé de lâcher de façon préventive et régulière N. cucumeris et N. californicus. Lorsque les échantillonnages montraient une augmentation au dessus du seuil de nuisibilité de ces ravageurs, des lâchers complémentaires de «nouveaux auxiliaires» pour le thrips, ou de P. persimilis (en curatif) pour les acariens, ont été réalisés. Sur ces bases, des lâchers réguliers de N. cucumeris ont été effectués : semaines 23, 26, 29, 32, 35, 38, 41 à la dose de 250 sachets (1 sachet/2m 2 ) pour chaque lâcher et semaines 44 et 46 à la dose 500 sachets (soit 1 sachet/m 2, dose préconisée par Olivier et al., 2005). Pour lutter contre les tétranyques, des lâchers de N. californicus, dose 4000 (semaines 25, 28, 31, 34, 37, 40, 43, 46 et 50) ont été réalisés. P. persimilis a été lâché en curatif à la dose 2000 (semaines 35 et 46) et dose 4000 (semaines 23 et 40). 32
45 Par ailleurs, la serre a été traitée contre l oïdium (Sphaerotheca pannosa) avec des lampes à soufre (soufre sublimé, 3 heures par nuit pendant toute la période). Trois fongicides ont également été appliqués contre l oïdium : myclobutanil (Systhane) le 30 septembre, bupirimate localisé au Nord Est de la serre (Nimrod) le 13 octobre et dodémorphe (Mehltaumittel) le 22 décembre. Résultats et discussion Lutte contre Frankliniella occidentalis Avec la stratégie utilisée, les populations de F. occidentalis ont été maintenues à un seuil inférieur ou égal à 0,5 thrips par fleur du mois de juin au mois de novembre 2005 (Figure 1), soit plus de six mois avec les lâchers d auxiliaires (N. cucumeris en préventif et avec des essais de Franklinothrips vespiformis Crawford et d Amblyseius swirskii Athias Henriot). L action combinée des traitements systématiques (N. cucumeris) et des interventions ciblées (F. vespiformis et A. swirskii) lors des fortes attaques de thrips a permis un bon contrôle de ce ravageur jusqu à l automne. Les interventions ciblées ont eu lieu lorsque des captures importantes (pièges jaunes englués) étaient enregistrées dans la serre (données non présentées ici) ou lorsque le seuil de 0,5 thrips par fleur était atteint. A l automne, nous avons observé une entrée massive de thrips dans la serre. Malgré des lâchers renforcés de N. cucumeris (1 sachet/m 2 ), semaines 44 et 46, nous n avons pas pu contrôler les fortes populations de thrips. Cette augmentation de la dose avait pour but de pallier leur faible dispersion (Vänninen & Linnamaki, 2002), mais cela n a pas permis de maintenir les populations en dessous du seuil de nuisibilité (0,5 à 1 thrips par fleur). Ceci confirme les résultats de Pizzol et al., (2005). Pendant la période allant de juin à novembre, les «nouveaux auxiliaires» (F. vespiformis et A. swirskii) ont contribué à maintenir les thrips à un niveau assez bas. Ces auxiliaires n ont pas été testés à l automne car il sont surtout actifs à des températures plus élevées que celles enregistrées dans cette serre à cette époque. Concernant les essais avec les «nouveaux auxiliaires» (F. vespiformis et A. swirskii), ils sont préliminaires et les résultats sur l efficacité relative de chacun des auxiliaires feront l objet d une note ultérieure après avoir été complétés. Cette forte infestation automnale est consécutive à une taille massive effectuée dans les autres serres du site. Notre culture était alors la seule culture de rose ayant un stade réceptif pour le thrips (roses aux stades pré-récolte et récolte). Lorsque le seuil de 1,5 thrips (larves et adultes), par fleur a été atteint, un traitement chimique (Vertimec, 50 ml/hl, matière active abamectine) a été fait (le 22 décembre 2005). Ce traitement chimique a été décidé pour ne pas laisser pendant l hiver des populations résiduelles trop importantes, auxquelles auraient pu s ajouter les nouvelles entrées au printemps suivant. 33
46 Nombre moyen de thrips/fleur.. 2,00 1,50 1,00 0,50 0, Semaines Thrips/fleur Seuil N. cucumeris Lâchers F. vespiformis et/ou A. swirskii Traitement chimique Figure 1. Dynamique des populations de Frankliniella occidentalis (sur fleurs) et lâchers d auxiliaires dans la serre (moyenne des 3 chapelles), Juin - Décembre 2005 Dynamics of Frankliniella occidentalis (on flowers) and releases of predatory mites in the integrated protection greenhouse (mean of the 3 compartments), June December 2005 Lutte contre Tetranychus urticae Lors de la prise en charge de tous les ravageurs dans la serre en juin (8/06/05), nous avions une attaque importante de tétranyques qui a pu être contrôlée avec des lâchers de N. californicus complétés par des lâchers de P. persimilis sur foyers (Figure 2). Les lâchers de N. californicus ont ensuite été effectués toutes les trois semaines. Des lâchers ponctuels et complémentaires de P. persimilis ont été faits sur des foyers décelés lors des contrôles hebdomadaires des folioles. Ces lâchers de P. persimilis étaient décidés après observation de foyers d acariens pendant deux à trois semaines consécutives. Les conditions qui doivent déclencher les lâchers curatifs sont à préciser. Les lâchers de P. persimilis étaient faits en priorité sur les foyers localisés. Le surplus était ensuite distribué de la manière la plus homogène possible sur l ensemble de la serre. Avec cette méthode et grâce à une brumisation adaptée, nous avons contrôlé de manière très satisfaisante T. urticae dans la serre pendant toute la période considérée. De Juin à fin Décembre la stratégie utilisée contre T. urticae a donné de très bons résultats, puisque l acarien a été parfaitement contrôlé (Figure 2). Lutte contre les autres ravageurs Les cochenilles. Des foyers localisés de la cochenille Coccus hesperidum L., ont été observés à trois reprises. Ces foyers ont été supprimés manuellement et aucun auxiliaire n a été lâché. Les aleurodes et pucerons. Dans cette serre, Trialeurodes vaporariorum, n a pas posé de problème en Nous n avons donc fait aucun lâcher d auxiliaire contre ce ravageur. De même, nous n avons pas eu d attaque par les pucerons sur cette culture de Juin à Décembre. Il faut rappeler que cette serre est équipée de filets «anti-bemisia» qui limite l entrée de certains ravageurs. 34
47 % de feuilles attaquées Semaines % de feuilles attaquées Lâchers de N. californicus Lâchers de P. persimilis Figure 2. Dynamique des populations de Tetranychus urticae sur rosiers et lâchers d auxiliaires dans la serre (moyenne des 3 chapelles), Juin - Décembre 2005 Dynamics of Tetranychus uticae (on roses) and releases of predatory mites in the integrated protection greenhouse (mean of the 3 compartments), June December 2005 Les lâchers préventifs d auxiliaires (N. cucumeris et N. californicus) complétés par des lâchers d auxiliaires ponctuels et curatifs ont permis un très bon contrôle des ravageurs pendant plus de six mois, notamment durant l été, période où le risque est maximal sur rosier dans le sud de la France. Cette protection uniquement biologique a été possible du fait que ces recherches ont été conduites dans une serre où la prophylaxie est maximale et où une brumisation adaptée a favorisé l installation des auxiliaires. Ce suivi est à poursuivre pendant plusieurs années pour confirmer les résultats. Il sera ensuite nécessaire d optimiser les doses et la fréquence des lâchers en fonction des périodes à risques pour chaque ravageur, afin de diminuer puis de déterminer les coûts de cette PBI (Protection Biologique Intégrée). De telles expérimentations montrent qu en floriculture il est possible d obtenir des résultats aussi bons que ceux obtenus en cultures maraîchères et de diminuer de manière importante le nombre de traitements chimiques sur cultures ornementales et plus particulièrement sur le rosier, notre modèle d étude. Remerciements Nous tenons à remercier M. J. M. Rabasse pour la relecture du document, M. J. F. Germain du L.N.P.V. de Montpellier pour la détermination des cochenilles et tous les techniciens des serres ou stagiaires (A. Bout, J. M. Drapier, P. Julien, A. Rimbaud, A. Settembrino), qui ont contribué à cet essai. Nous remercions également l Union Européenne (UE INTERREG III Alcotra France-Italie) et la Région PACA pour leur soutien financier. Bibliographie Astredhor, 2004: Maladies et ravageurs des cultures ornementales : Raisonner la protection des plantes. Actes des Journées Techniques Astredhor, 13 et 14 janvier 2004,
48 2004 : 224 pp. Bertaux F., Poncet C. & Pionnat J.C., 2003: Les maladies et parasites. In : Brun R., Mary L. La rose sous serre pour la fleur coupée. INRA Editions ASTREDHOR, Bournier, A. & Bournier, J.-P. 1987: L introduction en France d un nouveau ravageur : Frankliniella occidentalis. Phytoma, 388: Brun, R., Bertaux, F., Metay, C., Blanc, M.-L., Widzienkonski, C. & Nuée S. 2004: Stratégie de protection intégrée globale sur rosier de serre. PHM Revue Horticole, 461: Brun, R., Blanc, M. L., Metay C. & Wdziekonski C. 2005: Maîtrise des acariens en culture de rosier sous serre conduite en protection intégrée. PHM Revue Horticole, 465 : Olivier, N., Gros, P., Boujot, Y., Cambournac, L., Champoussin, J.-N., Francheschini, C., Lanza, R. & Paris, B. 2005: P. B. I. en culture de rose fleur coupée : bilan de 5 années d expérimentations dans les Alpes-Maritimes. PHM Revue Horticole, 469 : Pizzol, J., Brun, R., Marchal, C., Metay, C., Blanc, M. L. & Poncet, C. 2005: Efficacité de Neoseiulus cucumeris Oudemans sur les populations de thrips, Frankliniella occidentalis Pergande, en culture de rosiers sous serre. AFPP 2 ème Colloque International sur les Acariens des Cultures Montpellier Octobre 2005 : 9pp. Poncet, C., Brun, R. & Onesto, J.P. 2003: La production des Plants.. In : Brun R., Mary L. La rose sous serre pour la fleur coupée. INRA Editions ASTREDHOR, Pralavorio, M. 1976: Etude de mise au point d une méthode de lutte intégrée en serre de rosier. Bull. OILB / SROP, 1976 / 4, Ramakers, P. M. J. & Rabasse, J. M. 1995: I. P. M. in protected cultivation. In: R. Reuveni ed.: Novel approaches to integrated pest managment. Lewis Publishers, Boca Raton, Van Lenteren, J. C. & Loomans, A. J. M. 1998: Is there a natural ennemy good enough for biological control of thrips? Proceedings British Crop Protection Conference, Brighton UK, 1998, 5B-3: Vänninen, I. & Linnamaki, M. 2002: Performance of Neoseiulus cucumeris as a biocontrol agent of the Western Flower Thrips in cut roses. Bull. OILB/SROP 25(1): Preventive IPM for greenhouse roses in the South of France Abstract: Frankliniella occidentalis Pergande and Tetranychus urticae Koch are the most serious pests on roses in greenhouses, particularly under Mediterranean climate. During the summer of 2005, a preventive biological control strategy founded on a well-planned succession of releases of predatory mites was tested. This strategy made it possible to avoid any chemical treatment against these pests from the beginning of June until December, more than six months including the period of maximum risk. This result has to be confirmed in various situations and the control method has to be optimized for cost/efficiency ratio. Nevertheless, the level of protection already obtained shows real possibilities for using such a strategy on roses under glass. Key-words: Integrated Pest Management, preventive releases, Frankliniella occidentalis, Tetranychus urticae, Neoseiulus cucumeris, Neoseiulus californicus, Phytoseiulus persimilis, greenhouses, rose 36
49 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 p. 37 Reduction of pesticide use on plastic house cucumber in Varamin and Shahriar region of Tehran Province of Iran through Participatory IPPM Abbas Nasrollahi, Mohammad S. Moghaddam Department of Biotechnology, Iranian Research Organization for science & Technology, POBox Tehran Iran Abstract: Use of pesticides on greenhouse cucumbers in Iran has increased dramatically because of high infestation of diseases and pests, and due to lack of knowledge by growers. Use of IPM techniques by teaching the growers or selecting a strategy for pest control by extension workers usually does not work very well, unless the growers can see something such as use of biocontrol agents or etc. and then believe it. Even though, several thousands hectares of land are under greenhouse cultivation and the main crop is seedless cucumber. In general the average sizes of greenhouses are not very big and most of them are less than one hectare. Implementation of IPM programmes is a hard task because growers would not accept readily what you tell them, and they can not employ an agriculture specialist in their fields. For this reason we shifted from conventional methods to IPPM methods (Integrated Participatory Protection and Production Management) through FFS (Farmer Field Schools) and we followed the following criteria: 1) Production of healthy crop; 2) Protection of ecosystem by stakeholders; 3) Empowerment of stakeholders to understand what is going on in the field where they work; 4) Growers become experts. In the process of the work one plant protection specialist were assigned as facilitator and then we chose several agricultural graduates to do the job as TOTs (training of trainers) under the supervision of a facilitator and at the same time they could work in greenhouses to disseminate the IPPM knowledge to the growers. Greenhouse structure in these regions usually is not standard and ventilation does not take place very efficiently. Powdery mildew and downy mildew are big problems, and impose sever loses. Growers have to spray many times to overcome these losses. In the Varamin region growers usually do not do soil testing and rarely soil sterilization. They sow the seeds directly in the soil, but by implementing transplanting procedure damping off will be reduced in this way by almost 90%. Therefore, the use of fungicides will be eliminated except? in severe cases because they usually will use sterile soil in transplanting. In general, the problem which now exists throughout Iran is the dependency of all growers on chemicals pesticides for greenhouse crop protection, which poses severe contamination on harvested crops that can be eliminated to large extent by just only educating the growers. The mentioned problems are true for all regions in Iran, but nowadays people are realizing the danger of chemical pesticide residues in their food and the demand is toward chemical-free? goods. We believe the best solution for cutting pesticide use at the present time is to educate growers and let them to experiment and then believe they decide then proper measure for controlling the pests and diseases by IPPM method in greenhouses. Key words: IPM, IPPM, Farmers Field School, greenhouse cucumbers, extension 37
50 38
51 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Mechanical distribution of beneficials: laboratory tests Giovanna Tropea Garzia 1, Lucia Zappalà 1, Gaetano Siscaro 1, Giacomo Blandini 2, Sabina Failla 2, Giuseppe Manetto 2 1 Dipartimento di Scienze e Tecnologie Fitosanitarie, University of Catania, Via Santa Sofia 100, Catania, 2 Dipartimento di Ingegneria agraria, University of Catania, Via Santa Sofia 100, Catania, Italy. Abstract: A first version prototype was projected and evaluated, in order to mechanically release Phytoseiulus persimilis and Orius laevigatus, commonly employed in biological and integrated control programs on protected vegetable crops. This machine was used to release the beneficials which where placed, together with the carrier material present inside the bottles (vermiculite and buckwheat husk), into a hopper equipped with a distributor. Preliminary trials were conducted under laboratory conditions to evaluate the vitality and the damages eventually caused to the predators and to determine the range of action of the machine at different working rates. No physical damages to the arthropods limiting their mobility and vitality were observed. Also the distribution, evaluated on a horizontal surface, seems to be suitable for applications in biological control programs. Key words: biological control, plant protection machines, beneficials, laboratory Introduction Biological control of agricultural pests involves seasonal inoculative or inundative releases of natural enemies; this strategy turned out to be effective both in conditioned and cold greenhouses of northern and southern Italy. Among the various antagonists employed on horticultural crops, the predatory mite Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae) and the bug Orius laevigatus (Fieber) (Hemiptera: Anthocoridae) are valid instruments to keep the two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) and the western flower thrips Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) under control. Given the success in several experiments, these predators are reared and distributed by commercial insectaries. The manual-sprinkling method on infested crops is time consuming and doesn t allow uniform spreading. Studies on potential mechanization of these processes are rather limited. On extensive crops release tests of natural enemies (phytoseiids, chrysopids and trichogrammatids) were conducted using small planes (Pickett et al., 1987; Maini et al., 1988; Drukker et al., 1993) or air-flow devices carried by tractors (Giles et al., 1995; Morisawa & Giles, 1995; Gardner & Giles, 1996; Gardner & Giles, 1997; Wunderlich & Giles, 1999). More recently, mechanical blowers were tested to release natural enemies in protected crops (Gill et al., 1999; Pezzi et al., 2002; Van Driesche et al., 2002; Opit et al., 2005). The results of preliminary laboratory tests on beneficials mechanical release using a new kind of device are reported. 39
52 Material and methods First laboratory prototype A detailed description of the prototype cannot be supplied because of the patent procedure started by action of the University of Catania.The prototype used in the laboratory tests was mounted on a frame, keeping a fixed location and standard distribution conditions of the organisms and their carrier material. The device is equipped with a hopper, provided with a distributor which regulates the quantity of product to be dispensed. The reservoir is placed above the releasing component which is positioned at a height of 1050 mm from the ground and is actioned by a motor using a belt-driven transmission system, which allows working conditions to be modify. A 3 speed gear was projected in order to obtain corresponding rotational velocities of the gear shaft [600, 1100 and 1600 revolutions/min (rpm)]. In the text the authors will refer to these speeds to indicate the different working conditions of the prototype, although no trials were conducted at 1600 rpm, considered too extreme. Laboratory tests The mechanical distribution tests were carried out using the predatory mite P. persimilis and the anthocorid O. laevigatus. Both beneficials were purchased from a single commercial insectary (Bioplanet S.c. a r.l., Martorano di Cesena-FC, Italy), in 500 cm 3 plastic bottles. P. persimilis is commercialized in two different kinds of bottles said to contain 1000 (used in the trials) or 2000 mites with vermiculite as dispersal material in both cases. O. laevigatus is shipped in a mixture of vermiculite (20%) and buckwheat husk (80%), each bottle containing approximately 500 specimens. The humidity rate of vermiculite is around 40%, and this parameter was taken into consideration during the distribution tests because variations in moisture contents may affect the performance of the dispensing system. Preliminary observations were conducted per each shipment, aiming at defining some quality parameters of the employed antagonists (quantity, mortality, sex-ratio, fecundity and longevity) and to verify their suitability to the tests, following the procedures described in the guidelines developed by the IOBC working group on Quality Control of Mass-Reared Arthropods (AMRQC), the International Biocontrol Manufacturers Association (IBMA) and the Association of Natural Bio-control Producers (ANBP) and recently published (van Lenteren et al., 2003). The effects of the mechanical distribution, using the above described prototype, were evaluated using 150 cm 3 of product (carrier material plus natural enemies) per each trial. Mortality of the antagonists was firstly assessed both as they came out from the hopper and after the release at two working conditions corresponding to different rotational speeds of the gear shaft (600 and 1100 rpm). In the trials concerning the effects of the hopper, the material was collected into a container, while the release tests were conducted spreading the product on a horizontal surface (140 cm wide and 180 cm long), placed 40 cm below the level of the release device. The material was observed under a dissecting microscope to record the number of live and dead individuals and to highlight the presence and degree of eventual physical damages caused by the mechanical release. Subsequent tests were carried out to evaluate the quantity of beneficials collected after the release and their distribution on the same surface employed in the previous trials. This was subdivided into 15 sectors (5 contiguous rows made of 3 sectors each) (Figure 1) using rectangular plastic trays (36 47 cm) for collecting. In P. persimilis release tests the trays contained a thin layer of water in order to make the collection of both carrier material and beneficials more effective. In this case, the contents of each tray were filtered using a siever (80 mesh) and the mites present in the separated material were then counted under a dissecting microscope. 40
53 Further observations were conducted using the dispersal substrate without the beneficials and the trays without the water film, in order to evaluate the amount of material distributed outside the surface considered. 140 cm cm Figure 1. Design and size of the horizontal release surface with reference numbers of the sectors (the arrow indicates the exit of the beneficials from the prototype). Results and discussion Phytoseiulus persimilis The number of mites present inside each bottle varied in relation to the shipping data and the bottle itself. An average number of 1550 mites per container was calculated, with the only exception of one bottle which contained a lower number than indicated on the label. These starting data were used to define the quantity of beneficials released with the volume used in each trial. Also the natural mortality rate observed in the bottles received from the commercial insectary was considerably different, varying between 7.90 and 26.81%. Preliminary tests on fecundity and longevity of the beneficials didn t show values significantly different from the standards, making them suitable to be employed in laboratory trials. Observations on the integrity and vitality conditions of natural enemies in all the phases of mechanical distibution were successful: no physical damage was noted on live specimens, both out of the hopper and after the release. Mortality levels appeared similar to those observed in the bottles, reaching 20.7% out of the hopper and 24.7% after the release at 1100 rpm; only at 600 rpm the value observed was higher (28%) compared to the initial maximum level. Only the data on distribution at the lowest velocity (600 rpm) are reported, because of the high dispersion of carrier material outside the horizontal surface, otherwise observed. Moreover, at both speeds, due to the particle-size of the substrate and the different mass of the beneficials versus the inert material, the mites together with the lighter fraction of vermiculite landed quite close to the prototype reaching a maximum distance of about 40 cm (Table 1). The amount of individuals was lower compared to the quantities released (Table 2). On the contrary, the carrier material was distributed more uniformly on the whole surface, mainly concentrated between 40 and 140 cm (Table 1); also in this case, a difference was registered in the quantity of vermiculite distributed and collected on the horizontal surface (Table 3). 41
54 Orius laevigatus The mean number of O. laevigatus observed was equal to 466 individuals per bottle. The population was formed by adults (79.70%) and young stages (20.30%) and the mean mortality rate was 3.35%. The sex-ratio recorded was equal to 47 and 53, similarly to what reported in the literature. Fecundity and longevity tests didn t show significant differences from the standard values. The distribution tests highlighted no physical damage out of the hopper and after the release on the horizontal surface both on young stages (with a soft integument) and adults (winged and therefore more susceptible to be harmed). Out of the hopper a mortality level of 8% was registered, while after the release at 600 rpm a rate of 7% was recorded; no mortality was observed at 1100 rpm. A clear difference was seen in the horizontal distribution of insects and carrier material, at both working conditions. At the lower velocity the beneficials were concentrated in sectors 4 to 8, while at 1100 rpm some individuals were found up to 140 cm (Table 1). At the higher speed, the number of specimens collected after the mechanical release (63) is much lower than the assumed one (175), at 600 rpm a reduced loss of individuals was observed (Table 2). Instead, the carrier material was always found on all the whole horizontal investigated area (Table 1); no relevant difference appeared between the quantity of substrate collected and distributed, at 600 and 1100 rpm (Table 3). Table 1. Amount of predators and carrier material collected on the surface after the releases. Species (n.) Carrier material (g) Vermiculit Buckwheat Sector P. persimilis O. laevigatus e husk+vermiculite 600 rpm 600 rpm 1100 rpm 600 rpm 600 rpm 1100 rpm Total
55 Table 2. Number of individuals distributed with 150 cm 3 of carrier material and collected after the releases. Species rpm distributed (n.) collected (n.) P. persimilis O. laevigatus O. laevigatus Table 3. Quantity of carrier material distributed and collected on the horizontal surface. Carrier material rpm distributed (g) collected (g) Vermiculite Buckwheat husk+vermiculite Buckwheat husk+vermiculite Dosing and distributing mechanisms of this first laboratory prototype, as well as the release system of the product (carrier material plus beneficials) appeared suitable for the use in greenhouses.moreover, the possibility of employing an electric motor, with low absorbed power, makes the machine easier to handle and minimizes the costs as well as the environmental impact. In laboratory tests the two antagonists didn t show any damage, both going through the hopper and after the mechanical distribution. Although the higher velocity didn t cause any problem to the arthropods, this working condition in protected crops could involve the spreading of vermiculite (without beneficials) on a larger surface while the animal organisms would fall always in the same area close to the machine. In fact, 90% P. persimilis was distributed in the first cm near the machine at both velocities, while O. laevigatus reached higher distances (nearly 100 cm at the lower speed and 140 cm at the other one). The carrier material, indeed, was distributed more uniformly at both working conditions. This is probably due to the different mass of the beneficials and the substrate that influences the machine range on horizontal plan. The number of individuals collected after the release on the surface was low if compared to the distributed one, especially in the case of the anthocorid for which, at the higher speed, a 64% decrease in the theorical number of individuals released was recorded. Nevertheless, the differences observed between the quantity of carrier material and beneficials distributed and collected on the horizontal surface didn t correspond to a real loss of antagonists, but to a distribution on a wider area, outside the investigated one. Therefore, the tests carried out up to now suggest to employ the lower working velocity, that seems optimal for both antagonists, in order to limit the waste of biological products and obtain an effective distribution. Further trials will be carried out to better orientate the machine throw, to dose the quantities of product and to adapt the prototype to be used directly in greenhouses. Acknowledgements The project is supported with funds from Italian Ministry of Education, University and Research (PRIN 2004) and is entitled Mechanical distribution of arthropods in biological 43
56 control of protected crops. The authors equally contributed to the work with reference to the own specific competence. References Drukker, B., Yaninek, J.S. & Herren, H.R. 1993: A packaging and delivery system for aerial release of Phytoseiidae for biological control. Exp. Appl. Acarol. 17: Gardner, J. & Giles, D.K. 1996: Handling and environmental effects on viability of mechanically dispensed Green Lacewing eggs. Biological Control 7: Gardner, J. & Giles, D.K. 1997: Mechanical distribution of Chrysoperla rufilabris and Trichogramma pretiosum: survival and uniformity of discharge after spray dispersal in an aqueous suspension. Biological Control 8: Giles, D.K., Gardner, J. & Studer, H.E. 1995: Mechanical release of predacious mites for biological pest control in stawberries. Transaction of the ASAE 38 (5): Gill, S., Reeser, R. & Speaker, J. 1999: Can biological control of thrips work in a spring bedding plant crop? Maryland Cooperative Extension Website: Lenteren, J.C. van, Hale, A., Klapwijk, J.N., Schelt, J. van & Steinberg, S. 2003: Guidelines for quality control of commercially produced natural enemies. In: Quality Control and Production of Biological Control Agents. Theory and Testing Procedures, Cap. 19, J. C. van Lenteren ed., CABI Publishing, Wallingford, Oxon, UK: Maini, S., Gattavecchia, C. & Libé, A. 1988: Impiego di velivolo ultraleggero per i lanci di Trichogramma maidis Pint. e Voeg. contro Ostrinia nubilalis (Hb.). Atti Giornate Fitopatologiche 2: Morisawa, T. & Giles, D.K 1995: Effects of mechanical handling on Green Lacewing Larvae (Chrysoperla rufilabris). Transaction of the ASAE 11: Opit, G.P., Nechols, J.R., Margolies, D.C. & Williams, K.A. 2005: Survival, horizontal distribution, and economics of releasing predatory mites (Acari: Phytoseiidae) using mechanical blowers. Biological Control 33: Pezzi, F., Rondelli, V. & Baraldi, G. 2002: Mechanical distribution of Phytoseiids in greenhouse crops. Rivista di Ingegneria Agraria 33 (3): Pickett, C.H., Gilstrap, F.E., Morrison, R.K. & Bouse, L.F. 1987: Release of predatory mites (Acari: Phytoseiidae) by aircraft for the biological control of spider mites (Acari: Tetranychidae) infesting corns. J. Econ. Entomol. 80 (4): Van Driesche, R.G., Lyon, S., Sanderson, J., Smith, T., Lopes, P., MacAvery, S., Rusinek, T. & Couch, G. 2002: Greenhouse trials in Massachusetts and New York with Amblyseius cucumeris: effects of formulation and mechanical application. IOBC/WPRS Bull 25 (1): Wunderlich, L.R. & Giles, D.K. 1999: Field assessment of adhesion and hatch of Chrysoperla eggs mechanically applied in liquid carriers. Biological Control 14:
57 Whiteflies 45
58 46
59 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Bumble bees: An effective delivery system for microbial control agents for arthropod pest and disease management Les Shipp, Jean Pierre Kapongo, Peter Kevan, John Sutton, Bruce Broadbent Agriculture and Agri-Food Canada, Harrow, Ontario, Canada N0R 1G0; University of Guelph, Guelph, Ontario, Canada N1G 2W1; Agriculture and Agri-Food Canada, London, Ontario, Canada N5V Abstract: Greenhouse trials were conducted to determine the optimal concentration of Beauveria bassiana (Botanigard 22WP formulation) as vectored by the bumble bee (Bombus impatiens) pollinator for control of greenhouse sweet pepper pests (tarnished plant bug, Lygus lineolaris and green peach aphid, Myzus persicae) and the tomato pest (greenhouse whitefly, Trialeurodes vaporariorum), and the impact of Beauveria on B. impatiens. Three concentrations of Beauveria: low (9x10 9 ), mid (6.24x10 10 ), and high (2x10 11 ) conidia/g of inoculum and two control treatments were evaluated. Infection levels in all pests species by Beauveria were the greatest at the two higher concentrations of Beauveria. However, bee mortality was significantly greater at the highest Beauveria concentration (2.0 x conidia/g of inoculum). Thus, the concentration of 6.24 x Beauveria conidia/g of inoculum provided optimal pest control and had the least negative impact on the bees. Also, B. impatiens demonstrated the ability to effectively disseminate two fungal control agents (B. bassiana and Clonostachys rosea) simultaneously for both insect pest and plant pathogen control. Key words: bee-vectored microbial control agents, Beauveria bassiana, Clonostachys rosea Introduction In the 1990 s, honey bees were evaluated as vectors of fungal control agents against gray mold on strawberries in Ontario, Canada (Peng et al., 1992) and later, against the same pathogen on raspberries using both honey bees and bumble bees (Yu & Sutton, 1997). This technology was also tested using the antagonistic fungus Trichoderma harzianum against grey mold on strawberries with honey bees in Italy (Maccagnani et al., 1999) and with honey bees and bumbles in the USA (Kovach et al., 2000). The first report of using bee pollinators to deliver fungal biocontrol agents against insect pests was by Butt et al. (1998) who showed that honey bees effectively vectored Metarhizium anisopliae to canola for the control of the pollen beetle (Meligethes aeneus) in the UK. In greenhouse vegetable production, Al-mazra awi et al. (2006) showed that bumble bees (Bombus impatiens) can disseminate Beauveria bassiana from hive-mounted dispensers to greenhouse sweet pepper plants for control of tarnished plant bug, Lygus lineolaris, and western flower thrips, Frankliniella occidentalis. A concentration of 1x10 9 colony forming units (CFU) of B. bassiana/g of inoculum resulted in infection levels of 34-45% for L. lineolaris and 34-40% for F. occidentalis. The present study determined the optimal concentration of the commercial formulation of B. bassiana (Botanigard 22WP ) for control of insect pests on greenhouse sweet pepper and tomato. In addition, greenhouse trials were conducted to determine if bumble bees can simultaneously vector both an entomopathogenic control agent (Beauveria) and a plant fungal 47
60 agent (Clonostachys rosea) for pest control (Lygus) and disease suppression (grey mold) on sweet pepper. Material and methods Beauveria concentration trials The trials were conducted in two greenhouse compartments (7 x 13 m) with two or three finemesh screened cages (520 x 240 x 220 cm high) per greenhouse. Each cage contained 64 potted sweet pepper or tomato plants with a minimum of two flower sets per plant. The cages were maintained at o C and 80-85% RH throughout the trials. A completely randomized design was applied for the five treatments with four replicates (sweet pepper) and five replicates (tomatoes) over time. The treatments were: low concentration: 9x10 9 conidia/g of inoculum and one hive of 50 bumble bee workers, mid concentration: 6.24x10 10 conidia/g and one hive, high concentration: 2x10 11 conidia/g and one hive, heat inactivated Beauveria and one hive, and a control treatment where neither Beauveria nor bumble bees were present. Each colony of bumble bees was introduced 1 day after the plants were placed in the cages to allow acclimation to their hive and the cage. On day 2, 256 adult L. lineolaris and 960 green peach aphids, Myzus persicae, were released per treatment for the pepper trials and 1000 adult greenhouse whiteflies, Trialeurodes vaporariorum, in the tomato trials. Wooden dispensers filled with the appropriate inoculum (BotaniGard 22WP and corn flour mixture) were placed in front of the hives on day 3. The first sample (50 adult Lygus, T. vaporariorum and M. persicae) was collected on day 6 after which the dispensers were removed. The dispensers were refilled and replaced day 10 and a second sample was collected 3 days later. Five bees were also sampled from each treatment at each sampling date to determine the amount of Beauveria that the bees were carrying. Ten Lygus and T. vaporariorum and 25 M. persicae per sample were surface sterilized following Shipp et al. (2003) to estimate internal infection levels. The samples were then placed on water agar plates, incubated at 25 o C and 80%RH for 5 days before checking for sporulation using a dissecting microscope. Another 10 Lygus and T. vaporariorum, 25 M. persicae and 5 bees per treatment sample were washed to determine the amount of conidia that individual pests carried using the protocol of Al-mazra awi et al. (2006). The remaining Lygus and T. vaporariorum were used to assess the percentage mortality caused by Beauveria. Each insect was placed in an aerated Petri dish (9 cm Ө) and fed fresh organic lettuce leaves for 7 days. The T. vaporariorum were placed individually in an aerated sealed plastic vial on the growing point of a tomato plant for 7 days. The tomato growing points were replaced every second day. All bioassay cages were inspected daily for insect death. The appearance of white mycelium on cadavers indicated Beauveria infection. Aphid mortality could not be determined this way because of high handling mortality. Mortality of the bumble bees exposed to the Beauveria treatments was determined after the second sampling. Each bumble bee colony was transferred back into the commercial hive boxes and fed pollen patties mixed with 50% wt/wt sugar solution for 5 weeks. The percentage of bumble bee mortality was recorded weekly. Simultaneous vectoring of insect and plant pathogen control agents These trials were conducted on sweet pepper using a similar experimental design as described above. In these trials, there were 32 plants per cage, 25 worker bees per hive and only 125 Lygus per cage were introduced. Three treatments were evaluated: 6.24x10 10 conida of B. bassiana x10 8 conidia of C. rosea/g of inoculum + bees, heat inactiviated inoculum + bees and control treatment with no inoculum and no bees. The plants were inoculated with 48
61 grey mold spores, Botrytis cinerea, and infested with L. lineolaris 2 days before the inoculum dispensers were placed in front of the bee hives. The Lygus samples were processed to determine CFU per bug and to assess mortality levels due to infection with Beauveria. A subset of the flower and leaf samples were processed to determine the presence of Beauveria and Clonstachys, and the rest of the samples were used to determine the impact of C. rosea on disease suppression of grey mold. These samples were placed individually on moistened filter paper lined in Petri dishes and incubated at 25ºC and 80% RH for 5 days for the flowers and 12 days for the leaves. The samples were then assessed for percentage of disease incidence. Results and discussion Sweet pepper Beauveria concentration trials Ninety-four and 35% of the bee and Lygus samples from the three Beauveria treatments contained detectable concentrations of Beauveria conidia. The greatest numbers of CFU per sample were found at the 6.24x10 10 and 2.0x10 11 conidia/g inoculum concentrations (Table 1). Table 1. Mean (± SE) number of CFUs per insect for the different Beauveria treatments on greenhouse sweet pepper. Treatment CFU counts Bumble bee Lygus Aphid x 10 9 conidia/g 3.3 x10 5 ± 1.1x10 5 a 77.3 ± 29.3b 39.0 ± 9.1b 6.24 x conidia/g 1.7 x10 6 ± 9.0x10 5 a ± 25.3a ± 30.5a 2.0 x conidia/g 1.6 x10 6 ± 7.4x10 5 a ± 29.7a ± 41.6a Heat inactivated inoculum 0 ± 0.0b 0 ± 0.0c 0 ± 0.0c No inoculum 0 ± 0.0b 0 ± 0.0c 0 ± 0.0c Within a column, means followed by different letters are significantly different at P< Aphids were pooled in groups of five for CFU counts. The highest mortalities for Lygus occurred at the concentrations (Figure 1). Mycosis occurred on 70% of the dead Lygus exposed to the lowest concentration of Beauveria and on 85-90% of the dead Lygus at the other two concentrations. The mean (± SE) internal infection levels by Beauveria were 55 ± 5% for Lygus and 26.0 ± 3.8% for M. persicae. Tomato Beauveria concentration trials Seventy-six percent of the bee samples over the five trials contained detectable concentrations of Beauveria conidia. CFU counts on the bees and T. vaporariorum exhibited similar trends as observed for sweet pepper and were the highest at the 6.24x10 10 and 2.0x10 11 conidia/g of inoculum concentrations. Like Lygus, the highest mortalities for T. vaporariorum occurred at the concentrations (Figure 1). Mycosis occurred on 70% of the dead T. vaporariorum that were exposed to the lowest concentration of Beauveria and on 82% of the dead T. vaporariorum at the other two concentrations. The mean (± SE) internal infection levels by Beauveria were approximately twice as high for the adult T. vaporariorum (36.8 ± 3.4%) at compared to the 10 9 conidia/g of inoculum. 49
62 Mortality (%) L. lineolaris T. vaporariorum x10 9 conidia/g 6.24x10 10 conidia/g 2.0x10 11 conidia/g Treatment heat inactivated inoculum no inoculum Figure 1. Mean (± SE) percentage mortalities of adult L. lineolaris and T. vaporariorum over the two sampling dates. Bumble bee mortality levels Bee mortality levels at the different treatments exhibited similar trends for both the sweet pepper and tomato trials. Mean percent mortality ranged from % for all treatments except the highest concentration which had a mortality rate of 42-45%. % Incidence of grey mold T1: 6.24x10 10 conidia of B. bassiana x10 8 conidia of C. rosea/g of inoculum T2: Heat inactivated inoculum T3: No inoculum Flowers Plant samples Leaves % mortality or mycosis Treatments Lygus mortality Lygus mycosis T1 T2 T3 Figure 2. Mean percentage mortality/mycosis of L. lineolaris and incidence of grey mold on flower and leaves exposed to the simultaneous bee vectoring of Beauveria and Clonostachys. 50
63 Simultaneous bee vectored insect pest control and disease suppression Washings of the Lygus, flower and leaf samples showed that most samples contained CFU of either Beauveria or Beauveria and Clonostachys. Mean CFU of Beauveria were , 5.8x10 3 and 6.96x10 4 for Lygus, flowers and leavers, respectively. Mean CFU of Clonostachys were 2.1x10 3 (flowers) and 2.66x10 3 (leaves). Mortality and mycosis levels for the Lygus for the different treatments were similar to what was observed in the earlier sweet pepper trials (Figure 2). Disease suppression of grey mold for the flower and leaf samples ranged from 54-73% (Figure 2). In summary, 6.24x10 10 conidia/g of inoculum seems to be the optimal concentration for control of L. lineolaris, M. persicae and T. vaporariorum on greenhouse sweet pepper and tomato. This concentration also had minimal impact on bee mortality. In addition, greenhouse cage trials have demonstrated that bumble bees can simultaneously delivery more than one fungal control agent for insect pest control and disease suppression. Acknowledgements Study was funded in part with a grant from Improved Farming Systems and Practices Initiative, Agriculture and Agri-Food Canada. References Al-mazra awi, M.S., Shipp, L., Broadbent, B. & Kevan, P. 2006: Biological control of Lygus lineolaris (Hemiptera: Miridae) and Frankliniella occidentalis (Thysanoptera: Thripidae) by Bombus impatiens (Hymenoptera: Apidae) vectored Beauveria bassiana in greenhouse sweet pepper. Biological Control: (in press). Butt, T.M., Ibrahim, L., Ball, B.V. & Clark, S. J. 1994: Pathogenicity of the entomogenous fungi Metarhizium anisopliae and Beauveria bassiana against crucifer pests and the honey bee. Biocontrol Sci. Technol. 4: Kovach, J., Petzoldt, R. & Harman, G.E. 2000: Use of honey bees and bumble bees to disseminate Trichoderma harzianum to strawberries for Botrytis control. Biological Control 18: Maccagnani, B., Mocioni, M., Gullino, M.L. & Ladurner, E. 1999: Application of Trichderma harzianum by using Apis mellifera for the control of gray mold of strawberries: first results. IOBC/WPRS Bull. 22: Peng, G., Sutton, J.C. & Kevan, P.G. 1992: Effectiveness of honeybees for applying the biocontrol agent Gliocladium roseum to strawberry flowers to suppress Botrytis cinerea. Can. J. Plant Pathol. 14: Shipp, J.L., Zhang, Y., Hunt, D.W. & Ferguson, G. 2003: Influence of humidity in greenhouse microclimate on the efficacy of Beauveria bassiana (Balsamo) for control of greenhouse arthropod pests. Environ. Entomol. 32: Yu, H. & Sutton, J.C. 1997: Effectiveness of bumble bees and honey bees for delivering inoculum of Giocladium roseum to raspberry flowers to control Botrytis cinerea. Biological Control 10:
64 52
65 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Current situation of Bemisia tabaci in protected vegetables crops in the South of France. Yannie Trottin-Caudal 1, Catherine Chabrière 2, Christine Fournier 1, Jean-Michel Leyre 1, Leen Schoen 3 1 Centre Technique Interprofessionnel des Fruits et des Légumes, BP 32, Bellegarde, trottiny@ctifl.fr, fournier@ctifl.fr, leyre@ctifl.fr 2 Association Provençale de Recherche et Expérimentation Légumière, Route de Mollégès, St Rémy de Provence, chabriere@aprel.asso.fr. 3 Centre Expérimental des Fruits et Légumes du Roussillon, Torreilles, France, l.schoen@centrex66.com Abstract: In 2001, IPM protected crops represented about 1800 ha in France, mainly tomatoes and cucumbers. The whitefly Bemisia tabaci, a pest and virus vector, was detected in 2001 in the South of France on different protected crops (tomato, cucumber but also on melon, courgette, eggplant and pepper). Its presence forced us to take measures and to provide more information to growers. Studies have been rapidly carried out in experimental and commercial greenhouses to better understand this pest and develop adequate control strategies. First results seem to prove that only the combination of beneficials with preventive measures could be efficient. Key words: Bemisia tabaci, tomato, biological control, greenhouse, Integrated crop protection Introduction The whitefly Bemisia tabaci was recorded in 2001 in Pyrénées-Orientales and in Provence- Alpes-Côte-d Azur. It is present on many crops in the South of France (tomato, sweet pepper, melon, etc.) and it is, moreover, vector of serious viruses on solanaceous crops, such as Tomato yellow leaf curl virus (TYLCV), Tomato chlorosis virus (ToCV) and on cucurbitaceous crops Cucumber yellow stunting disorder virus (CYSDV). Its presence on vegetables, especially on protected tomato crops, mobilized technical and professional organizations. Experimental studies were carried out to have a better knowledge on this new pest and to develop adapted strategies of protection. Situation of Bemisia tabaci Low populations of B. tabaci were detected in 2002 on several farms in Languedoc- Roussillon region. In contrast, very high populations of B. tabaci were present in 2003 when temperatures were very high in summer and autumn. Populations decreased in 2004 and On the major production sites, the species Trialeurodes vaporariorum was mixed with B. tabaci, at least at the beginning of the crop. To face this situation, a group was set up in 2001, made up of growers, professional organizations, official research and extension services. Warnings were also published in the regional press. In Provence-Alpes-Côte d Azur, high populations of B. tabaci were present in 2003 on tomato crops but also on sweet pepper, courgette, aubergine, etc. In 2004 and 2005, population levels were drastically lower even if an increase in populations was observed in 53
66 autumn in some commercial greenhouses, especially on sweet pepper. To sensitize those involved in the vegetable industry and to issue technical information, a paper was published in 2004 on prevention of virus disease risks on tomato by the official Services in close cooperation with professional organizations and grower representatives. It was reviewed in Moreover, a survey of the viruses transmitted by B. tabaci was carried out on various farms throughout the area. In Aquitaine, an assessment of B. tabaci populations was set up in 2004 on vegetable crops, according to the same protocol as in Provence Alpes Côte d Azur. B. tabaci was only identified in one commercial greenhouse, in early July. In 2005, B. tabaci was not observed (Favé, personal communication). The B. tabaci biotype was identified by CIRAD, using two micro-satellite markers. From the different samples taken in greenhouses, the study revealed the presence of the Q biotype with some polymorphism. This work is ongoing (Peterschmitt, personal communication). Studies in experimental greenhouses A few experiments were carried out with entomopathogenic fungi and chemical products compatible with IPM. For example, first results with kalium-based products, described in another paper for this meeting, are promising (Schoen, 2006). In the meantime, other trials were conducted at the Ctifl centre of Balandran, for a better understanding of biology and damage of B. tabaci on tomato crops as well as to study various biological control strategies. Trials were carried out in 60 m² insect-proof experimental glasshouses equipped with double rooms at the entrance. Treatments were applied in a separate compartment. Tomato plants were grown in cocofibre growing bags on the basis of 12 plants per row (density of 1.5/m²). Biology and damage of B. tabaci (2004) Material and methods. The infestation of B. tabaci was homogeneous on the plants: 40 or 110 per compartment (on 72 plants); two tomato varieties: Emotion (Syngenta Seeds) and Palmiro (De Ruiter). Observations were carried out on a weekly basis starting from the first B. tabaci infestation, on a random sample of 12 plants among the 48 plants in the 4 central rows. On each plant, adults and L4-pupae were counted or estimated on all the leaves. At harvest, fruits with symptoms of Tomato Irregular Ripening (TIR) were counted (100 to 300 fruits per harvest of the 48 central plants per compartment). Results. Average daily temperatures varied from C in April to 23 C in August. Emergence of B. tabaci adults took place after 4 or 5 weeks from the first infestation. This was confirmed in 2005 with a 34-day period between the first eggs and the adults at 20.7 C. The first symptoms of TIR were observed on both varieties 3 months after the first infestation of B. tabaci. Yellow star-shaped spots were observed at the apical part of pink or red fruits along with longitudinal marks, either yellow or orange, on the red fruit. Direct damage was also observed: when high populations of B. tabaci were present, with white punctures on small green fruits becoming yellow after ripening, and honeydew and sooty mold. In the compartment where more than 500 individuals (adult and L4-pupae) per plant were counted, more than 90% of the harvested fruit had TIR symptoms. On the other hand, in the compartment with a population of about 400 individuals of B. tabaci per plant, where the predator Macrolophus caliginosus (syn. M. melanotoma) fed on young larval stages, it appears that percentage of TIR fruit did not exceed 35 %. Percentage of fruit with TIR symptoms was less than 5% at the last harvest only in the compartment in which whitefly populations were under an average of 30 individuals per plant. It thus appeared that about 30 to 60 adults and L4-pupae per plant can cause TIR symptoms 4 to 5 weeks later. A relation between the density of larvae and pupae of B. tabaci and TIR symptoms was already 54
67 described by Shuster (2001), with a duration of 1 to 3 weeks between the level of B. tabaci populations and the appearance of symptoms under the experimental conditions of this study. Behavior of B. tabaci and T. vaporariorum on the plant (2004) Material and methods. Two specific counts were carried out to better define the distribution of the two whitefly species present on tomato plants. Adults and L4-pupae were counted on each leaf (about 25 leaves per plant) on a random sample of 12 plants per compartment. Results. On average 104 T. vaporariorum individuals versus 109 B. tabaci were observed per plant. B. tabaci adults were present on all the leaves but were more concentrated on the top of the plants and less on the apex when compared to T. vaporariorum. When the plants were divided into three levels, high, medium and low, 60%, 26% and 14% of B. tabaci adults and 87%, 11% and 2% of T. vaporariorum adults were observed respectively. For the L4-pupae stages, B. tabaci was mainly present on the bottom half part of the plants whereas T. vaporariorum was distributed on a more significant number of leaves, on the medium and superior levels of the plants. Biological control strategies against B. tabaci (2004 and 2005) Material and methods. The following treatments were studied: Control (without releases of beneficials), included in each trial; Eretmocerus mundus at 3/m² and 6/m² (in 2004), 1.5/m² and 3/m² (in 2005); E. eremicus at 1.5/m² and 3/m² (in 2005); Encarsia formosa at 3/m² reared on T. vaporariorum (in 2005, 2 nd trial); E mundus (1.5/m²) + E formosa (1.5/m²) in 2005, 2 nd trial; M. caliginosus 2x1.5/m² (in 2004). M. caliginosus 2x1.5/m² + E mundus at 3/m² (in 2004). Two experiments were carried out each year. Tomato plants cv. Palmiro, were grown in coco bags on the basis of 12 plants per row (density of 1.5/m²). They were planted on 11/03 in 2004 (1 st and 2 nd trial on the same plants) and in 2005, on 17/03 (1st trial) and on 26/07 (2 nd trial). Treatments were carried out in a random fashion. Plants were artificially infested with B. tabaci females along with a few males: in 2004, 40 per compartment for each trial and in 2005, 60 for each trial. Infestations were carried out on 01/04, 6/04, 16/04 and 21/04 (1 st trial) and 31/05, 6/06 (2 nd trial) in 2004 and on 30/03, 7/04 14/04 (1 st trial) and 1/08, 8/08, 16/08 (2 nd trial) in First release of E. mundus (from Koppert) was done as soon as eggs and first larval instar were observed (on 17/04, trial 2004) or later in 2005, either on the 2 nd /3 rd larval stages (on 17/08, 2 nd trial) or on the late instar (on 4/05, 1 st trial). For the other parasitoids, releases were carried out at the two late instar stages. The 3 other releases were carried out at 7-day intervals. All parasitoids were introduced in bulk at late instar stages with 3 to 4 points of release per compartment. M. caliginosus was introduced on 15/04 and 20/04 as female and male adults after sexing under a binocular lens (sex-ratio 1:1). Observations were carried out on a weekly basis on a random sample of 12 plants among 48 plants in the 4 central rows, starting from the date of B. tabaci infestation. On each plant the number of B. tabaci adults (along with L4- pupae, but only in 2004) and M. caliginosus adults and larvae were assessed. Moreover, in 2005 the effectiveness of parasitoids was monitored by observation of pupae under a binocular lens on a sample of leaves: the number of larvae parasitized, dead (natural mortality and predation) or alive, were counted. Three samples were taken during the 2 nd trial at about 15-day intervals and only one sample at the end for the 1 st trial. The parasitoid adult emergence ratio as well as the sex-ratio were also monitored on all commercial batches, either in laboratory conditions (random sample of 125 pupae in a commercial bottle) or in the greenhouses (on 2x75 pupae) 15 days after delivery. Results. In 2004 the control compartment was heavily infested for 3 months and the monitoring of B. tabaci populations was quickly stopped. Five chemical treatments applied when populations were too high (an average of more than 2000 individuals per plant) and the use of yellow sticky traps was insufficient to manage the pest population. When E. mundus was used alone provided good control on early B. tabaci populations but it required several 55
68 introductions every week for adequate parasitism to be obtained. The dose effect of introduction (3 or 6/m²) could not be detected under our conditions. Probably, B. tabaci population could not increase because of predation exerted by E mundus on the first larval stages. When M. caliginosus was introduced alone seemed effective in controlling B. tabaci but too late; this is probably due to its slow biological development. Indeed, the predator settled after 10 weeks and then controlled B. tabaci population. A strategy based on the mixture of E mundus and M. caliginosus proved to be effective throughout the experiment: E mundus controlled early B. tabaci populations and the predator was efficient once well established. In 2005, in laboratory conditions, the mean percentage of parasitoid adult emergence was 79.7% and 89.4% for E. eremicus and 80.5% and 86 % for E. mundus respectively in the 1 st and 2 nd trial. These values were similar to those obtained in greenhouses. However, E. eremicus adult emergence began a little later than that of E. mundus. Mean sex-ratio was 61.1% and 63.5% of females for E. eremicus, 63.4% and 68.4% of females for E. mundus and 100% of females for E. formosa respectively in the 1 st and 2 nd trial. These results are similar to those referred in several references. In the control treatment (2 nd trial), the mean number of adults per plant was significantly higher than that in the two treatments using E formosa from late August-early September (Kruskall Wallis Test, 5%). From 27/09 onwards this number doubled each week (Figure 1). Monitoring was stopped on 11/10, when the mean number of B. tabaci adults was 169 per plant. E. mundus or E. eremicus (reared on T. vaporariorum) at 1.5/m² or 3/m² were both effective parasitoids and B. tabaci populations were similar in both treatments. However, during the parasitoid establishment, the first whitefly generations were less efficiently controlled at the rate of 1.5/m² than in the other treatments. Indeed, on 13/09 and 20/09, the number of B. tabaci adults did not differ significantly in the two treatments (introduction of 1.5 parasitoids/m²) from that observed in the control treatment. Mean number of B. tabaci per plant (control) Control E. mundus 3/m² E. mundus 1,5/m² E. eremicus 1,5/m² E. eremicus 3/m² E. mundus 1,5/m² et E. formosa 1,5/m² E. formosa 3/m² /08 23/08 07/09 20/09 04/10 20/ Mean number of B. tabaci per plant Figure 1. Mean number of B. tabaci adults per plant in 7 treatments (2 nd trial). Random samples of 12 plants with observations on all the leaves (about 20 to 25 leaves per plant) were taken. On the other hand, the whitefly population could not establish on the plants in the two strategies using E formosa, and less than 2 adults per plant were present in late October. Population was always significantly lower than that in the other treatments (Kruskall Wallis test, 5%). This is partly due to the higher rate of introduced parasitoids because this species is 56
69 only composed of females. Concerning the effectiveness of parasitoids, two groups emerged; the first group with E. mundus and E. formosa that parasitized about 70% of the pupae present, and the second group with E. eremicus that parasitized 50% of the pupae (Figure 2). Moreover, some mortality was observed in pupae in all the treatments using parasitoids, almost reaching a percentage of 40% in the E. mundus + E. formosa treatment. This mortality was mainly due to predation exerted by parasitoids because natural mortality in the control was very low (not more than 5%). 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 100 Control Puparium parasitized by E. formosa Puparium parasitized by E. eremicus or E. mundus Puparium non parasitized E. mundus 1,5/m² E. mundus 3/m² E. eremicus 1,5/m² E. eremicus 3/m² E. mundus 1,5/m² + E. formosa 1,5/m² E. formosa 3/m² 5%(599) 22%(1056) 18%(737) 20%(1414) 19%(916) 36%(523) 32%(340) Figure 5. Percentage of parasitism by E. mundus, E. eremicus and E. formosa on B. tabaci pupae in the 7 treatments. Observations were made at the end of the test (2 nd trial), from 13/10 to 21/10 (according to the treatments). Percentage mortality is indicated for each treatment; it is calculated on all the pupae (alive, parasitized and dead) and this number is indicated in brackets. Observations in commercial greenhouses B. tabaci populations in Provence have been monitored since It appeared that a combination of control strategies with sanitation measures, releases of beneficials and, if necessary, a supervised chemical control, could effectively manage the two whitefly species. The two strategies with parasitoids (E. formosa, E. eremicus, E. mundus), either released alone or in combination with M. caliginosus could give effective control against whiteflies. On the other hand, it is important to note that growers often introduce parasitoids very early with regular releases in order to have good results. Furthermore, releases of M. caliginosus are often carried out later and in lower numbers even if this predator controls both whitefly species. The bug Nesidiocoris tenuis, regularly observed in tomatoes in Provence-Alpes-Côte d Azur (Chabrière and Trottin-Caudal 2001), performs well as whitefly predator but it very often causes damage to the plants and it is mainly considered a pest which can disrupt integrated protection (Trottin-Caudal et al., 2006). Trials were carried out either in insect-proof plastic tunnels (Diatex 0.80 mm 0.20 mm mesh at doors and openings) or in glasshouses. Nets showed their effectiveness on controlling 57
70 pest populations but it is necessary to install them before planting. It is also very important to combine this technique with preventive measures such as weed control and double doors at the entry of the greenhouse, even if this last measure is not always easy to apply, especially under plastic tunnels. A quicker development of whitefly populations has also been observed in greenhouses equipped with nets, probably caused by an increase in temperature. Under plastic greenhouses, a greater risk of Botrytis cinerea was also noted. Conclusions In the South of France, the presence of B. tabaci as a pest and virus vector has led to a more complex situation in protected crops. T. vaporariorum and B. tabaci are both present on the plants, so it is necessary to monitor those two species in order to adapt the biological control strategy. Following the various studies of biological protection and observations carried out since 2002, interesting results have been obtained on the biology and the harmfulness of B. tabaci, as well as the effectiveness of commercially-available beneficials or release strategies. Under our experimental conditions, the three parasitoids E. mundus, E. eremicus and E. formosa even reared on T. vaporariorum, provided good control against B. tabaci. These results remain to be confirmed in commercial greenhouses. References Chabrière, C. & Trottin-Caudal, Y. 2001: Tomate sous serre. Deux ravageurs en extension. Réussir Fruits et Légumes, 196, Schoen, L. 2006: Evaluation of two potassium based products on Bemisia tabaci (Gennadius) in greenhouse tomatoes. In this volumen of the IOBC/WPRS Bull. Shuster, D.J. 2001: Relationship of Silverleaf Whitefly population density to severity of Irregular Ripening of Tomato. Hortscience 36 (6), Trottin-Caudal, Y., Chabrière, C., Fournier, C. & Leyre, JM. 2006: Etude expérimentale sur la punaise Nedisiocoris tenuis Reuter, ravageur de la tomate sous serre dans le Sud-est de la France : nuisibilité, action prédatrice et efficacite de stratégies de limitation des populations. In : 3 ème Conférence Internationale sur les Moyens Alternatifs de Protection 58
71 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Natural parasitism of Bemisia tabaci and Trialeurodes vaporariorum in a horticultural area of Sardinia, Italy Mauro Nannini 1, Lorena Manca 1, Massimo Giorgini 2 1 Centro Regionale Agrario Sperimentale, V.le Trieste 111, Cagliari, laiu@cras.sardegna.it 2 Istituto per la Protezione delle Piante Sezione di Portici, C.N.R., Via Università 133, Portici (NA), Italy, giorgini@ipp.cnr.it Abstract: A three-year survey was carried out in southern Sardinia (Pula, Cagliari) to assess natural whitefly parasitism on greenhouse tomatoes and to evaluate the contribution of parasitic wasps as biocontrol agents for Bemisia tabaci on other crops and weeds. The rates of apparent parasitism on protected tomato crops were low, reaching 18.8% in the most favourable of cases. The role played by parasitoids for the control of B. tabaci varied greatly among cultivated and weed species. Seven species of Aphelinid parasitic wasps were found to be associated with whiteflies. On this basis, the agroecosystem examined could be regarded as quite rich in terms of parasitoid species. This positive precondition did not, however, make a decisive contribution to whitefly management. Key words: Aphelinidae, biological control, greenhouse tomato, Hymenoptera, parasitic wasps, protected crops, vegetables, weeds, whiteflies. Introduction In the 1990 s, the Regional Administration for the implementation of IPM practices in greenhouse tomato production made great efforts to optimise Sardinian horticultural production (Nannini & Ortu, 2003). In this context, a combination of insecticide treatments, exclusion nets and inoculative releases of predators and parasitoids succeeded in controlling whitefly and the whitefly-transmitted viral disease tomato yellow leaf curl. However, in the last few years an intensification of TYLCD has been observed in southern Sardinia, causing great concern to tomato growers and renewing the demand for effective and environmentally safe means to solve whitefly-related problems. Before introducing any biological control programme for whitefly, a study was undertaken in order to evaluate the contribution of naturally-occurring parasitic wasps to whitefly control on greenhouse tomato. Here we report the results of a three-year survey on parasitism on Bemisia tabaci (Gennadius) and Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae) in one of the major horticultural areas of the island (Pula, Cagliari). We also investigated the role of the parasitoid complex in limiting the development of B. tabaci populations on several other cultivated plants and also on weeds. Materials and methods Thirty tomato-growing greenhouses were monitored from 2003 to Long-cycle (from August-October to June) and short-cycle (autumn cultivations, from August to December- January; spring cultivations, from December-February to June-July) crops were sampled. On 18 greenhouse farms, whitefly management measures included the release of beneficials species; Macrolophus caliginosus, Encarsia formosa and Eretmocerus mundus, at 17, 2 and 4 sites, respectively. In most cases (25 of 30) 40 mesh anti-insect nets were used for screening greenhouse vents and windows. Each crop was sampled once or twice, between October and January (autumn sampling period) or between April and July (spring sampling period). Each 59
72 sample consisted of tomato leaves infested by immature whitefly instars, each of which was collected from a different plant. Natural parasitism of B. tabaci infesting crops other than tomato and weed species was investigated from August to October 2004 and from January to November In 2004 samples were collected from ten greenhouse farms, whereas in 2005 samples were taken from only a single site. Weeds were mainly recovered from uncultivated surroundings of tomato fields and in some cases from inside cultivated greenhouses. Each sample consisted of 10 plants per species and at least three leaves per plant were randomly collected. On each leaf, we counted the total number of apparently parasitized (no dissection was carried out) and non-parasitized whitefly 4 th nymphs and pupal cases. Both live and dead (due to any cause other than parasitization) non-parasitized whiteflies were counted, as were empty pupal cases for whitefly or wasp adult emergence. For each sample, we calculated the mean density of total immature whitefly per leaf (greenhouse tomato) or sample plant (hosts other than tomato) and the rate of apparent parasitism. Apparent parasitism was expressed as a ratio of parasitized to total number of 4 th nymphs and pupal cases. Parasitized pupal cases were isolated in ventilated Petri dishes in order to identify the species of adult wasps emerging. Results and discussion On greenhouse tomato, natural whitefly parasitism (pooling data from T. vaporariorum and B. tabaci) was lower in autumn-winter than in spring-summer (Tables 1 and 2), reaching maximum values of 3.1% and 18.8% respectively, at sites where no parasitoids were released. In general, parasitism was scarce (on average 2.1% in fall-winter and 4.0% in springsummer), even in greenhouses where inoculative releases of parasitoids were carried out. Five species of parasitic wasps (Hymenoptera: Aphelinidae) emerged from whitefly pupae: Encarsia lutea (Masi), Encarsia mineoi Viggiani, Encarsia pergandiella Howard, Encarsia tricolor Förster and Eretmocerus mundus Mercet (Table 3). From autumn samples, in which the most abundant whitefly species was B. tabaci (averaging 85.5% of observed whitefly), 21 parasitic wasps were collected from tomato crops where E. mundus was not inoculated. E. tricolor and E. mundus, with an incidence of 47.6% and 33.3% respectively, were the most prevalent indigenous species. In greenhouses in which E. mundus was released, this species represented 94.6% of the 92 parasitic wasps collected. From spring samples, mainly infested by T. vaporariorum (88.2%), E. pergandiella was the dominant species, representing 73.4% of 128 parasitic wasps collected. Encarsia formosa was not recovered in the two greenhouses in which it was inoculated. The parasitoid complex associated with whiteflies infesting greenhouse tomato was quite rich, including species such as E. pergandiella and E. mundus, which have been found to play an important role in whitefly control in the Mediterranean region (Viggiani, 2000; Arnó et al., 2005). E. mineoi was recorded for the first time in Italy; it was found to be associated only with B. tabaci. On open field horticultural crops (eggplant, pepper, sweet pepper, melon and watermelon) apparent parasitism of B. tabaci pupae ranged from 0.0% to 40.0% (Table 4), with very low values on heavily infested samples. Higher levels of natural parasitism were achieved on greenhouse cultures, including melon (12.8%) and ornamental crops like lemon verbena (Lippia triphylla) (20.0%) and poinsettia (Euphorbia pulcherrima) (28.5%). 60
73 Table 1. Apparent parasitism rates observed on whiteflies infesting greenhouse tomato in autumn-winter; n = 25 leaves/sample. 2003/ / mesh nets natural enemies released planting month rates of parasitism (%) mean number of 4 th instar whitefly nymphs and pupal cases/leaf Oct Dec Jan Oct Dec Jan X M Aug X M Aug X Aug Aug X Aug X Aug X Aug X M,E Aug *0.0 *6.5 *3.8 *8.0 X M,E Aug *13.8 *5.8 *20.0 *42.0 X Aug X M,E Aug *8.2 *1.8 *11.8 *24.1 X M,E Aug *0.0 *0.6 *5.0 *18.6 Legend: each line represents a greenhouse at a different site; * values refer to greenhouses in which parasitoids were released; M: M. caliginosus; E: E. mundus. Table 2. Apparent parasitism rates observed on whiteflies infesting greenhouse tomato in spring-summer; n = leaves/sample. 2003/ / mesh nets natural enemies released planting month rates of parasitism (%) 61 mean number of 4 th instar whitefly nymphs and pupal cases/leaf Apr May Jun Jul Apr May Jun Jul X M Aug X Aug X M Sep X M Sep X M,F Jan *0.0 *4.2 *2.8 *4.7 X M Jan X M Jan F Jan *0.4 *13.1 *10.6 *130.8 Aug Sep X M Sep X Sep X M Oct Oct X M Dec X M Feb X Feb X M Feb Legend: see Table 1
74 Table 3. Parasitoid complex associated with whiteflies in different crops. (Incidence is given as percentage) host plants E. hispida E. inaron greenhouse tomato* greenhouse tomato* E open field vegetables other protected crops weeds Legend: *both B. tabaci and T. vaporariorum were considered; E: values refer to greenhouses in which E. mundus was released. E. lutea E. mineoi E. pergandiella E. tricolor E. mundus Table 4. Apparent parasitism rates observed on Bemisia tabaci infesting open field and protected crops and mean number of 4 th instar whitefly nymphs and pupal cases/plant (between brackets); n = 10 plants/sample. host species Aug 04 Sep-Oct 04 Jun 05 Jul-Aug 05 Capsicum annum of 9.8 (4.1) 0.9 (31.7) Capsicum chinense of 12.5 (0.8) 7.5 (4.0) 40.0 (0.5) Citrullus lanatus of 0.9 (314.2) Cucumis melo of 0.1 (496.2) Solanum melongena of 0.1 (280.9) 3.9 (178.8) 0.0 (4.4) 2.9 (255.8) Cucumis melo gh 12.8 (27.3) Euphorbia pulcherrima gh 1.9 (5.3) 28.5 (18.6) Lippia triphylla gh 20.0 (35.5) Legend: of = open field; gh = greenhouse A quite different parasitoid complex composition was found to characterize open field crops and protected horticultural and ornamental crops (Table 3). Eretmocerus mundus was the prevalent parasitic wasp on field crops (63.9%), whereas no dominant species was observed on greenhouse melon and ornamental crops. On weeds, rates of apparent parasitism ranged from 0.0% to 39.4% (Table 5). The highest values were found on Amaranthus spp. (39.4%), tomato grown as a weed (33.9%) and Solanum nigrum (33.3%), on plants with low densities of whiteflies. On samples infested by high numbers of whiteflies the rate of parasitism was very low: % on samples with more than 100 pupal cases/plant. The relative abundance of parasitoid species on weeds is reported in Table 3. The results of the present study seem to indicate that, despite the high richness of Aphelinid species in the area examined, indigenous parasitic wasps played a secondary role in controlling the whitefly that infested greenhouse tomato crops. The observed rates of 62
75 parasitism were considerably lower than those reported for greenhouse tomato in other Mediterranean horticultural areas, such as southern Italy (20-49%) (Giorgini & Viggiani, 1994; Viggiani, 2000) and northeast Spain (30-37%) (Gabarra et al.,1999). Table 5. Apparent parasitism rates observed on Bemisia tabaci infesting weeds and mean number of 4 th instar whitefly nymphs and pupal cases/plant (in brackets); n=10 plants/sample host species Aug Sep Jan Mar Jun Jul Oct Feb May Aug Amaranthus spp (10.9) (2.5) Aster squamatus (6.6) (4.6) (15.6) Convolvulus arvensis Conyza spp. (6.9) 8.1 (10.4) Conyza spp. gh (77.9) (115.5) (9.2) (1.7) (1.1) (211.4) 0.0 (0.4) Datura stramonium (34.7) (3.9) Dittrichia 2.3 graveolans (4.3) Euphorbia humifusa (113.3) (64.5) Geranium spp (0.2) Heliotropium europaeum (3.2) (6.3) (3.4) Lycopersicon esculentum (54.2) (0.7) Malva sylvestris (33.2) (26.0) (0.5) Oxalis spp gh (0.3) Picris echioides (14.1) (8.7) Solanum nigrum (26.5) (63.3) (6.8) (0.9) (2.7) (272.0) Solanum 18.6 nigrum gh (22.0) Solanum 6.2 rantonetii (6.5) Sonchus oleraceus (13.7) (196.4) Sonchus spp (19.3) (108.3) (0.9) (245.2) Legend: gh = samples collected from greenhouses Sep 0.0 (0.6) 8.7 (123.3) 2.7 (25.6) 3.9 (23.0) Oct Nov 25.0 (9.6) 0.0 (0.1) 0.0 (0.5) 0.0 (2.7) This might depend on differences in agronomic and environmental conditions between the different zones. In Sardinia, it is likely that normal cultivation practices (use of persistent wide-spectrum insecticides, intense plant de-leafing) and climatic conditions inside 63
76 greenhouses are unfavourable to parasitoid colonization for a large part of the cropping season. The same factors could also reduce the efficacy of inoculative releases of E. formosa and E. mundus. Different levels of natural parasitism and differences in the composition of the parasitoid complex associated with B. tabaci were recorded on host plants different from greenhouse tomato. Parasitism on open field horticultural crops was scarce. This was presumably due to insecticide treatments, which might also have influenced the relative abundance of natural enemies, reducing early colonization of crops by parasitic wasps active since spring (i.e. E. pergandiella) in favour of species that were more abundant in summer (i.e. E. mundus). However, low rates of parasitism also occurred on weeds, which in general were sampled in areas that were not directly influenced by agricultural practices. This suggests that the activity of parasitic wasps could be limited by other factors, such as climate and intraguild predation. This latter hypothesis could be supported by the results of complementary observations carried out on Conyza spp., Sonchus spp. and S. nigrum, where a consistent number of Nesidiocoris tenuis and other predators were detected and where overall mortality rates for immature whitefly were approximately 6 to 7 times higher than apparent parasitism (data not shown). Mortality of Aphelinids caused by predators differentially related to specific host plants together with parasitoid preferences for different plant species could explain the notable differences in parasitism rates recorded among different species of wild weeds. The rich species composition of the parasitoid complex associated with B. tabaci in the horticultural area surveyed was presumably a consequence of limited land exploitation for agricultural purposes and the presence of extensive areas covered by natural vegetation. Even so, this positive precondition did not make a decisive contribution to whitefly management. Even if the application of measures for the conservation of natural enemies could enhance biocontrol on greenhouse tomato, at the moment, in consideration of the large diffusion in Sardinia of TYLCD, the adoption of such pest management tactic, without integration of other means of crop protection, does not seem adequate to avoid economic damage related to whitefly infestations. However, to improve whitefly management, further research is necessary to investigate the reasons for the relatively limited contribution of parasitoids to whitefly control on natural vegetation, and the role of indigenous predators like Mirid bugs in the population dynamics of both whiteflies and wasps. References Arnó, J., Matas, M., Marti, M., Ariño, J., Roig, J. & Gabarra, R. 2005: Coexistence between Trialeurodes vaporariorum and Bemisia tabaci and impact of natural enemies in tomato crops under Mediterranean conditions. IOBC/wprs Bull. 28(1): 1-4. Gabarra, R., Arnó, J., Alomar, O., & Albajes, R. 1999: Natural occurring populations of Encarsia pergandiella (Hymenotpera: Aphelinidae) in tomato grenhouses. IOBC/wprs Bull. 22(1): Giorgini, M. & Viggiani, G. 1994: Risultati di lotta integrata al Trialeurodes vaporariorum (Westwood) (Homoptera: Aleyrodidae) su pomodoro da mensa in coltura protetta (secondo raccolto). Informatore Fitopatologico 7-8: Nannini, M. & Ortu, P. 2003: Implementation of IPM strategies for the control of greenhouse whiteflies in protected tomato crops: the Sardinian experienc e. Actes du Colloque International "Tomate sous abri - Protection intégrée - Agriculture biologique", Avignon, France, settembre 2003: Viggiani, G. 2000: Note biologiche e distribuzione in Campania di Encarsia pergandiella, parassitoide di Bemisia tabaci e Trialeurodes vaporariorum. Informatore Fitopatologico 12:
77 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Alternative whitefly biocontrol options for greenhouse tomatoes Mauro Nannini, Francesco Foddi, Giovanni Murgia, Riccardo Pisci, Francesco Sanna, Salvatore Spanedda Centro Regionale Agrario Sperimentale, Viale Trieste 111, Cagliari, Italy, Abstract: An experiment has been carried out in Southern Sardinia (Pula, Cagliari) to evaluate the effectiveness of alternative options for whitefly biocontrol in greenhouse tomatoes. Four release programmes were carried out on different spring crops: (1) 2.0 Macrolophus caliginosus/m 2, (2) 1.6 M. caliginosus/m 2 provided with Ephestia kuehniella eggs as alternative food source, (3) 1.0 M. caliginosus, 8.0 Encarsia formosa and 8.0 Eretmocerus mundus/m 2, and (4) 16.0 E. formosa and 16.0 E. mundus / m 2. The establishment of natural enemies was poor (mirid bugs) or failed completely (parasitic wasps), presumably owing to unfavourable climatic conditions, low prey/host availability and severe plant de-leafing. Consequently, the beneficials released to control whitefly did not produce satisfactory results. Differences in predatory bug abundance were observed in greenhouses with different leaf removal regimens. The results suggest that further work is necessary to adapt whitefly biocontrol tactics to the cultural practices and climatic conditions existing in tomato greenhouses in Sardinia. Key words: Encarsia formosa, Eretmocerus mundus, Macrolophus caliginosus, natural enemies, protected crops Introduction Funded by the Sardinian Regional Authorities, during the 1990s the use of natural enemies for managing tomato pests became widespread practice in the greenhouse districts of the island, reaching peaks of approximately 10 million Encarsia formosa and 1.1 million Macrolophus caliginosus per season. In recent years the situation has changed significantly and releases of biocontrol agents (BCAs) have been reduced substantially. The reasons for this are both economic and technical (Nannini, 2003). Nevertheless, the most competitive tomato growers still argue that the use of natural enemies is an attractive option for protecting their most valuable greenhouse crops. Over the last 15 years the number of commercially available BCAs has increased considerably (Weintraub & Cheek, 2005), potentially enhancing pest management but also making growers decisions more difficult. Even limiting the choice to the more widely used species for whitefly control (i.e. M. caliginosus, E. formosa and Eretmocerus mundus), many options have to be considered regarding release rates, combinations of different species, modes of introduction, etc. In 2004 an experiment was conducted in Southern Sardinia for a preliminary evaluation of four alternative solutions for the biological control of whiteflies in tomato greenhouses. The cost of each option was fixed at approximately 0.26 euros / m 2. Materials and methods Four commercial spring tomato crops (one per treatment), cultivated from week 4-5 to week in the horticultural district of Pula (Cagliari), were selected for hosting the experiment. 65
78 Tomatoes (cvs. Cupido and Core Boi) were grown in glasshouses of 2,000 to 3,300 m 2 following standard cultural practices, which included 3 insecticide treatments for whitefly control, applied within the first two weeks after transplanting, and one treatment, or the release of Diglyphus isaea per m 2, for leafminer control. The following options were evaluated: greenhouse 1, 2.0 M. caliginosus/m 2 ; greenhouse 2: 1.65 M. caliginosus/m 2 provided with Ephestia kuehniella eggs as alternative food source; greenhouse 3: 1.0 M. caliginosus, 8.0 E. formosa and 8.0 E. mundus/m 2 ; greenhouse 4: 16.0 E. formosa and 16.0 E. mundus/m 2. E. kuehniella eggs and parasitic wasps were supplied by Koppert Biological Systems, The Netherlands; M. caliginosus by Biobest Biological Systems, Belgium. The mirid bugs were introduced on weeks 11 and 12, in clusters of approximately 200, 165 and 100 predators per release point, respectively in greenhouses 1, 2 and 3. One hundred introduction spots per hectare were set up, each consisting of a 3- (in greenhouse 1 and 3) or 4-stem plant (in greenhouse 2). E. kuehniella eggs were dispensed on points of release at a rate of 67 g / ha, four times starting from week 11, at 7-day intervals. Parasitoids were introduced three times on weeks 12, 14 and 16. Two different release patterns were followed: in greenhouse 3 E. formosa was introduced earlier than E. mundus (E. formosa: approximately 70% on week 12 and 30% on week 14; E. mundus: 30% on week 14 and 70% on week 16), whereas in greenhouse 4 equal amounts of the two species were released at the same time (15, 32 and 53% of the total amount on weeks 12, 14 and 16 respectively). Whitefly and mirid populations were monitored by means of fortnightly counts of the number of individuals present on the terminals of 50 sample plants per site (adult whiteflies on the top 3 leaves; nymph and adult mirids on the top 7 leaves). Whitefly species composition and mortality of immature stages were estimated in May and July by counting the number of live, preyed and parasitized whitefly on samples of 50 infested leaves randomly selected among those where pupae had started to hatch. For this purpose, only fourth instar nymphs, pupae and empty pupal cases were detected and examined under a stereomicroscope. Parasitized pupae were isolated in ventilated Petri dishes for identification of adult parasitic wasps. In order to study the factors influencing the establishment of M. caliginosus, 6 release areas per greenhouse were monitored. Each area consisted of 13 crosswise-arranged plants, the central plant coinciding with a release point. The number of whiteflies (4 th instar nymphs, pupae and adults) and M. caliginosus (nymphs and adults) present on one stem per plant was counted on weeks 12, 14, 17, 20, 23 and 26. The predatory bug abundance on weeks 12 and 20 was compared in greenhouses 1 and 3 by one-way ANOVA, after log-transformation of data, and means were separated by Tukey s HSD procedure. In the same sites, the number of leaves (more than 2 cm long) present and removed from sample plants was also recorded, on weeks 12, 14, 17 and 20, so as to evaluate the effects of plant deleafing on mirid bug population growth. For each sampling interval the ratio of the average number of leaves removed between week n and week n+1 to the average number of leaves present on week n was calculated. Results and discussion Throughout the entire cropping period, infestation levels of whitefly were remarkably low in all experimental sites (Figure 1). T. vaporariorum was found to be the predominant whitefly species (Table 2). On account of whitefly species composition and abundance, TYLCD incidence was also very low (Table 2). The efficacy of the pest control measures adopted combined with the unseasonably cold weather in late spring 2004, were likely responsible for the marked delay observed in the development of whitefly infestation. 66
79 mean number of adult whiteflies / plant whiteflies M. caliginosus greenhouse 1 greenhouse 2 release of M. caliginosus release of parasitic wasps greenhouse 3 greenhouse 4 1 0,9 0,8 0,7 0,6 0,5 0,4 0,3 0,2 0,1 mean number of nymph and adult Mirids / plant week number Figure 1. Mean number of whiteflies (adults) and M. caliginosus (nymphs and adults) per plant on samples of 50 plants per site. 0 Table 2. TYLCD incidence (in percentage) recorded 1, 2 and 4 months after transplanting; whitefly species composition and mortality of immature stages. TYLCD incidence (%) whitefly species composition and immature stages mortality rates 1 month 2 months 4 months week % B. tabaci % T. vaporariorum % of parasitized whiteflies % of preyed whiteflies greenhouse greenhouse greenhouse greenhouse
80 Mirid bug density was also negligible (Figure 1). Nevertheless in greenhouse 3 M. caliginosus abundance achieved maximum values approximately 10-fold higher, on average, than in greenhouses 1 and 2. The parasitic wasps released between March and April failed to colonize the crops. The only evidence of parasitoid establishment was detected in greenhouse 4, where 21 T. vaporariorum pupae parasitized by E. formosa were found on week 21. Because of the unsatisfactory growth of BCA populations, mortality rates were low, in all cases less than 17% (Table 2). The highest level of parasitism was achieved in greenhouse 4 (16.5%), where large numbers of the naturally occurring Aphelinid Encarsia pergandiella were observed. Apart from this species, which showed an incidence of approximately 95% of the total amount of adult parasitoids emerging from T. vaporariorum pupae (n = 184), the parasitoid complex included Encarsia tricolor (5% of incidence). The survey of the release point surroundings revealed differences in mirid bug abundance in the greenhouses examined (Figure 2). The highest numbers of M. caliginosus per plant were observed in greenhouse 3: as this site showed the lowest levels of whitefly infestation, and no alternative food source was supplied to predators, it seems plausible that the differences observed in Mirid population growth did not depend on food availability. mean number of whiteflies and Mirids (immature stages and adults) / plant 3 2,5 2 1,5 1 0,5 0 whiteflies M. caliginosus release of M. caliginosus greenhouse 1 greenhouse 2 greenhouse week number Figure 2. Mean number of whiteflies (4 th instar nymphs, pupae and adults) and M. caliginosus (nymphs and adults) per release area. Concerning plant deleafing, observations revealed differences in how this was practised in greenhouses 1 and 3 (Table 3). No leaves were removed for one week after the release of M. caliginosus on week 11. During the intervals week 12 - week 14, plant defoliation in greenhouse 1 was more severe than in greenhouse 3; the inverse was observed between week 14 and week 17. Furthermore, the ratio of the number of leaves removed between week 12 and week 20 to the number of leaves present on week 12 was similar in greenhouses 1 and 3. 68
81 Table 3. Deleafing severity expressed in terms of mean percentage of leaves removed between week n and week n+1 with respect to leaves present on week n; M. caliginosus abundance recorded in greenhouses 1 and 3 before and after deleafing, in 6 release areas per site (13 plants per area). percentage of leaves removed mean number of M. caliginosus per release area (± SE) weeks weeks weeks weeks weeks week 12 week 20 greenhouse ± 0.4 a 3.7 ± 0.2 a greenhouse ± 0.1 a 10.6 ± 0.3 b Means within columns followed by a common letter do not differ significantly according to Tukey s HSD test (p = 0.05). Predatory bug abundance in greenhouses 1 and 3, before and after the application of different deleafing regimens, are reported in Table 3. Whereas no difference was observed on week 12 in the average number of Mirids per release area (F=1.4; df=1, 10; P>0.05), on week 20 the number of predatory bugs was significantly larger in greenhouse 3 (F=8.9; df=1, 10; P<0.05). Possibly the more severe leaf removal observed during the first 3 weeks after mirid introduction into greenhouses 1 disrupted the establishment of the predators, resulting in lower abundance. The results of the present experiment suggest that BCAs provided poor whitefly control in the greenhouses examined. Unfavourable weather conditions and low prey availability were presumably the major limiting factors to the establishment of M. caliginosus on tomato crops, as reported in previous works (Carboni et al., 2002; Nannini et al., 2004). Nevertheless feeding the predator with Ephestia eggs did not result, in the present test, in an increase in abundance. Early plant deleafing probably affected mirid bug establishment: the impact of this practice on M. caliginosus populations has already been observed (Bonato et al., 2004), evidencing the need for its rational application. Both E. formosa and E. mundus failed to establish on the crops. Considering the weather conditions and cultural practices in the experimental sites, as well as the thermal requirements and host preference of E. mundus, the parasitic wasp was probably introduced prematurely. E. formosa presumably stood more chances of successfully colonizing the crop, but the severe leaf removal practised in all sites, along with the presence of the hyperparasitoid E. pergandiella in greenhouse 3, may have disrupted the establishment of this parasitoid (Calabretta et al., 1989; Loomans et al., 2002). Finally, the present experiment suggests that further work is needed to adapt whitefly control tactics to the climatic and cultural conditions which occur in Sardinian tomato greenhouses. More specifically, the application of all cultural practices which may disrupt the establishment and the development of BCA populations should be critically reconsidered. For the time being, even if the implementation of biocontrol measures remains difficult in the horticultural areas where TYLCD is present, the reduction of insecticide treatments and the use of selective active ingredients could enhance biological control of whiteflies on spring crops, which are less exposed to viral infections. 69
82 Acknowledgements This work was funded by the Regione Autonoma della Sardegna within the project Difesa e valorizzazione delle produzioni ortive in coltura protetta attraverso l impiego di ausiliari. The authors would like to thank Dr. M. Giorgini (Istituto per la Protezione delle Piante del C.N.R. Portici, Italy) and M. Lorena (C.R.A.S.) for parasitoid identification, and the Cooperativa S. Margherita Terra e Sole for the collaboration. References Bonato, O., Ridray, G. & Jeannequin, B. 2004: Impact of Macrolophus caliginosus populations in protected tomato crops. Abstracts Compendium of the 2 nd European Whitefly Symposium, Cavtat, 5-9 October 2004: Calabretta, C., Calabrò, M. & Nucifora A. 1989: Lotta biologica con lanci di Encarsia formosa (Gahan) su pomodoro in colture protette nel ragusano. Informatore Fitopatologico 6: Carboni, M.A., Marchi, S. & Nannini, M. 2002: Fattori che influenzano l'efficacia dei lanci inoculativi di Macrolophus caliginosus Wagner (Heteroptera, Miridae) nelle serre di pomodoro della Sardegna meridionale. Atti Giornate Fitopatologiche (1): Loomans, A.J.M., Huang, Y., Bukovinszkiné-Kiss, G. & van Lenteren, J.C. 2002: Heteronomous hyperparasitoids for biological control of whiteflies: balancing benefits and risks. IOBC/WPRS Bull. 25 (1): Nannini, M. 2003: Experiments on the use of Macrolophus caliginosus Wagner in Sardinian tomato greenhouses. Actes du Colloque International "Tomate sous abri - Protection intégrée - Agriculture biologique", Avignon, France, Septembre 2003: Nannini, M., Carboni, M.A. & Marchi, S. 2004: Osservazioni sull insediamento in serre di pomodoro del Miride Macrolophus caliginosus Wagner introdotto con lanci inoculativi autunnali. XIX Congresso Nazionale Italiano di Entomologia, Catania, giugno 2002: Weintraub, P. & Cheek, S. 2005: Need for new biocontrol agents in greenhouse IPM a European perspective. IOBC/WPRS Bull. 28 (1):
83 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Releases of Eretmocerus mundus and Macrolophus caliginosus for controlling Bemisia tabaci on spring and autumn greenhouse tomato crops Rosa Gabarra, Rafael Zapata, Cristina Castañé, Jordi Riudavets, Judit Arnó Plant Protection Department, IRTA Centre de Cabrils, E Cabrils (Barcelona), Spain Abstract: Bemisia tabaci is an important tomato pest in Mediterranean greenhouses and open field crops because it can build up important populations and can vector some important viral diseases. Biological control is necessary to improve B. tabaci control since this whitefly has become resistant to many available insecticides. We studied the efficacy of using two natural enemies, Eretmocerus mundus and Macrolophus caliginosus, either individually or in combination to control this whitefly on greenhouse tomatoes in spring and autumn crop cycles. E. mundus was effective in reducing whitefly populations in both crop cycles. However, the greatest reduction in terms of both adults and nymphs of B. tabaci was achieved with the combined use of E. mundus and M. caliginosus, especially in spring and with high whitefly populations. Releases of both natural enemies prevented adult whitefly emergence and the establishment of predators permitted to control subsequent crop colonization by whitefly. Key words: Bemisia tabaci, Eretmocerus mundus, Macrolophus caliginosus, tomato, greenhouses Introduction Bemisia tabaci is an important tomato pest in Mediterranean greenhouses and open field crops because it can build up important populations and can vector some important viral diseases. The development of a biological control strategy for tomato crops is much needed, among other reasons because this whitefly has become resistant to many available insecticides. Moreover, tomato coexists with other neighbouring vegetable crops and uncontrolled B. tabaci populations may increase the risk of virus transmission and impede successful application of area-wide IPM programs (Gerling et al. 2001). Eretmocerus mundus is a very effective biological control agent of B. tabaci (Greenberg et al. 2002, Stansly et al. 2005a). This parasitoid is well adapted to the Mediterranean area and natural populations are abundant in insecticide-treated and untreated vegetable and ornamental crops (González-Zamora et al. 1996, Stansly et al. 2005a, Arnó et al. 2005). Macrolophus caliginosus is an effective whitefly predator and is widely used on greenhouse tomatoes. This omnivorous predator is also well adapted to Mediterranean conditions. It feeds on B. tabaci (Barnadas et al. 1998) and its use with the parasitoid Encarsia formosa improves greenhouse whitefly control on spring greenhouse tomato crops (Castañé et al. 2004). Nevertheless, no known trials have assessed the complementary effectiveness of M. caliginosus and E. mundus for B. tabaci control on greenhouse tomatoes produced in the Mediterranean basin. In this temperate area, tomatoes are produced all year round in non-heated greenhouses, mainly in two crop cycles; spring (from March to July) and autumn (from August to December). The objective of our study was to determine the efficacy of these two natural enemies of B. tabaci, both individually and in combination, to control this whitefly on greenhouse 71
84 tomatoes. Since climatic conditions may affect the establishment and efficacy of natural enemies we tested them in two different seasons: autumn and spring. Material and methods Two successive tomato crops were grown in an experimental greenhouse divided into 12 exclusion compartments (4 x 6 m) and covered with mesh (10 x 20 threads/cm 2 ) to prevent insect movement between compartments. Forty-eight tomato plants (cv. Bodar) were transplanted in each cage in August (autumn crop) of one year and in March (spring crop) of the following year in standard growing bags. Plants were supported with plastic twine and trained to a single stem by removing all lateral shoots. A standard fertirrigation plan for tomatoes was used and no insecticide or fungicide treatments were applied. Daily maximum and minimum temperatures were registered with a data logger during both trials. In each crop cycle, four release treatments were applied: (1) E. mundus (12 adults/plant in 6 releases); (2) M. caliginosus (0.6 adults/plant in 2 releases); (3) E. mundus and M. caliginosus (12 E. mundus and 0.6 M. caliginosus/plant in 6 and 2 releases, respectively); and (4) control without releases of natural enemies. Treatments were randomly assigned to each compartment in a Completely Randomized Block Design with 3 replications. Three B. tabaci adults per plant were introduced into each compartment during the same week as the transplant. Fourteen plants were randomly selected from each cage every week and adult whitefly and mirids (adults and nymphs) were counted on the 7 youngest leaves. To assess the density of B. tabaci alive pupae, 30 leaflets were randomly collected at fortnightly intervals from the strata where B. tabaci adults were emerging. Percentage reduction of whitefly (pupae and adults) versus control at the end of the tomato crop achieved with the inoculation of natural enemies was calculated. At the end of the crop, mirid densities were compared among treatments using an ANOVA, after previously transforming data by Log (x+0.5) in order to homogenize variances. Results and discussion The maximum adult whitefly density in control cages receiving no natural enemies was more than 60 times higher for the spring than the autumn crop cycle, with 3700 as opposed to 60 adults per plant, respectively, at the end of the crop. Similarly, the average number of nymphs per leaflet was 6 in spring and 0.4 in autumn. During the spring cycle, temperatures ranged between 26ºC to 31ºC. However, during the autumn cycle, mean greenhouse temperatures dropped from 32ºC in August to 16ºC in November. The two treatments including releases of E. mundus were effective in reducing whitefly populations in both crop cycles (Figure 1). In the autumn crop, the percentage adult whitefly reduction was greatest (>98% reduction) for treatments including E. mundus releases. The combined use of E. mundus and M. caliginosus produced the same amount of reduction. On the contrary, the reduction in the number of whitefly pupae was just about 80%, and in this case the control was improved when E. mundus and M. caliginosus were combined. In the spring crop, there was a reduction of at least 80 % in adult whitefly associated with the release of E. mundus and M. caliginosus, whether individually or in combination. Similar results were obtained for pupae control. The greatest reduction in terms of both adults and pupae of B. tabaci was achieved with the combined use of E. mundus and M. caliginosus (99% of adults and nymphs). Although, the use of E. mundus alone and in combination with M. caliginosus gave similar percentages of reduction for the two crop cycles, it is necessary to keep in mind that 72
85 the whitefly populations were very different in the two crop seasons. Whereas whitefly populations were reduced to fewer than one adult per plant with E. mundus in autumn, in spring the number of adults per plant in compartments where the parasitoid was used was around 60. In compartments with M. caliginosus releases, there was a large reduction in whitefly adults and pupae in the spring crop, but in autumn a reduction of only 20% of adult whitefly was achieved. Better mirid establishment were obtained in spring than in autumn, regardless of whether the mirid was released alone or in combination with E. mundus. However, in spring, the presence of the parasitoid significantly reduced the number of mirids per plant at the end of the crop (F=8.25; df 1.5; P=0.04) (Figure 2). Fall crop Spring crop % of whitefly reduction vs. control Adults Pupae Em Mc Em + Mc Figure 1. Percentage reduction of whitefly (pupae and adults) versus control at the end of the tomato crop achieved with the inoculation of E. mundus (Em), M. caliginosus (Mc) or with both natural enemies (Em+Mc) Although the initial level of infestation was the same for both crop cycles, much greater populations of B. tabaci were established in the spring than in the autumn crop. This was probably due to the high temperature,. but also to the photoperiod. B. tabaci is very well adapted to high temperatures (Dorst et al. 1998) and greenhouse temperatures remained within the optimal range for this pest throughout the spring crop. Conversely, in autumn, temperatures dropped below 16ºC in November. With these falling temperatures, the increase in the B. tabaci population was slower. 73
86 Mc Mc + Em No. mirid bugs/plant % of plants with bugs Autumn crop Spring crop Figure 2. Percentage of tomato plants with mirid bugs and number (mean ± SE) of mirid bugs per plant after releasing M. caliginosus and M. caliginosus + E. mundus for the autumn and spring crop cycles. According to our results, 12 E. mundus per plant were clearly sufficient to control whitefly populations in autumn. This is a much lower release rate than that proposed by Stansly et al. (2005b), who claimed that 82.5 wasps per plant were needed to control whitefly in autumn. Although the percentage of whitefly reduction was at least 80 % in spring, the increasing temperatures and photoperiod favoured the build up of B. tabaci rather than E. mundus populations. As a result, at the end of the crop, there were too many adult whitefly on the plant. Stansly et al. (2005b) also found that controlling B. tabaci with E. mundus was less effective in spring than in autumn, even with very high release rates. However, when lower release rates of E. mundus were made in commercial greenhouses, whitefly control was observed although with great variability among greenhouses (Stansly et al. 2005a). This was possibly due to the spontaneous colonization of natural populations of E. mundus that complemented the dose of released parasitoids. The different size of these natural populations caused the observed variability. Under our conditions, whitefly populations were not satisfactorily controlled by the M. caliginosus released in spring, in spite of the important reduction in whitefly populations obtained. The M. caliginosus release rate used in this essay (0.6 adults/plant) is the recommended dose for whitefly control in spring greenhouse tomatoes in our area (northeast Spain) where large populations of native M. caliginosus colonize greenhouses and 74
87 complement the dose of released predators (Albajes et al. 2003). However, in greenhouses without predator colonization this release rate may be too low since Malausa & Trottin Caudal (1996) found that almost one M. caliginosus per plant was needed to effectively control greenhouse whitefly. In the autumn crop, the establishment of M. caliginosus was much poorer than in spring. These differences are probably not only explained by those between mean temperatures in the two crop cycles (26º C in autumn and 29º C in spring) but also by the reduced availability of prey, which has already been discussed in relation with the abundance of B. tabaci and crop cycles. Our results indicate that releases of M. caliginosus combined with E. mundus improved B. tabaci control in the spring crop cycle when climatic conditions were more favourable to the pest. Probably, the combined use of a parasitoid and a predator made possible to control the whitefly population by the parasitoid during the time required for mirid bugs to complete their first generation in the crop (Malausa & Trottin Caudal, 1996). The control subsequently benefited from the complementary action of the two natural enemies. Under Mediterranean conditions, control of B. tabaci on tomato crops tends to focus more on reducing virus transmission than on preventing direct fruit damage, since a viruliferous whitefly adult is able to efficiently transmit Tomato Yellow Leaf Curl Virus with one phloem contact lasting <2 min (Jiang et al. 2000). Consequently, biological control of B. tabaci by releases of E. mundus and M. caliginosus could be a good control strategy that would prevent adult whitefly emergence. The establishment of predators could permit to control subsequent crop colonization by whitefly. Acknowledgements The technical assistance of Pilar Hernandez, Francisca Oliver and Victor Muñoz is gratefully acknowledged. This work was funded by the Spanish Research Agency (AGL ). References Albajes, R., M.J. Sarasúa, J. Avilla, J. Arnó & R. Gabarra. 2003: Integrated Pest Management in a Mediterranean Region: the case of Catalonia. In K. S. Maredia, D. Dakouo and D, Mota-Sanchez (eds) Integrated Pest Management in the Global Arena, CAB International Publisher, UK, pp Arnó, J., Matas, M., Martí, M., Ariño, J. & Gabarra, R. (2005). Coexistence between Trialeurodes vaporariorum and Bemisia tabaci and impact of natural enemies in tomato crops under Mediterranean conditions. IOBC/WPRS Bull. 28(1): 1-4. Barnadas, I., R. Gabarra & R. Albajes 1998: Predatory capacity of two mirid bugs preying on Bemisia tabaci. Entomol. Exp. Appl. 86: Castañé, C., Alomar, O., Goula M. & Gabarra R. 2004: Colonization of tomato greenhouses by the predatory mirid bugs Macrolophus caliginosus and Dicyphus tamaninii. Biological Control 30(3): Dorst, Y.C., van Lenteren, J.C. & van Roermund, H.J.W. 1998: Life-history parameters of different biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae) in relation to temperature and host plant: a selective review. Bull. Entomol. Res. 88: Gerling, D., Alomar, O. & Arnó, J Biological control of Bemisia tabaci using predators and parasitoids. Crop Prot. 20:
88 González-Zamora, J.E., Moreno Vázquez, R., Rodríguez Rodríguez, M.D., Rodríguez, M.P., Mirasol, E., Lastres, J. & Manzanares Ruiz, C. 1996: Evolución del parasitismo en Bemisia tabaci (Genn.) y Trialeurodes vaporariorum (West.) (Hom.: Aleyrodidae) en invernaderos de Almería. Bol. San. Veg. Plagas 22: Greenberg, S.M., Jones, W.A. & Liu, T.-X. 2002: Interactions among species of Eretmocerus (Hymenoptera : Aphelinidae), two species of whiteflies (Homoptera: Aleyrodidae), and tomato. Environ. Entomol. 31(2): Jiang, Y.X., De Blas, C., Barrios, L & Fereres, A. 2000: Correlation between whitefly (Homoptera: Aleyrodidae) feeding behavior and transmission of Tomato Yellow Leaf Curl Virus. Ann. Entomol. Soc. Am. 93(3): Malausa, J.C. & Trottin-Caudal, Y. 1996: Advances in the strategy of use of the predaceous bug Macrolophus caliginosus (Heteroptera: Miridae) in glasshouse crops. In: Zoophytophagous heteroptera: Implications for life history and Integrated Pest Management, eds. Alomar, O. & Wiedenmann, R. N. Entomol. Soc of America Maryland Stansly, P.A., Calvo, J., & Urbaneja, A a: Augmentative biological control of Bemisia tabaci biotype Q in Spanish greenhouse pepper production using Eretmocerus spp. Crop Prot. 24: Stansly, P.A., Calvo, J., & Urbaneja, A b: Release rates for control of Bemisia tabaci (Homoptera: Aleyrodidae) biotype Q with Eretmocerus mundus (Hymenoptera: Aphelinidae) in greenhouse tomato and pepper. Biological Control 35(2):
89 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Amblyseius swirskii (Acari: Phytoseiidae) as a biological control agent of the tobacco whitefly Bemisia tabaci (Hom.: Aleyrodidae) in protected sweet pepper crops in Southern Spain. Javier Calvo 1, Paloma Fernández 1, Karel Bolckmans 2, José Eduardo Belda 1. 1 Departamento I+D, Koppert Biological Systems, P.O. BOX 286, Águilas (Murcia), Spain jcalvo@koppert.es. 2 Koppert Biological Systems, P.O. BOX 155, 2650 AD Berkel en Rodenrijs, The Netherlands. Abstract: The tobacco whitefly Bemisia tabaci is a key pest in horticultural crops in Southern Spain including sweet pepper. The parasitoid E. mundus is now the basis of biological control of tobacco whiteflies in biocontrol based IPM programs in sweet pepper crops in Spain. Nevertheless in case of high infestation levels of B. tabaci in sweet pepper crops chemical pesticides are usually needed in combination with E. mundus. Using incompatible pesticides can seriously hamper the establishment and development of E. mundus and other natural enemies in the crop. Therefore more effective biological control programs are needed. The focus of this study was to investigate the possibilities of the phytoseiid predatory mite A. swirskii as a biological control agent of B. tabaci in protected sweet pepper crops in Southern Spain. First the efficacy of A. swirskii was tested under semi-field conditions by releasing a total of 8 adults of B. tabaci/plant and comparing three release rates of the predatory mite A. swirskii (0, 25 and 100 A. swirskii/m 2 ). In a second trial conducted under semi-field conditions, 4 different biological control strategies were compared: strategy EM, receiving 24 E. mundus/m 2 ; strategy NT, receiving 24 E. mundus /m 2 and 2 N. tenuis/plant; strategy AS, receiving 12 E. mundus /m 2 and 50 A. swirskii/m 2 ; and strategy MIX, receiving 12 E. mundus/m 2, 50 A. swirskii/m 2 and 2 N. tenuis/plant. The plants were infested by releasing a total of 50 adults of B. tabaci per plant. Finally, the efficacy and establishment of A. swirskii against B. tabaci in sweet pepper were studied under field conditions. Under semi field conditions A. swirskii at a release rate of 25 and 100 mites/m 2 was able to suppress almost totally, a initial infestation of 8 B. tabaci adults per plant. The combination of 50 A. swirskii/m 2 and 12 E. mundus/m 2, was the most efficient strategy against an initial infestation of 50 adults of B. tabaci per plant. These results were confirmed under field conditions, where the population of B. tabaci was always controlled very well in the plots where A. swirskii was released compared to the plots where the mite was not released. According to these results, A. swirskii proves to be a great candidate as biological control agent against B. tabaci in protected sweet pepper crops in Southern Spain making the control of B. tabaci possible by only using biocontrol agents, even in case of high infestation levels. Key words: Amblyseius swirskii, Bemisia tabaci, biological control, sweet pepper, greenhouses. Introduction The tobacco whitefly Bemisia tabaci Genn. (Hom.: Aleyrodidae) is a key pest in protected sweet pepper crops in Southern Spain. The parasitic wasp Eretmocerus mundus Mercet (Hym.: Aphelinidae) has been described as the most abundant naturally occurring parasitoid of B. tabaci in greenhouses in Southern Spain (Rodríguez-Rodríguez et al., 1994) and the most efficient in this conditions on this host (Stansly et al, 2005) and it is now the basis of biological control of tobacco whiteflies in biocontrol based IPM programs in sweet pepper crops in Spain. On the other hand, it is usually, in crops which are planted during summer, which is the case in Almería, pest pressure from outside can be very high and pest population 77
90 development very fast. In case of high infestation levels, it is needed to apply pesticides in combination with E. mundus to keep the population of B. tabaci under control. Usage of incompatible pesticides can seriously hamper the establishment and population development of biological control agents. The combination of other biocontrol agents with E. mundus can potentially reduce the need to use pesticides, improving the biological control of B. tabaci and the biological control system in sweet pepper IPM programs in general. Recent studies in the Netherlands have shown that the predatory mite Amblyseius swirskii Athias-Henriot (Acari.: Phytoseiidae) is an efficient natural enemy of B.tabaci and Trialeurodes vaporariorum (Nomikou et al., 2001; Hoogerbrugge et al., 2005). A. swirskii can also feed on pollen (Ragusa & Swirski, 1975; Nomikou et al., 2004; van Houten, et al., 2005) making possible its preventive release prior to the occurrence of pests in pollen producing crops such as sweet peppers (Hoogerbrugge et al., 2005). The goal of the present experiments was to test the efficacy of A. swirskii as a biological control agent of B. tabaci in protected sweet pepper crops in Southern Spain. Material and methods Semi-field trial (I): Establishment of A. swirskii and efficacy against B. tabaci. The experiment was conducted in a 40 x 10 m twin-skin plastic tunnel located at the Koppert Biological Systems facilities in Aguilas (Murcia, Spain), provided with pad and fan cooling and heating. The research greenhouse was divided in 36 walk-in cages of 2 x 4 x 3.5 m (l x w x h), of which 12 were used in the present experiment. Cages are entirely constructed from anti-thrips polyethylene screening of 220 x 331 µm and the floor is covered with 2 mm thick white polyethylene ground cloth. Two release rates (25 and 100 A. swirskii/m 2 ) and an untreated control were compared using a complete randomized block design of 3 treatments and 4 replicates. Ten sweet pepper plants were placed in each cage. To calculate the releases of A. swirskii per plant a density of 2.5 plants/m 2 was used. Plants were infested releasing a total of 8 B. tabaci adults/plant divided over three sequential releases ( adults/plant) at a weekly interval (weeks 0, 1 and 2). The predatory mites were released in week 1, one week after the first release of whiteflies. Semi field-trial (II): Comparison of 4 different strategies for biological control of B. tabaci. This experiment was conducted in the same greenhouse as the previous experiment. Twelve cages were used for this trial. The efficacy of 4 different biological control strategies was compared using a complete randomized block design of 4 treatments and 3 replicates. The strategies were: E. mundus EM, N. tenuis NT, A. swirskii AS, and mixture MIX. The releases rates and weeks of natural enemies and pest are summarized in the Table1. Field-trial: First results of establishment and efficacy of A. swirskii against B. tabaci in Spanish sweet pepper crops The experiment was conducted in 4 commercial greenhouses, 2 Parral plastic greenhouses of 2000 m 2 in Campo de Cartagena (Murcia, Spain) planted in February with sweet pepper cv. Herminio and 2 Parral plastic greenhouses of 4000 m 2 in Dalías (Almería, Spain) planted with sweet pepper cv. Requena also in February. In both places a complete randomized block design of 2 treatment and 4 replicates was used to test the efficacy of A. swirskii against B. tabaci. The treatments were E. mundus alone and E. mundus + A. swirskii. One release at a rate of 50 and 75 A. swirskii/m 2 was done in Campo de Cartagena the 5 th of May (week 0) and Almería one day after respectively. A total of 5 E. mundus/m 2 were released in Campo de Cartagena and Almería in the A. swirskii treatment and a total of 5 and 27 E. mundus/m 2 were released in the E. mundus treatment in Campo de Cartagena and Almería respectively. 78
91 Pyridaben, soap and pyriproxifen were sprayed two times during the trial in the control treatment in Almería. Table 1. Releases of B. tabaci and natural enemies in each strategy. Strategy Release Rate E. mundus N. tenuis A. swirskii Mix B. tabaci (Adult/plant) (5X; 0-4) * 50 (5X; 0-4) 50 (5X; 0-4) 50 (5X; 0-4) E. mundus (pupae/m 2 ) 6 24 (4X; 1-4) 24 (4X; 1-4) 12 (2X; 1-2) 12 (2X; 1-2) N. tenuis (Adult/plant) 2-2 (1X; 2) - 2 (1X; 2) A. swirskii (mites/m2) (1X; 1) 50 (1X; 1) *Total of individuals released (Number of releases; weeks in which the release were done). Results and discussion Semi-field trial (I): Establishment of A. swirskii and efficacy against B. tabaci The number B. tabaci nymphs per leaf reached higher levels in the control treatment than the treatments with releases of A. swirskii (Figure 1A). The highest number of nymphs per leaf found in the control treatment was 55.14±15.16 at the end of the trial, while in the treatments receiving 25 and 100 A. swirskii/m 2 this number was 1.50±0.48 and 0.57±0.18 respectively (Figure 1A). At both release rates the A. swirskii population level increased fast (Fig. 1B), rising up to 3.42±0.47 in the sixth week with 25 A. swirskii / m 2 and 1.96±0.22 in the fifth week with 100 A. swirskii/m 2 (Figure 1B). At the beginning of the trial the higher values of mites per leaf were observed in the treatment receiving 100 A. swirskii / m 2 but in the last three weeks the treatment receiving 25 A. swirskii/m 2 showed the higher numbers of mites per leaf (Figure 1B). A. swirskii was also observed in the control cages, and its population rose up to 2.68±0.46 mites per leaf in the last week of the trial (Figure 1B). Semi field trial (II): Comparison of 4 different strategies Lower levels of nymphs of B. tabaci per leaf were always observed in the treatments receiving A. swirskii. In the treatment EM (E. mundus alone) and NT (E. mundus + N. tenuis) the highest number of nymphs per leaf observed was 35.5±5.98 and 29.3±4.00 respectively while in the treatments where A. swirskii was released this value reached to 12.5±4.27 in the treatment AS (E. mundus + A. swirskii) and 10.9 ± 2.32 in the treatment MIX (E. mundus + N. tenuis + A. swirskii) (Figure 2). Furthermore, no significant reductions in the number of nymphs of B. tabaci per leaf was observed between the treatments with and without N. tenuis releases (Figure 2), in other words, between EM and NT and AS and MIX. Field trials The evolution of B. tabaci nymphs population was similar in both places but the number of nymphs per leaf in the E. mundus alone treatment in Almería was about 30 times higher than in Campo de Cartagena. In Almería 65.5±6.10 nymphs per leaf were counted while in Campo de Cartagena the highest number of nymphs of B. tabaci per leaf observed was 2.15±0.52 (Figure 3 A and C). In the plots where A. swirskii + E. mundus was released the number of nymphs of B.tabaci was always lower than 1 and 0.1 in Almería and Campo de Cartagena respectively, 79
92 while in the treatments without release of the predatory mite the number of nymphs per leaf increased steeply, rising up to the values showed before (Figure 3 A and C) A 3 B Ind / leaf 40 Ind / leaf W0 W1 W2 W3 W4 W5 W6 W7 W8 0 W0 W1 W2 W3 W4 W5 W6 W7 W8 Week trial Week trial Control 100 A. swirskii/m2 25 A. swirskii/m2 Control 100 A. swirskii/m2 25 A. swirskii/m2 Figure 1. Nymphs of B. tabaci (A) and mites (B) (mobile forms) per leaf (mean ± standard error) observed in each week in the treatments receiving 0 (Control), 100 and 25 A. swirskii/m Ind / Leaf W0 W1 W2 W3 W4 W5 W6 Week trial W7 W8 W9 W10 W11 W12 EM NT AS MIX Figure 2. Nymphs of B. tabaci per leaf (mean ± standard error) observed every week in each strategy. Strategies: EM (24 E. mundus/m 2 ); NT (24 E. mundus/m N. tenuis/plant); AS (12 E. mundus/m A. swirskii/m 2 + MIX); MIX (12 E. mundus/m A. swirskii/m N. tenuis/plant). A. swirskii was able to establish in both crop areas, keeping high population levels during the entire experiment (Figure 3 B and D). In Campo de Cartagena the population of A. swirskii increased steeply and reached 5.1±0.30 while in Almería, where the trend was more constant, a maximum of 2.24±0.16 A. swirskii per leaf was observed (Figure 3 B and D). In the control 80
93 treatment A. swirskii also appeared and its population rose up to 1.87±0.19 and 0.17±0.10 mites per leaf in Campo de Cartagena and Almería respectively. Ind / leaf W0 W1 W2 W3 W4 W5 W6 0 A W7 Control W8 W9 Week Trial A. swirskii W10 W11 W12 W13 W14 Ind / leaf W0 W1 W2 W3 W4 W5 W6 1 0 B W7 Control W8 W9 Week Trial A. swirskii W10 W11 W12 W13 W14 Ind / leaf W 0 W1 W2 W3 W4 W 5 W6 0 C Control W7 W 8 W9 Week Trial A. swirskii W10 W11 W12 W13 W14 Ind / leaf W0 W1 W2 W 3 W4 W5 W6 0 D W7 Control W8 W9 Week Trial A. swirskii W10 W11 W12 W13 W14 Figure 3. Number of nymphs of B. tabaci per leaf (mean ± standard error) in each week in the treatment control and A. swirskii in Campo de Cartagena (A) and Almería (C). Number of mites (mobile forms) per leaf (mean ± standard error) in each week in the treatment control and A. swirskii in Campo de Cartagena (B) and Almería (D). The phytoseiid predator A. swirskii was able to establish and reproduce in sweet pepper under semi field conditions, rising to high population levels. This results were also confirmed by the experiment carried out in Campo the Cartagena and Almería in commercial sweet pepper greenhouses, where A. swirskii was able to establish and maintain a population during the entire experiment, and were similar than obtained by Hoogerbrugge et al. (2005) in the Netherlands. In semi field conditions A. swirskii was able to suppress an initial infestation of 8 B. tabaci adults per plant with a release of 25 and 100 A. swirskii/m 2. In the second semi-field trial, a release of 50 A. swirskii/m 2 and 12 E. mundus/m 2 was able to control efficiently an infestation of 50 B. tabaci adults per plant. These results are consistent with the results obtained by Nomikou et al. (2001) where under laboratory conditions, and using cucumber as 81
94 a plant host, A. swirskii was also able to suppress efficiently to B. tabaci. Furthermore, under semi field conditions the strategy combining E. mundus and A. swirskii was the most efficient compared with strategies receiving only E. mundus or strategies receiving E. mundus and the mirid bug N. tenuis, so it would be the best strategy to control B. tabaci. This was confirmed by the trials conducted in commercial greenhouses, where the population of B. tabaci was always significantly lower in the plots where both A. swirskii and E. mundus had been released compared to the plots receiving only E. mundus. These results show the great possibilities of the predatory mite A. swirskii as an augmentation biological control agent of the tobacco whitefly B. tabaci in protected sweet pepper crops in Southern Spain. The use of the phytoseiid predatory mite A. swirskii will improve the biological control of B. tabaci in particular, reducing the use of chemical insecticides. Based on he efficacy of A. swirskii against Tobacco Whitefly B. tabaci, and against Western Flower Thrips Frankliniella occidentalis (van Houten et al., 2005), the implementation of A. swirskii in greenhouse sweet peppers in Southern Spain will be a major step forward towards sustainable crop production in this area. References Hoogerbrugge, H., Calvo., J., van Houten, Y. & Bolckmans, K. 2005: Biological control of the tobacco whitefly Bemisia tabaci with the predatory mite Amblyseius swirskii in sweet pepper crops. IOBC/WPRS Bull. 28 (1): Nomikou, M., Janssen, A., Schraag, R. & Sabelis, M.W. 2001: Phytoseiid predators as potential biological control agents for Bemisia tabaci. Exp. Appl. Acarol. 25: Nomikou, M., Janssen, A., Schraag, R. & Sabelis, M.W., 2004: Vulnerability of Bemisia tabaci inmatures to phytoseiid predators: Consequences for oviposition and influence of alternative food. Entomol. Exp. Appl Ragusa, S., Swirskii, E. 1975: Feeding habits, development and oviposition of the predacious mite Amblyseius swirskii Athias-Henriot (Acarina: Phytoseiidae) on pollen of various weeds. Israel J. Entomol. 10: Rodríguez-Rodríguez, M.D., Moreno, R., Téllez, M.M., Rodríguez-Rodríguez, M.P. & Fernández-Fernández, R. 1994: Eretmocerus mundus (Mercet), Encarsia lutea y Encarsia transvena (Timberlake) (Hym., Aphelinidae) parasitoides de Bemisia tabaci (Homoptera: Aleyrodidae) en los cultivos hortícolas protegidos almerienses. Bol. San. Veg. Plagas 20: Stansly, P.A., Calvo, F.J. & Urbaneja, A. 2005: Augmentative biological control of Bemisia tabaci biotype Q in Spanish greenhouse pepper production using Eretmocerus spp. Crop Prot. 24: Van Houten, Y.M., Ostlie, M.L., Hogerbrugge, H. & Bolckmans, K. 2005: Biological control of western flower thrips on sweet pepper using the predatory mites Amblyseius cucumeris, Iphiseius degenerans, A. andersoni and A. swirskii. IOBC/WPRS Bull. 28(1):
95 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Efficacy of the Beauveria bassiana strain ATCC (Naturalis ) against whiteflies on protected crops Fernando Mayoral ¹, Massimo Benuzzi², Edith Ladurner² ¹Agrichem Bio S.A., Plaza Castilla 3-14 A, E Madrid, Spain; ²Intrachem Bio Italia S.p.A., R&D Department, Via Calcinaro 2085/7, I Cesena, Italy Abstract: In 2004, the efficacy of the B. bassiana-based bioinsecticide, applied at different doses, in controlling whiteflies was tested on protected tomato in Italy and on greenhouse eggplant in Spain. In Italy on tomato, all B. bassiana treatments, irrespective of the dose (doses tested: 125, 250, and 300 ml/hl), significantly reduced the whitefly infestation compared to the untreated control, with their efficacy ranging from 72.3 to 82.8%. Similar results were obtained in Spain on eggplant: all B. bassiana treatments significantly reduced the whitefly infestation compared to the untreated control, but a significant dose-response effect was recorded. The efficacy of Naturalis alone at 200 ml/hl was lower than that of the chemical imidacloprid-based reference product, of the bioinsecticide applied at the same dose in tank mixture with an adjuvant, and of the biocontrol agent alone at 300 ml/hl. However, when the product was applied at 200 ml/hl in tank mixture with the adjuvant and alone at 300 ml/hl, its efficacy was always comparable to that of the chemical standard. These trials, conducted on two different crops in two different mediterranean countries, confirm the results of previous trials carried out in other countries and on other crops: the B. bassiana-based product Naturalis is an efficient tool for controlling whitefly infestations on protected crops. Key words: entomopathogen, bioinsecticide, Aleurodidae, biocontrol, integrated control Introduction Beauveria bassiana (Balsamo) Vuillemin (Deuteromycetes, Moniliales) is an entomopathogenic fungus, recognized in 1835 by Agostino Bassi as the causal agent of the white muscardine disease of the silk worm. It can affect a wide range of arthropod pests, such as coleopterans, mites, homopteran and heteropteran hemipterans, and all their developmental stages (eggs, immature stages, and adults), provided that these are present on the outside of the plants (Knauf, 1992; Wright & Kennedy, 1996; Lacey et al., 1999; Benuzzi et al., 2001). The microrganism acts primarily by contact. Once attached to the insect s cuticle, its conidia germinate producing penetration hyphae, which penetrate the cuticle and proliferate in the insect s body. High ambient humidity and free water favour conidial germination (Hallsworth & Magan, 1999; Benuzzi & Santopolo, 2001), but B. bassiana conidia are invasive and pathogenic at all ambient humidity levels (Ramoska, 1983; Lecuona et al., 2001). Infection can take between 24 and 48 hours, depending on the temperature (BCPC, 2004). The proliferation of the fungus inside the insect s body leads to the insect s death within 3-5 days. The B. bassiana mycelium multiplies by feeding on the host and consuming its nutrients. After the insect s death, conidiospores can be produced on the outside of its body and new conidia may be released on the outside of the insect cadaver (Pekrul & Grula, 1979; Feng et al., 1994; BCPC, 2004), with temperature and humidity affecting the sporulation of B. bassiana, and thus the transmission of the fungus to other insects (Fargues & Luz, 1998; Luz & Fargues, 1998). 83
96 The formulated bioinsecticide Naturalis is a concentrated suspension of spores of the B. bassiana strain ATCC The original strain has been isolated from Anthonomus grandis (Boheman), the cotton boll weevil, in the Lower Rio Grande valley, Texas, USA (BCPC, 2004). The penetration hyphae perforate the cuticle by means of special enzymes, causing the dehydration and finally the death of the insect (Benuzzi & Vacante, 2004). Several years of experimental applications of this entomopathogenic B. bassiana strain against the boll weevil showed that also other insect pests, such as whiteflies, thrips and leafhoppers, are susceptible to infection (Knauf, 1992; Wright & Kennedy, 1996). The formulated product (henceforth f.p.) contains at least 2.3 x 10 7 viable spores / ml f.p. Since this bioinsecticide is a suspension of conidiospores in natural oil, it can be used also in organic farming. In 2005, Intrachem Bio International S.A. (Geneva, Switzerland) acquired the production rights for Naturalis from Troy Biosciences, and the commercial product is now formulated in Europe. Whiteflies (Homoptera, Aleurodidae) are major agricultural pests worldwide: in subtropical and tropical areas, on both protected and field crops in the Mediterranean climates, and in greenhouses in the temperate zones (Denholm & Horowitz, 2000; Benuzzi & Vacante, 2004). They suck phloem sap, excrete sticky honeydew, cause yellowing or death of leaves, and can act as vectors of several important plant virus diseases (Schuster et al., 1996). Most whiteflies, especially the most common pest species Trialeurodes vaporariorum and Bemisia spp., have a wide host range that includes many weeds and crops. Because of their fast reproduction rate (several overlapping generations a year), inadequate physical barriers and management of crop residues, and low infestation threshold values, the management of heavy whitefly infestations is extremely difficult. In many situations, natural enemies may provide adequate control, but outbreaks frequently occur when natural enemies are disrupted or destroyed by insecticide applications (Benuzzi & Vacante, 2004). Due to the negative side effects of pesticides on beneficials, the increased number of pesticides no longer registered for greenhouse productions in several countries, residue issues, and especially due to the development of resistance in whiteflies to a wide range of chemical control agents (Denholm & Horowitz, 2000; Elbert & Nauen, 2000; Gorman, 2005), there is increasing demand for alternative methods for whitefly control and urgent need for effective resistance management strategies. In this paper, we report the results of efficacy trials with the B. bassiana-based bioinsecticide against whiteflies, conducted in 2004 in greenhouses on eggplant and tomato under the supervision of the Spanish and Italian national distribution companies of Intrachem Bio International S.A., Agrichem S.A. and Intrachem Bio Italia S.p.A. Material and methods B. bassiana against whiteflies on protected tomato The trial was carried out on greenhouse tomato (Lycopersicon esculentum Miller) cv Naomi in Molfetta, Bari, Italy, by a certified Italian testing facility in compliance with the EPPO guidelines and with the Principles of Good Experimental Practices (GEP). The efficacy of applications of the B. bassiana-based bioinsecticide Naturalis (a.s. B. bassiana strain ATCC 74040; SC 7.16%) at 125, 250, and 300 ml/hl, was compared to that of an untreated control and of the chemical reference insecticide Confidor 200 SL (a.s. imidacloprid; SL 17.8%), applied at 75ml/hl. To compare the five treatments, a fully randomized block design was used with 4 replicates per treatment, and with 10 plants per plot. The B. bassiana-based treatments were applied at 6-7-day intervals for a total of 7 applications per treatment, while the chemical 84
97 standard was applied at 14-day intervals for a total of 4 applications. The treatments were applied starting from the beginning of fruit colour change (August 26) to the second pick (October 10). Four days after the application of the last treatment, the number of living whitefly nymphs (1 st - 4 th nymphal instars) was counted on 20 randomly selected leaves per plot, and the efficacy according to Abbott (Abbott, 1925) of the different treatments in reducing the number of living nymphs on leaves was calculated. The total number of living nymphs was compared across treatments using one-way ANOVA, followed by the Student-Newman-Keuls test for posthoc comparison of means. B. bassiana against whiteflies on protected eggplant The trial was conducted on greenhouse eggplant (Solanum melongena L.) cv Cristal in La Mojonera, Almeria, Spain, by Agrichem S.A. in compliance with the EPPO guidelines and with the Principles of Good Experimental Practices (GEP). The efficacy of applications of the B. bassiana-based bioinsecticide Naturalis (a.s. B. bassiana strain ATCC 74040; SC 7.16%) alone at 200 and 300 ml/hl, respectively, and at 200 ml/hl in tank mixture with the adjuvant NuFilm 17 (a.s. di-1-p-menthene; EL 904 g/l) at 100 ml/hl was compared to that of an untreated control and of the chemical reference insecticide Confidor 200 SL (a.s. imidacloprid; SL 17.8%), applied at 75ml/hl. To compare the five treatments, a fully randomized block design was used with 5 replicates per treatment, and with 10 plants per plot. All treatments were applied twice (May 18 and 25), starting from the beginning of flowering. In each plot, the number of living adults was counted on 2 randomly selected leaves per plant on 4 successive assessment dates: the day of the first application (May 18, to verify whether the whitefly infestation level was homogeneous among plots), three days after the first application (May 21), 7 days after the first application (May 25), and 7 days after the second application (June 1). For the last three assessment dates, the efficacy according to Abbott (Abbott, 1925) of the different treatments in reducing the number of living adults per leaf was determined. For the last three assessment dates, the numbers of living adults per leaf were compared across treatments using one-way ANOVAs, followed by the Student-Newman-Keuls test for posthoc comparison of means. To improve homoscedasticity of variances, data were arsen (x+0.5)-transformed. Results and discussion B. bassiana against whiteflies on protected tomato Differences among treatments in the number of living whitefly nymphs were significant: it was highest in the untreated control plots, intermediate in the plots treated with the B. bassiana-based product at the different doses, and lowest in the plots treated with the reference insecticide (Table 1). Irrespective of the dose applied, all B. bassiana treatments significantly reduced the whitefly infestation compared to the untreated control, with their efficacy ranging from 72.3 to 82.8%. The chemical imidacloprid-based reference treatment showed highest efficacy, and significantly differed from all the other treatments. Resistance to this neonicotinoid did therefore not yet occur in the target whitefly population. B. bassiana against whiteflies on protected eggplant Within the greenhouse, all plots showed similar whitefly infestation levels prior to the application of the first treatment (Table 2). On the three successive assessment dates, differences among treatments in the number of living whitefly adults per leaf were always significant. For all treatments, the whitefly infestation decreased over time, while it increased in the untreated control (Table 2). The 85
98 mean number of whitefly adults per leaf was always lowest in the plots treated with the chemical imidacloprid-based standard, the bioinsecticide applied in tank mixture with the adjuvant ( ml/hl), and with the bioinsecticide alone at 300 ml/hl (with the 3 treatments never differing significantly), intermediate in the plots treated with the B. bassiana-based product at 200 ml/hl, and highest in the untreated control plots (Table 2). At the dose of 200 ml/hl, the bioinsecticide alone significantly reduced the whitefly infestation compared to the untreated control, but its performance was significantly lower than that of the chemical reference product, the bioinsecticide at 200 ml/hl in tank mixture with the adjuvant, and that of the bioinsecticide alone at 300 ml/hl (Table 2). However, when applied at the same dose in tank mixture with the adjuvant, and when applied at a higher dose (300 ml/hl), the efficacy of the B. bassiana-based product was always comparable to that of the chemical standard. In conclusion, the trials carried out on two different vegetable crops (eggplant and tomato) in two different mediterranean countries confirm the results of previous trials conducted on other crops and in other countries (Knauf, 1992; Brownbridge et al., 1996; Murphy et al., 1998; Siongers & Coosemans, 2003): the B. bassiana-based product Naturalis is highly efficient in controlling whitefly infestations on protected crops. Furthermore, the trial carried out in Spain indicates that the efficacy of the product may be increased by adding an adjuvant to the tank mixture. Applications of the B. bassiana-based product can thus be an important and useful tool not only in organic farming but also for conventional growers, because the inclusion of B. bassiana in integrated pest management strategies of whiteflies can help to develop effective resistance management strategies, and to avoid the presence of inadequate levels of residues in the final produce (in both Italy and Spain, Naturalis has no pre-harvest interval). References Abbott, W.S. 1925: A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18: BCPC 2004: Beauveria bassiana biological insecticide (fungus). In: The Manual of Biocontrol Agents, 3 rd edition, ed. L.G. Copping: Benuzzi, M. & Santopolo, F. 2001: Naturalis: bioinsetticida a base di Beauveria bassiana. Informatore Fitopatologico 4: Benuzzi, M. & Vacante, V. 2004: Difesa fitosanitaria in agricoltura biologica, ed. Il Sole 24 Ore Edagricole S.r.l., Bologna, Italy: 297 pp. Benuzzi, M., Albonetti, E. & Baldoni P.G. 2001: Prova di lotta biologica su fragola contro il ragnetto rosso (Tetranichus urticae) con un formulato a base di Beauveria bassiana. Notiziario sulla protezione delle piante 13: Brownbridge, M., Adamowicz, A., Skinner M. & Parker, B.L. 1996: Management of silverleaf whitefly and western flower thrips with Beauveria bassiana: effect of spray techniques on efficacy, pp In: Abstracts Society for Invertebrate Pathology, 29th Annual Meeting, 1-6 Sept. 1996, Cordoba, Spain. Denholm, I. & Horowitz, R. 2000: Insecticide resistance in whiteflies. Paper presented at the 34 th Meeting of IRAC International, Almeria, Spain, April, Elbert, A. & Nauen, R. 2000: Resistance in Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides in Southern Spain with special reference to neonicotinoids. Pest Manag. Sci. 56:
99 Fargues, J. & Luz, C. 1998: Effects of fluctuating moisture and temperature regimes on sporulation of Beauveria bassiana on cadavers of Rhodnius prolixus. Biocontrol Sci. Technol. 8: Feng, M.G., Proprawski, T.J., Khachatourians, G.G. 1994: Production, formulation and application of the entomopathogenic fungus Beauveria bassiana for insect control: current status. Biocontrol Sci. Technol. 4: Gorman, K. 2005: Resistencia de la mosca blanca a los insecticidas, Bemisia tabacci y Trialeurodes vaporariorum. Phytoma 173: Hallsworth, J.E. & Magan, N. 1999: Water and temperature relations of growth of the entomogenous fungi Beauveria bassiana, Metarhizium anisopliae, and Paecilomyces farinosus. J. Invert. Pathol. 74: Knauf, T.A. 1992: Naturalis-L: a biorational insecticide for boll weevil and whitefly control. Proc. Beltwide Cotton Conference 1: Lacey, L.A., Horton, D.R., Chauvin, R.L. & Stocker, J.M. 1999: Comparative efficacy of Beauveria bassiana, Bacillus thuringiensis, and aldicarb for control of Colorado potato beetle in an irrigated desert agroecosystem and their effects on biodiversity. Entomol. Exp. Appl. 93: Lecuona, R.E., Edelstein, J.D., Berretta, M.F., La Rossa, F.R. & Arcas, J.A. 2001: Evaluation of Beauveria bassiana (Hyphomycetes) strains as potential agents for control of Triatoma infestans (Hemiptera Reduviidae). J. Med. Entomol. 38 (2): Luz, C. & Fargues, J. 1998: Factors affecting conidial production of Beauveria bassiana from fungus-killed cadavers of Rhodnius prolixus. J. Invertebr. Pathol. 72: Murphy, B.C., Morisawa, T.A., Newman, J.P., Tjosvold, S.A. & Parrella, M.P. 1998: Fungal pathogen provides control of western flower thrips in greenhouse flowers. California Agriculture 52 (3): Pekrul, S. & Grula, E.A. 1979: Mode of infection of the corn earworm (Heliothis zea) by Beauveria bassiana as revealed by scanning electron microscopy. J. Invert. Pathol. 34: Ramoska, W.A. 1983: The influence of relative humidity on Beauveria bassiana infectivity and replication in the chinch bug, Blissus leucopterus. J. Invertebr. Pathol. 43: Siongers, C. & Coosemans, J. 2003: Biological control products of greenhouse whitefly (Trialeurodes vaporariorum) with fungal insecticides. Communications in Agricultural and Applied Biological Sciences 68: Schuster, D.J., Stansly P.A. & Polston, J.E. 1996: Expressions of plant damage by Bemisia. In: Bemisia 1995: taxonomy, damage, control and management. Intercept Ltd., Andover, Hants, UK, eds. D. Gerling & R.T. Mayer: Wright, J.E. & Kennedy, F.G. 1996: A new biological product for control of major greenhouse pests. Proc. Brighton Crop Protection Conference. Pest & Disease:
100 Table 1. Total number of living whitefly nymphs (Mean ± S.D.) in the different treatments, and mean efficacy (%) of the treatments in reducing the number of living nymphs. No. Treatment Dose No. living nymphs * % efficacy (a.s.) (ml/hl) (m±s.d.) (mean) 1 Naturalis (B. bassiana) ±57.6 b Naturalis (B. bassiana) ±8 b Naturalis (B. bassiana) ±16.2 b Confidor 200SL (imidacloprid) ±26.0 c Untreated control - 1,483.3±225.3 a * Different letters indicate statistically significant differences (Student-Newman-Keuls-test: P<0.05). Table 2. Number of living adults per leaf (Mean ± S.D.) in the different treatments, and mean efficacy (%) of the treatments in reducing the number of living adults per leaf on the four successive assessment dates. No. Treatment, Assessment date (dd/mm/yy) dose (ml/hl) 18/05/04 21/05/04 25/05/04 01/06/04 No. living adults per leaf (m±s.d.)* 1 Naturalis, ± ±0.2 b 1.4±0.2 b 1.0±0.2 b 2 Naturalis+NuFilm, ± ±0.3 bc 0.7±03 c 0.5±0.2 c 3 Naturalis, ± ±0.4 bc 0.9±0.5 bc 0.6±0.4 c 4 Confidor, ± ±0.3 c 0.5±0.2 c 0.4±0.2 c 5 Untreated control 2.8± ±0.3 a 3.6±0.6 a 4.0±0.4 a % efficacy (mean) 1 Naturalis, Naturalis+NuFilm, Naturalis, Confidor, * Different letters within the same column indicate statistically significant differences (Student-Newman- Keuls-test: P<0.05). 88
101 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Efficacy of the technical grade product and commercial formulation based on Paecilomyces fumosoroseus for controlling whitefly under laboratory conditions Ángeles Padilla-Cubas 1, Soledad Amador 1, Estrella Hernández-Suárez 1, Aurelio Carnero 1, José Manuel Lara 2, Carolina Fernández 2 1 Entomology Dpt., Instituto Canario de Investigaciones Agrarias (ICIA), Crta del Boquerón, La Laguna, Tenerife, 2 Research and Development Dpt., FuturEco S.L., Aragón 264, 5º 2ª, Barcelona, Spain The effects of the BCA Paecilomyces fumosoroseus strain FE 9901 on eggs and immature stages (N 1 and N 4 ) of whiteflies Trialeurodes vaporariorum and Bemisia tabaci, was assessed under laboratory conditions. The ability to produce infections was compared between the technical grade product of BCA (freeze dried blastospores) and the commercial formulation (freeze dried blastospores plus adjuvants) named as FUTURECO NOFLY. The immature whitefly individuals were exposed to fungus on tomato leaf discs sprayed with water suspensions of both products. The fungus susceptibility of immature stages of both whitefly species was the same except in eggs, where T vaporariorum was more susceptible (70-80%) than B. tabaci (40-60%) and there were statistical differences between the formulation and the technical product in the capacity to infect B. tabaci eggs. The efficacy of P. fumosoroseus strain FE 9901 to produce mortality on N 1 and N 4 nymphal stages, was essentially the same (98-100%) when applied as either technical product (freeze dried blastospores), or as a commercial formulation. The selected adjuvants neither interfere with the mode of action of the fungus, nor have insecticide properties by themselves. Key words: Paecilomyces fumosoroseus, biological control, bioinsecticide, blastospore, Bemisia tabaci, Trialeurodes vaporariorum. Introduction In the Canary Islands 18 different whitefly species have been reported (Hernández-Suárez, 1999). Of these, Bemisia tabaci (Gennadius) and Trialeurodes vaporariorum (Westwood) are major pests in commercial tomato greenhouses of the Canary Islands (Hernández-Suárez y Carnero, 2000). Majority of damage is caused by transmition of different virus species, including the Tomato Yellow Leaf Curl Virus (TYLCV), the Tomato Yellow Leaf Curl Sardinia Virus (TYLCSV), Cucurbit Yellow Stunting Disorder Crinivirus (CYSDV) and the Tomato Chlorosis Virus (ToCV) (Espino et al., 2003; Wisler et al., 1999; EWSN, Newsletter, 1999). An increasing number of growers in the archipelago employ beneficial insects to control pests, but effective control of these whiteflies represent a handicap to the implementation of these type of pest management programs. Moreover, pest populations have also developed resistance to commonly used pesticides, even to the recently introduced ones. The lack of effectiveness from traditional treatments, along with the implementation of ecological friendly control strategies, has lead to the research and development of new biological treatment alternatives. Within these practices, the use of Biological Control Agents (BCAs) is seen as an adequate alternative to synthetic pesticides. There are reported different entomopathogenic fungi able to produce infections on whiteflies (Lacey & Fransen, 1995) 89
102 such as Beauveria bassiana (Balsamo) Vuillemin, Paecilomyces fumosoroseus (Wize) Brown and Smith, and Verticillum lecanii (Zimm) Viegas. Many products based on entomopathogenic microorganisms have been very effective in the laboratory only, and fail at some stage in the field. Common causes of this demise are poor stability of the product during storage prior to application, too little active ingredient actually reaching the field target, and rapid degradation of the active material on the target. Formulation plays a vital role in helping to solve these problems and in making an organism effective in practice. The formulation comprises aids to preserving survival forms of the microorganisms (e.g. spores), to delivering them to their targets and -once there- to improving their activities. The most critical decisions about formulations of mass-produced fungus material, like Paecilomyces fumosoroseus, are made at the end point of the operation, i.e. the application of spores to insects in different habitats and the need to ensure their survival in field conditions. The selection of substances is crucial as the spores, as living organisms, should not be subjected to harsh chemical or physical treatment during the formulation process. The only way to check out the suitability of a new formulation is to compare its effectiveness to infect target insects against the technical concentrate from which it was formulated. The objective of this study was to compare the efficacy for producing fungal infections on different stages of B. tabaci and T. vaporariorum life cycle, between the technical grade material (freeze dried blastospores) and a commercial formulation (FUTURECO NOFLY TM ) under laboratory conditions. Material and Methods The laboratory work was conducted at the Instituto Canario de Investigaciones Agrarias (ICIA), placed in Tenerife, Canary Islands, Spain. The project was developed under a Cooperative Research and Development Agreement between the ICIA and the company FuturEco S.L. (Barcelona, Spain) Entomological material The whitefly isolates used were initially collected in La Laguna, Tenerife, Spain. For insect rearing a dome type crystal insect cabinet containing entomological cages covered with anti-trip screens was used. Trialeurodes vaporariorum was reared on tobacco plants (Nicotina tabacum L.) while Bemisia tabaci was reared on tobacco and aubergine plants (Solanum melongena L.). The whitefly immature stages were obtained after artificial infestation of tomato plants (Lycopersicum sculentum var. Boludo ) grown in a climatic chamber inside two controlled cabinets (one for each specie), on 12-cm diameter pots containing a mixture of sand, volcanic ash, and peat (1:1:1) as a substrate. Fertilization consisted of periodic foliar irrigation with aminoacids and minor elements (Wuxak AA Aventis Crop Science, Spain) at a dose of 0.2 % (v/v). When plants were 30 cm high, five small plastic containers (10mL, with a 120 mesh screen for ventilation) retainig whitefly adults each, were clipped on 5 different leaflets of each plant. Groups of five plants were used per each whitefly species. After 48 hours, the containers were removed from the plants and the leaflets were labeled. The evolution of the biological cycles for each one of the species was daily registered on the selected leaflets. Temperature and relative humidity were recorded. For the bioassays, leaf discs containing 20 individuals of eggs, N1 or N4 larvae were taken and placed into an evolutionary camera with a wet filter paper in a laminar flow hood. Fungal products Fungal material was produced at the Barcelona University (Spain) using a previously 90
103 developed proprietary liquid fermentation method and formulation ingredients (FuturEco S.L.). The technical concentrate consisted of a powder containing 1.05 x 10 9 freeze dried blastospores (as CFU/g) of P. fumosoroseus strain FE The formulation was a claybased wettable powder (WP) containing 0.83 x 10 9 CFU/g. The preparations were stored at 4ºC until usage. Previous to each bioassay, viability of blastospores was verified by the Spread Plate Technique using Potato Dextrose Agar (25 ±1ºC, 72 h). The concentration of blastospores in the technical concentrate, formulation and water suspensions was measured in a standard hemacytometer. Bioassays Each bioassay consisted of three different treatments: Control (water only), Technical Concentrate and Formulation. Five leaf discs per each stage were sprayed on the underside at a rate of 2.0 x 10 5 blastospores/cm 2 or water only. After removing the excess of water in a laminar flow hood, treated discs were placed in 5.5 cm Petri dishes containing 12 g fine sand wetted with 2.6 ml sterile water. Then, Petri dishes were incubated for 24 h, in the dark at 26 ºC (T max 27 ºC, T min 23 ºC) and 62 % RH (RH max 86%; RH min 39%). After the first 24h observation, the Petri lids were replaced by lids with a central hollow (1cm, covered with a 120 mesh screen) to allow aeration and avoid condensation. Then, dishes were returned to the incubation chamber and the evolution of individuals was observed, recording daily the number of alive and died individuals, the symptoms of fungal infection and the general aspect. Mortality of the treated individuals was considered as the percentage of dead organisms out total organisms in a single disc. Effects of fungus were observed in a short term, at the stage where fungus was applied, and in a long term, at the further life stage. The specific observations consisted of: 1) Eggs: direct mortality and mortality of the N 1 nymphs emerged from eggs. 2) N 1 nymphal stage: direct mortality on the N 1 nymphs treated and effects on the subsequent transformed N 2 nymphs. 3) N 4 nymphal stage: direct mortality and effects on the adults emerged. Statistical analysis Mortality data in all stages were analysed using software SPSS for Windows. The results from the 5 replicates (20 individuals each) were transformed through the formula arcsine x, and Kolmogorov-Sminorv test was used to verify the normal distribution of data. Those values adjusted to normal distribution were compared through an Analysis of Variance (ANOVA, P 0.05) and Tukey test (P 0.05) was applied when there were significant differences among them. Results and Discussion Effects on eggs Short term effects (Direct Mortality). In the bioassay of B. tabaci, the fungal infection over eggs caused by the Formulation was higher than that of the Technical Concentrate (60% vs 40% mortality) (Figure 1a). In the T. vaporariorum bioassays there were not statistically significant differences (70-80% eggs mortality), although the infection process seems to be quicker with the technical product (Figure 1b). The direct effect produced by P. fumosoroseus strain FE 9901 on eggs seems to be higher on T. vaporariorum than on B. tabaci. Long term effects: survival of N 1 emerged from treated eggs. All the hatched eggs from the control treatment reached N 1 nymph stage by the end of the bioassays. Although 40-60% of the B. tabaci treated eggs on the bioassay 1 were able to hatch, none of the N 1 nymphs emerged reached the N 2 nymphs stage (Figure 2a). In the case of T. vaporariorum, where 20-91
104 30% of the eggs were able to develop into N 1 nymphs, only 20% of the N 1 nymphs were able to develop into N 2 nymphs (Figure 2b). a % Mortality Technical Product a a b a Formulation Days after application b a b a b a b % Mortality Technical Product Formulation a a a a Days after application b a a Figure 1. Percentage of mortality on eggs treated with a technical or a formulated product based on P. fumosoroseus strain FE 9901: a) B.tabaci; b) T. vaporariorum. Mean mortalities in a sampling date not followed by the same letter are significantly different (ANOVA, P 0.05), by Tukey test, P % Living nymphs a Control TP FM % Living nymphs b Control TP FM Days after hatching Days after hatching Figure 2. Long term effects on eggs treated with a bioinsecticide based on P. fumosoroseus strain FE 9901., a) Survival of N 1 B. tabaci nymphs hatched from treated eggs, b) Survival of N 1 T. vaporariorum nymphs hatched from treated eggs. Effects on N 1 nymphs Short term effects (Direct Mortality). The effects of the fungus on the N 1 nymphs were similar in both species (Figures 3a and 3b). Symptoms of positive infection appeared since the second day and consisted of dehydration and presence of mycelium. All the treated N 1 nymphs died before the fourth day. There were not statistically significant differences in the mortality caused by the formulation or the technical product in both bioassays. The mortality in control treatment was less than 2% in B. tabaci and 0 % in T. vaporariorum. 92
105 a % Mortality Control Technical Product Formulation a a a a Days after treatment a b a b b a b a a % Mortality b Technical Product a a Formulation a a a a Days after treatment Figure 3. Percentage of mortality on N 1 nymphs treated with a technical or a formulated product based on P.fumosoroseus strain FE 9901: a) B. tabaci; b) T. vaporariorum. There were not statistically significant differences in the mortality caused by the techynical or formulated product in both bioassays. a % Mortality Technical Product a a a Formulation Days after treatment aa b % Mortality Technical Product a b Formulation a b Days after treatment a b Figure 4. Percentage of mortality on N 4 nymphs treated with a technical or a formulated product based on P. fumosoroseus strain FE 9901: a) B. tabaci; b) T. vaporariorum. Mean mortalities in a sampling date not followed by the same letter are significantly different (ANOVA, P 0.05), by Tukey test, P Effects on N 4 nymphs Direct Mortality. The N 4 nymphs of both species treated with P. fumosoroseus, revealed the presence of fungus from the first day. By the third day, the fungus had completely colonised the body of the infected nymphs. The N 4 nymphs that could not reach the pupae stage showed dehydration symptoms, and those transformed into pupae had brownish colour and vague red coloured eyes. The mortality on N 4 nymphs at the end (3-5 days) of both bioassays was 100%, and not significant differences were noted between the technical and the formulated product (Figures 4a and 4b). To conclude, it can be emphasize that the efficacy of P. fumosoroseus strain FE 9901 to produce mortality on the different stages of both species of whitefly, was essentially the same 93
106 when applied as either a Technical Concentrate (freeze dried blastospores), or as a Commercial Formulation (freeze dried blastospores plus adjuvants). The selected adjuvants neither interfere with the mode of action of the fungus, nor have insecticide properties by themselves. Furthermore, the absence of significant differences in the time for producing infections between the technical and formulated product may suggest that the concentration of the active ingredient (blastospores) in the formulation could be reduced, allowing a major yield from the manufacturing process. References Lacey L. A. & Fransen J.J., 1995: Global distribution of naturally ocurring fungi of Bemisia, their biologies and use as biological control agents. In: Gerling, D. & R.T. Mayer (eds.). Bemisia: 1995 Taxonomy, biology, damage, control and management pp. Intercept Ltd, UK. Hernández Suárez, E. 1999:. La familia Aleyrodidae y sus enemigos naturales en las Islas Canarias. Tesis doctoral. Departamento de Biología Vegetal, Universidad de La Laguna. Hernández Suárez, E. & Carnero Hernández A. 2000: Enemigos naturales de las moscas blancas (Hemiptera: Aleyrodidae) de importancia económica en Canarias (I): depredadores. La Granja 7: Espino, A. I.; Montero Gómez N.; Hernández-Suárez E.; Carnero Hernández A.; Rodríguez Rodríguez J. M.; Martín Suárez R.; Galván Sintes F. & Estévez Gil J. R. 2003: Situación actual de la población de las especies de Geminivirus asociadas a la enfermedad del rizado amarillo del tomate (TYLCD) en cultivos de tomate de exportación en Tenerife y Gran Canaria. La Granja, 10: Wisler, G.C.; Duffus J.E. & Liu H.Y.1999: Expansion of tomato-infecting criniviruses into new areas. VII th International Plant Virus Epidemiology Symposium. Aguadulce (Almería), España de Abril. EWSN Newsletter. Canary Island Worshop. Noviembre,
107 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Evaluation of the effect of two potassium based products on Bemisia tabaci in greenhouse tomatoes L. Schoen Sica CENTREX, Torreilles, France Abstract: Since the introduction of Bemisia tabaci Q type combined with several transmitted viruses in France the need of corrective compounds to manage the populations of this insect is very important. Several trials have been conducted with a multitude of pesticides and two Kalium-based products have been chosen for further investigations. The advantage of these products is their low toxicity for mammals and their possible integration in IPM programs under certain conditions. The results of the presented trials showed an interesting possible use of these products against B. tabaci and an important side-effect on Oidium neolycopersici. Keywords: tomato, Bemisia tabaci, insecticidal soap, IPM, Oidium neolycopersici. Introduction Since the introduction in 2001 of Bemisia tabaci associated with several viruses (Trottin Caudal et al. 2005, Trottin-Caudal et al. 2002) in the Perpignan region (South of France), the need for practical solutions to manage whitefly populations, especially at the end of the culture cycle under greenhouses, is very important. On demand of local vegetable producers the experimental station CENTREX undertook a screening of different commercial products (Schoen, 2003) for their efficacy against this insect. Some of these products were retained like «potentially interesting». The most important criteria were their efficacy against adult whiteflies, their low mammalian toxicity and a low effect on beneficials for the next culture cycle. Some products based on Kalium derivates (soaps) give interesting results (Schoen, 2005, Schoen & Decoin, 2005). Liquid soaps have been used for a long time against aphids in home gardens with various results. In some countries these so-called «insecticidal soaps» are proposed for professional use (Butler et al. 1993, Gills & Raup, 1990, Puri et al. 1994, Tong-Xian & Stansly 2000, Smith & Chancy 2001). Insecticidal soaps are composed of Kalium (Potassium) and fatty acids. The mechanism causing the death of insects is not well-known, but it seems that fatty acids influence the permeability of the cell membranes, which starts leaking. Another effect is the partly destruction of the waxy layer of insect cuticles, which makes the insect more sensitive to entomopathogens and insecticides. These soaps are efficient against soft insects such as aphids, whiteflies, soft scales etc., but also against mites. They seem to have little impact on several beneficials like coccinellids and hymenopteran parasitoids. Once the product is dry, the impact on pest and beneficials is very low. Potassium based products are totally biodegradable and do not persist in the environment. Because of their mechanical mode of action, development of resistance seems to be improbable. Since 2003 different commercially available products have been tested with more or less the same results. For the trials proposed in this paper we had chosen «Bioshower» of Biobest N.V. Recently, another product based on Potassium Phosphate (Agri50, distributed in France by Desangosse) is proposed with interesting effects on whiteflies (Hanafi et al. 2002) and had been added to our 95
108 tests. Another important effect has been observed on different pathogenic fungi, especially on Oidium neolycopersici.the practical use of these compounds is not without some constraints. The use of «hard» water for the tank mix causes important deposit of calcium/magnesium ions which causes problems in the spray machines. Also, they can be phytotoxic under certain conditions (Cranshaw, 2004). Material and methods In an insect proof greenhouse at the CENTREX, tomato plants (cv. Cheers) were planted on 21/02/2005 (trial I) and 21/06/2005 (trial II). The same day an artificial infestation had been carried out with 5 (trial 1) or 8 (trial11) B. tabaci adults per plant. The tested compounds had been sprayed with a «Pulvexper» knapsack sprayer with a constant pressure of 3 bars, at 800 l/ha at: 07/04/2005, 11/04/2005 and 19/04/05 (Trial I: 18 plants/plot): 13/07/2005, 18/07/2005 and 24/07/2005 (Trial II: 16 plants/plot). Treatments were arranged in a randomized block design with 3 replicates, and are detailed in Table 1. Table 1. Treatments applied. Trial I Trial II Commercial name Dose Active ingredient potassic soap (code SP) Bioshower 20l/ha potassium phosphate (code SPA) Agri 50 10l/ha potassic soap (SP) Bioshower 10l/ha Potassium phosphate (Agri50/6) Agri 50 6l/ha Potassium phosphate (Agri50/3) Agri 50 3l/ha The number of B. tabaci adults on 4 middle leaves in 10 plants/plot was scored. Counting dates: Trial I: 06/04/05, 11/04/05, 12/04/05 and 19/04/05. Trial II: 23/06, 27/06, 05/07, 11/07, 12/07, 18/07, 19/07, 20/07, 25/07, 26/07, 27/07.The number of larvae (all instars): (Only Trial I: 11/04/05 and 25/04/05). Evaluation of the effect on O. neolycopersici. (0= 0%, 1= 1 to 20%, 2= 21 to 50%, 3=51 to 75%, 4=100% of leaves with at least one powdery mildew spot). Results and discussion Trial I Effects on B. tabaci One day after the first treatment (11/04/2005) both compounds showed a reduction in the number of adults compared to the non-treated plots (Figures 1 and 2). A day after the second treatment the counting shows a clear «knockdown» effect with a spectacular effect on adults. The difference between compounds is not statistically significant (Newman-Keuls, 5%). Observations after the third treatment confirms this effect: one day after treatment the population fell down considerably. The last counting (25/04/2005) showed an important reduction of adults on the untreated plots, probably caused by a «generation effect». Mortality of larval stages is shown in Figure. 3. The compound SPA showed an important effect, compared to the compound SP. This confirmed former observations. 96
109 12,00 10,00 8,00 6,00 4,00 2,00 0,00 6/04/05 7/04/05 8/04/05 9/04/05 10/04/05 11/04/05 12/04/05 13/04/05 14/04/05 15/04/05 16/04/05 17/04/05 18/04/05 19/04/05 SPA SP tˇmoin Figure. 1. Number of B. tabaci adults per leaf (average of 3 replicates). 20/04/05 21/04/05 22/04/05 23/04/05 24/04/05 25/04/05 100,00 90,00 80,00 70,00 60,00 50,00 40,00 30,00 20,00 10,00 0,00 SPA 10-avr 11-avr 17/04/ /04/ /04/2001 SP Figure. 2. Percentage efficacy on adults of B. tabaci (average of 3 replicates) 97
110 SPA SP untreated 11/04/ /04/2005 Figure 3. Percentage of larval mortality of B. tabaci (average of 3 replicates) Effect on Oidium neolycopersici A very interesting secondary effect of both compounds had been observed on O. neolycopersici (Figure 4). Both compounds reduced very significantly the number of O. neolycopersici in plots. The evaluation counted by «notes» shows an efficacy of about 95% for SPA and 60% for SP. The use of SPA caused a certain number of phytotoxical symptoms. Necrotic spots were observed on leaves and, which is more important for commercial purposes, on fruits. The reasons of this phytotoxicity can have different origins: the number of treatments, the applied doses, the tank mix concentration, and/or the application conditions (especially whether conditions). Trial II has been designed for testing lower application rates of both compounds. 3,00 2,50 2,00 1,50 1,00 0,50 0,00 SPA SP untreated 01/05/ /05/2001 Figure 4. Effect on O. neolycopersici, (average of 3 replicates, notes from 0 to 4) 98
111 Trial II Effects on B. tabaci 9,00 8,00 7,00 6,00 5,00 4,00 3,00 2,00 1,00 0,00 22/06/ /06/ /06/ /06/ /06/ /07/ /07/ /07/ /07/ /07/ /07/ /07/ /07/ /07/ /07/ /07/ /07/ /07/2001 SP Agri50/6 Agri50/3 untreated Figure 5. Number of B. tabaci adults per leaf (average of 3 replicates) 100,00 90,00 80,00 70,00 60,00 50,00 40,00 30,00 20,00 10,00 0,00 SP Agri50/6 Agri50/3 12/07/05 18/07/05 19/07/05 20/07/05 25/07/05 Figure 6. Percentage efficacy on adults of B. tabaci (average of 3 replicates) In the presented trials, «Agri50» at different application rates show an important efficacy on B. tabaci on tomatoes. The reduction of the adult population and also on the larvae is very important. Bioshower confirms the results of former tests with an important effect on the adults, but has less effect on larvae. No significant differences (Newman-Keuls, 5%) had been found between the different application rates tested. The lowest tested rates will be chosen for further trials. The effect on O. neolycopersici of both compounds seemed very interesting but has to be confirmed in later tests. 99
112 References Butler, G. D., T. J. Henneberry, P. A. Stansly, & D. J. Schuster. 1993: Insecticidal effects of selected soaps, oils, and detergents on the sweetpotato whitefly: (Homoptera: Aleyrodidae). Florida Entomol. 76(1): Cranshaw W.S., 2004: Insect Control : Soaps and detergents. Fact sheet Colorado State University Cooperative Extension. Gills, S. & Raup, M. 1990: Use of Insecticidal Soap and Neem for the control of Azalea Lacebug. University of Maryland Cooperative Extension Service/ Mycogen Corporation 1991 Technical fact sheets. Hanafi A., Bouharroud R. & Murphy B. 2002: Evaluating a new non-toxic pesticide forintegrated control of Bemisia tabaci in protected agriculture in Morocco, IOBC/WPRS Bull. 25 (1): Puri, S. N., B. B. Bhosle, M. Ilyas, G. D. Butler, & T. J. Henneberry 1994: Detergents and plant-derived oils for control of the sweet potato whitefly on cotton. Crop Prot. 13(1): Schoen, L., 2005: Le savon potassique, une alternative dans la lutte contre les aleurodes sur culture sous serre? Rencontres Techniques phytosanitaires Légumes et Fraises INH Angers. Schoen, L. & Decoin, M. 2005: Légumes sous serre, des alliés inattendus. Un savon potassique face aux aleurodes. Phytoma-Défense des Végétaux N. 579: Schoen, L : Evaluation de l'efficacité biologique de deux bio-insecticides sur Bemisia tabaci. International Symposium on Greenhouse Tomato, Integrated Crop Protection and Organic Production. Ctifl/OILB Avignon. Smith R. & Chancy B. 2001: Guidelines for Insecticidal Soap. Fact sheet University of California Cooperative Extension, Farm Advisors, Monterey County. Tong-Xian, L., & P. A. Stansly. 2000: Insecticidal activity of surfactants and oils against silverleaf whitefly (Bemisia argentifolii) nymphs (Homoptera: Aleyrodidae) on collards and tomato. Pest Manag. Sci. 56(10): Trottin-Caudal Y., Chabrière C. & Schoen L., 2005: Présence de Bemisia tabaci Gennadius dans le Sud-Est de la France en cultures légumières sous abri : études réalisées et situation actuelle de la protection intégrée. In 7 e Conférence Internationale sur les Ravageurs en Agriculture, Montpellier, oct Trottin-Caudal Y., Trouvé C., Pacheco C., Monnet Y. & Schoen L., 2002 : Bemisia tabaci et Trialeurodes vaporariorum - Eléments du diagnostic, Infos-Ctifl, 184:
113 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 p. 101 Imidacloprid as a soil application against whitefly Bemisia tabaci in greenhouse cucumber V. Baniameri, A. Sheikhi Plant Pests and Diseases Research Institute (PPDRI), P. O. Box: 19394/1454, Tehran, Iran. Abstract: Bemisia tabaci is a serious pest of cucumbers in all greenhouses in Iran. The effect on B. tabaci of three different concentrations of Imidacloprid (Confidor 350 SC) (0.25, 0.50, and 0.75 ml/l) and a control treatment was tested in soil application. A Completely Randomized Block Design with 4 replications was applied. The percent mortality of larvae and decrease of eggs were analyzed after correcting by the Henderson-Tilton formula. After 7 days, the mean effect of 0.25ml/l of Imidacloprid on eggs and larvae were 58.87±6.42 % and 43.81±6.9 % mortality respectively. For the dose 0.75 ml/l were 67.03±5.83 % and 50.56±5.5 % mortality respectively. The mean decrease of egg numbers in leaf in concentration 0.75 ml/l was 98.09±0.48 % in 30 days. The comparison of means showed that there is no significant difference among treatments, but there is a significant difference with the control. According to our results, a dosage of 0.25 ml/l of imidocloprid was recommended in soil application, only at planting time and gets the final results 30 days after application. Key words: Bemisia tabaci, imidacloprid, soil application, greenhouse cucumbers, Iran. 101
114 102
115 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 p. 103 Laboratory screening of pesticides against Lecanoideus floccissimus Estrella Hernández-Suárez, Carina Ramos-Cordero, Aurelio Carnero Entomology Department, Instituto Canario de Investigaciones Agrarias (ICIA). Ctra del Boquerón s/n, La Laguna, Tenerife-(Canary Islands), Spain Abstract: Lecanoideus floccissimus Martin et al. is a major pest in commercial banana greenhouses of the Canary Islands. Infestations on banana can be extremely heavy all year around, causing enormous damage to the plants and fruits. The most representative damage of this whitefly species is the development of black sooty mould fungi on plants due to the copious white flocculent wax secreted by nymphs, as well as the abundant excretion of whitefly honeydew. Although, chemical control against L. floccissimus is not practicable for long term and stable management of this pest, information on effective pesticides is important for commercial banana growers before a Classical Biological Control Programme can be implemented. We present the effects of eighteen foliar applied pesticides on L. floccissimus eggs and nymphal stages. Detached leaf-disc bioassays were conducted under controlled conditions (25±2ºC, 60±5% R.H., 16L:8D) in the laboratory. In the study we evaluated only active ingredients authorized for Integrated and Organic banana production in the Canary Islands, i.e.: insecticidal soaps, horticultural mineral and vegetable oils, neem products containing azadirachtin, malathion and chlorpyrifos. Results indicated that there was significant difference (P<0.05) in the effectiveness of the different products evaluated but dependent on the whitefly instar. L. floccissimus eggs were much less affected by any of the products used than nymphal stages. Among the eighteen products evaluated, malathion 50% p/v and chlorpyrifos 48% gave the best egg control. However insecticidal soaps (made of the potassium salts of fatty acids) were more effective for nymphs and 100% mortality was observed with treatments of first nymphal instar. The highest mortalities of fourth nymphal instar were recorded with vegetable oil, insecticidal soaps and Pyrethrum extract treatments. Key words: Spiralling whitefly, Lecanoideus floccissimus, pesticides efficacy. 103
116 104
117 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 p. 105 Does the PGPR Bacillus subtilis induce plant resistance to whiteflies and Phythium spp. in greenhouse tomato? A. Hanafi, K. Fellah Integrated Production and Protection Unit, Department of Plant Protection, Complexe Horticole IAV Hassan II, BP: Cité Balnéaire Agadir, Morocco. Abstract: The development of biological products based on beneficial micro-organisms can extend the rage of options for maintaining the health yield of crops. It has been recognized that the application of the rhizobatcterium Bacillus subtilis (BS) confers vitality as well as the capacity of the plant to face the conditions of biotic and abiotic stress such as drought and salinity. These stress conditions prevail in the Mediterranean regions, where a proportion of farmers use saline water for the irrigation of their crops. In our experiments, we evaluated the impact of BS, used in salinity stress and non stress conditions, on the resistance and tolerance of soilless tomatoes grown under greenhouse, to the whitefly Bemisia tabaci and the root fungal pathogen Pythium Sp. Two levels of salinity (EC = 6dS/m and EC= 2.4 ds / m) were used for irrigation of tomato plants inoculated and non inoculated with BS. Two cultivars of tomato were evaluated in this experiment. On one hand, the cultivar Durinta (Western seeds) is relatively tolerant to salinity, whereas the cultivar Tyjico (Syngenta Seeds) is sensitive to salinity. On the other hand, the cultivar Durinta is sensitive to tomato Yellow leaf curl virus (TYLCV). In contrast, the cultivar Tyjico is tolerant to TYLCV. TYLCV is a Gemini virus exclusively transmitted by the whitefly B. tabaci. TYLCV is by far the number one virus disease of greenhouse tomatoes in the Mediterranean region. The objective of this study was to evaluate the of BS in stress (E C 6) and non stress (E C2.4) conditions on the whitefly B. tabaci population dynamics in a salinity tolerant cultivar (Durinta) and a salinity sensitive cultivar (Tyjico). This study concluded that irrigation of tomato with saline water (E C 6) makes the plant less sensitive to the whitefly B. tabaci. On the other hand, the inoculation of plants with BS appears to induce plant resistance to B. tabaci, whether in salinity or non salinity stress conditions. Regarding the incidence of Pythium, the inoculation of plants with BS induced resistance to this root pathogen, especially under saline conditions. Key words: tomato, soilless, salinity, resistance, tolerance, whitefly, Bemisia tabaci, Bacillus subtilis, Pythium. 105
118 106
119 Mites
120 108
121 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Crop loss assessment of Pediculaster fletchmani (Acari: Pygmephoridae) on button mushrooms Katayun Kheradmand 1, Karim Kamali 1, Yaghoub Fathipour 1, Ebrahim M. Goltapeh 2 Ali R. Nemati 1 1 Department of Entomology, Faculty of Agriculture, Tarbiat Modarres University, P. O. Box , Tehran. 2 Department of Plant Pathology, Faculty of Agriculture, Tarbiat Modarres University, P.O.Box , Tehran, Iran. Abstract: Certain fungivorus pygmephoride mites (red pepper mites, pygmy mites) periodically swarm in commercial mushroom houses, often carpeting the surface of the compost. Yield loss on button mushroom caused by Pediculaster fletchmani was studied at different mite densities (0, 10, 50, 100, 150 and 200 mites per kg compost). A significant difference between the control and the higher mite density was obtained. Regression analysis showed a significant negative relationship between mite density and mushroom yield (P<0.05), with yield decreasing when mite density increased. The slope of the line was , so the yield loss per mite (LPM) was g/kg compost. The fact that these mites are becoming increasingly widespread, together with the losses caused by them throughout the mushroom-growing region, mean that they are considered of major threat to mushroom yield. Information provided in this study may also be used to develop alternative action levels in IPM programmes for P. fletchmani in cultivating button mushrooms in Iran. To provide a sustainable alternative to pest control, a variety of control methods must be integrated into pest control programs. Keywords: Pediculaster fletchmani, Mushroom, crop loss assessment Introduction Yield-loss relations are a key component of Integrated Pest Management (IPM) systems, and although it is widely appreciated that yield loss may depend on various aspects of the cropping system (Poston et al., 1983; Gutierrez, 1987), this level of complexity is often disregarded in practice. Nominal thresholds, which assume that economically significant yield loss will occur when pests exceed a certain fixed density, remain in widespread use, while simple or calculated thresholds, although more flexible, assume that yield loss is primarily a function of pest density (Andow & Hidaka, 1998). A crop loss model is a mathematical method used to describe the relationship between yield reduction and the intensity of harmful organisms (Chiarappa, 1981; Hughes, 1996). Such quantitative relationships between yield reduction and the intensity of harmful organisms (often referred to as damage curves) are an important component of crop protection strategies. Insect and mite pests of cultivated mushrooms can severely limit yields unless controlled (Rinker et al., 1984; Hussey, 1981; Clift & Toffolon, 1981). The main pests include Lycoriella auripila (Winnerz), Lycoriella mali (Fitch), Megaselia haltereta (Wood), Brennandania lambi (Kcrzal) and the Pediculaster spp. The cosmopolitan distribution of some fungus and dung inhabiting species of the genus Pediculaster could well have occurred in conjunction with the expansion of synantropic habitats (Camerik & Coetzee, 1997). Certain fungivorus pygmephoride mites (red pepper mites, pygmy mites) periodically swarm in commercial mushroom houses, sometimes carpeting the surface of the compost (Gurney & Hussey, 1967; Clift & Toffolon, 1981). Pygmephorid mites have previously been considered 109
122 of doubtful pest status (Clift & Toffolon, 1981). Some species e.g. Microdispus lambi (Krczol) feed directly upon the mycelium of crop mushrooms and commonly cause % yield loss, and occasionally even total crop loss on some farms in China (Gao & Zou, 2001; Zou et al, 1993) and up to 30% yield loss in Australia (Clift & Toffolon, 1981; Ferragut et al, 1988). Pediculaster fletchmani Wich is a cosmopolitan nuisance pest in the cultivation of mushrooms Agaricus bisporus (Lange). The objective of this study was to quantify the yield loss on button mushroom attributable to P. fletchmani over several crop harvests. Materials and methods Mushroom culture Commercially prepared compost was used to study the relationship between button mushroom yield and various densities of P. fletchmani. Inoculated compost was maintained at C until the mushroom mycelium had colonized the compost (12-14 days). The casing layer, composed of neutralized peat moss, was then applied. Button mushrooms were cultivated on the compost in 18.5cm high and 14cm diameter containers. Infestation Procedure Pygmy mites were originally collected from mushroom compost at Mallard, near Tehran. They were inoculated into Trichoderma mycelium in glass Petri dishes (9cm) and cultured at 22 C, with a relative humidity of 70±5% and photoperiod of 16L:8D hours. P. fletchmani infestations were established by releasing 0 (control), 10, 20, 50, 100 and 200 adult mites per kg compost when the mycelium had spawn run. Pygmy mites were introduced in each container after the casing stage. Precautions were taken to ensure that the compost used for crop loss assessment experiments was free of P. fletchmani. The containers were put in a growth chamber at 17 C, with a relative humidity of 85 % and a photoperiod of 0L: 24D hours. This procedure was replicated five times for each treatment. The zero infestation level provided information about maximum yield under our specific experimental conditions. Differences between the yield of the control and of the different treatments reflected the levels of yield loss. Sampling was carried out on the emergence of the first flash of button mushroom and undertaken following a daily program. The mushrooms produced in each replication were harvested and weighed when the diameter of their caps was about 6cm. This procedure continued for 2 months after mite release. Mushroom yields were expressed as g/kg compost. Data analysis There are many different methods and techniques for assessing crop losses and determining the relationship between levels of pest damage or pest infestation and crop loss. We used the Small-plot technique, which is probably the most commonly used to generate crop loss data (Chiarappa, 1981). In this technique, small replicated plots, situated side by side, are exposed to different levels of pest density. In some cases, investigators artificially infest plots with laboratory-reared insect pests or disease inoculums, or manipulate natural infestations. In order to determine loss per mite (LPM), we used the linear regression equation between various densities of P. fletchmani and yield of button mushroom as follows: y = a + bx in which y is yield, x is density of mites, a is the intercept and b is the slope of the regression line that indicates loss per mite (LPM). Button mushroom yields between the control treatment and the different densities of P. fletchmani were compared by applying a two sample t-test. 110
123 Results and Discussion The effect of various densities of P. fletchmani on button mushroom yields is shown in Figure 1. Significant differences were only observed between 200 mites per kg density and the control treatment (Figure 1). The intercept value of the regression line was which provided potential yields at each phenological crop stage in the case of zero damage by P. fletchmani. In this way, the highest predicted yield was g/kg compost (Figure 2). Regression analysis showed a significant negative relationship between mite density and mushroom yield (P value <0.05), with yield decreasing as mite density increased. The slope of the line was , so the yield loss per mite (LPM) was g/kg compost. We also found that P. fletchmani carried Trichoderma spores (a mushroom parasite) that Terras & Hales (1995) had also reported. The average loss in mushroom production due to Trichoderma infection was 8.5 kg/m 2, as also reported by Clift & Terras (1991). To the best of our knowledge, there have been no previous experimental studies to assess quantitative crop loss associated with P. fletchmani. As expected, the results clearly indicated that P. fletchmani significantly affected mushroom yield. In New South Wales, Clift (1978) and Clift & Terras (1991, 1994), reported crop loss data for Siteroptes mesembrinae (Canestrini), Histiostoma feroniarum (Dufour) and phorid flies. The values that they reported were considerably greater than those found in this study. For example, they reported that the yield loss caused by S. mesembrinae was 200 kg/tonne and 198 kg/tonne for H. feroniarum, which was about 20 kg/tonne more than ours. These differences might have been due to different experimental conditions. Bhattacharyya et al (1993) examined crop loss caused by several major mushroom pests. Their results indicated that various mushroom pests such as Lycoriella mali (Fitch) (Diptera: Sciaridae), Megaselia halterata (Wood) (Diptera: Phoridae), S. mesemberinae (Acari: Pygmephoridae) and other minor pests caused button mushroom yields to fall by as much as 49%. These findings agreed with those of White, 1986, who also reported the extent of the damage to the mushroom crop as ranging from 15 to 75%, depending on the sciarid species involved. Jess & Klipatrick (2000) studied an integrated approach to the control of L. mali. The potential for Hypoaspis miles (Berlese) and Steinernema feltiae (Poinar) to reduce L. mali infestations by 82% at different stages of mushroom production has been clearly demonstrated. Chemical control of mushroom pests has been successfully achieved (Al-Amidi et al, 1991), but in recent years it has been shown that some species have developed resistance to organophosphorus insecticides (Fletcher et al, 1989). Some of these chemicals can be phytotoxic, causing yield losses of up to 30% (Hussey & Wyatt, 1959). These factors, coupled with the general desirability of reducing the use of chemical pesticides in food crops, have increased interest in the biological control of mushroom pests. To provide a sustainable alternative strategy to conventional pest control, a variety of new control methods must be integrated into pest control programmes and it is essential that the effects of these control systems on specific biological control agents are evaluated. 111
124 Yield (g/kg compost) a a a a a b Mite density Figure 1. Comparison of button mushroom yields at different infestation densities (mites/kg compost) of Pediculaster flechmani and control treatment. Different letters indicate significant (P < 0.05) differences between control (0) and other mite densities (t-test) y = x Yield (g/kg compost) Mite density Figure 2. Relationship between button mushroom yield and different densities of Pediculaster flechmani. References Al-Amidi, A.H.K., Dunne R. & Downes, M.J. 1991: Parasitus bitubersus (Acari: Parasitidae): An agent for control of Lycoriella solani (Diptera: Sciaridae) in mushroom crops. Exp. Appl. Acarol. 11:
125 Andow, D.A. & Hidaka, K. 1998: Yield loss in conventional and natural rice farming systems. Agric. Ecosyst. Environ. 70: Bhattacharyya, P., Adhikary, P.K. & Bordoli, D.N. 1993: Damage potential and biology of mushroom-infesting sciarid. J. Food Sci. Technol. Mysore, 30: Camerik, A.M. & Coetzee S.H. 1997: The phoretic female of Pediculaster austrelis spec. nov. (Acari: Pymephoridae) from South Africa and new synonyms for P. morelliae Rack, 1975, Entomologie, 67: Chiarappa, L. 1981: Crop Loss Assessments Methods. Commonwealth Agricultural Bureaux, Slough, UK. 123pp. Clift, A.D. 1978: The identity, economic importance and control of insect pests mushrooms in New South Wales, Australia, Mushroom Science X (2): Clift, A.D. & Terras, M.A. 1991: Effects of pesticides on the yield and production pattern of three standard and three hybrid strains of cultivated mushrooms in NSW. Australian J Exp. Agric. 31: Clift, A.D. & Terras, M.A. 1994: The consequences of mite's surviving the composting process, Proceeding of 2 nd AMGA/ ISMS INt. Workshop. Clift, A.D. & Toffolon, R.B., 1981: Distribution of larvae of Lycoriella agarici Louden (Diptera: Sciaridae) within mushroom beds in commercial culture of Agaricus bisporus and Agaricus bitorquis in N. S. W. J. Austr. Entomol. Soc. 20: Ferragut, F., Gea, F.J. & Garcia-Morras, J.A. 1997: The mushroom mite Brennandania lambi (Acari: Pygmephoridae): introduction in Spain, economic importance and distinction of related species, Bol. San. Veg. Plagas 23(2): Fletcher, J.T.; White P.F. & Gaze R.H. 1989: Mushroom: Pest and Disease Control, 2nd edition, 174 pp. Gao, J.R. & Zou, P. 2001: Biology, life table and host specificity of the mushroom pest, Brennandania lambi (Acari: Pygmephoridae), Exp. Appl. Acarol. 25: Gurney, B. & Hussey, N.W Pygmephorus species (Acarina: Pyemotidae) associated with cultivated mushrooms, Acarologia, 9(2): Gutierrez, A.P. 1987: System analysis in integrated pest management. In: Delucchi, V. (Ed.), Integrated Pest Management, an International Perspective. Parasitis 86, Geneva, pp Hughes, G. 1996: Incorporating spatial pattern of harmful organisms into crop loss models. Crop Prot. 15 (5): Hussey, N.W. 1981: Cultural innovation: its implication for mushroom pest control. Mushroom Sci. 11(1): Hussey, N.W. & Wyatt, I.J. 1959: Cecid control by incorporation of insecticides in composts. Mushroom Sci. 4: Jess, S. & Kilpatrick, M. 2000: An integrated approach to the control of Lycoriella solani during production of the cultivated mushroom (Agaricus bisporus). Pest Management Sci. 56: Poston, F.L., Pedigo, L.P. & Welch, S.M. 1983: Economic injury levels: reality and practicality. Bull. Entomol. Soc. Am. 29: Rinker, D.L., Finely, R.J., Wuest, P.J., Snetsinger, R.J. & Royes, D.J., 1984: The influence of phorid and sciarid populations on mushroom yield at commercial mushroom farms in Pennsylvania. Proc. Penn. Academy of Science, 58: Terras, M.A. & Hales D.F. 1995: Red pepper mites are vector of Trichoderma, Science and cultivation of edible fungi, Volume 2: Proceedings of the 14 th international Congress, Oxford, UK, September,
126 White, P.F. 1986: The effect of sciarid larvae (Lycoriella auripilla) on cropping of the cultivated mushroom (Agaricus bisporus). Ann. Appl. Biol. 109: Zou, P., Gao, J.R. & Ma, E.P. 1993: Preliminary studies on the biology of the pest mite Luciaphorus auriculariae (Acari: Pygmephoridae) infesting Jew's ear mushroom Auricularia polytricha in China. Exp. Appl. Acarol. 17(3):
127 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Biology of the two spotted spider mite, Tetranychus turkestani (Acari: Tetranychidae) on four common varieties of eggplant in Iran Ebrahim Soleimannejadian 1, Alireza Nemati 2, Parviz Shishehbor 1, Karim Kamali 2, Valiallah Baniameri 3.1 Plant Protection Department, Agricultural College, Shahid Chamran University, Ahwaz. 2 Department of Entomology, College of Agriculture, Tarbiat Modares University, Tehran. 3 Plant pest and Disease Research Institute, Tehran, Iran. Abstract: Spider mites are important pests for various crops throughout the world. Host plants can exert profound effects on biological and ecological characteristics of tetranychid mites. In the southern provinces of Iran the major tetranychid species is Tetranychus turkestani, which is called the strawberry or Atlantic mite. The life history parameters of T. turkestani on four aubergine varieties, Esfahani, Qasri (two local varieties), Blackbeauty and Inerash (imported varieties) were studied on the lower surface of eggplant leaves under laboratory conditions. Survival rates of eggs on Qasri and Inerash cultivars were 100%, while on Esfahani and Blackbeatuy they were 93% and 88% respectively. The larval developmental period of T. turkestani differed on the four cultivars. Inerash had the shortest development time, while those of the other three cultivars were not significantly different. The pre-oviposition period of the mite was significantly different for Esfahani, Qasri, Inerash and Blackbeauty, with the longest pre-oviposition period relating to Inerash. There were major variations in the total number of eggs laid by each female mite. The total numbers of eggs laid by each female mite on the four cultivars during the oviposition period were 1-67 on Qasri, 3-95 on Esfahani, 2-61on Blackbeauty and on Inerash. Given its survival rate and the duration of the development time of its immature stages, the total longevity of adults, and the total number of eggs per female per day, it was concluded that the Qasri cultivar is not as suitable as a host as the other three cultivars in the south of Iran. Key words: Tetranychus turkestani, eggplant, life history parameters Introduction Spider mites are important pests for various crops throughout the world (Jepson et al., 1975; Carey & Bradley, 1982; Leigh, 1985). The biological characteristics of the spider mite are affected by various factors, for example, with host plants often exerting several profound effects on the biology of tetranychid mites (Henderson & Holloway, 1942; Watson, 1964; Wilson, 1994). These effects may be manifested as differences in developmental rate, survival, reproduction and longevity attributable to differences among plant species, varieties or cultivars (Nemati, 2005). In the southern provinces of Iran, the major tetranychid species is Tetranychus turkestani Ugarov & Nikolski, the strawberry or Atlantic mite (Kamali, 1988). So far, no studies have been carried out on the biological and ecological characteristics of the mite on the eggplant in Iran (Nemati, 2005). As an initial step in our investigation into population biology, we examined the effects of four aubergine varieties (Blackbeauty, Inehrash, Qasri and Esfahani), acting as host plants, on some biological characteristics of T. turkestani. 115
128 Material and methods The stock culture of T. turkestani used in this study was initiated by mites obtained from the eggplant field of Shahid Chamran University during the summer of All experiments were conducted under laboratory conditions at 30±1 0 C, 55± 10% relative humidity and with a photoperiod of 16:8(L:D). To study the life history parameters of T. turkestani on four aubergine varieties, the lower surface of a leaf disk was maintained on water soaked cotton. A water saturated barrier of cello-cotton was placed along the leaf periphery and down the mid-vein of each leaf. This area was then subdivided into 6 8 narrower small rectangular cells using strips of cellocotton. One female was placed in each of the cells and removed after 8h whether it had laid eggs or not. All but one of the eggs were removed from each cell. Observations were made on a daily basis at 8 a.m., 4p.m. and 8p.m. until each mite had completed its development. Two mature males were confined with each virgin female throughout her pre-oviposition period (Tanigoshi et al., 1975). The number of eggs laid by the mated females was recorded every day (at 24 hour intervals). One way ANOVA was used for statistical analysis. Means were separated by the Duncan Multiple Range Test (α=0.05). Results and discussion Development times for immature stages of T. turkestani are shown in Table 1. Analysis of variance showed that the mean egg development time differed significantly among varieties (F=281.28, df=176, P<0.0001). The shortest period was on Qasri and the longest was on Esfahani. Since there was no nutritional relationship between the eggs and the substrate (host leaf), it is probable that the difference was due to females acquiring nutrition from the host plant. Survival rates for eggs on Qasri and Inerash cultivars were 100%, and on Esfahani and Blackbeauty, they were 93% and 88%, respectively (Table 2). The developmental periods for T. turkestani larvae on the four cultivars were significantly different (F=281.28, df=176, P<0.0001). Inerash had a shorter development time for larvae than the other three cultivars, while no significant differences were observed among Esfahani, Blackbeauty and Qasri (Table 1). Table 1: Mean (±SD) development time (days) for T. turkestani immature stages on four eggplant cultivars under laboratory conditions (30 C, 55%±10 R.H and16: 8L:H). Cultivars Life stage Esfahani Qasri Inerash Blackbeauty Egg Larva First chrysalis Protonymph Second chrysalis Deutonymph Third chrysalis 3.88± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± = ± ± ± ± ± ± ± ±0.06 Development times for the first chrysalis stage on Qasri and Inerash were different, but no significant difference was observed on the other two cultivars. The shortest development 116
129 period for the first chrysalis stage and protonymph was on Inerash, and the shortest for the second chrysalis and deutonymph was on Blackbeauty. There was a significant difference in total development time from egg to third chrysalis on all cultivars. However, Blackbeauty and Quasar were in the same statistical group. Table 2: Percentage survival of T. turkestani eggs and immature stages on four eggplant cultivars under laboratory conditions (30 C, 55%±10 R.H and16: 8L:H). Cultivars Life stage Esfahani Qasri Inerash Black beauty Egg Active immature Total immature Table 3 shows the development time of three periods of the adult stage of T. turkestani on the four cultivars. Total time for development of the adult stage was also calculated. The pre-oviposition period was significantly different (F=5.15, df=137, P=0.0021) on Esfahani, Qasri, Inerash and Blackbeauty and the longest pre-oviposition period was on Inerash. The oviposition period of adults on three of the cultivars was different, but no significant difference was seen between Inerash and Qasri (Table 3). There was also a significant difference in the post-oviposition period of the mite on the four cultivars. Although there was variation in the total development period of the adult stage, no significant differences were seen in the lifespan of adults living on all cultivars. Table 3: Mean (±SD) development time (days) of T. turkestani adults on four eggplant cultivars under laboratory conditions (30 C, 55%±10 R.H and16: 8L:H). Cultivars Adult stage Esfahani Qasri Inerash Blackbeauty Pre-oviposition Oviposition Post-oviposition Total adult stage 0.77±0.35b 5.88±1.93b 0.97±0.38b 7.03±2.19a 0.86±0.30b 4.41±2.34b 0.24±0.24c 5.52±2.25a 1.12±0.29a 6.47±2.74b 0.84±0.20b 7.85±2.88a 0.67±0.33b 9.30±3.93a 1.45±0.93a 8.88±5.60a There were large variations in the total number of eggs laid by each mite. The total number of eggs laid by each female on four cultivars during the oviposition period was 1-67 on Qasri, 3-95 on Esfahani, 2-61on Blackbeauty and on Inerash. Changes in the total number of eggs per day corresponding to all the female mites used in the experiment and the mean number of eggs per day laid by each female are shown in Figures 1 and 2. Analysis of variance showed a significant difference in total eggs per mite on different cultivars (F=2.92, df=61, P=0.04). The Duncan test indicated that the number of eggs per mite on Qasri was smaller than the number of eggs laid by other mites feeding on the other three cultivars: Esfahani, Blackbeauty and Inerash. 117
130 14 12 Esfahani Inehrash Qasri Blackbeauty Egg/female/day Time(day) Figure 1. Mean number of eggs laid daily by T. turkestani females on four eggplant varieties under laboratory conditions (30 C, 55%±10 R.H and16: 8L:H). Due to the survival rate and duration of the development time of the immature stages, the total longevity of adults, and the total number of eggs per female per day, it was concluded that the Qasri cultivar is not as suitable a host as the other three cultivars. This was confirmed by comparing demographic parameters of T. turkestani reared on different cultivars (Nemati 2005). Cagle (1956) and Andres (1957) (as reported in Carey & Bradley, 1982) were the first researchers to examine the biological and demographical characteristics of T. turkestani at different temperatures. Carey and Bradley (1982) studied the demographic parameters of the Atlantic mite on cotton under different temperature regimes. There were no reports of biological traits of T. turkestani on aubergine varieties. Due to the important influence of the host plant on the biological parameters and differences in experimental procedures, we cannot directly compare our results with those of other researchers. However, such studies could be conducted in different parts of the word under varying conditions in order to find varieties resistant to T. turkestani, and thereby reduce the need for insecticide applications. References Carey, J.R. & Bradley, J.W. 1982: Developmental rates, vital schedules, sex ratios, and life tables for Tetranychus utricle, T. turkestani and T. picnics (Acarina : Tetranychidae) on cotton. Acarologia, 23 (4): Henderson, C.F. & Holloway, J.K. 1942: Influence of leaf age and feeding injury on the citrus red mite. J. Econ. Entomol. 35 (5): Jeppson, L.R., Keifer, H.H. & Buker, E.W. 1975: Mites Injerious to Economic Plants University of California Press. Berkeley, 614 pp. Kamali, K. 1988: Tetranychid mites (Acari: Tetranychidae) injurious to plants in Khuzestan, Iran. Proceeding 18 th International Congress of Entomology. Vancouver, Canada (Abstract). P
131 Leigh, T.F. 1985: Cotton. In: Helle, W. and Sabelis, M. W. (Eds). Spider Mite, Their Biology, Natural Enemies and Control. Vol. 1A. Elsevier, Amsterdam, pp Nemati, A. 2005: Population Dynamics and Life History Parameters of Two-spotted Spider Mite Tetranychus turkestani Ugarov & Nikolski (Acari: Tetranychidae) in Ahwaz. Ph.D. dissertation, Shahid Chamran University, Ahwaz, Iran, 492pp. Tanagoshi, L.K., Hoyt, S.C., Browne, R.W. & Logan, J.A. 1975: Influence of temperature on population increase of Tetranychus Mcdanieli (Acari: Tetranychidae). Ann. Entomol. Soc. Am. 68 (6): Wilson, L.J.1994: Plant quality effect on life history parameters of the two spotted spider mite (Acari: Tetranychidae) on cotton. J. Econ. Entomol. 87 (6): Watson, T.F. 1964: Influence of host plant condition on population increase of Tetranychus telarius (Linnaeus) (Acarina: Tetranychidae). Hilgardia, 35(11):
132 120
133 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Efficacy of Phytoseiulus persimilis versus Neoseiulus californicus for controlling spider mites on greenhouse sweet pepper Phyllis Weintraub 1, Sophia Kleitman 1, Nurit Shapira 2, Yael Argov 3, Eric Palevsky 4 1 Agricultural Research Organization (ARO), Gilat Research Center, DN Negev, 85280; E- mail: phyllisw@volcani.agri.gov.il; 2 Research and Development, Sapir Center, D.N. Arava, 86825; 3 Israel Cohen Institute for Biological Control, Plant Production and Marketing Board, Citrus Division, POB 80 Bet Dagan, 50250, Israel; 4 ARO, Newe Ya ar Research Center, Ramat Yishay, 30095, Israel Abstract: The efficacy of two predatory phytoseiid mites, Phytoseiulus persimilis and Neoseiulus californicus were compared for the biological control of Tetranychus urticae (TSSM) on greenhouse sweet peppers. We developed a simple and expedient rearing method for N. californicus solely on pollen. At the beginning of the trial there was a low-level spider mite infestation. Cohorts of P. persimilis (10/plant) and N. californicus (20/plant) were released in separate tunnels when the TSSM population was low and in additional tunnels 2 weeks thereafter. These treatments were compared with acaricide-treated and non-treated plants. Samples of leaves from the top and middle sections of plants from each tunnel were taken. There was no significant difference between the number of TSSM in the acaricide-treated and 1 st N. californicus-release tunnels, but the number of TSSM in the acaricides-treated and P. persimilis-release tunnels were significantly different. At the higher spider mite population, there were significantly more TSSM in the P. persimilis release tunnels than in the N. californicus-release tunnels. We have confirmed laboratory trials and have shown that N. californicus is a superior TSSM predator at low humidities. Key words: Neoseiulus californicus, Phytoseiulus persimilis, mite rearing Introduction Tetranychus urticae Koch, the two spotted spider mite (TSSM), is an important polyphagous pests in greenhouse, field and orchard crops worldwide. In Europe and Israel, these spider mites are pests of more than 80 plants. Climatic conditions within greenhouses in temperate climates, especially in the Mediterranean Basin, and screen houses in subtropical climates usually allow for continuous development of spider mites all year. The biological control of spider mites with predatory mites on vegetables has been studied. The most widely used and commercially sold phytoseiid is Phytoseiulus persimilis. This is a specialized predator, preying only on mites belonging to the genus Tetranychus, and requiring high humidity and moderate temperature to establish and thrive. While it is an excellent predator on many crops in environmentally controlled greenhouses, in Mediterranean climates where greenhouses or screen houses are under ambient conditions, its efficacy in many cases is unacceptable. Neoseiulus californicus is another phytoseiid, native to the Mediterranean area and similar climates world-wide. Neoseiulus californicus is a type II-III predator (Croft et al., 1998); feeding on a select group of arthropods and is tolerant to low humidity environmental conditions (Rott & Ponsonby, 2003). Small-scale experiments on gerbera demonstrated that N. californicus has a potential to effectively suppress spider mites on greenhouse grown crops comparable to P. persimilis (Schausberger & Walzer, 2001). Furthermore, recent studies 121
134 suggested that N. californicus and P. persimilis may even have synergistic effects in spider mite suppression (Walzer & Schausberger, 2005). In this paper we report on a comparison of the efficacy of N. californicus and P. persimilis against spider mites on sweet peppers. Materials and Methods All trials took place at the Yair Research and Development Farm in the Arava Valley (between the Dead Sea and Red Sea), Israel. Plants were fertilized and watered by a drip irrigation system, according to standard organic agricultural practices for the area. Twentyfour walk-in tunnels (7 x 15 m) were planted with variety of organically grown sweet pepper seedlings (red fruit) in three double row beds on 30 August At the beginning of the trials, there was a uniform low-level spider mite infestation in all tunnels. N. californicus were reared on Vallonea oak (Quercus itaburensis) pollen as the sole food source (Figure 1). A 5 cm diameter hole was cut from the bottom of a clear, disposable plastic food container (270 ml) and replaced with nylon mesh (pore size 37µm) secured with glue. A loose mat (ca. 3 mm) of synthetic fibers was placed in the bottom of the container for egg laying. A standard Petri dish (open side up) was placed on the fiber and sprinkled with pollen. To prevent mites from escaping a moat (fashioned from the lid of the container) was secured to the top of the container and partially filled with castor oil (barrier). Containers were placed on wet cotton. This allowed the mites a source of free water and created high humidity between the bottom of the Petri dish and the bottom of the container. Castor oil Petri dish Pollen Synthetic fibers Screening Wet Cotton Figure 1. Mite rearing container. For release, mites were dusted off the Petri dish and mixed into moistened vermiculite. Approximately 20 N. californicus were released on the top leaves of each plant in 4 tunnels on 11 October and in another 4 tunnels on 31 October. P. persimilis were obtained from Bio-Bee Biological Systems, Kibbutz Sedeh Eliyahu, Israel. Approximately 10 P. persimilis were released on the top leaves of each plant in 4 tunnels on 11 October and in another 4 tunnels on 31 October. One set of 4 tunnels was treated with bigenazate (Floramite, Uniroyal, USA) and one set of 4 tunnels was left untreated. Samples of 20 leaves from the upper and 20 leaves from the middle portion of plants from each tunnel were taken weekly from immediately before the first release until the end of the trial. Sampled leaves were placed in jars containing 80% EtOH and 0.5% sodium hypochlorite (to dissolve spider mite webbing). In the 122
135 laboratory, the leaves were removed and the contents of each container were examined under a dissecting microscope at 25x for presence of mites. Temperature and humidity inside the greenhouse were recorded hourly on a data logger, which was protected from direct sunlight, throughout the course of this trial. Mite data were analyzed using JMPIN (SAS Institute, Inc., 2002). The weekly number of observed spider mites was converted to mite-days before statistical analysis. Analysis of replicates was done by completely randomized multiple ANOVA. Means were separated by Tukey HSD at α= Results and discussion Daily temperatures at the beginning of the trial were in the low 30 s (30-33ºC) and gradually dropped to the mid- 20 s (24-26 ºC) by the end of the trial. Nighttime temperatures were 17-19ºC at the beginning of the trial and down to ºC by the end of the trial. On a daily basis the relative humidity dropped below 45% for at least 2-3 hours (sometime much longer) and sometimes dropping as low at 25%. Night time RH was generally reached a maximum of 80-85%. Table 1. Mean number of mite-days of spider mites, and standard error, observed in each treatment. First predator release was on low (< 0.1 TSSM/leaf) populations. The second predator release was on higher (1.2 TSSM/leaf) populations. Treatment Significance Mean Mite-Days Std Error 2 nd P. persimilis A Non-treated B nd N. californicus B st P. persimilis B C st N. californicus C D Acaricide D At the first release there was an average of > 0.1 TSSM/leaf, while at the second release there was ~ 1.2 TSSM/leaf. Multiple ANOVA, on the number of mite-days, (main effects: height of leaf sample, treatment) showed that there was no positional effect; therefore, mite counts from leaves in the upper and middle portion of the plants were pooled. Results of the number of spider mites collected weekly are shown in Figure 2. On the days that predators were released, leaf samples were taken prior to release. There was a significant difference between treatments (df = 5, 426, F = , P < ) over the course of the trial (Table 1). The mite rearing system offered distinct advantages over other rearing methods: 1. A humid environment was created which was highly advantageous to egg and mite development, mimicking the domatia where predatory mites often lay eggs on leaf surfaces (Romero & Benson, 2005). 2. Pollen is separated from, and no longer exposed to, humid environments, thus virtually eliminating mold development. 3. Since the mites were reared solely on pollen, no other contaminating and potentially phytophagous mites, such as Tyrophagous spp. (Nakao, 1991) or TSSM (Lee & Lo, 1999), were released with the predators. 123
136 4.5 4 A 3.5 Average # Spider Mites B Average # Spider Mites Oct 15-Oct 20-Oct 25-Oct 30-Oct 4-Nov 9-Nov 14-Nov 19-Nov 24-Nov N. c. P. p. Acar. Figure 2. The average number of spider mites per leaf recovered weekly. A. Release of predatory mites at low TSSM populations. B. Release of predatory mites at higher TSSM populations. Downward arrows indicate date of predatory mite release. Upward arrows indicate date of acaricides treatment. The Arava Valley is a unique and geographically isolated region in Israel. With moderate winters and hot dry summers typical of the Mediterranean Basin climate, agricultural pests often thrive, but not all natural enemies can perform well. Currently, the predatory mite, P. persimilis, is the only biological control agent available for the control of spider mites. While this is a specific predator, it requires high humidity and moderate temperatures to 124
137 establish and thrive. Conditions in the Arava Valley are good for this predator only in greenhouse situations for a limited time frame; once temperatures increase in the spring, spider mite populations increase and P. persimilis does not control them. The average daytime RH was 47%. Neoseiulus californicus is a type II-III predator (feeding on various mites and plant pollen); as demonstrated in this study it possesses the ability to regulate prey populations and is highly adaptable to different biotic and abiotic conditions. Laboratory studies (Rott & Ponsonby, 2001) have clearly demonstrated the ability of N. californicus to pursue its prey at low humidities. When both predatory mites were release on low TSSM populations (> 0.1 mite/leaf) N. californicus outperformed P. persimilis, but not significantly so. Under moderate spider mite populations (~1.2 mites/leaf), P. persimilis were overwhelmed and N. californicus exerted significantly better control. Even though there was no significant difference between the acaricides-treated and early release of N. californicus, there were still some hot-spots of TSSM. More work and possibly multiple releases will have to be examined in the future. Acknowledgements The authors would like to thank Yoav Ben-Rubi for his technical assistance. This research was funded by a grant from the Chief Scientist of the Ministry of Agriculture of Israel, and by a grant from the European Commission, contract COOP-CT EUROMITE. This paper is a contribution from the Agricultural Research Organization, Institute of Plant Protection, Bet Dagan, Israel. References Croft, B.A., Monetti, L.N. & Pratt, P.D. 1998: Comparative life histories and predation types: are Neoseiulus californicus and N. fallacies (Acari: Phytoseiidae) similar type II selective predators of spider mites? Environ. Entomol. 27: Lee C-Y. & Lo, K.-C. 1999: Mass rearing of Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae). Chinese J. Entomol. Special publ. 12: Nakao, H. 1991: Studies on acarid mites (Acari: Astigmata) damaging vegetable plants. II. Damage to vegetable seedlings. Japan. J. Appl. Entomol. Zool. 35: Romero, G.Q. & Benson, W. W. 2005: Biotic interactions of mites, plants and leaf domatia. Curr. Opin. Plant Biol. 8: Rott, A.S. &. Ponsonby D.J. 2001: Control of two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) on edible crops in glasshouses using two interacting species of predatory mite, pp In R. B. Halliday, D. E. Walter, H. C. Proctor, R. A. Norton and M. J. Colloff [eds.], Acarology: Proceedings of the 10th International Congress. CSIRO Publishing; Collingwood; Australia. Schausberger, P. & Walzer, A. 2001: Combined versus single species release of predaceous mites: predator-predator interactions and pest suppression. Biol. Cont. 20: Walzer A. & Schausberger P. 2005: Are two better than one? Combined effects of the predatory mites Phytoseiulus persimilis and Neoseiulus californicus (Acari: Phytoseiidae). IOBC/WPRS Bull. 28:
138 126
139 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Identification of Pollens for Small Scale Mass Rearing of Neoseiulus californicus and a Novel Method for Quality Control Yael Argov 1, Martin Berkeley 1 Silvi Domeratzky 1, Eti Melamed 1, Phyllis Weintraub 2, Eric Palevsky 3 1 Israel Cohen Institute for Biological Control, Plant Production and Marketing Board, Citrus Division, POB 80 Bet Dagan, 50250, yael@jaffa.co.il; 2 Agricultural Research Organization, Gilat Research Center, DN Negev, 85280, phyllisw@volcani.agri.gov.il; 3 Agricultural Research Organization (ARO), Newe-Ya ar Research Center, Ramat Yishay, 30095, Israel, palevsky@volcani.agri.gov.il Abstract: In this study we set out to identify indigenous pollen for small scale mass rearing of Neoseiulus californicus. From ca. 30 plant pollen evaluated, six species were found to be suitable: Zea mays, Quercus ithaburensis and four Pistacia species, P. atlantica, P. vera, P. lentiscus and P. palestina, the latter yielding the shortest duration of development and the highest fecundity. An improved method for monitoring development and fecundity of phytoseiid species on pollens using polyacrylamide gel (PAG) as a water source and barrier was evaluated. The proportion of replicates lost using the PAG method was substantially lower than the conventional wick method. Moreover the wick method took twice as long to complete each evaluation. We propose that the polyacrylamide gel method be adopted for quality control of phytoseiids reared by commercial insectaries. Key words: Neoseiulus californicus, pollen, Zea, Quercus, Pistacia, arenas, polyacrylamide, quality control. Introduction Neoseiulus californicus (McGregor) is a type II to type III phytoseiid predator (Croft et al., 1998) capable of feeding on various mites, insects and pollens and is used commercially to control different species of spider mites. This predator naturally occurs in open fields, vegetables cultivated in green houses and various tree crops in arid and humid ecosystems in temperate and subtropical regions (Castagnoli & Simoni, 2003). Recently, within the frame work of an European project, we imported this dynamic predator to Israel to improve spider mite control in vegetable and tree crops. Our first objective in this study was to identify indigenous pollens suitable for small scale mass rearing of N. californicus. Once identified these pollens could also be utilized to improve the establishment of this predator when prophylactic releases are conducted (Van Rijn et al., 2002). Development and fecundity evaluations are usually conducted in small arenas with one individual mite per replicate. A problem that is frequently encountered as the experiment takes its course is that these individual mites go astray, thus the loss of replicates. Mites will often be found drowned in the water source (free water must be provided otherwise the mites will not survive) or in the barrier that was set up to prevent their escape. While closed cells do prevent escape they are problematic because pollen must be provisioned 2 to 3 times a week, and molts and eggs must be collected every day when monitoring immature development and fecundity, respectively. Consequently an additional focus of this work was to improve the methodology used for the evaluation of development and fecundity of phytoseiid species on pollens. 127
140 Materials and methods One shipment of a few thousands mites of N. californicus were received from Koppert (Berkel en Rodenrijs, Netherlands) at the Israel Cohen Institute for Biological Control (ICIBC), November, This strain was originally collected in California, reared by Biotactics (Romoland, California) and then sent to and subsequently cultured by Koppert, Netherlands (KNL). Pollen from ca. 30 plant species consisting of wild and cultivated hosts were collected. Mite cultures, consisting of 50 females, were established on black plastic arenas (Argov et al., 2002) and fed pollen 3 times per week. At least two arenas were established for each pollen species. Pollen was scored qualitatively according to survival and egg laying using the following index: 1) No mites survived. 2) Mediocre - tens of eggs. 3) Suitable - more than hundreds of eggs. In the second phase of our trials we evaluated the six pollen species, Pistacia palestina (Eastern Terebinth), P. vera (pistachio), P. atlantica (Atlantic pistachio), P. lentiscus (mastic tree), Quercus ithaburensis (Tabor oak) and Zea mays (maize) that were classified as suitable (group 3) and particularly easy to collect in comparison to eggs of Tetranychus cinnabarinus, thought to be their ideal food. Experiments were conducted in growth chambers at 25±1 0 C, 72±4% RH, 16:8 L:D. Two experimental setups were used to monitor development and fecundity of individual mites, both utilizing Agfa (Agfa-Gevaert N.V., Belgium) 35 mm black film containers. 1. Wick method: we used the canister as a water reservoir and the cover (upside down) as the arena. Water was provided via a 3 mm water wick of synthetic fibers threaded through a hole in the center of the cap. To prevent escapees castor oil was injected with a fine syringe into the moat (Reuveny et al., 1996). Following the first set of evaluations (2 months after rearing was initialized) it became apparent that this setup had a number of drawbacks. Replicates would be lost due to drowning in the water source or the castor oil. Another difficulty was that eggs would be laid within the strands of the water wick making them very difficult to find, thereby extending the time needed to complete each evaluation. 2. PAG Method: the cover, right side up, was placed on the surface of polyacrylamide gel (PAG), which served both as a water source and a barrier against escape. Groups of five arenas were placed in trays (19*14*5). To obtain a homogenous texture 3 g of PAG ALCOSORB AB3C (Ciba, UK) were added to 1 l of water, allowed to stand for a few hours, mixed until smooth in a blender, then poured into trays. For each pollen species, the development of forty individuals were assessed, one individual per arena (comprising a replicate). Mite development was monitored daily from eggs 12 ± hr old to adults by observing egg hatch and exuviae of each life stage. Upon reaching adulthood single pairs were placed on one arena and the fecundity was monitored every day for the first ten days. These trials were conducted at least 6 months after rearing was initialized on each pollen source. Data were analyzed using linear fit and ANOVA procedures with JMP5.0.1a (SAS Institute, Inc.). Results and Discussion Pollens that were disqualified in the qualitative screenings (groups 1-2). Mites did not survive on the pollens (group 1, sorted alphabetically) of Acacia saligna (wattle/acacia), Actinidia deliciosa v. bruno (kiwi), Ephedra sp. (joint pine), Helianthus annua (sunflower), Hibiscus sinensis (Hibiscus), Lilium sp. (ornamental lily), Olea europaea (olive), Opuntia ficus-indica (sabra), Phoenix dactylifera (date palm), Pinus canariensis 128
141 (Canary pine) and Tamarix sp. (tamarisk). Fecundity was low (group 2) on the pollens of Banksia sp. (banksia), Carpobrotus edulis (ice plant), Carya illinoinensis (pecan), Quercus boissieri (Cyprus oak), Quercus calliprinos (Calliprinos oak), Ricinus communis (castor bean), Sorghum vulgare (sorghum), Typha domingensis (common reed), Typha latifolia (bulrush) and honey bee pollen loads. Although the fecundity on the pollen of Punica granatum (pomegranate) was high we did not continue to evaluate it quantitatively for two reasons: first, the pollen was too labor intensive to collect, and second due to the direct yield loss as the removal of each flower would be the loss of one fruit. Development and fecundity on six suitable pollens (group 3) Duration of development (egg to adult) was significantly affected by pollen (P=0.0001; F=5.33; df=5,11) and sex (P<0.0001; F=56.21; df=1,11), with no significant interaction (P=0.07; F=2.04; df=5,11). While males developed significantly faster than females on all pollen species, duration of male development was not affected. Female development was affected by pollen (P<0.0001; F=9.36; df=5), Z. mays yielding the longest duration of development and P. palestina the shortest, the latter differing significantly from Q. ithaburensis and Z. mays. (Figure 1). Development was similar on all four species of Pistacia. Comparable results in the pollen ranking were obtained for the fecundity of N. californicus, with Z. mays having the lowest number of eggs/day and P. palestina the highest, coming very close to the fecundity of individuals fed on Tetranychus cinnabarinus, supposedly the optimal food for this predator (P<0.0001; F=55.38; df=6) (Figure 2). Figure 1. Mean duration of development (plus standard errors) of N. californicus from egg to adult fed on pollens of six plant species, Zea mays, Quercus ithaburensis, Pistacia atlantica, P. vera, P. lentiscus and P. palestina. Columns followed by a common letter do not differ significantly at P<0.05 (Tukey s test). Reports in the literature on the fecundity of N. californicus on different pollen species vary. In Egypt on date palm and castor oil pollen, no oviposition was recorded (El-Laithy & El-Sawi, 1998), while our rating of date palm pollen in group one supports the above study, 129
142 our castor oil pollen rating (group 2) does not. In an earlier study conducted in Israel mean fecundity of N. californicus on maize was less than 1egg/day (Swirski et al., 1970) whereas our results found it to be < 2 eggs/day. We believe that these low values from the earlier work conducted in Israel could have been improved by extending the pre-conditioning period necessary for adapting to a new food source (Castagnoli & Simoni, 2003). In this study we observed a significant increase in fecundity on pollen of Z. mays and Q. ithaburensis from the 2 nd to the 12 th month of rearing, whereas the fecundity of individuals fed on T. cinnabarinus did not improve (Figure 3). Our results on Quercus ithaburensis are very similar to those published for Quercus spp. pollen endemic to Tuscany (Castagnoli & Simoni, 2003). It is of interest to note that the Israeli oak species differed considerably between them, Quercus boissieri and Q. calliprinos were placed in group 2 (unfitting for mass rearing) whereas Q. ithaburensis was found to be a very suitable pollen for development and egg laying. In contrast all species of Pistacia evaluated in this study were suitable for mass rearing, albeit P. palestina seems to be the most promising. The proportion of replicates lost using the PAG method was substantially lower than the conventional wick method (Table 1). Moreover the wick method took twice as long to complete each evaluation because of the time needed to find the eggs hidden between the strands of the wick. Another advantage to the PAG method is that the eggs were only laid in the moat surrounding the top surface of the arena thereby facilitating the counting and collecting of the eggs. We believe that the PAG method could be adopted for quality control of phytoseiids reared by commercial insectaries. Figure 2. Mean Fecundity (plus standard errors) of N. californicus fed on pollens of 6 plant species, Zea mays, Quercus ithaburensis, Pistacia atlantica, P. vera, P. lentiscus and P. palestina in comparison to the fecundity of individuals fed on Tetranychus cinnabarinus. Columns followed by a common letter do not differ significantly at P<0.05 (Tukey s test). 130
143 Figure 3. Mean Fecundity (plus standard errors) of Neoseiulus californicus fed on pollens of Zea mays and Quercus ithaburensis in comparison to the fecundity of individuals fed on Tetranychus cinnabarinus after 2 and 12 months of rearing. Columns followed by a common letter, within the same food source, do not differ significantly at P<0.05 (Tukey s test). Table 1. Proportion of replicates lost during the course of development and fecundity experiments on the two types of experimental arenas. Experiment Water Source Percent n Development Polyacrylamide gel 6% 240 Wick 26% 80 Fecundity Polyacrylamide gel 14% 226 Wick 35% 424 In summary in this study we have identified the pollens of a number of indigenous species to the Mediterranean region suitable for small scale mass rearing of N. californicus. The most promising being Q. ithaburensis and the four Pistacia species. As far as we know this is the first time that species of Pistacia have ever been evaluated as a food source for phytoseiid mites. Acknowledgements We are indebted to Dr. Hillary Voet for her statistical guidance. This research was funded by the European Commission, contract COOP-CT EUROMITE. This manuscript is a contribution of the Institute of Plant Protection, Volcani Center, ARO, Israel. 131
144 References Argov, Y., Amitai, S., Beattie, G.A.C. & Gerson, U. 2002: Rearing, release and establishment of imported predatory mites to control citrus rust mite in Israel. BioControl 47: Castagnoli, M. & Simoni, S. 2003: Neoseiulus californicus (McGregor) (Acari Phytoseiidae): survey of biological and behavioural traits of a versatile predator. Redia LXXXVI: Croft, B.A., Monetti, L.N. & Pratt, P.D. 1998: Comparative life histories and predation types: are Neoseiulus californicus and N. fallacis (Acari: Phytoseiidae) similar type II selective predators of spider mites? Environ. Entomol. 27: El-Laithy, A.Y.M. & El-Sawi, S.A. 1998: Biology and life table parameters of the predatory mite Neoseiulus californicus fed on different diet. Zeitschrift Fur Pflanzenkrankheiten Und Pflanzenschutz-Journal Of Plant Diseases and Protection 105: Reuveny, H., Palevsky, E. & Gerson, U. 1996: Laboratory life history studies of the predaceous mite Typhlodromus athiasae (Acari: Phytoseiidae). Sys. Appl. Acarol. 1: Swirski, E., Amitai, S. & Dorzia, N. 1970: Laboratory studies on the feeding habits, postembryonic survival and oviposition of the predacious mites Amblyseius chilenensis Dosse and Amblyseius hibisci Chant [Acarina: Phytoseiidae] on various kinds of food substances. Entomophaga 15: Van Rijn, P.C.J., Van Houten, Y.M. & Sabelis, M.W. 2002: How plants benefit from providing food to predators even when it is also edible to herbivores. Ecology 83:
145 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Are spider mites aware of the presence of thrips? Banafsheh Ahmadi, Seied Hossein Goldansaz, Ahmad Ashouri Plant Protection Dep., Faculty of Horticulture & Plant Protection Science, College of Agriculture & Natural Resources (UTCAN), University of Tehran, Karaj, Iran. Abstract: Onion thrips (Thrips tabaci) and spider mites (Tetranychus urticae) are serious pests of greenhouse crops and they co-occur on the same crop plants. Thrips are known to be omnivores and co-occurrence with their competitors (spider mites) on the same plant parts will cause them to eat mite eggs. We were eager to find out whether or not mites were aware of the presence of their competitors and predators. Key words: Tetranychus urticae, Thrips tabaci, defensive behavior, Cucumeris sativus, interaction. Introduction Onion thrips (Thrips tabaci) and spider mites (Tetranychus urticae) are important pests for the same crops (Helle & Sabelis, 1985; Lewis 1997), so they compete for the same resources. Thrips feed on mite eggs (Trichilo & Leigh, 1986; Wilson & Bauer, 1996; Milne & Walter, 1998; Agrawal & Klein, 2000). Egg feeding has two advantages for thrips: eliminating a competitor (Trichilo & Leigh, 1986; Wilson et al., 1991; Agrawal et al., 1999; Agrawal & Klein, 2000) and improving the nutritional value of their diet (Trichilo & Leigh, 1988; Milne & Walter, 1998). Through experimental observations of our spider mite and onion thrips biotypes we tested the hypothesis Do spider mites do anything to protect their progeny? Material and methods Insect rearing Thrips were collected from stored red onions (Allium haematochiton) and cultured on planted onions; Tetranychus urticae were collected from greenhouse sugar beets and cultured on cucumber plants. Colonies were maintained under controlled conditions of 16h light: 8h darkness and 25±1 C. Experiment Leaves of six-week-old cucumber plants were used in the experiment. Experimental units were glass clip cages (10 x 10 cm) with a round open area (diameter 3.5 cm) on the top, covered with a No thrips mesh (~ 150 µm). Between the two glass parts there was a cotton layer, a cucumber leaf and another cotton layer (figure 1). Forty young mated females (about 4 days old) of T. urticae were selected and separately placed on each of the experimental units. Twenty second larval stages of T. tabaci were added to half of these units (one per cage). Cages were kept under controlled conditions (25±1 C, 60±5 RH, 16 L: 8 D) and checked after 6, 24 and 48 hours. The number of eggs and their location on the arena were recorded. Since T. urticae females lay egg masses on spinning webs, we searched the leaf 133
146 arena to find them. Data were analyzed with the SAS program using ANOVA and Duncan s test (P < 0.05). Lower glass Iron ring (fixing the cover) Upper glass with a round open area The glasses covered with cotton layers internally Clips Cucumber leaf between cotton layers No thrips mesh Figure 1. Clip cage details. Results and discussion No eggs were seen in the first observation period, 6 hours after the start of the experiment. The second observation period (24 h later) showed no significant differences between treatments, either in egg numbers (7.3 and 6.5 eggs per female, respectively, in presence and absence of thrips larvae), or in egg position. But in the third observation period (48 h later), the presence of thrips caused a significant difference (P=0.0037; df=1, 37; F=9.62) in the number and location of egg masses within the leaf arena (Figures 2 & 3). In order to prevent predation on their offspring, females must select a safe oviposition site. Mothers do not always lay eggs on the best site available because they must balance their own reproductive success. Availability and suitability of food and risk of predation are factors affecting this balance that depend on the needs of the mother and her offspring. Faraji (2001) showed that the predatory mite Iphiseius degenerans oviposited close to pollen in the absence of thrips (Frankliniella occidentalis) but oviposited further away from the food source in their presence. Pallini (1998) showed that T. urticae avoided plants that were previously infested with F. occidentalis, in both olfactometer and greenhouse experiments, to prevent intraguild predation and competition on the food source. Grostal & Dicke (1999) showed that given a choice after 24h, fewer adults and eggs were found on leaf discs that had previously contained the specialist spider mite predator, Phytoseiulus persimilis, than on discs unexposed to this predator. Our data did not show any reduction in fecundity, but showed differences in the number and location of egg masses within the leaf arena (Figures 2 & 3). The difference between our experiment and that of Grostal & Dicke (1999) may have been that our predator, T. tabaci, is an omnivore that turns to egg feeding to eliminate its opponents, whereas Ph. persimilis is a specialist predator. As T. tabaci is an egg predator (Trichilo & Leigh, 1986; Wilson & Bauer, 1996; Milne & Walter, 1998; Agrawal & Klein, 2000; Faraji, 2001), and both thrips and spider mites colonize the same site for feeding (Wilson et al., 1991; Wilson & Bauer, 1996) and ovipositing (beneath the leaf and near the ribs), putting their eggs in different places in order 134
147 to reduce the risk of predation constitutes a critical form of defensive behavior on the part of spider mites. Egg masses With thrips larva Without thrips larva Figure 2. Egg dispersion of Tetranychus urticae in the presence and absence of thrips larvae on the leaf disc after 48 hours. 2 Number of egg masses a b 0 With thrips larva With out thrips larva Figure 3. Number of Tetranychus urticae egg masses in the presence and absence of thrips larvae. Acknowledgments We would like to thank Dr. Shekarforoush and Dr. Moazzeni, both from the College of Veterinary Science of Shiraz University, for their kind help. This research was supported by the Faculty of Agriculture, University of Tehran and the College of Agriculture, Zanjan University. 135
148 References Agrawal, A.A. & Klein, C.N. 2000: What omnivores eat: direct effects of induced plant resistance on herbivores and indirect consequences for diet selection by omnivores. J. Anim. Ecol. 69: Agrawal, A.A., Kobayashi, C &. Thaler, J.F. 1999: Influence of prey availability and induced host-plant resistance on omnivory by western flower thrips. Ecology. 80 (2): Faraji, F. 2001: How counter-attacking prey influence foraging and oviposition decision of a predatory mite. Ph.D. dissertation. University of Amesterdam. 76 pp. Grostal, P. & Dicke, M. 1999: Direct and indirect cues of predation risk influence behavior and reproduction of prey: a cause for acarian interactions. Behav. Ecol. 10 (4): Helle, W., & Sabelis, M.W. 1985: Spider mites. Their biology, natural enemies and control. Elsevier Amsterdam. Lewis, T. 1997: Thrips as crop pests. CAB International. 740 pp. Milne, M. & Walter, G.H. 1998: Significance of mite prey in the diet of the onion thrips Thrips tabaci Lindeman (Thys.: Thripidae). Austr. J. Entomol. 37: Trichilo, P.J. & Leigh, T.F. 1986: Predation on spider mite eggs by the Western Flower Thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), an opportunist in cotton agroecosystem. Ann. Environ. Ecol. 15: Trichilo, P.J. & Leigh, T.F. 1988: Influence of resource quality on the reproductive fitness of flower thrips (Thys.: Thripidae). Ann. Entomol. Soc. Am. 81: Wilson, L.J. & Bauer, L.R. 1996: Phytophagous thrips are facultative predators of twospotted spider mites. (Acari: Tetranychidae) on cotton in Australia. Bull. Entomol. Res. 86: Wilson, L.T., & Trichilo, P.J. & Gonzalez, D. 1991: Natural enemies of spider mites (Acari: Tetranychidae) on cotton: density regulation or causal association? Environ. Entomol. 20 (3):
149 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 p. 137 Effect of different types of prey on fecundity of the phytoseiid mite Typhlodromips swirskii, a potential biological control agent for horticultural greenhouse pests in the Canary Islands Estrella Hernández-Suárez 1, Mª del Cristo Velásquez 1, Mª Inmaculada Paz-González 1, Francisco J. González 2, Aurelio Carnero 1, Francisco J. Ferragut 3 1 Entomology Department, Instituto Canario de Investigaciones Agrarias. Ctra del Boquerón s/n, La Laguna Tenerife (Canary Islands), Spain. 2 Koppert Canarias, C/ Roble s/n, Polígono Industrial de Arinaga, Agüimes-Gran Canaria (Canary Islands), Spain. 3 Instituto Agroforestal Mediterráneo, Dpt. Ecosistemas Agroforestales, Universidad Politécnica, C/ de Vera 14, Valencia, Spain Abstract: An increasing number of commercial greenhouse growers from the Canary Islands employ beneficial insects to control pests, but effective biological control of some horticultural pests represent a handicap to the implementation of Integrated Pest Management Programs in our archipelago. That is the case of the western flower thrips Frankliniella occidentalis in cucumber, the broad mite Polyphagotarsonemus latus in sweetpepper, and the tomato ruset mite Aculops lycopersici and the glasshouse whitefly Trialeurodes vaporariorum in tomato. Typhlodromips swirskii has shown a high efficacy against the sweetpotato whitefly Bemisia tabaci, and it has been evaluated as biocontrol agent of many other pests (Tetranychus urticae, Eriophyes dioscoridis, Thrips tabaci, Aphis gossypii, etc.). The present work aimed to study whether T. swirskii could be used for the biological control of different key pests in commercial horticultural greenhouses in the Canary Islands. As the first step, preliminary laboratory trials have been conducted to study the suitability of different types of prey and non-prey food substances (pollen) on the biology of a commercially available strain of the phytoseiid predator T. swirskii. We present the effect of the food type on the predator fecundity. The preys included in the study were F. occidentalis, T. vaporariorum, P. latus and A. lycopersici. The study has been conducted under a joint ICIA- Koppert Biological Systems research project. Key words: Typhlodromips swirskii, biological control, Canary Islands 137
150 138
151 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Feeding rate of Stethorus gilvifrons on Tetranychus urticae in three greenhouse cucumber cultivars with different resistance levels Neda Kheradpir 1, Jafar Khalghani 2, Hadi Ostovan 1, Mohammad Reza Rezapanah 3. 1 Isalamic Azad university, Science and Research Branch, Faculty of Agriculture and Natural Sources, Department of Entomology. Poonak, Tehran, 2 Plant Pests and Diseases Research Institute, Velenjak, Teharn, 3 Biological Control Research Department, Plant Pests and Diseases Research Institute, Velenjak, Tehran, Iran. Abstract: The aim of this study was to determine the effect of three greenhouse cucumber cultivars (Sina, Sultan and Fadia) on the feeding rate of S. gilvifrons on two-spotted spider mites. The experiment was conducted in a growth chamber (25±0.5 C, RH= 65±5% and 16L:8D) on leaf disks with 5 replications. The results were compared among different developmental stages of the predator and the prey. Sultan had the highest predator feeding rate and Fadia had the lowest. Females were observed to have the highest feeding rate, consuming adult mites, while larvae had the lowest feeding rate, mainly consuming mite eggs. Keywords: cucumber, Stethorus gilvifrons, Tetranychus urticae, predation rate Introduction Spider mites from the family Tetranychidae are the most common pests found in greenhouses around the world. Among its varied species, Tetranychus urticae Koch, is the most serious pest affecting greenhouse vegetable crops throughout the world. In Iran, it especially affects greenhouse cucumber, the crop with the highest planting area with respect to other vegetables (Mersino, 2002; Yano et al. 1998). Due to increased concern about pesticide residues in vegetables the use of other pest control methods, like biological control, has increased in importance during the last decade. Stethorus species (Coleoptera: Coccinellidae) could be one the best candidates for controlling this pest over wide areas including the Mediterranean countries and some parts of Europe and Asia (Ofek et al., 1997). In Iran, Stethorus gilvifrons Mulsant is the most common species, especially around Tehran. Three common cucumber cultivars grown in Iranian greenhouses are Sina, Sultan and Fadia. Each of these cultivars affects the biological and demographic traits of T. urticae in different ways (Kheradpir et al., 2005). The mouthparts of Stethorus are particularly suitable for sucking the contents of tetranychid mites. There are few reports about the feeding rates of ladybirds on this genus of tetranychid mites, however according to Islamizadeh and Pourmirza (2001), Chazeau (1974) and Rott and Ponsonby (2000), the feeding rate of this predator increases with larval development and progress to adulthood. These predators are able to slip through the webs that surround the mites (Yoder et al., 2003). Fournier et al. (2003) reported the effect of host plant and leaf structure on the searching efficiency and predation rate of S. siphonulus. The aim of this study was to evaluate the feeding rate of S. gilvifrons on T. urticae on these three cultivars. 139
152 Materials and Methods Experimental conditions Experiments were conducted at the Biological Control Research Department of the Plant Pest and Disease Research Institute in Tehran. The experimental site was a growth room with a temperature of 25±0.5 C, RH= 65±5% and a photoperiod of 16:8 (L:D). All stages included mite and plant rearing and the experiments took place in the growth room. Plant material The plants selected for this experiment, Sina (RS-189), Sultan and Fadia, were 3 cucumber cultivar F 1 hybrids used in the largest greenhouse cropping areas in Iran.. Their seeds were respectively provided by representatives of the Royal Sluis Co., Petoseed Co. and Enza Zaden Co. in Tehran. Cucumber plants were grown in small clear plastic pots that were 75 mm in diameter and 110 mm in height. The pots were irrigated daily and fertilized twice in the first week with a 20:20:20 (N-P-K) formulation at a concentration of 200 ppm. The plants used for the experiment were at the 5 th leaf stage. Predator and prey colonies The natural food for rearing the predator ladybird used in the experiment was T. urticae Koch, which was reared on cow bean plants. The ladybirds were kept in plastic containers, 80 mm in diameter and 40 mm in height, on berry leaf disks that were soaked and placed on wet cotton. Four or five ladybirds of both sexes (3 females and 2 males) were kept in each container and cow bean leaves contaminated with T. urticae were added on a daily basis to nourish the predators. To prevent cannibalism, newly hatched larvae were removed and transferred to another container. Two-spotted spider mites Tetranychus urticae Koch (Acari: Tetranychidae) were reared on lima bean plants. These plants were grown in 900ml plastic pots. Bean plants were infected at the 3-leaf level with 10 adult female mites. Mites of the fifth generation were used for the experiments. Experiment One plant from each cultivar hybrid was infected with T. urticae. After 5 days, 4-5 day old female and male adult predators and 4 th instar larvae were used to conduct the test. The predators were isolated and starved for 24 hours before the test in an empty container. Then 100 mm 2 leaf disks from each infected cucumber cultivar were prepared in 5 replications and put on wet cotton in a tray. The mites on each leaf disk were counted and only 20 adult mites, 20 nymphs, 20 larvae and 20 eggs were kept: the others were discarded. The predators were released individually for 3 hours. The eaten and remaining mites were subsequently counted. There were 5 replications for each cucumber cultivar and predator developmental stage. The results were analyzed with ANOVA and compared using the Duncan test with SPSS 13.0 software. Results and discussion The results from this experiment were first separately analyzed for female, male and 4 th instar larvae and then compared. The mean number of eaten mites on different cucumber cultivars per female predator showed significant differences (P=0.017; Figure 1). The female predators on Sultan ate the most mites and those on Fadia ate the fewest. The greatest number of prey eaten by the female predator was for adult female mites and the smallest number was for mite eggs (Figure 2). The difference between the number of mites eaten at different developmental stages on different cultivars was significant (egg: P=0.031, larvae: P=0.113, nymph: P=0.202 and adult: P=0.001). 140
153 There were significant differences between the feeding levels of male predators on the mites on the different cultivars (P=0.015). The highest level of feeding was recorded on Sultan and the lowest on Fadia (Figure 1). On the other hand, differences between feeding rates at different developmental stages of T. urticae were not significant (egg: P=0.245, larvae: P=0.422, nymph: P= and adult: P=0.010). However, female mites were the ones most eaten and mite eggs were least eaten by male predators (Figure 3). There was no significant difference in the number of mites eaten among the cultivars (P=0.096; Figure 1) or among the different developmental stages of the two-spotted spider mite (egg: P=0.106, larva: P=0.940, nymph: P=0.351 and Adults: P=0.300; Figure 4). It was assumed that there would be a direct relationship between the different developing larval stages and the mean feeding rate of the predator. In other words, the overall feeding capacity would increase until the highest feeding level would be seen in adult predators. On the other hand, Hadjizadeh (2004) registered the highest feeding rate of tetranychids in S. gilvifrons for the adult female predator. This was followed by the 4 th instar larvae, and finally the male predator. He also stated that predator larvae preferred to eat mite eggs and that adult predators preferred to consume adult mites. Chazeau (1974) reported the mean daily feeding rate of Stethorus ladybirds as being about 40 adult mites for adult predators and 100 eggs for predator larvae. The host plant and the physical structure of the plant leaves were the most important factors in determining the searching efficiency of Stethorus predators and this made differences between cultivars significant. Acknowledgments Special thanks to the Biological Control Research Department of the Plant Pest and Disease Research Institute for providing the laboratory and growth room in which this experiment was carried out. References Islamizadeh, R. & Pourmirza, A.A. 2001: Biology and predatory ability of S. punctillum and Orius minutus on Panonychus ulmi in laboratory. J. Agr. Sci. 22(1): Chazeau, J. 1974: Evaluation de l`action predatrice de Stethorus madecassus sur Tetranychus neocaledonicus. Entomophaga, 19: Fournier, V., Rosenheim, J.A., Brodeur, J., Laney, L.O. & Johnson, M.W. 2003: Herbivorous mites as ecological engineers: indirect effects on arthropods inhabiting papaya foliage. Oecologia, 135: Hajizadeh, J. 2004: The effect of prey density on feeding rate and developmental time in ladybird Stethorus gilvifrons. 15 th. Iran Plant Protection Conference, p: 335. Kheradpir, N., Khalghani, J., Ostovan, H. & Rezapanah, M.R. 2005: Effect of different cucumber cultivar hybrids Cucumis sativus on developmental time, fecundity and intrinsic rate of increase (r m ) of Tetranychus urticae Koch. IOBC/WPRS Bull. 28(1): Mersino, E. 2002: Mites on ornamentals: Miscellaneous pests. Dec. 2002: MP-2. Hawaii University Cooperative Extension. 3 pp. Ofek, O., Huberman, G., Izhar, Y., Wyasoki, M., Kuzlitzky, W. & Reneh, S. 1997: Control of oriented red scale (A. orientalis) and of the California red scale (A. aurantii) in Mango orchards in Hevel Maon, Israel. Phytoparasitica, 25(2):
154 Rott, A.S. & Ponsonby, D.J. 2000: Improving the control of Tetranychus urticae on edible glasshouse crops using a specialist coccinellid (Stethorus punctillum) and a generalist mite (Amblyseius californicus) as biological agents. Biological Science and Technology, 10: Yano, S., Wakabayashi, M., Takabayashi, J. & Takafuji, A. 1998: Factors determining the host plant range of the polyphagous mite Tetranychus urticae (Acari: Tetranychidae): a method for quantifying host plant acceptance. Exp. Appl. Acarol., 22: Yoder, J.A., Pollock, D.A. & Benoit, J.B. 2003: Moisture requirements of the ladybird beetle Stethorus nigripes in relation to habitat preference and biological control. Entomol. Exp. Appl. 109: totalf totalm totall Mean Fadia Sultan Sina Figure 1. Total feeding rate of predators (female, male and larvae) on three cucumber cultivars eggf larvaef nymphef adultf Mean Fadia Sultan Sina Figure 2. Feeding rate (mean ± SE) of female predator on T. urticae at different developmental stages. 142
155 eggm larvaem nymphem adultm 8.00 Mean Fadia Sultan Sina Figure 3. Feeding rate (mean ± SE) of male predator on T. urticae at different developmental stages eggl larvael nymphel adultl 5.00 Mean Fadia Sultan Sina Figure 4. Feeding rate (mean ± SE) of larvae predator on T. urticae at different developmental stages. 143
156 144
157 Thrips, leafminers, aphids and lepidoptera pests
158 146
159 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Development of traps for monitoring thrips (Thysanoptera: Thripidae) Chang-chi Chu 1, Matthew A. Ciomperlik 2, Marcus Richards 3, Thomas J. Henneberry 2 1 USDA, ARS WCRL, Phoenix, AZ 85040; 2 USDA, APHIS, PPQ, CPHST, PDDML, Edinburg, TX 78541; 3 MAF, St. Vincent and the Grenadines, W.I., USA. Abstract: Scirtothrips dorsalis was identified as an invasive pest in the Caribbean area in 2004 and was recognized as established in Florida in The species is recognized as a significant threat to agriculture and trade in both regions and potentially worldwide. Methods are urgently needed to detect, monitor, and manage the pest. We developed traps for detection and monitoring Frankliniella occidentalis in cotton Gossypium hirsutum and alfalfa Medicago sativa, and S. dorsalis and other thrips on chilli peppers Capsicum chinense in the field. Key Words: Scirtothrips dorsalis, CC traps, Blue-D traps, LED-yellow sticky card traps, Caribbean Introduction Scirtothrips dorsalis (Hood) was described in 1916 as a new species collected from castor bean and chili plants in Coimbatore, Southern India (Hood 1919). S. dorsalis is a polyphagous species with habitats range from temperate to tropical climate regions in India, Pakistan, the Caribbean, Taiwan, Japan and Australia (Mound & Palmer 1981) and with more than 100 host plant species from more than 40 families (Meissner et al. 2005). Almost all the plant families occur in the United States. Host plant species include tea, cashew nut, citrus, avocado, mango, grapes, chili, cotton, tomato, tobacco, peanut, and castor bean. The insect has been reported by the U.S. Animal Plant Health Inspection Serviceas one of the thirteen most important pest species that could become a serious threat to U.S. agricultural crops if it becomes established in the country (USDA-APHIS 2004). The insect has also been an A2 quarantine pest (EPPO 2004). Live S. dorsalis were found on chili peppers Capsicum chinense (L.) shipped from St. Vincent and the species identity verified in St. Vincent and St. Lucia (Ciomperlik & Seal 2004). Monitoring to obtain information on the spread of the species to new areas has received high priority attention. To obtain specimens for taxonomic and genetic studies, leaf samples from plant terminals can be collected, stored in ziplock bags, and washed with ethanol to obtain intact specimens (Ciomperlik & Seal, 2004). The CC traps also collected intact S. dorsalis specimens from a peanut filed in India in 1996 (Chu et al. 2000). Traps can be installed in commercial crop production areas where the pest has been found or suspected and serviced periodically for long periods of time. We have since tested different colored sticky card traps and modified CC traps, and modified a commercially available 2,2-dichlorovinyl dimethyl phosphate (DDVP, Hot Shot No-Pest Strip, United Industries Corp., St. Louis, Mo) dispenser for detection and monitoring S. dorsalis and other thrips under field conditions in Caribbean inlands (Chu et al. 2005, Chu et al. 2006). This report summarizes results of studies conducted in cotton and commercial alfalfa field with Frankliniella occidentalis (Pergrande) in Maricopa, AZ, and chili pepper fields with S. dorsalis in St. Vincent in
160 Materials and Methods Frankliniella occidentalis (Pergande), Maricopa, AZ Sticky card traps, CC traps, and trap types. The experiments were conducted in a randomized complete block design with 8, 15, and 10 replicates for sticky card traps, CC traps, and trap types, respectively at Maricopa, AZ. The sticky card traps experiment was conducted in cotton field. Plots were not treated with pesticides. Blue sticky card trap tested were: Sensor, blue Bug-Scan, and Takitrap. Traps spaced 1 m apart were placed 8 cm above plant canopy tops. Traps were replaced weekly from 22 June to 13 July Trapped F. occidentalis were counted with aid of stereoscopes. The CC traps experiment was conducted in in a commercial cv. Cebola alfalfa, field. DDVP (1 cm 2 x 0.5 cm cubes) were placed one cube in each trap to kill and retain the F. occidentalis that entered blue base CC traps. Trap catches of were counted daily. The same DDVP cubes were used during the entire experimental period, from 15 to 21 July The trap types experiment was conducted in the same alfalfa field mentioned above in a randomized complete block design with ten replicates. Treatments were blue base CC traps with one cm 2 DDVP cube, commercial DDVP dispensers, and blue stripe modified DDVP dispensers (Blue-D). Vertically oriented blue plastic strips (two 2.5-cm wide strips spaced at 2.5 cm apart) were attached inside the front and back surfaces of the DDVP dispenser packages. Traps were fixed on 30 cm long garden stakes that were held in place on 120-cm long wooden stakes with 5-cm wide binder clips. The trap bottoms were at the level of alfalfa terminals. Scirtothrips dorsalis and other thrips species, Georgetown,St Island Sticky card traps, CC traps and trap types. The experiments were conducted in a randomized complete block design with 10 replicates each for sticky card traps and trap types. The experiments were conducted about one mile inland from the Caribbean Sea in two (~ 0.2 ha each) geographically separated chili pepper farms during the dry season in Plants were set 1 m apart within and between rows in both fields. White, blue, and yellow sticky card traps were each 10.0 by 10.5 cm in size. Traps were placed with wire loops attached to 25 cm long wooden stakes 5-10 cm above the plant terminal. The trap types were installed in the chili farms and held on 120 cm long wooden stakes similar to the one described above. Statistical Analysis Numbers of thrips caught in traps were averaged over sampling periods for each experiment. Data were analyzed using ANOVA orthogonal comparisons or three factor factorial analyses (Anonymous 1989). Means were separated using Tukey s HSD. Results and discussion Frankliniella occidentalis (Pergande) - Maricopa, AZ Sticky card traps, CC traps, and trap types. F. occidentalis catches in cotton plots were highest on blue sticky cards with peak wavelength reflectance of 458 nm compared with traps with 444 nm or 446 nm wavelength reflectances (Table 1). More F. occidentalis were caught per day in white or blue base CC traps compared with yellow base CC traps. Blue stripe modified commercial DDVP dispenser (Blue-D traps) caught more F. occidentalis compared with unmodified DDVP dispenser and the blue base traps with one cm 2 DDVP cubes. 148
161 Table 1. Mean numbers of Frankliniella occidentalis caught on blue sticky card traps (with different peak wavelength reflectance), on CC traps and on Trap Types in a cotton field, Maricopa, AZ, No. thrips/ trap/ day Variable Mean Range Blue sticky card Sensor (444 nm) a 5.7±1.0c b 2.1± ±2.3 Blue Bug-Scan (446 nm) 38.9±6.9b 18.1± ±26.1 Takitrap (458 nm) 60.3±5.9a 40.9± ±13.4 CC traps White 0.7±0.1a a 0.0± ±0.8 Blue (458 nm) 0.8±0.1a 0.1± ±0.7 Yellow (552 nm) 0.2±0.1b 0.0± ±0.7 Trap types CC trap + one cm 2 DDVP cube 32.6±3.6b 11.2± ±10.3 DDVP dispenser alone 37.5±2.8b 6.0± ±5.5 DDVP dispenser + blue stripes 53.4±5.1a 7.5± ±7.9 a Peak wavelength. b Means in a column of a variable not followed by the same letter are significantly different by Tukey s HSD, P=0.05. Scirtothrips dorsalis and other thrips species, Georgetown, St. Vincent Island Sticky card traps, CC traps, and trap types. Significantly more S. dorsalis were caught on yellow sticky card traps compared with white and blue sticky card traps (Table 2). More T. palmi, Frankliniella sp. and M. abdominalis were caught on blue sticky card traps compared with white and yellow sticky card traps. Blue-D traps caught more T. palmi, Frankliniella sp., and M. abdominalis than CC traps (Table 3). Mean captures of S. dorsalis for the two trap types were not significantly different. Capture of S. dorsalis in CC traps with different trap color bases were not significantly different. White and blue base traps caught more T. palmi, Frankliniella sp. than yellow base traps. Blue base traps caught more Frankliniella sp. and M. abdominalis than yellow base traps. DDVP cubes in traps resulted in increased captures of all thrips species. Ethylene glycol increased trap catches. Thrips species captured in traps with ethylene glycol were well preserved, with less damage to antennae and less desiccation than traps without ethylene glycol. Blue trap base traps with DDVP cubes caught greater numbers of all thrips species. Blue trap base traps with ethylene glycol also caught more Frankliniella sp than other trap base and ethylene glycol cominbations. Color attraction is an innate characteristic of insects. It is well demonstrated in the literature known that yellow is attractive to a great numbers of insect, eg. Asterochiton vaporarorium (Westwood), since early 1900 s (Lloyd, 1921). Results of our studies indicate that yellow and blue sticky card traps are useful for trap thrips in greenhouse environments. For S. dorsalis, yellow sticky traps appear to be a better choice than blue sticky traps. Chen et al. (2004a, 2004b) reported that blue sticky card traps equipped with blue light emitting diode (LED) caught more F. occidentalis than without LED in the field and yellow sticky card equipped with yellowish green LED trapped more pest insects than without LED under greenhouse conditions. 149
162 Table 2. Seasonal means (±SE) of Scirtothrips dorsalis and other thrips species caught on white, blue, and yellow sticky card traps in commercial chili pepper fields, Georgetown, St. Vincent, dry season, 23 March to 4 May No. thrips/ trap/ week Sticky trap Microcecolor S. dorsalis T. palmi Frankliniella sp phalothrips sp White 1.41±0.11b a 8.04±1.45b 2.08±0.34b 4.39±0.57b Blue 3.72±0.37b 27.11±2.20a 8.75±0.85a 21.40±1.79a Yellow 14.10±1.06a 7.38±0.62b 1.73±0.18b 4.30±0.45b a Means in a column not followed by the same letter are significantly different by Tukey s HSD, P=0.05. The blue-d trap, a modified commercial available DDVP dispenser caught more species of thrips than the CC traps on St. Vincent Island. The blue-d traps also caught more S. dorsalis in Taiwan compared to the CC traps (Chu et al. 2006). The addition of ethylene glycol to CC traps increased trap catches of S. dorsalis and M. abdominalis on St. Vincent Island. Although the CC trap captures fewer S. dorsalis than sticky card traps, the quality of the captured specimens is high. They are easily recovered from the trap and stored in ethanol for later taxonomic and genetic analysis. Studies of S. dorsalis behavior, including the development of attractants and pheromones as potential lures, are being conducted to develop more efficient trap systems for detection and monitoring of this insect pest. The spread of S. dorsalis from the Caribbean to Florida through agricultural trade and tourism (Meissner et al. 2005) was predicted, however, the speed with which this occurring was not realized. The potential for rapid spread of this pest thrip species to other continent in field and greenhouses is a distinct possibility. The potential impact of this species on agriculture in the United States alone has been estimated at approximately $3.6 to $6.0 billion a year (Lynn Garrett, USDA APHIS PPQ CPHST, personal communication). Effective survey and detection methods are needed to monitor the spread, and manage populations of S. dorsalis in the Caribbean, US, and other geographical areas. 150
163 Table 3. Seasonal means (±SE) of Scirtothrips dorsalis and other thrips species caught in commercial chili pepper fields, Georgetown, St. Vincent, dry- season, 23 March to 4 May No. thrips/ trap/ week Microcephalothrips S. dorsalis T. palmi Frankliniella sp. abdominalis Trap type Blue-D a 0.05±0.03a b 1.06±0.33a 0.21±0.09a 0.33±0.06a CC trap 0.09±0.02a 0.49±0.08b 0.08±0.02b 0.17±0.03b CC trap (base color) White 0.06±0.02a 0.53±0.13a 0.08±0.02ab 0.16±0.04ab Blue 0.13±0.04a 0.78±0.16a 0.14±0.04a 0.25±0.06a Yellow 0.08±0.02a 0.17±0.04b 0.02±0.01b 0.10±0.03b Dichlorvos in CC traps Yes 0.13±0.03a 0.82±0.12a 0.14±0.03a 0.31±0.04a No 0.05±0.01b 0.17±0.03b 0.02±0.01b 0.04±0.01b Base-DDVP White-yes 0.08±0.04b 0.89±0.18b 0.13±0.03b 0.28±0.06b White-no 0.04±0.02b 0.18±0.07c 0.03±0.02bc 0.03±0.03c Blue-yes 0.23±0.06a 1.35±0.19a 0.26±0.06a 0.46±0.07a Blue-no 0.03±0.01b 0.22±0.06c 0.02±0.01c 0.04±0.03c Yellow-yes 0.08±0.02b 0.22±0.06c 0.04±0.02bc 0.18±0.05bc Yellow-no 0.08±0.03b 0.12±0.04c 0.01±0.01c 0.03±0.01c Ethylene glycol in CC traps Yes 0.13±0.02a 0.51±0.09a 0.11±0.03a 0.22±0.04a No 0.05±0.02b 0.48±0.12a 0.05±0.02b 0.12±0.04b Base-ethylene glycol White-yes 0.10±0.04a 0.57±0.15a 0.10±0.03ab 0.24±0.07a White-no 0.03±0.02a 0.50±0.21a 0.06±0.03b 0.08±0.03a Blue-yes 0.15±0.05a 0.74±0.21a 0.21±0.07a 0.27±0.08a Blue-no 0.11±0.05a 0.83±0.26a 0.07±0.03b 0.23±0.09a Yellow-yes 0.13±0.02a 0.23±0.06a 0.02±0.01b 0.16±0.05a Yellow-no 0.03±0.01a 0.11±0.04a 0.03±0.02b 0.05±0.02a a DDVP dispenser plus blue stripes (Blue-D). b Means in a column of the same variable not followed by the same letter are significantly different by orthogonal comparison for Blue-D vs. CC traps and by Tukey s HSD for CC traps, P =
164 References Cited Anonymous. 1989: MSTATC. A microcomputer program for the design, management, and analysis of agronomic research experiments. Michigan State Univ. Chen, T.Y., C.C. Chu, E.T. Natwick, & Henneberry, T.J a: Trap Evaluations for thrips (Thysanoptera: Thripidae) and hoverflies (Diptera: Syrphidae). J. Environ. Entomol.: Chen, T.Y., C.C. Chu, T.J. Henneberry, & Umeda, K. 2004b. Monitoring and trapping insects on poinsettia with yellow sticky card traps equipped with light-emitting didoes. HortTech. 14: Chu, C.C., Pinter, P.J.Jr., Henneberry, T.J., Umeda, K., Natwick E.T., Wei, Y.A., Reddy, V.R. & Shrepatis, M. 2000: Use of CC traps with different trap base colors for silverleaf whiteflies (Homoptera: Aleyrodidae), thrips (Thysanoptera: Thripidae), and Leafhoppers (Homoptera: Cicadellidae). J. Econ. Entomol. 93: Chu, C.C., Ciomperlik, M., Chen, T.Y., Tuck, S., Alexander, P. & Henneberry, T.J. 2005: Variations in CC trap catches of thrips associated with different colors with or without dichlorvos cubes, pp In P. Dugger and D. Richter [eds.] Proc. Belt. Cotton Conf., New Orleans, LA. Chu, C.C., Ciomperlik, M.A., Chang, N.T., Richards, M. & Henneberry, T.J. 2006: Developing and evaluating traps for monitoring Scirtothrips dorsalis (Thysanoptera: Thripidae). Florida Entomol. (in press). Ciomperlik, M.A., & Seal, D. 2004: Surveys of St. Lucia and St. Vincent for Scirtothrips dorsalis (Hood), Jan , USDA APHIS PPQ, Technical Report. 19 pp. EPPO. 2004: EPPO Standard PM1/2(13) A1 and A2 Lists of pests recommended for regulation as quarantine pests - adopted in ( lists.htm), European and Mediterranean Plant Protection Organization. Hood, J.D. 1919: On some new Thysanoptera from southern India. Insecutor Inscit. Menstir. 7., Lloyd, L. 1921: Notes on a colour tropism of Asterichiton (Aleurodes) vaporariorum, Westwood. Bull. Entomol. Res. 12: Meissner, H., Lemay, A,. Borchert, D., Nietschke, B., Neeley, A., Magarey, R., Ciomperlik, M., Brodel, C. & Dobbs T. 2005: Evaluation of Possible Pathways of Introduction for Scirtothrips dorsalis Hood (Thysanoptera: Thripidae) from the Caribbean into the Continental United States. Plant Protection and Quarantine. Center for Plant Health Science and Technology. 124 pp. Mound, L.A., & Palmer J.M. 1981: Identification, distribution and host plants of the pest species of Scirtothrips (Thysanoptera: Thripidae). Bull. Entomolo. Res. 71: USDA. 2003: Port Information Network (PIN-309): quarantine status database. US Department of Agriculture, Animal and Plant Health Inspection Service, Plant Protection and Quarantine, Riverdale, MD. Restricted access database. USDA Offshore Pest Information System. 2004: Scirtothrips dorsalis Confirmed as Present in Trinidad and Tobago, reported December 23, 2004, USDA Restricted access database. 152
165 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Preliminary field trials with the synthetic sexual aggregation pheromone of Frankliniella occidentalis on protected pepper and tomato crops in South-east Spain M. Gómez 1, F. García 1, R. GreatRex 2, M. Lorca 1, A. Serna 1 1 Syngenta Bioline España. 2 Syngenta Bioline Ltd. Telstar Nursery. Holland Road, Essex, CO16 9QG - UK Abstract An aggregation pheromone produced by males of Frankliniella occidentalis has recently been identified and synthesised. Results of two trials of a commercial product containing this pheromone are presented here. In protected Sweet Pepper and Tomato crops grown in South East Spain, the addition of pheromone to blue sticky traps produced a significant increase in the numbers of thrips caught. The role of this pheromone as a monitoring tool within IPM programmes is discussed. Key words: Frankliniella occidentalis, pheromones, sticky traps, ICM, monitoring Introduction Western Flower Thrips, Frankinella occidentalis, is considered to be one of the main pests in horticultural crops in Spain, including flowering ornamentals, salad vegetables and fruits (Lacasa & Llorens, 1998). It is one of the main phytosanitary problems on tomato and pepper crops, which are the most important protected crops in Almeria, not only because of the direct damage that it causes on fruit and flowers, but also because of the eficiency with which it transmits Tomato Spotted Wilt Virus (TSWV). During the season 2004 to 2005, 9,100 ha of tomato and 8,955 ha of pepper were grown in Almeria, with a production of 632,957 million tonnes and 451,185 million tonnes respectively (Anuario de la Agricultura Almeriense, 2005). Since Tomato Spotted Wilt Virus was first reported in May 1990 in pepper greenhouses at the Campo de Cartagena (Lacasa et al., 1991), several measures have been taken in order to reduce the devastating effects of this virus disease. These measures include the use of antithrips mesh, coloured sticky traps, the introduction of biological control agents, and the use of virus tolerant varieties. Blue sticky traps are widely used for the capture and monitoring of F. occidentalis because the blue colour provides the highest visual attraction according to the great majority of authors. (Vernon & Gillespie, 1990; Rodrigo & Carnero, 1992; Lacasa & Llorens, 1998). These traps are an essential tool for monitoring the evolution of thrips populations, not only in protected crops but in the open field as well. Their use allows growers to introduce biological control agents at the appropriate time, and to make insecticide treatments according to need rather than according to the calendar, and thus to reduce the total number of insecticide applications made to crops. It has been reported that the first adult thrips entering sweetpeppers greenhouses are best detected on sticky traps rather than by direct plant sampling (Belda et al., 1992, Cabello et al., 1993). In the same way, the trap height affects the number of captures: traps must be placed above the level of the crop because reduction of light and colour intensity inside the vegetable canopy reduces the number of thrips caught (Rodrigo & Carnero, 1992). 153
166 Teerling et al (1993) and McDonald et al (2002, 2003) have reported that F. occidentalis larvae, but not adults, produce an anal droplet containing an alarm pheromone which has many effects on the behaviour of both larvae and adults. Several species of thrips are known to form mating aggregations on flowers (Terry, 1997). Several authors have now demonstrated the existence of a male produced pheromone which is involved in this aggregation behaviour (Hamilton & Kirk, 2003, Kirk & Hamilton, 2004, de Kogel & van Deventer, 2003). Hamilton et al, (2005) subsequently identified an aggregation pheromone produced on demand by male thrips during mating displays. The authors used gas chromatography/mass spectrometry to demonstrate that two major odor components and five minor components were present in male odor and not in female odor of adult F. occidentalis. This male odor attracts virgin females and previously mated females, as well as adult males, to aggregations in which mating takes place (Hamilton et al, 2005). This is the first such pheromone identified from the order Thysanoptera. The major components of this male-produced aggregation pheromone have subsequently been identified and synthesised (Hamilton et al, 2005). In field trials, synthetic compounds were tested in sweet pepper greenhouses with natural thrips populations in Spain. Captures of adult male and female F. occidentalis were increased on blue and yellow sticky traps treated with these compounds. Material and methods The trials reported here were carried out in sweet pepper and tomato greenhouses located in the area of protected crops in the west of Almeria. They form part of a series of trials carried out in several European countries and in the USA. Sweet-pepper greenhouse The sweet-pepper greenhouse was located in Dalías. It was a typical Almeria pepper greenhouse with a total surface area of 1500 m 2. Vents on the sides and roof of the greenhouse were all covered with mesh (10x14 threads/cm 2 ) to limit thrips entry. The trial was conducted on an area of 675 m 2 at the northern end of the greenhouse. The sweet pepper grown was the cultivar Baltasar, of the californian red type, at a density of 2 plants/m 2. Plants were transplanted on June 7 th 2005 and grown according to normal grower practice. Amblyseius cucumeris (Amblyline cu CRS ) and Orius laevigatus (Oriline l ) were released for the control of F. occidentalis, and no chemical treatments were necessary against this pest. Blue plastic sticky traps (22.5 x 20 cm) (Trapline ), coated with glue on both surfaces, were suspended vertically with the base 30 cm above crop height. Twenty traps were used in total, 10 treated and the rest without pheromone (control). A single lure was stuck to the center of one side of each of the treated traps. Commercially packed pheromone lures consisting of a rubber septum dosed with 30µg of the F. occidentalis synthetic aggregation pheromone were used. Treated traps and control traps were placed 7 metres apart, and a minimum of 5 metres from the edge of the greenhouse. The trial started on August 22nd 2005, when the crop was flowering well. Sixty five percent of the plants sampled had thrips, with an average thrips population of 1.4 adults per flower overall. Cumulative captures of F. occidentalis on each trap were recorded one hour, 2 days and 7 days after trap placement (T+1h, T+2 and T+7). 154
167 Tomato Greenhouse The tomato greenhouse was sited at El Ejido (Estación Experimental de Cajamar Las Palmerillas ). This is a multi-tunnel metal structure consisting of eight bays, with lateral ventilation on north and south edges and roof ventilation on each bay. All vents were protected by 10 x 20 thread/cm 2 mesh to limit thrips entry. The tomato cultivar was Pitenza, a truss tomato variety, grown on a perlite substrate at a density of 1.8 plants/m 2. No specific chemical treatments were applied for thrips control. Blue plastic sticky traps (22,5x40 cm) (Trapline ), coated with glue on both surfaces, were suspended vertically with the base 30 cm above crop height. The pheromone lures were stuck to the center of one side of each trap. Ten blue traps were placed in two adjacent bays, 5 of them with pheromone lures and 5 without lures as controls. The traps were 8.3 m apart along the rows and 15 m apart between the bays. The traps were placed on January 11th The cumulative adult captures of F. occidentalis for each trap were recorded 30 minutes after trap placement, then again after 1 hour, 1 day, 5 days and 12 days (T+30, T+60, T+1, T+5 and T+12). The data was statistically analysed using t tests from the statistical analysis package incorporated with Microsoft Excel. Using this test, P values of 0.05 or below indicate a significant difference between the treatments. At the end of each trial two traps were selected at random for thrips identification, to confirm that the majority of the thrips were F. occidentalis. Results and discussion Sweet-pepper greenhouse Table 1 shows that the average numbers of thrips are aproximately three times higher on the traps treated with the pheromone than on the control traps. The differences are statistically significant despite high variability between the catches on traps in different areas of the crop. The highest ratio between the thrips catches on treated traps and control traps, just one hour after trap placement (3.56), gives an indication of the quick response of F. occidentalis to the pheromone. This rapidity of response has also been noted by growers. Although the numbers of male and female thrips were not recorded during this trial, both sexes were present on the traps. Both sexes are attracted by the pheromone (Kirk & Hamilton, 2004), and approximately equal numbers of males and females have been recorded in previous trials (Hamilton et al, 2005, internal trials). Tomato Greenhouse Table 2 shows the average of the cumulative thrips captures per trap and the ratios between treated and control traps. As in the sweet-pepper greenhouse, traps treated with the aggregation pheromone catch significantly more thrips than the control untreated traps at all intervals except 30 minutes after trap placement. The ratios between treated and untreated are lower than in the pepper crop. The biggest ratio between control and treated traps is recorded 1 day after the pheromone traps were placed in the crop (2.46). The high value (2.15) 60 minutes after introduction of the pheromone indicates, as in the pepper crop, that thrips respond rapidly to the presence of the pheromone. The results of these field trials show that the synthetic commercial version of the aggregation pheromone significantly improves the attractivness of blue sticky traps to F. occidentalis. Similar results have been obtained in trials in other European countries and in the USA. This increase in sensitivity of traps allows earlier and more sensitive detection of 155
168 the arrival of thrips in protected crops, and consequently better timing of biological or chemical applications for the control of thrips. The attractiveness of the pheromone to males and females of F. occidentalis, and the use of the pheromone together with a visual stimulus, such as a flower or other distinctive object, for the formation of mating aggregations, makes this pheromone different from the Lepidopteran pheromones which are familiar monitoring tools in agriculture Table 1. Average thrips captures and ratio between treated traps and control. Treated traps Control T + 1 hour T + 2 days T + 7 days Average/trap Standard Deviation (SD) Average/trap SD P Ratio treated:control Thrip adults / trap Control trap Treated trap T + 1 hour T + 2 days T + 7 days Figure 1: Cumulative catches of WFT in peppers Further trials are in progress to determine the direct impact of the pheromone on thrips populations, and the implications of this product for the transmission of TSWV within crops. It is hypothesised that earlier detection of the first viruliferous thrips entering a crop, and more rapid treatment of that initial population, will help reduce the rapidity of spread of TSWV within the crop. 156
169 Acknowledgements We are grateful to Mr. Diego Arévalo and Estación Experimental de Cajamar Las Palmerillas for access to their greenhouses to conduct our experiments. Thanks also to Mrs Ana Belen Arévalo and Juan Carlos Gázquez for help counting thrips in the tomato trial. Thripline ams is being marketed under agreement with Keele University, England, who own the patents on the aggregation pheromone. Table.2: Average thrips captures and ratios between treated and control traps T + 30 T + 60 T + 1 days T + 5 days T+12 days Control AVERAGE SD Treated AVERAGE traps SD P ns Ratio treated:control Thrip adults / trap Control trap Treated trap T + 30 min T + 60 min T + 1 T+5 T+12 Figure 2: Cumulative catches of thrips in tomatoes References Anuario de la Agricultura Almeriense. 2005: La Voz de Almeria. Pag Belda, J, Cabello, T., Ortiz, J. & Pascual, F. 1992: Distribución de Frankliniella occidentalis (Pergande) (Thysanoptera; Thripidae) en cultivo de pimiento bajo plástico en el sureste de España. Bol. San. Veg. Plagas, 18 (1). 157
170 Cabello, T., Carreño, R., Belda, J. E. & Pascual, F. 1993: Modelo de Distribución espacial y temporal de Frankliniella occidentalis (Thys.; Thripidae) en cultivo de pimiento en invernadero. IV Jornadas Científicas de la Sociedad Española de Entomología. Tenerife. de Kogel, W.J. & van Deventer, P. 2003: Intraspecific attraction in Western Flower Thrips, Frankliniella occidentalis: indications for a male sex pheromone. Entomol. Exp. Appl. 107; Hamilton J.G.C. & Kirk, W.D.J. 2003: Method of monitoring/controlling thrips. Priority date 21Dec International patent application published under the Patent Cooperation Treaty WO 03/ World Intellectual Property Organisation, Geneva. Hamilton, J., Hall, D., & Kirk, W. 2005: Identification of a male-produced aggregation pheromone in the western flower thrips Frankliniella occidentalis. Journal of Chemical Ecology, 31(6): Kirk, W. & Hamilton, J. 2004: Evidence for a male-produced sex pheromone in the western flower thrips Frankliniella occidentalis. J. Chem.l Ecol. 30(1): Lacasa, A; Torres, J. & Martínez, M. C. 1991: La implicación de Frankliniella occidentalis en la diseminación del virus del bronceado del tomate (TSWV) en los cultivos murcianos. Agrícola Vergel, 112. Lacasa, A. & Llorens, J. M. 1998: Trips y su Control Biológico. Vol. I and Vol. II. MacDonald, K.M., Hamilton, J., Jacobson, R. & Kirk, W. 2002: Effects of alarm pheromone on landing and take-off by adult western flower thrips. Entom. Exp. Appl MacDonald, K.M., Hamilton, J., Jacobson, R. & Kirk, W. 2003: Analysis of anal droplets of the western flower thrips Frankliniella occidentalis. J. Chem. Ecology 29. Rodrigo, P. & Carnero, A. 1992: Efecto del color y la altura de trampas pegajosas sobre Frankliniella occidentalis (Pergande) (Thys.; Thripidae) en calabacín bajo invernadero. Investigación Agraria Prod. Prot. Veg. 7 (3). Teerling, C.R., Pierce, H.D., Jr. Borden, J.H., & Gillespie, D.R Identification and bioactivity of alarm pheromone in the western flower thrips, Frankliniella occidentalis. In Kirk, W. and Hamilton, J., Evidence for a male-produced sex pheromone in the western flower thrips Frankliniella occidentalis. J. Chem. Ecology 30(1). Terry, L.I. 1997: Host selection, communication and mating behaviour. pp In Thrips as Crop Pests. Lewis. T. (Ed) CAB International Thripline ams. Hojas Técnicas Syngenta Bioline, In Vernon, R.S. & Gillespie, R.S : Spectral responsiveness of Frankliniella occidentalis (Thys.; Thripidae ). In Moreno, R. (Ed), Sanidad Vegetal en la Horticultura Protegida. Cursos Superiores 1/94. Consejeria de Agricultura y Pesca. Junta de Andalucia. 158
171 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp The addition of potassium silicate to the fertilizer mix to suppress Liriomyza leafminers attacking chrysanthemums Michael P. Parrella, Tom Costamagna Department of Entomology, University of California, Davis, CA 95616, USA Abstract: Silicon is the second most abundant element in soils, and is essentially the mineral substrate for most of the world s plant life. This material has long been associated with increasing a plant s ability to withstand attack by pests, but data in this area are limited. We examined whether the addition of potassium silicate to potted chrysanthemum plants would reduce development of the leafminer, Liriomyza trifolii. A range of silicate concentrations were made on a regular basis for the full duration of a potted chrysanthemum crop, and then individual plants were repeatedly challenged with mated pairs of leafminers in caged studies. At silicon concentrations of 200 ppm and higher we observed a significant reduction in leafminers emerging from treated plants vs. the control. These results suggest that the addition of silicon may increase the chrysanthemum s ability to withstand attack by leafminers, and may be an additional cultural tool in the IPM arsenal. Key words: potassium silicate, chrysanthemumus, cultural control, Liriomzya trifoli Introduction Silicon is the second most abundant element in soils, and is essentially the mineral substrate for most of the world s plant life. Water in soil contains silicon, mainly as salicic acid H 4 SiS0 2 at concentrations similar to that of potassium, calcium, and other major plant nutrients, and well in excess of those of phosphate. In spite of this prominence of silicon as a mineral constituent of plants, it is usually not counted among the elements or nutrients that are deemed essential for plant growth. For that reason it is often not in the formulation of any of the commonly used nutrient solutions. In an excellent review which focused on this anomaly regarding silicon in plant biology, Epstein (1993) points out that silicon has the potential to afford protection of plants from both abiotic (mineral toxicity and salinity) as well as biotic stress (fungi, insects and weeds). This was first observed in 1940, where silicon applications were shown to protect agricultural crops from fungal attack. Today, there are potassium silicate solutions available for use in the greenhouse (Zacsil AG and Pro-Tekt) which act as a silica supplement and ph buffer, and they are also touted as helping control powdery mildew on some plants. These are not registered as pesticides, but the literature suggests they may play a role in reducing attacks by insects and diseases and certainly the literature provided by the manufacturers supports this potential added benefit. We became intrigued by the possibility that potassium silicate could be useful in increasing a plant s ability to resist attack by insects and fungi in greenhouses for two reasons: 1) several highly respected growers in California use these products and claim overall healthier plants and increased protection against plant pests, and 2) we began using these products in our greenhouses at UC Davis and our observations appeared to support what growers were finding. There are many products on the market, both pesticides and fertilizers, for which there are label claims that may or may not be borne out in practice. This is probably more common 159
172 with fertilizers, since they do not have to go through the rigorous labeling system that is required for pesticides. The label for one potassium silicate product (Pro-Tekt by Dynagro) reads as follows: Pro-TeKt (0-0-3) provides increased potassium and silicon to build strong cell walls that protect against piercing insects and invading fungi. This formula improves photosynthesis, stem strength, heat and drought tolerance, plus it helps plants recover quickly from insect damage. Use as a supplement with other fertilizers and amendments. We found ourselves in the unenviable position of supporting this product and growers testimonials based on our observations without actual proof that the material does what is says on the label. The role of the university should be to objectively evaluate products and provide unbiased data as to their effectiveness. Therefore, we decided to scientifically analyze the situation by asking the question Will a potassium silicate supplement improve a plant s ability to resist attack by insects and diseases? Materials and methods We established a replicated study in greenhouses on the Davis campus where various rates of the supplement where applied to potted chrysanthemums and leafminers (Liriomyza trifolii) were used as the bioassay organism. Individual chrysanthemum plants were potted in 6 inch pots, grown for 2 weeks and set up on a greenhouse bench fed by an injector system that allowed a series of pots to receive the following treatments: no potassium silicate (control) or one of 5 doses ranging from 1x = 40 ml/gal to 10x = 400 ml/gal. Pots were randomized on the benches and there were 18 pots (replicates) for each treatment. Plants were watered and fertilized daily with a (% N-P-K) liquid fertilizer solution (Nelson 1998). After 3 weeks, pots were covered with a ventilated cage, and one pair of mated leafminers was released onto each plant. One week later, an additional two mated pairs were released into each cage. Two weeks after this (when plants were in full flower), they were cut at the soil line, placed on the soil surface and the pot with the cut plant was placed in an upright brown paper bag with one small, yellow sticky card at its apex. One week later, all leafminers from the pot (and plant) caught on the sticky card were counted. An identical set of plants was established at the initiation of the trial and leaf samples were taken from these for an analysis of silicon as the trial progressed. In order to compare the various impact of various levels of silicate on leafminers, we used Kruskal-Wallis one-way analysis of variance on ranks. Pairwise comparisons were performed using Dunn s method (Zar, 1999). Results and discussion We observed a significant reduction in leafminers emerging from chrysanthemums treated with potassium silicate at 200 ppm (Figure 1). Higher rates reduced the number of emerging leafminers when contrasted with the control, but this was not statistically significant from the 200 ppm rate. Residual analysis of silicon in chrysanthemum leaves (Figure 2) shows that a level of silicon sufficient to control leafminers was achieved at 200ppm after 6 weeks. At the higher rates, more silicon was found, but this did not significantly reduce emerging leafminers. We are excited about these results and plan to do more work with silicon against pests such as aphids, whiteflies and western flower thrips. In addition, we plan to do examine the compatibility of silicon with natural enemies. While not providing 100% control, this material may enhance a plants ability to withstand insect attack and thus add to the overall IPM arsenal. 160
173 References Cited Epstein, M. 1993: The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. 91: Nelson, P.V. 1998: Greenhouse operation and management. Prentice Hall, Upper Saddle River, NJ. Zar, J.H. 1999: Biostatistical analysis. Prentice-Hall, Upper Saddle River, NJ. 161
174 Number of Emerging Leafminers a b c c ANOVA on SR-transformed data : F = 11.95, P < c c Silicon (ppm) Figure 1. The impact of various concentrations of silicon on leafminers (Liriomyza trifolii) emerging from chrysanthemum plants. Concentration of Potassium Silicate Applied (ppm) Week 6 Week 4 Week % Silicon in Chrysanthemum Leaves Figure 2. Residual accumulation of silicon in chrysanthemum leaves over time under different application rates. 162
175 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Natural parasitism of Liriomyza sativae (Diptera: Agromyzidae) on cucumber under field and greenhouse conditions Yaghoub Fathipour 1, Mostafa Haghani 1, Ali A. Talebi 1, Valiollah Baniameri 2, Abbas A. Zamani 1 1 Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, P. O. Box: , Tehran, fathi@modares.ac.ir. 2 Department of Agricultural Entomology, Plant Pests & Diseases Research Institute, P. O. Box: , Tehran, Iran. Abstract: Liriomyza sativae is one of the most important pests affecting cucumber in Iran and is mainly controlled by applying chemical products. The main purpose of this study was to determine the rate of natural parasitism of L. sativae in the Tehran region, Iran. From September to December 2004 surveys were carried out on cucumbers in sprayed fields and also in sprayed and non-sprayed greenhouses in Varamin and Pishva, near Tehran. A sampling program was designed to estimate the population density and parasitism rate of L. sativae larvae on cucumber leaves. In this study we determined the average percentage and trend of parasitism associated with each parasitoid species. The results showed that in the study area L. sativae larvae were parasitized by three eulophid parasitoid species including Diglyphus isaea, Hemiptarsenus zilahisebessi and Closterocerus formosus. D. isaea was the most frequent species at all three sampling sites, followed by C. formosus and H. zilahisebessi. The overall percentages of parasitism on cucumber caused by these three parasitoids in the field and in sprayed and non-sprayed greenhouses were ( ), ( ) and ( ), respectively. These results indicated that sequential application of insecticides on cucumber led to a significant reduction in the rate of natural parasitism in the field and in sprayed greenhouses. The highest rate of parasitism was observed in non-sprayed greenhouses. It was finally concluded that illogical and inappropriate use of insecticides for controlling pests affecting field and greenhouse crops can reduce the population density and efficacy of natural enemies. Keywords: Liriomyza sativae, parasitism, greenhouse cucumber, Tehran, Iran Introduction Cucumber is the main greenhouse vegetable in Iran and this crop is attacked by various pests including leafminers. The vegetable leafminer, Liriomyza sativae Blanchard, is a major pest of a wide variety of vegetables and ornamental crops throughout the world (Parrella, 1983; Reitz & Trumble, 2002). This fly thrives in suitable warm temperature habitats and especially in temperature-controlled greenhouses where reproduction is almost uninterrupted (Spencer, 1986). For many years, chemical insecticides have been used to control leafminers in Iran and they are still the main tool employed in this and many other countries. However, applications of broad spectrum insecticides have led to a decline in vectors of natural pest control such as parasitism (van Lenteren, 2000). Alternative strategies such as biological control within a framework of Integrated Pest Management (IPM) programs are essential for effectively reducing leafminer populations (Minkenberg & van Lenteren, 1986; Minkenberg, 1989; Heinz & Parrella, 1990). Agromyzid leafminers are found almost everywhere in the world and are economically important pests of many agricultural crops (Spencer, 1986; Nicoli & Pitrelli, 1994). The main damage caused by agromyzids is due to larval feeding, although leaf punctures caused by 163
176 females of the species can also be significant (Johnson et al., 1980; Parrella et al., 1981, 1983; Minkenberg & van Lenteren, 1986; Spencer, 1986). Biological control as part of IPM programs is a reliable and economically viable crop protection strategy for growers of greenhouse crops (van Lenteren & Woets, 1988; Jervis & Copland, 1996). The introduction and support of a number of natural enemies in situations where chemical control is insufficient, impossible or undesired, is also a powerful pest control option (van Lenteren, 1995). Liriomyza species have many natural enemies, particularly in their native area. Forty-one species of parasitoids belonging to four different families have been found in Asia (Murphy & LaSalle, 1999). However, under natural conditions, the parasitism rate is usually low in early crop development and gradually increases as the crop matures (Parrella, 1987). At least 23 species of parasitoids have been used in biological control programs against L. trifolii and L. sativae in Senegal, California, Hawaii, Barbados, Marianas, Tonga, Taiwan and Guam (Johnson, 1993). Using biological pest control on greenhouse crops it should be possible to produce the same crops without the need to use conventional pesticides. This would also imply a cleaner environment and satisfy consumer demands for pesticide-free food and sustainable crop protection (van Lenteren, 2000). Our study was conducted to demonstrate the natural parasitism and seasonal activity of native parasitoids of L. sativae in field and vinyl greenhouse cucumber production at Varamin and Pishva (near Tehran, Iran). Materials and methods From September to December 2004 surveys were carried out on cucumber grown in the field and also in sprayed and non-sprayed greenhouses. Numerous insecticides were applied in the fields and sprayed greenhouses of the study region. A sampling program was designed to estimate the population density and parasitism rate of L. sativae larvae on cucumber leaves. Field and greenhouses were monitored 4-5 times per month throughout the growing season by randomly collecting cucumber leaves. First, we counted the number of larvae on each leaf and then the leaves were placed in separate transparent plastic containers (15 10 cm) covered with a fine mesh screen for ventilation. These leaves, containing leafminer larvae and their parasitoid, were kept in a growth chamber at 25±1 C, 65 ± 5% RH, with a photoperiod of 16:8 (L: D) h. for two or three weeks. We then recorded the number of emerged parasitoids. All parasitoids were isolated in small vials with 75% ethanol for identification. Statistical analysis Statistical analysis was carried out using one-way ANOVA to determine differences between mean numbers of parasitoid species per leaf at each site. The percentage of parasitism was calculated for each parasitoid and also for all parasitoid species. Results and Discussion In the province of Tehran, cucumber is produced in open fields and greenhouses, where it is often attacked by L. sativae. A total of three hymenoptera parasitoid species were collected from the study region as natural enemies of L. sativae. These included Diglyphus isaea (Walker), Hemiptarsenus zilahisebessi (Erdos) and Closterocerus formosus (Westwood). All three species are larval parasitoids belonging to the Eulophidae family and attack many of the hosts that live in confined places (Gauld & Bolton, 1988). No egg or pupal parasitoids were collected during the sampling period. The previously mentioned parasitoids play an important role in controlling the population of L. sativae in non-sprayed and less-sprayed areas. The 164
177 parasitoid complex of leafminers often depends on the predominant Liriomyza species, the host crop and the particular region (Palumbo et al., 1994). Table 1.Mean (±SE) number and percentage of parasitism (emerged adults per leaves) of L. sativae larvae reared on cucumber leaves collected from three different sites at Varamin and Pishva during the period September-December, Sampling site Field Mean Percentage Range Sprayed greenhouse Mean Percentage Range Non-sprayed greenhouse Mean Percentage Range Parasitoid species D. isaea H. zilahisebessi C. formosus 0.60±0.08 a ( ) 0.30±0.04 a 4.04 ( ) 2.23±0.36 a ( ) 0.41±0.05 ab 7.02 ( ) 0.24±0.04 a 3.16 ( ) 1.21±0.21 a 8.98 ( ) 0.23±0.03 b 3.93 ( ) 0.22±0.02 a 3.06 ( ) 1.79±0.24 a ( ) Total 1.24±0.15 B ( ) 0.76±0.09 C ( ) 5.23±0.68 A ( ) Means in the same row followed by the same small letters are not significantly different. Means in the last column followed by the same capital letter are not significantly different. All significant differences identified by the LSD test are at the 0.05 level. The average percentage and trend of parasitism caused by D. isaea, C. formosus and H. zilahisebessi are shown in Table 1 and Figure1. Analysis of variance (ANOVA) showed a significant difference between parasitism rate of the three parasitoid species under field conditions (F=10.77; df=2, 30; P<0.001) and non-significant difference under greenhouse conditions (in sprayed greenhouses: F=1.46; df=2, 24; P=0.252, in non-sprayed greenhouses: F=3.33; df=2, 24; P=0.053). ANOVA also revealed a significant difference between total parasitism rate in the three sampling sites (F=39.12; df=2, 26; P<0.001) (Table 1). Overall rates of parasitism in the cucumber field, and in sprayed and non-sprayed cucumber greenhouses were 21.36% ( ), 10.26% ( ) and 39.31% ( ), respectively. These results indicated that sequential application of insecticides on cucumber led to a significant reduction in the rate of natural parasitism in field and sprayed greenhouses. The highest rate of parasitism was observed in non-sprayed greenhouses. D. isaea was the most frequently found species at all three sampling sites, followed by C. formosus and H. zilahisebessi. D. isaea has already been reported parasitizing L. cicerina Rondani in Iran (Adldoost, 1995). This is a primary ectoparasitoid of various Agromycidae (Diptera). It has also been reported on other hosts, including Lyonetidae (Lepidoptera) and Tephritidae (Diptera) (LaSalle & Parrella, 1991; Zhu et al., 2000). H. zilahisebessi has been collected on L. sativae, L. bryoniae Kaltenbach, L. congesta (Becker), and L. trifolii (Diptera: Agromyzidae). The host range of C. formosus is not restricted to Diptera; other host species include Coleoptera, Hymenoptera, and Lepidoptera. (Noyes, 2004; Talebi et al., 2005). 165
178 a Non-sprayed greenhouse b % Parasitism Sep Oct Nov Sampling Date Sprayed greenhouse Field Dec % Parasitism Sep Oct Nov Sampling Date Dec % Parasitism c % Parasitism d Sep Oct Nov Dec Sep Oct Nov Dec Sampling Date sampling Date Figure.1. Percentage of parasitism of L. sativae populations on cucumber leaves in the field and in sprayed and non-sprayed greenhouses. a) overall parasitism; b) by D.isaea; c) by H. zilahisebess; d) by C. formosus. Our results showed that chemical insecticides are lethal to leafminer parasitoids and that these compounds can reduce the efficiency and population density of parasitoids on cucumber. The population density and parasitism rate of parasitoids of L. sativae larvae in non-sprayed greenhouses were significantly higher than those found in sprayed fields and greenhouses. It was finally concluded that illogical and inappropriate use of insecticides for controlling pests affecting field and greenhouse crops can reduce the population density and efficacy of natural enemies. There are various indigenous natural enemy communities of Liriomyza spp., particularly parasitoids in their native ranges and there is evidence that in pesticide-free areas these can regulate leafminers (Benuzzi & Nicoli, 1988; Benuzzi & Raboni, 1992). They may also vary in type and number in their adventive ranges in continental areas, as invading Liriomyza spp. quickly attract local parasitoids and polyphagous arthropod predators (Heinz & Parrella, 1990). The main reason for the use of biological controls in the 1960s was the presence of resistance to pesticides amongst several key pests in greenhouses (van Roermund et al., 1997). 166
179 Acknowledgments We would like to thank Dr. Saulius Pakalniskis (Institute of Ecology of Vilnius University) for his help in identifying the agromyzid leafminer. This research was supported by the Department of Entomology, Tarbiat Modares University. References Adldoost, H. 1995: Study of population dynamics of rainfed chickpea leafminer in West Azarbaijan. Proc. XII. Plant Protection Congress, University of Tehran, Iran. p Benuzzi, M. & Nicoli, G. 1988: Lotta Biologica e Integrata Nelle Colture Protette. Centrale Ortofrutticola, Cesena. Benuzzi, M. & Raboni, F. 1992: Diglyphus isaea. Inf. Fitopat. 11: Gauld, I. & Bolton, B. 1988: The Hymenoptera. Oxford University Press, New York. 332pp. Heinz, K.M. & Parrella, M.P. 1990: Holarctic distribution of the leafminer parasitoids Diglyphus begini (Hym., Eulophidae) and notes on its life history attacking Liriomyza trifolii (Dip., Agromyzidae) in chrysanthemum. Ann. Entomol. Soc. Am. 88: Jervis, M.A. & Copland M.J.W. 1996: The life cycle. pp In: Insect Natural Enemies., practical approaches to their study and evaluation. Jervis, M. and Kidd, N. (eds) Chapman and Hall, London, UK. Johnson, M.W., Oatman, E.R., Wyman, J.A. & Van Steenwyk. R.A. 1980: A technique for monitoring Liriomyza sativae in fresh market tomatoes. J. Econ. Entomol. 73: Johnson, M.W. 1993: Biological control of Liriomyza leafminers in the Pacific Basin. Micronesica Suppl. 4: LaSalle, J. & Parrella, M.P. 1991: The Chalcidoid parasites (Hymenoptera: Chalcidoidea) of economically important Liriomyza species (Diptera: Agromyzidae) in North America. Proc. Entomol. Soc. Washington. 93(3): Minkenberg, O.P.J.M. 1989: Temperature effects on the life history of the eulophid wasp Diglyphus isaea, an ectoparasitoid of leafminers (Liriomyza spp.), on tomatoes. Ann. Appl. Biol. 115: Minkenberg, O.P.J.M. & van Lenteren J.C. 1986: The leafminers Liriomyza trifolii and L. bryoniae (Diptera: Agromyzidae), their parasites and host plants: a review. Agric. Univ. Wageningen Pap. 86: Murphy, S.T. & LaSalle, J. 1999: Balancing biological control strategies in the IPM of New World invasive Liriomyza leafminers in field vegetable crops. Biocontrol News and Information 20: Nicoli, G. & Pitrelli, P. 1994: Parasitization and predation in Diglyphus isaea. In: Nicoli, G., Benuzzi, M., Leppla, N.C. (Eds.), Proceedings of the Quality Control of Mass Reared Arthropods, Rimini (I), September 13 16, 1993, pp Noyes, J. 2004: Universal chalcidoidea database. The natural history museum [on-line]. Available on the Palumbo, J.C., Mullis, C.H. & Reyes, F.J. 1994: Comparison, seasonal abundance, and parasitism of Liriomyza (Diptera: Agromyzidae) species on lettuce in Arizona. J. Econ. Entomol. 87: Parrella, M.P., Robb, K.L. & Bethke J. 1981: Oviposition and pupation of Liriomyza trifolii (Burgess) (Diptera: Agromyzidae). Proc. IFAS Industry Conference on biology and control of Liriomyza leafminers. Lake Buena Vista, Fla. pp Parrella, M.P., 1983: Intraspecific competition among larvae of Liriomyza trifolii (Diptera: Agromyzidae): effects on colony production. Environ. Entomol. 12:
180 Parrella, M.P., Rob, K.L. & Bethke. J. 1983: Influence of selected host plants on the biology of Liriomyza trifolii (Diptera: Agromyzidae). Ann. Entomol. Soc. Am. 76: Parrella, M.P., 1987: Biology of Liriomyza. Annu. Rev. Entomol. 32: Reitz, S.R. & Trumble, J.T. 2002: Intraspecific and interspesific differences in two Liriomyza leafminer species in California. Entomol. Exp. Appl. 102: Spencer, K.A.1986: Manual of the Agromyzidae (Diptera) of the United States. United States Department of Agriculture. Agricultural Handbook, Nº 638. Talebi, A.A., Asadi, R., Fathipour, Y., Kamali, K., Moharramipour, S. & Rakhshani, E. 2005: Eulophid parasitoids of agromyzid leafminers genus Liriomyza (Dip.: Agromyzidae) in Tehran, Iran. IOBC/WPRS Bull. 28(1): van Lenteren, J.C. & Woets, J. 1988: Biological and integrated pest control in greenhouses. Annu. Rev. Entomol. 33: van Lenteren, J.C. 1995: Integrated pest management in protected crops. In: Dent, D. (ed.), Integrated Pest Management. Chapman & Hall, London, pp van Lenteren, J.C. 2000: A greenhouse without pesticides: fact or fantasy? Crop Prot. 19: van Roermund, H.J.W., van Lenteren, J.C. & Rabbinge, R. 1997: Biological control of greenhouse whitefly with the parasitoid Encarsia formosa on tomato: an individual-based simulation approach. Biol. Control. l 9: Zhu, C.D., LaSalle, J. & Huang, D.W. 2000: A Review of the Chinese Diglyphus Walker (Hymenoptera: Eulophidae). Oriental Insects, 34:
181 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp First data on the population dynamics of aphidophagous syrphids in Mediterranean pepper greenhouses Ana Pineda, Mª Ángeles Marcos-García University of Alicante, Research Institute of Biodiversity CIBIO, San Vicente del Raspeig, Spain Abstract: Aphids are an important pest in pepper greenhouses and one group of their natural enemies are hoverflies (Diptera, Syrphidae). This work presents the first results about the main aphidophagous syrphid species found in Mediterranean greenhouses in Spain and certain aspects concerning the monitoring of their mobility from and to greenhouses. Eupeodes corollae was the first species to emerge from a two-year sampling of immature stages. It was followed by Episyrphus balteatus and finally by Sphaerophoria rueppellii. A modified Malaise trap was tested and revealed an adult syrphid population dynamic comparable with data obtained from direct sampling of immature stages. We suggest placing these modified Malaise traps outside greenhouses in order to understand what proportion of adult syrphids go inside greenhouses. This could be a method for monitoring syrphid populations in a given area, for both outdoor and protected crops. Keywords: Syrphids, pepper greenhouses, aphid natural enemies, Malaise trap Introduction After Andalusia, the southeast of Spain is the second largest region in terms of the total surface area of sweet pepper crops with 75% of this surface growing under cover (MAPA, 2004). In sweet pepper greenhouses, aphids are the most destructive pest. They have not traditionally been a major concern because of the presence of Pirimicarb, a selective aphicide (Ramakers, 2004) and also due to the great diversity of natural enemies frequent in these crops. Larvae of aphidophagous hoverfly (Diptera, Syrphidae) are aphid predators and their effectiveness has been amply demonstrated in several works (Chambers & Adams, 1986). They are one of the primary natural enemies of aphids, together with coccinellids, chrysopids and parasitoid hymenopterans, and artificial releases of all of them are being done. Before those introductions are carried out on a crop, it is vital to establish which species are the most abundant in the area and their specific phenology in order to understand which species to introduce during each period of the year. In Mediterranean areas, greenhouse isolation conditions are less controlled than in northern Europe, because of their flexible polyethylene covers, which are less hermetic than more rigid and glass versions. Furthermore, to prevent extreme temperatures and humidity, the sides and zenithal windows of greenhouses are generally left open. Most previous studies about syrphids in European crops have been performed in northern and central Europe. To the best of our knowledge, the results of this work are therefore the first relating to Mediterranean greenhouses. The monitoring of natural enemies is essential for estimating their presence and effectiveness (Urbaneja & Jacas, 2004), and effective sampling methods are required to minimise application times and costs. Malaise traps offer an effective method for capturing adults of Diptera (Cecidomyiidae and Syrphidae) and Hymenoptera (Braconidae and Aphelinidae), groups to which some of the main aphid natural enemies belong. On the other hand, Malaise traps have not previously been used in agro-ecosystems, due both to their 169
182 complexity and to the high number of captures, which could potentially reduce natural enemy populations. Platt et al (1999) designed a simpler modified Malaise trap for monitoring crop insects. In this paper, we present the first results of its use to monitor syrphids. The objectives of this study were to: 1) identify the most important aphidophagous syrphid species present in pepper greenhouses in the southeast of Spain and study their population dynamics; 2) determine the relation between syrphid populations outside and inside greenhouses; 3) test the effectiveness of a modified Malaise trap for monitoring syrphid populations. Materials and methods Experimental site This work was carried out in commercial greenhouses growing sweet pepper crops that belonged to the agricultural cooperative SURINVER S.C. located in Pilar de la Horadada (southeast Spain). Sampling was carried out in 2004 and 2005, during the pepper culture (January-July). In 2004, 20 greenhouses were selected, and in 2005 sampling took place in 28 greenhouses. All these greenhouses were of the Almería type and had total surface areas of between 1000m 2 and 12000m 2. External isolation was very limited, mainly due to side openings which were open most of the time. Sampling of immature stages After the pepper crop was planted, sampling was performed every two weeks until the first syrphids were observed. From this moment on, sampling was carried out on a weekly basis till the end of the culture. The sampling methodology started with the location of aphid focus, and the determination of a severity level, on a scale from 1 to 4. Counts were taken at random around the focus, through the rigorous observation of 200 leaves (sampling unit) of pepper plants per greenhouse. All leaves with syrphid eggs, larvae, or pupae were collected in rearing cages (21 x 15 x 9 cm), as were leaves with such high numbers of aphids that they did not permit rigorous observation in situ. Average leaf size was similar to those of the central leaves. Leaves on the terminal apex were given a value of one and those on the bottom part of the plant were given double value. The Syrphids were reared in a climatic room until the emergence of adults, which were identified to species level. Sampling of adult hoverfly with modified Malaise traps In 2005, a complementary study of adults was carried out using modified Malaise traps (Figure 1). The design was based on a trap described by Platt et al. (1999), but it had an aluminium structure and a smaller collecting container (100 ml) into which 70% ethanol was introduced to kill and conserve the insects. The experimental design consisted of placing 1 trap inside a greenhouse and 1 trap outside, in a total of 4 greenhouses with similar characteristics that were separated by more than 1 km. Collection was carried out every 2 weeks from week 6 until week 12, when the first syrphid was caught. Thereafter, collection was carried out on a weekly basis until week 27, when the crop was retired. Syrphids were subsequently identified to species level in the laboratory. Results and discussion Phenology of the main aphidophagous species In 2004, the most abundant species of the aphidophagous hoverfly larvae found were Eupeodes corollae (Fabricius, 1794) (32.07%), Episyrphus balteatus (De Geer, 1776) (38.83%) and Sphaerophoria rueppellii (Wiedemann, 1830) (27.57%). These species constituted 98.47% of a total of 1244 individuals collected as immature stages from aphid 170
183 colonies. The population dynamics of these species in 2004 (Figure 2a), show that hoverflies did not appear in the greenhouses before week 15 (5 th April), although aphid colonies were present from week 3 (15 th January). E. corollae was the first species observed, with a population maxim on week 15 (5 April). While its population decreased, E. balteatus population increased with a peak occurring in week 21 (20 th May), this same trend was followed by the population of S. rueppellii, which was most abundant during weeks (June). The last observation of hoverflies was in week 26, and the last aphid colony was seen in week m 0.52 m 0.52 m m Figure 1. Graphic of the modified Malaise trap used in this work and based on the trap designed by Platt et al. (1999). Figure 2. Population dynamics of the main aphidophagous syrphid species observed as immature stages during pepper culture, in 2004 (a) and 2005 (b). The sampling unit consisted of 200 leaves per greenhouse. In 2005, the total number of immature individuals observed was 225 hoverfly, 80% fewer than observed in The respective percentages of the three main species also changed in 2005; E. balteatus was less abundant with only 0.89% of the overall total, followed by E. corollae with 31.11%. The most abundant species was S. rueppellii with an abundance of 68%. We can observe (Figure 2b) that the population peaks for E. corollae and for S. rueppellii were similar to results for The greatest difference in the phenology of 171
184 these species for the two consecutive years was observed for E. balteatus, whose population also peaked in week 21 in 2005, but with just 0.33 ± 0.33 individuals per leaf, for the 200 leaves observed. The changes observed in 2005 could have been mainly due to the more arid conditions was the driest year in Spain since climatic measurements began (Instituto Nacional de Meteorología, 2005) and the drought could have directly affected the syrphid community structure. This is the most likely explanation for the decrease in the E. balteatus population and the corresponding increase and relative abundance of S. rueppellii, a well adapted species. Dry conditions can also have an effect on aphid predators and indirectly on the aphidophagous syrphid community, both of which are factors directly related to aphid abundance. With the same sampling effort, only half the number of aphid colonies were observed in 2005 and most of these belonged to the lowest level of severity. It might therefore be expected that a lower amount of larvae can be observed. The phenology of both years suggests a time succession for the three main aphidophagous syrphid species. This seems to be related to climatic conditions because E. corollae and E. balteatus are also the most frequent species found on central European crops (Cowgill, et al., 1993). On the other hand, S. rueppellii is a typical Mediterranean species and, as already reported, the most abundant when temperatures and drought conditions are extreme. These results suggest when artificial syrphid releases should be made, and also which species are most appropriate, although only E. balteatus is currently commercialized. Comparison between inside and outside greenhouse populations The total number of syrphid adults caught and studied in Malaise traps placed outside and inside greenhouses did not reveal any significant differences between the 4 sampling locations (H=3.33; df=3; P= 0.34). When we compared the proportion of syrphids that went inside the greenhouse with those captured outside, we observed (Figure 3) a variation of between 11.11% and 20%, with the outdoor populations being greater than those inside. These data are the first relating to syrphid presence in Mediterranean greenhouses and suggest that natural populations of syrphids could be established inside greenhouses. This movement of insects is further proof that the semi-open structure of Mediterranean greenhouses constitutes an intermediary situation between open fields and conventional glass greenhouses. Typical control strategies for outdoor crops could therefore also be effective in these greenhouses. These could include the introduction of floral resources in order to attract natural populations. Table 1. Percentage of samples from a two-year sampling, with a severity level from 1 to 4, with presence or absence of immature syrphid stages, and with different aphid species (Mp, Myzus persicae; Me, Macrosiphum euphorbiae; Ag, Aphis gossypii; As, Aulacorthum solani). Number of Severity level Syrphid presence Aphid species Year samples yes no Mp Me Ag As
185 Figure 3. Syrphid abundance based on captures from modified Malaise traps placed inside and outside greenhouses. Bars represent the mean (±SE) number of adult syrphids per trap. Numbers above bars show the percentage of adults caught inside compared to those caught outside. The population dynamics of the three main aphidophagous species observed from sampling with Malaise traps inside greenhouses in 2005 (Figure 4a), showed a similar pattern to that obtained from direct sampling of immature hoverfly (Figure 2b). Although E. balteatus was never caught by the modified Malaise trap, this species is often caught by traditional Malaise traps. E. balteatus was observed in small numbers through immature stage sampling in 2005 and we assume that its relatively small total population explains why it was not caught in the traps. Effectiveness of modified Malaise traps The modified Malaise traps yielded maximum rates of capture, with a mean of 1.25 ± 1.25 individuals being caught per greenhouse: this should not affect aphid control. Moericke traps have traditionally been used to catch syrphids in agrarian ecosystems, but they do not seem to be very effective in greenhouses, with capture rates being relatively low (1 adult in 3 sampling years in the same area) (J.A. Sánchez, personal communication). The characteristics of the modified Malaise trap were used to devise an effective method for monitoring the populations of the main syrphid species in Mediterranean greenhouses. This is something that had not been previously achieved. In contrast, the fact that the number of syrphid species caught inside greenhouses was equal to that caught outside and with a similar population dynamics (Figure 4b) was significant, as was the fact that we were able to discover the proportion between syrphid adults caught inside and outside greenhouses. With data obtained from Malaise traps placed outside greenhouses, it should therefore be possible to estimate the population inside. This monitoring approach would, to a certain extent, affect populations of natural enemies in Mediterranean greenhouses; so a larger amount of data is needed to draw more reliable conclusions. 173
186 Figure 4. Population dynamics of the main aphidophagous syrphid species captured as adults by the modified Malaise trap during pepper culture in Traps were placed inside (a) and outside (b) greenhouses. Acknowledgements This work has been supported by the Ministerio de Ciencia y Tecnología, INIA (Nº RTA C2). A. P. was supported by a student grant from the Generalitat Valenciana, Conselleria d Empresa, Universitat i Ciència (CTBPRB/2004/081). References Chambers, R.J. & Adams, T.H.L. 1986: Quantification of the impact of hoverflies (Diptera: Syrphidae) on cereal aphids in winter wheat: an analysis of field populations. J. App. Ecol. 23: Cowgill, S.E., Wratten, S.D. & Sotherton, S.W. 1993: The effect of weeds on the numbers of hoverfly (Diptera: Syrphidae) adults and the distribution and composition of their eggs in winter wheat. Ann. Appl. Biol. 123 (3): Instituto Nacional de Meteorología September 5. El año más seco. Accessed 2006 January 10. MAPA 2004: Anuario de estadística agroalimentaria, Ministerio de Agricultura, Pesca y Alimentación: p Platt, J., Caldwell, J.S. & Kok, L.T. 1999: An easily replicated, inexpensive Malaise-type trap design. J. Entomol. Sci. 34 (1): Ramakers, P.M.J. 2004: IPM program for sweet pepper. In: Biocontrol in protected culture, eds. Heinz, Van Driesche and Parrella: Urbaneja, A. & Jacas, J. 2004: Métodos de muestreo de enemigos naturales. Phytoma España, 164:
187 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Evaluation of two aphid parasitoids as candidates for biocontrol of aphid pests in protected cultivation in Brazil Vanda H. P. Bueno 1, Marcus V. Sampaio 2, Joop C. van Lenteren 3 Bruno F. De Conti 1, Robson J. Silva 1, Sandra M. M. Rodrigues 4, Ariana B. Carnevale 1 1 Federal University of Lavras, Department of Entomology, P.O.Box 3037, Lavras, MG, 2 Federal University of Uberlandia, Faculty of Agriculture, Uberlandia, MG, Brazil; 3 Laboratory of Entomology, Wageningen University, Wageningen, The Netherlands; 4 Embrapa-Algodão, Primavera do Leste-MT, Brazil. Abstract: Several aphid species can be important pests in greenhouse vegetables and ornamentals and biological control is a desirable option to control aphids in protected cultivation in Brazil. To select and evaluate the local aphid parasitoids, Aphidius colemani and Lysiphlebus testaceipes, tests are performed involving host suitability and preference, and evaluation of biological parameters of both species at different temperatures. Host preference indicated that A. colemani and L. testaceipes prefer A. gossypii above M. persicae as a host. Both aphid hosts are suitable for the development of both parasitoid species. However, A. gossypii is a more suitable host than M. persicae for L. testaceipes. A temperature of 22ºC for A. colemani and 25ºC for L. testaceipes are the most adequate for reproduction and mass rearing. Host quality for A. colemani was decreasing as follows: M. persicae = R. maidis > S. graminum > A. gossypii. The development time, rate of parasitism and longevity of L. testaceipes were decreasing from S. graminum = R. maidis > M. persicae. Honey is an important food and increases the longevity of A. colemani, and water is a good complement to the diet of this parasitoid. The two parasitoid species are good candidates to use as biocontrol agents in protected crops in Brazil. Key words: Lysiphlebus testaceipes, Aphidius colemani, host suitability, host preference, food sources Introduction Aphids, particularly, the cotton aphid Aphis gossypii Glover and the green peach aphid Myzus persicae (Sulzer), play an important role as pests in several crops under protected cultivation in Brazil (Bueno et al. 2003; Bueno, 2005). These aphids have a wide plant host range, and their rapid population growth and resistance to pesticides make it very difficult to obtain good and long lasting control. Thus, it is necessary nowadays to search for new control methods to aphids, and biological control is an attractive option. Several biological control projects in Brazil have used native natural enemies that may be better adapted to local conditions. The aphid parasitoids Aphidius colemani Viereck and Lysiphlebus testaceipes (Cresson) are the most frequent aphid natural enemies occurring under greenhouse conditions in Brazil. According to Rodrigues et al. (2005) a satisfactory control of A. gossypii in two chrysanthemum cultivars was achieved when releasing the parasitic wasp L. testaceipes. Also, these aphid parasitoids can invade spontaneously the greenhouse and exert natural control (Bueno et al., 2003). However these parasitoids attack more than one aphid species and certain species are not accepted for oviposition by aphidiids. Therefore we need to study behavior of the parasitoids by studying several steps in the host selection process, which explains the final parasitization rates of the host population. Also, the temperature directly affects the development time and 175
188 behavior of the insects. The adaptability to climate conditions is a key point for the success of mass rearing and parasitoid establishment in biological control programs. A careful selection and evaluation process of natural enemies to minimize risks of failure of pest control programs in protected cultivation is very important. As preliminary work to develop mass rearing and use of the parasitoids L. testaceipes and A. colemani as biological control agents of aphid pests, this work aimed to evaluate the suitability of aphid hosts for A. colemani and L. testaceipes and the preference of both species for M. persicae and A. gossypii. This study also aimed at determining the development and parasitism of A. colemani and L. testaceipes on A. gossypii as a host at different temperatures, and determination of the longevity of A. colemani in different food sources. Material and methods Preference of L. testaceipes and A. colemani in non-choice and choice tests for M. persicae and A. gossypii Petri dishes (5cm) with a 1 cm layer of agar-water medium and 4 cm sweet pepper leaf discs with 20 M. persicae nymphs and/or A. gossypii were used. A single female of L. testaceipes or A. colemani was released on each disc and observations were performed under a stereoscopic miscroscope during 15 minutes. Suitability of aphid species to L. testaceipes and A. colemani The species of aphid were offered to the parasitoids separately. Small cages with a host plant leaf and 2 nd and 3 rd instar nymphs were used. One female parasitoid was released into the cage and removed after 2h. The aphids were transferred to Petri dishes (5cm) containing a 1% solution of water/agar and a leaf disc of the host plant. Development and parasitism of L. testaceipes and A. colemani on A. gossypii The experiments were carried out in controlled environmental chambers at 15, 20, 25 and 30±1ºC, RH 60±10% and 10 hr photophase. Third instar nymphs of A. gossypii attacked once by L. testaceipes were kept individually in Petri dishes (6cm diameter) on a leaf disc (2cm diameter) of chrysanthemum (Yellow Snowdon cultivar) on a layer of agar-water (1%). The experiments with A. colemani were carried out in controlled environmental chambers at 16, 19, 22, 25, 28 and 31±1ºC, RH 70±10% and 12 hr photophase. Second instar nymphs of A. gossypii attacked once by the parasitoid were kept individually in glass tubes (2.5cm x 8.5cm) containing a leaf disc of cucumber (2cm) and 1% water-agar solution. Effect of different food sources on the longevity of A. colemani The experiment was carried out in controlled environmental chamber at 22±1ºC, RH 70±10% and 12 hr photophase. Males and females of A. colemani reared on A. gossypii were submitted to different food sources: no food, water, pure honey, diluted honey at 50%, pure honey + water, diluted honey 50% + water. Results and discussion Occurrence of aphid parasitoids and aphid hosts in greenhouses and its neighborhood in the southeast region in Brazil The parasitoid A. colemani was found having as aphid hosts Aphis craccivora, A. gossypii, A. spiraecola, Eucarazzia elegans, Myzus persicae, Nasonovia ribisnigri, Rhopalosiphum padi, Schizaphis graminum, Sitobion avenae and Toxoptera aurantii. L. testaceipes was found parasitizing Aphis coreopsidis, A. craccivora, A. gossypii, Aphis nerii, A. spiraecola, M. persicae, Rhopalosiphum maidis, R. padi, S. graminum and Toxoptera citricidus. According to Sampaio et al. (2005a), the parasitoid L. testaceipes was found as a potential 176
189 competitor of A. colemani in aphids of the tribe Aphidini, and responsible for the nonoccurrence of A. colemani in certain periods of the year in southeast region in Brazil. Preference of A. colemani and L. testaceipes of M. persicae and A. gossypii Host acceptance by A. colemani of M. persicae and A. gossypii in non-choice tests demonstrate that there was no difference in the first visit and in the number of encounters between the parasitoid and the two species of aphids. The same was found in choice tests. That A. colemani prefers A. gossypii above M. persicae can be concluded from the higher number of attacked A. gossypii by the parasitoid (oviposition) and the higher number of parasitoid larvae found in this aphid. Host acceptance by L. testaceipes of M. persicae and A. gossypii in non-choice test showed that the average number of stings with the ovipositor (short touches), ovipositions (long touches) and the total number of larvae found after host dissection were higher in A. gossypii than in M. persicae. In the choice tests a difference between the numbers of encounters with the two host species was not observed. But the other parameters (probes, oviposition and number of larvae) were different in both hosts; they were higher for A. gossypii. These results indicate that A. colemani and L. testaceipes prefer A. gossypii above M. persicae as a host. Suitability and quality of aphid species for L. testaceipes and A. colemani Once the host is accepted for oviposition and an egg is deposited, the successful development of the parasitoid into an adult depends on host suitability. The parasitism rates of L. testaceipes were higher in A. gossypii (44.2%) than in Myzus persicae (6.7%). The percentage of emergence of L. testaceipes in M. persicae (100%) and in A. gossypii (92.6%) was the same, also the development time, 9.0 and 8.8 days, respectively, was the same. The longevity of L. testaceipes was shorter in M. persicae (3.9 days) when compared to it on A. gossypii (5.5 days). Rodrigues & Bueno (2001) found parasitism rates of 76% for S. graminum and 56% for A. gossypii for L. testaceipes. Parasitism rates of A. colemani were 52% and 75% for M. persicae and A. gossypii, respectively. No difference was found for the percentage of emergence, being 86% in A. gossypii and 81% in M. persicae. When the effect of competition for food among M. persicae aphids on the development time of males and females of A. colemani was evaluated, Sampaio et al. (2002) showed that increased competition for food in hosts does not result in differences in development time of both sexes, but it can extend the development time of the parasitoid. The aphid A. gossypii is more accepted than M. persicae by both parasitoids, and M. persicae is more accepted by A. colemani than L. testaceipes. We observed that both aphid hosts are suitable for the development of both parasitoids, and that A. gossypii is more suitable than M. persicae for L. testaceipes. The suitability of A. gossypii and the low suitability of M. persicae as host for L. testaceipes has also been show by van Steenis (1993). The suitability of other tested aphids showed that host quality for A. colemani was decreasing from M. persicae = R. maidis > S. graminum > A. gossypii. Larger hosts showed better quality than smaller ones for size and development of A. colemani. Concerning to L. testaceipes we found a development time, parasitism rate and longevity decreasing on S. graminum = R. maidis > M. persicae. Rodrigues et al. (2003b) stated that L. testaceipes has a high potential of population growth on S. graminum as a host, having a finite rate of increase of 1.67 females per day, a mean length of a generation of days and a time to duplicate the population of 1.35 weeks. When tested the effect of storage at 5ºC of S. graminum mummies parasitized by L. testaceipes, Rodrigues et al. (2003a) found no effect on reproductive capacity of the parasitoid emerging after exposition periods of 4, 6, 8 and
190 days. One-day-old mummies of S. graminum can be stored at 5ºC for a period of 6 days without fitness reduction of L. testaceipes. Development and parasitism of L. testaceipes and A. colemani in A. gossypii The development time of L. testaceipes was 26.9, 14.8, 11.3 and 12.2 days at 15, 20, 25 and 30ºC, respectively. Parasitism rates were 76, 68, 65 and 40% at 15, 20, 25 and 30ºC, and the emergence rates were 80, 61, 62 and 14% at these temperatures. The sex rate of L. testaceipes was 0.35, 0.43, 0.45 and 0.54 for the temperatures 15, 20, 25 and 30ºC, respectively. The females showed longevities of 12.3, 6.0, 3.5 and 2.0 days and the males 10.4, 5.4, 3.1 and 2.2 days at 15, 20, 25 and 30ºC. The combination of a low developmental time (11.3 days) and parasitism and emergency higher than 60% occurring at 25ºC, indicate that this temperature seems best for mass rearing and establishment of L. testaceipes. Rodrigues et al. (2001) found that L. testaceipes is capable of covering distances of at least 3.35 m during 24 hours from an open rearing system to parasitize A. gossypii in sweet pepper in greenhouse. The development time of A. colemani was 19.4, 16.2, 12.6, 10.5 and 10.7 days, respectively at 16, 19, 22, 25 and 28ºC, and decreased with the increase of the temperature in an interval of 16 and 25ºC. The parasitism rates (93.3, 91.7, 91.4, 86.5 and 73.3%) and the emergence (89.7, 94.2, 94.5, 86.4 and 26.8%) of the parasitoid A. colemani, and its longevity (21.3, 18.4, 14.1, 8.3 and 1.7 days, respectively, at 16, 19, 22, 25 and 28ºC) decreased with the increase of the temperature. There was no mummy formations at 31ºC. The temperature of 22ºC was optimal for the development time of A. colemani. The threshold temperature for development was 5.94ºC and the thermal constant was 200 day-degrees for A. colemani. When the response to temperature of A. colemani from three climatic regions of Minas Gerais, Brazil was evaluated, Sampaio et al. (2005b) found that the more suitable temperature for the development of this parasitoid from Lavras was higher than those for individuals from Juramento and Sao Gotardo. The individuals from Lavras showed highest emergence at 28ºC, and this demonstrated the existence of individuals of A. colemani with tolerance to higher temperatures. The authors concluded that these results open news perspectives for the use of different strains of A. colemani in biological control of A. gossypii in protected cultivation in Brazil. The parasitoid A. colemani showed higher longevity having as food pure honey + water and honey 50% + water (P 0.01). The longevities of male and female of A. colemani were significantly different when the parasitoid was feeding on pure honey (4.2 days; 6.7 days) and diluted honey 50% (4.5 days; 6.9 days) (P 0.01) (Table 1). In conclusion, the aphids M. persicae and A. gossypii are suitable for the development of both parasitoid species. However, A. gossypii is more suitable for L. testaceipes and M. persicae for A. colemani. The parasitoids L. testaceipes and A. colemani prefer A. gossypii both in non-choice and in choice tests. The temperature of 22 and 25ºC, respectively for A. colemani and L. testaceipes, are the most adequate temperatures for reproduction and mass rearing. Honey is important food to increase longevity of A. colemani, and water a good complement to the diet of the parasitoid. Water together with honey (pure or diluted) promotes a high longevity of A. colemani. Host suitability and host preference together demonstrate that parasitoids respond to a series of steps, which influence the parasitoid-host-interaction. As an augmentation approach to biological control in greenhouses in Brazil becomes more important, there are increased interests in the factors involved in successful parasitism, in the hope that these can be manipulated for more effective utilization of parasitoids in protected cultivation. It would be convenient if the parasitoids used for biological control of one aphid species also could be used in biological control of other important aphid species. The evaluation of local or native 178
191 species as candidates for biological control is a goal in Brazil, and L. testaceipes and A. colemani seem to be promising and suitable candidates to use them as biological control agents. Table 1. Longevity (days) of A. colemani on different food sources Food sources Longevity of A. colemani Female Male Without food 1.13 ac 1.50 ac Water 1.07 ac 1.67 ac Pure honey 6.71 ab 4.23 bb Diluted honey 50% 6.93 ab 4.58 bb Pure honey + Water aa 9.67 aa Honey 50% + Water 9.36 aa aa Means followed by the same letter in the columns and rows do not differ from one another by the Tukey Test (P 0.01). Acknowledgements We would like to thank to CNPq for grants to the authors and for financial support of these studies. References Bueno, V.H.P., van Lenteren, J.C., Silveira, L.C.P. & Rodrigues, S.M.M. 2003: An overview of biological control in greenhouse chrysanthemums in Brazil. IOBC/WPRS Bull. 26 (10): 1-5. Bueno, V.H.P. 2005: Controle biológico de pulgões ou afídeos-praga em cultivos protegidos. Informe Agropecuário 26: Rodrigues, S.M.M. & Bueno, V.H.P. 2001: Parasitism rates of Lysiphlebus testaceipes (Cresson) (Hym.: Aphidiidae) on Schizaphis graminum (Rond.) and Aphis gossypii Glover (Hym.: Aphididae). Neotrop. Entomol. 30(4): Rodrigues, S.M.M., Bueno, V.H.P. & Bueno-Filho, J.S.S. 2001: Desenvolvimento e avaliação do sistema de criação aberta no controle de Aphis gossypii Glover (Hem.: Aphididae) por Lysiphlebus testaceipes (Cresson) (Hym.: Aphidiidae) em casa de vegetação. Neotrop.l Entomol. 30(3): Rodrigues, S.M.M., Bueno, V.H.P. & Sampaio, M.V. 2003a: Armazenamento de múmias de Schizaphis graminum (Rondani) parasitadas por Lysiphlebus testaceipes (Cresson) (Hymenoptera, Aphidiidae) em baixa temperatura. Bol. San. Veg. Plagas 29: Rodrigues, S.M.M., Bueno, V.H.P. & Sampaio, M.V. 2003b: Tabela de vida de fertilidade de Lysiphlebus testaceipes (Cresson, 1880) (Hymenoptera, Aphidiidae) em Schizaphis graminum (Rondani, 1852) (Hemiptera, Aphididae). Rev. Bras. Entomol. 47 (4): Rodrigues, S.M.M., Bueno, V.H.P. & Sampaio, M.V. 2005: Efeito da liberação inoculativa sazonal de Lysiphlebus testaceipes (Hym.: Aphidiidae) na população de Aphis gossypii (Hem.: Aphididae) em cultivo de crisântemo em casa de vegetação comercial. Bol. San. Veg. Plagas 31(2):
192 Sampaio, M.V., Bueno, V.H.P., Rodrigues, S.M.M. & Soglia, M.C.M. 2002: The effect of competition among aphids on development time of Aphidius colemani Viereck (Hymenoptera: Aphidiidae), p In: Simon, J.C.; Dedryver, C.A.; Rispe, C.; Hullé, M. (eds.), Aphids in a new millennium. INRA Editions, France, 549p. Sampaio, M.V., Bueno, V.H.P., De Conti, B.F., Rodrigues, S.M.M. & Soglia, M.C.M. 2005a: Co-occurrence of Aphidius colemani and other aphid parasitoids in some localities of Southeastern Brazil. IOBC/WPRS Bull. 28(1): Sampaio, M.V., Bueno, V.H.P., Rodrigues, S.M.M. & Soglia, M.C.M. 2005b: Resposta à temperatura de Aphidius colemani Viereck (Hymenoptera, Braconidae, Aphidiinae) originário de três regiões climáticas de Minas Gerais, Brasil. Rev. Bras. Entomol. 49(1): van Steenis, M.J. 1993: Suitability of Aphis gossypii Glov., Macrosiphum euphorbiae (Thom.), and Myzus persicae Sulz. (Hom.: Aphidiidae) as host for several aphid parasitoid species (Hym.: Braconidae). IOBC/WPRS Bull. 16(2):
193 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Host stage preference by Aphidius colemani and Aphidius matricariae (Hymenoptera: Aphidiidae) as parasitoids of Aphis gossypii (Hemiptera: Aphididae) on greenhouse cucumber Ali Asghar Talebi 1, Abbas Ali Zamani 1, Yaghoub Fathipour 1, Valiollah Baniameri 2, Katayun Kheradmand 1 and Mostafa Haghani 1 1 Department of Entomology, Faculty of Agriculture, Tarbiat Modares University, P. O. Box: , Tehran, 2 Department of Agricultural Entomology, Plant Pests & Diseases Research Institute, P. O. Box: , Tehran, Iran. Abstract: Aphidius colemani Viereck and A. matricariae (Haliday) (Hymenoptera: Aphidiidae) have a high potential as effective biological control agents for the cotton aphid, Aphis gossypii Glover in greenhouse crops. In this study, the host stage preferences of A. colemani and A. matricariae were determined in a growth chamber at 25 C, 65 % relative humidity and during a photoperiod of 16L: 8D hours. In choice and no-choice experiments, individual females of A. colemani and A. matricariae were offered first, second, third and forth instar nymphs of A. gossypii, as well as adults. The results showed that there were significant differences between the mean percentages of parasitized hosts at different stages (P<0.05). Under no-choice conditions, the highest value was registered for parasitism of third instar nymphs of A. gossypii by A. colemani and of third and fourth instar nymphs by A. matricariae. In choice experiments, females of A. colemani and A. matricariae consistently preferred third and fourth instar nymphs of A. gossypii. Mean percentages for parasitism by A. colemani and A. matricariae on the third nymphal instar of A. gossypii were 23.6 ± 3.69 and 18.4 ± 2.42, respectively. The progeny body size of A. colemani and A. matricariae at emergence increased with host stage at the time of parasitization. The greenhouse release of parasitoids would be best timed to coincide with the period when third and fourth nymphal instars of cotton aphid are most abundant. Keywords: Aphidius colemani; Aphidius matricariae; host stage preference; cotton aphid; greenhouse cucumber Introduction The cotton aphid, Aphis gossypii Glover (Homoptera: Aphididae) is a cosmopolitan, polyphagous species widely distributed in tropical, subtropical and temperate regions (Kersting et al., 1999). In many regions, A. gossypii is one of the most important greenhouse crop pests (Havelka, 1978; Kersting et al., 1999). Chemical control has generally been used as the major tool for controlling aphids on cucumbers (Parrella et al., 1999). However, insecticide resistance and growing concerns about environmental hazards due to frequent use of insecticides have prompted the development of biological control of aphids. Biological control of aphids is increasingly applied on greenhouse crops (Parrella et al., 1999; van Lenteren and Woets, 1988). The two aphid parasitoids, Aphidius colemani Viereck and Aphidius matricariae (Haliday), have been considered good candidates for biological control of aphids such as A. gossypii (Bennison, 1992; van Steenis, 1993; Goh & Yoo, 1997), and their biology and use as biological control agents have been extensively reviewed (Goh & Yoo, 1997; Toussiddou et al., 1999; van Lenteren & Woets, 1988). Aphid parasitoids may show considerable potential 181
194 as biological control agents for A. gossypii since A. colemani parasitism rates of up to 70% have been reported in June surveys of melon greenhouses in South Korea (Goh & Yoo, 1997). Understanding the host stage preferences of parasitoids is a precondition for successful biological control programs, as it helps mass-rearing efforts and may also help to explain observed outcomes in the field. Both A. colemani and A. matricariae attack different stages of A. gossypii. It is not however known which stage is preferred for oviposition. This knowledge is essential for the mass-rearing of these parasitoids. We therefore sought to determine the stage range during which A. gossypii are vulnerable to attack from A. colemani and A. matricariae under no-choice and choice conditions. Materials and methods Rearing methods and experimental conditions Insects were reared and experiments performed in a growth chamber at 25±1 C, 65±5% Relative Humidity and with a photoperiod of 16L: 8D hours. The cotton aphid and its parasitoids, A. colemani and A. matricariae, were originally collected from cucumber greenhouses in Tehran, Iran, in June Aphids were reared on cucumber (Cucumis sativus cv. Negin). Seedlings of C. sativus were grown to the 4-5 leaf stage in a mixture of sand (33%), clay (33%) and peat moss (33%) in 25cm pots. Aphidius colemani and A. matricariae were separately reared on A. gossypii in cm cages. Colonies of A. colemani and A. matricariae were replenished with field-collected individuals in spring and autumn. Aphids and parasitoids were reared on cucumber in the laboratory for several generations before they were used in the experiments. No-choice test Cucumber leaves containing 50 aphids at various stages (first, second, third and fourth instar nymphs and newly emerged adults) were placed in an experimental arena made of translucent glass containers ( cm). Recently-mated A. colemani and A. matricariae females were separately introduced into the test arena for 24 hours. After 24 hours exposure, parasitoids were removed. Two weeks after parasitoid exposure, the number of mummies was recorded. Parasitism percentage (number of mummies/total number of hosts) was calculated for each stage and parasitoid species. The procedure was repeated 5 times for each parasitoid species. Choice test To determine which aphid nymphal instars were accepted for oviposition by A. colemani and A. matricariae, 50 aphids - 10 of each instar - were randomly placed on cucumber leaves inside an experimental arena made of translucent glass containers ( cm). One recently-mated female wasp from each species was separately introduced into the test area for 24 hours, and then removed. After one day, second to fourth instar nymphs and adult aphids were moved to other leaves, surrounded by translucent plastic containers using a soft brush, and allowed to develop. The number of mummies was recorded. The procedure was repeated 5 times for each parasitoid species. Statistical analysis Data from experiments were analyzed using ANOVA and means were compared by Duncan s multiple range test (SAS Institute 2003). Standard error values are provided for all means. Results Under no-choice and choice tests, both A. colemani and A. matricariae females were able to parasitize all nymphal instars and the apterous adults of the cotton aphid (Figure 1, A & B). 182
195 However, the percentages of aphids parasitized by the two parasitoid species varied significantly between the different aphid stages. Under no-choice conditions, A. matricariae showed a greater preference for third and fourth instars of the cotton aphid than for other stages (Figure 1A). Percentage of parasitism by A. colemani was highest on third instar A. gossypii, indicating that the third instar of cotton aphid was the most easily utilized by A. colemani for oviposition. In the choice test, both parasitoid species showed a preference for the third and fourth instars. When A. colemani and A. matricariae were exposed to the third instar of cotton aphid in no-choice conditions, mean percentages of parasitism were 23.6±3.69 and 18.4±2.42, respectively (Figure 1A). There was a significant species effect on percentage of parasitism when parasitoids were exposed to the second instar of cotton aphid under choice conditions (F=7.538; df=9; P<0.05), whereas this relationship was not significant for the first (F=0.182; df=9; P>0.05), third (F=1.44; df=9; P>0.05), and fourth nymphal instars (F=1.11; df=9; P>0.05) or at the adult stage (F=1.00; df=9; P>0.05) (Figure 1B). The body size of A. colemani and A. matricariae progeny increased with host stage at the time of parasitization. Discussion Female wasps of A. colemani and A. matricarie oviposited in all 4 nymphal instars and also in recently-emerged adult cotton aphids. The high percentage of parasitism in the third and fourth nymphal instars in the choice experiment shows that female wasps preferred older nymphs for oviposition. Defensive behaviour in these instars therefore had no significant effect on parasitism. There were no significant differences between percentages of parasitism in the first and second nymphal instars and adults (Figure 1B). Similarly, the walnut aphid parasitoid, Trioxys palliodus (Haliday), preferred third and fourth instar nymphs of the walnut aphid, Chromaphis juglandicola (Kaltenbach), (Rakhshani et al., 2004). In contrast, Binodoxys angelicae (Haliday, 1833) preferred first instar nymphs of Aphis pomi de Geer (Cierniewska, 1976). Trioxys cirsii (Curtis) and Monoctonus pseudoplatani (Marshall) showed a preference for younger nymphs of the sycamore aphid, Drepanosiphum platanoides (Shrank) and rarely parasitized older nymphs (Hamilton, 1974). Although older nymphs are generally the most hazardous, aphids parasitized in the 3rd and 4th instar nymphs can emerge as adults and reproduce before being killed (Stary, 1988). Lin and Ives (2003) showed that the preference of A. colemani for large hosts resulted in the greatest reduction in the population growth rate of Aphis glycines Matsumura, because by attacking adults, A. colemani kills the aphids that have the greatest immediate effect on the population growth rate. Thus, parasitism of the aphid instar plays an important role in biological control. Differences in parasitoid preference for the host stage depend on several factors. Mackauer (1983) notes that host stage preference is not constant: it is affected by experimental conditions and is a functional response of the parasitoid to population density. Host stage selection can also considerably affect the population growth of both the host and parasitoid species and can therefore have a definitive effect on whether potential hosts are successfully controlled by parasitoids (Hagvar and Hofsvang, 1991). Further laboratory studies are required to determine the effect of different factors (e.g. various temperatures, relative humidity etc.) on the preference for and effect of various stages of the host on functional response and other aspects of aphid parasitoid behaviour. 183
196 30 x a (A) x a x b x a A. colemani A. matricariae Mean percentage of parasitized hosts x c x b x b x b x bc x b x b y b x a A. colemani A. matricariae 1 st 1 instar 2 nd instar 2 3 rd 3 instar 4 th instar 4 adult 5 x a Aphid Stages x a x a x bc x b x b x b (B) Figure 1. Percentage (mean ± SE) of different life stages of Aphis gossypii parasitized by Aphidius colemani and Aphidius matricariae in no-choice (A) and choice (B) tests. Letters a, b, and c indicate differences within species; letters x and y indicate differences between species (P<0.05). Acknowledgments We are grateful to the Department of Entomology, Tarbiat Modares University for supporting this project. References Bennison, J.A. 1992: Biological control of aphid on cucumber, use of open rearing systems or banker plants to aid establishment of Aphidius matricariae and Aphidoletes aphidomyza. Med. Fac. Landbouw. Univ. Gent. 57(2b):
197 Cierniewska, B. 1976: Studies on the ecology of Trioxys angelicae (Hymenoptera: Aphidiidae) a parasite of the green apple aphid, Aphis pomi (Homoptera: Aphidiidae). Rocz. Nauk Roln. 6: Goh, H.G. & Yoo, J.K. 1997: Controlling cotton aphid, Aphis gossypii, with the parasitoid Aphidius colemani, on banker plant in greenhouse. Abstract presented at the 6 th European congress of Entomology. Ceske Budejovice, pp: 754. Hagvar, E.B. & Hofsvang, T. 1991: Aphid parasitoids (Hymenoptera: Aphidiidae): biology, host selection and use in biological control. Biocontrol News Inform. 12: Hamilton, P.A. 1974: The biology of Monoctonus pseudoplatani, Trioxys cirsii and Dyscritulus planiceps, with notes on their effectiveness as parasites of sycamore aphid, Drepanosiphum platanoides. Ann. Entomol. Soc. Fr. 10: Havelka J. 1978: Carnivorous gall midge Aphidoletes aphidomyza (Diptera: Cecidomyiidae): The bionomic, mass laboratory rearing and use against aphids on greenhouses crops. Ph.D. Thesis, 259pp. Kersting U., Satar S. & Uygun N. 1999: Effect of temperature on development rate fecundity of aptrous Aphis gossypii Glover (Homoptera: Aphididae) reared on Gossypium Hirsutum. J. Appl. Entomol. 123: Lin, L.A. & Ives, A.R. 2003: The effect of parasitoid host-size preference on host population growth rates: an example of Aphidius colemani and Aphis glycines. Ecol. Entomol. 28: Mackauer, M. 1983: Determination of parasite preference by choice tests: the Aphidius smithi (Hymenoptera: Aphidiidae)-pea aphid (Homopetare: Aphididae) model. Ann. Entomol. Soc. Am. 76: Parrella, M.P., Hansen, L.S. & van Lenteren, J.C., 1999: Glasshouse environments. In: Fisher, T.W., ellows, T.S., Caltagirone, L.E., Dahlstein, D.L., Huffaker, C.B. & Gordh, G. (Eds.), Handbook of Biological Control. Academic Press, New York, pp: Rakhshani, E., Talebi, A.A., Kavallieratos, N. & Fathipour, Y. 2004: Host stage preference, juvenile mortality and functional response of Trioxys pallidus (Hymenoptera: Aphidiinae). Biologia 59(2): SAS Institute. 2003: JMP: a guide to statistical and data analysis, version 5.0.1, Cary, NC. Stary, P Aphidiidae. In: Minks, A.K. & Harewijn, P. (eds), Aphids, their biology, natural enemies and control. Vol. B. Elsevier Publisher, Amsterdam. Toussidou, M., Williams, M. & Leather. S. 1999: Life history parameters of Aphidius colemani on sweet pepper in different temperature regimes. IOBC/WPRS Bull. 22(1): van Lenteren, J.C. & Woets, J. 1988: Biological and integrated pest control in greenhouse. Annu. Rev. Entomol. 33: van Steenis, M.J. 1993: Suitability of Aphis gossypii, Macrosiphum euphorbiae and Myzus persicae as host for several aphid parasitoid species. IOBC/WPRS Bull. 16(2):
198 186
199 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp The number of Cotesia plutellae needed to suppress the population growth of diamondback moth, Plutella xylostella, in a greenhouse Satoru Urano 1, Junichiro Abe 2, Masaya Matsumura 1, Koukichi Nagasaka 2, Junji Takabayasi 3 1 National Agricultural Research Center for Kyushu Okinawa Region; 2421 Suya, Nishigoshi, Kikuchi, Kumamoto , Japan, 2 National Agricultural Research Center for Western Region; 200 Ueno, Ueno, Ayabe, Kyoto , Japan, 3 Center for Ecological Research, Kyoto University; , Hirano, Otsu, Shiga , Japan Abstract: We have been studying the use of host plant volatiles that attract carnivorous natural enemies of herbivorous pest (diamondback moth larvae, Plutella xylostella) from the viewpoint of integrated pest management. Here, we showed the number of Cotesia plutellae, a parasitoid of P. xylostella larva, which is needed to suppress the population growth of P. xylostella in a greenhouse. By using the mathematical model and the previously reported data on the biology of both P. xylostella and C. plutellae, we evaluated the number of wasps needed for the management of P. xylostella to be 7 wasps in one greenhouse (ca. 150 m 2 ). We then conducted the experiments in two greenhouses to confirm the evaluation. When no wasps were released, the growth rates between two generations of P. xylostella were 5 27 times. By contrast, when the wasps were released, the rate remained , suggesting that constant recruitment of a relatively small number of the wasps (ca. 20) from the surroundings of a greenhouse could suppress the population growth rates of P. xylostella. Key words: Cotesia plutellae, Plutella xylostella, population growth, greenhouse, integrated pest management, attractants Introduction The conditions of greenhouse cropping in Japan are similar to those in Mediterranean area in the following points: (1) located in temperate zones, (2) the variation of seasonal temperature is large, (3) the greenhouse structure is similar (open windows in the roof), (4) family farming, (5) the size of the greenhouses are 200 to 2,000 m 2, and (6) the surroundings are rich in biodiversity. Thus, both areas may face similar problems in integrated pest managements in greenhouse. For example, the scale of farming is relatively small and isolated, and it is economically and technically difficult to use a large number of natural enemies that are commercially available. Instead, surroundings harbor many native natural enemies of pest insects. We have been studying the use of plant volatiles that attract carnivorous natural enemies of herbivorous pests as a novel method in integrated pest management in the agroecosystems mentioned above. Cotesia plutellae (Kurdjumov) is a dominant native parasitoid of diamondback moth larvae, Plutella xylostella (Linnaeus) in Japan. They were found in many crop fields and their surroundings (Matsuura, 1977; Yamada & Yamaguchi, 1985; Okada, 1989; Noda et al., 1996; Uematsu & Yamashita, 1999). This species was introduced in many countries such as Australia and Malaysia for integrated pest management (Talekar & Shelton, 1993). P. xylostella is an important pest insect of crucifer crops and occurs from early spring to late autumn. In Japan, the economic injury level by the larvae in greenhouse is quite low, and thus the larvae are serious herbivorous pest insects in greenhouses. Shiojiri et al. (2000) 187
200 reported that cabbage plants infested by P. xylostella larvae attract C. plutellae. In our group, the attractants emitted by the infested cabbage were identified and an artificial blend of the attractants is going to be used in integrated pest management of P. xylostella. To develop the method of the use of natural enemy attractants for integrated pest management, it is important to evaluate the number of natural enemies that is needed to suppress the pest population under the economically injured levels. Small-scaled annual crop farming in temperate zone does not necessarily require the stable low density of pest insect throughout the season (Urano et al., 2003). Rather, in the case of (1) very short cropping intervals, (2) the invasion of herbivorous pests to a greenhouse is by chance, and (3) the case where the very low density of the herbivorous pests results in the serious damages, it is important to know the conditions (=number of natural enemies attracted) in which the population of a herbivorous pest stays under an economic injury level. The number of a natural enemies used for integrated pest managements had been based on the rule thumb, rather than evaluation based on experiments. In this paper, we first obtained the parasitism rate that was necessarily to suppress the population growth of P. xylostella by using a theoretical model. We then evaluated the number of C. plutellae needed to suppress the population growth of the host by using the already reported data of both the parasitoid and the host. In addition, we conducted the greenhouse experiments to evaluate the effective ratio of the wasps and the host larvae. Material and methods We obtained the parasitism rate of C. plutellae needed to suppress the population growth of P. xylostella by using mathematical model (Urano et al. in preparation). To test whether C. plutellae suppress the population growth of P. xylostella in a greenhouse, we conducted the release experiments three times ( ) in Ayabe City, Kyoto, Japan. The size of each greenhouse was m. The wasps were released in one greenhouse and the other was used as a control (no wasps released). The entrance and the openings of the greenhouses were covered with insect net (0.6 mm mesh), thereby the released wasps could not go out of the greenhouse. Two ridges were made to grow Komatsuna plants (Brassica rapa). Sixteen to 25 days after sowing one ridge of plants, we sowed the other ridge. Under this condition, P. xylostella could finish two generations in the greenhouses. We grew ca. 2,000 plants in one ridge. When the plants in the first ridge opened one or two true leaves, we inoculated the second stadium larvae of P. xylostella (total 100 larvae in 2000 plants; 0.05 larva per plant). When plants grew to have 10 leaves per plant, we harvested. In this process, some larvae on the harvested plants might have taken away from the greenhouse. One to three days after the inoculation of the larvae, we released the wasps (the first experiment, April 4th, 2004; the second experiment, September 14th, 2004; the third experiment, March 15th, 2005). In both the released greenhouse and the control greenhouses, we put honey (100 ml; 50% aqueous solution) impregnated into yellow sponge cellulous on a small table 50 cm up to the ground. The number of the wasps released in each experiment were 15 females and 10 males in the first experiment, 10 females and 10 males in the second experiment, and 5 females and 5 males in the third experiment. Wasps were mated one day after adult emergence and were released at 10 a. m. Once a week, we checked plants in the greenhouses to count the number of larvae (3rd and 4th stadium). We estimated the number of larvae in the greenhouses with these data. To evaluate the generation growth rate, we used the Kuno's method of estimating generation boundaries and mean population density for each generation. 188
201 Results and discussion Our model showed that parasitism rates need to suppress the population growth of pest insects were described as more than 1-1/R (R = the generation growth rate of pest insects). Based on Noda et al. (2000) and the additional unpublished data, we calculated the maximum generation growth rate of P. xylostella to be R=7.3. Thus, we estimated that parasitism rates needed to suppress the population growth of P. xylostella was 86 %. The incidence of parasitism by C. plutellae was 19.2 (with honey as food) and 7.6 (no food). If the average number of plants in a greenhouse was 9,000, and the control threshold was 0.05 larva / plant, we need 7 (with food) or 50 (no food) wasps to obtain the parasitism rate to be 86 %. In three greenhouse experiments, the generation growth rates in the control greenhouse were 27 (Experiment 1), 5.2 (Experiment 2) and 9.1 (Experiment 3) (Figure 1, A-C). By contrast, in the released greenhouse, the growth rates were 0.8 (Experiment 1), 0.13 (Experiment 2) and 0.4 (Experiment 3), showing that the wasps decreased the P. xylostella population. In these experiments, we offered honey as food. Based on our model, we could calculate that the number of wasps needed was 7. However, in the greenhouse experiments, we released 15 (Experiment 1), 10 (Experiment 2) and 5 (Experiment 3) wasps. The release rates were 1:6.7, 1:11 and 1:22 respectively, while the calculated release rate was 1:64. Thus, in wasp-larva ration base, we released ca. 3 to 10 times more wasps in the experiments. The results suggested that at least three to ten times more wasps than the model expectation would be sufficient to control P. xylostella larvae in Komatsuna greenhouses in Japan. Acknowledgements We thank Dr. T. Noda, Dr. K. Takashino, Dr. H. Kanno, Dr. T. Mitsunaga, Dr. T. Shimoda, Dr. K. Sano, Dr. Y. Ohara, Dr. S. Kugimiya, Drs. K. Shiojiri and Dr. M. Uefune for thier useful suggestions. We are also grateful to Ms. K. Ito and Ms. H. Tateishi. This study was supported by Bio-oriented Technology Research Advancement Institution. References Matsuura, M. 1977: Parasites of the diamondback moth, Plutella xylostella (Linnaeus), their species and seasonal fluctuations. Bull. Fac. Agric. Mie Univ: Noda, T., Miyai, S., Yamada, S. & Konishi, K. 1996: Larval and pupal parasitoids of diamondback moth, Plutella xylostella (L.) in cabbage fields in Morioka, Japan. Jpn. J. Appl. Entomol. Zool. 40: Noda, T., Miyai, S., Takashino, K. & Nakamura, A. 2000: Density suppression of Plutella xylostella (Lepidoptera: Yponomeutidae) by multiple releases of Diadegma semiclausum (Hymenoptera: Ichneumonidae) in cabbage fields in Iwate, northern Japan. Appl. Ent. Zool. 35: Okada, T. 1989: Parasitoids of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae): species and seasonal changes of parasitism in cabbage fields. Jpn. J. Appl. Entomol. Zool. 33: Shiojiri, K., Takabayashi, J., Yano, S. & Takafuji, A. 2000: Flight response of parasitoids toward plant-herbivore complexes: A comparative study of two parasitoid-herbivore systems on cabbage plants. Appl. Ent. Zool. 35(1): Talekar, N.S. & Shelton, A.M. 1993: Biology, ecology and management of diamondback moth. Anuu. Rev. Entomol. 38:
202 Uematsu, H. & Yamashita, T. 1999: Seasonal occurrence of diamondback moth, Plutella xylostella (L.) and parasitoids in some commercial cruciferous fields in Miyazaki, southern Japan. Jpn. J. Appl. Entomol. Zool. 43: Urano, S., Shima, K., Hongo, K. & Suzuki, Y. 2003: A simple criterion for successful biological control on annual crops. Population Ecology 45: Yamada, H. & Yamaguchi, T. 1985: Notes on the parasites and predators attacking the diamondback moth, Plutella xylostella (L.). Jpn. J. Appl. Entomol. Zool. 29: Greenhouse with No wasp release Wasp release A Total no. of diamondback moth larvae Generation 1 Generation 2 Greenhouse with No wasp release Wasp release Generation 1 Generation 2 Greenhouse with No wasp release Wasp release B C Generation 1 Generation 2 Figure 1. Generational changes in the total number of diamondback moth larvae: (A) Experiment 1, release rate 1: 6.4; (B) Experiment 2, release rate 1: 11; (C) Experiment 3, release rate 1:
203 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 p. 191 Functional response of Chelonus oculator (Hym.: Braconidae) in greenhouse conditions: influence of the parasitoid release ratio M. García-Martín 1, M. Gámez-Cámara 2, A. Torres-Ruiz 1, T. Cabello 1 1 Dpto. Biología Aplicada, 2 Dpto. Estadística y Matemática Aplicada, Universidad de Almería, Almería, Spain Abstract: C. oculator is an endoparasitoid of Lepidoptera. In laboratory conditions, this parasitoid exhibited a type III functional response. This is the unique one that is considered adaptive, host density dependent and being able to regulate the pest population in a conventional program of Biological Control. The objective of this work was to verify the parasitoid functional response in greenhouse conditions, as well as the influence of the parasitoid release ratio and, therefore, to evaluate its potentiality as a natural enemy of beet armyworm Spodoptera exigua. The trials were carried out from May to June of 2005 in greenhouse crops placed at F.I.A.P.A. research station. Several parasitoid release ratios and host densities were employed. The data were fitted to type I, II and III models, to a newly developed equation (type III with 2 parameters) and to a functional response surface. Total parasitization and percentage of discovered egg masses were significantly greater in the highest release ratio (1.5 /m 2 ). Moreover, in field conditions the functional response was of type III. The best fit was obtained with the new equation, with exception for the density of 1.5 /m 2. The parameters handling time (T h ) and parasitism potential (α) were well correlated to those found in our previous laboratory test. Our results show this parasitoid species could be a good pest control agent in greenhouse crops in southeast Spain. Keywords: functional response; Chelonus oculator, Ephestia kuehniella, greenhouse crops 191
204 192
205 Generalist predators
206 194
207 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp When does alternative food promote biological pest control? Maurice W. Sabelis 1, Paul C. J. Van Rijn 2 1 Section Population Biology, Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, Amsterdam, sabelis@science.uva.nl; 2 Institute of Terrestrial Ecology, Heteren, The Netherlands, p.vanrijn@nioo.knaw.nl Abstract. That alternative food, whether or not provided by a plant or introduced artificially, promotes biological pest control via its effect on predators, is not immediately obvious. On the one hand it enhances survival, reproduction and searching. On the other hand it may reduce the rate of predation, which is the case when alternative food and prey are substitutable as opposed to complementary food sources. Moreover, it is not immediately obvious how the impact of alternative food on the outcome of biological pest control differs depending on the type of dynamics (e.g. equilibrium vs transient dynamics), the type of predator (e.g. stage-related consumption and life history effects of alternative food) and food web structure (presence of hyperpredators or intraguild predators). We review the conditions under which alternative food can lead to either pest extermination, to a decline of the pest towards a positive, asymptotic density or to no effect on pest density at all. Key words: biological control, predators, phytophagous arthropods, alternative food, pollen. Introduction. Carnivorous arthropods such as predators and parasitoids supplement their diet with plantprovided food (PPF), such as nectar or pollen. Depending on the arthropod taxa under consideration, this type of omnivory may occur in all or only in specific feeding stages. It usually stimulates survival, development and/or reproduction, thereby promoting the numerical response of the carnivore to the density of prey or host. Whether this numerical effect on the carnivore also translates in reduced prey/host densities, is not immediately obvious because consumption of PPF may negatively affect consumption of prey, because prey consumption and PPF consumption vary with life history stage, because prey and PPF vary in spatial distribution and because prey and PPF may be eaten by other members of the food web. Assessing the conditions, under which PPF reduces density at the second trophic level via its impact on the third, is of crucial importance to designing strategies for biological control of crop pests. Here, we summarize the results of theoretical exercises with consumer-resource models presented elsewhere (Van Rijn & Sabelis, 2005). First, we consider the lessons from simple consumer-resource models that ignore stage or spatial structure. Second, we discuss results from stage-structured consumer-resource models to investigate how the impact of PPF on herbivory may depend on the life history and feeding requirements of the carnivorous arthropods. We compare parasitoids and predators, as well as predators with different types of omnivory. Finally, we step beyond consumer-resource interactions and ask when food web complexities (omnivory, hyperpredation, intraguild predation, competition) alter the predictions from simple consumer-resource models. All this will be discussed with a keen eye for how the theory can be applied to reduce crop damage by herbivores. 195
208 Well-mixed, unstructured consumer-resource models. To understand how PPF through its effect on the carnivore affects herbivore abundance, consider a system where a predator population directly controls a herbivore population, where individuals of each species are identical (e.g. no (st)age structure) and populations are well mixed (no spatial structure). These conditions apply to Lotka-Volterra or Rosenzweig- MacArthur models. The addition of PPF will initially result in an increase of the predator population, simply because there is more food available. This increase will come to a halt at equilibrium, i.e. when births exactly compensate for deaths. At this point, the herbivore population has decreased to an extent that compensates for the supply of PPF. Thus, adding food will lead to a decrease in the herbivore population via the consumers they share. This effect of a non-reproducing food source is very similar to the case where adding a second, reproducing prey species causes a decline of the first via the predator they share (Holt & Lawton, 1994; Van Baalen et al., 2001; Van Rijn et al., 2002). The effect of one prey species on the other looks like competition, but in fact it is only apparent, because the mechanism is predator-mediated. This is why Holt (1977) termed it apparent competition, to create a contrast with resource and interference competition. From the equilibrium equations of the one-predator-two-prey or predator-ppf-prey models, some counterintuitive conclusions emerge. If prey and PPF are substitutable food sources for the predator (Tilman, 1982), addition of PPF will reduce equilibrium prey density, even when per capita consumption of PPF leads to a decrease in per capita consumption of prey. This insensitivity of the herbivore equilibrium to per capita prey consumption arises because equilibrium implies that the predators will increase to larger densities to achieve an overall prey mortality rate that compensates the overall prey birth rate. As long as the per capita predation rate exceeds zero, increasing the amount of PPF can even lead to extinction of the prey. At prey extinction, the predator population is maintained only by PPF. This prediction of prey extinction by adding PPF does not hold when food and prey are not substitutable, but complementary. Two food types are thought to complement each other when they influence different components of the predator s life history. As an extreme example, consider the case where prey affects reproduction and PPF affects mortality. Then, increasing the amount of PPF also reduces herbivore density, but can never lead to extinction of the prey. This is because herbivore density declines asymptotically to a fixed level determined by background (= minimum) predator mortality. As long as assimilation of PPF and prey will have a positive effect on predator reproduction and survival (which is why PPF should be eaten by the predator anyway!), PPF will reduce equilibrial prey densities irrespective of its effect on the prey consumption rate. Thus, it does not matter whether the predator switches to PPF at low prey densities or whether it becomes satiated for PPF at another level of ingested biomass than for prey (Van Baalen et al., 2001; Van Rijn, unpublished data). It even does not matter whether the herbivore consumes PPF and therefore survives, reproduces or develops faster (Van Rijn et al., 2002)! At equilibrium, the enhanced herbivore performance due to PPF will be compensated by predation from a larger predator population. The equilibrium approach holds when environmental conditions, such as climate, availability of PPF to predators and plants to herbivores, remain unchanged over a sufficiently long period. How long the conditions need to be constant to approximate the equilibrium depends on the initial densities of the interacting populations, their generation times, and other traits of predator and prey that determine the dynamics around the equilibrium. For carnivorous mites and herbivorous thrips with generation times of about 3 weeks, populations were already within the 10% range of their equilibrium level after 12 weeks following their 196
209 introduction in a cucumber crop (Van Rijn et al., 2002). After this period the impact of a regularly supplied food source on mite and thrips populations can adequately be predicted from equilibrium equations only. Arthropods larger than mites and thrips generally have longer generation times and their populations require more time to settle around the equilibrium (Sabelis, 1992). For insects with only one or two generations per year and with food sources available only during part of the year, an equilibrium approach is unlikely to hold. In that case, one should rather focus on the dynamics displayed before the system approaches its equilibrium state (so-called transient dynamics). Models of such systems require proper representation of developmental delays and age-dependent reproduction and this may make them mathematically less tractable. Although numerical techniques are available to simulate transient dynamics (Caswell, 1989; Nisbet, 1997; De Roos & Persson, 2001), obtaining transparent insight requires simplification. We reduced complexity by focusing on the first generation after predator release (Van Rijn & Sabelis, 2005). We assumed predators to be subject to a constant (i.e. herbivore-independent) per capita mortality rate, whereas the per capita prey mortality depends on predator density. Solving the integral over the first generation of the predators yields an expression relating prey density to background (= predator-independent) prey mortality, predation rate and predator mortality. If prey and PPF are complementary food sources, PPF may reduce predator mortality without affecting the predation rate. Then, reducing predator mortality by adding PPF translates into reduced prey density. If, however, PPF and prey are substitutable, then adding PPF may reduce predator mortality as well as predation rate. To make herbivore density go down requires that PPF reduces predator mortality more than it reduces the predation rate. If PPF is not only utilized by predator, but also by prey, adding PPF as a complementary food to prey will only reduce herbivore density when its reducing effect on predator mortality is disproportionally larger than that on background prey mortality. This demand will be even more extreme when substitutability of foods is assumed and therefore consumption of PPF will likely go at the expense of that on prey. Thus, for prey density to go down in the first generation after predator release adding PPF is subject to stringent conditions. It is then critically important to know whether foods are substitutable or complementary and whether herbivores utilize PPF as well. These conclusions are quite complex, yet they are intuitively much more obvious, than the simple general conclusion drawn for the case of equilibrium conditions stating that: PPF always reduces herbivore density irrespective of its effect on predation and irrespective of its utilization by the herbivore. Stage-structured consumer-resource models. Real-world predator prey and parasitoid host interactions differ in the life stages that are affected by prey (or host) density and/or PPF. In general, three scenarios can be observed that differ in whether (A) adult performance (survival, attack, oviposition), (B) juvenile performance (survival, development) and (C) both adult and juvenile performance are affected by prey density. The different types of PPF (nectar, pollen) can also have three different effects as it may (1) promote survival, (2) provide fuel (when rich in sugars as in nectars) for searching and (3) enhance assimilation processes (when rich in amino acids as in pollen), and thereby development and oviposition. As defined above, PPF and prey can be substitutable or complementary food sources and they are called essential if their absence causes the carnivore population to decline even at the highest abundance of the other food source (i.e. reproduction does not compensate mortality or the basic reproduction ratio R 0 < 1). Assuming the prey/host is always essential, PPF is never essential when substitutable, but when complementary it can be either essential or not. 197
210 Scenario A applies to parasitoids. Here, the larva is carnivorous. It feeds in or on a single host. So there is no need to search for hosts. The adult female searches for hosts and she makes decisions on whether to lay eggs in hosts. Usually the adult female feeds on PPF, but some species also feed on hosts. Thus, host density affects the oviposition rate of the adult parasitoid, but not the survival or development of the larvae. Scenario B comes close to hoverflies. Here, the purely carnivorous larvae actively search for prey, whereas the adult females feed on PPF and lay eggs near areas with prey. Thus, prey density will here most strongly affect juvenile performance. Scenario C is best illustrated by ladybeetles, predatory bugs, earwigs and predatory mites, where carnivory and search for prey occurs in all active stages. Here, prey density will affect the juvenile, as well as adult performance. Lacewings have predatory larvae, but as adults only some species are carnivorous. Thus, they represent either scenario B or C. If the oviposition rate of hoverflies strongly depends on how much prey the area harbours, then they are more close to scenario C than to B. Scenario C also becomes more applicable to parasitoids when they kill and feed on hosts to obtain nutrients essential for egg maturation. For each of the three scenarios on stage-related prey density dependence models were developed that incorporate the effect of substitutable or complementary PPF affecting (1) survival, (2) searching or (3) development and reproduction, plus any combination of 1, 2 and 3. Parameters ranges were based on literature data. From the equilibrium equations of those models we derived how equilibrium prey density changes with an increase in PPF. Such calculations are particularly meaningful when populations return to the equilibria after perturbation (i.e. equilibria are stable). This is likely when not all prey stages are vulnerable to predator attack (Murdoch et al., 1987), which holds for many arthropod predator-prey systems (Sabelis, 1992; Sabelis & Van Rijn, 1997). These calculations show that increasing PPF whether substitutable or complementary, essential or not essential causes equilibrium prey density always to decline under all scenarios, but the mode and quantitative details of the decline depend on the scenario under consideration. Extinction above a critical level of PPF availability can only be achieved when the predators eat PPF and prey as substitutable foods in all life stages (and are thus true omnivores) whereas both mortality and reproduction are affected by PPF. In all other cases increasing PPF can never drive the prey population to extinction. Instead, prey density will asymptotically approach some positive value set by the level of background (= minimum) predator mortality. Under each of the three scenarios (A, B and C) the strongest decline in prey density is achieved when PPF is substitutable (and hence non-essential) and when PPF promotes both survival and reproduction of the predator; effects of PPF on survival alone come second in prey suppression efficiency and effects of PPF on searching alone come third. When PPF is essential (and hence complementary), there is a minimum amount of PPF required for the predator population to persist and thereby to suppress the prey population. Beyond consumer-resource models In this article we provided a review of the conditions under which PPF gives rise to herbivore suppression via a shared consumer. In particular, we considered how the impact of PPF is modified by stage- and space-related interactions. With few exceptions, the overall pattern is that PPF somehow promotes herbivore/prey suppression. The underlying assumption was that the system consists of one species at the third trophic level, one species at the second trophic level and PPF, as an influence from the first trophic level. In reality, herbivore and carnivore are part of a much more complex food web of species interacting with each other, (Polis & 198
211 Strong, 1996). How will these interactions affect the conditions under which PPF leads to herbivore suppression? Consider first the presence of a fourth trophic level. Carnivores may have their own suite of (hyper-)predators, (hyper-)parasitoids, and pathogens (Rosenheim, 1998; Sullivan & Volkl, 1999). Trophic cascade models predict that the top-carnivore will at equilibrium control the primary carnivore, so that the herbivore is released from top down control (Oksanen et al., 1981). The equilibrium density of the primary carnivore would be determined by the traits of the top-carnivore, whereas the herbivore would grow to a density where it is limited from bottom up. This implies that at equilibrium, food provided to the primary carnivore would no longer affect the density of the primary carnivore, nor that of the herbivore! In some cases, PPF can (also) be used by the top-predator or hyperparasitoid (Chang et al., 1994). PPF will now likely reduce the density of the primary carnivore, and consequently have a negative rather than a positive impact on biological control of the herbivores. Thus, the presence of a fourth trophic level may dramatically alter the predictions for the impact of PPF on herbivore suppression. At the third trophic level, competition for herbivores as prey and intraguild predation may alter the species composition and thereby the impact on herbivore suppression (Polis et al., 1989; Polis & Holt, 1992; Rosenheim, 1998). PPF may change the outcome of competition and intraguild predation by promoting one species more than others (e.g. Evans & England, 1996). If PPF supports the species that in absence of PPF is a worse competitor but a good intraguild predator, PPF may reduce the density of the better competitor and promote herbivore density (Rosenheim et al., 1995; Holt & Polis, 1997; Briggs & Collier, 2001; Mylius et al., 2001; Rosenheim, 2001; Snyder & Ives, 2001; Hunter et al., 2002). However, beyond the PPF level that results in exclusion, PPF will have the same effects as predicted from simple carnivore-herbivore models. Thus, rather restrictive conditions are required for competition and intraguild predation to alter the predictions for the impact of PPF on herbivore suppression obtained from simple predator-prey models. References Briggs, C.J. & Collier, T.R. 2001: Autoparasitism, interference, and parasitoid-pest population dynamics. Theor. Popul. Biol. 60: Caswell, H. 1989: Matrix Population Models. Sinauer, Sunderland, Massachusetts. Chang, Y.D., Lee, J.Y., & Youn, Y.N. 1994: Primary parasitoids and hyperparasitoids of the soybean aphid, Aphis glycines Matsumura (Homoptera: Aphididae). Korean. J. Appl. Entomol. 33: De Roos, A.M. & Persson, L. 2001: Physiologically structured models - from versatile technique to ecological theory. Oikos 94: Evans, E.W. & England, S. 1996: Indirect interactions in biological control of insects: Pests and natural enemies in alfalfa. Ecol. Appl. 6: Holt, R.D. 1977: Predation, apparent competition and the structure of predator-prey communities. Theor. Popul. Biol. 12: Holt, R.D. & Polis, G.A. 1997: A theoretical framework for intraguild predation. Am. Nat. 149: Holt, R.D., & J. H. Lawton. 1994: The ecological consequences of shared natural enemies. Annu. Rev. Ecol. Evol. S. 25: Hunter, M.S., Collier, T.R. & Kelly, S.E. 2002: Does an autoparasitoid disrupt host suppression provided by a primary parasitoid? Ecology 83:
212 Murdoch, W.W., Nisbet, R.M., Blythe, S.M., Gurney, W.S.C. & Reeve, J.D. 1987: An invulnerable age class and stability in delay-differential parasitoid-host models. Am. Nat. 129: Mylius, S.D., Klumpers, K., de Roos, A.M. & Persson, L. 2001: Impact of intraguild predation and stage structure on simple communities along a productivity gradient. Am. Nat. 158: Nisbet, R.M. 1997: Delay-differential equations for structured populations. In Structuredpopulation models in marine, terrestrial, and freshwater systems (S. Tuljapurkar and H. Caswell, eds), pp , Chapman & Hall, New York. Oksanen, L., Fretwell, S.D., Arruda, J. & Niemela, P. 1981: Exploitation ecosystems in gradients of primary productivity. Am. Nat. 118: Polis, G.A., & Strong, D.R. 1996: Food web complexity and community dynamics. Am. Nat. 147: Polis, G.A. & Holt, R.D. 1992: Intraguild predation - the dynamics of complex trophic interactions. Trends Ecol. Evol. 7: Polis, G.A., Myers, C.A., & Holt, R.D. 1989: The ecology and evolution of intraguild predation - potential competitors that eat each other. Annu. Rev. Ecol. Evol. S. 20: Rosenheim, J.A. 1998: Higher-order predators and the regulation of insect herbivore populations. Annu. Rev. Entomol. 43: Rosenheim, J.A. 2001: Source-sink dynamics for a generalist insect predator in habitats with strong higher-order predation. Ecol. Monogr. 71: Rosenheim, J.A., Kaya, H.K., Ehler, L.E., Marois, J.J. & Jaffee, B.A. 1995: Intraguild predation among biological-control agents - theory and evidence. Biol. Control 5: Sabelis, M.W. 1992: Arthropod predators. In Natural Enemies, The Population Biology of Predators, Parasites and Diseases (M. J. Crawley, ed.), pp , Blackwell, Oxford. Sabelis, M.W. & van Rijn, P.C.J. 1997: Predation by insects and mites. In Thrips as Crop Pests (T. Lewis, ed.), pp , CAB-International, London. Snyder, W.E. & Ives, A.R. 2001: Generalist predators disrupt biological control by a specialist parasitoid. Ecology 82: Sullivan, D.J. & Volkl, W. 1999: Hyperparasitism: Multitrophic ecology and behavior. Annu. Rev. Entomol. 44: Tilman, D. 1982: Resource Competition and Community Structure. Princeton University Press, Princeton, New Jersey. van Baalen, M., Krivan, V., van Rijn, P.C.J. & Sabelis, M.W. 2001: Alternative food, switching predators, and the persistence of predator-prey systems. Am. Nat. 157: van Rijn, P.C.J., van Houten, Y.M. & Sabelis, M.W. 2002: How plants benefit from providing food to predators even when it is also edible to herbivores. Ecology 83: van Rijn, P.C.J. & Sabelis, M.W. 2005: Impact of plant-provided food on herbivore-carnivore dynamics. In Plant-Provided Food and Herbivore Carnivore Interactions (F.L. Wäckers, P.C.J. van Rijn & J. Bruin, eds). pp , Cambridge University Press, Cambridge, UK. 200
213 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Feeding behavior of the rove beetle, Atheta coriaria (Coleoptera: Staphylinidae) under laboratory conditions Eva M. Birken, Raymond A. Cloyd Department of Natural Resources and Environmental Sciences, University of Illinois, Urbana, IL 61801, USA, Abstract: Laboratory choice tests were conducted to determine the feeding behavior, based on food preference, of the adult and larval stage of Atheta coriaria when presented with both fresh moistened oatmeal and second instar fungus gnat, Bradysia sp. nr. coprophila larvae. In all tests, Atheta coriaria adults and larvae preferred to feed on fungus gnat larvae more so than oatmeal based on the amount of oatmeal and number of fungus gnat larvae consumed after 4 and 6 hours. Key words: Atheta coriaria, feeding behavior, biological control, integrated pest management Introduction The rove beetle, Atheta coriaria Kraatz (Coleoptera: Staphylinidae) is a soil-dwelling predator that has the capability to be an effective biological control agent for certain greenhouse pests such as fungus gnats, Bradysia spp. (Diptera: Sciaridae) (Carney et al., 2002), which are important insect pests in greenhouses (Hamlen & Mead, 1979). Plant damage mainly occurs from the larvae feeding on root tissue in the growing medium (Wilkinson & Daugherty, 1970). If populations are excessive, this may result in economic losses. Atheta coriaria is a small beetle, approximately 3 to 4 mm in length, and dark brown in color (Miller & Williams, 1983). Adults are very mobile and are capable of flying long distances, however, they tend to spend most of their life span in growing media (Helyer et al., 2003). The larvae are white in color during the early instars, turning a yellowish-brown color in the later instars. Both the larvae and adult may feed on various life stages of a wide-range of arthropod pests including fungus gnats (eggs, larvae, and pupae), shore flies, Scatella spp. (eggs, larvae, and eggs), and thrips (pupae and second stage larvae) (Helyer et al., 2003). However, relatively little is known regarding the interaction between A. coriaria and one of their primary food sources, fungus gnats. In addition, there are no documented reports indicating that A. coriaria will feed on alternative food sources in the absence of fungus gnats. The objective of this study was to observe and quantify the feeding behavior, based on food preference, of rove beetle adults and larvae under laboratory conditions. Materials and methods In this study, rove beetle adult and larval food preferences were evaluated using choice tests, which included fresh oatmeal (The Quaker Oats Company, Boston, MA) and 2 nd instar fungus gnat larvae. The fungus gnats were obtained from a laboratory colony established using flies from the growing medium of a potted geranium (Pelargonium sp.). Specimens from the colony have been identified as Bradysia sp. nr. coprophila. The colony is maintained in moist growing medium supplemented with shredded potato and oatmeal (Cloyd & Zaborski, 2004). The method of obtaining a cohort of same-aged fungus gnat larvae is described in Cloyd & 201
214 Dickinson (2005). Rove beetles used in this study came from a laboratory-reared colony and have been identified as Atheta coriaria (Kraatz); however, recently this species was transferred to a closely related genus, Dalotia Casey so the correct citation should be Dalotia coriaria (Kraatz). In this study, we will use the commonly cited species name, Atheta coriaria. Adult choice tests Rove beetle adults (age and sex unknown) were obtained from established laboratory colonies, created on October 1, 2002 and November 10, 2003 by sieving a sample of growing medium through an 850-micron sieve and then agitating the sieve for less than one minute, which separated the adults from the growing medium. The adults were collected on a 425- micron sieve and then aspirated into 7 x 3-cm plastic vials. A single adult was placed in the center of a petri dish (55-mm in diameter) lined with moistened Whatman No. 1 filter paper (Whatman, Maidstone, England). On one side of the petri dish were 5 or ten 6-day old 2 nd instar fungus gnats and on the other side were two to three pieces of fresh moistened oatmeal. The adult rove beetles were placed equidistant from both food sources. All petri dishes were sealed with elastic film (Parafilm ) and placed into an environmental growth chamber (Sherer Environmental Chamber model CEL-36-10, Warren/Sherer Division of Kysor Industrial Corp., Marshall, MI) set at 22±3ºC, 50 to 55% relative humidity, and 0:24 (L:D) hour photoperiod. There were three separate choice tests for the adults with 10 adults per test. Each petri dish containing an adult was a replicate. Data were recorded after four and six hours, then the adults were removed from the petri dishes with an aspirator. The amount of oatmeal and number of fungus gnats consumed by each rove beetle adult was objectively assessed based on percent missing (0 to 100%). Larvae choice tests A similar choice test was conducted using rove beetle larvae. The larvae used were obtained from another study evaluating rove beetle adult female fecundity. Newly hatched larvae (<24 hours old) were only fed two to three pieces of fresh moistened oatmeal before they were used in the choice tests. In each petri dish (55-mm in diameter), which was lined with moistened filter paper, five to seven 7-day old 2 nd instar fungus gnats were positioned on one side and two to three pieces of fresh moistened oatmeal were located on the other side of the petri dish. A single rove beetle larva was placed in the center of each petri dish, equidistant from both food sources, using a soft-bristled camel paintbrush. There were three separate choice tests for the larvae with five to seven larvae per test. Each petri dish containing a larva was a replicate. Petri dishes were sealed with an elastic film (Parafilm ) and placed into an environmental growth chamber (described above) set at 22±3ºC, 50 to 55% relative humidity, and 0:24 (L:D) hour photoperiod. Data were recorded after four and six hours. Larvae were then removed from the petri dishes using an aspirator. The amount of oatmeal and number of fungus gnats consumed by each rove beetle larva was objectively assessed based on percent missing (0 to 100%). Results and discussion Results of the choice tests indicate that rove beetle adults preferred to feed on fungus gnat larvae more so than oatmeal (Tables 1 through 3). Rove beetle larvae also preferred to feed on fungus larvae compared to oatmeal (Tables 4 through 6). The results of this study clearly indicate that rove beetle larvae that have been fed oatmeal as their only food source tend to prefer fungus gnat larvae. After four hours, both the adults and larvae consumed most of the fungus gnat larvae, with no major differences evident six hours after releasing the adults and larvae into the petri dishes. It has been suggested that since A. coriaria has such a wide-host 202
215 range that it is capable of switching from preferred hosts to non-preferred hosts (Miller & Williams, 1983). However, this study is the first to quantitatively demonstrate that A. coriaria larvae, even when initially feeding on an alternative food source, in this case oatmeal, will still feed on fungus gnat larvae. We have been rearing rove beetles in our laboratory for approximately four years and supply them with both fungus gnat larvae and oatmeal as food sources. However, in our experience, long-term rearing (>3 generations) of rove beetles may lead to the inadvertent development of astigmatid mite, Sancassania aff. sphaerogaster (Acari: Acaridae) and/or potworm (Oligochaeta: Enchytraeidae) populations. Since A. coriaria is a generalist predator, it is likely that the adults and larvae are feeding on these organisms as well as fungus gnat larvae and oatmeal. This suggests that alternative food sources should be tested to assess whether rove beetles, initially feeding on a substitute food source or prey, will still provide control of fungus gnats when released into greenhouses. As the use of natural enemies increases in greenhouses, there is a need for basic information regarding biology and feeding behavior. This includes mass-rearing, laboratory, and field studies to better understand the potential value of the rove beetle, A. coriaria. Acknowledgments We thank Amy Dickinson and Theresa Meers (University of Illinois, Urbana, IL) for providing technical support. References Carney, V. A., J. C. Diamond, G. D. Murphy, & D. Marshall. 2002: The potential of Atheta coriaria Kraatz (Coleoptera: Staphylinidae), as a biological control agent for use in greenhouse crops. IOBC/WPRS Bull. 25(1): Cloyd, R. A., & A. Dickinson. 2005: Effects of growing media containing diatomaceous earth on the fungus gnat Bradysia sp. nr. coprophila (Lintner) (Diptera: Sciaridae). HortScience 40(6): Cloyd, R. A., & E. R. Zaborski. 2004: Fungus gnats, Bradysia spp. (Diptera: Sciaridae), and other arthropods in commercial bagged soilless growing media and rooted plant plugs. J. Econ. Entomol. 97: Hamlen, R. A., & F. W. Mead. 1979: Fungus gnat larval control in greenhouse plant production. J. Econ. Entomol. 72: Helyer, N., K. Brown, & N. D. Cattlin. 2003: A colour handbook of biological control in plant protection. Manson Publishing, London, England. 224 pgs. Miller, K. V., & R. N. Williams. 1983: Biology and host preference of Atheta coriaria (Coleoptera: Staphylinidae), an egg predator of Nitidulidae and Muscidae. Ann. Entomol. Soc. Am. 76: Wilkinson, J. D., & D. M. Daugherty. 1970: The biology and immature stages of Bradysia impatiens (Diptera: Sciaridae). Ann. Entomol. Soc. Am. 63:
216 Table 1. Adult rove beetle, Atheta coriaria, choice test with five 2 nd instar fungus gnats (Bradysia sp. nr. coprophila) and two to three pieces of fresh moistened oatmeal per petri dish. Percentages are based on amount of oatmeal and number of fungus gnat larvae consumed after 4 and 6 hours (09/14/2005). Replicates 4 hours 4 hours 6 hours 6 hours Fungus gnat Fungus gnat Oatmeal Oatmeal larvae larvae 1. Adult - 100% 10% 100% 2. Adult 10% 100% 10% 100% 3. Adult 10% 100% 10% 100% 4. Adult 25% 100% 25% 100% 5. Adult 25% 80% 25% 100% 6. Adult 10% 100% 10% 100% 7. Adult 25% 80% 25% 100% 8. Adult - 100% 10% 100% 9. Adult 10% 100% 10% 100% 10. Adult - 100% 10% 100% Table 2. Adult rove beetle, Atheta coriaria, choice test with ten 2 nd instar fungus gnats (Bradysia sp. nr. coprophila) and two to three pieces of fresh moistened oatmeal per petri dish. Percentages are based on amount of oatmeal and number of fungus gnat larvae consumed after 4 and 6 hours (10/14/2005). Replicates 4 hours 4 hours 6 hours 6 hours Fungus gnat Fungus gnat Oatmeal Oatmeal larvae larvae 1. Adult Adult 5% 100% 5% 100% 3. Adult 10% 90% 10% 100% 4. Adult 5% 100% 5% 100% 5. Adult - 100% - 100% 6. Adult 10% 70% 10% 90% 7. Adult - 80% 5% 100% 8. Adult 5% 50% 5% 70% 9. Adult 5% 70% 10% 90% 10. Adult - 100% - 100% 204
217 Table 3. Adult rove beetle, Atheta coriaria, choice test with ten 2 nd instar fungus gnats (Bradysia sp. nr. coprophila) and two to three pieces of fresh moistened oatmeal per petri dish. Percentages are based on amount of oatmeal and number of fungus gnat larvae consumed after 4 and 6 hours (10/17/2005). Replicates 4 hours 4 hours 6 hours 6 hours Fungus gnat Fungus gnat Oatmeal Oatmeal larvae larvae 1. Adult 10% 100% 10% 100% 2. Adult 5% 90% 5% 90% 3. Adult 10% 100% 10% 100% 4. Adult 10% 100% 10% 100% 5. Adult - 100% - 100% 6. Adult 5% 100% 5% 100% 7. Adult 10% 90% 10% 100% 8. Adult 5% 100% 5% 100% 9. Adult 5% 90% 5% 90% 10. Adult 10% 80% 10% 100% Table 4. Rove beetle, Atheta coriaria, larvae choice test with five 2 nd instar fungus gnats (Bradysia sp. nr. coprophila) and two to three pieces of fresh moistened oatmeal per petri dish. Percentages are based on amount of oatmeal and number of fungus gnat larvae consumed after 4 and 6 hours (09/14/2005). Replicates 4 hours 4 hours 6 hours 6 hours Fungus gnat Fungus gnat Oatmeal Oatmeal larvae larvae 1. Larvae - 80% - 100% 2. Larvae - 60% - 100% 3. Larvae - 40% - 80% 4. Larvae - 80% - 100% 5. Larvae - 100% - 100% 6. Larvae - 80% - 80% 205
218 Table 5. Rove beetle, Atheta coriaria, larvae choice test with five 2 nd instar fungus gnats (Bradysia sp. nr. coprophila) and two to three pieces of fresh moistened oatmeal per petri dish. Percentages are based on amount of oatmeal and number of fungus gnat larvae consumed after 4 and 6 hours (09/20/2005). Replicates 4 hours 4 hours 6 hours 6 hours Fungus gnat Fungus gnat Oatmeal Oatmeal larvae larvae 1. Larvae 10% 80% 10% 60% 2. Larvae 10% 100% 10% 100% 3. Larvae 10% 60% 10% 80% 4. Larvae 10% 100% 10% 100% 5. Larvae 10% 100% 10% 100% Table 6. Rove beetle, Atheta coriaria, larvae choice test with five 2 nd instar fungus gnats (Bradysia sp. nr. coprophila) and two to three pieces of fresh moistened oatmeal per petri dish. Percentages are based on amount of oatmeal and number of fungus gnat larvae consumed after 4 and 6 hours (09/27/2005). Replicates 4 hours 4 hours 6 hours 6 hours Fungus gnat Fungus gnat Oatmeal Oatmeal larvae larvae 1. Larvae - 100% 10% 100% 2. Larvae - 100% 10% 100% 3. Larvae - 100% 10% 100% 4. Larvae 10% 100% 10% 100% 5. Larvae 10% 100% 10% 100% 6. Larvae 10% 100% 10% 100% 7. Larvae - 60% 10% 100% 206
219 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp The predatory bug Orius niger: its biology and potential for controlling Thrips tabaci in Iran Valiollah Baniameri 1, Ebranhim Soleyman-nejadian 2, Jafar Mohaghegh 1 1 Plant Pest and Disease Research Institute (PPDRI), P. O. Box: 19395/1454 Tehran, 2 Plant Protection Department, University of Sh. Chamran, Ahwaz, Iran Abstract: We measured the basic life-history parameters of Orius niger on a diet of corn pollen and eggs of the moth Ephestia kuehniella at 3 constant temperatures (26, 29 and 32 o C under 16L 8D, 65±5 % RH) to provide information for the application of this predator in biocontrol programs. Bean pods were supplied as oviposition substrates. Results showed that the mean development times from egg to adult were 16.71, and days, sex ratios were 0.59, 0.56 and 0.52 females, respectively. The observed rates of sterile females were 20, 25 and 5 % and the longevity of adult females was 38, 25 and 24 days, respectively. This species preys upon thrips and is commonly found attacking T. tabaci throughout Iran. It was concluded that O. niger is an ideal predator to use as a biological agent in greenhouse crops which have enough pollen, or when they are mixed with banker plants. Key words: Orius niger, biology, thrips, biological control Introduction Greenhouse crops are currently grown on 6,008 ha., and are mainly located in southern and central parts of Iran. More than 50 pest species and diseases affect these greenhouse crops, of which 6 are thrips species. Two of these, Thrips tabaci Lind. and Frankliniella occidentalis Pregande, are important pests (Baniameri, 2005). In recent years, the use of predatory bugs of the genus Orius as biological control agents of thrips has greatly increased in many greenhouse crops (vegetables and ornamentals) (Cocuzza et al., 1997; Van den Meiracker & Ramakers, 1991; Van den Meiracker, 1999). In 2001, F. occidentalis was imported from the Netherlands with gerbera seedlings and is now widely found in various areas, including Tehran province. Thrips cause direct crop damage by feeding on parenchyma and puncturing plant tissue for oviposition, resulting in a reduction in photosynthetic capacity (Van Rijn, 2002; Royer et al., 1986). They also cause indirect damage by transmitting plant viruses, such as TSWV and INSV (Broadbent et al., 1987; German et al., 1992). Orius niger Wolff is a Palaearctic species (Loomans et al., 1995) recorded in many parts of Iran (Baniameri, 2003). This species preys upon thrips and is commonly found attacking T. tabaci throughout Iran, suggesting that it is adapted to various different climatic conditions found within the country. The biology of O. niger was studied using Ephestia eggs as prey under laboratory conditions. The objectives of the current study were to determine the development time, sex ratio, oviposition period and total number of eggs per female of O. niger at three different temperatures. These data would be useful in biocontrol programs using O. niger as a predator of thrips. 207
220 Materials and Methods Insect colonies The predatory bugs used in the experiments were collected from marguerite, Chrysanthemum frutescens L. (Compositae) flowers, in Tehran, Iran. O. niger was separated and identified using the single rearing method (Yasunaga, 1997). Individuals were reared at 26 C, 70±5 % RH and with a photoperiod of L16:D8 hours with eggs of E. kuehniella as food. Green bean pods and corn pollen were used as oviposition sites and supplementary food, respectively. Life history parameters The experiment was carried out at the Agricultural Entomology Dept. of the PPDRI. Fresh bean pods were placed in rearing jars of O. niger for oviposition. After four hours, pods with eggs were incubated at 26ºC, 29ºC or 32 C, 70±5 % RH and L16:D8 in plastic soft-drinks bottles (150 ml) lined with paper tissue, and closed with a screw cap. Three cohorts of 159, 103 and 96 eggs were used at 26ºC, 29ºC and 32 C, respectively. Ventilation was through two lateral holes covered with fine nylon mesh. Water was supplied in a small glass test tube closed with cotton wool. E. kuehniella eggs were provided for nymphs and adults on a small piece of paper which was replaced every day. Bottles were checked daily for egg hatching, egg and nymph survival, and adult emergence. Sex ratio, total nymph survival and development time from egg to adult were recorded at all three temperatures. A total of 30 females emerged at 26 C and 20 females emerged at the other two temperatures. They were separated and kept individually with 2-3 males in a small test tube for one hour to ensure mating. Mated females and their accompanying males were transferred to a plastic cup (100 ml) with a lid and two lateral holes covered with mesh to provide ventilation. The total number of eggs laid by the females, oviposition period and number of sterile females were all recorded. Results and discussion The duration of development for the eggs, nymphs and the total life cycle of O. niger at the three temperatures is presented in Table 1. The lowest mean development time from egg to adult was days at 29 C. The development time and survival rate of the Orius species on Lepidoptera eggs had previously been studied. According to these studies, the duration of nymphal development at about 25 C varied from 10 to 18 days (Van den Meiracker, 1999). Nymphal survival was between 60% and 63% at the three temperatures. Nymphal survival for the Orius species was usually greater than 70% on average (Van den Meiracker, 1999), which is comparable with the results of this study. The highest sex ratio was 0.59 females at 26 C. The longevity of adult females was 38, 21 and 19 days, and the rate of sterile females was respectively 20%, 25% and 5%, at the three temperatures. The oviposition period and the number of eggs laid per female at the three above-mentioned temperatures was 33, 16 and 14 days, and 76.70, and eggs, while the hatching rates were 0.79, 0.90 and 0.88, respectively. Results showed that the mortality rate decreased with age. The highest mortality was during the nymphal period and the lowest occurred in pre oviposition and in the early stages of the oviposition period. In another study, Baniameri et al. (2003) recorded the mean parameters of the predation rate for O. niger, which were 6.22±1.18, 8.60±0.76 thrips larvae per hour; the encounter rate was 0.525±0.08, 0.724±0.029 per hour; the success ratio was 0.973±0.14, 0.852±0.041 and the handling time was 4.095±0.672, 3.006±0.520 larvae per minute with densities of 1 and 4 females, respectively. Of these parameters, the encounter rate and success ratio were significantly higher at densities of 4 than 1. O. niger preys upon thrips and is commonly 208
221 found attacking T. tabaci throughout Iran, suggesting that it is adapted to the different climatic conditions found across the country. From late June onwards, O. niger naturally colonizes crops such as Taget, Marguerite flowers and Sunflower in the field, and Carnation and Chrysanthemum in greenhouses. Table 1. Life history parameters of Orius niger at three temperatures. T. o C ± ± ± =No. eggs examined, 2=Egg incubation (days), 3= Nymphal development time (days), 4=Nymphal survival (proportion), 5=Egg hatching (proportion), 6=Pre-oviposition period (days), 7=Total oviposition period (days), 8=Total life cycle (days), 9=Sex ratio (proportion females). Similar cases have been observed at locations in other countries around the world, such as in Italy (Tavella et al., 2003) and Belgium (Van de Veire & Degheele, 1992). Because of poor levels of pollen availability, O. niger are rarely found in some vegetable plants like greenhouse cucumber. Van Schelt (1993) showed that second generation of O. niger could not successfully establish on cucumber. In greenhouses of ornamental plants in Iran (Chrysanthemum and Carnation) O. niger was the most common species in Mahallat and Tehran (Pakdasht area). On the basis of our results and observations, it was concluded that O.niger is an ideal predator to use as a biological agent in greenhouse crops which have enough pollen, or alternatively it could be mixed with banker plants. Acknowledgements We would like to thank the Plant Pests and Diseases Research Institute (PPDRI) for funding this work. References Baniameri, V. 2003: Investigation on biological characteristics of the predatory bug Orius niger Wolff and its predatory behavior on onion thrips Thrips tabaci Lind. under laboratory condition. Ph.D thesis, University of Sh. Chamran Ahwaz, pp Baniameri, V., 2005: Evaluation of integrated control of ornamental plants pests and diseases in Iran, challenges and strategies. 1 th symposium on Perspective of ornamental plants industry development of Iran. p. 42. Baniameri, V., Soleyman-nejadian, E., Janssen, A., Mohaghegh, J. & Sabelis, M.W. 2003: Study on predatory behavior on Orius niger Wolff on onion thrips Thrips tabaci Lind. in laboratory conditions. Appl. Entomol. Phyt. 70 (2). Broadbent, A.B., Allen, W.R. & Foottit, R.G., 1987: The association of Franklineilla occidentalis (Pergande) (Thysanoptera; Thripidae) with greenhouse crops and the tomato spotted wilt virus in Ontario. Can. Entomol. 119:
222 Cocuzza, G.E., De Clercq, P., Lizzio, S., Van de Veire, M,. Tirry, L., Degheele, D. & Vacante, V. 1997: Life table and predation activity of Orius laevigatus and Orius albidipennis at three constant temperatures. Entomol. Exp. Appl. 85: German, T.L., Ulmann, D.E. & Mayer, J.W. 1992: Tospoviruses: diagnosis, molecular biology, phenology, and vector relationships. Annu. Rev. Phytopathol. 30: Loomans, A.J.M.; Lenteren, J.C. van; Tommasini, M.G.; Maini, S. & Riudavets, J. 1995: Biological control of thrips pests. Wageningen Agricultural University Papers, 201 pp. Royer, T.A., Edelson, J.V. & Catwright, B. 1986: Damage and control of Thrips tabaci Lindeman on spring onions. J. Rio Grande Valley Hort. Soc. 39: Tavella, L., Basco, L. & Faure, E. 2003: Distribution and population dynamics of Orius spp. in sweet pepper greenhouses in north-west Italy. IOBC/WPRS Bull. 26(10): Van de Veire, M. & Degheele, D. 1992: Biological control of the western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), in glasshouse sweet peppers with Orius spp. (Hemiptera: Anthocoridae). A comparative study between O. niger (Wolff) and O. insidiosus (Say). Biocontrol Sci. Technol. 2: Van den Meiracker, R.A.F. 1999: Biocontrol of western flower thrips by heteropteran bugs. Ph.D thesis, University of Amsterdam, The Netherlands. 147 pp. Van den Meiracker, R.A.F. & Ramakers, P.M.J. 1991: Biological control of the western flower thrips Frankliniella occidentalis, in sweet pepper, with the anthocorid predator Orius insidiosus. Med. Fac. Landbouw. Rijksuniv. Gent 56: Van Rijn, P.C.J. 2002: The impact of supplementary food on a prey-predator interaction. Ph.D Thesis, University of Amsterdam, The Netherlands, 251 pp. Yasunaga, T. 1997: The flower bug genus Orius Wolff (Heteroptera: Anthocoridae) from Japan and Taiwan, part III. Appl. Entomol. Zool. 32:
223 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Predatory capacity and longevity of adults of Orius thyestes (Hemiptera, Anthocoridae) on different prey Elizabeth C. Pedroso, Vanda H. P. Bueno, Robson J. Silva, Alessandra R. Carvalho, Alexandre J. F. Diniz, Murilo P. F. Silva, Lívia M. Carvalho Laboratory of Biological Control, Department of Entomology, Federal University of Lavras, P.O.Box 3037, , Lavras, MG, Brasil. Abstract: The predator Orius thyestes was recently found in Brazil but knowledge of its biology and food consumption including its different prey is still limited. The purpose of this work was to evaluate consumption and male and female longevity of O. thyestes preying upon Aphis gossypii, Schizaphis graminum and Caliothrips phaseoli, and eggs of Anagasta kuehniella. The experiment was conducted in a climatic chamber at 28 ± 1C, 70±10% RH and with photophase of 12h. Both male and female O. thyestes showed highest levels of consumption (P< 0.05) for A. kuehniella eggs, these were followed by C. phaseoli and the aphids A. gossypii and S. graminum. As for C. phaseoli and A. kuehniella eggs, consumption was higher amongst females than males. Nevertheless, there was no difference in male and female consumption when aphids were the prey. There was greater longevity amongst females than males when they were fed on C. phaseoli and A. kuehniella eggs. A. kuehniella eggs and C. phaseoli seemed to be more nutritionally fit to the predator, which was shown by good results for the biological parameters analyzed. Key words: Predation, aphids, thrips, Lepidoptera eggs Introduction The presence of the predator Orius thyestes Herring was recently registered in Brazil (Silveira et. al., 2003a), but knowledge of its biology is currently limited. Orius spp. are associated with several field crops: corn, sunflower, beans, alfalfa, sorghum; protected cultivation (chrysanthemum, tango, carthamus); and also weeds (farmer s friend, amaranth and Joseph s coat) which offer alternate food sources and habitat for their prey, such as thrips (Silveira et al., 2003b). Anthocorids can feed on a large number of pest-prey, although they probably prefer one or a more limited number of species, and this may influence their biological cycle (Richards & Schmidt 1996). It is therefore necessary to make a careful evaluation of the nutritional ecology of generalist predators before using them in pest management programs. According to Bueno (2000), information about nutrition and ecology constitutes an important element of studies aimed at increasing and optimising the use of Orius spp. as biological control agents. This work presents the results of studies into the predatory capacity of males and females of O. thyestes and their longevity in relation to different prey. It aims to promote their use as biological control agents mainly on protected cultivations. Material and methods The specimens used in this study originated from stock reared by the Biological Control Laboratory of the Department of Entomology at the Federal University of Lavras, Minas Gerais, Brazil. The experiment was carried out in a climatic chamber at 28±1ºC, 70 ± 10% RH and with a photophase of 12 hours. 211
224 Predation of aphids and thrips Adult male and female O. thyestes were individualized in 5-cm diameter Petri dishes containing cucumber (Cucumis sativus L.) var. Caipira, sorghum (Sorghum bicolor L.) var. BR 303 or Canavalia ensiformes (L.), to evaluate predator consumption of Aphis gossypii Glover, Schizaphis graminum (Rondani) and Caliothrips phaseoli (Hood), respectively. The leaf discs were maintained over a 1% agar layer with ten adult aphids (A. gossypi or S. graminum) or ten adult thrips (C. phaseoli) per disc, according to the test. Twenty replicates were made for each treatment. The number of prey consumed (body contents partially or totally removed leaving only the integument) was recorded on a daily basis. After each evaluation, all prey were removed and substituted by fresh adults. The longevity of both male and female predators was also evaluated in relation to consumption of different prey. Consumption of eggs of Anagasta kuehniella Male and female O. thyestes were individualized in 5-cm diameter Petri dishes containing 20 A. kuehniella eggs as prey with 20 replicates. Eggs were placed on cucumber leaf discs over a 1% agar layer for better visualization of consumed and unconsumed eggs. Eggs were considered preyed upon during the daily evaluations when only their chorion was left or when their content had been partially removed. The remaining eggs were removed on a daily basis and substituted. Both sexes were evaluated for egg consumption. Results and discussion Prey consumption Both male and female O. thyestes consumed the largest number of A. kuehniella eggs (P<0.05), followed by C. phaseoli, A. gossypii and S. graminum (Table 1). The highest level of consumption by O. thyestes females was observed when C. phaseoli and A. kuehniella were used as prey as opposed to aphids (S. graminum and A. gossypii) (Table 1). No statistical difference (P< 0.05) was found in daily consumption between male and female O. thyestes when fed with S. graminum and A. gossypii. The preference for C. phaseoli may be explained by the fact that many species of Orius are found in fields associated with several species of thrips (Silveira et al., 2005). Nevertheless, the lesser preference for aphids by the predator may indicate, as observed by Evans & Gunther (2005), that predators use aphids as survival prey in periods of food shortage: this may also be true for certain heteropteran bugs. Mendes et al. (2002) observed that nymphs of Orius insidiosus (Say) eat four times more A. gossypi than A. kuehniella eggs and twice as many as thrips. Table 1. Daily consumption of O. thyestes adults fed with different prey (28± 1 o C, 70± 10% RH and with a 12-h photophase). Means followed by the same lower case letter in columns and capital letters in rows are not statistically different by the Scott-Knott test (P< 0.05). Prey Males Females Adults of S. graminum 0.7 ± 0.4 ca 0.7 ± 0.3 ca Adults of A. gossypii 0.7 ± 0.5 ca 0.6 ± 0.4 ca Adults of C. phaseoli 1.5 ± 0.9 bb 2.0 ± 0.7 ba Eggs of A. kuehniella 2.7 ± 1.0 ab 4.0 ± 1.1 aa 212
225 Table 2. Longevity (days) of O. thyestes adults fed with different preys (28± 1 o C, 70± 10% RH and with a 12-h photophase). Means followed by the same lower case letter in columns and capital letters in rows are not statistically different by the Scott-Knott test (P< 0.05). Prey Males Females Adults of S. graminum 7.0 ± 3.7 aa 9.0 ± 5.6 ba Adults of A. gossypii 8.0 ± 3.7 aa 8.0 ± 3.9 ba Adults of C. phaseoli 9.0 ± 4.0 ab 13.0 ± 4.5 aa Eggs of A. kuehniella 11.0 ± 6.6 ab 14.0 ± 7.4 aa This indicates that there may be different nutritional requirements between these two species of Orius and also between stages of the predator: nymphs and adults. This lack of difference in the consumption of the two species of aphids by O. thyestes was also observed by Mendes et al. (2003) for the consumption of A. gossypii by male and female O. insidiosus. According to Bilde & Toft (2001), many generalist predators feed on a wide diversity of prey which frequently offer better nutritional qualities than aphids. The results for male and female O. thyestes and for the consumption of thrips were similar to those reported by Mendes & Bueno (2001), who found consumption rates of 5.1 thrips/male/day and 7.6 thrips/female/day for O. insidiosus. Female ate more A. kuehniella eggs than males. The reason for this may be a major nutritional requirement for gonads to mature and for oviposition to take place: the same hypothesis was discussed by Saucedo & Reyes (1987). Egg consumption by O. thyestes females was greater than that observed by Shchenikova & Stepanycheva (2005) for O. laevigatus preying on eggs of Sitotroga cerearella (Olivier) (2.4 eggs). Meanwhile, Carvalho et al. (2005) reported a consumption of 10.6 and 15.3 A. kuehniella eggs per day by the 4th and 5th instars of O. thyestes, respectively. Longevity There were no statistical differences for the prey offered to male O. thyestes (P< 0.05) but the longevity of females was greater (P< 0.05) when fed with thrips or A. kuehniella eggs (Table 2). Greater longevity was observed by Tommasini & Nicoli (1993) for O. insidiosus fed A. kuehniella eggs (42 days) as opposed to F. occidentalis (17.1 days). The high nutritional quality of Lepidoptera eggs is related to their high Nitrogen index: nitrogen is an important nutrient throughout the metabolic process (Parra, 1999) and this explains their use by several laboratories throughout the world for the mass rearing of Orius spp. Yano et al. (2002) reported that females had a longevity of up to 20.3 days when fed with T. palmi nymphs and 27.9 with A. kuehniella eggs. According to Ruberson et al. (2000), female anthocorids store more lipids than their male counterparts and this may contribute to their greater longevity. The results obtained in this experiment with O. thyestes indicate that their voracity varies with the prey used. We can speculate about possible preferences of the prey evaluated, but other studies should be conducted in order to determine their potential role as biological control agents, given the need to demonstrate sustainability in research projects investigating methods for controlling thrips in existing projects involving protected crops in Brazil. Acknowledgments We would like to thank the CNPq for grants to the authors and for financially supporting this study. 213
226 References Bueno, V.H.P. 2000: Controle biológico de pragas: produção massal e controle de qualidade. Ed. UFLA: 207p. Bilde, T. & Toft, S. 2001: The value of three aphid species as a food for generalist predator. Phys. Entomol. 26: Carvalho, L.M., Bueno, V.H.P. & Mendes, S.M. 2005: Desenvolvimento, consumo ninfal e exigências térmicas de Orius thyestes Herring (Hemiptera: Anthocoridae). Neotrop. Entomol. 34: Evans, E.W. & Gunther, D.I. 2005: The link between food and reproduction in aphidophagous predators: a case study with Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 102: Mendes, S.M. & Bueno, V.H.P. 2001: Biology de Orius insidiosus (Say) (Hemiptera: Anthocoridae) alimentado com Caliothrips phaseoli (Hood) (Thysanoptera: Thripidae). Neotrop. Entomol. 30: Mendes, S.M., Bueno, V.H.P., Argolo, V.M. & Silveira, L.C.P. 2002: Type of prey influences biology and consumption rate of Orius insidiosus (Say) (Hemiptera, Anthocoridae). Rev. Bras. Entomol. 46: Mendes, S.M., Bueno, V.H.P., Carvalho, L.M. & Silveira, L.C.P. 2003: Efeito da densidade de ninfas de Aphis gossypii Glover, 1877 (Hemiptera, Aphididae) no consumo alimentar e aspectos biológicos de Orius insidiosus (Say, 1832) (Hemiptera, Anthocoridae). Rev. Bras. Entomol. 47: Parra, J.R.P. 1999: Técnicas para criação de insetos para programas de controle biológico. Ed. FEALQ: 137. Richards, P.C. & Schmidt, J. 1996: The effect of selected dietary supplements on survival and reproduction of Orius insidiosus (say) (Hemiptera: Anthocoridae). Can. Entomol. 128: Ruberson, J.R., Shen, Y.J. & Kring, T.J. 2000: Photoperiodic sensitivity and the diapause in predator Orius insidiosus (Heteroptera: Anthocoridae). Ann. Entomol. Soc. Am. 93 (5): Saucedo, G.J. & Reyes, V.F. 1987: Resposta funcional de Orius insidiosus (Hemiptera: Anthocoridae) sobre Caliotrips phaseoli (Thysanoptera: Tripidae). Fol. Entomol. Mex. 71: Shchenikova, A. & Stepanycheva, E.N. 2005: Rearing of predator bug Orius laevigatus (Fieb.) (Heteroptera, Anthocoridae) with alternative food and its application against Frankliniella occidentalis (Pergande). IOBC/WPRS Bull. 28(1): Silveira, L.C.P., Bueno, V.H.P., Lousada, J.N.C. & Carvalho, L.M. 2005: Percevejos predadores (Orius spp.) (Hemiptera: Anthocoridae) e tripes (Thysanoptera): Interação no mesmo habitat? Rev. Árv., Viçosa, 29 (5): Silveira, L.C.P., Bueno, V.H.P., Pierre, L.S.R. & Mendes, S.M. 2003a: Record of two species of Orius Wolff (Hemiptera, Anthocoridae) in Brasil. Rev. Bras. Entomol. 47: Silveira, L.C.P., Bueno, V.H.P., Pierre, L.S.R. & Mendes, S.M. 2003b: Plantas cultivadas e invasoras como habitat para predadores do gênero Orius spp. (Heteroptera, Anthocoridae). Brag., Campinas, 62 (2): Tommasini, M.G. & Nicoli, G. 1993: Adult activity of four Orius species reared on two preys. IOBC/WPRS Bull. 16(2): Yano, E., Nagai, K., Watanabe, K. & Yara, K. 2002: Biological parameters of Orius spp. for control of thrips in Japan. IOBC/WPRS Bull. 25(1):
227 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Disinfection of oviposition substrate with sodium hypochlorite: effects on some biological traits of Orius thyestes Alexandre J. F. Diniz, Vanda H. P. Bueno, Alessandra R. Carvalho, Elizabeth C. Pedroso, Robson J. Silva, Lívia M. Carvalho Laboratory of Biological Control, Department of Entomology, Federal University of Lavras, P.O.Box 3037, , Lavras, Minas Gerais, Brazil. Abstract: The predator Orius thyestes (Hemiptera: Anthocoridae) was recently found in Brazil and its potential as a biological control agent as well as the possibilities of rearing it under laboratory conditions are currently being investigated. Rearing implies using farmer s friend inflorescences (Bidens pilosa), previously disinfected with sodium hypochlorite, before introducing females of this predator into the rearing chamber. This work aimed to evaluate the effects of applying commercial formulations and analytical sodium hypochlorite on biological traits of O. thyestes. The experiment was carried out in a climatic chamber at 27±1º C, 70±10% RH and with a photophase of 12 hours. For disinfection, the substrates were immersed in 0.12% of commercial solutions of sodium hypochlorite (Tandira and Q boa brands) and analytical sodium hypochlorite (PA) as treatments, and in distilled water as a control. Tandira proved inadequate for our purposes due to its negative interference with the reproduction and longevity of the predator. Key words: Orius thyestes, sodium hypochlorite, Bidens pilosa, Tandira, Q boa. Introduction Predators of genus Orius have been investigated as biological control agents for thrips in protected crops in Brazil (Carvalho et al., 2005; Mendes et al., 2005) and Orius thyestes Herring, which has recently been found in this country (Silveira et al., 2003), is a potential candidate for this purpose. Studies also focus on several biological and behavioural aspects. Besides serving as shelter and protection, several plant species are also food sources (pollen) or provide an oviposition substrate for Orius spp. These insects exhibit endophytic oviposition and according to Isenhour & Yeargan (1981) and Coll (1998) species with this type of behaviour are very specific in their oviposition sites. Viability of O. insidiosus (Say) eggs depends on the oviposition site and the characteristics of the substrate. Its consistency and humidity may also influence its acceptance by the predator (van den Meiracker & Sabelis, 1993). Mendes et al. (2005) found farmer s friend inflorescences, Bidens pilosa L., to be better than other oviposition substrates for O. insidiosus in laboratory rearing. High density insect populations in laboratory rearing favour the development of fungi and bacteria which can be easily decimated if control measures, such as appropriately cleaning the rearing site, are not implemented to prevent such problems (Greenberg, 1970; Funke, 1983). In our laboratory rearing, oviposition substrates for Orius spp. were previously disinfected with sodium hypochlorite, but it seems that some commercial formulations may interfere with certain biological traits of this predator. This work aimed to evaluate two commercial formulations and analytical sodium hypochlorite (PA) when used to disinfect oviposition substrates for O. thyestes. 215
228 Material and methods The experiment was conducted in the Biological Control Laboratory of the Department of Entomology, Federal University of Lavras, Minas Gerais, Brazil. It was carried out in a climatic chamber at 27±1ºC, RH 70±10% and with a 12 hours photophase. All insects used in the experiment were from our laboratory stock rearing and were raised under similar conditions. Nymphs and adults were fed Anagasta kuehniella (Zeller) eggs. Twigs of Bidens pilosa were used as oviposition substrate with the inflorescences removed to prevent the availability of pollen as an alternate food source. Twigs were dipped into a 0.12% sodium hypochlorite solution for four minutes and then washed in distilled water before daily submission to females. Two commercial brands (Tandira and Q boa ) and analytical sodium hypochlorite (PA) were used for the treatments and distilled water was used as a control. Male and female O. thyestes were kept in single 5 cm diameter Petri dishes containing B. pilosa twigs and A. kuehniella eggs, with 25 replicates per treatment. The number of eggs laid was counted until the females died. The pre-oviposition and oviposition periods, and the longevity of the predator were also evaluated. Results and discussion Our results indicated different effects on insects reared in oviposition substracts when they were disinfected with Tandira and Q boa, although the commercial formulations of sodium hypochlorite tested listed the same compositions on their labels. This confirmed observations made during routine stock rearing in our laboratory. Results indicated that the use of Tandira may jeopardize O. thyestes rearing since females subjected to this treatment laid fewer eggs than in the other treatments (Table 1). Longer preoviposition and shorter oviposition periods were also observed. When this brand was used as a disinfectant for oviposition substrates, significantly shorter (P<0.05) female longevity (7.92 days) was observed compared with the other treatments. No statistically significant differences were found among the remaining treatments. For male longevity the results showed no statistically significant difference (P<0.05) with respect to the control, 5.13 and 5.45 days, respectively. A shorter preoviposition period (2.29 days) was observed when Q boa was used: this was very similar to that of the control treatment (2.69 days). Moreover, the oviposition period was significantly longer from the control and the remaining treatments (P<0.05). When the number of eggs/female was analysed, Q boa produced similar results to analytical sodium hypochlorite (PA), which may provide opportunities for reducing costs, as the commercial product is less expensive than PA s. Thus, it is possible to minimize the occurrence of contaminant microorganisms without prejudicial effects to insects or at least reducing these effects to acceptable levels. It is easier to attain the security threshold for eggs of insect pests, since natural enemies are more sensitive to the effects of compounds with disinfecting properties (Grenier & Liu, 1990). Analytical sodium hypochlorite (PA) negatively interfered in the preoviposition and oviposition periods indicating that the concentration used may have been high. Carvalho et al. (1998) also found that commercial and analytical sodium hypochlorite used to eliminate egg pedicells of Chrysoperla mediterrânea (Hozel) (Neuroptera: Chrysopidae), reduced the hatching of eggs when used in high concentrations. 216
229 Table 1. Biological parameters of O. thyestes on treated laboratory rearing oviposition substrates with sodium hypochlorite formulations. Parameters Evaluated Control PA Sodium Hypochlorite Tandira Q'boa Mean number of eggs/female/day 3.15a 2.23a 1.74b 2.65a Preoviposition period (days) 2.69a 3.00b 3.28b 2.29a Oviposition period (days) 7.46b 6.04b 7.22b 9.81a Male longevity (days) 5.45b 6.46a 5.13b 7.92a Female longevity (days) 9.85a 9.08a 7.92b 10.77a Means followed by the same letter in the same row are not statistically different by the Scott-Knott (P<0.05) test. These results indicate that certain factors should be considered when decontaminants are used in insect rearing. Differences found between the two commercial brands, despite identical formulations being listed on their labels, demonstrate that products may contain contaminants or different concentrations of the active ingredient from those stated on the label. This indicates that careful research is required to obtain appropriate products in order to avoid undesired effects upon the insects to be reared. Singh & House (1970) and Singh & Bucker (1971) advise that products should be chosen taking into account the microorganisms targeted and insect tolerance to what would constitute effective concentrations of the product in question. This would help to optimize rearing of Orius under laboratory conditions. Acknowledgements We would like to thank the CNPq for research grants and the FAPEMIG for providing financial support for the present work. References Carvalho, L.M., Bueno, V.H.P. & Mendes, S.M. 2005: Response of two Orius species to temperature. IOBC/WPRS Bull. 28(1): Carvalho, C.F., Canard, M. & Alauzet, C. 1998: Destruction of egg pedicels by sodium hypochlorite and its effects on the hatching of eggs of Chrysoperla mediterranea (Hozel) (Neuroptera: Chrysopidae). Acta Zool. Fennica 209: Coll, M. 1998: Living and feeding on plants in predatory Heteroptera. In: Coll, M. & Ruberson, J.R. Predatory Heteroptera: their ecology and use in biological control. Lanham: Funke, B.R. 1983: Mold control for insect-rearing media. Bull. Entomol. Soc. Am. 29: Greenberg, B. 1970: Sterilizing procedures and agents, antibiotics and inhibitors in mass rearing of insects. Bull. Entomol. Soc. Am. 16: Grenier, S. & Liu, W.H. 1990: Antifungals: mold controland safe levels in artificial media for Trichogramma (Hymenoptera: Trichogrammatidae). Entomophaga 35: Isenhour, D.J. & Yeargan, K.V. 1981: Effect of crop phenology on Orius insidiosus populations on strip-cropped soybean and corn. J. Georgia Entomol. Soc. 16:
230 Mendes, S.M., Bueno, V.H.P. & Carvalho, L.M. 2005: Adequabilidade de diferentes substratos à oviposição do predador Orius insidiosus (Say) (Hemiptera: Anthocoridae). Neotrop. Entomol. 34 (3): Silveira, L.C.P., Bueno, V.H.P., Pierre, L.S.R. & Mendes, S.M. 2003: Record of two species of Orius Wolff (Hemiptera, Anthocoridae) in Brasil. Rev. Bras. Entomol. 47: Singh, P. & Bucher, J.E. 1971: Efficacy of safe levels of antimicrobial food additives to control microbial contaminants in a synthetic diet for Agria affinis. Entomol. Exp. Appl. 14: Singh, P. & House, H.L. 1970: Antimicrobial agents: their detrimental effects on size of an insect, Agria affinis. Can. Entomol. 102: van den Meiracker, R.A.F. & Sabelis, O. 1993: Oviposition sites of Orius insidiosus in sweet pepper. IOBC/WPRS Bull. 16(2):
231 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Intraguild predation between Orius insidiosus and Aphidius colemani, and biological control of Aphis gossypii Leonardo S. R. Pierre¹, Vanda H. P. Bueno¹, Marcus V. Sampaio 2, Joop C. van Lenteren 3, Bruno F. De Conti¹, Murilo P. F. Da Silva¹, Luís Cláudio P. Silveira 4, ¹Universidade Federal de Lavras, Departamento de Entomologia, Caixa Postal 3037, Lavras, MG, 2 Universidade Federal de Uberlândia, Caixa Postal 593, , ICIAG, Campus Umuarama, MG, Brazil; 3 Laboratory of Entomology, Wageningen University, Wageningen, The Netherlands, 4 APTA Regional Centro Norte, Caixa Postal 24, Pindorama, SP, Brazil. Abstract: Biological control of pests in greenhouses crops may involve the introduction of more than one species of natural enemies, and this can result in intraguild predation. This work aimed at evaluating the effect of the intraguild predation between Orius insidiosus (Say) and Aphidius colemani Viereck, natural enemies of Aphis gossypii Glover. Aphids were kept on caged cucumber plants and the treatments were: a) control without natural enemies; b) parasitoid only; c) predator only; d) parasitoid + predator. The parasitoid A. colemani appeared to be an effective biological control agent of A. gossypii even in the presence of the predator. The average number of A. gossypii on the plants in the presence of the predator O. insidiosus was similar to that as in the treatment with only aphids. The predator O. insidiosus did have no effect on the action of the parasitoid A. colemani in the reduction of A. gossypii and on the numbers of mummies, percentages of parasitism and emergence of the parasitoid. Key words: parasitoid, predator, aphids, greenhouse, cucumber plants Introduction The parasitoid Aphidius colemani Viereck is a natural enemy that may control Aphis gossypii Glover on greenhouse crops (Steenis, 1993; Sampaio et al., 2001; Saiki & Wada 2005). However, the predator Orius insidiosus (Say) is often used to control Frankliniella occidentalis (Pergande), a pest that may occur together with the aphids on the same crop (Silveira et al., 2004; Bueno, 2005). Predators like Orius of the family Anthocoridae may feed on parasitized aphids, characterizing the intraguild predation (Meyling et al., 2002). As a consequence, biological control of aphids by the parasitoid A. colemani might be negatively affected by the introduction of Orius predators to the crop system. This work aimed to evaluate the effect of intraguild predation of O. insidiosus on the effectiveness of A. colemani in its control of A. gossypii on caged cucumber plants in the laboratory. Material and Methods The aphid A. gossypii and the natural enemies A. colemani and O. insidiosus came from our laboratory rearing. The tests were carried out in a climatized room at 25 ± 1 o C, RH 70 ± 10% and 12hr photophase. Cucumber plants (Cucumis sativus L. cv. Caipira) 28 days old were used. Each plant was kept individually in an acrylic cage (30 x 30 x 60 cm), and infested with 50 aphids A. gossypii (20 adults and 30 nymphs, 10 nymphs of 4 th instar, 10 nymphs of 3 rd 219
232 instar and 10 nymphs of 2 nd instar). This procedure was adopted to reach the natural age distribution of the aphid colonies on the plants in greenhouses. The aphids were placed on the three oldest leaves of the plant. The high number of aphids ensured that excess amounts were present throughout the experimental period. The natural enemies were released on the superior part of the plant five hours after the aphid infestation on the plants. Four treatments were evaluated: a) control (only A. gossypii), b) one female of A. colemani + one female of O. insidiosus, c) one female of A. colemani and d) one female of O. insidiosus. The number of A. gossypii on the plants in each treatment was counted each two days for a period of 12 days, resulting in six counts. The number of mummies and adult aphids found on the 12 th day were used to calculate the total number of mummies and the percentage of parasitism. The number of emerged adult parasitoids was observed until the 18 th day on five plants on the treatments with the presence of A. colemani only. The tests were conducted randomly in a factorial scheme of 4 x 7 (four treatments and seven evaluations during the time) and six replicates. The results are submitted to an analysis of variance. To evaluate the number of aphids on the plants for different treatments, the averages were compared with the Tukey test. To evaluate the number of mummies, the percentages of parasitism and the emergence of parasitoids, the averages were compared with a t test. Results and discussion An interaction was found between treatment time and level of aphid infestation. The number of aphids in the plants on the control treatment and on treatments with natural enemies was not significantly different until the 6 th day after the A. gossypii infestation. From the 8 th day onwards, the treatments with the parasitoid A. colemani showed the lowest number of aphids on the plants. The number of aphids on the plants was not significantly different on the treatments with A. colemani alone or together with O. insidiosus. The number of aphids in the treatment with O. insidiosus alone was different from the control treatment at the 10 th day (Table 1). The number of aphids on the plants was increasing from the first count in the control treatment and in the treatment with only O. insidiosus. The increase can be characterized by a quadratic equation. However, the treatments with A. colemani do not show a significant increase of aphids over time (Figure 1). No significative differences were found in the number of mummies, the percentages of parasitism and emergence of A. colemani in the treatments with the parasitoid alone or together with O. insidiosus (Table 2). Biological control of aphids by the parasitoid A. colemani begins with a toxin injection in the host during the act of oviposition by the female parasitoid, and this can lead to a decrease in fecundity of the adult aphids and to prevent the parasitized nymphs to reproduce, even those that stay alive until the reproductive phase (van Steenis & El-Kawass 1995). The population of A. gossypii on the cucumber plants has probably been kept stable in the treatments with the parasitoid A. colemani due the action of toxins of the parasitoid on the reproductive system of the aphid. It is expected that the next generations of A. colemani will keep the aphid population at low levels. Saiki & Wada (2005) wrote that if mummies of A. colemani are found 2 weeks after the first introduction on strawberry crop and aphids do not increase rapidly, it can be concluded that aphids are almost controlled. One adult of O. insidiosus eats on average 11 nymphs of A. gossypii per day (Mendes et al. 2003) and can eat both on parasitized and unparasitized aphids (Pierre, 2005). However the number of mummies and the percentages of parasitism and the emergence of the parasitoid 220
233 were not affected by the presence of the predator O. insidosus. This may mean that the next generation of A. colemani will not be affected by the presence of the predator O. insidiosus. Table 1. Mean number of Aphis gossypii on cucumber plants with different combinations of natural enemies and different days after plant infestation. Days Average number of Aphis gossypii/plant (N= 6)* Control A. colemani + O. insidiosus A. colemani O. insidiosus 0 50±0.00 a 50±0.00 a 50±0.00 a 50±0.00 a ±7.80 a 133.3±15.46 a 98.2±14.43 a 117.2±9.75 a ±21.72 a 172.3±23.25 a 132.2±22.55 a 335.0±23.61 a ±23.56 a 127.5±26.25 a 130.3±22.2 a 586.2±50.34 a ±79.56 a 83.8±18.72 b 77.2±23.47 b ±97.73 a ± a 83.0±19.07 c 86.3±42.89 c ± b ± a 72.8±43.21 b 131.8±99.85 b ± a Means followed by the same letters in the rows are not significantly different (Tukey test P 0.05) Number of Aphids Control O. insidiosus + A. colemani Days Number of Aphids Orius insidiosus Aphidius colemani Days Figure 1. Mean number of Aphis gossypii on cucumber plants in the absence of natural enemies (control; y = x² x ; R² = 0.946) and in the presence of natural enemies Aphidius colemani + Orius insidiosus (y = mean), O. insidiosus (y = x² x ; R² = 0.998) and A. colemani (y = mean). The presence of the predator O. insidiosus did not affect the A. gossypii numbers on the plants and did not decrease mummy formation or the emergence of A. colemani. Thus, the results indicate that the predator O. insidiosus does not negatively interfere with biological 221
234 control of A. gossypii by the parasitoid A. colemani on caged plants in the laboratory. Experiments at the greenhouse scale and for extended periods of time are now necessary to evaluate the interaction between these two natural enemies under practical conditions Table 2. Mean number of mummies and percentages of parasitism and emergence of Aphidius colemani on Aphis gossypii on cucumber plants with and without Orius insidiosus. Natural enemies Mummies (N)* % Parasitism (N)* % Emergence (N)* A. colemani+o. insidiosus 80.3 ± 1.66 A (6) 68.1 ± 2.26 A (6) 93.2 ± 1.19 A (5) A. colemani 94.3 ± 3.21 A (6) 69.0 ± 2.43 A (6) 72.8 ± 1.85 A (5) *Means followed by the same letters on the rows are not significantly different by t test (P 0.05). Acknowledgements We thank to CNPq for a research grant to the second author and FAPEMIG for financial support to this study. References Bueno, V.H.P. 2005: Controle biológico de tripes: pragas sérias em cultivos protegidos. Informe Agropecuário 26: Mendes, S.M., Bueno, V.H.P., Carvalho, L.M. & Silveira, L.C.P. 2003: Efeito da densidade de ninfas de Aphis gossypii Glover, 1877 (Hemiptera: Aphididae) no consumo alimentar e aspectos biológicos de Orius insidiosus (Say, 1832) (Hemiptera: Anthocoridae). Rev. Brasil. Entomol. 47: Meyling, N.V., Brodsgaard, H.F. & Enkegaard, A. 2002: Intraguild predation between the predatory flower bug, Anthocoris nemorum, and the aphid parasitoid, Aphidius colemani. IOBC/WPRS Bull. 25: Pierre, L.S.R. 2005: Interação entre Orius insidiosus (Say) (Hemiptera: Anthocoridae) e Aphidius colemani Viereck (Hymenoptera, Braconidae, Aphidiinae). Lavras, UFLA (MS Thesis). Saiki, Y. & Wada, T. 2005: Biological control in strawberry in Japan. IOBC/WPRS Bull. 28 (1): Sampaio, M.V., Bueno, V.H.P. & van Lenteren, J.C. 2001: Preferência de Aphidius colemani Viereck (Hymenoptera: Aphidiidae) por Myzus persicae (Sulzer) e Aphis gossypii Glover (Hemiptera: Aphididae). Neotrop. Entomol. 30: Silveira, L.C.P., Bueno, V.H.P. & van Lenteren, J.C. 2004: Orius insidiosus as biological control agent of Thrips in greenhouse chrysanthemums in the tropics. Bull. Insectol. 57: van Steenis, M.J. 1993: Suitability of Aphis gossypii Glov., Macrosiphum euphorbiae (Thom.) and Myzus persicae Sulz. (Hom.: Aphididae) as host for several aphid parasitoid species (Hym.: Braconidae). IOBC/WPRS Bull. 26: van Steenis, M.J. & El-Kawass, K.A.M.H. 1995: Life history of Aphis gossypii on cucumber: influence of temperature, host plant, and parasitism. Entomol. Exp. Appl. 76:
235 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Differentiation of Macrolophus pygmaeus (Rambur 1839) and Macrolophus melanotoma (Costa 1853) (Heteroptera: Miridae) based on molecular data Jose Isidro Martinez-Cascales, José Luis Cenis, Juan Antonio Sanchez Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (IMIDA), Department of Biotechnology and Crop Protection, c/ Mayor s/n, La Alberca, Murcia, Spain. juana.sanchez23@carm.es Abstract: Several species of the Macrolophus genus are well known predators of small pests such as whiteflies, aphids and mites. Macrolophus melanotoma (Costa 1853) (Syn. Macrolophus caliginosus Wagner 1951) and Macrolophus pygmaeus (Rambur 1839) are the most economically important species. In spite of their economic importance the classification of these two species has not received comprehensive treatment until recently. The lack of discontinuity in the taxonomical characters used to differentiate M. melanotoma from M. pygmaeus made it difficult to define species boundaries and conspecificy was not excluded. In this work we use molecular information to clarify the identity of M. melanotoma and M. pygmaeus. The phylogenetic analysis using cytochrome b sequences grouped the specimens studied in two distinct clades, corresponding to insects classified as M. melanotoma and M. pygmaeus using key morphological characters. Commercial insects, labelled and sold as M. caliginosus, were classified as M. pygmaeus. Based on molecular phylogenetic and morphological evidences we conclude that M. melanotoma and M. pygmaeus are two different species. The biology, ecology and role as pest control agents of Macrolophus melanotoma and M. pygmaeus may have been messed up due to misidentification. Key words: Macrolophus spp., M. melanotoma, M. caliginosus, M. pygmaeus, DNA, phylogeny, cytochrome b, Miridae, Dicyphine, IPM. Introduction Several species of the Macrolophus genus are well known predators of small pests in vegetable crops. Macrolophus melanotoma (Costa 1853) [Syn. Macrolophus caliginosus Wagner 1951, (Carapezza, 1995)] and Macrolophus pygmaeus (Rambur, 1839) are the most economic important species (Malausa & Trottin-Caudal, 1996; Perdikis & Lykouressis, 2000; Alomar et al., 2003). Macrolophus caliginosus is used for pest control in vegetable crops worldwide. In spite of their economic importance, the taxonomical classification of these two species has not received comprehensive treatment until recently (Martinez-Cascales et al., 2006). Josifov (1992) based the differentiation of M. melanotoma and M pygmaeus in the black band-shape macula behind the eye and the colour of the first antennal segment. He reported several major findings, as follows: (1) M. melanotoma, band-shape macula behind the eye thin and colour of the first antennal segment clear in the middle part; (2) M. pygmaeus, macula as high as half the width of the eye and first antennal segment entirely black. However, several authors pointed out the inconsistency of the characters used to differentiate the two species and conspecifity was not excluded (Kerzhner & Josifov, 1999; Goula et al., 2002). The paucity of morphologic characters leads to the use of molecular techniques to clarify 223
236 ²the issue. The use of sequence variation in mitochondrial genes has proven most useful in establishing species boundaries (Simon et al., 1994; Sanchez et al., 2006; Martinez-Cascales et al., 2006). The aim of this work was to investigate whether M. melanotoma and M. pygmaeus were two different species. We also tested the identity of a Macrolophus species labelled and sold as M. caliginosus. Materials and methods Samples Macrolophus were collected on Dittrichia viscosa and tomato (Lycopersicon esculentum) in the Iberian Peninsula and Canary Islands. Only specimens that could undoubtly be classified, either as M. melanotoma or M. pygmaeus, according to the key characters listed by Josifov (1992) were used in the phylogenetic analysis: (1) insects with a thin band-shape macula behind the eye and middle part of the first antennal segment clear (M. melanotoma); (2) insects with a wide macula and first antennal segment entirely black (M. pygmaeus). Two females and two males of each group were used for phylogenetic analyses (Table 1). A female and two males of a commercial Macrolophus labelled as M. caliginosus were also analysed (Table 1). Table 1. Sample codes, GenBank accession numbers, localities and host plants, for all taxons used in the phylogenetic analyses. Commercial = commercial Macrolophus labelled as "M. caliginosus". Localities: Murcia (Southeast Spain); Valverde (Portugal); Tenerife (Canary Islands). Plants: D. viscosa = Dittrichia viscosa, L. esculentum = Lycopersicon esculentum. Species Sex Specimen code GenBank number Locality Host plant M. melanotoma M017-2 DQ Murcia D. viscosa M. melanotoma M121-5 DQ Tenerife D. viscosa M. melanotoma M037-2 DQ Murcia D. viscosa M. melanotoma M125-2 DQ Tenerife D. viscosa M. pygmaeus M004-3 DQ Murcia L. esculentum M. pygmaeus M015-2 DQ Murcia L. esculentum M. pygmaeus M012-2 DQ Murcia L. esculentum M. pygmaeus M016-2 DQ Murcia L. esculentum Commercial M115-5 DQ Commercial M115-3 DQ Commercial M115-4 DQ M. costalis M240-1 DQ Valverde Cistus monspeliensis D. tamaninii D014-1 DQ Murcia Epilobium hirsutum D. cerastii D003-1 DQ Murcia Cucurbita maxima N. tenuis N006-1 DQ Murcia L. esculentum C. geniculata C001-1 DQ Murcia Ononis natrix Macrolophus costalis Fieber was used as an outgroup. This species can be clearly differentiated from other Macrolophus species by the black apex of the scutellum. Other dicyphines were also used as outgroups: Dicyphus cerastii Wagner, Dicyphus tamaninii 224
237 Wagner, Nesidiocoris tenuis (Reuter), Cyrtopeltis geniculata Fieber. DNA analysis A fragment of the mitochondrial cytochrome b region (320 bp) was amplified by PCR and sequenced as described by Sanchez et al. (2006) and Martinez-Cascales et al. (2006). Sampling information and GenBank sequence accession numbers are given in Table 1. DNA sequences were first explored using MEGA 3.1 (Kumar et al., 2004). All sequences and alignments were checked manually to ensure their correction. Maximum parsimony analysis was carried out using PAUP* 4.Ob10 version (Swofford, 2002). Initial parsimony trees were obtained by branch and bound search. Additionally, Neighbor-Joining method with Kimura 2-parameter substitution model was performed using MEGA 3.1. Nodal support in both analyses was estimated by 2000 bootstrap replicates. Results and discussion The insects classified as M. melanotoma and M. pygmaeus based on the black band-shape macula behind the eye and the colour of the first antennal segment, grouped in two distinct clades in the phylogenetic analyses (Figure 1). M. pygmaeus is a sister species of M. costalis, and M. melanotoma joins the group formed by those two species. Parsimony and Neighbor- Joining analyses produced trees with similar topologies. Nodes at the species level were supported by high bootstrap values. Figure 1 shows the maximum parsimonious tree inferred from cyt b sequence data. The phylogenetic analyses using molecular data confirm the species identity of M. melanotoma and M. pygmaeus. The commercial Macrolophus, labelled as M. caliginosus (Syn. M. melanotoma), clustered within the M. pygmaeus group (Figure 1); they also shared the wide macula and the entirely black first antennal segment. This result reveals that some of the commercial products are labelled with the wrong species name and we suspect this could be more general for other insects commercialised as M. caliginosus. This unfortunate event may have been a great source of mistakes in characterizing the biology, ecology and role as pest control agents of Macrolophus species because, many times, insects used in experiments came from natural enemy producers. The correct classification of the species is one of the key factors for the success of biological pest control programs. Failures in biological control programs due to misidentification of natural enemies are well documented (DeBach 1964). There are several reasons that make the identification of Macrolophus species highly important: (1) being zoophytophagous they feed on plants and the degree of phytophagy may greatly vary between species. Goula & Alomar (1994) argued that plant damage information for M. melanotoma and M. pygmaeus may have been assigned to the wrong species due to misidentification; (2) the characterization of Macrolophus species ecology is prior to developing pest management programs based on natural biological control. Many species of Dicyphus and Macrolophus are restricted to a low number of host plants (Sanchez et al., 2006; Martinez-Cascales et al., 2006). Host plant restrictedness in Macrolophus species may be driven by plant preference behaviour, like it has been reported for other dicyphines such as Dicyphus hesperus (Sanchez et al., 2004). We predict that differences in host plant preference between M. melanotoma and M. pygmaeus would limit the movements of each species to the most preferred hosts. The lack of discontinuous taxonomical characters may sometimes difficult the differentiation between M. melanotoma and M. pygmaeus using morphological traits (Martinez-Cascales et al., 2006). Molecular techniques can be used to solve the species identity when morphology characters alone were irresolute. 225
238 93.9 Dicyphus cerastii Dicyphus tamaninii Nesidiocoris tenuis 84.9 Cyrtopeltis geniculata M.melanotoma M121-5 M.melanotoma M125-2 M.melanotoma M017-2 M.melanotoma M037-2 Macrolophus costalis M.pygmaeus M Commercial M115-3 Commercial M Commercial M115-5 M.pygmaeus M012-2 M.pygmaeus M015-2 M.pygmaeus M004-3 Figure 1. Phylogeny of Macrolophus species based on cyt b sequences. Maximum parsimony tree with bootstrap values. Outgroups: Macrolophus costalis, Cyrtopeltis geniculata, Nesidiocoris tenuis, Dicyphus cerastii and Dicyphus tamaninii. Acknowledgements We thank Gerry Cassis (Australian Museum, Sydney) for useful comments on dicyphine taxonomy and biology. We thank Fernando Cánovas and Pilar de la Rúa for advice on molecular analyses. Estrella Hernández and Alfredo Reyes-Betancort helped collecting 226
239 specimens in the Canary Islands. Fundación Séneca (Agencia Regional de Ciencia y Tecnología. Región de Murcia, Ref /EE2/05) granted a predoctoral fellowship to J. I. Martinez-Cascales and granted a research stay of J. A. Sanchez at the Australian Museum (Sydney). This work was funded by the CICYT Project Nº AGL CO3-03. References Alomar, O., Riudavets, J. & Castañé, C. 2003: Macrolophus caliginosus in the biological control of Bemisia tabaci in greenhouse melons. IOBC/WPRS Bull. 26(10): DeBach, P. 1964: Biological control of insect pests and weeds. Chapman and Hall Ltd. London. Carapezza, A. 1995: The specific identities of Macrolophus melanotoma (A. Costa, 1853) and Stenodema curticolle (A. Costa, 1853) (Insecta Heteroptera, Miridae). Naturalista XIX Supl. IV(3-4): Goula, M. & Alomar, O Miridos (Heteroptera: Miridae) de interés en el control integrado de plagas en tomate. Guía para su identificación. Bol. San. Veg. Plagas. 20(1): Goula, M., Alomar, O. & Castañé, C. 2002: Predatory mirids in tomato crops. European Whitefly Studies Network ( Ed.: John Innes Centre, U.K. Josifov, M. 1992: Zur Taxonomie der paläarktischen Macrolophus-Arten (Insecta, Heteroptera: Miridae). Reichenbachia 29 (1):1-4. Kerzhner I.M. & Josifov, M. 1999: Cimicomorpha II: Miridae. In: Catalogue of the Heteroptera of the Palaearctic Region. Vol III, eds. Aukema & Rieger: 577. Kumar, S., Tamura, K. & Nei, M. 2004: MEGA 3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Briefings in Bioinformatics 5: Malausa, J.C. & Trottin-Caudal, Y. 1996: Advances in the strategy of use of the predaceous bug Macrolophus caliginosus (Heteroptera: Miridae) in glasshouse crops. In: Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management, eds. Alomar & Wiedenmann: Martinez-Cascales, J.I., Cenis, J.L., Cassis, G. & Sanchez, J.A. 2006: Species identity of Macrolophus melanotoma (Costa 1853) and Macrolophus pygmaeus (Rambur 1839) (Insecta: Heteroptera: Miridae) based on morphological and molecular data and bionomic implications. Submitted to Insect Systematic and Evolution. Perdikis, D. & Lykouressis, D. 2000: Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Control 17 (1): Sanchez, J. A., Gillespie D. R. & McGregor R. R. 2004: Plant preference in relation to life history traits in the zoophytophagous predator Dicyphus hesperus. Entomol. Exp. Appl. 112: Sanchez, J.A., Cassis, G. & Martinez-Cascales, J.I. 2006: Description of a new species of Dicyphus Fieber (Insecta: Heteroptera: Miridae) from Portugal based on morphological and molecular data. Submitted to Insect Systematics and Evolution. Simon, C., Frati, F., Beckenbach, A., Crespi, B., Liu, H. & Flook, P. 1994: Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 87 (6): Swofford, D.L. 2002: PAUP*. Phylogenetic Analysis Using Parsimony (* and other methods). Version 4.0b10. Sunderland, Massachusetts (USA): Sinauer Associates, Inc. 227
240 228
241 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Reproductive traits of the generalist predator Macrolophus caliginosus Cristina Castañé 1, Oscar Alomar 1, Jordi Riudavets 1, César Gemeno 2 1 Departament de Protecció Vegetal, IRTA-Centre de Cabrils, Ctra. Cabrils s/n, Cabrils, Barcelona, 2 Universitat de Lleida, Centre UdL-IRTA, Rovira Roure 191, Lleida, Spain. cristina.castane@irta.es Abstract: The objectives of this study were to describe the copulatory behaviour and maturation of the sexual organs of the predator Macrolophus caliginosus with the aim of implementing its use in augmentative releases in greenhouses. Results indicate that males actively pursue females but that courtship is almost non-existent; more mating take place during night hours and in the first half of the day than during the second half of the day; 5 and 7 day-old females tend to mate more than younger ones; and the mating propensity of males is not affected by age. This is consistent with the physiological maturation of sexual organs. Key words: mating, development of sexual organs, copulation frequency, mirid bugs Introduction Macrolophus caliginosus Wagner (Heteroptera: Miridae) is a polyphagous predator native to the Mediterranean region where it spontaneously colonizes open field and protected vegetable crops. Several companies produce this natural enemy that is widely used in Europe. Nevertheless, its use in greenhouses is somewhat unpredictable and a rather slow installation of the population is observed in some circumstances. Due to a lack of knowledge of its reproductive behaviour, it is difficult to overcome many of the difficulties that tend to be associated with releasing this predator. Its reproduction characteristics remain relatively unknown and this affects the performance of the predator both in the greenhouse and in commercial mass rearing. In laboratory fertility trials we observed a large variation in female fertility, with up to 30% of non egg-laying females. In this work we have studied the mating behaviour and maturation of the sexual organs of M. caliginosus with the aim of implementing its use in augmentative releases in greenhouses. Material and methods All M. caliginosus individuals used were derived from our permanent colonies that are maintained on tobacco plants (Nicotiana tabacum L.) with Epestia kuehniella Zeller (Lepidoptera: Pyralidae) eggs as prey. Rearing was done in a climatic chamber maintained at 25 ± 1 C, with 70 ± 10% R.H., and with a 16:8 h (L:D) photoperiod. Mating behaviour Five to seven day old virgin females where isolated in individual ventilated cages (4 x 7 cm diameter) containing a 5 cm diameter tobacco leaf disc on top of a 0.5% agar layer and with E. kuehniella eggs as prey. After 24 hours two virgin males were gently introduced through a hole in the cage. Their behaviour was monitored for one hour by both video camera and direct observation. 229
242 Diel pattern and effect of age on mating frequency Observations were performed on a small tobacco plant with only one remaining leaf, using E. kuehniella eggs as prey. The plant was covered with a Plexiglas cylinder to prevent the insects from escaping. We chose three time intervals to determine whether mating frequency was affected by time of day: two with daylight (from 8 a.m. to 4 p.m., and from 4 p.m. to 12 p.m.) and one with darkness (from 12 p.m. to 8 a.m.). In this last case, a red fluorescent light was used to avoid disturbing the insect s behaviour. Four (6- to 8-day-old) females were placed on the plant the day before the experiment was performed in order to habituate them to the setup. On the day of the experiment four males were transferred to a plastic test tube (1.5 x 6 cm) where they stayed for min. The test tube was then gently tapped above the plant so that the males fell onto it. To study the effect of age on female mating, cylinders were prepared with three 1-, 3-, 5-, or 7-day-old females the day before the experiment. On the day of the experiment three males were aspirated into a glass tube (5 mm x 7 cm) where they were left for 45 min. They were then gently encouraged, with the help of a plunger, to walk onto the plant. To study the effect of age on male mating, cylinders were prepared containing three 6- to 8-day-old females the day before the observation. On the day of observation three 1-, 3-, 5-, or 7-day-old males were aspirated into glass tubes and released as before. The behaviour of males and females was examined for 30 minutes by direct observation, recording the number of contacts and copulas between males and females, and the duration of each copula. Each treatment (period of day, female age and male age) was replicated 20 times, with each cylinder constituting one replication. Insects were discarded after being used. Physiological development of sexual organs We used females and males of different ages to evaluate the physiological development of the sexual organs. Insects were dissected in saline solution. Their abdomens were opened laterally and their sexual organs removed, teased apart, and observed under a binocular microscope. In the case of males, the seminal vesicle was squashed with a cover slip and the presence of mobile spermatozoa observed under a compound microscope. Results and discussion Mating behaviour Only ten of 55 females actually mated in this set-up. In general, females stood still on the leaf during the observation period whereas males were more active and walked around the plant. Males and females often stayed close to each other without any activity for long periods of time: this suggested that if, as in other mirids, females produce sex pheromones, these were either not released continuously or played only a minor role once the insects were in close contact. However, when contact was made, usually because the male touched the female, the male became very active: this suggested the presence of contact pheromones. The female generally rejected the male at first, running away or shaking her abdomen, but after a few more contacts the female finally accepted. Courtship was almost non-existent. The male walked on top of the female, slid his abdomen downwards always by her right side, inserted his genitalia on hers, and turned to the right, so that they remained united in the same direction but opposed to one another (Figure 1). This process happened very rapidly, in a few seconds. This mating position is common in plant bugs (Wheeler, 2001). Once in this position they remained still during the whole mating period. Towards the end of mating the female became more active, kicking the male with her hind legs and pulling the male behind her. The copula lasted an average of 5.53 ± min. 230

243 Figure 1. Position of a mating pair of Macrolophus caliginosus: female on the left, male on the right. Diel pattern and effect of age on mating frequency Maintaining natural conditions as much as possible seems to be important for mating. In these trials, more copulas were observed than in the previous trial with small cages. Diel pattern. The number of contacts between males and females was similar for all three times of day considered, indicating that the level of activity was relatively constant. Although there were copulas in all three of the periods considered, there was a greater tendency for mating to occur during the dark period (12 p.m. to 8 a.m.) and during the first half of the day (8 a.m. to 4 p.m.) than during the second half of the day (4 p.m. to 12 p.m.) (Figure 2). This pattern of higher activity during the dark period has also been observed in a closely related species, Dicyphus hesperus, whose females are more active, and find and consume prey at higher rates at night than during the day (VanLaerhoven et al., 2003). % Females that mate pm<8am 8am<4pm 4pm<12pm Figure 2. Percentage of 6-to 8-day-old M. caliginosus females mated according to diel pattern when offered males. Effect of age. Young females were very reluctant to mate and the number of contacts required before they accepted males was higher in young females than in older ones. The percentage of 231
.")
 Figure 3. Percentage of M.")
, vitellogenic follicles of a threeday-old female (b), and detail of a chorionated follicle (c) of M. caliginosus.")
244 males that mated was hardly affected by age, although older males mated more frequently than younger ones. Conversely, the percentage of females that mated varied according to age: fewer copulas were observed with 1 and 3 day-old females than with those that were 5 and 7 days old (Figure 3). In both trials, we observed that after mating, all females vigorously rejected any further attempts at mating by males. On the other hand, males were able to mate again immediately afterwards. % Adults that mate Age of adults (days) Figure 3. Percentage of M. caliginosus males and females that mated with 6-8 day-old individuals of the opposite sex, according to their age. a b c Figure 4. Previtellogenic ovarioles of a one-day-old female (a), vitellogenic follicles of a threeday-old female (b), and detail of a chorionated follicle (c) of M. caliginosus. Physiological development of sexual organs Females. Each ovary consisted of seven ovarioles, as in most other Miridae, and was of the telotrophic type, which is usual in Hemiptera (Wightman, 1973). One-day-old females had 232
 that became chorionated follicles when the chorion was")
245 previtellogenic ovarioles (Figure 4a) in which oocytes were still connected to the germarium by nutritive cords and vitellogenesis had not yet begun (Ma & Ramaswamy, 1987). As females aged, oocytes enlarged by receiving nutrients through the follicular epithelium and forming vitellogenic follicles (Figure 4b) that became chorionated follicles when the chorion was produced (Figure 4c). Chorionated follicles could already be seen in three-day-old females and a mean of 5.6 were found in four-day-old females. These eggs were fully formed and ready to be fertilized and deposited in the plant tissues. These results indicate that ovary maturation and the sexual acceptance of males for mating must be closely related in M. caliginosus. Lygus hesperus females also become sexually receptive when their first egg matures (Strong et al., 1970). Males. The seminal vesicle was a bright yellow spot that could be seen with naked eye at the end of the abdomen of non dissected individuals. It contained the sperm and was accompanied by four white accessory glands (Figure 5). Although many males exhibited mobile sperm inside their seminal vesicle soon after the last nymphal molt, their four accessory glands were not completely filled. However, three days later, more than 90% of males had mobile sperm and most of them had their accessory glands completely filled (Figure 5). These results indicate that the development of accessory glands in males may be related to their mating success. Development of accessory glands in Lygus hesperus males has been related with their sexual aggressiveness (Strong et al., 1970). sv ag Figure 5. Seminal vesicle (sv) and four accessory glands (ag) of M. caliginosus males. In conclusion, preliminary data on the reproductive characteristics of this predatory bug have been determined in this study. The success of inoculative releases in greenhouses could be implemented by taking into account the bug s mating behaviour, the development of its sexual organs according to female s age and the relationship between both characters. The release of young females imply a high probability that they will be unmated and the difficulty 233
246 of finding a male to mate in the greenhouse will probably determine that few progeny will be produced. Acknowledgements We would like to acknowledge Miguel Angel Mejías and Carles Solá for their dedication and patience in performing some of the laboratory experiments for this study. This work was funded by the Spanish Research Agency INIA (project RTA03-062) and the Spanish Ministry of Education and Science (project AGL ). References Ma, W.K. & Ramaswamy, S.B. 1987: Histological changes during ovarian maturation in the tarnished plant bug, Lygus lineolaris (Palisot de Beauvois)(Hemiptera: Miridae). Int. J. Insect Morphol. & Embryol. 16(5/6): Strong, F.E., Sheldahl J.A., Hughes, P.R. & Hussein E.M.K. 1970: Reproductive biology of Lygus hesperus Knight. Hilgardia 40(4): VanLaerhoven, S.L., Gillespie D.R. & Roitberg B.D. 2003: Diel activity pattern and predation rate of the generalist predator Dicyphus hesperus. Entomol. exp. appl. 107: Wheeler, A.G.Jr. 2001: Biology of the Plant Bugs (Hemiptera: Miridae): Pests, Predators, Opportunists. Cornell University Press, Ithaca, NY. 507 pp. Wightman, J.A. 1973: Ovariole microstructure and vitellogenesis in Lygocoris pabulinus (L.) and other mirids (Hemiptera: Miridae). J. Ent. (A) 48,
247 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Rearing predatory bugs with the brine shrimp Artemia sp. as alternative prey food Jordi Riudavets, Judit Arnó, Cristina Castañé Plant Protection Department, IRTA. Centre de Cabrils, Ctra. Cabrils s/n, Cabrils (Barcelona), Spain. Abstract: We compared the number of individuals of the mirid Macrolophus caliginous and the anthocorid Orius majusculus mass-reared in the laboratory, using dry cysts of Artemia sp. or frozen Ephestia kuehniella eggs as food sources. Numbers of M. caliginosus individuals produced using the two diets were similar. However, the survival of O. majusculus adults and the number of O. majusculus produced were greater when fed with E. kuehniella eggs than with Artemia sp. cysts. The alternative food prey, Artemia sp., seems to offer a feasible and economical alternative to the mass rearing of M. caliginosus but not to that of O. majusculus. Key words: insect rearing, predators, alternative diets Introduction Several predatory bug species are commercially mass reared for use in horticultural crops. Among these species, Macrolophus caliginosus Wagner (Heteroptera: Miridae) and Orius spp. (Heteroptera: Anthocoridae) are supplied in large numbers by several European biofactories. The mirid M. caliginosus is traditionally reared on tobacco plants with Ephestia kuehniella (Lepidoptera: Pyralidae) eggs as prey. Orius spp. is reared using green bean pods as an oviposition substrate and E. kuehniella eggs as prey food (Castañe & Zapata, 2005; van Lenteren & Tommasini, 2003). The price of the E. kuehniella eggs used in the mass rearing of these predators is an important component of their high production cost. Moreover, the need to store E. kuehniella eggs at sub-zero temperatures in order to conserve them carries with it the risk of changes in their nutritional quality, especially if the cold chain is frequently broken. Alternative diets for the mass rearing of generalist predators have been tested in the laboratory with partial success (Castañé & Zapata, 2005; Coudron et al., 2002; Grenier et al., 1989). The brine shrimp Artemia sp. (Branquiopoda: Artemiidae) is used as prey in the larviculture of fish for aquaculture. Its cysts are comparatively cheaper and easier to store than frozen E. kuehniella eggs. Artemia sp. cysts were recently tested as an alternative food for M. caliginosus (Castañé et al., 2006) and Orius laevigatus (Fieber) (Arijs & De Clercq, 2001). Although, under laboratory experiments, Artemia sp. cysts proved excellent alternative prey for M. caliginosus, the two hydrated cysts and decapsulated and rehydrated cysts of Artemia sp. prolonged nymphal development time, reduced adult weight and size, and reduced the fecundity and longevity of O. laevigatus females in relation with predators fed E. kuehniella eggs. The objective of our work was to compare the number of individuals produced in laboratory mass rearing of the mirid M. caliginous and the anthocorid O. majusculus (Reuter) using dry Artemia sp. cysts or frozen E. kuehniella eggs as food sources. 235
248 Material and methods Insect colonies and experimental conditions Stock colonies were raised and experiments carried out in a climatic chamber at a temperature of 25±2ºC with a photoperiod of 16:8 L:D. Insects were obtained from various locations in the area around Barcelona and reared for several generations with E. kuehniella eggs as prey food. During the experiments, we used dry cysts of the brine shrimp Artemia sp. from the Great Salt Lake (Utah, USA) as a comparative food source. Comparison of rearing diets for Macrolophus caliginosus Seventy-five adults were isolated for one week in transparent plastic boxes (37x44x53 cm) with 2 tobacco plants, each of which had 4 or 5 leaves, and with E. kuehniella eggs or dry cysts of Artemia sp. The adults were then removed and the plants were kept in the same climatic chamber for 5 weeks. Twice a week, a small spoonful of eggs or cysts (0.2 g) was distributed amongst the leaves to provide prey food. The number of newly emerged adults and nymphs and the number of dead individuals were recorded at the end of the rearing period. There were 3 replications per prey food source and each experiment was replicated twice. Comparison of rearing diets for Orius majusculus Twenty-five adults were placed in two litres glass jars containing a green bean pod as an oviposition substrate, moistened cotton wool as a water source, paper tissue to increase the surface area and prevent cannibalism, and E. kuehniella eggs or dry Artemia sp. cysts as prey. Bean pods were replaced three times a week for fifteen days and pods containing eggs were isolated in 0.5 l ventilated glass cages. Water and 0.2 g of eggs or cysts were supplied to the neonate nymphs three times a week. The number of newly emerged adults and nymphs and the number of dead individuals were recorded for each cage. Four replications per prey source and 6 bean pods with eggs were isolated per replica. t tests (SAS Institute, 2000) were performed on the total number of individuals produced to test for significant differences between prey sources (α=0.05). Results and discussion Macrolophus caliginosus Approximately 90% of the adults placed in the plastic boxes and fed with the tested diets were still alive after being isolated for one week. As seen in Figure 1, total numbers of M. caliginosus individuals produced in this set-up with dry Artemia sp. cysts and E. kuehniella eggs were not significantly different (first experiment, t=2.22; df=4; P=0.09; second experiment, t=0.07; df=4; P=0.95). The number of dead individuals was very low (<35) but similar for the two diets. Our results agreed with those obtained by Castañe et al. (2006) in laboratory trials using small plastic cages. They succeeded in rearing more than nine successive generations of M. caliginosus on cysts of two different strains of Artemia sp. with Sedum rubrotictum R. T. Clausen (Crassulaceae) leaf cuttings as the plant source. On the other hand, we observed a high level of variability in the number of descendents of M. caliginosus obtained in our experiment, as we had also previously observed in laboratory mass rearing of this predator. Using the same set up used in the present experiment for more than four successive years, a mean production of 381 adults per box was obtained with a standard deviation of (n=234 generations) (unpublished results). Among possible causes for this variability, the quality and freshness of prey appeared to be an important factor. Other than the different qualities of E. kuehniella eggs supplied by the various producers, eggs quality may also be easily altered by the broke of the cold chain in the transportation from the producer to the end user. In a previous experiment we tested 236
249 commercial E. kuehniella eggs of the same batch, feeding bugs with eggs thawed at different periods of time: every day, every two days, every three days or every seven days. We observed that there were no differences in the number of progeny obtained, but again, there was a high of variability among replications (unpublished results). Tibia length was the only parameter studied that was significantly greater for individuals that were fed more frequently First experiment N.S. Second experiment N.S. No. individuals E. kuehniella Artemia sp. E. kuehniella Artemia sp. dead alive Figure 1. Total number of M. caliginosus adults and nymphs (dead and alive) produced on tobacco plants (± SE) with E. kuehniella eggs or dry Artemia sp. cysts as prey. Orius majusculus Survival of the parental generation of O. majusculus was significantly higher when fed with E. kuehniella eggs than with Artemia sp cysts (first experiment, t=6.23; df=6; P<0.001; second experiment, t=7.84; df=4; P<0.001). (Figure 2). In both experiments, adult survival was less than 30% when the alternative diet was supplied. There were no significant differences in the progeny produced (evaluated as O. majusculus adults) when dry Artemia sp. cysts and E. kuehniella eggs were used as prey, as observed in the first experiments (t=0.94; df=6; P=0.38)(Figure 3). However, in the second experiment, the number of individuals produced with E. kuenhiella was significantly higher than the number of individuals produced with Artemia sp cysts (t= 11.29; df=6; P<0.001). Again, as with M. caliginosus, the results obtained were highly variable. As seen in Figure 4, the cumulative number of individuals produced in each trial was different, but the production pattern was similar throughout the two weeks period. Arijs and De Clercq (2001) and De Clercq et al. (2005) previously tested Artemia sp. cysts as prey for O. laevigatus, another anthocorid bug species. Although they found that Artemia sp. cysts were adequate for the development and reproduction of the bug s first generation, after three successive generations they noted longer nymphal development time, lower adult weight, smaller size and reduced female fecundity and longevity with respect to predators fed on E. kuehniella eggs. 237
250 25 20 First experiment * Second experiment * No. individuals E. kuehniella Artemia sp. E. kuehniella Artemia sp. Figure 2. Number of O. majusculus adults alive (± SE), corresponding to the parental generation, after two weeks in glass jars with a green bean pod, moistened cotton wool and E. kuehniella eggs or dry Artemia sp. cysts as prey First experiment N.S. Second experiment * No. individuals E. kuehniella Artemia sp. E. kuehniella Artemia sp. Figure 3. Comparison of O. majusculus progeny produced (total number of adults) in glass jars with a green bean pod, moistened cotton wool and E. kuehniella eggs or dry Artemia sp. cysts as prey 238
251 No. individuals E. kuehniella 1 E. kuehniella 2 Artemia sp 1 Artemia sp 2 Figure 4. Accumulated number of O. majusculus produced in the two experiments (1 and 2) in glass jars with a green bean pod, moistened cotton wool and E. kuehniella eggs or dry Artemia sp. cysts as prey. In conclusion, Artemia sp. cysts seemed to offer a feasible and economical alternative prey to mass rearing M. caliginosus. However, Artemia sp. cysts could not be used in the mass rearing of O. majusculus. Their combined use with other food supplements such as flower pollen should be investigated in future research. Acknowledgements We would like to thank Pilar Hernández and Victor Muñoz for their help in conducting the lab experiments. This work was supported by a grant from the Spanish Research Agency INIA (project RTA03-062). References Arijs Y. & De Clercq P. 2001: Rearing Orius laevigatus on cysts of the brine shrimp Artemia franciscana. Biological Control 21: Castañé C. & Zapata R. 2005: Rearing the predatory bug Macrolophus caliginosus on a meatbased diet. Biological Control 34: Castañé C., Quero R. & Riudavets J. 2006: The brine shrimp Artemia sp. as alternative prey for rearing the predatory bug Macrolophus caliginosus. Biological Control (submitted). Coudron T.A., Wittmayer J. & Kim Y. 2002: Life history and cost analysis for continuous rearing of Podisus maculiventris (Say)(Heteroptera: Pentatomidae) on a zoophytophagous artificial diet. Journal of Economic Entomology 95(6): De Clercq P., Arijs Y., Van Meir T., Van Stappen G., Sorgeloos P., Dewettinck K., Rey M., Grenier S. & Febvay G. 2005: Nutritional value of brine shrimp cysts as a factitious food for Orius laevigatus (Heteroptera: Anthocoridae). Biocontrol Sci. Technol. 15(5):
252 Grenier S., Guillaud J., Delobel B. & Bonnot G. 1989: Nutrition et élevage du predateur polyphage Macrolophus caliginosus (Heteroptera: Miridae) sur millieux artificiels. Entomophaga 34(1): SAS Institute. 2000: PROC user s manual, version 8.02 ed. SAS Institute, Cary, NC. van Lenteren J.C. & Tommassini, M.G. 2003: Mass production, storage, shipment and release of natural enemies. Pp In: Quality control and production of biological control agents, Theory and testing procedures. JC van Lenteren. CAB International. UK. 240
253 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Investigation on the efficiency of a predator: a strategy beyond satiation Dionyssios Perdikis 1, Panagiotis Labropoulos 2, Dionyssia Maselou 2, Argyro Fantinou 2 1 Laboratory of Agricultural Zoology and Entomology; 2 Laboratory of Ecology and Environmental Sciences, Agricultural University of Athens, Iera Odos 75, , Athens, Greece Abstract: The predation efficiency of the polyphagous predator Macrolophus pygmaeus (Hemiptera: Miridae) was investigated using the aphid Myzus persicae (Homoptera: Aphididae) as prey. One 5 th instar nymph of the predator that had been caged on an eggplant without prey for 24h prior to the experiments was released in a plastic Petri dish of 9cm in diameter. In each dish an eggplant leaf was placed on which 2, 4, 8, 12, 16, 20 or 24 nymphs of only one of the 4 instars of the aphid were introduced. The experiments were conducted at 20, 25 and 30 C, under a L16:D8 photoperiod and 65±5% R.H. In each case 10 replicates (predators) were performed. The sucked (consumed) aphids were measured 24h after the introduction of the predator in the dish. It was observed that apart from the consumed aphids, killed but unconsumed aphids were also present in the dish. The number of these killed aphids was highest at 20 C and decreased with temperature increase whereas were much more frequently recorded in the 4 th and the 3 rd than in the younger instars. Most interestingly, these aphids were recorded mainly in the higher densities of prey used. Therefore, it seems that apart from killing aphids for direct consumtion this predator causes aphid mortality by killing them without sucking them. This behavior increases the rate of pest killed with considerable implications in the effectiveness of this predator in biological control. Key words: biological control, killing rate, Macrolophus pygmaeus, Myzus persicae Introduction Field studies revealed that the predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae) was the main natural enemy of aphids on several vegetable crops (Lykouressis et al ). This predator is polyphagous feeding on aphids, whiteflies and mites (Perdikis & Lykouressis 2000, Hommes & Horst 2002). All nymphal stages and adults of this predator, and in particular 5 th stage nymphs and females, have been found to consume relatively high numbers of Myzus persicae (Sulzer) (Homoptera: Aphididae) nymphs on eggplant and pepper (Perdikis et al. 1999). The aim of this study was to investigate the predation rates of the predator M. pygmaeus at different temperatures when different densities of all instars of its prey, the aphid M. persicae, were available. Materials and methods M. pygmaeus were reared on potted eggplants (cv. Bonica) infested with M. persicae in wood framed cages, and were kept in a controlled temperature glasshouse under natural lighting. Experiments were conducted in plastic Petri dishes (9 cm diameter, 1.5 cm height) with a round hole (3 cm diameter) in their top, covered with fine muslin to reduce the high humidity inside the dish. A leaf of eggplant was placed with the abaxial surface up on a layer of moistened cotton wool and with water resting on the bottom of each dish. On each leaf 4, 8, 12, 16, 20 or 24 aphids for each aphid instar were placed gently and left undisturbed for 1h to 241
254 settle. Then, one fifth instar nymph of the predator was introduced in the dish. Each fifth instar nymph of M. pygmaeus had been developed from first or second instar nymphs which had been transferred from the rearing wood framed cages to potted eggplants infested with an excess of M. persicae. Each fifth instar nymph was caged on an eggplant at the above conditions and deprived of prey for 24h prior to its use in the experiments. The sucked (eaten) aphids by the predator in each dish were recorded 24h after the introduction of the predator in the dish. The number of killed but not sucked aphids was also recorded in each treatment. For each aphid density, 10 replicates (predators) were used. The experiments were carried out at 20, 25 and 30 C, under a L16:D8 photoperiod and 65±5% R.H. Consumption rates were analysed using ANOVA. Results and discussion The number of killed but not sucked aphids was found to increase with temperature, aphid instar and aphid density (F 2,648 =7.62, P<0.001; F 3,648 =35.97, P<0.001; F 5,648 =7.85, P<0.001, respectively). The number of killed aphids was expressed as a percentage of the total number of dead (sucked plus killed) aphids in each replicate. This percentage was found to be highest in the fourth instars at 20 C in the density of 8 aphids averaging 29%. Among temperatures it was highest at 20 C averaging 0.07% and among instars it was highest in the fourth reaching to 0.09%. Among instars and densities this percentage was highest in the fourth instars at the highest density reaching to 0.12%. The results revealed that apart from the prey consumed by M. pygmaeus, this predator could further reduce the population of its prey by killing without eating it. Such a feeding behaviour has been also recorded for the predator Aphidoletes aphidimyza Rondani (Diptera: Cecidomyiidae) which does not consume a percentage of the prey it kills and this percentage increased with the size and the number of the available prey (Uygun 1971). In the case of M. pygmaeus the percentage of killed but not sucked prey was highest at higher prey densities and therefore, when prey was abundant the predatory capacity of M. pygmaeus can be enlarged contributing therefore to a more effective exploitation and reduction of prey numbers. References Hommes, M. & Horst, S.T. 2002: Development and life span of Macrolophus pygmaeus Rambur at different temperatures and influence of host plants and prey. IOBC/WPRS Bull. 25(1): Lykouressis, D.P., Perdikis D.Ch. & Chalkia, Ch.A : The effects of natural enemies on aphid populations on processing tomato. Entomol. Hellenica 13: Perdikis, D.Ch. & Lykouressis, D.P. 2000: Effects of various items, host plant and temperature on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Control 17: Perdikis, D.Ch., Lykouressis, D.P. and Economou, L.P. 1999: The influence of temperature, photoperiod and plant type on the predation rate of Macrolophus pygmaeus on Myzus persicae. BioControl 44: Uygun, N. 1971: Der Einfluss der Nahrungsmenge auf Fruchtbarkeit und Lebensdauer von Aphidoletes aphidimyza (Rond.) (Diptera: Itonididae). Z. ange. Entomol. 69:
255 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Zoophytophagy of the plantbug Nesidiocoris tenuis in tomato crops in southeast Spain. Juan Antonio Sanchez, Modesto del Pino-Pérez, Mª del Mar Davó, José Isidro Martinez- Cascales, Alfredo Lacasa Department of Biotechnology and Crop Protection, Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (IMIDA), c/ Mayor, s/n, La Alberca, Murcia, Spain. juana.sanchez23@carm.es Abstract: Nesidiocoris tenuis (Heteroptera: Miridae) is one of the most abundant zoophytophagous plantbugs in tomato crops in southeast Spain. It preys on whiteflies and other small pests such as spider mites, leafminers and early instars of Lepidoptera. It also feeds on plants producing necrotic rings in stems and leaves, flower abortion and fruit blemishing. However, the intensity of the damage seems to vary from one geographical region to another. The aim of this assay was to determine the role of N. tenuis as a whitefly predator and to quantify damage in tomato crops in southeast Spain. Assays were carried out in four mesh-houses from July to October. Two of the greenhouses had screened vents while in the other two, the vents were unscreened. Trialeurodes vaporariorum was much more abundant than Bemisia tabaci. There were no differences in whitefly and N. tenuis population dynamics between greenhouses with screened and unscreened vents. Damage to flowers and stems followed N. tenuis population dynamics. N. tenuis caused the abortion of around 50% of the flowers during the four weeks following the population outbreak. We advise caution when using this plantbug as a biological control agent until management strategies have been better defined. Key words: Nesidiocoris tenuis, Miridae, plantbugs, whitefly, Bemisia tabaci, Trialeurodes vaporariorum, tomato, damage, flower abortion. Introduction Nesidiocoris tenuis (Reuter) (Heteroptera: Miridae) is the one of the most abundant dicyphines in tomato crops in southeast Spain. This zoophytophagous insect is commonly found in tomato crops along the southern Mediterranean coast when the use of pesticides is reduced (Sanchez et al., 2003a, 2003b). It is a very effective whitefly predator able to reduce the pest population in a short time. Calvo and Urbaneja (2003) considered N. tenuis a key predator for the control of Bemisia tabaci Gennadius and Trialeurodes vaporariorum (Westwood) in southern Spain and the Canary Islands. It also preys on other tomato pests such as spider mites, leafminers and early instars of Lepidoptera (Torreno 1994; Urbaneja et al., 2005). Augmentative release and conservation of N. tenuis have been encouraged for pest control in tomato crops in the Mediterranean area and the Canary Islands (Calvo & Urbaneja, 2003; Calvo & Urbaneja, 2004). N. tenuis also feeds on plants, producing necrotic rings in stems and leaves, flower abortion and fruit blemishing. However, the intensity of the damage varies from one geographical region to another; for instance, whilst in the Canary Islands N. tenuis damage has not been reported and the bug is a welcome pest control agent (Carnero et al., 2000), in southeast Spain it causes damage to tomato plants and its role is controversial. Calvo and Urbaneja (2004) suggested an approximate intervention threshold for tomato crops, but neither the factors involved nor the quantity of damage produced have ever been determined. 243
256 The aim of this assay was to determine the role of N. tenuis as a whitefly predator and to quantify damage in tomato crops in southeast Spain. In this area, meshes have been widely used to cover vents and prevent B. tabaci and T. vaporariorum immigration because of the high incidence of virus (e.g. Tomato yellow leaf curl TYLCV- and Tomato chlorosis virus ToCV-) in the last few years. Insect-excluding screens are also considered to negatively influence crop colonization by natural enemies. In this work we also compared whitefly and N. tenuis population dynamics in greenhouses both with and without screened vents. Material and methods Crop history Assays were carried out in four mesh-greenhouses (10 x 20 threads/cm 2 ) located in Mazarrón (Murcia, Spain), from the beginning of July until the end of October of In two of the greenhouses, 480 plants were arranged in eight double rows of 30 plants. In the other two, 360 plants were arranged in six double rows of 30 plants: in these last two greenhouses, the lateral vents had no screens. Tomato seedlings were transplanted on July 10 th, Distance between plants was 0.5 m, and 1 m both within lines and between double rows. Tomato yellow leaf curl Sardinian virus (TYLCSV) tolerant varieties Boludo (Petoseed) and Tyrade (S&G) were used. No chemicals were applied to control pests or diseases. Insect and plant sampling N. tenuis and whitefly population dynamics were monitored by weekly sampling 30 randomly selected plants in each greenhouse. N. tenuis, adults and nymphs, were counted in situ on one leaf from the apex, one from the middle, and one from the lower part of the plant. Middle leaves were bagged, brought to the laboratory and observed under a stereo microscope for scale counting. Whitefly immatures were only counted on alternate leaflets. B. tabaci and T. vaporariorum pupae were recorded separately, while nymphs were all counted together and not differentiated. Damage produced by N. tenuis in shoots and flowers was quantified for each of the 30 randomly selected plants. The number of necrotic rings was counted in the upper 20 cm of each plant. Both the total and the number of aborted flowers were also counted in one bunch of flowers from each plant. Data analysis Abundance of N. tenuis was expressed as the average number of individuals per leaf (the average of the three sampled leaves per plant). Whitefly abundance was expressed as the average number of immatures per leaf (nymphs + pupae), without differentiating between B. tabaci and T. vaporariorum. Damage was expressed as the average number of necrotic rings and the proportion of aborted flowers. Results and discussion Insect population dynamics The whitefly population reached a very high density soon after the transplant, due to the high rate of whitefly immigration. In greenhouses with unscreened lateral vents, a maximum of 194 scales per leaf was reached at the end of July (Figure 1a). In the following six weeks the whitefly population progressively decreased to 3.3 scales per leaf (Figure 1a). The most abundant whitefly species was T. vaporariorum, while only 5% of all pupae were B. tabaci. In greenhouses with screened vents whitefly reached a maximum of 292 scales per leaf in 244
257 about the second week of August (Figure 2a). In the following five weeks, the number of scales descended to 2.8 per leaf. T. vaporariorum was the most abundant whitefly species during the sampling period, representing 90% of all pupae. N. tenuis spontaneously colonized the crops about three weeks after the transplant. In houses with unscreened vents, the N. tenuis population started to increase and reached a peak of 5.4 individuals per leaf in about the fourth week of August (Figure 1a). Thereafter, the number of N. tenuis decreased following the same trend as for whitefly. In greenhouses with screened vents N. tenuis population dynamic was very similar to that of unscreened greenhouses; N. tenuis reached a maximum of 4.4 individuals per leaf at the beginning of September and numbers decreased thereafter. In all greenhouses, N. tenuis density stabilized at around 1 individual per leaf after the fall in population (Figure 1a and 2a). There were almost no differences between greenhouses with respect to the establishment and population dynamics of N. tenuis either with screened or unscreened vents. This was also the case for whitefly, whose initial population and evolution were very similar for both types of greenhouses. Contrary to what we had expected, screens did not represent an obstacle to the immigration of insects to crops. However, the absence of significant differences may have been due to poor mesh setting at junctions, leaving holes that facilitated the entry of insects. Crop damage The number of necrotic rings in the apex of tomato plants and the number of aborted flowers evolved parallel to N. tenuis population dynamics (Figure 1 and 2). In both types of greenhouse, the greatest damage was observed at N. tenuis population peaks. The highest numbers of necrotic rings were around 6 and 4 for greenhouses with unscreened and screened vents, respectively (Figure 1b and 2b). In greenhouses with unscreened laterals, the percentage of aborted flowers remained around 50% during the four weeks following N. tenuis outbreaks (Figure 1b). In screened vent greenhouses, the percentage of aborted flowers ranged between 43 and 52% in the weeks following N. tenuis outbreaks. The damage inflicted by N. tenuis is of great concern when using this zoophytophagous plantbug for pest control in tomato crops in southeast Spain. The number of necrotic rings produced by N. tenuis, even at high population densities, did not seem to limit plant growth and loss from feeding on stems and leaves is expected to be of very little economic significance. However, necrotic rings sometimes weaken the apex, leading to the decapitation of the plant. The biggest problem with N. tenuis was that it caused the abortion of a high number of flowers. Although yield losses were not been quantified in this work, the high rate of flower abortion produced by N. tenuis was quite likely to have a significant economic impact on the produce. The abortion of flowers is especially troublesome in cluster tomato varieties. Damage to fruits has also to be quantified because it may downgrade yield. On the basis of these results we advise growers to be very cautious in the use of N. tenuis in tomato crops in southern Spain. The phytophagous behaviour of N. tenuis has to be characterized and intervention thresholds established before we can be confident about the use of this plantbug as a pest control agent in tomato crops. Acknowledgements We would like to thank Celestino Méndez from the COARA growers association and also José Francisco Pastor and Fernando Cánovas, who helped with sampling. This work was funded by research project AGL C J.I. Martinez-Cascales had a pre-doctoral fellowship granted by the Fundación Séneca (Agencia Regional de Ciencia y Tecnología. Región de Murcia, Ref /EE2/05). 245
258 References Calvo, J & Urbaneja, A. 2003: Nesidiocoris tenuis Reu. (Het.: Miridae) en tomate: Amigo o Enemigo? AlmeríaVerde 4: Calvo, J & Urbaneja, A Nesidiocoris tenuis un aliado para el control biológico de mosca blanca. Horticultura Internacional 44: Carnero, A., Díaz, S., Amador, S., Hernández, M. & Hernández E. 2000: Impact of Nesidiocoris tenuis (Heteroptera, Miridae) on whitefly populations in protected tomato crops. IOBC/WPRS Bull. 23: 259. Sanchez, J.A., Martinez-Cascales, J.I. & Lacasa, A. 2003a: Abundance and wild host plants of predator mirids (Heteroptera: Miridae) in horticultural crops in the Southeast of Spain. IOBC/WPRS Bull. 26(10): Sanchez, J.A., Martinez-Cascales, J.I. & Lacasa, A. 2003b: Distribución geográfica, abundancia y plantas hospedantes de míridos depredadores (Heteroptera: Miridae) de interés para el control biológico de plagas en cultivos hortícolas de la región de Murcia. Phytoma España 154: 55. Torreno H. 1994: Predation behavior and efficiency of the bug Cyrtopeltis tenuis (Hemiptera: Miridae), against the cutworm, Spodoptera litura (F.). Philippine Entomol. 9: Urbaneja, A., Tapia, G. & Stansley, P. 2005: Influence of host plant and prey availability on developmental time and survivorship of Nesidiocoris tenuis (Het.: Miridae). Biocontrol Sci. Technol. 15:
259 a 6 5 Scales per leaf Nesidiocoris per leaf Jul 28-Jul 4-Aug 11-Aug 18-Aug 25-Aug 1-Sep Whitefly 8-Sep 15-Sep 22-Sep Nesidiocoris 29-Sep 6-Oct 13-Oct 20-Oct 0 Necrotic rings 4,5 4 3,5 3 2,5 2 1,5 1 0, Jul 28-Jul 4-Aug 11-Aug 18-Aug Feeding damage 25-Aug 1-Sep 8-Sep 15-Sep 22-Sep 29-Sep 6-Oct 13-Oct 20-Oct Aborted flowers Figure 1. Mesh greenhouse with unscreened lateral vents. (a) Population dynamics of Nesidiocoris tenuis and whiteflies (T. vaporariorum and B. tabaci). (b) Damage to tomato plants. b 0,6 0,5 0,4 0,3 0,2 0,1 0 Proportion of aborted flowers 247
260 a 6 5 Scales per leaf Nesidiocoris per leaf Jul 28-Jul 4-Aug 11-Aug 18-Aug 25-Aug 1-Sep Whitefly 8-Sep 15-Sep 22-Sep Nesidiocoris 29-Sep 6-Oct 13-Oct 20-Oct ,6 Necrotic rings Jul 28-Jul 4-Aug 11-Aug 18-Aug Feeding damage 25-Aug 1-Sep 8-Sep 15-Sep 22-Sep 29-Sep 6-Oct 13-Oct 20-Oct Aborted flowers Figure 2. Mesh greenhouse with screened lateral vents. (a) Population dynamics of Nesidiocoris tenuis and whiteflies (T. vaporariorum and B. tabaci). (b) Damage to tomato plants. b 0,5 0,4 0,3 0,2 0,1 0 Proportion of aborted flowers 248
261 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Characterization of damage to tomato plants produced by the zoophytophagous predator Nesidiocoris tenuis Judit Arnó, Cristina Castañé, Jordi Riudavets, Job Roig, Rosa Gabarra Departament de Protecció Vegetal, IRTA-Centre de Cabrils, E Cabrils (Barcelona), Spain. Abstract: Nesidicoris tenuis is a zoophytophagous predator that is native to the Mediterranean. It is known to prey on different pests of vegetables but also to damage plants. As a consequence, whereas some authors regard it as an interesting biological control agent, for others it is a pest. To asses the importance of damage caused by this mirid bug, we surveyed commercial crops and conducted greenhouse and laboratory experiments to characterize the kind of damage caused in both the presence and absence of prey. Although, necrotic rings in the apex, leaf and leaflet petioles have been observed in crops with established N. tenuis populations, the densities reported for commercial fields in our region do not seem to cause severe damage to tomato plants. Greenhouse experiments showed that nymphs potentially caused more damage than adults. Even so, the damage caused by N. tenuis feeding tended to disappear or attenuate with time and did not affect plant growth. In laboratory experiments, the presence of prey significantly reduced the number of necrotic rings caused. In this preliminary study, no flower abortion was observed but more specific experiments need to be done to address this question of capital importance. Key words: Nesidiocoris tenuis, zoophytofagy, tomato Introduction Nesidicoris tenuis is a zoophytophagous predator that is native to the Mediterranean region and present in many areas were tomato and other vegetables are grown. It is very abundant in the Southeast of Spain and the Canary Islands (Carnero et al. 2000, Sánchez et al. 2003). In the Northeast of Spain it is only abundant in late summer-autumn crops. This mirid bug spontaneously colonizes tomato crops, both in greenhouses and open fields, especially when IPM programs are used. The role of N. tenuis in tomato crops is not clear. While for some authors this mirid is an interesting biological control agent for whitefly (Carnero et al. 2000, Vacante & Tropea Garzia 1994), for others it is a pest (Raman et al. 1984, Trottin-Caudal & Millot 1997). The damage mentioned in the literature includes cell necroses produced by stylet insertion in plant tissues and loss of sap causing the appearance of brown rings around stems and leaf petioles. These rings can lead to a withering of leaflets or even of the apex. Flower abortion, fruit blemish and discolouration punctures in leaflets have also been described (Raman et al. 1984, Vacante & Tropea-Garzia 1994). The aim of this study was to obtain preliminary data on the importance of damage inflicted on tomato crops by N. tenuis under the conditions typically found along the northeast coast of Spain. We therefore carried out the field surveys and greenhouse and laboratory experiments reported in this paper. 249
262 Material and methods Field surveys: evaluation of damage incidence To assess the importance of the damage caused by N. tenuis, eight commercial (including greenhouse and open field) tomato crops, in which IPM advisors had detected its presence were sampled from the end of August until mid-november. The number of visits to each crop depended on its phenological stage and varied from 1 to 5. On each sampling date, 30 plants were selected at random. One terminal shoot and ten leaflets were examined for each plant. The number of Trialeurodes vaporariorum and Bemisia tabaci adults and the number of N. tenuis adults and nymphs were separately recorded. The number of necrotic rings around the shoot apex and around the leaf and leaflet petioles was also registered. The mean number of N. tenuis adults or nymphs and the mean number of whitefly adults were regressed against the mean number of injuries per plant. Greenhouse experiments: damage characterization. The experiment was conducted in an experimental greenhouse at IRTA-Cabrils. The N. tenuis used in the experiment came from laboratory colonies originally collected from vegetable crops in the province of Barcelona. Tomato plants (cv. Bodar) were transplanted into pots and grown until they had two flower clusters. Side shoots were individually caged in muslin bags with N. tenuis adults and nymphs. No prey was present on the shoot in order to force predators to feed on the plant. Three treatments were set up according to the number and stage of mirid bugs released in each bag: 10 or 20 adults and 20 nymphs. There were 23 or 24 replicas for each treatment. After 48 h, the muslin bags were opened and the bugs were removed. Between 0 and 24 h after removal of N. tenuis, the number of necrotic rings on the stems and on the leaf, leaflets and flower petioles of each shoot were counted and data were pooled. A second observation was carried out between 3 and 5 days after removal, and for some of the 10 adults and 20 nymphs treatment repetitions, a third observation was made 7 days after mirid removal (n=12 and 20, respectively). The effect of mirid treatment on the mean number of injuries per shoot was analyzed with an ANOVA of the transformed data ( X ). The Tukey test (P<0.05) was used to determine differences between means. Laboratory experiments: damage in relation to prey density The experiment was conducted in a climatic chamber at 25ºC and 16:8 (L.D) h. A tomato shoot was inserted into a hole made in the bottom of a transparent plastic cup (300 cc) which was covered with a piece of muslin cloth to allow ventilation. This cup was then placed inside a second plastic cup containing water to keep the tomato shoot turgid. Five adults or five nymphs of N. tenuis laboratory from the IRTA laboratory colony were then introduced into the cage together with B. tabaci adults. Two different treatments were set on the basis of the whitefly density: with no prey and with 50 B. tabaci adults. Insects were kept in the cage for 3 days. They were subsequently removed and the number of injuries (necrotic rings) was immediately recorded. Between 9 and 14 replicas were made. Data on the number of injuries per shoot was analyzed with a two-way ANOVA to evaluate the effect of mirid stage and the presence of prey. When significant differences were found, the Tukey test (P<0.05) was used to determine differences between means. Results and discussion Field surveys: evaluation of damage incidence In our surveys, N. tenuis populations fluctuated from 0.13 to 5.23 individuals per plant. On average, nymphs accounted for 74% of the mirid population. In the surveyed crops, 94% of the observed necrotic rings were found around the leaf and leaflets petioles and just 6% were 250
263 around the stem. The mean number of N. tenuis nymphs (independent variable) was consistently related to the mean number of necrotic rings around the petioles (dependent variable) (Y= 0.84 X -0.09; F=84.65; df=1, 18; P<0.001; R 2 =0.83) (Figure 1). A consistent relationship was also found between the mean number of mirid bug adults and the mean number of injuries in the leaf and leaflet petioles, but in this case, a low proportion of variation in the amount of necrotic rings is explained by variation in the number of adults (Y= 1.22 X ; F=6.84; df=1, 18; P=0.02; R 2 =0.29). In different samplings on commercial crops, whitefly populations ranged from 0.1 to 22.3 adults per plant. T. vaporariorum was the predominant whitefly species and the mean proportion of B. tabaci was 4% of the total number of adults registered. No consistent relationship was found between the pooled average density of the two species of whitefly adults and the number of necrotic rings on leaf and leaflet petioles (F=1.45; df=1, 18; P=0.25). No. injuries / plant NYMPHS R 2 = # N. tenuis /plant ADULTS R 2 = # N. tenuis /plant Figure 1: Regression model with the number of N. tenuis as the independent variable and the number of injuries to leaf and leaflet petioles as the dependent variable. No. injuries / plant Greenhouse experiments: damage characterization. In the first observation after mirid removal, N. tenuis nymphs produced significantly a higher number of necrotic rings on tomato shoots than adults (F=39.90; df=2, 60; P<0.001) (Figure 2). No differences in the number of injuries were detected between the treatments with 10 and 20 adults caged on a shoot. However, the number of injuries detected between 0-24 h after mirid removal decreased during the first week. Three to 5 days after mirid removal, 63% and 76% of the necrotic rings had respectively disappeared in the 10 adults and 20 adults treatments, but in the 20 nymphs treatment the reduction was negligible (1.4%). Furthermore, only 1 necrotic ring was observed in the 10 adults treatments, but none in the 20 nymphs treatments, 7d after the mirids were removed. Although flower abortion was not specifically registered, flower clusters developed as normal, fruit set occurred and fruits did not fall. 251
264 No. of injuries / shoot a b b 10 adults Nt 20 adults Nt 20 nymphs Nt Figure 2: Mean (±SE) number of necrotic rings caused by N. tenuis on tomato shoots < 24 h after the mirids were removed. Laboratory experiments: damage in relation to prey density Figure 3 shows the influence of prey density on the number of necrotic rings. Two-way ANOVA show no interaction between the two factors: mirid stage and prey density (F=0.32; df=1, 41; P=0.58). The number of injuries per shoot was not significantly different between nymphs and adults of N. tenuis (F=0.47; df=1, 41; P=0.50), but the presence of B. tabaci adults significantly reduced the number of injuries detected on each shoot (F=11.48; df=1, 41; P=0.002). According to the results presented in this work, nymphs cause more damage to young shoots and to leaf and leaflet petioles than adults. A high proportion of the variation in damage observed in commercial fields could be explained by the abundance of nymphs but not by the number of adults recorded on the plants. This may have been due to the greater number of nymphs recorded in our samplings and was probably related to the sampling procedure itself, since adults are mobile and therefore more difficult to localize on plants. However, greenhouse experiments showed that nymphs potentially caused more damage than adults. Injuries caused by N. tenuis feeding on tomato stems and petioles tended to disappear or attenuate with time and did not affect plant growth. Consequently, the damage observed in commercial fields was probably a response to continuous feeding on the same growth point. Raman & Sanjayan (1984) reported that, under field conditions, 4 to 5 individuals feeding for at least hours produced the withering of stems or petioles. The results of our field surveys showed that, under our conditions, no reduction of plant growth was observed in relation to the density of N. tenuis on these crops (up to 5 individuals on the young apex and ten leaflets per plant). Raman et al. (1984), Vacante & Tropea-Garzia (1994) and Calvo & Urbaneja (2004) reported the appearance of necrotic rings in flowers and fruits that sometimes lead to the abortion of flowers. Our observations show that, at the N. tenuis densities tested and with 48 hours of interaction between the mirids and the plant, the zoophytophagous predator did not cause the abortion of flowers even when immature flower truss were caged. 252
265 No. of injuries / shoot No. of B. tabaci adults offered nymphs adults Figure 3: Mean number of necrotic rings (±SE) caused by adults or nymphs of N. tenuis in the presence and absence of B. tabaci adults. N. tenuis require animal prey to reach adulthood, although they can survive up to the third nymphal instar on a strict tomato plant diet (Urbaneja et al. 2005). According to our laboratory experiments, the absence of prey coincided with an increase in the number of necrotic rings detected on shoots and petioles. These necroses were a consequence of frequent stylet insertion and the loss of plant sap along the stylet track (Raman et al. 1984). Presumably, when no prey is present more frequent stylet insertion in plant tissues occurr, reflecting attempts to obtain more nutrients from the plant. The fact that in our field survey we did not observe any relationship between the number of whitefly adults and number of necrotic rings might have been because whitefly were not the only prey present in the field. This polyphagous predator feeds on whitefly (Vacante & Tropea-Garzia 1994, Carnero et al. 2000), but also on lepidoptera eggs and worms (Torreno 1994) and also on other small insects such as spider mites, aphids and leafminers (Vacante & Benuzzi 2002). Even in cases in which necrotic rings that appeared on crops that had established N. tenuis populations, the densities reported for commercial fields in our region did not seem to cause severe damage to tomato plants. In this preliminary study no cases of flower abortion were observed but more specific experiments need to be done to address this question of capital importance. Acknowledgements We would like to thank Virginie Thiriaud, Clea Henzen, Victor Muñoz and Pilar Hernández for their technical support during the experiment. The study was financially supported by a project of the Ministerio de Educación y Ciencia (AGL ) and was made possible by a collaboration agreement between the IRTA, SELMAR and ADV Baix Maresme. References Calvo, J. & Urbaneja, A. 2004: Nesidiocoris tenuis, un aliado para el control biológico de mosca blanca. Horticultura Internacional 44: Carnero, A., Díaz, S., Amador, S., Hernández, M. & Hernández, E. 2000: Impact of Nesidiocoris tenuis (Heteroptera, Miridae) on whitefly populations in protected tomato crops. IOBC/WPRS Bull. 23 (1):
266 Raman, K. & Sanjayan, K.P. 1984: Histology and histopathology of the feeding lesions by Cyrtopeltis tenuis Reut. (Hemiptera: Miridae) on Lycopersicon esculentum Mill. (Solanaceae). Proc. Indian Acad. Sci. (Anim. Sci.) 93 (6): Raman, K., Sanjayan, K.P. & Suresh, G. 1984: Impact of feeding injury of Cyrtopeltis tenuis Reut. (Hemiptera: Miridae) on some biochemical canges in Lycopersicon esculentum Mill. (Solanaceae). Current Sci. 53 (20): Sánchez, J.A., Martínez, J.I. & Lacasa, A Abundance and wild host plants of predator mirids (Heteroptera:Miridae) in hosrticultural crops in the Southeast of Spain. IOBC/WPRS Bull. 26 (10): Torreno, H.S. 1994: Predation behavior and efficiency of the bug Cyrtopeltis tenuis (Hemiptera:Miridae), against the cutworm, Spodoptera litura (F). Philipp. Ent. 9(4): Trottin-Caudal, Y. & Millot, P. 1997: Etude de deux mirides en culture de tomate: Macrolophus caliginosus Wagner et Nesidiocoris (Cyrtopeltis ) tenuis Reuter. Infos-Ctifl 131: Urbaneja, A., Tapia, G. & Stansly, P. 2005: Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenius (Het.: Miridae). Biocontrol Sci. Techn. 15: Vacante, V. & Benuzzi, M. 2002: Pomodoro, la difesa biologica e integrata. Colture Prottete 8: Vacante, V. & Tropea-Garzia, G. 1994: Nesidiocoris tenuis: antagonista naturale di aleurodidi. Informatore Fitopatologico. 4:
267 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Susceptibility to virus infection of candidate plants used to enhance predatory dicyphine (Heteroptera: Miridae) Ignacio Morales 1, Juan Antonio Sanchez 2, Oscar Alomar 3, Alfredo Lacasa 2, Alberto Fereres 1 1 Instituto de Ciencias Agrarias (CCMA-CSIC). Serrano 115 dpdo, Madrid. 2 Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario, IMIDA La Alberca, Murcia. 3 Departament de Protecció Vegetal, IRTA-Centre de Cabrils, Cabrils, Barcelona, Spain. Abstract: Selecting candidate plants in order to enhance predators for conservation biological control not only has to consider their contribution to enhancement of predators, but also avoid the risk of acting as reservoirs of potential pests or diseases. Mirid bugs are effective biological control agents of tomato pests and the introduction of insectary plants has been shown to be an effective means for the conservation of these important natural enemies. We studied the degree of susceptibility and the role of selected insectary plants as reservoirs of the most important viruses found in tomato crops in Spain. A virus survey conducted in the regions of Murcia and Barcelona, and a series of transmission experiments conducted under controlled conditions indicated that the species Dittrichia viscosa and Marrubium vulgare are susceptible and may play a significant role in Potato virus Y and Cucumber Mosaic Virus epidemics, whereas virus were never detected in Carlina corymbosa and Ononis natrix collected in the field and were immune to the transmission of both viruses by aphids. Our work suggests that there is a potential risk for virus epidemics to occur if M. vulgare and D. viscosa are introduced as insectary plants to attract and enhance biological control by predatory mirids. Further research is needed to confirm the role that M. vulgare and D. viscosa and Verbascum thapsus may play in the epidemics of PVY and CMV in tomato crops. C. corymbosa and O. natrix appear to be the species that show the lowest risk as reservoirs of viruses that commonly infect tomato crops. Keywords: Heteroptera, Miridae, Dyciphinae, Marrubium vulgare, Dittrichia viscosa, Ononis natrix., Carlina corymbosa, Verbascum thapsus, insectary plants, PVY, CMV, TSWV, TYLCV, tomato Introduction The introduction of insectary plants has been shown to be effective for the conservation of natural enemies in the proximity of crops and to facilitate their establishment (Arnó et al., 2000; Alomar et al., 2002; Sanchez et al., 2003a; Pascual-Villalobos et al., 2006). However, the selection of plants also has to avoid or minimize the risk of plants enhancing pests or diseases that are common in the target cropping area (or avoid plants that augment the inoculum pressure). Vegetable production in Spain is threatened by a series of insect pests and insect-transmitted virus diseases that cause significant economic yield losses every year. Tomato crops are particularly susceptible to a wide range of virus diseases. Tomato spotted wilt virus (TSWV) has caused severe epidemics since the introduction of Frankliniella occidentalis (Pergrande) (Thysanoptera: Thripidae) in Spain in the late 80s (Lacasa, 1990). Important viruses are also Potato virus Y (PVY), Cucumber Mosaic Virus (CMV), and Tomato yellow leaf curl virus (TYLCV), this last one is transmitted by the whitefly Bemisia tabaci Gennadius, that causes the most destructive disease of tomato crops throughout the Mediterranean region (Gafni, 2003). 255
268 Several species of predatory mirids contribute to the control of important pests of vegetables in the mediterranean regions (Gabarra, 2002). Macrolophus caliginosus Wagner is an effective predator of Trialeurodes vaporariorum (Westwood) and B. tabaci in several crops, especially in tomato (Castañé et al., 2000; Albajes & Alomar, 1999).) The proximity of refuge plants like Dittrichia viscosa (L.) Greuter, favoured the colonization of tomato crops by M. caliginosus (Alomar et al., 2002). Mirid bugs have been also observed in other plant species like Marrubium vulgare L., Ononis natrix L. and Carlina corymbosa L. (Sanchez et al., 2003b). However, natural vegetation may play a very important role in the epidemiology of plant viruses such as TSWV that cause severe virus epidemics in tomato crops (Kahn et al., 2005). The objective of our work was to analyze the degree of susceptibility of selected candidate insectary plant species to the major viruses that infect tomato crops in Spain, and to study their likely role as virus reservoirs. Materials and methods Virus survey of insectary plants A virus survey of selected insectary plant species was conducted in two separate vegetable producing regions of Spain (Murcia and Barcelona). A total of 104 samples of M. vulgare, D. viscosa, O. natrix and C. corymbosa were collected in spring of 2004 from Mazarrón, Murcia. Other 97 samples from O. natrix and D. viscosa were collected in autumn of 2004 and 2005 from various locations at Barcelona. All samples were analysed by ELISA to test for the presence of CMV, PVY, TSWV and TYLCV, which are the main viruses commonly found in the tomato crops in the regions under study. No obvious symptoms of virus infection were observed in the plants sampled, and therefore, samples were collected at random from the margins or across the field in the selected locations. Transmission experiments The viruses that were found present in our survey (CMV and PVY) were inoculated by aphids to healthy insectary test plants to assess the role of such plants as virus reservoirs. Both viruses were inoculated by aphids to several healthy test plants of M vulgare, D. viscosa, C. corymbosa, O. natrix and V. thapsus. CMV was inoculated by Myzus persicae (Sulzer) and Aphis gossypii Glover, and PVY was inoculated by M. persicae only. Transmission experiments were conducted under laboratory conditions (21±2 ºC) using a procedure described by Fereres et al. (1993) with slight modifications. In summary, aphids were subjected to a 5 min acquisition access period on infected-pepper source plants, and then groups of aphids were transferred to each test plant for a 2h inoculation access period. Test plants were sprayed with imidacloprid and tested by ELISA to assess virus infection. The insectary test plants that became infected by either PVY or CMV were used as source plants to run a second series of transmission tests to test their role as virus reservoirs. For such tests, the same transmission protocol was used and the viruses were transmitted by aphids from each infected insectary plant to healthy tomato test plants (28 replicates). ELISA-assays Viruses were identified by ELISA-assay using specific commercial antibodies against the following four virus species: CMV, TSWV, PVY and TYLCV. The procedure used was a DAS-ELISA (Clark & Adams, 1977). Plants were considered infected when absorbance values reached three times the mean value of the healthy controls. Healthy samples of D. viscosa, M. vulgare, C. corymbosa and V. thapsus were obtained from seeded plants grown in virus-free environmental chambers. 256
269 Results and discussion Virus survey of insectary plants The only two viruses detected in the plants sampled were CMV and PVY (Table 1). PVY was detected in a single M. vulgare from Murcia and in 3 D. viscosa plant samples collected in Barcelona. CMV was found in 3 D. viscosa plants in samples collected in Barcelona. The natural occurrence of CMV infecting M. vulgare has been reported by Fletcher (2001) who found asymptomatic infected plants in a field survey conducted in New Zealand. These results show that both M. vulgare and D. viscosa appear to be infected in the field by PVY and CMV. The two other viruses (TYLCV and TWSV) included in our survey were never found in none of the insectary plants sampled. Also, none of the samples obtained from C. corymbosa and O. natrix appeared to be infected with any of the viruses included in our survey. However, TSWV was found in natural infections on M. vulgare in field surveys conducted in North America (Sclar et al., 2001) and Australia (Latham & Jones, 1997). Natural infections of TSWV were also found in V. thapsus in North America (Sclar et al., 2001). Table 1. Virus detection in selected insectary plants sampled in different locations of Spain during 2004 and 2005 Viruses present a Locality Host plant N b CMV PVY TSWV TYLCV D. viscosa Murcia M. vulgare C. corymbosa O. natrix Barcelona D.viscosa O. natrix Total a number of infected plants; b number of plants sampled. Transmission experiments The results obtained in the transmission experiments of the viruses CMV and PVY to insectary test plants are shown in Table 2. Transmission of CMV was positive in the case of D. viscosa and M. vulgare and PVY was readily infecting V. thapsus (20/28) but none of the other insectary plants tested became infected. However, M. vulgare and D. viscosa were found infected by PVY in our field survey although we failed to inoculate the virus when using M. persicae as a vector. It is possible that other aphid species are involved in the transmission of PVY to M. vulgare and D. viscosa in the field. We found that C. corymbosa and O. natrix never became infected with PVY or CMV after aphid inoculation although the number of test plants was low and further experiments are needed to confirm this result. The results on the role of insectary plants as sources of inoculum of both CMV and PVY are shown in Table 3. Tomato test plants became infected by CMV when using D. viscosa or M. vulgare as infected source plants. However, tomato plants did not become infected with PVY when the virus was acquired from V. thapsus infected plants. This result suggests that V. thapsus might be susceptible to PVY inoculation but may not be a natural effective source 257
270 for virus acquisition by aphid vectors. Table 2. Transmission of CMV and PVY to selected insectary plant species by A. gossypii and M. persicae. Virus source Test plant Vector CMV-infected pepper PVY-infected pepper NT: not tested A. gossypii M. persicae D. viscosa 1/14 0/14 M. vulgare 1/13 1/13 O. natrix 0/3 0/4 C. corymbosa 0/4 0/4 V. thapsus 0/14 0/14 D. viscosa NT 0/47 M. vulgare NT 0/18 O. natrix NT 0/7 C. corymbosa NT 0/7 V. thapsus NT 20/28 Table 3. Transmission of CMV and PVY from infected insectary plants to tomato test plants by A. gossypii and M. persicae Virus-Source CMV-infected D. viscosa CMV-infected M. vulgare PVY-infected V. thapsus NT: not tested. Vector A. gossypii M. persicae 1/15 0/15 0/14 1/14 NT 0/28 In summary, our results suggest that susceptibility of M. vulgare to virus transmission by aphid vectors is low, although it is likely to occur when a significant number of viruliferous aphids land on the plant. Therefore, there is a potential risk for virus epidemics to occur if M. vulgare and D. viscosa are introduced as insectary plants to attract and enhance biological control by mirid bugs such as M. caliginosus. However, more experiments are needed to confirm the role that M. vulgare and D. viscosa may play in the epidemics of PVY and CMV in tomato crops. We also found that V. thapsus was susceptible to the transmission of PVY by aphids although it was not a source of inoculum for acquisition of the virus. Therefore, this species needs to be investigated further before its release as an insectary plant because it has been reported as a susceptible host species of TSWV. Finally, we have shown that O. natrix and C. corymbosa were never found infected by none of the viruses tested in our field survey and were not susceptible to virus inoculation by aphids under laboratory conditions and 258
271 therefore, they are good candidates to be used as insectary plants from the point of view of avoiding insect-transmitted virus diseases. Acknowledgements This work has been supported by the Spanish Ministry of Education and Science, research grant no. AGL C03. References Albajes, R. & Alomar, O. 1999: Current and potential use of polyphagous predators. In: Integrated pest and disease management in greenhouse crops, eds. Albajes, Gullino and Van Lenteren & Elad: Kluwer Academics Publishers, Dordrecht, The Netherlands. Alomar, O., Goula, M. & Albajes, R. 2002: Colonization of tomato fields by predatory mirid bugs (Hemiptera: Heteroptera) in northern Spain. Agr. Ecosyst. Environ. 89: Arnó, J., Ariño, J., Español, R., Martí, M. & Alomar, O. 2000: Conservation of Macrolophus caliginosus Wagner (Het.: Miridae) in commercial greenhouses during tomato crop-free periods. IOBC/WPRS Bull. 23(1): Castañé, C., Alomar O., Goula, M. & Gabarra, R. 2000: Natural populations of Macrolophus caliginosus and Dicyphus tamaninii in the control of the greenhouse whitefly in tomato crops. IOBC/WPRS Bull. 23(1): Clark, M.F. & Adams, A.N. 1977: Characteristics of the microplate method of enzyme-linked inmunoabsorbent assay for the detection of plant viruses. J. Gen. Virol. 34: Fereres, A., Pérez, P., Gemeno, C., & Ponz, F. 1993: Transmission of Spanish Pepper- and Potato-PVY Isolates by Aphid (Homoptera: Aphididae) Vectors: Epidemiological Implications. Environ. Entomol. 22: Fletcher, J.D. 2001: New hosts of Alfalfa mosaic virus, Cucumber mosaic virus, Potato virus Y, Soybean dwarf virus, and, Tomato spotted wilt virus in New Zealand. New Zealand Journal of Crop and Hort. Sci. 29(3): Gabarra, R. 2002: Control integrado de moscas blancas y pulgones en cultivos de invernadero. 12º Symposium internacional. Ecología y Producción integrada en cultivos hortícolas de invernadero. Phytoma España 135: Gafni, Y. 2003: Tomato yellow leaf curl virus, the intracellular dynamics of a plant DNA virus. Mol. Plant Pathol. 4(1): Kahn, N.D., Walgenbach, T.F. & Kennedy, G.G. 2005: Summer weeds as host for Frankliniella occidentalis and Frankliniella fusca (Thysanoptera: Thripidae) and as reservoirs for tomato spotted wilt tospovirus in North Carolina. J. Econ. Entomol. 98(6): Lacasa A. 1990: Un trienio de Frankliniella occidentalis en España, evolución temporal y espacial de una plaga importada. Phytoma España 6: 3-8. Pascual-Villalobos, M.J., Lacasa, A., González, A., Varó, P. & García, M.J Effect of flowering plant strips on aphid and syrphid populations in lettuce. Eur. J. Agron. 24: Sanchez, J.A., Gillespie, D.R. & Mcgregor, R.R. 2003a: The effects of mullein plants (Verbascum thapsus) on the population dynamics of Dicyphus hesperus (Heteroptera : Miridae) in tomato greenhouses. Biol. Control 28(3):
272 Sanchez, J.A., Martínez-Cascales, J.I., Lacasa, A. 2003b: Abundance and wild host plants of predator mirids (Heteroptera: Miridae) in horticultural crops in the Southeast of Spain. IOBC/WPRS Bull. 26(10): Sclar, C., Anisko T. et al. 2001: Tospovirus Host List, Longwood Gardens, Kennett Square PA. 3pp. 260
273 Plant diseases
274 262
275 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Biological Control of Fusarium oxysporum f.sp. melonis race 1.2 by cross protection Hela Chikh-Rouhou, José Maria Álvarez, Rafael González-Torres Centro de Investigación y Tecnología Agroalimentaria de Aragón, Unidad de Tecnología en Producción vegetal, Unidad de Sanidad Vegetal, P.O. Box 727, Zaragoza, Spain Abstract: The melon cultivar Charentais T susceptible to Fusarium oxysporum f.sp. melonis (Fom) race 1.2 was pre-inoculated with a non-pathogenic isolate of Fom. The biocontrol agent was only effective when it was applied one or two days before a challenge inoculation with the pathogen. Simultaneous inoculation did not provide any durable protection. The effectiveness of cross-protection was not evident in the case of the pathogenic isolate belonging to pathotype 1.2Y, since all plants died by the end of the test. However, this did not happen when the pathogenic isolate was from pathotype 1.2W. Protection, consisting of a delay in the expression of symptoms and a reduction in the disease index, was apparent from the beginning of the experiments. Similarly, the ratio between inducer and challenger ( spores/ml : spores/ml) gave the highest cross protection. These results indicate that cross-protection required a certain time and an adequate concentration of the inducer organism to become effective. The need for a time interval between induction and challenge inoculation for maximum protection suggests that protection is conditioned by prolonged fungus-plant interaction and regulated by the consequent metabolic changes in the plant. Key words: Fusarium wilt, melon, Cucumis melo, biocontrol agent, physiological races. Introduction Fusarium wilt of melon (Cucumis melo L.) caused by Fusarium oxysporum f.sp. melonis (L and C) Synd. and Hansen (Fom) inflicts an important yield loss throughout the world (Katan et al., 1994). The pathogen causes wilting by colonizing the plant vascular system and eventually results in seedling or adult plant mortality. Four races of the pathogen - 0, 1, 2, and are known to be virulent in melon cultivars (Risser et al., 1976). Resistance to races 1 and 2 is conferred by two single dominant genes Fom-1, and Fom-2 respectively. Both genes also confer resistance to race 0 (Risser et al., 1976; Zink & Thomas, 1990). Race 1.2, which is able to overcome these two resistant genes, is subdivided into race 1.2Y, which causes symptoms of yellowing, and race 1.2W, which causes wilt symptoms (Risser et al., 1976). A number of melon accessions from the Far East show partial resistance to Fom race 1.2 (Risser & Rode, 1973). It has recently been shown that this potential resistance is due to polygenic control (Perchepied et al., 2005). This type of resistance is difficult to introduce into commercial cultivars, and only a few have been developed by incorporating resistance to Fom; most are only used as rootstocks. This is why other disease control methods, like crossprotection, must be explored. Cross-protection is considered useful for the biological control of soil borne diseases. Cross-protection against F. oxysporum has been previously reported in several hosts, having been induced by different Verticilium spp. (Jorge et al., 1992), saprophytic F.oxysporum isolates (Fuchs et al., 1997), pathogenic formae specialis of other hosts (Biles & Martyn, 1989) or avirulent races of the same formae specialis (Martyn et al., 1991). The proposed 263
276 mechanisms involved in the biological control of Fusarium spp. include (i) competition for infection sites (Mandeel & Baker, 1991), (ii) competition for nutrients (Alabouvette, 1990), and (iii) local and systemic induced resistance (Biles & Martyn, 1989). Success in obtaining biological control depends upon a number of factors, including the particular formae specialis of F. oxysporum used as the cross-protection agent, the concentration at which it is applied, and the period separating the challenge and the inducing inoculation. This study attempted to determine the effectiveness of cross-protection in the control of race 1.2 of Fusarium oxysporum of melon, by prior inoculation with a non-pathogenic Fusarium oxysporum isolate, and also some of the factors affecting this phenomenon. Material and methods Isolate is a non-pathogenic isolate (NPI) originating from a Fom race 1.2W isolate, which lost its pathogenicity after conservation at -20ºC since Isolates 37mls and Fom0125 are pathogenic isolates (PI) belonging to races 1.2 W and 1.2Y respectively. The plant material was melon cv Charentais T, which is susceptible to all races of Fom. Artificial inoculation was performed by continuously dipping the roots of intact seedlings in plastic pots containing 200 ml of a nutrient solution and 10 ml of a conidial suspension of the appropriate fungus isolate. Pots were then placed in an orbital shaker operating at 120 r.p.m in a growth chamber at 28/20ºC (day/night), and with a light intensity of µeistein/m 2 for 14h/day. Inoculation of plants with the appropriate PI was performed at 0h, 4h, 24h and 48h, after inoculation with the NPI. The spore concentrations of the two isolates were 3x10 6 spores/ml for the PI, and the three different concentrations for the NPI were 3x10 7 spores/ml, 3x10 6 spores/ml and 3x10 5 spores/ml. The experimental design was a randomized complete block design; four replications were used per treatment with four plants per replication. Plants were examined 15, 30 and 45 days after inoculation, and the number of yellowed, wilted or dead seedlings was recorded. Disease severity was assessed using the following index: (0) no apparent symptoms; (1) first signs of wilt symptoms in leaves; (2) heavily wilted leaves; (3) all leaves completely wilted; (4) dead plant. For statistical analysis, two-way ANOVA was performed, using the SAS software. Results Both pathogenic isolates, Fom0125 and 37mls, were pathogenic on Charentais T, producing yellowing and wilting, respectively, followed by plant death. The Fom0125 isolate (1.2Y) appeared to be much more aggressive than 37mls (1.2W) (Table 1). By 45 days after inoculation with Fom0125 all plants had died, although plants pre-inoculated with NPI and then inoculated with 37mls were still alive. Given the different behaviour of the two PI, the statistical analyses of the data from each isolate were performed separately. Simultaneous inoculation Simultaneous inoculation of Charentais T with the NPI did not provide any protection against the pathogenic isolate Fom0125 (Table 1). However, when the PI was 37mls, crossprotection resulted in both a delay in symptom expression and a reduction in the disease index (Table 1). Thirty days after inoculation with the PI there was no cross-protection for any PI and NPI/PI concentration ratio, and all of the Charentais T plants had either died or were about to die.. 264
277 Table 1: Severity reaction of Charentais T plants after pre-inoculation with the nonpathogenic isolate (NPI) and further inoculation with the pathogenic isolates (PI) Fom 0125 or 37mls of Fusarium oxysporum f.sp. melonis at different concentrations and intervals between inoculations. Days after inoculation with the PI Interval between inoculations Treatments 0h 4h 24h 48h 0h 4h 24h 48h 0h 4h 24h 48h Control 0.0 x NPI Fom 0125 (Y) 4.0a y a a NPI(3x10 7 ) Z +Y(3x10 6 ) 3.9a 3.1b 0.0d 0.0d 4.0a 4.0a 3.0b 3.1b 4.0a 4.0a 4.0a 4.0a NPI(3x10 6 ) +Y(3x10 6 ) 4.0a 3.8a 0.5d 0.1d 4.0a 4.0a 3.3b 3.0b 4.0a 4.0a 4.0a 4.0a NPI(3x10 5 ) +Y(3x10 6 ) 4.0a 3.8a 2.0c 3.2b 4.0a 4.0a 3.8a 4.0a 4.0a 4.0a 4.0a 4.0a 37mls (W) 3.6d a a NPI(3x10 7 )+W(3x10 6 ) 0.8a 0.4b 0.0c 0.0c 3.8a 0.6c 0.4c 1.3d 4.0a 1.6c 1.4c 2.3d NPI(3x10 6 )+W(3x10 6 ) 0.8a 0.8a 0.0c 0.1c 4.0a 4.0a 1.7b 2.1b 4.0a 4,0a 2.9b 3.0b NPI(3x10 5 )+W(3x10 6 ) 0.8a 0.7a 0.0c 0.1c 4.0a 4.0a 4.0a 4.0a 4.0a 4.0a 4.0a 4.0a ( x )Disease severity was recorded using a scale from 0 to 4 (0= no symptoms; 4= dead plant). Figures are the means from 16 plants per treatment. ( y ) For each PI (Y or W) and number of days since inoculation with PI, means followed by the same letter are not significantly different (P = 0.05). ( Z ) Concentration: spores/ml. Effect of time separation In general, the level of protection offered to the Charentais T cultivar against the PI increased in line with the time between inoculations with the NPI (inducer) and the PI (challenger) (Table 1). This effect was most evident when the PI was 37mls (W): in this case, protection increased when separation between the two inoculations increased from 0h to 48h, regardless of the number of days since inoculation with PI. Nevertheless, when the PI Fom0125 was used, there was no protection at all by 45 days after inoculation with PI. Furthermore, the degree of protection was always less evident than that observed when the isolate 37mls was used. The best cross-protection results were observed when the intervals between the application of the NPI and the PI were 24h or 48h; cross-protection resulted in a delay in symptom expression and also in a reduction in the disease index. Effect of inoculum concentration A dosage-response effect related to the NPI/PI ratio of the conidia concentration was observed. Concentrations of the NPI equal to or higher than that of the PI resulted in protection, whereas concentrations of the NPI lower than that of the PI did not. This effect was much more evident when 37mls was used as the PI. The PI Fom0125 appeared to be more aggressive and was able to produce death or severe damage to the plants as early as 30 days after PI inoculation, regardless of the concentration ratio. 265
278 Discussion It was possible to protect melon plants from the effect of a virulent strain of Fom by preinoculation with a non-pathogenic isolate. The cross-protection provided by the nonpathogenic isolate was expressed by a delay in the appearance of symptoms, regardless of the pathogenic isolate used. In the case of the pathogenic isolate Fom 0125 (1.2Y), all plants had died by the end of the test. This did not happen when the pathogenic isolate was 37mls (1.2W). This different behaviour for the two isolates could be related to the greater aggressiveness of the former. Some authors have reported that pathogens closely related to the challenge isolate, such as avirulent races of formae specialis, were better inducers of resistance than non-pathogens or pathogens of unrelated hosts (Biles & Martyn, 1989; Martyn et al., 1991). Mas et al. (1981) with Fusarium wilt of melon reported that avirulent races of Fom (race 0 or 1) were effective inducers of resistance to a virulent race (race 1 or 2). This could explain why our NPI (originated from an isolate of pathotype 1.2W) was a better inducer of resistance when the PI was 37mls (pathotype 1.2W). A delay in the expression of symptoms in plants pre-inoculated with NPI, was observed by many authors (Wymore & Baker, 1982; Huertas-Gonzalez et al., 1999). The effect of the separation of the two inoculations (inducer and challenger) was also reported (Wymore & Baker, 1982; Davis, 1966, 1967). Only temporal separation of inoculations of the inducer and challenger provided significant protection. Simultaneous inoculation did not provide any protection, indicating that direct interaction between the two organisms at the site of infection was not required. The need for a time interval between the two inoculations in order for cross-protection to be effective also suggests that protection must be conditioned by inducer-plant interaction and regulated by consequent metabolic changes to the plant occurring after protective inoculation. Matta (1989) reported that time separation between the two inoculations did not allow the non-pathogenic isolate to acquire or increase its ability to compete with the pathogen for sites and nutrients. Our results therefore suggest that changes in plant metabolism were responsible for cross-protection. The degree of protection depended on the ratio between the respective spore concentrations of the inducer and challenger isolates. Other researchers have also reported the need for equal or greater amounts of inducer than challenger inoculum to provide significant protection to tomato against F.oxysporum f.sp. lycopersici (Wymore & Baker, 1982). Acknowledgments This research was supported by Project AGL CO2-02 of the Spanish Ministry of Education and Science, and the European Fund for Regional Development (FEDER). Hela Chikh-Rouhou has received a pre-doctoral fellowship from the Agencia Española de Cooperación Internacional (AECI). References Alabouvette C. 1990: Biological control of Fusarium wilt pathogens in suppressive soils. In: Biological Control of Soilborne Plant Pathogens. D. Homby, ed. CAB International, Wallingford, UK Biles C.L. & Martyn R.D. 1989: Local and systemic resistance induced in watermelons by formae specialis of Fusarium oxysporum. Phytopathology 79: Davis D. 1966: Cross infection in Fusarium Wilt diseases. Phytopathology 56:
279 Davis D. 1967: Cross-Protection in Fusarium Wilt diseases. Phytopathology 57: Fuchs J.G, Moënne-Locoz Y. & Defago G Nonpathogenic Fusarium oxysporum strain Fo47 induces resistance to Fusarium wilt in tomato. Plant Dis. 81: Huertas-González M.D, Ruiz-Roldán M.C. & Di Pietro A. 1999: Cross protection provides evidence for race-specific avirulence factors in Fusarium oxysporum. Physiol. Mol Plant Pathol. 54: Jorge P.E, Green R.J.J. & Chaney W.R. 1992: Inoculation with Fusarium and Verticilium to increase resistance in Fuarium-resistant tomato. Plant Dis. 76: Katan T, Katan J, Gordon T. & Pozniak D. 1994: Physiologic races and vegetative compatibility groups of Fusarium oxysporum f.sp melonis in Israel. Phytopathology 84: Mandeel Q. & Baker R. 1991: Mechanisms involved in biological control of Fusarium wilt of cucumber with strains of non-pathogenic Fusarium oxysporum. Phytopathology 81: Mas P, Molot P.M. & Risser G. 1981: Fusarium wilt of muskmelon. En: Nelson PE, ToussenTA, Cook RJ, eds. Fusarium: Disease, Biology and taxonomy. University Park, PA,USA: Pennsylvania State University Press, Matta A. 1989: Induced resistance to Fusarium wilt diseases. In: Tjamos E.C.& Beckman, C. Eds, vascular wilt Diseases of Plants: Martyn R.D, Biles CL. & Dillard E.A. 1991: Induced resistance to Fusarium wilt of watermelon under simulated field conditions. Plant Dis. 75: Perchepied L, Dogimont C. & Pitrat M. 2005: Strain specific and QTLs involved in the control of partial resistance to Fusarium oxysporum f.sp. melonis race 1.2 in a recombinant inbred line population of melon. Theor. Appl. Genet. 111: Risser G. & Rode J.C. 1973: Breeding for resistance to Fusarium oxysporum f.sp. melonis. In: Eucarpia: La selection du melon. Montfavet Avignon, Risser G, Banihashimi Z. & Davis D.W : A proposed nomenclature of Fusarium oxysporum f.sp. melonis races and resistance genes in Cucumis melo. Phytopathology 66: Wymore L.A. & Baker R. 1982: Factors affecting cross-protection in control of Fusarium wilt of tomato. Plant Dis. 66: Zink F. & Thomas C.E. 1990: Genetics of resistance to Fusarium oxysporum f.sp. melonis races 0, 1 and 2 in muskmelon line MR-1. Plant Dis. 80:
280 268
281 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Evaluation of susceptibility of varieties of Gerbera jamesonii to Fusarium wilt Andrea Minuto, Domenico Bertetti, Maria Lodovica Gullino, Angelo Garibaldi Center of Competence for Innovation in the agro-environmental sector, University of Torino, Via Leonardo da Vinci 44, Grugliasco, Italy. Abstract: This paper reports an evaluation of the sensitivity of gerbera cultivars to Fusarium wilt. The data obtained by inoculating 79 cultivars with a strain of Fusarium oxysporum (MASS1) originating from gerbera and with a strain of F. oxysporum f.sp. chrysanthemi (REP8) obtained from chrysanthemum allowed us to classify the gerbera strain as F. oxysporum f.sp. chrysanthemi. Moreover, the percentage of resistant and tolerant varieties was considered sufficient to satisfy grower needs in the cut flower sector. Key words: gerbera, Fusarium oxysporum f.sp. chrysanthemi, genetic resistance. Introduction In summer 2002 gerbera (Gerbera jamesonii cv Kaiki) plants exhibiting symptoms of a wilt disease were observed in a soilless cultivation system at Albenga (Savona) in northern Italy. A similar wilt was observed in summer 2002 on a commercial gerbera farm, in the province of Imperia (northern Italy), on cv Red Bull, Anedin and Gud finger grown in soil. In both cases, the planting material originated from the Netherlands. In 2003, wilted plants were repeatedly observed in other commercial greenhouses, in the same area, on cv Red Bull, Basic and Cirill (Garibaldi et al., 2004). The affected plants were stunted and had yellowed leaves with initially brown and eventually black streaks in their vascular systems. In some cases the leaves of affected plants turned red. Fusarium oxysporum was identified as the causal agent of the disease (Garibaldi et al., 2004). Several flower crops belonging to the Compositae family were inoculated and Fusarium oxysporum from gerbera was determined as a strain of the f.sp. chrysanthemi (Garibaldi et al., in preparation). Based on preliminary experimental trials, gerbera plants appeared to be more sensitive to the disease when grown in open and/or closed soilless systems on artificial/inorganic substrates and, particularly, under hot climate conditions (temperature > 25 C). This paper summarizes data collected during two years of testing the susceptibility of several cultivars of gerbera to the disease. Material and methods Experimental trials in open soilless systems were carried out at the Agricultural Experimental Center of Albenga (northern Italy). In the first (2004) and second trial (2005), 75-day-old gerbera plants grown in 4x4x5 cm rockwool cubes during the nursery stage were transplanted into plastic pots (19 cm diameter and 5 l volume) filled with a rockwool growing medium (Growcubes Grodan). Seventy-nine gerbera varieties were selected on the base of their market availability and their relative importance within the gerbera market. The plants were maintained on concrete benches with a drip irrigation system (with one water emitter for each plant and a flow rate of 6 l/hour) in an open soilless crop. A nutrient solution (N-NO mg 269
282 l -1 ; P-P 2 O 5 98 mg l -1 ; K-K 2 O 266 mg l -1 ; Mg-MgO 35 mg l -1 ; Ca 204 mg l -1 ) was delivered at least 5 times/day (200 ml nutrient solution/single irrigation per plant). Relevant data from trials 1 and 2 are synthetically reported in Table 1. Pathogen inoculation One strain of F. oxysporum isolated from the gerbera plant (MASS1) (Garibaldi et al., 2004) and one strain of F. oxysporum f.sp. chrysanthemi isolated from the chrysanthemum (REP 8) (Pasquali et al., 2003) were respectively tested for pathogenicity on gerbera and chrysanthemum. The inoculum was obtained by incubating each strain on casein liquid medium for days, placing it on a rotary shaker and then centrifuging. Artificial inoculation was carried out by suspending the homogenized pathogenic biomass (conidia, mycelium and chlamidospores) in deionised and sterile water to a final concentration of 10 7 CFU/ml and then directly dipping the roots of gerbera plants into the inoculum immediately before transplanting. Each variety was randomised in three independent cultural rows, with 5 plants per replicate. For each variety, three replicates of 5 plants were also grown as a noninoculated control. Evaluation of disease infection Evaluations of the severity of Fusarium wilt on inoculated gerbera plants were carried out every 4-7 days by counting and removing the dead plants. Sections of the crowns of diseased plants were regularly surface-disinfested and plated on potato dextrose agar amended with antibiotic and Fusaria selective medium (Komada, 1975) at 25 C for h, to check for the presence of F. oxysporum. At the end of the trial, all plants were uprooted and the presence of symptoms of Fusarium wilt was visually assessed using a 0 to 100 scale on which 0 indicated the absence of symptoms, 25 indicated the unilateral presence of yellow leaves, 50 indicated the unilateral presence of yellow leaves together with brown and eventually black streaks in the vascular system, 75 indicated the unilateral presence of yellow and eventually withered leaves with brown and eventually black streaks in the vascular system accompanied with initial collapse symptoms, and 100 indicated plant withering and death. Statistical analysis The data collected were statistically analyzed using SPSS for Windows software. Prior to analysis, percentage data were arcsine transformed to improve the homogeneity of variances. The sources of variation were treatments and blocks. The effects of disease in each variety were examined using ANOVA and when the F-test was significant at P<0.05, treatment means were compared using the Tukey HSD test. Table 1. Relevant dates of trials 1 and 2. Trial 1 Trial 2 Transplant and artificial inoculation 30/07/ /06/2005 First symptoms appearance 13/08/ /07/2005 Final evaluation 15/10/ /09/2005 Temperature average minimum maximum
283 Results and discussion In the two trials, cultivars tested for resistance/susceptibility gave similar results when inoculated with the two strains: there were few exceptions (Tables 2a, 2b, 2c). The susceptibility of the gerbera varieties to the strain of F. oxysporum originating from gerbera and to F. oxysporum f.sp. chrysanthemi obtained from chrysanthemum was similar. Among the tested cultivars, 59%, 22%, 5% and 14% % were respectively resistant, tolerant, susceptible and very susceptible to the strain isolated from gerbera, while 47%, 32%, 4% and 18% were respectively resistant, tolerant, susceptible and very susceptible to F. oxysporum f.sp. chrysanthemi. These data, together with other information collected from experimental trials (Garibaldi et al., in preparation), confirmed the identification of the strains of F. oxysporum obtained from gerbera as F. oxysporum f.sp. chrysanthemi. Only Von Arx had previously reported the presence of Fusarium wilt infections on gerbera plants in the Netherlands, back in 1948 (Arx, 1948). Gordon identified the strains of F. oxysporum isolated by Von Arx as f.sp. gerberae (Gordon, 1965). Unfortunately, as the Fusarium strains isolated by Von Arx are not now available, we cannot definitively clarify the relationship between those Dutch strains and the ones isolated in Italy. In conclusion, even if the varieties tested in our trials cannot be considered exhaustive for the cultivar currently available on the market, the percentage of resistant and tolerant varieties can be considered sufficient to fulfil grower needs in the cut flower sector. Acknowledgements This work was supported by grants from the University of Torino and the Regione Liguria. References Arx, J.A. Von 1948: De voetziekte van Gerbera, veroorzaakt door Fusarium oxysporum Schlecht. Tijdschr. PlZiekt. 58: 5-9. Garibaldi, A., Minuto, A., Bertetti, D. & Gullino, M.L. 2004: Fusarium wilt of gerbera in soil and soilless crops in Italy. Plant Dis. 88: 311. Gordon, W.L. 1965: Pathogenic strains of Fusarium oxysporum. Can. J. Bot. 43: Komada, H. 1975: Development of selective medium for quantitative isolation of Fusarium oxysporum from natural soil. Rev. Plant Prot. Res. 8: Pasquali, M., Acquadro, A., Balmas, V., Migheli, Q., Garibaldi, A. & Gullino, M.L. 2003: RAPD Characterization of Fusarium oxysporum isolates pathogenic on Argyranthemum frutescens L. J. Phytopathology 151:
284 Table 2a. Evaluation of the severity of Fusarium wilt on gerbera plants inoculated with strains from gerbera and chrysanthemum (% of surviving plants; disease index Albenga, ). Cultivar % of surviving plants disease index MASS1 REP8 MASS1 REP8 Trial 1 Trial 2 Trial 1 Trial 2 Trial 1 Trial 2 Trial 1 Trial 2 Belladonna 83 ab 92 a 67 a-c 50 b-d 30 a-d 30 a-c 33 a-c 53 cd Cadillac 100 a 100 a 100 a 75 a-c 45 a-e 8 ab 28 a-c 25 a-c Carillon 100 a 100 a 100 a 100 a 18 ab 0 a 23 ab 0 a Charleston^ 67 a-c 25 cd 89 a 0 f 43 a-e 80 d-e 30 a-c 100 f Conga^ 17 cd 42 bc 72 ab 39 c-e 93 ef 63 c-e 35 a-c 65 de Corazon 100 a 33 b-d 78 ab 0 f 43 a-e 80 d-f 48 a-c 100 f Cyrano + 89 ab 100 a 100 a 100 a 10 a 0 a 0 a 0 a Dino 89 ab 0 d 100 a 0 f 23 a-c 100 f 5 a 100 f Duchessa rosa 100 a 100 a 89 a 100 a 0 a 0 a 10 a 0 a Evergreen 100 a 100 a 100 a 100 a 0 a 0 a 0 a 0 a F a 100 a 100 a 100 a 15 a 0 a 5 a 0 a F a 100 a 75 ab 100 a 0 a 0 a 25 ab 0 a Floriade 100 a 100 a 100 a 100 a 8 a 0 a 0 a 0 a G a 100 a 89 a 100 a 0 a 0 a 10 a 0 a Happy Hour^ 17 cd 8 cd 44 a-c 8 ef 83 def 95 ef 55 a-c 93 ef High five^ 89 ab 100 a 89 a 100 a 10 a 0 a 10 a 0 a Marlene 100 a 100 a 67 a-c 100 a 0 a 0 a 33 a-c 0 a Maxx 100 a 100 a 100 a 100 a 5 a 0 a 5 a 0 a Napoli 100 a 100 a 67 a-c 100 a 0 a 0 a 33 a-c 0 a Natasha 100 a 100 a 100 a 100 a 5 a 0 a 5 a 0 a Passion 67 a-c 33 b-d 56 a-c 0 f 68 b-f 80 d-e 58 a-c 100 f Pinky springs x 83 ab 100 a 100 a 100 a 43 a-e 0 a 5 a 0 a Purple springs x 100 a 100 a 100 a 100 a 0 a 0 a 5 a 5 ab Red star 100 a 100 a 100 a 100 a 0 a 0 a 5 a 0 a Shakira 100 a 100 a 100 a 100 a 0 a 0 a 0 a 0 a Soleada 100 a 100 a 100 a 100 a 5 a 0 a 0 a 0 a Stephanie 83 ab 100 a 72 ab 100 a 20 ab 0 a 28 a-c 0 a 100 a 100 a 100 a 100 a 5 a 0 a 0 a 0 a Tucan Virginia 100 a 25 cd 100 a 8 ef 30 a-d 88 d-e 25 ab 95 ef Yellow springs x 72 a-c 92 a 83 a 75 a-c 30 a-d 8 ab 18 a 25 a-c Results in the same column that do not share a common letter are significantly different according to the Tukey HSD test (P=0.05). (, ^, +, x ) Cultivar type: Standard; ^ Germini; + Muppet; x Springs. 272
285 Table 2b. Evaluation of the severity of Fusarium wilt on gerbera plants inoculated with strains from gerbera and chrysanthemum (% of surviving plants; disease index Albenga, ). Cultivar % of surviving plants disease index MASS1 REP8 MASS1 REP8 Trial 2 Trial 2 Trial 2 Trial 2 Advance 67 ab 92 a 33 a-c 25 a-c Arianna 100 a 100 a 0 a 3 a Axxion 92 a 92 a 8 ab 8 ab Carambole 92 a 33 d-f 43 bc 83 ef Cirillo 0 d 0 f 100 f 100 f Cornice 100 a 83 ab 0 a 33 bc Dalma 0 d 0 f 100 f 100 f Dameblanche 100 a 100 a 0 a 0 a Dune 42 bc 8 ef 60 cd 95 ef Excellence 17 cd 0 f 88 d-f 100 f Finola 100 a 100 a 0 a 0 a Fredonzelle 100 a 100 a 0 a 0 a Ginger 100 a 100 a 0 a 5 ab Jaska 0 d 0 f 100 f 100 f Malibù 100 a 100 a 0 a 0 a Meriva 0 d 0 f 100 f 100 f Mexx 100 a 100 a 0 a 0 a Mont blanc 100 a 100 a 0 a 0 a Okey 100 a 100 a 0 a 8 ab Pink elegance 100 a 100 a 0 a 0 a Ruby red 100 a 100 a 0 a 8 ab Sunway 100 a 100 a 0 a 0 a Tropic blend 92 a 92 a 10 ab 10 ab Yucatan 0 d 0 f 100 f 100 f Zembla 100 a 100 a 0 a 0 a Results in the same column that do not share a common letter are significantly different according to the Tukey HSD test (P=0.05). (, ^, +, x ) Cultivar type: Standard; ^ Germini; + Muppet; x Springs. 273
286 Table 2c. Evaluation of the severity of Fusarium wilt on gerbera plants inoculated with strains from gerbera and chrysanthemum (% of surviving plants; disease index Albenga, ). Cultivar % of surviving plants disease index MASS1 REP8 MASS1 REP8 Trial 1 Trial 1 Trial 1 Trial 1 Alfred 100 a 67 a-c 13 ab 33 a-c Bahia a 100 a 0 a 0 a Black Jack 100 a 100 a 0 a 0 a Cariba 100 a 100 a 0 a 0 a Cassiana 100 a 100 a 3 a 0 a Floyd a 100 a 0 a 0 a Friends a 100 a 0 a 0 a Hamlet a 100 a 0 a 0 a Honduras 100 a 100 a 0 a 0 a Hope 100 a 100 a 0 a 0 a Isaac 100 a 100 a 0 a 0 a Paintball^ 100 a 100 a 0 a 0 a Primadonna 0 d 11 bc 100 f 93 c Red fashion 100 a 100 a 0 a 0 a Sahara 100 a 100 a 0 a 0 a Schumy 100 a 100 a 0 a 0 a Sharky 100 a 100 a 0 a 0 a Snowdance^ 100 a 100 a 0 a 0 a Soul^ 100 a 100 a 0 a 0 a Spitfire 100 a 100 a 0 a 0 a Terra Gina^ 100 a 100 a 0 a 0 a Varenne 100 a 100 a 0 a 0 a Zenith^ 100 a 100 a 0 a 0 a Zeppelin^ 100 a 100 a 0 a 0 a Results in the same column that do not share a common letter are significantly different according to the Tukey HSD test (P=0.05). (, ^, +, x ) Cultivar type: Standard; ^ Germini; + Muppet; x Springs. 274
287 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Efficacy of Ampelomyces quisqualis isolate M-10 (AQ 10 ) against powdery mildews (Erysiphaceae) on protected crops Massimo Benuzzi¹, Edith Ladurner¹, Fernando Mayoral², ¹ Intrachem Bio Italia S.p.A., R&D Department, Via Calcinaro 2085/7, I Cesena, Italy; ² Agrichem Bio S.A., Plaza Castilla 3-14 A, E Madrid, Spain Abstract: In the two mediterranean countries, Italy and Spain, the efficacy of the A.quisqualis-based biofungicide AQ10 for the control of powdery mildews has been evaluated for several years in numerous field and greenhouse trials. As an example of the trials conducted on protected crops, in this paper we report the results of two GEP trials carried out in 2004: one in Italy on protected cucumber and one in Spain on greenhouse pepper. In Italy, on protected cucumber, A. quisqualis treatments significantly reduced powdery mildew incidence and severity compared to the untreated control. The biofungicide showed highest efficacy in reducing both powdery mildew incidence and severity, when applied in tank mixture with a pine oil adjuvant, followed by the biocontrol agent applied alone. Similar results were obtained in Spain on pepper: A. quisqualis treatments resulted in a significant reduction of powdery mildew severity compared to the untreated control. In both trials, the efficacy of the A. quisqualis-based treatments was comparable to or higher than that of the chemical reference treatments. Key words: biofungicide, mycoparasite, Sphaerotheca fuliginea, Leveillula taurica, vegetable crops Introduction Powdery mildew fungi (Erysiphaceae) are an extremely conspicuous group of plant pathogens, comprising more than 500 species that attack more than 1500 plant genera (Braun, 1987). Important protected crops grown in the mediterranean area, such as cucurbits, tomato, pepper, and strawberries, are among the major targets of powdery mildew fungi, and despite extensive research, powdery mildew control remains one of the most important plant protection problems worldwide (Hewitt, 1998; Kiss, 2003). Powdery mildews are usually controlled by repeated applications of fungicides, but environmental and public concerns towards the use of pesticides and the development of powdery mildew strains resistant to different fungicides (Schepers, 1983; Ishii et al., 2001; Gullino & Garibaldi, 2003; Pérez-García et al., 2005) have reduced the appeal of agrochemicals. In greenhouse productions, powdery mildew control is even more difficult due to the compatibility problems of sulphur-based products with benificial insects, especially adults of parasitic hymenopterans (Hassan et al., 1988). All these problems have led to the search of alternative methods to control powdery mildews, and to the development of the biofungicide AQ 10, containing conidia (at least 5.0 x 10 9 viable spores/g) of an isolate of the pycnidial fungus, Ampelomyces quisqualis Cesati. A. quisqualis is a well-known mycoparasite of powdery mildews. Its germinating spores suppress the development of powdery mildews through hyperparasitism (BCPC, 2004). Under adequate conditions (R.H. 60%), the hyphae of A. quisqualis penetrate the hyphae of their fungal host in less than 24 hours, continue their growth internally, and produce their pycnidia in the cells of the host s hyphae, conidiospores, and immature cleistothecia within 3 to 5 days (biotrophic interaction) (Kiss, 2003; BCPC, 2004; Zanzotto et al., 2005). The pycnidia can 275
288 sporulate and produce new spores of the antagonistic fungus. A. quisqualis suppresses the sporulation of the attacked powdery mildew mycelia, and kills all the parasitized cells, causing a gradual degeneration of their cytoplasm without the production of any toxins (necrotrophic interaction) (Kiss, 2003). The formulated product AQ 10 consists of the A. quisqualis isolate M-10, discovered in Israel in 1984, which has been formulated and commercialized by Ecogen Inc. under the trade name AQ 10. In Europe, the product, registered in Italy and Slovenia and under registration in Spain, is commercialized by Intrachem Bio International S.A. In 2005, the company obtained the inclusion of the active substance of AQ 10 into Annex I of the European Directive 91/414/EEC, which lists the active substances authorized for incorporation in plant protection products in Europe. In this paper we report the results of efficacy trials with AQ 10 against powdery mildew conducted in 2004 on pepper in Spain and on cucumber in Italy under the supervision of the Spanish and Italian national distribution companies of Intrachem Bio International S.A., Agrichem S.A. and Intrachem Bio Italia S.p.A., respectively. Material and methods A. quisqualis against cucumber powdery mildew (Sphaerotheca fuliginea) The GEP trial was conducted in 2004 on protected cucumber (Cucumis sativus L.) cv Adone by a certified Italian testing facility in Pontegradella, Ferrara, Italy. The efficacy in reducing powdery mildew incidence and severity on cucumber of the A. quisqualis-based biofungicide AQ 10 (a.s. A. quisqualis isolate M-10; WG, 58%), applied alone at 7 g/hl and in tank mixture with the adjuvant NuFilm 17 (a.s. di-1-p-menthene; EL, 904 g/l) at 30 ml/hl, was compared to that of an untreated control and of the chemical reference fungicide Flint 50 WG (a.s. trifloxystrobin; WG, 50%) at 15 g/hl. To compare the different treatments, a fully randomized block design was used with 4 replicates per treatment, and with 10 plants per plot. All treatments were applied 3 times: on August 24 (crop stage BBCH 51), August 31 (crop stage BBCH 53), and September 8 (crop stage BBCH 71) (Anonymous, 2001). From the moment the first disease symptoms appeared in the untreated control (August 31), the day of the last application (September 8), and 8 and 15 days after the last application September 16 and 23), the number of powdery mildew-infected leaves was counted on 100 randomly selected leaves per plot (percent disease incidence), and for each leaf, the percentage of infected leaf area was assessed. To assess the percentage of leaf area affected, the following scale was used: 0=no disease, 1=0-1, 2=1-5, 3=6-20, 4=21-40, and 5=>40% leaf area infected. Data on infected leaf area were transformed into percent infection severity using the formula of Townsend Heuberger: Severity = (Σ ni * vi) / N * V * 100, where ni is the number of infected leaves within each class vi, N the total number of infected leaves, and V the highest infection class. Furthermore, on each assessment date, the efficacy according to Abbott (Abbott, 1925) of the different treatments in reducing percent disease incidence and severity was calculated. For each assessment date, the percentages of powdery mildew incidence and severity were compared across treatments using one-way ANOVAs, followed by the Student- Newman-Keuls test for posthoc comparison of means. A. quisqualis against pepper powdery mildew (Leveillula taurica) The GEP trial was conducted in 2004 by Agrichem S.A.on protected pepper (Capsicum annuum L.) cv P66 in Campo de Cartagena, Murcia, Spain. The efficacy in reducing powdery 276
289 mildew infection severity on pepper of the A. quisqualis-based biofungicide AQ10 (a.s. A. quisqualis isolate M-10; WG, 58%), applied at the dose of 70 and 80 g/ha, respectively, was compared to that of an untreated control and of the chemical reference fungicide Trifmine 30 WP (a.s. triflumizole; WP, 30%) at 80g/hl. To compare the different treatments, a fully randomized block design was used with 4 replicates per treatment, and with 20 plants per plot. Treatments were applied at 7-day intervals (November 11, December 4, December 11), for a total of three applications per treatment. On the 3 successive application dates, and on 2 additional assessment dates (December 16 and 21, respectively 5 and 10 days after the last application), 100 leaves were selected randomly in each plot, and for each leaf, the percentage of infected leaf area and powdery mildew infection severity were assessed as described above for cucumber. For each assessment date, the percentages of powdery mildew infection severity were compared across treatments using one-way ANOVAs, followed by the LSD test for posthoc comparison of means, and the efficacy according to Abbott (Abbott, 1925) of the different treatments in reducing percent infection severity was calculated. Results and discussion A. quisqualis against cucumber powdery mildew (Sphaerotheca fuliginea) Except for powdery mildew infection severity on the first assessment date, differences among treatments in powdery mildew incidence and severity were always significant. On all 4 assessment dates, the A. quisqualis-based product in tank mixture with the pine oil adjuvant showed highest efficacy in reducing both disease incidence and severity on cucumber, followed by the biofungicide alone, and finally by the chemical trifloxystrobin-based reference treatment (Table 1). On the last assessment date, the mean efficacy of the biofungicide in tank mixture with the adjuvant in reducing disease incidence was 54.1%, and in reducing disease severity 86.0%. Spray adjuvants promote survival of biocontrol agents and ensure a homogeneous coverage of the plant surface, and have already been shown to improve the performance of this biofungicide (Philipp & Hellstern, 1986; Marchetti & D Aurelio, 1999). With the biocontrol agent alone, both powdery mildew incidence and severity were significantly reduced compared to the untreated control (efficacy 19.9 and 76.6%, respectively). In the plots treated with the chemical standard, instead, on the last two assessment dates, powdery mildew incidence did not differ significantly from that in the untreated control. Both A. quisqualis-based treatments thus resulted in an evident reduction in severity of powdery mildew on protected cucumber, higher than the approximately 50% reduction obtained by Jarvis and Slingsby (1977) and Sztejnberg et al. (1989), and lower than the up to 98% reduction observed by Elad et al. (1998). This variability in efficacy is probably due to the humidity conditions within the greenhouses, limiting the growth of the mycorparasite and thus affecting its efficacy (Dik et al., 1998; Shishkoff & McGrath, 2002; Kiss et al., 2004). A. quisqualis against pepper powdery mildew (Leveillula taurica) Prior to the application of the first treatment, powdery mildew severity was comparable among treatments (no significant differences): it ranged from 1.3% in the A. quisqualistreated plots to 0.3% in the chemical reference plots, and 0.6% in the untreated control plots. On all 4 successive assessment dates, significant differences among treatments emerged. Powdery mildew infection severity was always significantly higher in the untreated control than in all the other treatments. With regard to the treated plots, infection severity was always highest in those treated with the chemical standard, intermediate in those treated with AQ10 at 277
290 70 g/ha, and lowest in those treated with the biofungicide at 80 g/ha, with differences among treated plots being significant on December 11 and 16, and failing significance on December 4 and 21 (Table 2). Table 1. Percentages of powdery mildew incidence and severity (m±s.d.) on cucmber in the different treatments on the four successive assessment dates, and mean efficacy of the treatments in reducing disease incidence and severity. No. Treatment % incidence (% efficacy) % severity (% efficacy) 1 st assessment (August 31, 2004) * 1 AQ ±0.8 (40.0) c 8.5±1.0 (8.1) a 2 AQ 10+NuFilm 4.0±0.8 (60.0) d 4.5±1.0 (51.4) a 3 Flint 50 WG 7.5±1.0 (25.0) b 7.3±2.6 (21.6) a 4 Untreated control 10.0±0.8 (-)a 9.3±5.4 (-) a 2 nd assessment (September 8, 2004) * 1 AQ ±5.9 (27.3) c 9.0±1.2 (52.0) b 2 AQ 10+NuFilm 43.5±6.7 (50.6) d 4.5±1.0 (76.0) b 3 Flint 50 WG 77.8±12.3 (11.7) b 9.5±4.2 (49.3) b 4 Untreated control 88.0±6.9 (-) a 18.8±4.8 (-)a 3 rd assessment (16 September 2004) * 1 AQ ±7.4 (26.1) bc 10.0±1.6 (48.7) bc 2 AQ 10+NuFilm 45.0±7.1 (49.0) c 6.0±2.5 (69.2) c 3 Flint 50 WG 78.8±13.2 (10.8) ab 12.5±4.1 (35.9) b 4 Untreated control 88.3±3.9 (-) a 19.5±4.2 (-) a 4 th assessment (23 September 2004) * 1 AQ ±8.9 (19.9) b 13.8±2.5 (76.6) b 2 AQ 10+NuFilm 45.0±7.1 (54.1) c 8.3±2.4 (86.0) b 3 Flint 50 WG 92.0±13.5 (6.1) a 16.3±6.3 (72.3) b 4 Untreated control 98.0±2.5 (-)a 58.8±10.3 (-) a * Different letters within the same column indicate statistically significant differences (Student-Newman-Keuls test: P<0.05). The efficacy of the A. quisqualis-based product applied at 80 g/ha ranged over time from 78.8 to 95.8%. Comparable, but slightly lower values (68.8 to 90.2%) were recorded for the product used at 70 g/ha, and for the chemical triflumizole-based standard ( %). To our knowledge, the number of previous studies on the biocontrol potential of Ampelomyces isolates on pepper is limited, but the obtained results were positive (Sztejnberg, 1997; Diop-Bruckler & Molot 1987; Olsen et al., 2001; Kiss et al., 2004). Given the highly promising results recorded in this study and the absence of negative side-effects (phytotoxicity) on the target crop, further studies on the use of the A. quisqualis isolate M-10 for powdery mildew control on pepper are definitely warranted. In conclusion, the applications of the A. quisqualis-based biofungicide in common plant protection strategies can be advantageous for several reasons: in organic farming, they constitute one of the few available alternatives to applications of sulphur-based products, while in integrated farming, they may be useful for the development of adequate resistance management strategies. Furthermore, the biocontrol agent has no pre-harvest interval, and therefore allows to prevent the presence of undesired residues in the final produce. Finally, contrary to sulphur- 278
291 based and some other fungicides, the product does not cause any phytotoxic effects (cucurbits are especially susceptible to phytotoxic effects), and is harmless to beneficials. Table 2. Percentages of powdery mildew infection severity (m±s.d.) on pepper in the different treatments on the last 4 successive assessment dates, and mean efficacy of the treatments in reducing disease severity. No. Treatment (dose) % severity (m±s.d.) % efficacy (mean) December 4, 2004 (2 nd application date) * 1 AQ 10 (70 g/ha) 2.8±1.2 b AQ 10 (80 g/ha) 1.9±0.7 b Trifmine 30 WP (80 g/hl) 3.8±1.4 b Untreated control 10.0±2.7 a - December 11, 2004 (3 rd application date) * 1 AQ 10 (70 g/ha) 2.8±1.2 c AQ 10 (80 g/ha) 1.9±1.6 bc Trifmine 30 WP (80 g/hl) 5.3±1.6 b Untreated control 31.9±3.8 a - December 16, 2004 (5 days after last application) * 1 AQ 10 (70 g/ha) 4.4±1.6 b AQ 10 (80 g/ha) 1.9±1.3 c Trifmine 30 WP (80 g/hl) 5.9±2.1 b Untreated control 45.0±2.7 a - December 21, 2004 (10 days after last application) * 1 AQ 10 (70 g/ha) 6.6±1.6 b AQ 10 (80 g/ha) 5.6±4.1 b Trifmine 30 WP (80 g/hl) 7.5±1.4 b Untreated control 62.5±4.2 a - * Different letters indicate statistically significant differences (LSD test: P<0.05). References Abbott, W.S. 1925: A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18: Anonymous, 2001: Growth stages of mono- and dicotyledonous plants, BBCH Monograph. 2 nd edition, ed. U. Meier, pp. BCPC, 2004: Ampelomyces quisqualis biological fungicide (fungus). In: The Manual of Biocontrol Agents, 3 rd edition, ed. L.G. Copping: 6-8. Braun, U. 1987: A monograph of the Erysiphales (powdery mildews). Nova Hedwigia Beihefte, Beiheft 89: Dik, A.J., Verhaar, M.A., Bélanger R.R. 1998: Comparison of three biological control agents against cucumber powdery mildew (Sphaerotheca fuliginea) in semi-commercial-scale glasshouse. Eur. J. Plant Pathol. 104: Diop-Bruckler, M. & Molot, P.M. 1987: Interét de quelques hyperparasites dans la lutte contre Léveillula taurica et Sphaerotheca fuliginea. OEPP Bull. 17:
292 Elad, Y., Kirshner, B. Yehuda, N. & Sztejnberg, A. 1998: Management of powdery mildew and gray mould of cucumber by trichoderma harzianum T39 and Ampelomyces quisqualis AQ10. Biocontrol 43: Gullino, M.L. & Garibaldi, A. 2003: La resistenza ai fungicidi in viticoltura: un aggiornamento sulla situazione italiana. Informatore Fitopatologico 4: Hassan, S.A., Bigler, F., Bogenschütz, H., Boller, E., Brun, J., Chilverton, P., Edwards, P., Mansour, F., Samsoe-Petersen, L., Staübli, A., Sterk, G., Tavares, K., Tuset, J.J., Viggiani, G. & Vivas, G. 1988: Results of the fourth joint pesticide testing programme carried out by the IOBC/WPRS-Working Group Pesticides and Beneficial Organisms. J. appl. Ent. 105: Hewitt, H.G. 1998: Fungicides in crop protection. CABI Publishing, Wallingford, UK, 232 pp. Ishii, H., Fraaije, B.A., Sugiyama, T., Noguchi, K., Nishimura, K., Takeda, T., Amano, T. & Hollomon, D.W. 2001: Occurrence and molecular characterization of strobilurin resistance in cucumber powdery mildew and downy mildew. Phytopathology 91: Jarvis, W.R. & Slingsby, K. 1977: The control of powdery mildew of greenhouse cucumber by water sprays and Ampelomyces quisqualis. Plant Dis. Rep. 61: Kiss, L. 2003: A review of fungal antagonists of powdery mildews and their potential as biocontrol agents. Pest Manag. Sci. 59: Kiss, L., Russell, J.C., Szentiványi O., Xu, X. & Jeffries, P. 2004:m Biology and biocontrol potential of Ampelomyces mycoparasites, natural antagonists of powdery mildew fungi. Biocontrol Sci. Technol. 14 (7): Marchetti, L. & D Aurelio, A. Z. 1999: Natural products against powdery mildew of lagerstroemia indica. Inform. Agrario 55: 99. Olsen, M.W., Oehler, J. & Rorabaugh, P. 2001: Evaluation of fungicides for control of powdery mildew of greenhouse pepper. Arizona College of Agriculture 2001 Vegeable Report, index at: Pérez-García, A., de Vincente, A., Fernández-Ortuño, D., López-Ruíz, F.J. & Torés, J.A. 2005: Resistencia a inhibidores de la biosíntesis de ergosterol y a estrobilurinas en oídio de cucurbitáceas. Phytoma 173: Philipp, W.D. & Hellstern, A. 1986: Biologische Mehltaubekämpfung mit Ampelomyces quisqualis bei reduzierter Luftfeuchtigkeit. Z. Pflanzenkr. Pflanzenschutz 93: Schepers, H.T.A.M. 1983: Decreased sensitivity of Sphaerotheca fuliginea to fungicides which inhibit ergosterol biosynthesis. Neth. J. Plant Pathol. 89: Shishkoff, N. & McGrath, M.T. 2002: AQ10 biofungicide combined with chemical fungicides or AddQ spray adjuvant for control of cucurbit powdery mildew in detached leaf culture. Plant Dis. 86 (8): Sztejnberg, A. 1997: Biological control of powdery mildew diseases. Phytoparasitica 25 (4): Sztejnberg, A., Galper, S., Mazar, S. & Lisker, N. 1989: Ampelomyces quisqualis for biological control of powdery mildews in Israel. J. Phytopathol. 124: Zanzotto, A., Bellotto, D. & Borgo, M. 2005: Efficacia antioidica di AQ 10 ed effetti sulla formazione di cleistoteci in vigneto. Informatore Fitopatologico 7-8:
293 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Quantification of powdery mildew removal by the mycophagous Beetle Psyllobora vigintimaculata (Coleoptera: Coccinellidae) Andrew M. Sutherland 1, Michael P. Parrella 2 1 Graduate Student Researcher, Entomology Graduate Program. 2 Professor of Entomology, Department of Entomology, University of California-Davis, USA Abstract: The coccinellid tribe Psylloborini (Halyziini) is entirely comprised of obligate consumers of powdery mildew (PM) fungi (Ascomycota: Erysiphales). The small ashy gray ladybird beetle, Psyllobora vigintimaculata, a North American species, is being evaluated for use as a biological control agent against PM in greenhouse systems. Individual larvae were reared on PM-infected leaf discs in a laboratory incubator to quantify the removal of PM due to this insect s feeding. The leaf area cleaned and the background growth of PM were measured with image analysis software using periodic digital images. Leaf discs exposed to neonate larvae for 192 hours showed a significant decrease in PM growth as compared to an untreated control, and leaf discs exposed to 3 rd instar larvae for 96 hours showed a significant decrease in infected leaf area. A simple model based on these data predicts that an individual larva cleans 6.3±3.3cm 2 leaf area of all visible PM hyphae and conidia from the time of egg eclosion until successful pupation. Key words: powdery mildew, Psyllobora vigintimaculata, biological control, functional response Introduction The fungi belonging to the Erysiphales (Ascomycota), commonly known as powdery mildews (PM), are all obligate biotrophs. As an order, they have been recorded to infect close to 10,000 species of angiosperm plants in 169 families (Amano, 1986). Since many of these host plants are valued as agricultural commodities, PM are collectively considered one of the most important plant pathogens worldwide. Chemically based disease management programs have historically led to resistance by the fungi to several key fungicides (Gubler et al, 1996; McGrath, 2001). Biological control of PM may offer solutions to this growing resistance problem in addition to issues such as residues in food crops, worker health and safety, and negative effects to nontarget organisms. There are several commercially available microbial biological control agents including the spore-forming bacterium Bacillus subtilis and the fungal hyperparasite Ampelomyces quisqualis Ces. Very little is known of the potential for arthropod agents to control or reduce the disease through consumption. All members of the beetle tribe Psylloborini Casey (Coleoptera:Coccinellidae) are obligate consumers of various PM conidia and hyphae in all mobile life stages (Gordon, 1985). The cosmopolitan genus Psyllobora Chevrolat is represented in natural and managed systems in temperate and subtropical regions worldwide, and may be utilized as a native biological control agent of PM (Cruz et al., 1989; Almeida & Milleo, 1998). Soylu et al. (2002) recorded a reduction in PM conidia of 92% when comparing leaf areas grazed upon by P. bisoctonotata Mulsant with non-fed-upon areas, suggesting a real and measurable PM removal through consumption. The small ashy gray ladybird, P. vigintimaculata Say, is native to northern and western North America. It has recently been recorded as locally common in Davis, California, feeding on various PM associated with horticultural and agronomic crops in the landscape as well as in protected greenhouse culture (Sutherland, 2005a). The overall 281
294 objectives of our work are the evaluation and development of P. vigintimaculata for biological control of PM, and the assessment of its compatibility with chemically managed systems such as greenhouse horticulture (Sutherland, 2005b). The success of any biocontrol agent depends directly on its ability to locate the pest, reproduce in response and in relation to the pest, and to function or consume based on the relative abundance or density of the pest (Solomon, 1949). We are interested in evaluating these responses for P. vigintimaculata. The prey item of this insect, spores and hyphae of PM fungi, sometimes exist in huge dense colonies of millions of spores and sometimes as minuscule patches in isolated areas on specific plants. Previous work has shown that adults of this species can be found associated with low density, isolated PM patches (Sutherland, 2005a), suggesting their ability to locate food by responding to long-range stimuli during flight. It is established that large coccinellids such as Coccinella septempunctata L. respond to olfactory cues from their aphid prey, and can even distinguish between aphid species based on volatiles (Sengonca & Liu, 1994). Additionally, landscape surveys have demonstrated strong positive correlations between presence and density of PM in the landscape and the presence and density of P.vigintimaculata life stages (Sutherland, 2005a), suggesting an increase in reproduction and/or aggregation by the insect in response to increases in its food item. Work presented here addresses aspects of the functional response of P. vigintimaculata. Specifically, we quantify the average amount of leaf area visibly cleaned of hyphae and conidia by a P. vigintimaculata individual during its entire larval development. Material and methods Organism rearing Plant material (Zinnia elegans) with PM was grown under high-pressure sodium lighting (600W) in a humidified (50-80% relative humidity) growth room utilizing ebb-and-flood hydroponics. Periodic inoculations with conidia from the PM fungus Erysiphe chicoracearum were made either by spore solution or by brushing from infected plants. A colony of P. vigintimaculata was maintained in the laboratory in a series of insect rearing cages. Sufficiently infected Zinnia plants were exposed to caged adults at regular intervals for egg deposition. Afterwards, the egg-laden plants were retained in the cage to facilitate larval development. Towards the end of the fourth larval instar the plants were moved to a final pupation cage and cut at media level. Adults were captured as they emerged and flew towards the light at the top of the cage, as in a reverse Berlese funnel. In this manner the colony provided harvestable eggs, larvae or adults of uniform age at five day intervals. Experimental units Egg masses deposited on the same day were removed from the insect colony and transferred to an incubator (Percival Scientific I-30 BL) kept at 20±5 o C and 50-90% relative humidity under fluorescent lights. Excised PM-infected Zinnia leaves from the hydroponic PM colony were cut with a scalpel to conform to a circular template (55mm diameter). These leaf discs were then placed on filter paper on top of ~10mm agar (USP, 150 mesh, Bio-serv ) in a vented petri dish enclosure (55mm X 30mm). Upon egg eclosion, the 1st instar larvae were individually transferred with a fine paintbrush to one half of the dish enclosures. The other half of the dishes received no insects and represented untreated controls, subject to normal fungal growth and development. All dishes were then assigned a reference number and returned to the incubator in a randomized fashion (random.org). This methodology maintained turgidity of leaf discs and allowed for development of insects and fungi for up to ten days. Larvae were removed and returned to the colony after they had completed two larval instars (approximatelt eight days). A similar set of experiments was established for newly molted 282
295 third instar larvae. Fresh PM-infected leaf discs were created as above and introduced into the dish enclosures and incubator. These larvae were allowed to develop until pupation, at which time the pupae were returned to the colony and observation was terminated. Together, the two cohorts of P. vigintimaculata larvae used in this feeding quantification trial represented one complete larval development period, from egg eclosion until sucessful pupation. Image analysis Software based on existing image analysis algorithms (Assess Image Analysis Software for Plant Disease Quantification, The American Phytopathological Society, 2002), coupled with digital photography, was used to quantify the PM present on Zinnia leaf discs throughout the study. A digital camera (Nikon D70, Japan) equipped with a macrophotograpy lens (Sigma 150mm 1:2.8 APO MACRO DG HSM D, Japan) mounted on an adjustable tripod (bogen, Manfrotto, Italy) was used to capture digital images of each leaf disc just prior to insect release and every 48 hours thereafter. The described software was used to separate PM from the background leaf disc based on pixel saturation (Lamari, 2005), and then to express disease as the percentage of leaf area covered by visibly discernable PM colonies (%PM). This derived percentage was used as the response variable at all times. Statistical analysis Experimental units were arranged as in a completely randomized design (CRD) within the incubator, and were re-randomized after each image acquisition event. Differences in %PM between units containing larvae and those left untreated were determined through use of the one-way analysis of variance (ANOVA) (JMP Start Statistics, SAS Institute, 2005). Data were collected to reflect the increase in leaf area affected ( %PM) and the percentage increase in relation to initial severity ( %PM/%PM initial ). These variables were also analyzed using ANOVA to detect treatment differences. All percentage-based variables could be expressed as leaf area (cm 2 ) by multipling by the area of an entire leaf disc (23.7cm 2 ). Model construction In order to estimate the total amount of leaf area cleaned (LAC) of PM by one P. vigintimaculata individual during larval development we constructed a basic model that included a measure of the normal PM growth (G) that should occur in the absence of feeding. Additionally, we treated the data resulting from the young larva (egg-2nd instar) and old larva (3rd instar-pupa) trials as continuous, with the sum representing the estimated response for the entire larval duration. Therefore, the constructed model is as follows: LAC total = LAC young + LAC old + G young + G old, where LAC = (%PM initial - %PM final ) * 23.7cm 2 for larva units, and G = mean(%pm final - %PM initial ) * 23.7cm 2 ± standard error for all untreated units. Results and discussion 1 st and 2 nd instar trial There was no significant initial treatment difference in %PM (df=1,18; F=0.18; p=0.68). Both treatments exhibited an increase in %PM until 96 hours after larval introduction, when %PM began to stabilize and decline (Figure 1). After 192 hours of observation %PM was significantly lower on leaf discs containing larvae as compared to untreated leaf discs (df=1,18; F=6.48; p=0.02). When larvae were removed after 192 hours the mean increase in infected leaf area in untreated units (4.38±0.95cm 2 ) was significantly higher (df=1,18; F=6.74; p=0.02) than that in larva units (1.17±0.8cm 2 ). Additionally, PM-affected areas 283
296 increased an average 54.3% relative to initial size in untreated units but only 19.5% relative to initial size in larva units n = untreated with one larva Time (hours) Figure 1. Percentage leaf area affected by powdery mildew on leaf discs (23.7cm 2 ) over time in the presence of neonate-2 nd instar larvae of Psyllobora vigintimaculata as compared to untreated. 3 rd and 4 th instar trial There was no significant treatment difference in %PM at the onset of observation (df=1,18; F=0.33; p=0.57) or 48 hours following release, but the mean %PM on leaf discs containing larvae began to decrease immediately (Figure 2). After 96 hours there was a significant difference in %PM between treatments (df=1,18; F=7.79; p=0.01), with a PM decrease in larva units as compared to a PM increase in untreated units (Figure 3). Area infected by PM increased an average 14.9% relative to initial size in untreated units but actually decreased 12.3% relative to initial size with larvae present; a significant difference (df=1,18; F=12.38; p=0.002). Consumption model Each larva cleaned an average of 3.22±1.8cm 2 of Zinnia leaf area of all visible traces of the PM from the time of egg eclosion until molt initiation at the end of the 2 nd instar during a period of 192 hours. PM in larva units actually increased during this time by 4.9% to give a calculated LAC young of 1.17cm 2 /unit, but this was offset by a PM background growth, G young, of 4.38 cm 2 /unit. In the second trial each larva cleaned an average of 3.10±1.5cm 2 leaf area of PM from the start of the 3 rd instar until pupation during a period of 96 hours. The LAC old was calculated as 1.69cm 2 /unit and the G old as 1.41cm 2 /unit. The background PM growth rate was similar in both trials, at and 0.015cm 2 /hour, respectively. The model predicts that an average P. vigintimaculata larva will clean 6.32±3.3cm 2 of Zinnia leaf area of PM spores and hyphae during its entire larval development 284
297 60 n = 20 untreated with one larva Time (hours) Figure 2. Percentage leaf area affected by powdery mildew on leaf discs (23.7cm 2 ) over time in the presence of 3 rd - 4 th instar larvae of Psyllobora vigintimaculata as compared to untreated. Change in PM-Affected Leaf Area (cm2) untreated n = 20 with one larva Figure 3. Overall change in area affected by powdery mildew on leaf discs (23.7cm 2 ) exposed to 3 rd -4 th instar Psyllobora vigintimaculata larvae for 96 hours as compared to untreated.. Based on this figure, it would take four larvae developing from egg to pupa to consume the PM covering an entire Zinnia leaf disc 55mm in diameter (23.7cm 2 ) if the disc were 100% covered with spores and hyphae. 285
298 This quantification of the PM consumption abilities of P. vigintimaculata is essential to its evaluation for use as an effective biological control agent against PM fungi. Members of this genus are known to feed on many genera of PM on many different host plants (Ahmad et al, 2003), and different PM species grow at different densities, rates and patterns, so it is important to realize that varying levels of effectiveness may be seen in different systems and with other Psyllobora species. References Ahmad, M.; Younis, G & Ali, N. 2003: Biology of the coccinellid Psyllobora (Thea) bisoctonotata Muls. (Coleoptera: Coccinellidae) a predator of powdery mildew fungi. Proceedings of the Eighth Arab Congress of Plant Protection, Elbeida City, Libya. Almeida, L.M. & Milleo, J. 1998: The immature stages of Psyllobora gratiosa Mader, 1958 (Coleoptera: Coccinellidae) with some biological aspects. J. New York Entomol. Soc. 106(4): Amano, K. 1986: Host range and geographical distribution of the powdery mildew fungi. Japan Scientific Societies Press, Tokyo. Cruz, B.; Gonzalez-Avila, M. & Soto, L. 1989: Psyllobora nana (Coleoptera: Coccinellidae), biological control agent of dusty mildew. Ciencias de la Agricultura 36: 151. Gordon, R. D. 1985: The Coccinellidae (Coleoptera) of America north of Mexico. J. New York Entomol. Soc. 93: Gubler, W.D.; Ypema, H.L.; Ouimette, D.G & Bettiga, L.J. 1996: Occurrence and development of resistance in Uncinula necator to triadimefon, myclobutanil and fenarimol in California grapevines. Plant Dis. 80: Lamari, L. 2005: ASSESS: Image analysis software for plant disease quantification. APS Press, St. Paul, MN, USA. McGrath, M.T. 2001: Fungicide resistance in cucurbit powdery mildew: Experiences and challenges. Plant Dis. 85(3), SAS Institute Inc. 2005: JMP Start Statistics third edition. A guide to statistics and data analysis using JMP and JMP IN software. Cary, NC: SAS Institute Inc. Sengonca, C. & Liu, B. 1994: Responses of the different instar predator, Coccinella septempunctata L. (Coleoptera: Coccinellidae), to the kairomone produced by the prey and non-prey insects as well as the predator itself. Z. Pflkrankh. Pflschutz. 101: Solomon, M.E. 1949: The natural control of animal populations. J. Anim. Ecol. 18: Soylu, S.; Yigit, A. & Shtienberg, D. 2002: Feeding of mycophagous ladybird, Psyllobora bisoctonotata, on powdery mildew infested plants. IOBC Proceedings of the 7 th Working Group Meeting, Kusadasi, Turkey 25, Sutherland, A.M. 2005a: Evaluation of Psyllobora vigintimaculata (Say) (Coleoptera: Coccinellidae) for Biological Control of Powdery Mildew Fungi (Erysiphales). M.S. thesis, University of California, Davis. Sutherland, A.M. 2005b: Effects of selected fungicides on a mycophagous ladybird (Coleoptera: Coccinellidae): Ramifications for biological control of powdery mildew. IOBC/WPRS Bull. 28(1):
299 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp UV interfering nets reduce TYLCD incidence and progress in tomato crops: influence of host genotype Leonardo Velasco 1, Dirk Janssen 3, Marisol Catalá 2, Joaquín Costa 2 1 Departamento de Biotecnología y Protección Vegetal. 2 Departamento de Horticultura. Instituto Murciano de Investigación y Desarrollo Agrario y alimentario (IMIDA). c/ Mayor s/n. E La Alberca Murcia, 3 CIFA Almería (IFAPA, CICE, Junta de Andalucía), Autovía del Mediterráneo, Km 420. E La Mojonera Almería, Spain. leonardo.velasco@carm.es Abstract: Four tomato cultivars, tolerant and non-tolerant to tomato yellow leaf curl disease, were grown under UV-blocking and non-uv-blocking nets and TYLCD incidence and progression were compared. Squash-blot followed by molecular hybridization was used to monitor disease incidence and severity. Severity was recorded as hybridization signal intensity. Results of AUDPC analysis showed that there were significant differences in disease incidence and severity between the two nethouses. Disease intensity was not enough to explain the results, where severity was a more explanatory parameter for quantifying disease incidence. The four cultivars showed lower mean AUPDC incidence and severity in the UV-blocking nethouse. Differences in TYLCD between tolerant or non-tolerant cultivars were only significant when severity was considered. There were also marked differences among cultivars: the tolerant cultivar Boludo showed no significant differences in incidence and severity when compared with the non-tolerant cultivar Izabella in the UV-blocking nethouse. Finally, all of the cultivars, including the tolerant ones, proved good sources of virus inoculum, which made them unsuitable for virus control. Keywords: Tomato yellow leaf curl virus, whitefly, greenhouse, nethouse, photoselective, UV nets, AUDPC, disease progress, disease incidence. Introduction Control of insect-borne diseases is an important task in Mediterranean horticulture. In southeastern Spain tomatoes are grown throughout the year, with marked differences depending upon the region. For example, in Almeria almost all crops are produced under greenhouse conditions, while in Murcia tomatoes are still grown in open air conditions. Crops are often grown under nets of different mesh sizes. An important economic limiting factor for tomato produced in these regions is insect-borne viruses, particularly Tomato yellow leaf curl virus (TYLCV) and Tomato yellow leaf curl Sardinian virus (TYLCSV) are responsible for tomato yellow leaf curl disease (TYLCD) in Spain (Moriones & Navas-Castillo, 2000). Both viruses are transmitted by the whitefly Bemisia tabaci, for which insecticidal control is increasingly ineffective in spite of the high doses applied. In addition, there is growing public health concern relating to the use of chemicals in agriculture where integrated pest management is becoming an alternative. The use of UV interfering screens or nets is a novel part of an IPM strategy to prevent insect-borne diseases. In previous reports, the use of UV-blocking screens interfering with insect vision and behaviour, and consequently with virus transmission, was proposed as a strategy to control both virus diseases and insect populations (Monci et al., 2004). As these screens have been increasingly introduced in south-eastern Spain, the present study evaluates 287
300 the use of UV-blocking nets, considering the influence of host genetic background with four representative tomato cultivars that are widely grown in the area. Material and Methods Tomato cultivars Four commercial cultivars were chosen in order to test TYLCD in different genetic backgrounds: Boludo (Petoseed) and Tyrade (S&G) are advertised as TYLCD tolerant whereas Izabella (Zeraim) and Durinta (Western seeds) are not. Crop cultivation and sampling procedure Two nethouses covered with 10 x 20 threads/cm 2 (50 mesh) nets were designed. Nets for both structures were provided by the same manufacturer (Klayman Meteor Ltd., Israel). The UVblocking net was BioNet TM, while the non UV-blocking net was Antivirus TM. Both greenhouses were organized in three continuous blocks, each with three rows of 24 plants of each of the cultivars. These were planted adjacent to one another on July 2 nd, 2003, and there were a total of 860 plants per greenhouse. In each greenhouse about one third of each cultivar (approximately 72 plants) was screened by hybridization. Eight of the 24 plants in each row were chosen for sampling and monitored throughout cultivation. Sampling was carried out every fifteen days over the course of four month trial period. The first sample was taken 3 days after the plantlets were transplanted (dpt) from the nursery. TYLCD detection TYLCD DNA probes were synthesized by PCR from plasmid constructions including the CP region of TYLCV and TYLCSV (Accotto et al. 2000). PCR for probe synthesis was carried out with 0.4 µm of TY1(+) and TY2(-) primer pair and 200 µm DIG-labelled dntp mix (Roche Molecular Biochemicals, Mannheim, Germany) in a final volume of 100 µl. Growing buds were squashed on Hybond N + (Roche) membranes and nucleic acids were UV crosslinked at 0.12 kj/m 2. Hybridization was performed at 55 ºC according to manufacturer s instructions with 25 ng/ml of the DIG-PCR probe in the DIG Easy hybridization solution (Roche). Washing steps and detection of the CDP-Star (Tropix Inc., Bedford, MA, USA) signal were performed as recommended by the manufacturer. Two parameters were determined: the percentage of plants that were positive after TYLCV/TYLCSV probe hybridization, and the severity of the disease, extrapolated from the intensity of the hybridization signal. This parameter was established as a three point scale: (1) = weak signal, (2) = positive signal and (3) = strong positive signal. Data analysis Data relating to disease incidence in both nethouses were used to determine the area under the disease progress curve (AUDPC inc ) using the midpoint rule method (Campbell & Madden, 1990). The AUDPC inc values were calculated using the formula: Σ n-1 i=1 [(t i+1 t i )(inc i + inc i+1 )/2], where t i is time in days since transplanting, inc i is the proportion of diseased plants at each record, and n is the number of records. In addition, disease severity was calculated as the scoring of severity (sev i ) for each cultivar and nethouse. Similarly, the AUDPC sev was calculated as Σ n-1 i=1 [(t i+1 t i )(sev i + sev i+1 )/2]. ANOVA of disease incidence (AUDPC inc ) and severity (AUDPC sev ) were performed for each nethouse and tomato cultivar. To construct disease progress curves, average disease incidence for each cultivar and nethouse were plotted for each sampling time. TableCurve2D v.5.01 (Systat Sofware, Inc.) was used to fit the following linear growth models: monomolecular, exponential, logistic, and Gompertz, in order to determine which model would best represent the relationship between disease incidence and time. For each regression model, coefficient of determination (R 2 ), mean square error (MSE) values, and plots of standardized residuals versus the predicted 288
301 values, were used to select the best model to describe disease progress curves for each greenhouse. Predicted disease values were back-transformed to the original scale and regressed on the observed disease values. Recalculated R 2 and MSE values (designated R* 2 and MSE*) were obtained following Campbell & Madden (1990) and compared with previous values. Finally, visual inspection of the plot of the residuals made it possible to discard unacceptable patterns. Results and Discussion The use of double insect exclusion systems based on both the use of physical barriers, such as high density nets, and of optical interfering screens is documented in the literature. According to Antignus et al. (1998), screens with low density mesh did not exhibit differences in protection with respect to UV-blocking and non UV-blocking screens. We therefore installed 50 mesh nets in the nethouses for the present study. Symptom recording is sometimes a quite subjective matter. In particular, TYLCD is not always recognized, especially when dealing with tolerant plant material. We consequently decided to use molecular hybridization for virus detection as it represented a more objective method for quantifying the incidence of disease in each nethouse. On the other hand, squash-blot followed by molecular hybridization is consi A B Incidence (%) Severity C D Incidence (%) Severity Days post transplant Figure 1. Disease incidence (A, C) and severity (B, D) progress of the four cultivars in the two nethouses: Antivirus (A, B); Bionet (C, D). ( ) Durinta, ( ) Boludo, ( ) Izabella and ( ) Tyrade. Each point represents the mean of three replications. 289
302 dered as the best method for assessing TYLCD diagnostics in breeding trials (Pico et al., 1999). A pre-sampling analysis, performed on the same day that the plantlets were transplanted from the nursery, resulted in 5% of tested plants registering as TYLCD positive. None of the cultivars in the two nethouses reached a 100% disease incidence at 115 dpt, although the non-tolerant cultivars registered higher incidence values than the tolerant ones (Figure 1A, C). Severity was much more intense for the non-tolerant cultivars under the Antivirus net than for the tolerant varieties (Figure 1B), although under the Bionet they showed a similar increase in severity (Figure 1D). AUDPC is a valuable parameter for assessing incidence and severity because it allows statistical analyses. Figures 2A and 2B show the mean AUDPC values for incidence and severity, respectively, for the four cultivars and for both nethouses. ANOVA results are displayed in Table 1. The difference between the two nethouses was significant (P=0.003) considering all of the different cultivars. When each variety was considered separately, differences in incidence were significant during the trial period, except for Tyrade. At the same time, differences in severity were all significant. When cultivars were grouped according to TYLCD tolerance, there were significant differences in incidence between the two nethouses, with P=0.029 and P=0.003 for tolerant and non-tolerant cultivars, respectively. These differences were more accused when severity was considered, with P= and P= for tolerant and non-tolerant cultivars, respectively. There were marked differences between tolerant and non-tolerant cultivars in each nethouse. While differences in incidence severities within each nethouse were significant, differences in disease incidences were not (P=0.655 and P=0.281). In addition, we were able to establish differences between cultivars. When comparing TYLCD incidence in Izabella and Tyrade, they were only significantly different for the Antivirus (P=0.022) but not in the Bionet nethouse. Severity was not significantly different in the two nethouses (P= and P=0.0005). When incidence was compared between Durinta and Tyrade, no significant differences were found in either nethouse, while severity was significantly different (P= and P=0.0013) A B AUDPC inc AUDPC sev Durinta Boludo Izabella Tyrade 0 Durinta Boludo Izabella Tyrade Figure 2. Means of AUDPC inc (A) and AUDPC sev (B) values per genotype for the two nethouses. Antivirus: plain white; dashed: Bionet. Bars represent the standard deviation of the mean. 290
303 When the incidences of Izabella and Boludo disease were compared, no significant differences were found in either of the nethouse. Differences in disease severities were significant for the Antivirus net (P=0.001) but not for the Bionet (P=0.139). When the incidence of disease in Durinta and Boludo were compared, no significant differences were found in either of the nethouse, although severity was significantly different between the two (P=0.031). In conclusion, the four cultivars behaved quite dissimilar. Although Durinta is a non-tolerant cultivar, it showed less disease incidence and severity than Izabella. Boludo, a tolerant variety, showed a higher incidence and severity than Tyrade. In fact, it was not possible to distinguish the incidence of disease for Izabella and Boludo and severity was not significantly different under Bionet. Analysis of growth models showed that the monomolecular curve best described TYLCD progress in tolerant cultivars in both nethouses, while disease progress in Durinta was exponential during the trial period (results not shown due to text length limitations). All the cultivars except Durinta reached a plateau before reaching 100% incidence, whereon they started to decline: this is typical of plant recovery after infection in adult stages. Table 1. Summary of ANOVA comparing incidence and severity as AUDPC inc and AUDPC sev for four tomato cultivars grown under Bionet or Antivirus nets. Source of variation AUDPC inc AUDPC sev F statistic P>F F statistic P>F Nethouse All cultivars Durinta Boludo Izabella Tyrade Tolerant Non-tolerant Tolerant vs. non-tolerant Bionet Antivirus Izabella vs. Tyrade Bionet Antivirus Durinta vs. Tyrade Bionet Antivirus Izabella vs. Boludo Bionet Antivirus Durinta vs. Boludo Bionet Antivirus The influence of UV filtering screens on insects seems to be due to the different attraction into greenhouses. Insects show a preference for entering non UV-blocking 291
304 greenhouses covered with polyethylene films, but these do not apparently interfere with normal flight or behaviour (Costa & Robb, 1999). However, other experiments suggest that UV-blocking screens not only affect the attraction of insects into greenhouses but also their mobility inside them, since most whitefly and diseases appear close to the UV-blocking tunnel entrance, while in non UV-blocking greenhouses they are dispersed throughout the tunnels. (Antignus et al., 2001). We also observed this effect (data not shown), suggesting that there is indeed an effect upon insect flight behaviour. Discrepancies between the results reported by Costa & Robb and those from Antignus and collaborators could be due to differences in the spectral absorption ranges of the UV-blocking plastics used and to the experimental design of the tunnels in the two sets of experiments: smaller internal tunnel volumes could overcome some insect flight conducts (Antignus et al., 2001). In conclusion, high mesh UV nets provide better protection against TYLCD in tomato than non UV nets. In addition, we found that it is very important to monitor disease severity, because incidence data alone do not provide enough information for comparisons. Furthermore, the genetic background of cultivars is critical when comparing incidence and severity. Finally, we found that even tolerant cultivars were a good source of viral inoculums in both greenhouses. None of these cultivars proved suitable for maintaining low virus levels in the field as Lapidot et al. (2001) observed. Non-resistant cultivars should therefore be used under UV-blocking nets since their associated disease levels are acceptable. On the other hand, resistant cultivars act as disease reservoirs and in most cases have worse agronomic properties. The combination of high density nets with UV interfering features offers a convincing strategy for IPM control of insect-borne viruses. References Accotto, G.P., Navas-Castillo, J., Noris, E., Moriones, E & Louro, D. 2000: Typing of Tomato yellow leaf curl viruses in Europe. Eur. J. Plant Pathol. 106: Antignus, Y., Lapidot, M. & Cohen, S. 2001: Interference with UV vision of insects: an IPM tool to impede epidemics of insect pests and insect associated virus diseases. In K.F. Harris, O. P. Smith and J. E. Duffus, (eds.) Virus-insect-plant interactions. Academic Press, New York, pp Antignus, Y., Lapidot, M., Hadar, D., Messika, Y. & Cohen, S. 1998: UV-absorbing screens serve as optical barriers to protect crops from virus and insect pests. J. Econ. Entomol. 91: Campbell, C. L. & Madden, L.V. 1990: Introduction to Plant Disease Epidemiology. John Wiley & Sons, New York City. 532 pp. Costa, H. & Robb, K.L. 1999: Effects of ultraviolet-absorbing greenhouse plastic films on flight behavior of Bemisia argentifolii (Homoptera: Aleyrodidae) and Frankiniella occidentalis (Tysanoptera: Thripidae). J. Econ. Entomol. 92: Lapidot, M., Friedmann, M., Pilowsky, M., Ben-Joseph, R. & Cohen, S. 2001: Effect of host plant resistance to Tomato yellow leaf curl virus (TYLCV) on virus acquisition and transmission by its whitefly vector. Phytopathology 91: Monci, F., García-Andrés, S., Sánchez, F., Moriones, E., Espí, E. & Salmerón, A. 2004: Tomato yellow leaf curl disease control with UV-blocking plastic covers in commercial plastichouses of southern Spain. Acta Hort. (ISHS) 633: Moriones, E. & Navas-Castillo, J. 2000: Tomato yellow leaf curl virus, an emerging virus complex causing epidemics worldwide. Virus Res. 71: Pico, B., Díez, M. J. & Nuez, F. 1999: Improved diagnostic techniques for tomato yellow leaf curl virus in tomato breeding programs. Plant Dis. 83:
305 Integrated Control in Protected Crops, Mediterranean Climate IOBC/wprs Bulletin Vol. 29 (4) 2006 pp Iris yellow spot virus (IYSV): a new disease in Spain Carmen Córdoba 1, Llucia Martínez-Priego 1, Ramona Muñoz 2, Concepción Jordá 1. 1 Departamento de Patología Vegetal-Unidad de Virologia, Universidad Politécnica de Valencia, Camino de Vera s/n, Valencia, ( mjordag@upv.es ); 2 ITAP: Instituto Tecnológico Agronómico Provincial. Albacete, Spain. Abstract: Iris yellow spot virus (IYSV) was detected recently in onions in the central region of Spain, causing straw colored ringspots or diamond-shaped lesions on leaves and flower stalks, and eventually leading to the death of the plant. The identification was carried out by double antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) of leaf sap extracted from sympthomatic plants, using specific antisera against IYSV. The ELISA result was verified with a one step reverse transcription-polymerase chain reaction assay (RT-PCR) using primers specific to the nucleocapsid gene of IYSV. The identity of the PCR product with an expected size of approximately 790 bp. was confirmed by sequencing. This is the first report on the detection of a new tospovirus in Spain that causes disease in onions. Key words: diagnosis, onion, RT- PCR, serology, Thrips tabaci, Tospovirus Introduction So far, only three viral diseases have been identified in onion crops grown in Spain. These are Tomato spotted wilt virus (TSWV), Onion yellow dwarf virus (OYDV) and Leek yellow stripe virus (LYSV). During September 2003, a new disease was observed in several onion plants (Allium cepa L.) in Albacete, Spain. Plants showed unusual virus-like symptoms as straw-coloured, dry, tan, diamond-shaped lesions on the leaves and stalks, sometimes with necrotic lesions, curled leaves and bulbs of minor size. Severely affected plants eventually died. The kind of symptoms described was also observed in 1994 in onion crops in Brazil. There the disease called sapeca often reached an incidence of 100%, resulting in a total loss of bulb and seed production (Pozzer et al. 1994). Later, the disease was found to be associated with the tospovirus Iris yellow spot virus (IYSV) (Pozzer et al. 1999). This virus was first identified in the Netherlands, where it was occasionally detected in cultivations of Iris hollandica Tub (Derks & Lemmers 1996) and leek (A. porrum L.) (Gera et al. 1998). The symptoms described consisted of typical chlorotic spots that, in a progressed stage of the disease, developed into yellow and necrotic spots. The causal agent of this iris disease was found to be a new member of the Tospovirus group, and hence named Iris yellow spot virus (IYSV) (Cortes et al. 1998). In 1991, a new disease of onion was first observed in the United States (Hall et al. 1993), which was later ascribed to an isolate of Iris yellow spot virus. Today, IYSV is endemic in Colorado, Washington, Idaho, Oregon, Utah, California, Arizona, Nevada and New Mexico. In 1997, the disease known to onion growers in Israel as straw bleaching, and whose incidence often reached 50 to 60% resulting in heavy losses of onionbulb production, was also found to be caused by Iris yellow spot virus (Gera et al. 1998). In 2002, IYSV-like symptoms were seen in spring onion and onion bulb crops in three Australian states (Coutts et al. 2003). At this moment the presence of IYSV is also confirmed in Slovenia (reported in 2002 on leek, onion and weeds) and Iran. 293
306 Material and methods ELISA tests In order to verify the identity of the disease found in the Spanish onions, DAS-ELISA was performed on leaf extracts of symptomatic onions using different antisera against the viruses that could affect onions in Spain: Onion yellow dwarf virus (OYDV), Leek yellow stripe virus (LYSV), Cucumber mosaic virus (CMV) (antibodies from Biorad Phyto-Diagnostics Marnes-La Coquette-France cat. no , 51228, 51219), Iris yellow spot virus (IYSV) and Tomato spotted wilt virus (TSWV) (antibodies from Loewe Biochemica Sauerlach-Germany cat. no , ). ELISA tests were carried out according to the antibodies manufacturer s protocols, using healthy onion tissue as a negative control. Positive controls of each virus were obtained from the antibodies manufacturers. ELISA values were determined with a Multiskan EX Reader (ThermoLabsystems, Vantaa, Finland). Absorbance values of at least two times higher than those of the healthy controls were considered positive. RT-PCR To confirm the ELISA results, viral RNA was extracted from some onion samples that were positive in ELISA against IYSV, a healthy onion plant and a positive control of IYSV provided by DSMZ Plant Virus Collection (Braunschweig-Germany)(cat. no. PV-0528), using the RNA Wiz Kit (Ambion cat. no. 9736). The extracted RNA was used in a One-Step RT-PCR assay using the SuperScript TM One-Step RT-PCR with Platinum Taq (Invitrogen Life Technologies, Barcelona-Spain, cat no ) in the presence of the oligonucleotides IYSV1S and IYSV1A specific to the nucleocapsid gene of IYSV (Counts et al., 2003). The reverse transcription reaction was carried out at 50ºC for 30 min. Samples were amplified in 35 cycles with a Gene Amp PCR System 2400 (Perkin Elmer, Norwalk, CT). Each cycle consisted of denaturation at 94ºC (60 s), primer annealing at 50ºC (60 s), extension at 72ºC (60 s), and final extension for 10 min at 72ºC. PCR-amplified products were separated by electrophoresis on a 1.5% agarose gel in 0.5xTBE buffer for 2 h at 80V and stained with ethidium bromide. Fragment sizes were determined by comparison with a 1-Kb DNA ladder (Invitrogen Life Technologies). Sequence analysis of PCR products In order to ascertain authenticity of the virus from onion, the PCR products were purified using the High Pure PCR Product Purification Kit (Roche Diagnostics, Mannheim-Germany, cat no ) in accordance with the manufacturer s protocol. The purified DNA was sequenced by the dideoxynucleotide chain termination method on an ABI-Prism Automatic Sequence machine. The nucleotide sequences obtained were analyzed and compared with the published IYSV sequences in the NCBI Genbank. Results and discussion Highly positive values were obtained with IYSV-specific antibodies with all samples of infected onion tissue. No reaction was observed with any sample using the polyclonal antisera against TSWV, OYDV, LYSV and CMV. The RT-PCR assay produced an amplicon of an expected size of 790 bp (Figure 1, lanes O1, O2 and IYSV positive control). No amplification products were observed when noninfected plants (O1 and O2) or water were used as templates in the RT-PCR reaction (Figure 1). The nucleotide sequences obtained were analysed and compared with the published IYSV sequences in the NCBI Genbank. This comparison showed that the sequence of our PCR product was 97% identical to the sequence of IYSV nucleocapsid gene deposited in the 294

307 Genbank database under the accession number AB that it corresponded the nucleocapsid gene of IYSV. M O 1 O2 IYSV HO NT M 790 Figure 1. RT-PCR detection of IYSV using the oligonucleotides IYSV1S-IYSV1A. Lanes O1-O2 correspond to onion symptomatic plant extracts, IYSV positive control (DSMZ PV- 0528), lane HO correspond to healthy onion plant extract and NT lane correspond to not template control. M: 1Kb DNA ladder (Invitrogen Life Technologies). IYSV is included in the European and Mediterranean Plant Protection Organization alert list of viruses ( Iris yellow spot virus is a virus with a host range limited to Lilliaceae family, principally causing disease in onions, but also infects other Allium crops, including leek (A. porrum), chive (A. schoenoprasum), garlic (A. sativum) and some flower species like iris (Iris hollandica), lisianthus (Eustoma russellianum) (Kritzman et al. 2000) and Hippeastrum hybridum (Kritzman et al., 2001). In 2005, the presence of IYSV has been confirmed in Tenerife on onion and leek. There are no reports that this disease affects bulb quality or marketability. However, heavy losses of onion bulb production are reported (Kritzman et al. 2001). IYSV belongs to the tospovirus group which is transmitted by various species of thrips (Thysanoptera: Thripidae) in a propagative manner. IYSV is transmitted by Thrips tabaci, and not by Frankliniella occidentalis or F. schultzei. The virus distribution within the onion plant is uneven. The highest titers are found in the internal leaves and in leaf segments close to the bulb. This corresponds to the area that is also most preferred by thrips as a feeding site, hence it offers the best opportunity for virus acquisition and/or infection by larvae and adults, respectively (Kritzman et al., 2001). T. tabaci is present in large populations in the oniongrowing areas in Spain but the efficiency of the Mediterranean ecotype in transmitting IYSV to susceptible plants is unknown. More research is needed to shed light on their potential danger that constitutes this virus to the Spanish onions. Due to the wide distribution of thrips among the natural vegetation and the detection of IYSV in onions, this virus may become an important constraint for onion production and other important bulb-crops in Spain as currently there are no effective measures to reduce the incidence of this disease. For an effective integrated management information on the occurrence and distribution of both virus and vector is necessary. Measures that should be taken in order to prevent the disease are: the use 295
IPM strategy for tomato
 When using chemicals, please check their compatibility with our beneficial insects on our website www.biobest.be Crop info sheet Tomato IPM strategy for tomato Crop info sheet Tomato Support your health
When using chemicals, please check their compatibility with our beneficial insects on our website www.biobest.be Crop info sheet Tomato IPM strategy for tomato Crop info sheet Tomato Support your health
Proceedings of the meeting at. Murcia (Spain) 14 th 18 th May 2006
 14 th 18 th May 2006") IOBC / WPRS Working Group Integrated Control in Protected Crops, Mediterranean Climate OILB / SROP Groupe de Travail Lutte Intégrée en Cultures Protegées, Climat Méditerranéen Proceedings of the meeting
IOBC / WPRS Working Group Integrated Control in Protected Crops, Mediterranean Climate OILB / SROP Groupe de Travail Lutte Intégrée en Cultures Protegées, Climat Méditerranéen Proceedings of the meeting
Integrated Pest Management of the exotic invasive pest Tuta absoluta
 Integrated Pest Management of the exotic invasive pest Tuta absoluta ABIM Luzern, 21-10-2009 Karel Bolckmans Tuta absoluta : passport Name : Tuta absoluta (Meyrick) Family : Lepidoptera: Gelechiidae, Leafmining
Integrated Pest Management of the exotic invasive pest Tuta absoluta ABIM Luzern, 21-10-2009 Karel Bolckmans Tuta absoluta : passport Name : Tuta absoluta (Meyrick) Family : Lepidoptera: Gelechiidae, Leafmining
FUTURECO NOFLY TM A NEW TOOL AGAINST WHITEFLIES IN IPM
 FUTURECO NOFLY TM A NEW TOOL AGAINST WHITEFLIES IN IPM José Manuel Lara and Carolina Fernández FuturEco BioScience Annual Biocontrol Industry Meeting - ABIM Lucerne, 20-21 October 2008 CONTENTS PART 1:
FUTURECO NOFLY TM A NEW TOOL AGAINST WHITEFLIES IN IPM José Manuel Lara and Carolina Fernández FuturEco BioScience Annual Biocontrol Industry Meeting - ABIM Lucerne, 20-21 October 2008 CONTENTS PART 1:
Status and Outlook for Beneficial Arthropods and Nematodes. Lynn LeBeck, Ph.D. Executive Director Association of Natural Biocontrol Producers
 Status and Outlook for Beneficial Arthropods and Nematodes Lynn LeBeck, Ph.D. Executive Director Association of Natural Biocontrol Producers INTERNATIONAL BIOCONTROL INDUSTRY SYMPOSIUM, MONTEREY, CA 2016
Status and Outlook for Beneficial Arthropods and Nematodes Lynn LeBeck, Ph.D. Executive Director Association of Natural Biocontrol Producers INTERNATIONAL BIOCONTROL INDUSTRY SYMPOSIUM, MONTEREY, CA 2016
Biological Control of Key Pests of Greenhouse and Nursery Production
 UC ANR Pesticide Use Report Analysis Workgroup Meeting Tuesday, June 9 th, 2015 PES3001, UC Davis Campus Biological Control of Key Pests of Greenhouse and Nursery Production We Edwin Lewis & Michael Parrella
UC ANR Pesticide Use Report Analysis Workgroup Meeting Tuesday, June 9 th, 2015 PES3001, UC Davis Campus Biological Control of Key Pests of Greenhouse and Nursery Production We Edwin Lewis & Michael Parrella
Advantages of ecological management of leafminers (Diptera: Agromyzidae) in commercial cultivation of crisphead lettuce in southern Brazil
 in commercial cultivation of crisphead lettuce in southern Brazil") Integrated Control in Protected Crops, Mediterranean Climate IOBC-WPRS Bulletin Vol. 80, 2012 pp. 223-227 Advantages of ecological management of leafminers (Diptera: Agromyzidae) in commercial cultivation
Integrated Control in Protected Crops, Mediterranean Climate IOBC-WPRS Bulletin Vol. 80, 2012 pp. 223-227 Advantages of ecological management of leafminers (Diptera: Agromyzidae) in commercial cultivation
Aphid control in pepper
 Aphid control in pepper Jeroen van Schelt Koppert Biological Systems Tobacco aphid or Red Myzus Myzus persicae nicotianae Dense colonies in top A. solani gives strong allergic reactions from the plant
Aphid control in pepper Jeroen van Schelt Koppert Biological Systems Tobacco aphid or Red Myzus Myzus persicae nicotianae Dense colonies in top A. solani gives strong allergic reactions from the plant
IPM Strategies and Techniques
 IPM Strategies and Techniques Learn the important strategies used to implement an effective IPM program. There is much more to IPM than oils and soaps! Rafael Andy Vega and Norman C. Leppla, Ph.D. UF,
IPM Strategies and Techniques Learn the important strategies used to implement an effective IPM program. There is much more to IPM than oils and soaps! Rafael Andy Vega and Norman C. Leppla, Ph.D. UF,
Lloyd Traven, M.Sc. Peace Tree. We ALL say we want it-- BUT.. Will THEY Buy it?
 Lloyd Traven, M.Sc Propagation @ Peace Tree We ALL say we want it-- BUT.. Will THEY Buy it? 1 The Journey We are Sustainable FIRST, SOME DEFINITIONS A GROWER S DEFINITION CUT TO THE CHASE!! I I TOOK IN
Lloyd Traven, M.Sc Propagation @ Peace Tree We ALL say we want it-- BUT.. Will THEY Buy it? 1 The Journey We are Sustainable FIRST, SOME DEFINITIONS A GROWER S DEFINITION CUT TO THE CHASE!! I I TOOK IN
Applied entomology in Spanish greenhouse horticulture
 Applied entomology in Spanish greenhouse horticulture Jan van der Blom Dpt. of Pest Control, Provincial Association of Horticultural Producers and Exporters, COEXPHAL, C/ Ronda, 11-1, 04004 Almeria, Spain,
Applied entomology in Spanish greenhouse horticulture Jan van der Blom Dpt. of Pest Control, Provincial Association of Horticultural Producers and Exporters, COEXPHAL, C/ Ronda, 11-1, 04004 Almeria, Spain,
RECENT PROGRESS IN INTEGRATED PEST MANAGEMENT OF VEGETABLES
 RECENT PROGRESS IN INTEGRATED PEST MANAGEMENT OF VEGETABLES Eizi Yano Division of Entomology, National Institute of Agro-Environmental Sciences, 3-1-1, Kannondai, Tsukuba, Ibaraki 305, Japan ABSTRACT Biological
RECENT PROGRESS IN INTEGRATED PEST MANAGEMENT OF VEGETABLES Eizi Yano Division of Entomology, National Institute of Agro-Environmental Sciences, 3-1-1, Kannondai, Tsukuba, Ibaraki 305, Japan ABSTRACT Biological
WP7: IPM solutions for protected vegetables
 WP7: IPM solutions for protected vegetables List of scenarios allowing the highest efficacy of pest control IOR Results Generalist predator efficacy The aim of the laboratory tests was to determine the
WP7: IPM solutions for protected vegetables List of scenarios allowing the highest efficacy of pest control IOR Results Generalist predator efficacy The aim of the laboratory tests was to determine the
Natural Enemies (Farmers' Friends) Introduction
 Introduction") Natural Enemies (Farmers' Friends) Introduction Beneficial living organisms which reduce pests and diseases are usually present in any crop unless broad spectrum pesticides (which kill a wide range of
Natural Enemies (Farmers' Friends) Introduction Beneficial living organisms which reduce pests and diseases are usually present in any crop unless broad spectrum pesticides (which kill a wide range of
Integrating Beneficials with Bio-Pesticides: The Strawberry IPM Example. Frank V. Sances Pacific Ag Group
 Integrating Beneficials with Bio-Pesticides: The Strawberry IPM Example Frank V. Sances Pacific Ag Group Pacific Ag Group Research Facilities by Region Pacific Ag Laboratories Hydroponic Greenhouses Entomology
Integrating Beneficials with Bio-Pesticides: The Strawberry IPM Example Frank V. Sances Pacific Ag Group Pacific Ag Group Research Facilities by Region Pacific Ag Laboratories Hydroponic Greenhouses Entomology
Implications of emerging plant pests for Integrated Pest Management
 Implications of emerging plant pests for Integrated Pest Management Johannette Klapwijk IBMA EFSA Scientific Colloquim XVI on emerging plant health risks 9, 10 june 2011, Parma International Biocontrol
Implications of emerging plant pests for Integrated Pest Management Johannette Klapwijk IBMA EFSA Scientific Colloquim XVI on emerging plant health risks 9, 10 june 2011, Parma International Biocontrol
Starting a Biocontrol Program for Cut Gerbera
 Starting a Biocontrol Program for Cut Gerbera Photos are courtesy OMAFRA unless otherwise stated Graeme Murphy IPM Specialist biological control solutions So how do we start a biocontrol program Most growers
Starting a Biocontrol Program for Cut Gerbera Photos are courtesy OMAFRA unless otherwise stated Graeme Murphy IPM Specialist biological control solutions So how do we start a biocontrol program Most growers
Biological Control in Brazil
 Biological Control in Brazil Vanda H. P. Bueno Laboratory of Biological Control, Department of Entomology, Federal University of Lavras (UFLA) Brazil Biological Control Industry in Brazil 21 Companies
Biological Control in Brazil Vanda H. P. Bueno Laboratory of Biological Control, Department of Entomology, Federal University of Lavras (UFLA) Brazil Biological Control Industry in Brazil 21 Companies
Special Research Report #215: Effects of Pesticide Mixtures in Controlling Arthropod Pests of Greenhouses
 Special Research Report #215: Effects of Pesticide Mixtures in Controlling Arthropod Pests of Greenhouses A. L. Willmott, graduate student; R. A. Cloyd, Professor and Extension Specialist; K. Y. Zhu, Professor,
Special Research Report #215: Effects of Pesticide Mixtures in Controlling Arthropod Pests of Greenhouses A. L. Willmott, graduate student; R. A. Cloyd, Professor and Extension Specialist; K. Y. Zhu, Professor,
Dr Mark Whittaker. Regulatory Affairs & Client Liaison. Regulatory affairs and risk assessment
 Regulatory affairs and risk assessment Dr Mark Whittaker Regulatory Affairs & Client Liaison Direct dial: 44 (0)1423 700236 Email: Mark.Whittaker@jsci.co.uk 1 Personal background Keele University Dr Mark
Regulatory affairs and risk assessment Dr Mark Whittaker Regulatory Affairs & Client Liaison Direct dial: 44 (0)1423 700236 Email: Mark.Whittaker@jsci.co.uk 1 Personal background Keele University Dr Mark
Using biopesticides and reduced-risk pesticides for insect control in high tunnel vegetable production
 Using biopesticides and reduced-risk pesticides for insect control in high tunnel vegetable production Gerald E. Brust, IPM Vegetable Specialist Feb. 2011 Summary: Previous work by the PI has shown that
Using biopesticides and reduced-risk pesticides for insect control in high tunnel vegetable production Gerald E. Brust, IPM Vegetable Specialist Feb. 2011 Summary: Previous work by the PI has shown that
CONTROL OF! GREENHOUSE WHITEFLY PUBLICATION C212 P1469. (1974 print) c.2. agriculture. Canada
 c.2. agriculture. Canada") CONTROL OF! H GREENHOUSE WHITEFLY PUBLICATION 1469 1972 630.4 C212 P1469 1972 (1974 print) c.2 agriculture Canada Copies of this publication may be obtained from INFORMATION DIVISION CANADA DEPARTMENT
CONTROL OF! H GREENHOUSE WHITEFLY PUBLICATION 1469 1972 630.4 C212 P1469 1972 (1974 print) c.2 agriculture Canada Copies of this publication may be obtained from INFORMATION DIVISION CANADA DEPARTMENT
Laura Ingwell Wednesday, September 5, 2018
 Biological control of insects in hydroponic production Laura Ingwell Wednesday, September 5, 2018 Prevention is key Inspect all material moving into the area Have order of entry protocols www.aliexpress.com
Biological control of insects in hydroponic production Laura Ingwell Wednesday, September 5, 2018 Prevention is key Inspect all material moving into the area Have order of entry protocols www.aliexpress.com
Integrating Beneficials with Bio-Pesticides: The Strawberry IPM Example. Frank V. Sances Pacific Ag Group
 Integrating Beneficials with Bio-Pesticides: The Strawberry IPM Example Frank V. Sances Pacific Ag Group Pacific Ag Group Research Facilities by Region Pacific Ag Laboratories Hydroponic Greenhouses Entomology
Integrating Beneficials with Bio-Pesticides: The Strawberry IPM Example Frank V. Sances Pacific Ag Group Pacific Ag Group Research Facilities by Region Pacific Ag Laboratories Hydroponic Greenhouses Entomology
ONLINE COURSE PEST AND DISEASE MANAGEMENT
 ONLINE COURSE From the Lead in your field series PEST AND DISEASE MANAGEMENT in vegetables, flowers and greenhouse crops https://cropaia.com/pest-disease-management-course/ General Details Course Code:
ONLINE COURSE From the Lead in your field series PEST AND DISEASE MANAGEMENT in vegetables, flowers and greenhouse crops https://cropaia.com/pest-disease-management-course/ General Details Course Code:
Controlling Tuta absoluta, a new invasive pest in Europe
 Controlling Tuta absoluta, a new invasive pest in Europe Judit Arnó and Rosa Gabarra, IRTA, Cabrils, Spain Clockwise from above: Healthy tomatoes, adult Tuta absoluta, leaf damage and damaged fruits (
Controlling Tuta absoluta, a new invasive pest in Europe Judit Arnó and Rosa Gabarra, IRTA, Cabrils, Spain Clockwise from above: Healthy tomatoes, adult Tuta absoluta, leaf damage and damaged fruits (
need for innovation in integrated control of greenhouse pests
 need for innovation in integrated control of greenhouse pests 8 th New Ag International Conference, Miami FL, March 2010 Pierre M.J. Ramakers INTEGRATED PEST MANAGEMENT ( IPM ) regulating pest populations
need for innovation in integrated control of greenhouse pests 8 th New Ag International Conference, Miami FL, March 2010 Pierre M.J. Ramakers INTEGRATED PEST MANAGEMENT ( IPM ) regulating pest populations
Development of fungal biopesticides for use against green vegetable bugs and mirids.
 Development of fungal biopesticides for use against green vegetable bugs and mirids. K M Knight, D G Holdom, and C Hauxwell Biopesticides Unit, QDPI, 80 Meiers Road Indooroopilly 4068 Introduction Beauveria
Development of fungal biopesticides for use against green vegetable bugs and mirids. K M Knight, D G Holdom, and C Hauxwell Biopesticides Unit, QDPI, 80 Meiers Road Indooroopilly 4068 Introduction Beauveria
Practical Challenges to Inundative and Inoculative Biocontrol
 Practical Challenges to Inundative and Inoculative Biocontrol Delemont, 17-01-2017 In order of appearance: Tom Groot: Markus Knapp: Bernd Wührer: production protected crops open fields Practical Challenges
Practical Challenges to Inundative and Inoculative Biocontrol Delemont, 17-01-2017 In order of appearance: Tom Groot: Markus Knapp: Bernd Wührer: production protected crops open fields Practical Challenges
Mechanical release of Phytoseiulus persimilis and Amblyseius swirskii on protected crops
 Bulletin of Insectology 70 (2): 245-250, 2017 ISSN 1721-8861 Mechanical release of Phytoseiulus persimilis and Amblyseius swirskii on protected crops Alberto LANZONI 1, Roberta MARTELLI 2, Fabio PEZZI
Bulletin of Insectology 70 (2): 245-250, 2017 ISSN 1721-8861 Mechanical release of Phytoseiulus persimilis and Amblyseius swirskii on protected crops Alberto LANZONI 1, Roberta MARTELLI 2, Fabio PEZZI
Integrated Pest Management
 Integrated Pest Management System or strategy Utilizes all methods of pest suppression Compatible Maintain pests below economically damaging level Environmentally sound Economically sound Biological Control
Integrated Pest Management System or strategy Utilizes all methods of pest suppression Compatible Maintain pests below economically damaging level Environmentally sound Economically sound Biological Control
Biologically-Based, Insect Pest Management in Tomato and Pepper
 Biologically-Based, Insect Pest Management in Tomato and Pepper Southern Wisconsin Vegetable Workshop February 1, 2013 Russell L. Groves Department of Entomology University of Wisconsin 1630 Linden Drive
Biologically-Based, Insect Pest Management in Tomato and Pepper Southern Wisconsin Vegetable Workshop February 1, 2013 Russell L. Groves Department of Entomology University of Wisconsin 1630 Linden Drive
Integrated. Pest. Dr Paul Horne IPM Technologies Pty Ltd. Management IPM
 Integrated Pest Management IPM Dr Paul Horne IPM Technologies Support growers to adopt IPM Independent advice Experience in a wide range of horticultural and broad-acre crops IPM research and training
Integrated Pest Management IPM Dr Paul Horne IPM Technologies Support growers to adopt IPM Independent advice Experience in a wide range of horticultural and broad-acre crops IPM research and training
Mirid Bugs as Biological Control Agents in Protected Tomato Crops in the Oeste Region
 Mirid Bugs as Biological Control Agents in Protected Tomato Crops in the Oeste Region E. Figueiredo 1, R. Prieto 1, A. Mexia 1, S. Rodrigues 2, C.A. Costa 3 and M.C. Godinho 4 1 CEER, Instituto Superior
Mirid Bugs as Biological Control Agents in Protected Tomato Crops in the Oeste Region E. Figueiredo 1, R. Prieto 1, A. Mexia 1, S. Rodrigues 2, C.A. Costa 3 and M.C. Godinho 4 1 CEER, Instituto Superior
IPM Components and Packages for Tropical Vegetable Crops
 IPM Components and Packages for Tropical Vegetable Crops R. Muniappan Director, Integrated Pest Management (IPM IL) Office of International Research, Education, and Development, Virginia Tech IPM Innovation
IPM Components and Packages for Tropical Vegetable Crops R. Muniappan Director, Integrated Pest Management (IPM IL) Office of International Research, Education, and Development, Virginia Tech IPM Innovation
P. NIHOUL AND T. HANCE Catholic University of Louvain Laboratory of Ecology and Biogeography Place Croix du Sud, 5 B-1348 Louvain-la-Neuve, Belgium
 IMPLICATIONS OF INTERCROPPING (SWEET PEPPER - BIOLOGICAL CONTROL OF PESTS IN GLASSHOUSES TOMATO) FOR THE P. NIHOUL AND T. HANCE Catholic University of Louvain Laboratory of Ecology and Biogeography Place
IMPLICATIONS OF INTERCROPPING (SWEET PEPPER - BIOLOGICAL CONTROL OF PESTS IN GLASSHOUSES TOMATO) FOR THE P. NIHOUL AND T. HANCE Catholic University of Louvain Laboratory of Ecology and Biogeography Place
Developing Insecticides with an Eye toward Beneficials Safety. Manuele Ricci, Thierry Merckling, Denise Manker, Dennis Long, Nicholas Wright
 Developing Insecticides with an Eye toward Beneficials Safety Manuele Ricci, Thierry Merckling, Denise Manker, Dennis Long, Nicholas Wright (Dir. 2009/128/UE) Member States would have to create the necessary
Developing Insecticides with an Eye toward Beneficials Safety Manuele Ricci, Thierry Merckling, Denise Manker, Dennis Long, Nicholas Wright (Dir. 2009/128/UE) Member States would have to create the necessary
The Role of Integrated Pest Management in USAID s Feed the Future Initiative
 The Role of Integrated Pest Management in USAID s Feed the Future Initiative R. Muniappan Director, IPM Innovation Lab Virginia Tech Horticulture Innovation Lab Annual Meeting, Lusaka, Zambia June 8, 2015
The Role of Integrated Pest Management in USAID s Feed the Future Initiative R. Muniappan Director, IPM Innovation Lab Virginia Tech Horticulture Innovation Lab Annual Meeting, Lusaka, Zambia June 8, 2015
IPM for Legumes. Big Solutions for Small Holder Farmers.
 IPM for Legumes Big Solutions for Small Holder Farmers Realipm www.realipm.com @Realipm Big solutions For small holder farmers Real IPM has also produced a mixture of other beneficial microbes to help
IPM for Legumes Big Solutions for Small Holder Farmers Realipm www.realipm.com @Realipm Big solutions For small holder farmers Real IPM has also produced a mixture of other beneficial microbes to help
Strawberry IPM Season Review Thursday 25th June Elizabeth Town Café 10:00 am till 2 pm
 Strawberry IPM Season Review Thursday 25th June Elizabeth Town Café 10:00 am till 2 pm Major pest management challenge Participants listed more than one pest challenge. These were rated by the order of
Strawberry IPM Season Review Thursday 25th June Elizabeth Town Café 10:00 am till 2 pm Major pest management challenge Participants listed more than one pest challenge. These were rated by the order of
Name of the contact person: Jan van der Blom Position: Coord. Dpt. Production Techniques
 Centre name : COEXPHAL, Association of producers and exporters of horticultural products from Almeria City : ALMERIA Country: SPAIN Telephone: +34 Website: www.coexphal.es Facebook page: Name of the contact
Centre name : COEXPHAL, Association of producers and exporters of horticultural products from Almeria City : ALMERIA Country: SPAIN Telephone: +34 Website: www.coexphal.es Facebook page: Name of the contact
Biological Control Starting in Propagation. Ronald Valentin Technical Lead Bioline North America
 Biological Control Starting in Propagation Ronald Valentin Technical Lead Bioline North America RValentin@biolineagrosciences.com Bio-control at Propagation: Bio-control developments at a global level:
Biological Control Starting in Propagation Ronald Valentin Technical Lead Bioline North America RValentin@biolineagrosciences.com Bio-control at Propagation: Bio-control developments at a global level:
Organic Crop Pest Management
 Organic Crop Pest Management Rick Foster, Dept. of Entomology Dan Egel, SW Purdue Ag Program Liz Maynard, NW Commercial Hort. Program, Dept. of Horticulture and LA Purdue University presented at Beginning
Organic Crop Pest Management Rick Foster, Dept. of Entomology Dan Egel, SW Purdue Ag Program Liz Maynard, NW Commercial Hort. Program, Dept. of Horticulture and LA Purdue University presented at Beginning
Evaluation of Living and Synthetic Mulches in Zucchini for Control of Homopteran Pests
 Evaluation of Living and Synthetic Mulches in Zucchini for Control of Homopteran Pests Daniel L. Frank Department of Entomology and Nematology University of Florida Cucurbit Production in Florida During
Evaluation of Living and Synthetic Mulches in Zucchini for Control of Homopteran Pests Daniel L. Frank Department of Entomology and Nematology University of Florida Cucurbit Production in Florida During
The Benefits of Insecticide Use: Spinach
 Crop Protection Research Institute The Benefits of Insecticide Use: Spinach Aphids on Spinach Parasitized Aphids Leafminer Damage Plowing Spinach Under Due to Aphid Presence March 2009 Leonard Gianessi
Crop Protection Research Institute The Benefits of Insecticide Use: Spinach Aphids on Spinach Parasitized Aphids Leafminer Damage Plowing Spinach Under Due to Aphid Presence March 2009 Leonard Gianessi
2017 Statewide IPM Report July 13, 2018 Managing Thrips in Commercial Eggplant Production Systems via Implementation of Macaranga Border Crops
 2017 Statewide IPM Report July 13, 2018 Managing Thrips in Commercial Eggplant Production Systems via Implementation of Macaranga Border Crops J. Sugano, K.H. Wang, J. Silva, J. Uyeda, S. Motomura, K.
2017 Statewide IPM Report July 13, 2018 Managing Thrips in Commercial Eggplant Production Systems via Implementation of Macaranga Border Crops J. Sugano, K.H. Wang, J. Silva, J. Uyeda, S. Motomura, K.
Greenhouse Climate: an Important Consideration when Developing Pest Management Programs for Greenhouse Crops
 Greenhouse Climate: an Important Consideration when Developing Pest Management Programs for Greenhouse Crops L. Shipp a Agriculture and Agri-Food Canada Greenhouse and Processing Crops Research Centre
Greenhouse Climate: an Important Consideration when Developing Pest Management Programs for Greenhouse Crops L. Shipp a Agriculture and Agri-Food Canada Greenhouse and Processing Crops Research Centre
Cucurbits Year-Round IPM Program Annual Checklist (10/11)
") www.ipm.ucdavis.edu Cucurbits Year-Round IPM Program Annual Checklist (10/11) Supplement to UC IPM Pest Management Guidelines: Cucurbits These practices are recommended for a monitoring-based IPM program
www.ipm.ucdavis.edu Cucurbits Year-Round IPM Program Annual Checklist (10/11) Supplement to UC IPM Pest Management Guidelines: Cucurbits These practices are recommended for a monitoring-based IPM program
EFFECTS OF INTRAGUILD PREDATION AND INTERSPECIFIC COMPETITITON AMONG BIOLOGICAL CONTROL AGENTS IN AUGMENTATIVE BIOLOGICAL CONTROL IN GREENHOUSES
 Intraguild Predation and Interspecific Competition Among Biological Control Agents EFFECTS OF INTRAGUILD PREDATION AND INTERSPECIFIC COMPETITITON AMONG BIOLOGICAL CONTROL AGENTS IN AUGMENTATIVE BIOLOGICAL
Intraguild Predation and Interspecific Competition Among Biological Control Agents EFFECTS OF INTRAGUILD PREDATION AND INTERSPECIFIC COMPETITITON AMONG BIOLOGICAL CONTROL AGENTS IN AUGMENTATIVE BIOLOGICAL
Key words: Thripinema nicklewoodi, Western flower thrips, nematodes, biological control
 IOBC/wprs Bulletin Vol. 25(1) 2002 IOBC/wprs Working Group Integrated control in protected Crops and IOBC/nrs Greenhouse, Nursery, & Ornamental Landscape IPM Working Group, Proceedings of the meeting at
IOBC/wprs Bulletin Vol. 25(1) 2002 IOBC/wprs Working Group Integrated control in protected Crops and IOBC/nrs Greenhouse, Nursery, & Ornamental Landscape IPM Working Group, Proceedings of the meeting at
Biological Control in Ornamental Propagation
 Biological Control in Ornamental Propagation Ronald Valentin Biological Crop Protection Specialist Technical Trainer and Coordinator Biobest USA Inc. Biobest Canada Ltd Rvalentin@biobest.ca Propagation
Biological Control in Ornamental Propagation Ronald Valentin Biological Crop Protection Specialist Technical Trainer and Coordinator Biobest USA Inc. Biobest Canada Ltd Rvalentin@biobest.ca Propagation
Scaling Horticulture and IPM Technologies
 Feed the Future Collaborative Research Innovation Lab Partners Workshop July 9, 2013. Accra, Ghana Scaling Horticulture and IPM Technologies IPM Innovation Lab Amer Fayad, PhD Associate Director, IPM Innovation
Feed the Future Collaborative Research Innovation Lab Partners Workshop July 9, 2013. Accra, Ghana Scaling Horticulture and IPM Technologies IPM Innovation Lab Amer Fayad, PhD Associate Director, IPM Innovation
Macro-organisms for biological control why biological control agents work how and when?
 Macro-organisms for biological control why biological control agents work how and when? Roma L Gwynn rgwynn@biorationale.co.uk Sustainable Use Directive alternatives & IPM Monitoring Insects EPN Predatory
Macro-organisms for biological control why biological control agents work how and when? Roma L Gwynn rgwynn@biorationale.co.uk Sustainable Use Directive alternatives & IPM Monitoring Insects EPN Predatory
Combating insecticide resistance in major UK pests: modelling section
 Rothamsted Research where knowledge grows Combating insecticide resistance in major UK pests: modelling section Joe Helps, Frank van den Bosch, Neil Paveley Start: 1/1/2013 End: 31/7/2016 Project aim Key
Rothamsted Research where knowledge grows Combating insecticide resistance in major UK pests: modelling section Joe Helps, Frank van den Bosch, Neil Paveley Start: 1/1/2013 End: 31/7/2016 Project aim Key
Expand your horizons. FMC Australasia Pty Ltd Phone:
 Expand your horizons Benevia provides highly effective cross spectrum protection that rapidly stops insects feeding. Further damage to the plant is reduced, ensuring maximum marketability of your crop.
Expand your horizons Benevia provides highly effective cross spectrum protection that rapidly stops insects feeding. Further damage to the plant is reduced, ensuring maximum marketability of your crop.
African Organic Agriculture Manual Booklet Series No. 8 Pest and Disease Management HOW CAN I MANAGE PESTS AND DISEASES NATURALLY?
 African Organic Agriculture Manual Booklet Series No. 8 Pest and Disease Management HOW CAN I MANAGE PESTS AND DISEASES NATURALLY? What do I need to know? It is important to know which pathogens create
African Organic Agriculture Manual Booklet Series No. 8 Pest and Disease Management HOW CAN I MANAGE PESTS AND DISEASES NATURALLY? What do I need to know? It is important to know which pathogens create
Preface... I Contents... III
 IOBC-WPRS Bulletin Vol. 80, 2012 Working Group Integrated Control in Protected Crops, Mediterranean Climate. Preceedings of the meeting at Catania, Sicily (Italy), 09-12 October, 2012. Edited by: Cristina
IOBC-WPRS Bulletin Vol. 80, 2012 Working Group Integrated Control in Protected Crops, Mediterranean Climate. Preceedings of the meeting at Catania, Sicily (Italy), 09-12 October, 2012. Edited by: Cristina
Integrated Pest Management Package on protected crops
 Integrated Pest Management Package on protected crops by Josef Tedeschini Plant Protection Laboratory, Agriculture University of Tirana The Integrated Pest Management (IPM) Package on protected crops is
Integrated Pest Management Package on protected crops by Josef Tedeschini Plant Protection Laboratory, Agriculture University of Tirana The Integrated Pest Management (IPM) Package on protected crops is
Pest Management in Organic Systems. Juan R. Anciso Extension Vegetable Specialist Texas AgriLife Extension Service
 Pest Management in Organic Systems Juan R. Anciso Extension Vegetable Specialist Texas AgriLife Extension Service IPM Integrated Pest Management Plan ahead (use preventive strategies where possible) Use
Pest Management in Organic Systems Juan R. Anciso Extension Vegetable Specialist Texas AgriLife Extension Service IPM Integrated Pest Management Plan ahead (use preventive strategies where possible) Use
Presented in 2012 by Brian Spencer Applied Bio-nomics Ltd
 Presented in 2012 by Brian Spencer Applied Bio-nomics Ltd Aphids have a very high Gross out factor When conditions are perfect, their lifecycle is extremely short It needs to be short, because everybody
Presented in 2012 by Brian Spencer Applied Bio-nomics Ltd Aphids have a very high Gross out factor When conditions are perfect, their lifecycle is extremely short It needs to be short, because everybody
NAPPO. John Goolsby Ph.D.
 NAPPO John Goolsby Ph.D. Research Entomologist Biological Control U.S. Dept. of Agriculture, Agricultural Research Service, Moore Airbase, Edinburg, TX, USA Proposed Action 1.1 Purpose of the release (reflects
NAPPO John Goolsby Ph.D. Research Entomologist Biological Control U.S. Dept. of Agriculture, Agricultural Research Service, Moore Airbase, Edinburg, TX, USA Proposed Action 1.1 Purpose of the release (reflects
#ZERO RESIDUE. Mealy bug and White fly IN ROSES SEE IT. BELIEVE IT. DO IT PART 5 ZERORESIDUE.
 . #ZERO RESIDUE IN ROSES SEE IT. BELIEVE IT. DO IT PART 5 Mealy bug and White fly ZERORESIDUE SEE IT. BELIEVE IT. DO IT Realipm @Realipm Realipm www.realipm.com Introduction Most growers complain that
. #ZERO RESIDUE IN ROSES SEE IT. BELIEVE IT. DO IT PART 5 Mealy bug and White fly ZERORESIDUE SEE IT. BELIEVE IT. DO IT Realipm @Realipm Realipm www.realipm.com Introduction Most growers complain that
Ecology and Management of Scirtothrips dorsalis
 Ecology and Management of Scirtothrips dorsalis Presented By: Dr. Joe Funderburk, UF/IFAS Contributing Authors: UF/IFAS: J. Funderburk,, and L. Osborne A.C. Hodges, UF/IFAS, SPDN (ed.) POPULATION ATTRIBUTES
Ecology and Management of Scirtothrips dorsalis Presented By: Dr. Joe Funderburk, UF/IFAS Contributing Authors: UF/IFAS: J. Funderburk,, and L. Osborne A.C. Hodges, UF/IFAS, SPDN (ed.) POPULATION ATTRIBUTES
From entomology in Bologna 1602 to pesticide free food in Joop C. van Lenteren Laboratory of Entomology Wageningen University The Netherlands
 From entomology in Bologna 1602 to pesticide free food in 2004 Joop C. van Lenteren Laboratory of Entomology Wageningen University The Netherlands From entomology in Bologna to pesticide free food What
From entomology in Bologna 1602 to pesticide free food in 2004 Joop C. van Lenteren Laboratory of Entomology Wageningen University The Netherlands From entomology in Bologna to pesticide free food What
Pesticide Applicator Recertification Credit
 Pesticide Applicator Recertification Credit The following states offer credit for this session. How To Combine Biocontrols And Conventional Pesticides GH313 Monday, July 17, 2017 11:00 AM to12:00 PM Arizona
Pesticide Applicator Recertification Credit The following states offer credit for this session. How To Combine Biocontrols And Conventional Pesticides GH313 Monday, July 17, 2017 11:00 AM to12:00 PM Arizona
On the control of the cotton aphid, Aphis gossypii Glov. (Hemiptera: Aphididae), on cucumber in greenhouses
, on cucumber in greenhouses") Eid et al. Egyptian Journal of Biological Pest Control (2018) 28:64 https://doi.org/10.1186/s41938-018-0065-9 Egyptian Journal of Biological Pest Control RESEARCH On the control of the cotton aphid, Aphis
Eid et al. Egyptian Journal of Biological Pest Control (2018) 28:64 https://doi.org/10.1186/s41938-018-0065-9 Egyptian Journal of Biological Pest Control RESEARCH On the control of the cotton aphid, Aphis
BioCeres WP AzaGuard. IR-4 Bio-Pesticide Workshop, Presented by: Manuel Campos
 BioCeres WP AzaGuard IR-4 Bio-Pesticide Workshop, 2016 Presented by: Manuel Campos BioCeres WP Active Ingredient: Beauveria bassiana strain-ant-03-20.0% w/w Shelf Life: 6 months Environmental Hazards:
BioCeres WP AzaGuard IR-4 Bio-Pesticide Workshop, 2016 Presented by: Manuel Campos BioCeres WP Active Ingredient: Beauveria bassiana strain-ant-03-20.0% w/w Shelf Life: 6 months Environmental Hazards:
Growing. Frequently asked questions & answers. 1. What is SIVANTO prime? 2. What is the mode of action of SIVANTO prime?
 Growing inunison Frequently asked questions & answers 1. What is SIVANTO prime? SIVANTO prime is the newest insecticide from Bayer CropScience for the control of major sucking insect pests and has an outstanding
Growing inunison Frequently asked questions & answers 1. What is SIVANTO prime? SIVANTO prime is the newest insecticide from Bayer CropScience for the control of major sucking insect pests and has an outstanding
Cucumbers (Pickles and Slicers) IPM Element Revised March, 2012
 IPM Element Revised March, 2012") Cucumbers (Pickles and Slicers) IPM Element Revised March, 2012 Edited by Jim Jasinski Contributing Authors: Celeste Welty (Entomology); Bob Precheur, Mark Bennett, Doug Doohan (Horticulture & Crop Science);
Cucumbers (Pickles and Slicers) IPM Element Revised March, 2012 Edited by Jim Jasinski Contributing Authors: Celeste Welty (Entomology); Bob Precheur, Mark Bennett, Doug Doohan (Horticulture & Crop Science);
Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenius (Het.: Miridae)
") Biocontrol Science and Technology, August 2005; 15(5): 513 /518 SHORT COMMUNICATION Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenius (Het.: Miridae)
Biocontrol Science and Technology, August 2005; 15(5): 513 /518 SHORT COMMUNICATION Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenius (Het.: Miridae)
IPM concepts in tobacco entomology
 IPM concepts in tobacco entomology Hannah J. Burrack Assistant Professor & Extension Specialist Department of Entomology, NC State 919.513.4344 hannah_burrack@ncsu.edu Quick Overview of Insect Relationships
IPM concepts in tobacco entomology Hannah J. Burrack Assistant Professor & Extension Specialist Department of Entomology, NC State 919.513.4344 hannah_burrack@ncsu.edu Quick Overview of Insect Relationships
IPM concepts in tobacco entomology
 www.anilrana13014.weebly.com www.k8449r.weebly.com IPM concepts in tobacco entomology Hannah J. Burrack Assistant Professor & Extension Specialist Department of Entomology, NC State 919.513.4344 hannah_burrack@ncsu.edu
www.anilrana13014.weebly.com www.k8449r.weebly.com IPM concepts in tobacco entomology Hannah J. Burrack Assistant Professor & Extension Specialist Department of Entomology, NC State 919.513.4344 hannah_burrack@ncsu.edu
Using biocontrol in vegetable & fruit crops. Celeste Welty Ohio State University August 2014
 Using biocontrol in vegetable & fruit crops Celeste Welty Ohio State University August 2014 Topics Cast of characters Types of biocontrol Conservation Augmentation Examples Greenhouse Field Biological
Using biocontrol in vegetable & fruit crops Celeste Welty Ohio State University August 2014 Topics Cast of characters Types of biocontrol Conservation Augmentation Examples Greenhouse Field Biological
THE IMPORTANCE OF SOIL ECOLOGY IN SUSTAINABLE AGRICULTURE
 THE IMPORTANCE OF SOIL ECOLOGY IN SUSTAINABLE AGRICULTURE Clive A. Edwards & Norman Q. Arancon The Soil Ecology Laboratory The Ohio State University Columbus, Ohio 1 SUSTAINABLE AGRICULTURE INTEGRATED
THE IMPORTANCE OF SOIL ECOLOGY IN SUSTAINABLE AGRICULTURE Clive A. Edwards & Norman Q. Arancon The Soil Ecology Laboratory The Ohio State University Columbus, Ohio 1 SUSTAINABLE AGRICULTURE INTEGRATED
10. Arthropod pests in poultry production
 67 10. Arthropod pests in poultry production 10.1 Prevalence of the chicken mite The chicken mite, Dermanyssus gallinae, is a well known ectoparasite of poultry. In May 1997, a questionnaire was sent to
67 10. Arthropod pests in poultry production 10.1 Prevalence of the chicken mite The chicken mite, Dermanyssus gallinae, is a well known ectoparasite of poultry. In May 1997, a questionnaire was sent to
5. Simple and Effective Integrated Pest Management Technique for Vegetables in Northeast Thailand
 5. Simple and Effective Integrated Pest Management Technique for Vegetables in Northeast Thailand Introduction Somchai Chuachin, Thawilkal Wangkahart, Suhas P Wani, TJ Rego and Prabhakar Pathak Insect
5. Simple and Effective Integrated Pest Management Technique for Vegetables in Northeast Thailand Introduction Somchai Chuachin, Thawilkal Wangkahart, Suhas P Wani, TJ Rego and Prabhakar Pathak Insect
CLIMEX models for selected glasshouse biological control agents
 CLIMEX models for selected glasshouse biological control agents Logan DP May 2012 A report prepared for Horticulture New Zealand DP Logan Plant & Food Research, Te Puke SPTS No. 6938 DISCLAIMER Unless
CLIMEX models for selected glasshouse biological control agents Logan DP May 2012 A report prepared for Horticulture New Zealand DP Logan Plant & Food Research, Te Puke SPTS No. 6938 DISCLAIMER Unless
Biological Control: The Basics. Ian Brown Ph.D. Georgia Southwestern State University
 Biological Control: The Basics Ian Brown Ph.D. Georgia Southwestern State University Biological Control What is it? Why use it? When does it work? Where does it work best? Types of Biological Control?
Biological Control: The Basics Ian Brown Ph.D. Georgia Southwestern State University Biological Control What is it? Why use it? When does it work? Where does it work best? Types of Biological Control?
Changes in Integrated Pest Management
 Changes in Integrated Pest Management Dr. GÉZA RIPKA Agricultural Office Plant Protection, Soil Conservation and Agro-environment Directorate Dr. JÁNOS MOLNÁR Ministry of Rural Development Agriculture
Changes in Integrated Pest Management Dr. GÉZA RIPKA Agricultural Office Plant Protection, Soil Conservation and Agro-environment Directorate Dr. JÁNOS MOLNÁR Ministry of Rural Development Agriculture
Biological Control of Wireworms. Todd Kabaluk, Research Biologist Integrated Pest Management Agassiz Research and Development Centre British Columbia
 Biological Control of Wireworms Todd Kabaluk, Research Biologist Integrated Pest Management Agassiz Research and Development Centre British Columbia Presentation Summary Monitoring wireworms to predict
Biological Control of Wireworms Todd Kabaluk, Research Biologist Integrated Pest Management Agassiz Research and Development Centre British Columbia Presentation Summary Monitoring wireworms to predict
Toward a Comprehensive Approach to the Biological Control of Wireworms
 Toward a Comprehensive Approach to the Biological Control of Wireworms Todd Kabaluk, Research Biologist Integrated Pest Management Agassiz Research and Development Centre British Columbia Microbial pesticides
Toward a Comprehensive Approach to the Biological Control of Wireworms Todd Kabaluk, Research Biologist Integrated Pest Management Agassiz Research and Development Centre British Columbia Microbial pesticides
Control of insect and mite pests in grains insecticide resistance and IPM. Paul Umina Svetlana Micic Laura Fagan
 Control of insect and mite pests in grains insecticide resistance and IPM Paul Umina Svetlana Micic Laura Fagan Insecticide Resistance and RLEM Paul Umina CESAR and The University of Melbourne 30 Flemington
Control of insect and mite pests in grains insecticide resistance and IPM Paul Umina Svetlana Micic Laura Fagan Insecticide Resistance and RLEM Paul Umina CESAR and The University of Melbourne 30 Flemington
Integrated Pest Management Improvements in California Melons from 2003 to 2016
 Integrated Pest Management Improvements in California Melons from 2003 to 2016 Matthew Baur Amanda Crump James Farrar Steve Elliott October 2018 1 EXECUTIVE SUMMARY Changes in pest management practices
Integrated Pest Management Improvements in California Melons from 2003 to 2016 Matthew Baur Amanda Crump James Farrar Steve Elliott October 2018 1 EXECUTIVE SUMMARY Changes in pest management practices
Pre-planting inoculation for early establishment of Dicyphus bolivari and D. errans on tomatoes
 https://doi.org/10.1007/s10526-018-09911-3 (0456789().,-volV) ( 04567 89().,-volV) BioControl (2019) 64:33 41 Pre-planting inoculation for early establishment of Dicyphus bolivari and D. errans on tomatoes
https://doi.org/10.1007/s10526-018-09911-3 (0456789().,-volV) ( 04567 89().,-volV) BioControl (2019) 64:33 41 Pre-planting inoculation for early establishment of Dicyphus bolivari and D. errans on tomatoes
IPM CRSP GOALS. Develop Improved IPM Technologies Develop Effective Institutional Changes
 IPM CRSP GOALS Develop Improved IPM Technologies Develop Effective Institutional Changes MAJOR OBJECTIVES Reduce crop losses Increase farmer s income Reduce pesticide use Reduce pesticide residues on crops
IPM CRSP GOALS Develop Improved IPM Technologies Develop Effective Institutional Changes MAJOR OBJECTIVES Reduce crop losses Increase farmer s income Reduce pesticide use Reduce pesticide residues on crops
DISEASE MANAGEMENT STRATEGIES
 DISEASE MANAGEMENT STRATEGIES Managing diseases effectively involves making the best possible decisions to reduce the risk of serious disease-related losses. The strategies upon which effective management
DISEASE MANAGEMENT STRATEGIES Managing diseases effectively involves making the best possible decisions to reduce the risk of serious disease-related losses. The strategies upon which effective management
Present practice, on-going research and future potential for non-chemical pest management in fruit and berry production in Denmark
 Present practice, on-going research and future potential for non-chemical pest management in fruit and berry production in Denmark Lene Sigsgaard Department of Plant and Environmental Sciences Section
Present practice, on-going research and future potential for non-chemical pest management in fruit and berry production in Denmark Lene Sigsgaard Department of Plant and Environmental Sciences Section
BLUE ALFALFA APHID: OLD PEST WITH NEW MANAGEMENT CHALLENGES
 BLUE ALFALFA APHID: OLD PEST WITH NEW MANAGEMENT CHALLENGES E. T. Natwick and M. I. Lopez E. Natwick is a UCCE Farm Advisors, UC ANR Cooperative Extension M. I. Lopez is a Laboratory Assistant, UC ANR
BLUE ALFALFA APHID: OLD PEST WITH NEW MANAGEMENT CHALLENGES E. T. Natwick and M. I. Lopez E. Natwick is a UCCE Farm Advisors, UC ANR Cooperative Extension M. I. Lopez is a Laboratory Assistant, UC ANR
Follow for more. tuta absoluta
 Follow us @Realipm for more tuta absoluta Real Solutions Real Farming Founded in 2013 by owners, Louise Labuschagne and Dr Henry Wainwright, Real IPM now employs 250 staff. Our BASIS-qualified field consultants
Follow us @Realipm for more tuta absoluta Real Solutions Real Farming Founded in 2013 by owners, Louise Labuschagne and Dr Henry Wainwright, Real IPM now employs 250 staff. Our BASIS-qualified field consultants
Greenhouse Tomato Production. Mary Peet North Carolina State University
 Greenhouse Tomato Production Mary Peet North Carolina State University Topics (mostly tomatoes) Statistics Pro s and Cons Decisions Production cycle for tomatoes Greenhouse types Greenhouse costs How Many
Greenhouse Tomato Production Mary Peet North Carolina State University Topics (mostly tomatoes) Statistics Pro s and Cons Decisions Production cycle for tomatoes Greenhouse types Greenhouse costs How Many
Monitoring and decision making strategies for soybean aphid management. Jordan Bannerman Department of Entomology University of Manitoba
 Monitoring and decision making strategies for soybean aphid management Jordan Bannerman Department of Entomology University of Manitoba Overview Soybean aphid (SBA) incidence and timing SBA economic threshold
Monitoring and decision making strategies for soybean aphid management Jordan Bannerman Department of Entomology University of Manitoba Overview Soybean aphid (SBA) incidence and timing SBA economic threshold
NORTHERN FEBRUARY 2017 SUNFLOWER CONTENTS
 NORTHERN FEBRUARY 2017 SUNFLOWER CONTENTS Sunflowers What s new Introduction A.1 Agronomy at a glance...xvi A.2 Crop overview...xvii A.3 Key management issues...xvii A.4 End uses...xviii 1 Planning/Paddock
NORTHERN FEBRUARY 2017 SUNFLOWER CONTENTS Sunflowers What s new Introduction A.1 Agronomy at a glance...xvi A.2 Crop overview...xvii A.3 Key management issues...xvii A.4 End uses...xviii 1 Planning/Paddock
CRANBERRY PEST MANAGEMENT IN THE FUTURE
 CRANBERRY PEST MANAGEMENT IN THE FUTURE 17 Daniel L. Mahr Department of Entomology University of Wisconsin - Madison The Food Quality Protection Act of 1996 (FQPA) is likely to have a profound impact on
CRANBERRY PEST MANAGEMENT IN THE FUTURE 17 Daniel L. Mahr Department of Entomology University of Wisconsin - Madison The Food Quality Protection Act of 1996 (FQPA) is likely to have a profound impact on
IPM in Brassica vegetables and oilseed rape: challenges and opportunities
 IPM in Brassica vegetables and oilseed rape: challenges and opportunities EIP focus group IPM in Brassica Monica Höfte Ghent University Inge Van Oost DG Agriculture and Rural development The European Innovation
IPM in Brassica vegetables and oilseed rape: challenges and opportunities EIP focus group IPM in Brassica Monica Höfte Ghent University Inge Van Oost DG Agriculture and Rural development The European Innovation
STATUS OF ENTOMOLOGICAL RESEARCH ON INSECT TRANSMITTED PLANT DISEASES IN CANADA
 STATUS OF ENTOMOLOGICAL RESEARCH ON INSECT TRANSMITTED PLANT DISEASES IN CANADA A report prepared by a Special Committee of the Entomological Society of Canada Status of Entomological Research on Insect
STATUS OF ENTOMOLOGICAL RESEARCH ON INSECT TRANSMITTED PLANT DISEASES IN CANADA A report prepared by a Special Committee of the Entomological Society of Canada Status of Entomological Research on Insect
Key notes. IPM Systems - 1 -
 IOBC/wprs Bulletin, Vol. 49, 2009 IOBC/WPRS Working Group Integrated Control in Protected Crops, Mediterranean Climate. Preceedings of the meeting at Chania (Greece: Crete), 6-11 September, 2009. Edited
IOBC/wprs Bulletin, Vol. 49, 2009 IOBC/WPRS Working Group Integrated Control in Protected Crops, Mediterranean Climate. Preceedings of the meeting at Chania (Greece: Crete), 6-11 September, 2009. Edited
High Tunnel Integrated Pest Management (IPM): Insects and Mites
: Insects and Mites") High Tunnel Integrated Pest Management (IPM): Insects and Mites Cathy E. Thomas* and Eric P. Burkhart** Pennsylvania Department of Agriculture* Department of Horticulture** Terrance T. Nennich, University
High Tunnel Integrated Pest Management (IPM): Insects and Mites Cathy E. Thomas* and Eric P. Burkhart** Pennsylvania Department of Agriculture* Department of Horticulture** Terrance T. Nennich, University
Horticultural Production in Nebraska Organic Vegetable Production Integrated, Sustainable Production Systems. Considering Organic Production?
 Horticultural Production in Nebraska 2002 Organic Vegetable Production Integrated, Sustainable Production Systems Laurie Hodges & Liz Sarno University of Nebraska-Lincoln Extension Vegetables, melons,
Horticultural Production in Nebraska 2002 Organic Vegetable Production Integrated, Sustainable Production Systems Laurie Hodges & Liz Sarno University of Nebraska-Lincoln Extension Vegetables, melons,
Benefits of Pyrethroids to Citrus
 PYRETHROIDS BENEFITS PROJECT The Pyrethroid Working Group contracted an extensive analysis of the benefits of to agriculture. A multitude of data was analyzed with different methodologies to determine
PYRETHROIDS BENEFITS PROJECT The Pyrethroid Working Group contracted an extensive analysis of the benefits of to agriculture. A multitude of data was analyzed with different methodologies to determine
Biological products: their significance in IPM strategies and risk management. Wilma Mac Pherson Minor crop workshop 11 April 2018
 Biological products: their significance in IPM strategies and risk management Wilma Mac Pherson Minor crop workshop 11 April 2018 Biological products Sustainable or not? 10 15 years ago Biological products
Biological products: their significance in IPM strategies and risk management Wilma Mac Pherson Minor crop workshop 11 April 2018 Biological products Sustainable or not? 10 15 years ago Biological products