THE POTENTIAL ROLE OF VH REPLACEMENT IN EDITING AND GENERATING AUTOREACTIVE ANTIBODIES RUN FAN
|
|
|
- Stephen Powell
- 5 years ago
- Views:
Transcription
1 THE POTENTIAL ROLE OF VH REPLACEMENT IN EDITING AND GENERATING AUTOREACTIVE ANTIBODIES by RUN FAN ZHIXIN ZHANG, MENTOR PETER D BURROWS LOUIS B JUSTEMENT JOHN D MOUNTZ HARRY W SCHROEDER A DISSERTATION Submitted to the graduate faculty of The University of Alabama at Birmingham, in partial fulfillment of the requirements for the degree of Doctor of Philosophy BIRMINGHAM, ALABAMA 2009
2 Copyright by Run Fan 2009
3 THE POTENTIAL ROLE OF VH REPLACEMENT IN EDITING AND GENERATING AUTOREACTIVE ANTIBODIES RUN FAN Department of Microbiology ABSTRACT V H replacement occurs through RAG-mediated secondary recombination between an upstream V H gene and an existing V H DJ H joint. In the first part of this dissertation, we focused on the potential regulation of V H replacement in the human B lineage EU12 cells μhc + population, which underwent spontaneous V H replacement during in vitro culture. We cloned one of the dominant IgH genes, A19H, expressed in the EU12 μhc + cells, and generated recombinant antibodies. We found that these recombinant antibodies recognize SM antigens. The anti-sm reactivity of A19H can not be efficiently neutralized by a randomly selected group of fourteen IgL chains from healthy controls, suggesting that the SM reactivity is mainly contributed by the A19H heavy chain. Indeed, the anti-sm reactivity was attenuated in the subsequent V H replacement product, A21H, cloned from the EU12 μhc + cells. These results indicated that V H replacement is essential to edit IgH genes encoding anti-sm antibodies. In the second part of this dissertation, we investigated the potential contribution of V H replacement in systematic lupus erythematosus (SLE). We performed single cell PCR assay and cloned 447 IgH genes with paired Igκ or Igλ genes from plasma cells (CD19 low CD27 high ) and naïve B cells (CD19 + CD27 - ) of three SLE patients and two i
4 healthy controls. Analyses of the IgL genes from SLE patients showed signs of excessive receptor editing of IgL genes, leading to elevated usages of downstream Jκ5 and Jλ3 genes. Strikingly, the frequency of V H replacement products in the IgH genes from all three SLE patients is significantly higher than in the control clones, especially in plasma cells. The V H replacement footprints preferentially encoded charged amino acids in the IgH CDR3 regions. Analyses of 12 recombinant antibodies derived from SLE plasma cells showed that 75% of recombinant antibodies recognize with cytoplasmic or nuclear antigens, in which more than half of them are encoded by V H replacement products. Taken together, the works presented in this dissertation suggested that V H replacement is normally employed to delete unwanted IgH genes; however, excessive V H replacement might generate IgH genes with extremely long CDR3 regions, which encode autoreactive antibodies. Keywords: V H replacement, EU12, autoreactive antibody, SM antigen, SLE ii
5 DEDICATION This dissertation is dedicated to my parents Wenfang Fan and Yueling Yan. They always cheer me up and stand by me through good and bad times in my life. This dissertation is also dedicated to my husband Yongliang Huo, for his support and encouragement throughout my PhD study. I am so grateful for his presence in my life. Although we were 1,000 miles apart geographically in my last year of PhD study, we felt never been closer. iii
6 ACKNOWLEDGEMENT This work was carried out in the Department of Microbiology, University of Alabama at Birmingham, during the years I would like to thank my advisor, Dr. Zhixin Zhang, for his guidance and providing me an excellent atmosphere for my PhD study. I would like to express my deepest gratitude to all my thesis committee members, Dr. Peter Burrows, Dr. Louis Justement, Dr. Harry Schroeder and Dr. John Mountz. They not only gave me suggestions on my projects, but also help me to become a scientist. They have consistently helped me keep perspective on what is important in life and shown me how to deal with reality. I thank everybody of Dr. Zhang lab including the lab members Jing Liu, Sangyong Hong and Huang Lin who moved from UAB to Nebraska; the previous lab members Zhihong Yu, Dr. Ti He, Dr. Himanshu Raikwar and Ashley Majors; the new members joined our lab after our lab moved to Nebraska Dr. Wail Hassan, Dr. Miles Lange, Dr. Kathy Nichol-Marran and Wanqin Xie. I also thank everyone from the former DCI (Division of Developmental and Clinical Immunology) of UAB. It was like a big family. They generously shared their research experiences and insights that supported and expanded my own work. Their knowledge challenged me and inspired me. I would like to thank Dr. Teresa Santiago, who as a good friend, is always willing to help me, giving me suggestions and encouraging me with her best wishes. I must acknowledge the many friends from Birmingham. All these good memories stay in my heart no matter where I go. iv
7 TABLE OF CONTENTS Page ABSTRACT...i DEDICATION...iii ACKNOWLEDGEMENTS...iv LIST OF TABLES...vi LIST OF FIGURES...vii LIST OF ABBREVIATIONS...ix INTRODUCTION B cell development...1 V(D)J recombination...9 Generation of autoreactive antibodies and negative selections...10 V H replacement...14 Systemic lupus erythematosus...31 EDITING OF IGH GENE ENCODING ANTI-SM ANTIBODY BY VH REPLACEMENT...35 POTENTIAL CONTRIBUTION OF VH REPLACEMENT PROUDUCTS TO THE GENERATION OF AUTOREACTIVE ANTIBODIES IN SYSTEMIC LUPUS ERYTHEMATOSUS...78 CONCLUSION GENERAL REFERENCES Appendix: IRB APPROVAL v
8 LIST OF TABLES Table Page INTRODUCTION 1 Unique motifs (V H replacement footprint) contributed by V H genes...24 EDITING OF IGH GENE ENCODING ANTI-SM ANTIBODY BY VH REPLACEMENT 1 Features of light chains used for neutralization experiments...59 POTENTIAL CONTRIBUTION OF VH REPLACEMENT PROUDUCTS TO THE GENERATION OF AUTOREACTIVE ANTIBODIES IN SYSTEMIC LUPUS ERYTHEMATOSUS 1 V H replacement products in SLE patients...95 vi
9 LIST OF FIGURES Figure Page INTRODUCTION 1 B cell development in the bone marrow and periphery Model of serial V H replacement Biological significance of V H replacement Potential contributions of V H replacement products to autoimmune diseases and anti-viral responses...30 EDITING OF IGH GENE ENCODING ANTI-SM ANTIBODY BY VH REPLACEMENT 1 Recombinant antibodies derived from EU12 cells are autoreactive Recombinant antibodies derived from EU12 cells react with SM antigens Changing the light chains cannot efficiently neutralize the anti-sm reactivity contributed by the A19H heavy chain Treatment of EU12 μhc + cells with purified SM antigen enhances V H replacement V H replacement product, A21H, reduces anti-sm reactivity contributed by A19H heavy chain Different effects of changing light chains on anti-sm reactivity contributed by the heavy chains from SLE B cells The model of anti-sm antibody production in EU12 μhc + cells...72 vii
10 POTENTIAL CONTRIBUTION OF VH REPLACEMENT PROUDUCTS TO THE GENERATION OF AUTOREACTIVE ANTIBODIES IN SYSTEMIC LUPUS ERYTHEMATOSUS 1 The CD19 low CD27 high plasma cells are significantly elevated in SLE patients The IgH gene repertoire in SLE patients IgH genes derived from SLE plasma cells have features of autoreactive antibodies IgH sequences derived from SLE plasma cells are highly mutated Excessive Jκ and Jλ editing in SLE patients Excessive V H replacement in SLE patients V H replacement footprints preferentially generate charged amino acids in the CDR3 region Recombinant antibodies derived from SLE plasma cells encode autoreactive antibodies and are positive for ANA staining The model of generation of autoreactive antibodies in SLE by V H replacement Supplementary 1 Jκ and Jλ gene usages for paring with V H replacement versus non-v H replacement products in SLE plasma cells viii
11 LIST OF ABBREVIATIONS AID BCR Blimp1 BLNK BM BSAP EBF CDR crss HEL GFP Igα Igβ IgH IgL Activation Induced Deaminase B Cell Receptor B Lymphocyte-Induced Maturation Protein-1 B cell Linker Bone Marrow B cell Specific Activator Protein Early B cell Factor Complementary-Determining Region criptic Recombination Signal Sequence Hen Egg-white Lysozyme Green Fluorescence Protein Immunoglobulin α Immunoglobulin β Immunoglobulin Heavy Chain Immunoglobulin Light Chain IRF4 Interferon-regulatory factor 4 MHC MZ RAG Major Histocompatibility Complex Marginal Zone Recombination Activating Gene ix
12 RSS SLE SM TdT Xbp1 Recombination Signal Sequence Systemic Lupus Erythematosus Smith Antigen Terminal Deoxynucleotidyl Transferase X-box Binding protein-1 x
13 INTRODUCTION B cell development B lineage lymphocytes are produced in the fetal liver in early human life and bone marrow throughout the adult life (Figure 1) (1). B lineage cells of different developmental stages can be separated according to the different surface markers. CD19 is B lineage cell specific surface marker expressed during B lineage cell development, except plasma cells (1). Pro-B cell is the earliest stage of committed B cell (2). The beginning stage of pro-b cell can be further specified as pre-pro-b cell. There s no rearrangement on immunoglobulin (Ig) genes for either heavy chain or light chain genes at pre-pro-b cell stage. Pre-pro-B cells do not express the surface B cell receptor (BCR), so this very first step of B cell commitment is independent of BCR or antigen (3). Transcription factors PU-1, E2A and EBF are important for early B cell development. PU-1 and E2A initiate EBF function and then EBF activates Pax5 gene expression (4). Pax5 is essential for maintaining B lineage commitment (5). Pax5 gene encodes B cell specific activator protein (BSAP). Pax5 -/- pro-b cells are not restricted to B lineage development and can differentiate into other cell types under appropriate conditions (5, 6). Pre-pro-B cell becomes pro-b cell when the first stage that V(D)J recombination is initiated during B cell development (2). The V(D)J recombination process is catalyzed by the RAG1 and RAG2 protein complexes (7, 8). RAG1 and RAG2 expression is tightly 1
14 regulated in a developmental stage specific manner in developing lymphocytes (9, 10). Ig gene recombination begins with D to J H segment rearrangements on one of the heavy chain loci. Following DJ H rearrangement, V H genes become accessible and V H to DJ H rearrangement in initiated (11, 12). If the first V H DJ H rearrangement is nonproductive, it will start again on the other chromosome. Bone marrow microenvironment is important for the differentiation from pro-b cells to pre-b cells. For mouse, stromal cells in bone marrow interact with pro-b cells and secret cytokines, such as IL-7, which is important for murine B cell development. However, for human pro-b cells, the equivalent growth factor has not been identified. 2
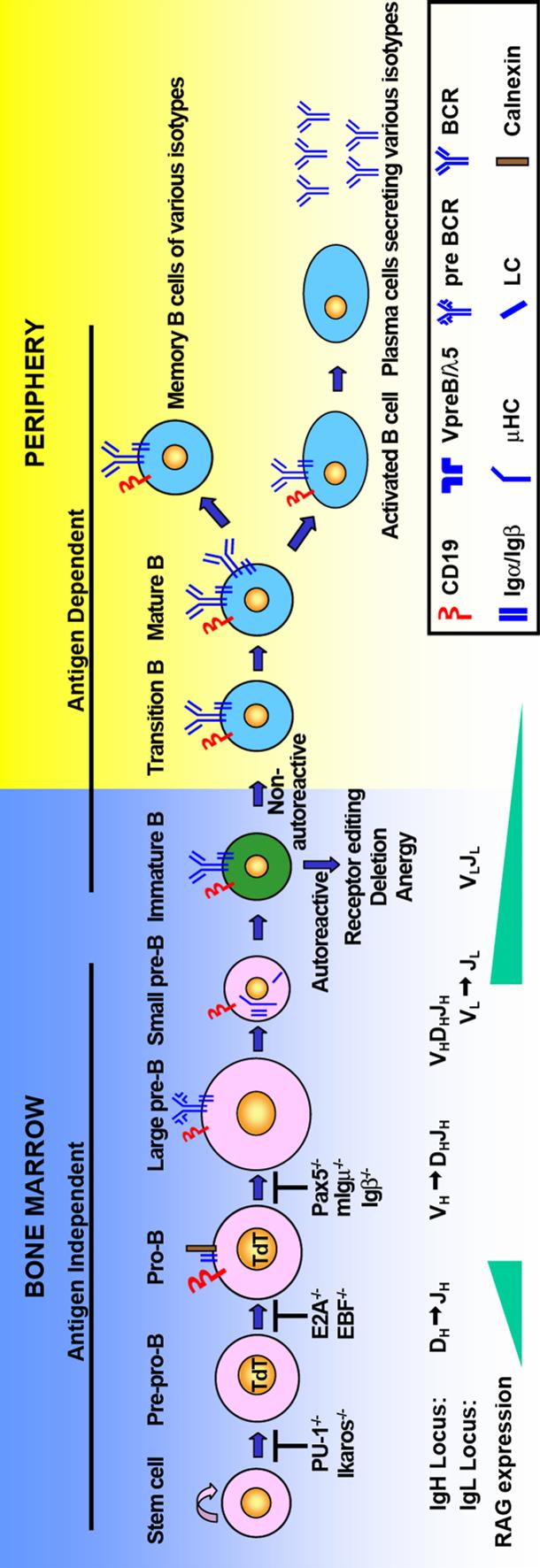
15 Figure1 3
16 Figure 1. B cell development in the bone marrow and periphery. B cell development begins from bone marrow stem cells. V(D)J recombination occurs in a stepwise manner. RAG gene expression initiates DJ H rearrangement in early lymphoid progenitor cell. Then the rearrangement proceeds to V H to DJ H accompanied with increased RAG expression. Successful expression of Igμ leads to the differentiation of pro-b cell. Functional Igμ rearrangement can pair with surrogate light chain VpreB and λ5. Igμ, surrogate light chain and Igα/Igβ heterodimer form pre-bcr. Expression of pre-bcr on the cell surface is critical for B cell differentiation. Pre-BCR expression downregulates RAG expression and pre-b cell starts to undergo proliferation. As the expression of surrogate light chain decreases, RAG gene expression is upregulated again and initiates Igκ transcription. Pre-B cell exits cell cycle and start differentiation. If the recombination on both Igκ alleles are nonfunctional, pre-b cells will initiate recombination on Igλ locus. Functionally expressed IgH and IgL chains pair with each other and form the B cell receptor (BCR). B cells with functional BCR differentiate into immature B cells. Immature B cell stage is important for selection of the B cell repertoire. Three mechanisms, namely receptor editing, clonal deletion and anergy, are employed to establish central tolerance. Immature B cells survival from negative selection acquires IgD expression and exit bone marrow as naïve B cells in the periphery. B cell maturation in the periphery is antigen-dependent. Upon encounter of antigen, mature B cells are activated to differentiate into antibody-secreting plasma cells and memory B cells. 4
17 Pre-B cell stage is marked by the expression of intracellular μhc and surface pre-bcr. The pre-bcr consist the μhc, Igα-Igβ heterodimer, and surrogate light chains. Surrogate light chain is composed of two proteins, V-like sequence (VpreB) and C-like sequence (λ5). VpreB and λ5 form light chain like structure with noncovalent bond. Signaling from the pre-bcr stimulates pre-b cell proliferate, differentiate, and initiate light chain recombination, although the expression level of pre-b cell receptor is very low. Pro-B cells with mutations in the signaling domain of Igμ or Igβ are completely blocked and cannot differentiate into pre-b cells (13-15). However, B cells with mutated Igα or surrogate light chains showed a leaky phenotype with only a partial block in allelic exclusion and B cell differentiation (16-18). After expression of functional Igμ, pre-b cells undergo proliferation and then are developmentally arrested. The second wave of RAG expression mediates light chain rearrangement (19, 20) and form functional κ or λ light chains, which pair with heavy chain and form B cell receptor to be expressed on the cell surface. Pre-BCR also important for allelic exclusion by down regulating RAG1/2 recombinases expression and changing the chromatin structure of V H locus, thus the accessibility of RAG1/2 to V H locus are much more restricted (10, 21). Pre-B cells differentiate into immature B cell in the bone marrow. Immature B cells are the first B lineage cells to express surface BCRs. B cell receptor composes of Ig heavy chains, Ig light chains and Igα-Igβ heterodimer. Terminal deoxyribonucleotidyl transferase (TdT) adds N-nucleotides to the D-J H and V H -DJ H junctions. Mice lacking 5
18 TdT activity resulted in a limited antibody repertoire compared with normal mice (22). Immature B cells express surface IgM but low level of IgD. Immature B cell is a very important developmental stage for negative selection in the bone marrow. B cells carrying self-reactive or non-functional BCRs undergo secondary Ig gene rearrangement on both heavy and light chain loci. B cells failed to edit themselves undergo clonal deletion or anergy (23). Immature B cells survived negative selection express both IgM and IgD on the cell surface, and differentiate into mature naïve B cells after exiting bone marrow. Naïve B cells mature in the spleen. Majority of them travel to primary follicular and mature as naïve follicular B cells. Small amount of naive B cells enter the marginal zone and mature as marginal zone B cells (24). Plasma cell is the terminal stage in B cell development functioning as antibody secreting cells. Two subsets of plasma cells are generated during immune responses, namely, long-lived plasma cells and short-lived plasmablasts (25). Long-lived plasma cells are generated in a T cell-dependent manner while plasmablasts are generated in T cell-independent manner. Long-lived plasma cells are produced in germinal center during immune reaction and migrate to bone marrow to produce antibodies for a prolonged period after the immune reaction. In contrast, plasmablasts still undergo proliferation and stay in the lymphoid tissues where they are produced. 6
19 Plasma cells can be generated by early immune response from B1 cells and marginal zone B cells. B1 B cells are CD5 positive and account for 5% of all B cells. B1 cells secret natural antibodies targeting common pathogens and are without further differentiation. The response of marginal zone B cells to foreign antigens is quick (24, 26). Mice without functional marginal zone B cells were vulnerable to bacteria infections (27). When circulating follicular B cells are challenged by foreign antigens, plasmablasts can form extrafollicular foci with the help of T cells and develop into antibody-secreting plasma cells. B1 cells, marginal zone B cells, and extrafollicular B cells all together are the first frontier of antibody protection. These antibodies secreting plasma cells are generated in early immune responses (usually the first week after antigen challenge). However, these plasma cells are not somatically mutated and undergo apoptosis within 3 days (28). Ten to fourteen days after antigen challenge, circulating follicular B cells can form foci in the secondary lymphoid organs, called germinal center. This process requires help of T cells (29, 30). Bcl6 is necessary for germinal center formation by promoting cell proliferation and inhibiting cell differentiation. Germinal center can be separated into light zone and dark zone by different activities. B cells in the light zone are called centroblasts, which expressing CXCR4; while B cells in the dark zone are called centrocytes, which expressing CXCR5. Germinal center B cells undergo proliferation 7
20 accompanying somatic hypermutation and class switch recombination (34). Germinal center is specialized for selecting plasma cells and memory B cells with high antigen affinity. Antigen-specific cells can exit germinal center while B cells carrying low antigen affinity BCRs fail the negative selection and undergo apoptosis. Memory B cells have high-affinity to antigen and they respond more rapidly comparing with naïve B cells in the circulation. Plasma cells and memory B cells proliferate dramatically on secondary encounter with the same antigen (31-33). Three transcription factors are important in the regulation of plasma cell development, namely, B lymphocyte-induced maturation protein 1 (Blimp1), interferon-regulatory factor 4 (IRF4), and X box-binding protein 1 (Xbp1). Blimp1 is the master regulator among these three factors. Blimp1 alone is sufficient for B cells to differentiate into plasma cells (35). However, initiation of plasma cell differentiation is independent of Blimp1 in that B cells deficient in Blimp1 still produce antibodies (125). Blimp1 and Bcl6 repress each other to balance germinal center formation and plasma cell differentiation (41, 42, 43, 44). Blimp1 induces IRF4 expression. IRF4 is important for plasma cell differentiation and is required for mature B cell proliferation after encountering foreign antigens (36). Xbp1 is the only transcription factor that is uniquely required for plasma cell differentiation, but not any other stages of B cells (37). Xbp1 is induced by Blimp1 and is responsible for the unfolded protein response, which enable plasma cells to produce, fold and secret large amount of protein (38). Blimp1 suppresses 8
21 the pax5 expression which is required for B cell commitment and maintenance until the plasma cell stage (39). Pax5 also represses Xbp1 expression (40). Plasma cells fail to induce Xbp1 expression undergo apoptosis due to the stress of unfolded protein response (126). The V(D)J recombination The variable region of Ig is generated by the recombination of variable (V), diversity (D) and joining (J) gene segments (11, 45, 46). The Ig heavy chain (IgH) variable region is composed of V, D, and J gene segments, while Igκ and Igλ light chain (IgL) variable regions are composed of V and J gene segments. Removal of D region in mice reduces the CDR3 diversity and results in a limited antibody repertoire (47). V(D)J recombination is a random process, which generates vast diversity of nascent B cell repertoire. V(D)J recombination is a site specific process that occurs between Ig segments flanked by conserved recombination signal sequences (48). RSS sites are composed of a conserved heptamer (5 -CACTGTG-3 ) and a nomamer (5 -ACAAAAACC-3 ) separated by nonconserved 12bp (12RSS) or 23bp (23RSS) spacers. Within the IgH locus, V H and J H genes are flanked by 23RSS and D H genes are flanked by 12RSS at both 5 and 3 ends. V(D)J recombination occurs preferentially between two gene segments flanked by different RSSs, referred as the 12/23 rule (11, 49). The V(D)J recombination and antibody gene assembly are mediated by RAG1 and RAG2 9
22 (50). Both RAG1 and RAG2 are absolutely required to initiate V(D)J recombination. RAG1 or RAG2 deficient mice ended up with a complete block in B cell development at the progenitor stage. The V(D)J recombination consists several steps. First, recombinase recognizes the RSS sites and brings two coding segments together. Then, RAG1/2 cut at the junction between signal end and coding end. Next, 3 -OH of the cut coding strand attacks the opposite DNA strand to form closed hairpin coding ends, leaving the signal end as double-stranded DNA break (51-53). Last, the hairpin is opened by Artemis endounclease to generate P-nucleotide at the coding end. Single-strand endonuclease trims a few nucleotides at the coding end. TdT adds up to fifteen N-nucleotides to junction region. TdT generates limitless diversification to the V(D)J junctions (54, 55). The two coding ends are ligated to form a new V(D)J joint. Ligation of the signaling ends forms an extrachromosomal circle released from the genome (56-58). Generation of autoreactive antibodies and negative selection The random V(D)J recombinant process can produce an almost infinitely diversified antibody repertoire. The disadvantage of this process is that majority of the antibodies are autoreactive. The frequency of autoreactive B cells in human bone marrow and peripheral blood B cells has been determined by single cell assay (59). The frequency of autoreactive B cells dropped from 75.9% to 43.1% from early immature B cells to immature B cells in human bone marrow; the frequency of autoreactive B cells dropped from 40.7% to 20.4% from new immigrant B cells to naïve B cells in human peripheral 10
23 blood (59). These results suggested two distinct checkpoints during early B cell development, one in bone marrow and the other in the periphery. These two checkpoints are also important for negative selection of polyreactive B cells. Studies of SLE (60) and RA (61) patients suggested that autoreactive B cells were failed to be removed from new immigrant B cells to mature naïve B cells in the periphery, which may cause the development of autoimmune disease. Random nature of V(D)J recombination generates nonfunctional rearrangement due to reading frame shift or induction of stop codon to the variable region. Because of the random joining process, theoretically only 1 out of 3 IgH rearrangements is functional. Three mechanisms, receptor editing, clonal deletion, and anergy, are employed to eliminate autoreactive B cells to establish central tolerance (62-65). About 10% of newly generated B cells can pass through all these selections and will be released into the periphery (17, 66, 67). Bone marrow immature B cell is a dedicated B cell stage for negative selection. Because immature B cells express RAG1/2 that allows secondary recombination, immature B cells are highly sensitive to apoptosis in response to strong BCR signaling (68-70). Receptor editing on light chain genes occurs through secondary rearrangement between an upstream V L gene and a downstream J L gene (64). The genetic organization of the κ and λ light chain genes allows multiple rearrangements. After the primary rearrangement, 11
24 upstream V L genes and downstream J L genes are still flanked by 12RSS and 23RSS which mediate successive rearrangement. V L to J L rearrangement starts from 5 end of the J L genes and extends to downstream genes. Since Igλ rearrangement always occurs after Igκ rearrangement, Igλ chain may serve as a salvage method for non-functional and autoreactive Igκ chain. After formation of a productive IgL chain, immature B cells terminate further rearrangement and continue B cell development (71). The evidence of light chain receptor editing came from two studies. In 1993, Tiegs et al built 3-83 antibody transgenic mice carrying anti-mhc (H2-K b and H2-K k ) reactivity (65). In these transgenic mice, B cells recognizing self antigen in the bone marrow showed an increased usage of endogenous light chains. RAG expression was elevated in bone marrow immature B cells of transgenic mice. Light chain receptor editing rescues autoreactive B cells from deletion. Another evidence of light chain receptor editing came from 3H9 transgenic mice built by Gay et al (64). 3H9 heavy and light chain combination recognizes double-strand DNA. The transgenic mice carrying both heavy and light chains from anti-dna antibody maintained almost normal B cell level in adult mice. All the transgenic heavy chains were paired with endogenous light chains instead of the transgenic light chain. The endogenous light chains that neutralized anti-dna activity showed more downstream Jκ usage and restricted V L gene repertoire. The frequency of receptor editing was measured in a mouse model with artificially 12
25 engineered κ alleles (72). One allele of the κ constant regions (mcκ) was replaced by the human κ constant region (hcκ). The usage of different constant regions can be measured by FACS analysis and mrna analysis. When the hcκ allele was combined with three different Vκ-Jκ joints, namely, 3-83κi, Vκ4R, and Vκ8R (71, 73), different Igκ knock-in mouse models showed that 25% of the peripheral B cells expressing the human Cκ. These results confirmed that light chain receptor editing plays an important role in both B cell development and antibody repertoire diversification. Another mechanism of bone marrow negative selection is clonal deletion. One of the major differences of bone marrow immature B cells from peripheral mature B cells is that immature B cells are susceptible to BCR induced deletion (23). The extent of B cell deletion depends on the strength of BCR signaling (74, 75). The third mechanism of bone marrow negative selection is anergy. Anergy is marked by the resistance of B cells to antigen stimulation. Anergy B cells usually have a short life span and impaired B cell differentiation (76, 77). The pioneer study on anergy was performed with different concentrations of anti-igm monoclonal antibody, E4. Murine bone marrow B cells underwent deletion when cultured with high concentrations of E4. However, low concentration of E4 antibody impaired both antibody production and B cell differentiation (78). The study about tolerance of peripheral B cells came from the Goodnow double-transgenic system (79). Double transgenic mice were generated from mice carrying HEL gene and anti-hel Ig gene. The number of mature B cells in the 13
26 spleen was reduced 2-4 fold in double transgenic mice. In the double transgenic mice, about half of the peripheral B cells have anti-hel reactivity. However, these highly selfreactive B cells remain functionally unresponsive. Very low level of anti-hel transgenic antibody was detected in the serum. V H replacement V H replacement occurs through RAG-mediated secondary rearrangement between an upstream unrearranged V H gene and a pre-formed V H DJ H joint (Figure 2). During primary V H DJ H rearrangement on heavy chain locus, RAG1/2 complex recognizes and cut at 23RSS (for V H and J H genes) and 12RSS (for D H genes). After the primary rearrangement, there is no 12RSS flanking D H genes any more, which makes the following rearrangement difficult or even impossible. The original evidence of V H replacement came from two independent studies on different cell lines (80, 81). One study is done on murine pre-b cell line derived from NIH/Swiss mice (81). Two clones were isolated from the cell line. Both of them carried V H DJ H - due to reading frame shift of V H and J H genes and failed to produce a μ-chain. However, after 2-4 weeks of culture, they started to become μ positive. Sequence analysis suggested that new V H DJ H genes were formed, while the identical D-N-J H rearrangement was kept. An upstream V H gene replaced the V H gene in the non-functional V H DJ H rearrangement to generate functional IgH genes. The same finding was observed in a murine leukemia cell line Ly1 + (80). V H gene replacement rescues non-functional V H DJ H rearrangement. 14
27 Sequence analysis suggests that V H replacement may be mediated by a heptamer-like sequence (5 -TACTGTG-3 ) embedded in the 3 end of V H coding sequence. This heptamer was identified as the cryptic RSS (crss). It differs from the heptamer from 12bp or 23bp RSS (5 -CACTGTG-3 ) by only one nucleotide (80). crss does not have an identifiable nonamer. crss is conserved among 91% of human V H genes and 70% of murine V H genes (80). This high conservation of crss suggests that V H replacement may play an important role during B cell development. V H replacement may occur with or without self-antigen stimuli to diversify B cell repertoire under normal conditions. The heptameric crss at the 3 end of V H gene presents not only in humans and mice, but also near the 3 end in other mammals, birds, and cartilaginous fish (82). V H replacement may be an important mechanism for B cell diversification and is positively selected during evolution. 15
 and crss (from the pre-formed V H DJ H joint, showed as grey triangle).")
28 Figure 2 Figure 2. Model of serial V H replacement. V H replacement occurs between a previous V H DJ H rearrangement and an upstream V H gene. V H replacement is mediated by 23bp RSS (from the invading V H gene, showed as white triangle) and crss (from the pre-formed V H DJ H joint, showed as grey triangle). RAG gene products can recognize and cleave at both sites. The DNA in between of 23bp RSS and crss is cut off and form an extrachromosomal circle. The upstream V H gene is ligated with the downstream genes to form a new V H DJ H joint. The previous V H gene is completely deleted except leaving a small stretch of DNA into the CDR3 region of new V H DJ H joint. This stretch of DNA is called V H replacement footprint. If serial of V H replacement occurs, the CDR3 region of the new IgH gene is extended. 16
29 Several mouse models were generated (Figure 3) to study the biological function of V H replacement. The 3H9 knock-in mouse model carries the V H DJ H joint with anti-dsdna activity, which is derived from an autoimmune-prone MRL/lpr mouse (83, 84). The IgH chain plays a dominant role in the antigen recognition and the anti-dsdna reactivity of 3H9 antibody can be neutralized by limited IgL chains (83). The 3H9 heavy chain has embedded crss within the 3 V H region. B cells with anti-dna reactivity were negatively selected in this transgenic mouse model. Three types of recombination on the 3H9 heavy chain were identified, namely, V H to V H DJ H, V H to D to V H DJ H, and D to V H DJ H. Among all the hybridomas isolated from transgenic mice, 25% of them were generated by V H to V H DJ H replacement. V H replacement in 3H9 mouse model occurred during the transition from pre-b cell to immature B cell in the bone marrow (83). 17


30 Figure 3 18
31 Figure 3. Biological significance of V H replacement comes from mouse models. (A) In 3H9 site-directed transgenic mouse model, the J H locus is replaced by 3H9 V H DJ H joint which encoding anti-dna reactivity. The other allele remains intact. Anti-DNA antibodies were negatively selected. Three types of rearrangements were detected: V H to V H DJ H, D H to V H DJ H, V H to D H to V H DJ H. (B) In the quasimonoclonal (QM) mouse, on one allele, the J H locus is replaced by V H DJ H joint encoding anti hapten (4-hydroxy-3-nitrophenyl) acetyl (NP) activity. J H locus was deleted for the other allele. QM mouse is kappa deficient. 20% of peripheral B cells lost anti-np reactivity through V H replacement. Upon virus infection, QM mouse also generates virus-specific neutralizing antibodies. (C) In this mouse model with nonfunctional IgH gene, D region and J H1, J H2 genes were replaced by a nonfunctional V H DJ H joint carrying stop codons at position 86 and 102. A diversified B cell repertoire was generated by V H replacement and directed V H to J H rearrangement. (D) Another mouse model carrying nonfunctional IgH gene. J H region was replaced by a nonfunctional V H DJ H joint carrying stop codon in the leader peptide. A vast number of B cells were generated by V H replacement. 19
32 The quasimonoclonal (QM) mouse model carries a functional rearranged segment on one IgH allele, and all J H segments on the other allele were deleted (85). The gene targeting V H DJ H rearrangement on the functional IgH allele was isolated from a hapten (4-hydroxy-3-nitrophenyl) acetyl (NP) specific hybridoma and recognized the NP when paired with Igλ chain. In QM mice, Igλ chain is the only type of IgL chains since Igκ chains on both alleles are nonfunctional. The primary repertoire of the QM mouse is genetically committed to be anti-np monospecific. However, 20% of peripheral B cells lost the anti-np reactivity despite of the genetic predisposition (86). These 20% of peripheral B cells produced over 90% of the serum antibody without NP binding specificity. These NP negative B cells were generated by V H to D to V H DJ H rearrangement and V H to V H DJ H rearrangement; also these B cells are class switched and somatic hypermutated. Because all other D genes were deleted during the primary V H DJ H recombination, the V H D to V H DJ H type of replacement could not happen under normal conditions. To study if V H replacement could generate antigen specific antibodies and further contribute to the immune response, QM mice were infected with vesicular stomatitis virus (VSV), lymphocytic choriomeningitis virus, or poliovirus mounted virus (87). Virus-specific neutralizing antibodies were produced by V H replacement in QM mice. Although the generation and expansion of V H replacement products may be due to the stimulation of viral antigens, VSV specific antibodies were isolated from naïve B cells of QM mice. This observation suggests that V H replacement not only mounts protective anti-viral response, but also diversifies the antibody repertoire. Sheep 20
33 erythrocytes (SRBC) were used to determine how QM mice respond to immunization (88). Similar levels of SRBC-reactive serum Ig were produced in QM mice compared with control. 50% of clones specific for SRBC were generated by V H replacement. Three clones with anti-srbc reactivity employing QM V H gene were paired with downstream λ1 gene, suggesting they have undergone extensive editing on λ light chain. Besides B cells with anti-np and anti-srbc reactivity, about 40% of B cells from the periphery carried other specificities. 88% of these B cells other specificities were generated by V H replacement. One mouse model was made by replacing the J H genes with a V H DJ H rearrangement carrying a stop codon in the leader peptide (89). In this mouse model all the pro-b cells produced are nonfunctional, so that the importance of Ig receptor signals in the induction of V H replacement can be studied. B cells developed beyond pro-b cell stage in bone marrow and spleen. The pro-b cells in the IgH VDJ-/VDJ- mice increased about 2-fold while pre-b and immature B cells from the bone marrow remain the same number compared with wild-type; on the other hand the mature B cells in the spleen reduced to 33% of normal level. Functional B lymphocytes were generated by two mechanisms, V H to V H DJ H rearrangement (5% of the sequences) and V H to D to V H DJ H rearrangement (95% of the sequences). The predominant incidence of V H to D to V H DJ H rearrangement could be due to the preference of RAG to recognize and cut between 12bp RSS and 23bp RSS. V H replacement can rescue nonfunctional pro-b cells and form a productive 21
34 rearrangement. V H replacement can occur in the absence of μhc signaling. V H replacement not only occurs in this abnormal transgenic mouse model with nonfunctional rearrangements, but also occurs in functional V H DJ H rearrangement transgenic mice (90). New V H DJ H rearrangement generated by V H replacement can be detected by single-cell PCR in pre-pro-b cells of functional V H DJ H rearrangement transgenic mice. This provides additional evidence that V H replacement occurs under physiological conditions and is independent of μhc expression. A mouse model with nonfunctional rearrangement was generated by deleting the D H locus and replacing the J H 1 and J H 2 genes with a V H 81X V H DJ H rearrangement. V H 81X V H DJ H joint is nonfunctional due to induction of two stop codons at position 86 and 102 (91). The homozygous D23 stop mutation generated about 20% of mature B cells compared with wild type mice. The numbers of immature B cells and mature B cells from bone marrow were substantially reduced compared with wild type mice. 55% of the Ig repertoire was generated by V H replacement, while 13% of the Ig repertoire was generated by direct V H to J H rearrangement. V H replacement is more efficient than V H to J H rearrangement in rescuing nonfunctional rearrangements. Since only one third V H replacement products have an identifiable V H replacement footprint, the actual frequency of V H replacement may be greatly underestimated by sequence analysis alone. In the D23 stop mice V H replacement preferentially added an arginine-encoding AGA codon to the new V H DJ H gene. Anti-DNA antibodies were produced in the serum of D23 stop mutant 22
35 mice, although at a lower level than autoimmune prone MRL/lrp mice. Both models demonstrate that V H replacement can rescue nonfunctional V H DJ H rearrangements from deletion. However, both models have limitations. First, in human bone marrow V H replacement can only be detected in immature B cells (92), while V H replacement can only be analyzed at pro-b cell stage in both mouse models. Second, both mouse models carry stop codon but not the condition of frame shift. The major problem in identifying V H replacement is that in wild type genome V H replacement products can be detected only by sequence analysis searching V H replacement footprints at the 3 end of V H genes downstream of crss. Human V H replacement footprints are shown in Table 1. Because V H replacement footprint is part of the most diversified CDR3 region, it is difficult to distinguish between V H replacement products and primary V H DJ H rearrangements. During the recombination and B cell differentiation, V H replacement footprints can be easily destroyed for many reasons. First, TdT adds nontemplate N-nucleotides to the V H -D and D-J H joint in early B cells. Second, the activity of exonuclease may chew-back the V H replacement footprint during the recombination. At last, Somatic hypermutation in activated B cells may destroy the footprints (54, 55). 23
36 Table 1 Unique motifs (V H replacement footprint) contributed by V H genes VH1 gene family VH3 gene family VH4 gene family VH1-18*01 TACTGTGCGAGAGA VH3-11*01 TACTGTGCGAGAGA VH4-28*01 TACTGTGCGAGAAA VH1-2*01 TACTGTGCGAGAGA VH3-11*03 TACTGTGCGAGA VH4-28*02 TACTGTGCGAGAAA VH1-2*02 TACTGTGCGAGAGA VH3-13*01 TACTGTGCAAGAGA VH4-28*03 TACTGTGCGAGAGA VH1-2*03 TACTGTGCGAGAGA VH3-13*02 TACTGTGCAAGAGA VH4-28*04 TACTGTGCGAGA VH1-2*04 TACTGTGCGAGA VH3-13*03 TACTGTGCAAGA VH4-30-2*01 TACTGTGCCAGAGA VH1-24*01 TACTGTGCAACAGA VH3-16*01 TACTGTGTGAGAAA VH4-30-2*03 TACTGTGCGAGACA VH1-3*01 TACTGTGCGAGAGA VH3-16*02 TACTGTGTGAGAAA VH4-30-2*04 TACTGTGCGAGAGA VH1-3*02 TACTGTGCGAGAGA VH3-20*01 CACTGTGCGAGAGA VH4-30-4*01 TACTGTGCCAGAGA VH1-45*01 TACTGTGCAAGANA VH3-21*01 TACTGTGCGAGAGA VH4-30-4*02 TACTGTGCCAGAGA VH1-45*02 TACTGTGCAAGA VH3-21*02 TACTGTGCGAGAGA VH4-30-4*05 TACTGTGCCAGAGA VH1-45*03 TACTGTGCAAGA VH3-23*01 TACTGTGCGAAAGA VH4-30-4*06 TACTGTGCCAGAGA VH1-46*01 TACTGTGCGAGAGA VH3-23*02 TACTGTGCGAAAGA VH4-31*01 TACTGTGCGAGAGA VH1-46*02 TACTGTGCGAGAGA VH3-23*03 TACTGTGCGAAA VH4-31*02 TACTGTGCGAGAGA VH1-46*03 TACTGTGCTAGAGA VH3-23*04 TACTGTGCGAAAGA VH4-31*03 TACTGTGCGAGAGA VH1-58*01 TACTGTGCGGCAGA VH3-23*05 TACTGTGCGAAA VH4-31*10 TACTGTGCGAGAGA VH1-58*02 TACTGTGCGGCAGA VH3-30*01 TACTGTGCGAGAGA VH4-34*01 TACTGTGCGAGAGG VH1-69*01 TACTGTGCGAGAGA VH3-30*02 TACTGTGCGAAAGA VH4-34*02 TACTGTGCGAGAGG VH1-69*02 TACTGTGCGAGA VH3-30*03 TACTGTGCGAGAGA VH4-34*04 TACTGTGCGAGAGG VH1-69*04 TACTGTGCGAGAGA VH3-30*04 TACTGTGCGAGAGA VH4-34*05 TACTGTGCGAGAGG VH1-69*05 TACTGTGCGAGA VH3-30*05 TACTGTGCGAGAGA VH4-34*09 TACTGTGCGAGAGA VH1-69*06 TACTGTGCGAGAGA VH3-30*06 TACTGTGCGAGAGA VH4-34*10 TACTGTGCGAGATA VH1-69*08 TACTGTGCGAGAGA VH3-30*07 TACTGTGCGAGAGA VH4-34*12 TACTGTGCGAGA VH1-69*09 TACTGTGCGAGAGA VH3-30*08 TACTGTGCGAGA VH4-34*13 TACTGTGCGAGAGG VH1-69*10 TACTGTGCGAGAGA VH3-30*09 TACTGTGCGAGAGA VH4-39*01 TACTGTGCGAGACA VH1-69*11 TACTGTGCGAGAGA VH3-30*10 TACTGTGCGAGAGA VH4-39*02 TACTGTGCGAGAGA VH1-69*12 TACTGTGCGAGAGA VH3-30*11 TACTGTGCGAGAGA VH4-39*06 TACTGTGCGAGAGA VH1-69*13 TACTGTGCGAGAGA VH3-30*12 TACTGTGCGAGAGA VH4-39*07 TACTGTGCGAGAGA VH1-8*01 TACTGTGCGAGAGG VH3-30*13 TACTGTGCGAGAGA VH4-4*02 TACTGTGCGAGAGA VH1-f*01 TACTGTGCAACA VH3-30*14 TACTGTGCGAGAGA VH4-4*06 TACTGTGCGAGAGA VH3-30*15 TACTGTGCGAGAGA VH4-4*07 TACTGTGCGAGAGA VH2 gene family VH3-30*16 TACTGTGCGAGAGA VH4-59*01 TACTGTGCGAGAGA VH2-26*01 TACTGTGCACGGATAC VH3-30*17 TACTGTGCGAGAGA VH4-59*02 TACTGTGCGAGAGA VH2-5*01 TACTGTGCACACAGACC VH3-30*18 TACTGTGCGAAAGA VH4-59*07 TACTGTGCGAGA VH2-5*04 TACTGTGTACGG VH3-30*19 TACTGTGCGAGAGA VH4-59*08 TACTGTGCGAGA VH2-5*05 TACTGTGCACACAGAC VH3-30-3*01 TACTGTGCGAGA VH4-59*09 TACTGTGCGAGAGG VH2-5*06 TACTGTGCACACAGA VH3-30-3*02 TACTGTGCGAAAGA VH4-59*10 TACTGTGCGAGATA VH2-5*08 TACTGTGCACACAGAC VH3-33*01 TACTGTGCGAGAGA VH4-61*01 TACTGTGCGAGAGA VH2-5*09 TACTGTGCACACAGAC VH3-33*02 TACTGTGCGAGAGA VH4-61*02 TACTGTGCGAGAGA 24
37 VH2-5*10 TACTGTGCACGG VH3-33*03 TACTGTGCGAAAGA VH4-61*03 TACTGTGCGAGAGA VH2-70*01 TACTGTGCACGGATAC VH3-33*04 TACTGTGCGAGAGA VH4-61*05 TACTGTGCGAGA VH2-70*09 TACTGTGTACGG VH3-33*05 TACTGTGCGAGAGA VH4-61*06 TACTGTGCCAGAGA VH2-70*10 TACTGTGCACGGATAC VH3-35*01 TACTGTGTGAGAAA VH4-61*07 TACTGTGCGAGACA VH2-70*11 TACTGTGCACGGATAC VH3-38*01 TACTGTGCCAGATATA VH4-61*08 TACTGTGCGAGAGA VH2-70*12 TACTGTGCACACAGAC VH3-38*02 TACTGTGCCAGATATA VH4-b*01 TACTGTGCGAGA VH3-43*01 TACTGTGCAAAAGATA VH4-b*02 TACTGTGCGAGA VH5 gene family VH3-43*02 TACTGTGCAAAA VH5-51*01 TACTGTGCGAGACA VH3-48*01 TACTGTGCGAGAGA VH5-51*02 TACTGTGCGAGACA VH3-48*02 TACTGTGCGAGAGA VH5-51*03 TACTGTGCGAGA VH3-48*03 TACTGTGCGAGA VH5-51*04 TACTGTGCGAGA VH3-53*01 TACTGTGCGAGAGA VH5-a*01 TACTGTGCGAGA VH3-53*02 TACTGTGCGAGA VH5-a*03 TACTGTGCGAGA VH3-53*03 TACTGTGCTAGGGA VH5-a*04 TACTGTGCGAGA VH3-64*01 TACTGTGCGAGAGA VH3-64*02 TACTGTGCGAGAGA VH6 gene family VH3-64*03 TACTGTGTGAAAGA VH6-1*01 TACTGTGCAAGAGA VH3-64*04 TACTGTGCGAGAGA VH6-1*02 TACTGTGCAAGAGA VH3-64*05 TACTGTGTGAAAGA VH3-66*01 TACTGTGCGAGAGA VH7 gene family VH3-66*02 TACTGTGCGAGA VH7-4-1*01 TACTGTGCGAGA VH3-66*03 TACTGTGCGAGAGA VH7-4-1*02 TACTGTGCGAGAGA VH3-66*04 TACTGTGCGAGACA VH7-81*01 TACTGTGCGAGATA VH3-7*01 TACTGTGCGAGAGA VH3-7*02 VH3-72*01 VH3-74*01 VH3-74*02 VH3-74*03 VH3-9*01 TACTGTGCGAGA TACTGTGCTAGAGA TACTGTGCAAGAGA TACTGTGCAAGA TACTGTGCAAGAGA TACTGTGCAAAAGATA 25
38 Table 1 DNA sequences show 3 end of germline functional V H genes. Black highlight indicates conserved cryptic RSS (crss). V H genes without crss and V H genes with less than 5 nucleotides at 3 end after crss are not shown here. Grey highlight indicates nucleotides can not be part of V H replacement footprint due to matches in the D genes. 5 -CACTGTG-3 in VH3-20*01 gene is identified to the heptamer from conventional RSS. The DNA fragment after crss is used as V H replacement footprint in the form of heptamer or hexamer. 26
39 The molecular mechanism of V H replacement comes from the study in human EU12 cells. EU12 cell line is a B cell line established from a childhood acute lymphoblastic leukemia (ALL) patient. EU12 cell repertoire is intraclonally diversified. EU12 cells undergo spontaneous differentiation from pro-b (μhc - ) to pre-b (CD34 - μhc -low SLC + ) and to immature B cells (CD34 - μhc high λhc + ) under normal tissue culture conditions. TdT is expressed in pro-b and pre-b cells, but not in immature B cells. Kappa locus is naturally deleted in EU12 cells. LM-PCR and excision circle PCR suggest that EU12 cells have ongoing V H replacement in the μhc + population. Sequence analysis suggests that EU12 clones have the same D 3-10 J H4 joint but employ different V H genes. EU12 cell line was transformed after DJ H rearrangement while recombinase is still expressed. Serial V H replacement products can be detected in this immortalized cell line. There is no N addition between D and J H gene segments. Four gene segments, V H2-5, V H3-7, V H3-11, and V H1-8, clustered in the D H proximal region of the heavy chain locus are used. The 3 V H gene used in EU12 μhc + cells is V H2-5, which is a nonfunctional rearrangement. The dominant clone within EU12 culture is V H3-7, which is adjacent to the V H2-5. The stimuli of V H replacement in EU12 cells are unclear. During the recombination between existing V H DJ H joint and an upstream V H gene, double-strand DNA break is temporally formed within the cell. Since this is only ectopically present within the cell, LM-PCR can be used to detect the ongoing V H replacement. V H3 gene specific double-strand DNA breaks can be detected only in 27
40 immature B cells from human bone marrow, which is the first evidence of occurrence of V H replacement in humans (92). Studies of EU12 cell line also suggest a method to identify potential V H replacement products by searching V H replacement footprints, which is the 3 end of V H germline genes (Table 1). Five nucleotides (pentamer) complete match or six nucleotides (hexamer) with one mismatch is accepted as V H replacement footprints. Of 300 IgH sequences from human peripheral blood, 5% of them are identified as V H replacement products containing at least one pentamer or hexamer (92). Using the same method, 10% of the 15,000 IgH sequences in IMGT data base are identified as V H replacement products (93). These results suggested that V H replacement contributes significantly to the diversity of human B cell repertoire. When the amino acids encoded by V H replacement footprint are analyzed, 80% of them encode charged amino acid regardless of the reading frames (92), majority of them are positively charged amino acids (94, 95). As shown in the EU12 model, V H replacement adds V H replacement footprints to the V H -D junction of the newly formed V H DJ H joint and extends the CDR3 region. Charged amino acids and extended CDR3 regions attributed by V H replacement are common features of autoreactive antibodies. These observations suggest the potential role of V H replacement in autoimmune diseases. IgH sequences from different autoimmune diseases were analyzed for V H replacement 28
41 frequency (Figure 4). In rheumatoid arthritis, the frequency of V H replacement is 30%; in Sjögren s syndrome s, the frequency is 22%. In SLE plasma cells (CD19 low CD27 high ), which associate with disease severity and anti-dna antibody production, frequency of V H replacement is 48%. The frequencies of V H replacement in these diseases are significantly higher than in healthy controls (93). Majority of V H replacement footprints encode Arginine in CDR3 regions. IgH sequences from anti-viral and anti-bacteria infection were also analyzed. The frequency of V H replacement is 33% among anti-hiv antibodies and 23% among IgH genes of mucosal plasma cells from HIV patients (93). The frequency of V H replacement is also elevated in hepatitis C virus disease which is 38% (93). In summary, V H replacement not only generates self-reactive antibodies, but also, in consensus with QM mouse study, involved in anti-viral responses. 29


42 Figure 4. Potential contribution of V H replacement products to autoimmune diseases and anti-viral responses. 30
43 Systemic lupus erythematosus (SLE) SLE is an autoimmune disease characterized by presence of autoantibodies against a vast variety of self antigens, including single-strand and double-strand DNA, histone, red blood cells, platelets, leukocytes, and clotting factors. Symptoms including fever, fatigue (50-100% of the patients), skin rashes (75% of the patients), and kidney dysfunction (16-38% of the patients) occur in the immune reactions between the autoantibodies and self antigens. The prevalence of SLE varies geographically. In the western countries about 0.04% of the population has SLE (96). Eleven criteria have been set by the American College of Rheumatology for SLE disease classification, namely, malar rash, discoid rash, photosensitivity, oral ulcers, nonerosive arthritis, pleuritis or pericarditis, renal disorder, neurologic disorder, hematologic disorder, immunologic disorder (positive for anti-dna or anti-sm or anti-phospholipid antibodies), and positive antinuclear antibody. For the purpose of identifying patients in clinic, a person shall be diagnosed to have systemic lupus erythematosus if any 4 or more of the 11 criteria are present, serially or simultaneously, during any interval of observation. 95% of the SLE patients are positive for anti-nuclear staining assay. Although he remainder is negative for anti-nuclear assay, they are positive for anti-ro 60 (SS-A). Thus, people negative for anti-nuclear staining assay or anti-ro60 are free of SLE (97). 31
44 Several factors may cause SLE. Sex hormone is believed to induce SLE based on the following observations. Around 90% of the SLE patients are female, resulting in the 9 to 1 of female to male patient ratio (98). The female SLE patients are usually at childbearing age (99). Pregnancy is an important risk factor to exacerbate the disease in SLE patients, especially those with nephritis or hypertension. All these facts suggest that female hormone is a potential factor to induce SLE while male hormone may be protective (99). Genetic background is a second determinant. In western countries the incidence of SLE in African-Americans is three times higher than that in the European-Americans (100). 25% of monozygotic twins are affected by SLE together while only 2% of dizygotic twins are affected together (101). The candidate genes involved in SLE pathogenesis include major histocompatibility complex (MHC), complement components (C1q, C2, or C4), C-reactive protein, IgG Fc receptor, OLF/EBF-associated zinc finger protein, poly-adp-ribose polymerase, programmed cell death 1, toll-like receptor 5, and tumor necrosis factor α ( ). Drugs including procainamide, hydralazine, and quinidine can induce SLE in humans (105). Epstein-Barr virus (EBV) infection is a forth factor to induce SLE. Since there is temporal connections between EBV infections and SLE onset, EBV may be important to induce SLE. Importantly, 99% of SLE patients have EBV antibodies and all SLE patients have EBV DNA, which are much higher than the frequencies in healthy controls, indicating an intrinsic connection between EBV infection and SLE may exist (106). Environmental factors like UV exposure can induce SLE. UV denatured DNA can pass through the basement membrane and go into the circulation 32
45 serving as self antigens. Autoreactive antibodies cause severe consequences in SLE disease. Pathogenic autoreactive antibodies in SLE patients include anti-double-strand DNA antibodies, nucleosome antibodies, C1q antibodies, N-methyl- D -aspartate receptor antibodies, Ro antibodies, phospholipids antibodies, Sm antibodies, La antibodies, and α-actin antibodies (18, ). Antibodies of cardiolipin, antinuclear antibodies, Ro, La, and SM in SLE patients may be secreted by long-lived plasma cells while anti-dna antibodies may be produced by short-lived plasma cells, based on the plasma antibody changes after anti-proliferative reagents treatment ( ). Autoreactive antibodies bind self antigens and form immune complexes, which is deposited on the small blood vessels. The immune complexes activate complement system and generate complement complexes that will damage the blood vessel and cause vasculitis and glomerulonephritis. Anti-red blood cell and anti-platelet antibodies can cause intramedullary lysis and result in hemolytic anemia and thrombocytopenia. Anti-double strand DNA antibodies are specific for SLE, presenting in 70% SLE patients while only less than half percent in other autoimmune diseases like RA. This provides a means for the diagnosis of SLE (114). Double strand DNA is released by apoptotic cells in the form of nucleosome. The serum level of anti-double strand DNA antibodies often reflects the severity of the disease (115). Anti-double strand DNA antibodies bind 33
46 nucleosomes to form immune complexes that deposit in the renal glomerular basement membrane. These antibody-antigen complexes activate complement system and cause glomerulonephritis (116). The anti-dna and anti-nucleosome antibodies can cross react with proteins in kidney as well and cause direct damage of renal cells. This may apply to other polyreactive antibodies that recognize antigens sharing similar surface structure with renal cells. Sm antigens include seven proteins (B/B, D1, D2, D3, E, F, G) that are part of the common core of U1, U2, U4 and U5 small nuclear ribouncleoprotein (snrnp). B/B, D1, and D3 are major antigens among these proteins. Anti-SmD antibodies are specific to SLE and anti-smd1 antibody levels correlate to lupus nephritis but not to the disease activity like anti-double strand DNA antibodies ( ). Corticosteroid, rituximab, and abetimus sodium are well studied drugs for SLE treatment. Corticosteroids were used to reduce the level of anti-double strand DNA antibodies with adverse side effects (120, 121). Rituximab is an antibody that is able to deplete CD20 expressing mature B cells. However, rituximab can not deplete antibody-secreting plasma cells, which do not have CD20 expression. Abetimus sodium, designed to deplete anti-double strand DNA antibody producing B cells, can engage the B cells by its four surface oligonucleotides. However, abetimus sodium has no epitopes to bind helper T cells (122). B cells arrested by abetimus sodium undergo apoptosis or are cleared from the circulation along with the drug. The exact mechanism of abetimus sodium in human patients needs to be exploited (123, 124). 34
47 EDITING OF IGH GENE ENCODING ANTI-SM ANTIBODY BY VH REPLACEMENT By RUN FAN, JING LIU, ZHIHONG YU, AND ZHIXIN ZHANG Manuscript in preparation for submission Format adapted for dissertation 35
48 ABSTRACT EU12 cells undergo spontaneous V H replacement and generate new IgH genes even after expression of functional B cell receptors on cell surface. To explore the molecular mechanism responsible for V H replacement recombination in EU12 cells, we cloned one of the dominantly expressed IgH genes, A19H, from the EU12 μhc + cells. We found that the A19H gene encodes anti-sm antigen antibodies when paired with two endogenous Igλ chains. When the A19H gene was artificially paired with a panel of 14 randomly selected light chains from healthy controls, 10 out of the 14 antibodies still reacted with the SM antigens; 5 of them even had higher affinities for SM antigens than the original antibody. Treatment of EU12 μhc + cells with purified bovine SM antigen strongly induced V H replacement, and SM antigen can be detected in the EU12 culture supernatant. Moreover, recombinant antibodies generated from the A21H gene, a V H replacement product, attenuate anti-sm reactivity. These results indicate that the SM reactivity is mainly contributed by the A19H IgH gene in the EU12 μhc + cells, which could not be effectively neutralized by light chain editing. V H replacement provides a unique mechanism to edit IgH genes encoding anti-sm antibodies. 36
49 INTRODUCTION EU12 cell line was established from bone marrow cells of pediatric lymphoblastic leukemia patients (1-3). Phenotypic analysis showed that EU12 cell line is a clonal B cell line containing three subpopulations along B cell differentiation path, including pro-b (CD34 + μhc - ), pre-b (CD34 - μhc - ), and B (CD34 - μhc + ) cell populations (4). EU12 cells undergo spontaneous differentiation from pro-b cell to pre-b cell and to B cells continually, which makes it an ideal model system for human B cell development study (5). During early B lineage cell development, an unlimited antibody repertoire is generated by the random V(D)J recombination. This V(D)J recombination process is catalyzed by the recombination activating gene products, RAG1 and RAG2, which recognize the recombination signal sequences (RSS) flanking each V, D, and J gene segments (8-10). The random V(D)J recombination also produces nonfunctional immunoglobulin genes or immunoglobulin genes encoding autoreactive antibodies, both of which need to be deleted from the B cell repertoire (7). Three mechanisms are employed to eliminate B cells producing autoreactive antibodies, namely receptor editing, deletion, and anergy (11, 12). Most previous studies of receptor editing have focused on the light chain genes (13-15). The organization of the V L and J L gene segments within the Igκ and Igλ loci allows continuous rearrangements by joining an upstream V L and a downstream J L gene segment through the 12/23 RSS sites. After each rearrangement, a new V L -J L joint is 37
50 formed and the previous V L -J L joint is deleted. In humans, B cells expressing Igλ have recombined Igκ genes, but B cells expressing Igκ only occasionally carry recombined Igλ genes, suggesting that light chain gene recombination occurs on Igκ alleles first then precedes to Igλ alleles (16 18). A cell with nonfunctional κ rearrangements on both Igκ alleles has the chance to rearrange its Igλ genes (13-15). Counting all the usable Jκ and Jλ genes in mice or humans, a B cell has up to 14 and 16 chances to edit its light chain genes under ideal conditions. Using a mouse carrying the knock-in human Cκ region on one of the Igκ alleles, it has been estimated that Igκ editing occurs in more than 25% of peripheral B cells. Up to 70% immature B cells change their light chain genes when challenged by anti-bcr antibodies. In humans, although the frequency of light chain editing cannot be directly measured, the occurrence of IgL editing can be estimated indirectly by the elevated usage of downstream Jκ or Jλ genes. The evidence of deletion came from in vitro culture of isolated immature B cells. When immature B cells were treated with anti-bcr antibodies, they underwent apoptosis within 16 hours (19). Immature B cells bearing a transgenic immunoglobulin receptor that encounter soluble antigen have been shown to enter a short-lived anergic state in which the cells are refractory to further antigen stimulation (20). V H replacement occurs through RAG-mediated secondary recombination between a cryptic RSS (crss) from a previously rearranged V H gene and a 23RSS from an upstream V H gene. In different mouse models, V H replacement can efficiently delete 38
51 knock-in IgH genes encoding anti-dna antibodies in 3H9 mouse model (21, 22) or antibodies against the hapten (4-hydroxy-3-nitrophenyl) acetyl (NP) in QM mouse model (23). V H replacement can also rescue B cells carrying two non-functional IgH alleles to generate almost normal numbers of mature B cells and a diversified antibody repertoire (24, 25). V H replacement not only diversifies the B cell repertoire, it also produces neutralization antibodies upon virus infection in the QM mouse model. In human, evidence of V H replacement has been found in the EU12 cells and in bone marrow immature B cells. By searching for the potential V H replacement footprints within the V H -D H junction of IgH genes obtained from IMGT database, 5% of the normal IgH sequences obtained from human peripheral blood B cells were found as potential V H replacement products (5), suggesting a significant contribution of V H replacement to the B cell repertoire in human. EU12 cells differentiate continuously from pro-b to pre-b and to B cells with ongoing serial V H replacement. Detailed sequence analysis of the IgH genes derived from the EU12 μhc + cells indicates that the first round of V H replacement deletes the V H2-5 D H3-10 J H4 non-functional rearrangement to generate functional IgH genes. However, V H replacement continuously occurs in the EU12 μhc + cells even after expression of functional BCRs. We speculate that the BCR expressed on the EU12 μhc + cells might react with certain antigens in the culture system and stimulate continuous V H replacement. Indeed, we found that one of the IgH genes expressed in majority of the EU12 μhc + cells, 39
52 A19H, encodes autoantibodies for SM antigens. Moreover, treatment of EU12 μhc + cells with purified bovine SM antigens enhances V H replacement. The anti-sm reactivity contributed by the A19H cannot be efficiently neutralized using a panel of 10 Igκ and 4 Igλ chains derived from naïve B cells of a healthy donor, but can be neutralized by changing the V H gene. Based on these results, we concluded that V H replacement offers a unique mechanism to delete IgH genes encoding anti-sm antibodies. 40
53 MATERIALS and METHODS Cell Culture and Treatment EU12 cells were cultured in RPMI 1640 Medium supplemented with 100 U/ml penicillin, 100 μg/ml streptomycin, 10% heat-inactivated fetal bovine serum, 2 mm L-Glutamine (GIBCO, Carlsbad, CA) and 25μM β-mercaptoethanol (Sigma, Saint Louis, MO). Viable cells were incubated with PE conjugated anti-cd34 monoclonal antibodies (Beckman) and Cy5 conjugated donkey F(ab ) 2 anti-μhc antibodies (Jackson Laboratory) for FACS. Subpopulations of the EU12 cells were purified by two rounds of sorting using a MoFlow (Cytomation, Fort Collin, CO) cell sorter. For SM-antigen treatment, EU12 μhc + cells were seeded in 6-well plates at cells per well in 1 ml complete RPMI1640 medium with or without the addition of 3μg/ml of purified calf thymus SM antigen (Immunovision, Springdale, AR). Cells were harvested after 24 hours to extract genomic DNA and cellular DNA for analysis of ongoing V H replacement. Antibody Production and Purification Human embryonic kidney fibroblast cell line 293H were cultured in DMEM supplemented with 10% ultra-low IgG FBS, 100 U/ml penicillin, 100μg/ml streptomycin (GIBCO, Carlsbad, CA) and 20 mm HEPES (Mediatech, Manassas, VA). Plasmid DNA encoding IgH and IgL chains (10μg) were co-tranfected by PEI reagent (Sigma, 41
54 Saint Louis, MO). Cells were washed with serum-free DMEM 24 hour after transfection and cultured in DMEM supplemented with 10% ultra-low IgG FBS (GIBCO, Carlsbad, CA). Cell culture supernatant containing antibodies was collected after 7 days of culture. For HEp-2 immunofluorescent assays, antibodies were purified on protein A Sepharose (Amersham Pharmacia Biosciences, Piscataway, NJ). Antibody expression was confirmed by Western Blot and ELISA assay using goat anti-human IgG antibodies (Jackson ImmunoResearch, West Grove, PA). Anti-Nuclear Antigen Immunofluorescence Assay HEp-2 cell coated slides (Bion Enterprises) were incubated with purified antibodies at μg/ml for 30 min at room temperature. After washing with PBS, bound antibodies were detected using FITC conjugated anti-human IgG antibodies. Slides were viewed and pictures were taken under fluorescent microscopy (Olympus). Ligation-Mediated PCR LM-PCR was performed as described previously (26, 27). Briefly, purified genomic DNA (0.1 μg) was ligated with double-strand DNA linkers at 14 o C overnight. Ligated DNA samples were subjected to two rounds of nested PCR amplifications with V H3 gene specific primers. The 1 st round PCR was performed under the condition 94 o C, 3 min; 94 o C, 30 sec; 58 o C, 30 sec; 72 o C, 30 sec for 30 cycles, 72 o C, 1 min and stopped at 4 o C. The 2 nd round PCR was performed under the condition 94 o C, 3 min; 94 o C, 30 sec; 58 o C, 42
55 30 sec, 72 o C, 30 sec for 30 cycles, 72 o C, 1 min and stopped at 4 o C. HotMaster Taq DNA Polymerase (Eppendorf, Germany) was used for both rounds of PCR. PCR product (10μl) was separated on 2% agarose gel and visualized under UV light after EtBr staining. The PCR products were cloned and sequenced to confirm that the double-strand linkers were directly ligated to the heptameric sequence of the V H3 crss site. Detection of V H Replacement Excision Circles Excision circles derived from V H replacement recombination was detected as described previously (5, 15, 28). Briefly, cellular DNA was prepared from untreated or treated EU12 μhc + cells. Two rounds of nested-primer PCR were performed to detect the V H3 to V H1 replacement excision circles. The sense primers were derived from the V H3 coding region. The anti-sense primer was derived from the spacer regions of the 23RSS site flanking the V H1-8 gene. The 1 st round PCR was performed under the condition 94 o C, 3 min; 94 o C, 30 sec; 58 o C, 30 sec; 72 o C, 30 sec for 30 cycles, 72 o C, 1 min and stopped at 4 o C. The 2 nd round PCR was performed under the condition 94 o C, 3 min; 94 o C, 30 sec; 58 o C, 30 sec, 72 o C, 30 sec for 30 cycles, 72 o C, 1 min and stopped at 4 o C. HotMaster Taq DNA Polymerase (Eppendorf, Germany) was used for both rounds of PCR amplification. ELISA Antibody concentrations in cell culture supernatants were determined by ELISA using human IgG1κ (Sigma, Saint Louis, MO) as a standard. The capture antibody is purified 43
56 mouse anti-human IgG1 (BD, San Jose, CA). To measure reactivity against SM antigen, microplates (Corning, Pittsburgh, PA) were coated with 5 μg/ml of purified SM antigen (Immunovision, Springdale, AR). Tissue culture supernatants were used at 10 μg/ml antibody concentrations and serial 1:2 dilutions in PBS with 1% BSA. Culture medium only without recombinant antibodies was used as negative control. Wells adding secondary antibody only were served as blank. Bound antibodies were detected with peroxidase-conjugated goat anti-human IgG antibodies (Jackson ImmunoResearch, West Grove, PA) and OPD substrate (Sigma, Saint Louis, MO). OD490nm was measured using a microplate reader (BMG). Single-Cell Reverse-Transcription PCR Purified blood samples were collected with IRB-approved protocols at the University of Alabama at Birmingham. Different subsets of B cells were purified as CD19 low CD27 high plasmablasts and CD19 + CD27 - naïve B cells from active SLE patients or healthy donors. Sorted cells were limiting diluted into 96-well PCR plates with one cell per well in 5 μl PBS with 1U RNasin (Promega, Madison, WI). All samples were stored at -70 C. IgH and IgL chain gene rearrangements were amplified in two rounds of PCR (50 cycles each) and subcloned into corresponding Igγ1, Igκ or Igλ expression vectors (gift of M.C. Nussenzweig, Rockefeller University, NYC). Single-cell RT PCR were performed as described previously (10). 44
57 Analysis of IgH Gene Sequences Immunoglobulin genes were analyzed using the IgBLAST program to assign the potential germline V, D, J genes. Mutation and junction analyses were conducted using the JOINSOLVER and IMGT/V-QUEST programs. Isoelectric points of CDRs were calculated based on CDR1, CDR2, and CDR3 amino acid sequences. 45
58 RESULTS BCR expressed on the EU12 cells recognizes SM antigens Analysis of the BCR repertoire in EU12 μhc + cells indicated limited intraclonal diversification of V H and V L gene usage. All the IgH sequences included the same D H3-10 J H4 joint, but different V H genes. Majority of the EU12 μhc + cells express the A19H gene containing a V H3-7 D H3-10 J H4 rearrangement (4). We cloned A19H gene V region into expression vector containing an immunoglobulin constant region. To produce functional antibodies the A19H heavy chain was paired with two endogenous Igλ chains, A15L or A18L, in a 293H cell based transient transfection system. The expression and secretion of recombinant antibodies with heavy and light chains was confirmed by western blot using anti-human IgG antibodies (Figure1A). The recombinant antibodies were purified and used to blot EU12 cell lysates. Both recombinant antibodies recognized multiple proteins from EU12 cell nuclear extracts (Figure1B), and weakly reacted with several peptides in the cytosolic extracts as well (Figure1C). The autoreactivities were further confirmed by immunofluorescent ANA assay using HEp-2 cell coated slides (Bion Enterprises, Ltd., IL). Both recombinant antibodies stain the nuclear or cytoplasmic antigens of HEp-2 cells as shown in Figure 1D. These results demonstrate that BCR expressed on the EU12 cells are autoreactive. The protein bands detected by the recombinant antibodies derived from EU12 cells range from 20 to 60kDa, which have similar running patterns as SM antigen(s) (29). SM 46
59 antigen is a non-histone nuclear protein complex composed of 6 small nuclear ribonucleoproteins (snrnp) (30). SM antibodies bind B /B (27kDa/26kDa), D (13kDa) and sometimes E (12kDa) and rarely F (11kDa) and G (9kDa) polypeptides that comprise the SM protein complex (31, 32). Purified SM antigens from calf thymus contain many peptides, which can be seen on 10% SDS-PAGE by coomassie blue staining (Figure 2A). Among all the peptides with SM antigen complex, the principle reactivity has been shown to reside in the B(26kDa), B (27kDa), and D(13kD) polypeptides. Next, we tested if the two recombinant antibodies derived from EU12 cells recognize purified bovine SM antigens. The two recombinant antibodies, A19H/A15L and A19H/A18L, recognized one of the major bands at 26kDa in the purified SM antigens, which can be recognized by the human anti-sm antibody (Immunovision, Springdale, AR) as well (Figure 2B). The true nature of SM antigens is the RNA splicing complex, which can be easily found in EU12 cell lysate as well as EU12 culture supernatant (Figure 2C and 2D) by human anti-sm antibody but not fresh culture medium. Thus SM antigen in the EU12 cell culture was presumably released from dead cells. These results indicate that SM antigens existing in the EU12 culture might be the initial stimuli for ongoing V H replacement targeting the functional A19H gene. Changing Light Chains cannot Neutralize the Anti-SM Reactivities Contributed by the A19H Heavy Chain Light chain receptor editing is an important mechanism for editing of autoreactive B 47
60 cells. EU12 cells have lost both Igκ alleles and can express Igλ chains only (4). When the A19H gene was paired with two endogenous Igλ chains from EU12 cells, both recombinant antibodies were SM reactive. It is possible that different light chains, especially κ chains, can neutralize the SM reactivity contributed by A19H gene. To test this possibility, we randomly picked 14 light chains from naïve B cells of one healthy donor. Ten of them are κ light chains and 4 of them are λ light chains (Table 1). All 14 light chain genes were cloned into Igκ or Igλ expression vectors. We paired these 14 light chains with A19H heavy chain and co-transfected into 293H cells to express recombinant antibodies. Antibody concentrations were measured by ELISA. Reactivity against purified SM antigen was tested by western blot, with recombinant antibody concentration of 10μg/ml. The two endogenous antibodies from EU12 cells were also included to compare changes in anti-sm reactivity. Ten of the 14 randomly paired recombinant antibodies cannot silence anti-sm reactivity; five of them have higher anti-sm reactivity. Only four recombinant antibodies successfully silenced the anti-sm reactivity (Figure 3A). Thus, 71% of randomly picked light chains cannot efficiently neutralize the anti-sm reactivity contributed by the A19H heavy chain. This result is confirmed by ELISA assay for anti-sm antigen reactivity (Figure 3B). ELISA plates were coated with 5μg/ml purified SM antigen. Fourteen recombinant antibodies together with the two endogenous antibodies from EU12 μhc + cells were used as primary antibodies at the concentration of 25μg/ml. Results showed only four of the 14 recombinant antibodies were negative to SM antigen. The others 10 antibodies showed either the same reactivity as antibodies 48
61 using endogenous light chains or higher reactivity to SM antigen. ELISA data confirmed that the anti-sm reactivity contributed by the A19H heavy chain cannot be efficiently neutralized by using different light chains. A19H heavy chain is the dominant factor for SM antigen recognition. Treatment of EU12 μhc + cells with purified bovine SM antigens enhances V H replacement EU12 cells differentiate continually with ongoing V H replacement even after formation of functional BCR. The occurrence of V H replacement in mouse models carrying knock-in IgH transgene encoding anti-dna antibodies or IgH gene encoding anti-np antibodies suggested that V H replacement could be induced upon binding of self-antigens (22, 40). We speculate that V H replacement in EU12 cells may be a result of SM-antigen stimulation in the cell culture. To test the effect of SM antigens on V H replacement in EU12 cells, we treated EU12 μhc + cells with purified bovine SM antigen. EU12 μhc + cells were seeded in six-well plate at cell/ml in complete growth media with or without purified SM antigen at concentration of 1μg/ml and 2μg/ml. Three days later, genomic DNA was harvested and analyzed by LM-PCR to detect double-strand DNA breaks at the V H3 crss border (26, 27). Stronger LM-PCR product was obtained from genomic DNA of SM antigen treated EU12 μhc + cells in comparison with untreated cells (Figure 4B). Following RAG-mediated cleavage at the 23bp V H1-8 RSS and V H3-7 crss, the intervening DNA fragment will be ligated to form an excision DNA circle and 49
62 released from the genome (15, 28). Using cellular DNA samples from SM antigen treated or untreated EU12 μhc + cells, we found that the V H3 to V H1 excision circles were produced more in the SM antigen treated EU12 μhc + cells in comparison with untreated cells (Figure 4C). These results showed that treatment of EU12 μhc + cells with purified bovine SM antigen induced V H replacement and suggested that SM antigen in the EU12 cell culture was the initial stimulus for the ongoing V H replacement. V H replacement product significantly reduced the anti-sm reactivity V H replacement product A21H was cloned from EU12 μhc + cells. A21H is using V H1-8 gene which is 5 adjacent gene of V H3-7 gene as used by A19H, while A21H is still keeping identical D-J H joint as A19H (Figure 5A). A21H gene was paired with two endogenous light chains (A15L and A18L) to express recombinant antibodies. Anti-SM reactivity was tested by ELISA assay at the antibody concentration of 25μg/ml. The average anti-sm reactivity for A19H/A15L was at OD490nm, while reactivity for A21H/A15L was at OD490nm (p<0.0001). The average anti-sm reactivity for A19H/A18L was at OD490nm, while the reactivity for A21H/A19L was at OD490nm (p=0.0341). Each experiment was repeated four times or more. Anti-SM reactivity was significantly reduced when paired with A21H compared with A19H for both lambda light chains (Figure 5B). Light Chain Editing cannot Silence Anti-SM Antibodies in SLE 50
63 Systemic lupus erythematosus (SLE) is a complicated autoimmune disorder characterized by production of pathogenic autoantibodies against nuclear antigens, including DNA, histones, and ribonucleic proteins (33). The factors that lead to a breakdown in B cell tolerance to these antigens remain elusive (34). Anti-SM antibodies are very rare in normal individuals but are found to occur about 1.5 years before diagnosis in SLE patients. It has been established that anti-sm antibodies are present in approximately 25% of the sera of SLE patients but absent in sera of patients with other rheumatic diseases. Presence of anti-sm antibody is a diagnostic criterion and a unique feature of SLE disease (33, 35). Apoptotic cell antigens are thought to be a source of self antigen in SLE. It was reported that SLE patients are related with deficiencies in the clearance of ribonucleoprotein SM from apoptotic cells which lead to the production and activation of anti-sm antibodies (36). In systemic lupus erythematosus patients, it has been shown that a subgroup of plasmablasts (CD19 low CD27 high ) in the peripheral blood correlates with the disease activity and is responsible for the production of autoantibodies (37). Analysis of the IgH sequences obtained from this subgroup of plasmablasts revealed a significantly higher frequency of V H replacement products (48%), indicating that V H replacement may contribute to the generation of autoantibodies in SLE patients (38). Similar to EU12 μhc + cells, peripheral blood B cells from SLE patients have anti-sm reactivity, elevated levels of V H replacement, and have encountered and been challenged by SM antigens released from apoptotic cells. The EU12 cells as a human model system 51
64 is useful to study how V H replacement edits IgH genes encoding anti-sm antibodies, which is helpful to understand the generation of anti-sm antibodies and disease pathogenesis in SLE patients. To obtain monoclonal recombinant antibodies from SLE patients, single-cell RT-PCR was used to amplify naturally paired IgH and IgL genes from single peripheral blood B cell of active SLE patients. Immunoglobulin variable regions of IgH and IgL genes were amplified and cloned into the Igγ1, Igκ or Igλ expression vectors as in EU12 assay. The original heavy chain and the light chain from one cell were co-transfected into 293H cells to produce functional antibody. ELISA assay was performed to screen for antibodies with anti-sm reactivity. We screened 12 recombinant antibodies from two SLE patients. Two antibodies, VH3#06 (from SLE patient S1212) (Figure 6A), VH3#19 (from SLE patient S4197) (Figure 6B) had different levels of reactivity to SM-antigen. Each heavy chain was used to pair with the fourteen κ and λ light chains as used in the previous neutralization experiment. Western blot and anti-sm ELISA assays were used to measure changes in anti-sm reactivity comparing with the original heavy and light chain combination in SLE patients. The VH3#06 heavy chain could pair with 10 of the 14 light chains and express recombinant antibodies. (Recombinant antibodies paired with κ29, λ16, λ27 and λ32 were not expressed.) As shown in Figure 6C, none of the 10 light chains could neutralize 52
65 the anti-sm reactivity contributed by the VH3#06 heavy chain. The VH3#19 heavy chain could pair with all 14 light chains and produced recombinant antibodies. As shown in Figure 6D, eight of the 14 (57%) light chains (κ18, κ21, κ23, κ33, κ34, λ16, λ20, λ27) silenced the anti-sm reactivity contributed by the VH3#19 heavy chain; 5 of 14 light chains (κ05, κ19, κ26, κ30, λ32) decreased the anti-sm reactivity contributed by the VH3#19 heavy chain; surprisingly, when VH3#19 heavy chain paired with light chain κ29, the anti-sm activity increased dramatically. Changing of light chains cannot neutralize the anti-sm reactivity of heavy chain VH3#06, while majority of light chains successfully neutralized the anti-sm reactivity of heavy chain VH3#19. IgL receptor editing has different effects on the two heavy chains from SLE plasmablasts. The elevated levels of V H replacement in peripheral B cells of SLE patients may be a result of SM-antigen stimulation in attempts to eliminate the autoreactive IgH genes. 53
66 DISCUSSION V H replacement is an important mechanism to eliminate autoantibodies. However, it is difficult to study triggers of V H replacement in humans. EU12 cell line undergoes continuous V H replacement along with pro-b to pre-b and to B cell differentiation, which provides a unique model to search for the potential stimuli of V H replacement. In this study, we cloned two recombinant antibodies from EU12 μhc + cells, A19H/A15L and A19H/A18L. Both recombinant antibodies were autoreactive and recognized SM antigen. The anti-sm reactivity is mainly from A19H heavy chain and cannot be neutralized by 71% of Igκ or Igλ light chains from human naïve B cells. Treatment of EU12 μhc + cells with purified SM antigen induces V H replacement. SM antigen can be detected within EU12 culture by human anti-sm antibody, presumably released from the dead cells in the culture. Chronic exposure to SM antigen induces the endogenous V H replacement in EU12 cells. Immunoglobulin genes are generated by somatic rearrangement of V, D (for heavy chain only), and J genes. The random nature of this process ensures production of a vast diversified B cell repertoire against infections. However, this random process generates non-functional rearrangement and autoantibodies. Negative selection is necessary to prevent production of autoantibodies. Three mechanisms are involved to eliminate autoreactivity, including clonal deletion, anergy and receptor editing. B cells with autoreactive B cell receptor undergo apoptosis when challenged by self-antigen. This is 54
67 called clonal deletion. If B cells remain unresponsive under antigen challenge, it is called anergy. Immature B cell is a dedicated developmental stage for negative selection, because immature B cells express RAG1/2 gene products which are necessary for V(D)J recombination and are vulnerable to BCR induced apoptosis. Most current studies of negative selection focus on light chain receptor editing. After the primary rearrangement, upstream V L genes and downstream J L genes, flanked by 12RSS or 23RSS, are still available. Secondary rearrangement can happen in the same mechanism as primary rearrangement when the recombination machinery is active. The usage of downstream J L genes, including Jκ4, Jκ5 and Jλ3 genes, is the only indication of receptor editing. Secondary rearrangement on heavy chain genes is hard to understand because no 12RSS flanking D region mediates further rearrangement after the primary rearrangement. Recently, the molecular mechanism of heavy chain secondary rearrangement was elucidated from the human EU12 cell study. V H replacement is a mechanism which renews the entire V H coding region while keeping the same D H J H joint. V H replacement occurs between crss in a rearranged V H gene and a RSS in an upstream V H gene. Double-strand DNA breaks at the crss border in immature B cells from human bone marrow can be detected by LM-PCR (5). This is the first evidence showing that V H replacement is a spontaneous process during early B cell development. Sequence analysis suggests that V H replacement diversifies the primary B cell repertoire (38). However, why V H replacement occurs spontaneously under normal condition and the regulation of V H replacement were not known for a long time. 55
68 The study of V H replacement begins from sequence analysis of a unique acute lymphoblastic leukemia (ALL) cell line, EU12. All the heavy chain genes in EU12 cells use the same D H3-10 J H4 joint. The primary rearrangement in EU12 μhc + cells is V H2-5 D H3-10 J H4, which is nonfunctional. To salvage B cells carrying nonfunctional rearrangement from deletion, EU12 cells undergo V H replacement and form the functional V H3-7 D H3-10 J H4 rearrangement, which is the dominant clone in EU12 μhc + cells. However, after formation of functional rearrangement, EU12 μhc + cells undergo further V H replacement spontaneously as detected by LM-PCR. We searched for the antigen inducing V H replacement, which was either released from dead cells in the culture or was a component of culture medium. By western blot, we found that SM antigen released from dead cells could be recognized by EU12 B cell receptor and triggered V H replacement. In humans, a B cell has up to 16 chances to edit IgL genes, including Jκ (Jκ1, Jκ2, Jκ3, Jκ4 and Jκ5) and Jλ (Jλ1, Jλ2 and Jλ3) genes on both alleles. Because EU12 cell line is κ deficient, the chance of light chain receptor editing is less than normal condition. All seven endogenous λ genes cloned from current studies are using Jλ3 gene (data not shown). The capacity of light chain receptor editing in EU12 cells is very limited, which further reduces the number of light chain genes that can be used to neutralize the autoreactive heavy chain. Thus, to edit the autoreactivity EU12 cells have to undergo V H replacement. It is also possible that upon SM antigen induction, EU12 cells undergo both V H replacement and IgL receptor editing simultaneously. 56
69 Secondary rearrangements are ceased when the anti-sm reactivity was eliminated. The last explanation is that V H replacement and receptor editing are sequential events. After binding of SM antigen, whether V H replacement or IgL receptor editing happens first in the autoreactive B cells is determined by BCR signaling strength. This strength varies due to affinity or avidity of antigen, the site and the electronic bond between the antigen and antibody, and the stage of B cell. The affinity between SM antigen and A19H heavy chain is low to medium based on western and ELISA data. It is possible that the BCR strength is not strong enough to induce receptor editing and results in V H replacement. Using EU12 cell as a model (Figure 7), we extended our study to the pathophysiology of SLE. Anti-SM response is specific for SLE, which is found in over 30% of SLE patients. In the antibody producing plasmablasts (CD27 high CD19 low ), the frequency of V H replacement is significantly higher than in healthy control. SM antigen presents constantly in the circulation of active SLE patients. One resource of SM antigen is the apoptotic cells because SLE patients have defects in complement system, which clears cell debris. The other resource of SM antigen in SLE patient is Epstein Barr virus (EBV) infection. The proline rich sequence PPPGMRPP, which repeats three times in the C terminus of the B peptide, is the major SM epitope in patient s sera. This sequence is similar to the PPPGRRP sequence in the Epstein Barr nuclear antigen 1 (EBNA-1) in EBV, which raises the hypothesis that EBV infection predisposes individuals to develop SLE through a cross reaction between the two Epitopes (39). This study showed that the 57
70 SM antigen, when bound by BCR, induces negative selection. However, light chain receptor editing itself is not sufficient to neutralize the autoreactivity. V H replacement is elevated, by which the heavy chain is edited, and may provide means to correct the malfunction of BCRs. 58
71 Table 1. Features of light chains used for neutralization experiments VL JL CDR3 pi I K#18 V1D-39 JK1 QQSYSTLWT 13 K#33 V3-15 JK5 QQYNNWPQT 13 K#34 V3-20 JK4 QQYGSSPPLT 13 L#20 V2-14 JL3 SSYTSSSTWV 13 II K#19 V3-20 JK3 QQYGSSLPT 13 K#21 V1-9 JK3 QQLNSYPPSLPL 13 K#23 V1D-33 JK4 QQYDNLPLT 13 K#30 V1D-39 JK1 QQSYSTPRT 9.25 L#16 V1-40 JL2 QSYDSSLSGYVV 13 III K#05 V2-30 JK5 MQGTHWPPA 7.5 K#26 V3-11 JK3 QQRSNWPPL K#29 V3-15 JK3 QQYNNWPPVT 13 L#27 V1-40 JL3 SSYTSSSVWV 13 L#32 V3-21 JL2 QVWDSSSDHEV 4.1 IV A15L V3-10 JL2 YSTDSSGNHSGV 5.45 A18L V3-10 JL2 YSTDSSGVV 13 Group I: Light chains completely neutralized the anti-sm reactivity contributed by A19H heavy chain; Group II: Light chains had similar anti-sm reactivity as endogenous light chains from EU12 cells; Group III: Light chains increased anti-sm reactivity compared with endogenous light chains; Group IV: Two endogenous light chains cloned from EU12 cells. 59

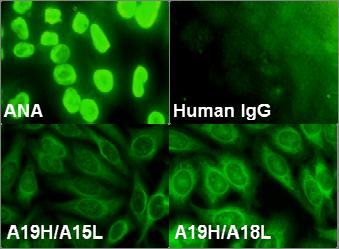
72 Figure 1 60
73 FIGURE 1. Recombinant antibodies derived from EU12 cells are autoreactive. A, Two recombinant antibodies, A19H/A15L and A19H/A18L, derived from EU12 cells. A19H, employing gene V H3-7 D 3-10 J H4, is expressed by the majority of EU12 cells. B, The recombinant antibodies recognize self-antigens in the EU12 cell nuclear extracts demonstrated by western blot analyses. The size of self-antigen is approximately 26kDa as indicated by the arrowhead. C, The recombinant antibodies weakly reacted with cytoplasmic proteins from EU12 cells. D, Anti-nuclear antigen staining by the recombinant antibodies shows that both antibodies recognize nuclear antigen(s). 61

74 Figure 2 62
75 FIGURE 2. Recombinant antibodies derived from EU12 cells react with SM antigens. A, Coomassie blue staining of purified SM antigen from calf thymus (Immunovision). Polypeptides B (26kDa) and B (27kDa) are indicated by the arrowhead. B, SM antigen (5μg per lane) was used for western blot. The two recombinant antibodies (A19H/A15L and A19H/A19L) were used as the 1 st antibody. Human anti-sm antigen antibody (Immunovision) served as the positive control. Both recombinant antibodies recognized the purified 26 kda SM antigens, as indicated by the arrowhead. C, When the human anti-sm antibody was used to blot nuclear and cytosolic extracts from EU12 cells, the 26kDa-peptide of SM antigen(s) could be detected in both extracts by western blot analyses. D, When the human anti-sm antigen antibody was used to blot culture medium of EU12 cells, the 26kDa-peptide of SM antigen(s) could also be detected by western blot analysis. However, there was no SM antigen detected in fresh medium. 63
76 Figure 3 A A19H/A15L A19H/A18L 2 nd Ab κ05 κ18 k19 κ21 κ23 κ26 κ29 κ30 κ33 κ34 λ16 λ20 λ27 λ32 A19H B OD490 nm Antibody (μg/ml) κ05 κ18 κ19 κ21 κ23 κ26 κ29 κ30 κ33 κ34 λ16 λ20 λ27 λ32 A19H/A15L A19H/A18L 64
77 FIGURE 3. Changing the light chains cannot efficiently neutralize the anti-sm reactivity contributed by the A19H heavy chain. The A19H heavy chain was artificially paired with 14 light chains from naïve B cells of one healthy control. Western analysis, (Panel A), and ELISA, (Panel B), were used to test anti-sm reactivity. Only 4 out of 14 recombinant antibodies reduced the anti-sm reactivity. 5 out of 14 recombinant antibodies remained the same activity level compared with the two endogenous antibodies from EU12 cells. However, 5 recombinant antibodies showed even stronger reactivity against SM antigen than the endogenous antibodies. Information of the light chains can be found in Table 1. 65
78 Figure 4 A V H V H3-7 D 3-10 J H4 B Cont SM V H3 crss GAPDH C Cont SM + V H3 V H1 Excision Circles GAPDH 66
79 FIGURE 4. Treatment of EU12 μhc + cells with purified SM antigen enhances V H replacement. A, Diagram of the ligation-mediated PCR and excision circle PCR assays. To determine if SM antigen treatment induced V H replacement in the EU12 μhc + cells, LM-PCR experiments were used to detect the double strand DNA breaks at the V H3 crss site after SM antigen treatment. Excision circle PCR was used to detect extrachromosomal DNA circles following RAG-mediated cleavage at the 12bp RSS and V H3 crss of V H replacement after SM antigen treatment. B, SM antigen treatment induced double stranded DNA breaks at the VH3 crss site. C, SM antigen treatment induced V H replacement excision circles in EU12 μhc + cells. 67
80 Figure 5 68
81 FIGURE 5. V H replacement product reduces anti-sm reactivity contributed by A19H. A. V H replacement product A21H was cloned from EU12 μhc + cells. DNA sequence of CDR3 was shown for both A21H and A19H. V H1-8 gene is the upstream adjacent gene of V H3-7 gene. V H replacement footprint was highlighted in A21H. B. V H replacement product reduces anti-sm reactivity. A19H and A21H were paired with two endogenous light chains A15L and A18L to product recombinant antibodies. Anti-SM reactivity was tested by ELISA. ELISA plates were coated with purified SM antigen at the concentration of 5μg/ml. Recombinant antibodies were used at 25μg/ml. 69
82 Figure 6 70
83 FIGURE 6. Different effects of changing light chains on anti-sm reactivity contributed by the heavy chains from SLE B cells. Systemic lupus erythematosus (SLE) is a complicated autoimmune disorder characterized by production of pathogenic autoantibodies against nuclear antigens, including DNA, histones, and ribonucleic proteins. Presence of anti-sm antibody is a diagnostic criterion and a unique feature of SLE disease. 25% of SLE patients develop anti-sm antibodies. It was also reported that SLE pathogenesis may be related to a deficiency in the clearance of ribonucleoprotein SM from apoptotic cells, leading to B cell activation and autoantibody production. We speculate that SM antigen in the circulation may also serve as a potential stimulus for V H replacement and anti-sm antibody production in SLE patients. We screened for anti-sm antibodies from SLE patient S1212 (A) and patient S4197 (B). Anti-SM reactivity was measured by ELISA. For IgH gene VH3#06, the anti-sm reactivity cannot be neutralized by all 14 light chains, (C). For IgH gene VH3#19, the anti-sm reactivity was efficiently neutralized by 8 out of 14 light chains, (D). 71

84 Figure 7 72
85 FIGURE 7. The model of editing anti-sm antibody in EU12 μhc + cells. Constant presence of SM antigen triggers Igλ receptor editing and V H replacement. Anti-SM reactivity cannot be completely neutralized by different endogenous Igλ genes, so EU12 μhc + cells undergo spontaneous V H replacement to eliminate the anti-sm reactivity from the V H gene. 73
86 REFERENCES 1. Zhou, M., L. Gu, C. D. James, J. He, A. M. Yeager, S. D. Smith, and H. W. Findley Homozygous deletions of the CDKN2 (MTS1/p16ink4) gene in cell lines established from children with acute lymphoblastic leukemia. Leukemia 9: Findley, H. W., L. Gu, A. M. Yeager, and M. Zhou Expression and regulation of Bcl-2, Bcl-xl, and Bax correlate with p53 status and sensitivity to apoptosis in childhood acute lymphoblastic leukemia. Blood 89: Zhou, M., L. Gu, A. M. Yeager, and H. W. Findley Sensitivity to Fas-mediated apoptosis in pediatric acute lymphoblastic leukemia is associated with a mutant p53 phenotype and absence of Bcl-2 expression. Leukemia 12: Wang, Y. H., Z. Zhang, P. D. Burrows, H. Kubagawa, S. L. Bridges, Jr., H. W. Findley, and M. D. Cooper V(D)J recombinatorial repertoire diversification during intraclonal pro-b to B-cell differentiation. Blood 101: Zhang, Z., M. Zemlin, Y. H. Wang, D. Munfus, L. E. Huye, H. W. Findley, S. L. Bridges, D. B. Roth, P. D. Burrows, and M. D. Cooper Contribution of Vh gene replacement to the primary B cell repertoire. Immunity 19: Bassing, C. H., W. Swat, and F. W. Alt The mechanism and regulation of chromosomal V(D)J recombination. Cell 109 Suppl:S Tonegawa, S Somatic generation of antibody diversity. Nature 302: Schatz, D. G., and D. Baltimore Stable expression of immunoglobulin gene V(D)J recombinase activity by gene transfer into 3T3 fibroblasts. Cell 53: Schatz, D. G., M. A. Oettinger, and D. Baltimore The V(D)J recombination activating gene, RAG-1. Cell 59: Wardemann, H., S. Yurasov, A. Schaefer, J. W. Young, E. Meffre, and M. C. Nussenzweig Predominant autoantibody production by early human B cell precursors. Science 301:
87 11. Nemazee, D. A., and K. Burki Clonal deletion of B lymphocytes in a transgenic mouse bearing anti-mhc class I antibody genes. Nature 337: Goodnow, C. C., J. Crosbie, S. Adelstein, T. B. Lavoie, S. J. Smith-Gill, R. A. Brink, H. Pritchard-Briscoe, J. S. Wotherspoon, R. H. Loblay, K. Raphael, and et al Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature 334: King, L. B., and J. G. Monroe Immunology. B cell receptor rehabilitation--pausing to reflect. Science 291: Nussenzweig, M. C Immune receptor editing: revise and select. Cell 95: Radic, M. Z., and M. Zouali Receptor editing, immune diversification, and self-tolerance. Immunity 5: Hieter, P. A., S. J. Korsmeyer, T. A. Waldmann, and P. Leder Human immunoglobulin kappa light-chain genes are deleted or rearranged in lambda-producing B cells. Nature 290: Farner, N. L., T. Dorner, and P. E. Lipsky Molecular mechanisms and selection influence the generation of the human V lambda J lambda repertoire. J Immunol 162: Brauninger, A., T. Goossens, K. Rajewsky, and R. Kuppers Regulation of immunoglobulin light chain gene rearrangements during early B cell development in the human. Eur J Immunol 31: Norvell, A., and J. G. Monroe Acquisition of surface IgD fails to protect from tolerance-induction. Both surface IgM- and surface IgD-mediated signals induce apoptosis of immature murine B lymphocytes. J Immunol 156: Fulcher, D. A., A. B. Lyons, S. L. Korn, M. C. Cook, C. Koleda, C. Parish, B. Fazekas de St Groth, and A. Basten The fate of self-reactive B cells depends primarily on the degree of antigen receptor engagement and availability of T cell help. J Exp Med 183: Chen, C., Z. Nagy, E. L. Prak, and M. Weigert Immunoglobulin heavy chain gene replacement: a mechanism of receptor editing. Immunity 3:
88 22. Chen, C., Z. Nagy, M. Z. Radic, R. R. Hardy, D. Huszar, S. A. Camper, and M. Weigert The site and stage of anti-dna B-cell deletion. Nature 373: Lopez-Macias, C., U. Kalinke, M. Cascalho, M. Wabl, H. Hengartner, R. M. Zinkernagel, and A. Lamarre Secondary rearrangements and hypermutation generate sufficient B cell diversity to mount protective antiviral immunoglobulin responses. J Exp Med 189: Koralov, S. B., T. I. Novobrantseva, J. Konigsmann, A. Ehlich, and K. Rajewsky Antibody repertoires generated by VH replacement and direct VH to JH joining. Immunity 25: Lutz, J., W. Muller, and H. M. Jack VH replacement rescues progenitor B cells with two nonproductive VDJ alleles. J Immunol 177: Roth, D. B., C. Zhu, and M. Gellert Characterization of broken DNA molecules associated with V(D)J recombination. Proc Natl Acad Sci U S A 90: Schlissel, M., A. Constantinescu, T. Morrow, M. Baxter, and A. Peng Double-strand signal sequence breaks in V(D)J recombination are blunt, 5'-phosphorylated, RAG-dependent, and cell cycle regulated. Genes Dev 7: Usuda, S., T. Takemori, M. Matsuoka, T. Shirasawa, K. Yoshida, A. Mori, K. Ishizaka, and H. Sakano Immunoglobulin V gene replacement is caused by the intramolecular DNA deletion mechanism. Embo J 11: Christensen, S. R., M. Kashgarian, L. Alexopoulou, R. A. Flavell, S. Akira, and M. J. Shlomchik Toll-like receptor 9 controls anti-dna autoantibody production in murine lupus. J Exp Med 202: Migliorini, P., C. Baldini, V. Rocchi, and S. Bombardieri Anti-Sm and anti-rnp antibodies. Autoimmunity 38: Zieve, G. W., and P. R. Khusial The anti-sm immune response in autoimmunity and cell biology. Autoimmun Rev 2: Tsokos, G. C In the beginning was Sm. J Immunol 176: Tan, E. M., and H. G. Kunkel Characteristics of a soluble nuclear antigen precipitating with sera of patients with systemic lupus erythematosus. J Immunol 76
89 96: Yurasov, S., H. Wardemann, J. Hammersen, M. Tsuiji, E. Meffre, V. Pascual, and M. C. Nussenzweig Defective B cell tolerance checkpoints in systemic lupus erythematosus. J Exp Med 201: Tan, E. M Antinuclear antibodies: diagnostic markers for autoimmune diseases and probes for cell biology. Adv Immunol 44: Qian, Y., H. Wang, and S. H. Clarke Impaired clearance of apoptotic cells induces the activation of autoreactive anti-sm marginal zone and B-1 B cells. J Immunol 172: Jacobi, A. M., M. Odendahl, K. Reiter, A. Bruns, G. R. Burmester, A. Radbruch, G. Valet, P. E. Lipsky, and T. Dorner Correlation between circulating CD27high plasma cells and disease activity in patients with systemic lupus erythematosus. Arthritis Rheum 48: Liu, Y., R. Fan, S. Zhou, Z. Yu, and Z. Zhang Potential contribution of VH gene replacement in immunity and disease. Ann N Y Acad Sci 1062: Arbuckle, M. R., M. Reichlin, J. B. Harley, and J. A. James Shared early autoantibody recognition events in the development of anti-sm B/B' in human lupus. Scand J Immunol 50: Cascalho, M., A. Ma, S. Lee, L. Masat, and M. Wabl A quasi-monoclonal mouse. Science 272:
90 POTENTIAL CONTRIBUTION OF VH REPLACEMENT PROUDUCTS TO THE GENERATION OF AUTOREACTIVE ANTIBODIES IN SYSTEMIC LUPUS ERYTHEMATOSUS By RUN FAN, KAIHONG SU, ASHLEY MAJORS, ZHIHONG YU, ROBERT P KIMBERLY, WINN CHATHAM, ROBERT CARTER, AND ZHIXIN ZHANG Manuscript in preparation for submission Format adapted for dissertation 78
91 ABSTRACT Systemic lupus erythematosus (SLE) is characterized by the production of high affinity autoantibodies against various nuclear antigens. To gain insight into the mechanisms of autoantibody production during the course of SLE, we performed single cell PCR analyses and obtained 447 IgH genes with paired Igκ or Igλ genes from the plasmablasts (CD19 low CD27 high ) and naïve B cells (CD19 + CD27 - ) of three SLE patients and two healthy controls. IgH genes from SLE patients have longer CDR3 regions that are enriched in Arginine residues, a common feature of anti-dna autoantibodies. SLE plasmablasts derived IgH and IgL genes are highly mutated, indicating their positive selection and active involvement in the abnormal immune responses. Interestingly, analyses of the IgL and IgH genes from SLE patients showed signs of excessive secondary recombination of IgL genes when compared with those from healthy donors, leading to elevated usage of downstream Jκ5 and Jλ3 genes. The frequency of V H replacement products in the IgH genes from all three SLE patients is significantly higher than that in the control donors. All the V H replacement footprints preferentially generate charged amino acids in the IgH CDR3 regions. Anti-nuclear antigen test showed nine of twelve (75%) recombinant antibodies derived from SLE plasma cells recognize perinuclear antigens. Importantly, five out of six V H replacement products encoded autoantibodies. These findings showed for the first time that V H replacement products are enriched in SLE plasmablasts, which directly contributes to the generation of autoantibodies. 79
92 INTRODUCTION Somatic rearrangement creates various immunoglobulin genes and allows the immune system to generate a repertoire of virtually unlimited antibodies against infectious agents. Somatic rearrangement is catalyzed by a pair of recombination activating gene products, the RAG1 and RAG2 proteins (1-3). The variable (V), diversity (D) (heavy chain only), and joining (J) gene segments intersperse in genome are joined together to form functional immunoglobulin genes in this process (4). A potential risk of V(D)J recombination is that autoantibodies are produced as well. B cells producing antibodies against self-antigens must be eliminated, anergized, or have their B cell receptors changed (5). Defects in any of these steps may result in the release of autoreactive B cells into the periphery (6). Receptor editing is the process of changing B cell receptors by secondary V(D)J recombination (7). Studies of receptor editing have been mainly focused on the IgL genes. V L and J L genes within Igκ or Igλ locus are flanked by two types of RSS sites (8). This genetic organization allows secondary rearrangement through joining of an upstream V L gene with a downstream J L gene segment, in a similar mechanism to the primary rearrangements (9). Secondary rearrangement can happen as long as the light chain locus remains accessible and the recombination machinery is active. In a mouse model carrying the human Cκ gene, secondary light chain rearrangement was observed in approximately 25% immature B cells (5, 10). 80
93 Analysis of the three dimensional structure of antibody-antigen complexes has shown that the Complementarity Determining Regions (CDRs) contact the antigen. In many complexes all six CDRs contact the antigen (11). In general, the residues in heavy chain CDRs are more important for contacting antigens than those in light chain CDRs. The V H domain contributes more to antigen binding than the V L domain, which indicates receptor editing of V L domain is enough to change the antibody specificity. However, in contrast to the relative ease of secondary light chain V L J L rearrangement, secondary rearrangement of an upstream V H to a preformed V H DJ H gene is difficult (12). After the primary V H to DJ H rearrangement, all the intervening D segments are deleted. This leaves no 12bp spacer RSS flanked gene segment to recombine with the 23bp spacer RSS flanked upstream V H genes or downstream J H genes (13, 14). Studies in mouse pre-b cell lines with nonfunctional IgH rearrangements (15, 16) and autoreactive VDJ gene knock-in mice (17-19) showed that V H replacement occurred through RAG-mediated recombination between a cryptic recombination signal sequence (crss) in a rearranged V H gene and a 23RSS in an upstream V H gene. In humans, 40 of 44 functional V H germline genes contain heptamer (TACTGTG) crss motifs, which can be targeted by V H replacement (20). V H replacement replaces the entire V H coding region, leaving a short stretch of nucleotides from the previous rearrangement as a V H replacement footprint in the V-D junction. Interestingly, these V H replacement footprints extend the IgH CDR3 regions preferentially with charged amino acids (21). About 5% of the normal IgH genes 81
94 in peripheral blood B cells are determined as potential V H replacement products by the potential V H replacement footprints within the V H -D H junction of these IgH genes (21), suggesting a significant contribution of V H replacement to the primary B cell repertoire. The study of biological function of V H replacement comes from transgenic and knock-in mouse. V H replacement can rescue B cells carrying anti-dna or non-functional VDJ rearrangement and generated a highly diverse heavy chain repertoire. The hallmark of systemic lupus erythematosus (SLE) is the production of high affinity autoreactive antibodies against nuclear antigens. This leads to the pathogenic immune complex formation and deposition in kidneys of SLE patients (22). Anti-DNA antibodies in the normal antibody repertoire have low affinity. In contrast, anti-dna antibodies in SLE possess high affinities for double strand DNA (dsdna), with somatic hyper-mutations in their variable regions and IgG isotype (23). The DNA binding specificity of autoreactive antibodies is determined mainly by the IgH CDR3 region. As shown by structural analysis, the IgH CDR3 loop directly contacts the DNA ligand (24). Analysis of immunoglobulin genes encoding anti-dna antibodies showed an overrepresentation of charged amino acid residues, especially Arginine, within the CDR3 regions (25). It is not clear how these high affinity antibodies are generated and enriched during the course of SLE. Previous studies have shown that the number and frequency of a subgroup of 82
95 plasmablasts (CD27 high /CD19 low /Ig low /CD38 + /CD138 + ) are elevated and significantly correlate with the disease activity and the titer of anti-dsdna antibodies in SLE patients (26). The plasmablasts produced anti-dsdna antibodies in vitro culture. Previous analysis of 17 IgH genes from SLE plasma cells showed that these IgH genes had abnormally long CDR3 regions enriched of charged amino acids. However, how these IgH genes that potentially encode autoantibodies are generated is not known. Further analysis of these 17 IgH genes showed that 8 of them are potential V H replacement products. Importantly, majority of the V H replacement footprints contribute Arginine to the IgH CDR3 regions. Positively charged amino acids can interact with the negatively charged phosphate backbone in nucleic acids, and long CDR3 regions enriched of aromatic residues may facilitate the interaction by stacking (24). Based on these observations, we hypothesize that V H replacement products contribute to the generation of autoreactive antibodies in SLE. By using single-cell PCR, we cloned IgH, Igκ and Igλ genes from three SLE patients and two healthy controls. Sequence analysis showed that IgH genes derived from SLE patients have higher frequency of V H replacement in both naïve B cells and plasmablasts. IgH genes from SLE plasma cells have longer CDR3 region and are enriched with Arginine residue within the CDR3 region. Expression of recombinant antibodies from SLE plasmablasts showed that 75% of them are autoreactive, especially V H replacement products. Signs of excessive light chain receptor editing also observed on Igκ or Igλ 83
96 locus in all three SLE patients. 84
97 METERIALS AND METHODS Cells Peripheral blood mononuclear cells were obtained from active SLE patients and healthy donors by Ficoll-gradient centrifuge. Cells were subjected to FACS analysis with CD19 and CD27 antibodies. Plasmablasts were identified as CD19 low CD27 high cells and naïve B cells were identified as CD19 + CD27 - cells (26). Plasmablasts and naïve B cells were purified by FACS sorting and deposited into 96-well PCR plates at one cell per well by limiting dilution. The same cell populations were purified from healthy donors as controls. Cells were stored at -80 C in 1U of RNase Inhibitor (RNasin, Promega) until use. Single cell RT- PCR The V(D)J joint regions of IgH, Igκ, or Igλ genes were amplified by single cell RT-PCR (27). RT-PCR and 1 st round PCR were performed with One Step RT-PCR kit (Qiagen) under the condition 50 C, 30 min; 94 C, 15 min; 94 C, 20 sec, 55 C, 30 sec, 72 C, 1 min for 50 cycles, 72 C, 10 min and stop at 4 C. 2 nd round PCR was performed with HotMaster Taq DNA Polymerase (Eppendorf) under the condition 94 C, 3 min, 94 C, 20sec, 57 C (IgH/Igλ) or 60 C (Igκ), 30 sec, 72 C, 45 sec for 50 cycles, 72 C, 5 min and stop at 4 C. The average single cell RT-PCR efficiency is 36.4%. Restriction enzyme digestion sites were introduced by the 2 nd round of single cell 85
98 RT-PCR. Digested IgH, Igκ and Igλ PCR products were purified by QIAquick PCR purification kit (Qiagen) and directly cloned into expression vectors containing human Igγ1, Igκ or Igλ constant regions. Sequence analysis Obtained immunoglobulin gene sequences were analyzed using the IgBLAST program to determine the potential V H, D H and J H germline gene usages ( The number of mutated nucleotides and amino acid sequences is determined by JOINSOLVER ( V H replacement products were identified by searching for V H replacement footprints (Table. 1). Expression and purification of recombinant antibodies Human embryonic kidney fibroblast cell line 293H was cultured in DMEM medium supplemented with 10% of low IgG FBS (GIBCO). 10 μg of each IgH and IgL expressing constructs were cotransfected using Fugene6 reagent (Roche) in 100mm culture plate. Supernatants were collected 7 days after transfection. The concentration of recombinant antibodies was measured by ELISA using anti-human IgG1 antibody. Human monoclonal IgG1 was used as standard (Sigma). Recombinant antibodies were purified and concentrated using protein A Sepharose beads (Amersham Pharmacia Biosciences) for immunofluoresence studies. 86
99 Anti-Nuclear Antigen Immunofluorescence Assay HEp-2 cell coated slides (Bion Enterprises, LTD.) were incubated at room temperature with purified antibodies at μg/ml for 30 min, washed in PBS and then incubated for another 30 min with FITC conjugated anti-human immunoglobulin. Samples were stored at 4 o C until pictures were taken by fluorescent microscopy. 87
100 RESULTS The CD19 low CD27 high plasmablasts are significantly elevated in SLE patients. CD19 low CD27 high plasmablasts from SLE patients secrete autoreactive antibodies in culture, especially anti-dna antibodies, suggesting that CD19 low CD27 high plasmablasts correlate with SLE disease activity (26). Mononuclear cells from peripheral B cells of SLE patients and controls were stained with CD19 and CD27 after Ficoll-gradient separation (Figure 1A). Naïve B cells were identified as CD19 + CD27 - to serve as control for plasmablasts. The same cell population was analyzed in 37 healthy donors, 18 active SLE patients and 4 inactive SLE patients. The frequency of CD19 low CD27 high plasmablasts is significantly higher in active SLE patients compared to healthy controls and inactive SLE patients (Figure 1B). The CD19 low CD27 high plasmablasts and CD19 + CD27 - naïve B cells were purified by FACS and deposited into 96-well PCR plates at one cell per well by limiting dilution. Single-cell RT-PCR was employed to amplify of the V(D)J joint regions for paired IgH, Igκ or Igλ genes. Obtained Ig gene sequences were analyzed using the IgBLAST program to determine the potential V(D)J germline gene usage. To exclude the effect of cross reaction between primers and Ig genes on gene assignment, the primer covered region of the sequences were not used for analysis. V H and J H gene repertoire in SLE patients and healthy controls 313 IgH genes were cloned from three SLE patients and 134 IgH genes were cloned from two healthy controls. The usage of genes V H1-69, V H3-30, V H3-15, and V H1-2 is significantly 88
101 higher in SLE patients than in healthy controls. The usage of genes V H3-66, V H1-46, V H4-34, V H3-23 and V H1-18 are significantly lower in SLE patients than in healthy controls (Figure 2A). J H gene usages in patients and controls are shown in Figure 2B. In both naïve B cells and plasmablasts, J H6 gene usage is lower in SLE patients than in healthy controls. None of the 90 heavy chain genes cloned from SLE patient S4197 and S1267 naïve B cells uses J H6 gene. IgH genes in SLE naïve B and plasmablasts have abnormally long CDR3 regions enriched of Arginine residue. IgH genes derived from autoreactive B cells have longer CDR3 region, which is enriched with charged amino acids. In anti-dna antibodies, which is present in 70-80% SLE patients and involved in SLE pathogenesis, the charged amino acid is usually positively charged Arginine. To determine if IgH genes from SLE patients have the features of autoreactive antibodies, the IgH genes were divided into four groups based on the CDR3 length (Figure 3A) for analysis. In healthy control, the IgH genes with CDR3 region longer than 58bp are negatively selected from naïve B cells to plasmablasts, with the frequency dropped from 14.1 % to 3.8 %. In SLE patients, the IgH genes with CDR3 region longer than 58bp remain the same frequency in patient S4197 (6.7%), and are positively selected in patient S1267 (2.5% in naïve B cells to 11.8% in plasmablasts) and S1212 (13.9% in naïve B cells to 29.7% in plasmablasts) (Figure 3A). 89
102 We further compared the number of Arginine residues in CDR3 region of IgH genes. IgH genes were divided into four groups based on the number of Arginine residues within their CDR3 regions. In healthy control, the IgH genes with three or more Arginine residues in their CDR3 regions were negatively selected from naïve B cells to plasmablasts. 5.8% IgH sequences in naïve B cells of healthy controls have 3 or more Arginine residues within the CDR3 regions, but none in plasmablasts. However, IgH genes with 3 or more Arginine residues within the CDR3 regions are enriched during B cell differentiation from naïve B cells to plasmablasts in SLE patients. Although the frequency of IgH genes with 3 or more Arginine residues within the CDR3 regions in naïve B cells from patient S4197 (2.2%) and S1267 (2.9%) is lower than in control naïve B cells, it increased to 7.1% (S4197) and 5.9% (S1267) in plasmablasts, significantly higher than in naïve B cells (Figure 3B). The frequency of IgH genes with 3 or more Arginine in S1212 naïve B cells is 10.8%, and it s increased to 14.1% in plasmablasts. In conclusion, the IgH genes from SLE patients have longer CDR3 region and are enriched of Arginine residues within the CDR3 region. They have features of anti-dna antibodies. IgH genes in SLE plasmablasts are highly mutated. We compared the average number of mutated nucleotides within the V H coding region of IgH genes derived from the plasmablasts and the naïve B cells of SLE patients. In patient S4197, the IgH genes carry 2.3bp mutations per gene averagely in naïve B cells, and 19.5bp mutations per gene averagely in plasmablasts. In patient S1267, the IgH genes 90
103 carry 2.3bp mutations per gene averagely in naïve B cells, and 15.6bp mutations per gene averagely in plasmablasts. In patient S1212, the IgH genes carry 2.9bp mutations per gene averagely in naïve B cells, and 14.6bp mutations per gene averagely in plasmablasts (Figure 4). Clearly, IgH sequences derived from SLE plasmablasts are highly mutated, indicating that they have been positively selected and actively involved in germinal center reaction, presumably due to abnormal immune response (28). Excessive editing on Jκ and Jλ loci in SLE patients Immunoglobulin light chain editing is an efficient way of eliminating autoreactive antibodies. To study the role of IgL receptor editing in SLE patients, usages of downstream Jκ and Jλ genes were analyzed. J L repertoire of 304 Igκ genes and 202 Igλ genes from SLE patients and healthy controls was analyzed. In patient S4197, Jκ5 gene usage is higher in both naïve B and plasmablasts than in healthy controls. In patient S1267, Jκ5 gene usage is increased in naïve B cells, but slightly decreased in plasmablasts comparing to healthy controls. In patient S1212, Jκ5 gene usage is significantly decreased in plasmablasts comparing to healthy controls (Figure 5A). Analysis of the Jλ gene repertoire is shown in figure 5B. In patient S4197, Jλ3 gene usage is slightly decreased in both naïve B and plasmablasts comparing to healthy controls. For patient S1267 and S1212, the Jλ3 gene usage is significantly higher than in healthy controls. In conclusion, all SLE patients had higher Jκ5 gene usage (S4197) or Jλ3 gene usage (S1267 and S1212) than healthy controls. SLE patients have excessive 91
104 editing on both Jκ and Jλ loci. Elevated frequency of V H replacement in IgH genes derived from SLE patients Although IgL receptor editing is elevated in active SLE patients (Figure 5), production of autoantibodies persists. V H replacement is another mechanism to change the affinity of antibodies. We next studied the role of V H replacement in SLE patients. V H replacement products were identified by searching footprints of formerly used V H genes at IgH gene V-D joint regions that match the 3 end of germline V H genes. Sequences with one mismatch to the germline sequences among 6 conserved nucleotides were also considered as footprints in a less stringent setting. As shown in Figure 6, 12% of the IgH genes derived from naïve B cells and 28.6% of the IgH genes derived from plasmablasts in patient S4197 are V H replacement products. In patient S1267, the ratio of V H replacement products is 8.7% in naïve B cells and 16.3% in plasmablasts. In patient S1212, the ratio of the V H replacement products is 21.5% in naïve B cells and 22.3% in plasmablasts. The frequencies of V H replacement products are significantly elevated in all SLE naïve B cells and plasmablasts of patient S4197 and S1212 in comparison with healthy controls, whose V H replacement frequency is 0 in naïve B cells and 8.5% in plasmablasts. Significant elevation of V H replacement in SLE patients suggests that V H replacement products might contribute to the generation of autoreactive antibodies in SLE. In addition, the frequencies of V H replacement in the naïve B cells of S4197 (SLEDI=6) and S1212 (SLEDI=4) are also significantly higher than that in S1267 (SLEDI=2), suggesting that 92
105 V H replacement might be abnormally regulated in active SLE patients. V H replacement footprints preferentially encode charged amino acids to CDR3 region. Previous sequence analysis suggests that V H replacement footprints preferentially contribute charged amino acids into the IgH CDR3 regions in a human B cell line, EU12 (21). We analyzed amino acids encoded by the V H replacement footprints in IgH sequences derived from SLE patients (Figure 7). Arginine (R), which is positively charged, occurs at the highest frequency among all amino acid residues. Two negatively charged amino acids, Aspartatic acid (D) and Glutamatic acid (E), are encoded at frequencies next to Arginine. This result is consistent with previous sequence analysis and confirms that the V H replacement footprints preferentially contribute charged amino acids into IgH CDR3 regions. Recombinant antibodies cloned from SLE plasmablasts are autoreactive. All the heavy chain and light chain genes were cloned into expression vectors and recombinant antibodies were produced from endogenously paired heavy and light chain genes. To determine the potential autoreactivity of the recombinant antibodies from SLE patients, we performed HEp-2 cell based ANA assays (27). Of the 12 recombinant antibodies derived from SLE plasma cells, 9 showed strong anti-nuclear or anti-cytoplasma staining patterns; 3 did not stain. Representative staining is shown in 93
106 Figure 8. Importantly, 5 out of 6 recombinant antibodies derived from the identified V H replacement products all displayed strong anti-nuclear staining. These results showed that the enriched V H replacement products in SLE plasmablasts encode autoantibodies. 94
107 Table 1. V H replacement products in SLE patients. VH V-D DH JH 1-2 TACTGTGCGAG GCCGAGcTCCTGCAAC TACTGTGCGAG GGGAGGCGGgAGGG TACTGTGCGAGAG TATTCGCTGCTCCGGCGTCCCGAAA TACTGTGTTAGAG GCCACAC TACTGTGCGAGA CAGACGCC CACAGACT TATTGTGCGA AATTTCGGAaA TAAAAAGAAGCCTC GCGTCCCACGG TACTGTGCGAGA CAAAAAGGGTCGA TACTGTGCGAGAGA GGCCCAACACCGTCT TACTGTACCAC TGAAGACCCCA TACTGTACCACA CCcGAGGCCGC TACTGTTCCACAG CGAGgG GGCTGTTTATTA TTGTACGAcAATCCT TACTGTGCGAGA TCgGACCCCG TACTGTGCGAGAG CCTATGCGgGAGC TCGACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAA 1 GAGCACCTCTGGGGGCACAGC 3-23 TACTGCGCGA GATTAGCCGTACGTCGAGCTTATATAGGCGAG TACTGTGCGA GGGACCTAGATCCCGGA TACTGTGCGA TCCCTGAAACA TACTGTGCGAAAG TACCAGgGGTCT TACTGTGCGA AAAGgGACCGAA TACTGTGCGA AAGAAGAGGGAC TACTGTGCGA AAGATC TACTGTGCGAAAG CACcAAAGG TACTGTGCGAAAG GTCAGACGCCTTATCGCAGGAC TACTGTGCGAAAGA TGTCGGGGCGGCACTAAA TACTGTGCGA AAGAGGAGGGG TACTGTGCGAGAGA CTTARTCGGATCG TACTGTGCGAGAGA TAGAAACCGCCTTACAATGGCG TAC GCCGTTTTCACGGGCGGACCCG TACTGTGCGAGAGA ACTGGAGAGG TACTGTGCGAGAGA CGCTCACCCACCTACGGTGG TACTGTGCGAGAGA TCCCAAAGAAAaACATGG TACTGTAC GGGGGGAAAAGAcACC TACTGTGTGAG CTGCTCGCTGCAGA TACTGTG TGCGAGACAACTTCC
108 3-7 TACTGTGCGAAAGA CCAAGATCTGGGC TACTGTGCGAGAGA TGAGGACGGAGTCATAT TACTGTGTGAGAGA TCTCGGAAAGAGTG TATTGTGAGA CACACGAAGGT TACTGCGCAA GGACGGATAATTGGAATTACGGAGGGGGTA TACTGTGCAAAA AGCCAGgGGGGGG TACTGTGTAAAAGATA CCGcGATGG CCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGC TACTGTGCGAGAGG CCTGCGAGtGGAA TACTGTGCG GCCCTAGACC TACTGTGCGAGA GAGTTACCGAAGAGGTCGGGAGAC TACTGTGCGAGAC CCgGAGGAAC TCCTATGAGCTGTGTGCTCCCTCAAGAC TACTATTGTGCGAGA CGTCAGACAGAGAGTGCAGTG TACTGTGCGAG GACCgGACCTGA TACTGTGCGAGAGA AGAAAGGCGCCTTGG TACTG CGCAAGATCAGACTGGGACAAAT TACTGTGCG GTTTCGCCAAGAACCGA TACTGTGCGAGA CTGGAGgTA TACTGTGCGAGA GTGAtAGGAG TACTGTGCGAGA TGCGACGtGAA TACTGTGCGCGA CCCACGGACGGAT TACTGTGCGAGACA AAAGGAGGATATgC V H replacement products were identified by analyzing IgH gene V-D joint sequences for footprints of formerly used V H genes in the form of pentameric nucleotides (as highlighted in red) that match the 3 end of germline V H genes. With a less stringent setting, 1 mismatch among 6 conserved nucleotides was also accepted (as highlighted in purple). 96

109 Figure 1 97
110 FIGURE 1. The CD19 low CD27 high plasmablasts are significantly elevated in SLE patients. A, FACS analysis of peripheral blood mononuclear cells from a non-sle healthy donor and a SLE patient. Plasmablasts were identified as CD19 low CD27 high and the naïve B cells were identified as CD19 + CD27 -. B, The frequency of CD19 low CD27 high plasmablasts is elevated in active SLE patients compared to healthy controls and inactive SLE patients. The CD19 low CD27 high plasmablasts and CD19 + CD27 - naïve B cells were purified by FACS and deposited into 96-well PCR plates at one cell per well by limiting dilution. Single-cell reverse-transcription PCR was employed to amplify of the V(D)J joint regions for paired IgH, Igκ or Igλ genes. Obtained Ig gene sequences were analyzed using the IgBLAST program to determine the potential VDJ germline gene usage. 98

111 Figure 2 99
112 FIGURE 2. The IgH gene repertoire in SLE patients. A, Comparison of the IgH V gene repertoire of SLE patients (n=313) with a healthy control (n=131). The frequency of V H1-69 (p=0.0009), V H3-30 (p=0.0073), V H3-15 (p=0.0123), and V H1-02 (p=0.013) genes was significantly increased (indicated in red bars). The frequency of V H3-66 (p=0.0032), V H1-46 (p=0.05), V H4-34 (p=0.0097), V H3-23 (p=0.0001), and V H1-18 (p=0.0031) genes was significantly decreased (indicated in black bars). B, Pie charts depict J H family usage in SLE patients and healthy controls. The absolute number of sequences analyzed in each B cell compartment is indicated in the center of the pie charts. In general, the frequency of J H6 genes in three SLE patients are less than in the healthy control for both naïve B cells and plasmablasts. A significantly decreased frequency of J H6 genes was seen in naïve B (p=0.0033) and plasmablasts (p=0.03) of SLE patient S4197, as well as in naïve B cells of SLE patient S1267 (p=0.0076) compared with the correlate group of cells in the healthy control. p values were calculate by two tailed Fisher s exact test. 100

113 Figure 3 101
114 FIGURE 3. IgH genes derived from SLE plasmablasts have features of autoantibodies. A, Bar graphs show frequency of IgH CDR3s with 27bp (yellow bars), 28-42bp (green bars), 43-57bp (purple bars) and 58bp (red bars). In healthy controls, IgH with long CDR3 (red bars) are counterselected during B cell development from naïve B cell to plasmablasts. While for three SLE patients, from naïve B cells to plasmablasts, the frequency of IgH with long CDR3 (red bars) are enriched. Also the frequency of IgH with long CDR3 in plasmablasts from three SLE patients is higher than in plasmablasts from healthy control, especially for plasmablasts from SLE patient S1212, in which the difference is highly significant (p=0.0002). B, The frequency of IgH genes with 0, 1, 2, or 3 Arg residues within the IgH CDR3 region. The total number of sequences analyzed in each B cell compartment is indicated in the center of the pie charts. For healthy control, as B cells develop from naïve B cells to plasmablasts, all the IgH with 3 Arg residues within the CDR3 regions are negatively selected, while for three SLE patients, the frequency of IgH with 3 Arg residues within the CDR3 regions is increased as B cells developing from naïve B cells to plasmablasts. Compared with plasmablasts from the healthy control, the frequency of IgH with 3 Arg residues within the CDR3 region is elevated in plasmablasts from all three SLE patients, especially S1212 (p=0.0043). p values were calculated by two-tailed Fisher s exact test. 102
115 Figure 4 FIGURE 4. IgH sequences derived from SLE plasmablasts are highly mutated. The number of mutated nucleotides was determined by JOINSOLVER ( The average number of mutated nucleotides within each cell population is showed by black bars. 103

116 Figure 5 104
117 FIGURE 5. Excessive Jκ (n=304) and Jλ (n=202) editing in SLE patients. A, The frequency of the Jκ5 gene is elevated in naïve B cells and plasmablasts of SLE patients S4197 and S1267, suggesting that there is excessive editing of the Igκ locus. However, in plasmablasts from SLE patient S1212, the frequency of the Jκ5 gene is significantly lower than plasmablasts from healthy control (p=0.01), which suggests that there is reduced receptor editing at the Igκ locus in plasmablasts from this patient compared with plasmablasts from healthy control. B, The frequency of the Jλ3 gene is elevated in naïve B cells and plasmablasts from SLE patients S1267 and S1212, suggesting that there is excessive editing at the Ig λ locus in these patients. The frequency of the Jλ3 gene is significantly higher in plasmablasts from SLE patient S1267 (p=0.03) and S1212 (p=0.018) compared with plasmablasts from a healthy control. The total number of sequences analyzed in each B cell compartment is indicated in the center of the pie charts. p values were calculated by a two-tailed Fisher s exact test. 105
118 Figure 6 FIGURE 6. Excessive V H replacement in SLE patients. By identifying the V H replacement footprint within the CDR3 region, the frequency of V H replacement was determined. p values were calculated by a two-tailed Fisher s exact test. V H replacement products from three SLE patients and a healthy control are listed in Table
119 Figure 7 FIGURE 7. V H replacement footprints preferentially generate charged amino acids in the CDR3 region. Among 69 V H replacement footprints from both SLE patients and a healthy control, 4 amino acids are present frequently. Two of them are positively charged, R (Arginine, red bar) and K (Lysine, blue bar). The other two residues are negatively charged, D (Aspartic Acid, grey bar) and E (Glutamic Acid, black bar). 107




120 Figure 8 FIGURE 8. Recombinant antibodies derived from SLE plasmablasts encode autoantibodies and are positive for ANA staining. A, Positive control showed anti-nuclear staining pattern. B, Negative control with human Ig. C-H, Staining with recombinant antibodies generated by V H replacement. I-N, Staining with recombinant antibodies don t have V H replacement footprint 108
121 Figure 9 FIGURE 9. The model of generation of autoantibodies in SLE by V H replacement. When immature B cells are challenged by self antigens, immature B cells undergo IgL receptor editing and V H replacement to change the autoreactivity before they are deleted from the B cell repertoire. The B cell repertoire ends up with high frequency of V H replacement products. However, V H replacement products have features of autoantibodies, with long CDR3 enriched with charged amino acids. Also V H replacement preferentially induces charged amino acid into the CDR3 region of IgH genes. Thus, autoantibodies are generated in SLE patients. 109

122 Supplementary Figure 1 Supplemental Figure 1 Jκ and Jλ gene usages for paring with V H replacement versus non-v H replacement products in SLE plasmablasts. 110