HMGB1 interacts with many apparently unrelated proteins by recognizing short amino acid sequences
|
|
|
- Pamela Anissa Todd
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on December 17, 2001 as Manuscript M HMGB1 interacts with many apparently unrelated proteins by recognizing short amino acid sequences Agnès Dintilhac and Jordi Bernués Departament de Biologia Molecular i Cel lular Institut de Biologia Molecular de Barcelona, CID Consell Superior d Investigacions Científiques Jordi Girona, Barcelona, Spain To whom correspondence should be addressed: Jordi Bernués Dept. Biologia Molecular i Cel lular Jordi Girona, Barcelona, Spain Tel.: Fax: jbmbmc@ibmb.csic.es Running Title: New factors interacting with HMGB1 Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
2 Summary The chromatin High Mobility Group protein 1 (HMGB1) 1 is a very abundant and conserved protein that is structured in two HMG-box domains plus a highly acidic C-terminal domain. From the ability to bind DNA unspecifically and to interact with various proteins, several functions in DNA-related processes have been assigned to HMGB1. Nevertheless, its functional role remains the subject of controversy. Using a phage display approach we showed that HMGB1 can recognize several peptide motifs. A computer search of the protein databases found peptide homologies with proteins already known to interact with HMGB1, like p53, and allowed us to identify new potential candidates. Among them, some transcriptional activators like heterogeneous nuclear ribonucleoprotein K (hnrnp K), repressors like Methyl-CpG binding protein 2 (MeCP2), and co-repressors like the Retinoblastoma susceptibility protein (prb), and Groucho-related gene proteins 1 (Grg1) and 5 (Grg5) can be found. A detailed analysis of the interaction of Grg1 with HMGB1 confirmed that the binding region contained the sequence homologous to one of the peptides identified. Our results lead us to propose that HMGB1 could play a central role in the stabilization and/or assembly of several multifunctional complexes through protein-protein interactions.
3 Introduction In the eukaryotic cell nucleus of all vertebrate cell types HMGB1 (formerly named HMG1, see (1) for a revised nomenclature) is one of the most abundant non-histone proteins. HMGB1 is essential since knock-out mice die 24 hours after birth (2). HMGB1 is highly conserved specially in mammals and to a lesser extent all through the animal kingdom. HMGB1 is structured into three domains: two basic HMG-boxes (HMG-domains A and B) and a highly acidic C-terminal domain that confer an overall dipolar appearance to this protein (see (3-5) for reviews). Each of the HMG-boxes is formed by two short and one long α-helix that upon folding produce an L- or V-shaped three dimensional domain structure (6-8). Whereas the acidic C-terminal domain is presumably involved in the modulation of HMGB1 activity, the HMG-box domains allow the protein to bind to linear DNA with moderate affinity and to highly structured (3- and 4-way junction DNA, cruciform DNA) or distorted DNA (bent or kink DNA, bulged DNA, cisplatinmodified DNA) with higher affinity but always without sequence specificity. The concave surface of the L- or V-shaped HMG-box domain contacts the DNA in the minor groove in two slightly different ways introducing important modifications in the structure of DNA, in particular a strong bend (reviewed in (5)). Presumably, these features will be of relevance for the biological functions in which HMGB1 has been involved (DNA repair, recombination, replication, and transcription). The activity of HMGB1 is not solely mediated by its ability to bind to DNA. Indeed, HMGB1 and the related HMGB2 protein can interact through their HMGbox domains with a broad range of proteins ranging from nuclear cell proteins to viral proteins. Interactions of HMGB1 have been described with the Recombination Activation Gene protein RAG1 (9), several transcription factors including the cellular tumor suppressor p53 (10), the octamer transcription factors Oct1, Oct2, Oct4 and Oct6 (11,12), some homeotic HOX proteins (13), the steroid receptors (progesterone PR, glucocorticoid GR, estrogen ER, and androgen AR) (14,15), the general initiation factor human TATA-binding protein (htbp) (16-18), and the viral replication proteins Rep78 and Rep68 (19). The consequences of these interactions are multiple. HMGB1 in general increases the DNA binding affinity of those factors and depending on the context and the assay conditions HMGB1 has shown to have a positive or a negative effect on transcription (14-17,20). In the case of RAG1 and Rep68/78 HMGB1 enhances the rate of
4 sequence-specific DNA cleavage reaction (19,21). Interestingly, HMGB1 can also stimulate the ATPase activity of Rep78 (19). The two HMG-box domains of HMGB1 appear to have a similar but not identical behaviour with respect to their protein interacting features. Thus, HMGbox A is the one important for binding to htbp and p53 whereas the binding to Oct factors, HOX factors, and hormone receptors can take place through boxes A or B (11,13,17,22). However, the interaction with RAG requires both HMG-box domains (9). To date, neither the HMG-box surface that is involved in the interaction with other proteins nor the amino acids of HMGB1 required are known. On the other hand, sequence analysis of the factors interacting with HMGB1 does not highlight any apparent homology or similarity among them. For instance, the interaction with RAG1, Oct and Hox factors occurs at the homeodomain. In the case of htbp it is the H2 α-helix of the core and with Rep78 two different regions are recognized. From these data, no consensus can be defined. The fast progress of genomics and proteomics has made obvious that an important focus in understanding biological processes is to characterize how proteins interact in macromolecular complexes. Attempts to define general rules for predicting specific recognition between proteins have been unsuccessful since each protein-protein interaction has its own properties. Nevertheless, some indications are emerging. The development of powerful tools has allowed to demonstrate that one type of recognition involves asymmetric interactions that occur between a particular domain and a short region, often less than 10 amino acids in length, within the other protein. A recent review (23) recapitulated several examples of protein domains involved in these kinds of interactions like SH3 (Src homology 3), phosphotyrosine-binding WW, EH (Eps15 homology), PDZ modules (PSD-95/dlg/ZO1), as well as prb, and the estrogen receptors (ER). Some of them, as for the ER, could interact in several modes with different peptides, in a way depending on the ligand bound. In the present study we have explored the molecular recognition properties of HMGB1 by ligand selection from a large library of heptapeptides displayed on phages. Our results do not give support to a unique strong consensus sequence but a few different kinds of peptides. Blast search allowed us to predict some new proteins that may interact with HMGB1. We have tested and confirmed the interactions with some of them and showed that HMGB1 can interact not only with transcriptional activators but also with repressors and co-repressors. Taken
5 together the data suggest a complex network of protein-protein interactions that will be discussed in order to understand the biological function(s) of HMGB1. Experimental procedures Expression and purification of recombinant HMGB1-box A, -boxb, and full-length HMGB1 The plasmids pt7-hmgb1ba, pt7-hmgb1bb, and pet14b-hmgb1 used for the expression of rat HMGB1 box A, box B, and full-length HMGB1 respectively, have already been described. The procedure used for the expression and purification of the three recombinant proteins was as described before (17). Native calf thymus HMGB1 was purified as described before (24). Peptide phage display analysis with HMGB1 boxes A and B A phage display heptapeptide library kit (New England Biolabs, Beverly, Ma) was used to screen for peptides binding to HMGB1-box domains A and B. The kit contained a random combinatorial collection of heptapeptides fused via a flexible linker sequence to the N-terminus of protein piii of bacteriophage M13. Each phage expressed at the tip of the cover 3 to 5 copies of the unique peptide it encoded. The library complexity contained all the possible combinations of the 20 natural amino acids taken as 7-mer sequences. For the phage biopanning process, we followed the kit instructions as indicated by the manufacturer. Four independent experiments were run in which 30-40µg HMGB1 box A or B were immobilized overnight at 4 C on 96-wells microtiter plates (Costar 3690). Wells were then blocked for 1 h at 4 C with TBS-0.1% Tween 20 and 2x10 11 plaque forming units (pfu) of the phage library were added per well and incubated for an additional hour at room temperature. Wells were washed ten times with TBS- 0.1% Tween 20 for the first round. For the subsequent rounds washing used TBS- 0.5% Tween 20 to increase stringency. Finally, phages were either eluted at room temperature by incubation with 0.2 M glycine-hcl, ph 2.2 for 10 min or affinity eluted by incubation for 1 h with 30-40µg of the respective HMGB1 box A or B. Phage amplification, titration, and purification were carried out according to the manufacturer protocol. Automatic phage DNA sequencing used the -96gIII primer (5 CCCCTCATAGTTAGCGTAACG3 ) and was performed by the "Serveis Científico-Tècnics" of the "Universitat de Barcelona". The search for homologies in the data banks with the sequences selected
6 in the phage display experiments was performed using the Blast (Basic Local Alignment Search Tool) program (NCBI, National Center for Biotechnology Information, USA) (25). The alignments were obtained using the MultAlin program (Multiple Sequence Alignment, INRA, Institut National de Recherche Agronomique, Toulouse, France) (26). GST-fusion constructions and protein purification All GST-constructs were prepared using the pgex-4t3 plasmid (Amersham Pharmacia Biotech). The twelve peptides selected to be further analyzed as GST fusions were amplified from the phages by using PCR with the following primers 5 TGGTACCTTTGAATTCTCACTC 3 and 5 TCAACAGTGTCGACCGAACC 3 which introduced EcoRI and SalI sites, respectively (underlined). The PCR products were inserted between these sites in the pgex-4t3 plasmid. pgex-grg1 and pgex-grg5 were constructed by inserting the NaeI-XhoI fragments obtained from the pbs-grg1 and pbs-grg5 (kindly provided by Dr. C. Lobe) into pgex-4t3 digested with SmaI and XhoI. pgex-grg Q-GP-CcN was produced by inserting a NaeI/SmaI fragment from pbs-grg1 into SmaI site of pgex vector. pgex-grg SP-WD was generated by inserting a SmaI/XhoI fragment of pbs-grg1 into the same sites of the pgex vector. pgex-grg Q was obtained by digesting pgex-grg1 with Bpu1102I and XhoI, made blunt and self-ligated. pgex-grg GP-CcN was prepared by digestion of pgex-grg1 Q-GP- CcN with BamHI and Bpu1102I, made blunt and self-ligated. pgex-grg GP-CcN was obtained by inserting a BamHI-XhoI fragment obtained by PCR from the pgex-grg1 construct by using the primers 5 TTCAGCCTCCTGGATCCCCG 3 and 5 GTTTTCTCGAGGTGAGTGTG 3 (restriction sites underlined) into pgex 4T3 digested with the same enzymes. pgex-grg SP was generated by inserting a SmaI-XhoI fragment obtained as above by PCR with primers 5 ACTCACCCCGGGAAAACG 3 and 5 GTTGATCTCTCGAGCATGTCG 3 into pgex 4T3 digested with the same enzymes. All constructions were verified by manual or automated DNA sequencing. pgex-mecp2 ( ) was obtained from Dr. A. Bird. pgex-kg-hn RNP K was a gift of Drs. S.K. Jong and J.H. Kim. pgex-prb ( ) and
7 pgex-p53 were obtained from Dr. M.A. Martínez-Balbás. All the GST fusions were expressed in E. coli BL21(DE3) and purified according to standard methods as suggested by the manufacturer (Amersham Pharmacia Biotech). Far-Western analysis and Western blotting GST-peptide fusions were separated by SDS-PAGE and electroblotted in transfer buffer (25 mm Tris, 40 mm Glycin, 0.05% SDS, 20% methanol) to nitrocellulose membranes (Optitran BA-85, Schleicher&Schuell). After blocking in PBST (PBS-0.1% Tween20) containing 5% non-fat dry milk for 1-2 h at room temperature, membranes were incubated overnight in 10 ml of D buffer without glycerol (20 mm HEPES, ph7.9, 0.2 mm EDTA, 100 mm NaCl, 0.5 mm DTT, 0.1 mm PMSF) containing 10-to-20 µg of either HMGB1 or its derivatives HMGB1 box A or B. After several washes in PBST membranes were incubated with a primary chicken anti-hmgb1 antibody raised in our lab against recombinant HMGB1 deleted of the C-ter domain, subsequently with a secondary anti-chicken IgY-HRP antibody (Jackson laboratories, USA) and detected using ECL reagents. Pull-down assays Glutathione-Sepharose beads (Amersham Pharmacia Biotech) were loaded with the different GST-fusion proteins as suggested by the manufacturer and washed 5 times with 450 µl of D buffer containing 20% glycerol. Then, they were incubated with HMGB1 box A or B for 1 h at 4 C in the same buffer and washed 6 times more with the same buffer. Beads were finally boiled in protein loading buffer, proteins were separated by SDS-PAGE and detected by Western blotting with specific antibodies. Results Preferential interaction of HMGB1-box domains A and B with some peptides In an attempt to identify targets that could be potentially recognized by HMGB1, and since other general approaches like yeast two-hybrid analysis were not possible (likely due to toxic effects of the expression of HMGB1-boxes in yeast, results not shown) a peptide library screening approach was carried out. A highly complex library containing the whole collection of natural heptapeptide sequences displayed on phage M13 was used. Four rounds of selection were
8 carried out for each of the four independent experiments that were performed using highly purified recombinant HMGB1 boxes A or B as bait proteins. During the biopanning process stringency was increased by using higher detergent concentrations in the washing buffer. Bound phages were recovered by either a non-specific acid-elution (exp. 1) or by affinity competition using HMGB1-boxes A or B free in solution (exps. 2-4). Several peptides were selected as potentially interacting with HMGB1 with some specificity (Table I). From those results, it became clear from their sequences that they were not related to a single strong consensus sequence, suggesting that either the interactions between HMGB1 and the peptides were weak or that HMGB1 boxes could interact with several unrelated motifs. We noted that frequencies of appearance of some amino acids in the selected peptides clearly deviated from the random theoretical level indicating that the selection process was successful. That is, if interactions of HMGB1 were specific they should be independent of the particular growth features of the phages, their statistics should clearly differ from those of the unselected phages in the biopanning assays and be due to their ligand-binding requirements. For example, positively and negatively charged amino acids usually presented frequencies lower than expected in the selected peptides suggesting that the interactions did not mainly rely on electrostatic forces despite the highly basic character of the HMGB1 boxes. Also, a high rate of aromatic residues was observed, in particular a high level of tryptophan in experiment number 4, which contrasts with the observation that this amino acid tends to decrease naturally without selection. A remarkable level of proline was also obtained in experiment number 3. Hydrophylic amino acids which tend to be involved in hydrogen-bond recognition, showed some decrease as well. These data indicated that selection had in fact occurred in the presence of the HMGB1 boxes although no clear-cut consensus could be easily drawn. Despite sequences selected were very variable as shown in Table I, the appearance of several copies of the same peptide in each experiment indicated a high enrichment and specificity in the screening. Note also that some peptides (e.g. HWGMWSY, HAIYPRH) were selected in independent experiments with both HMGB1 domains. Despite this variability in the peptide sequences it was also clear that amino acid distribution in the peptides was not at random (Table I). Thus, peptides could be grouped at least in two classes: some rich in proline and some others rich in tryptophan. Two peptides enriched in tryptophan, HWGMWSY and HSWLWWP, accounted for 50% of the clones in experiment 4 and
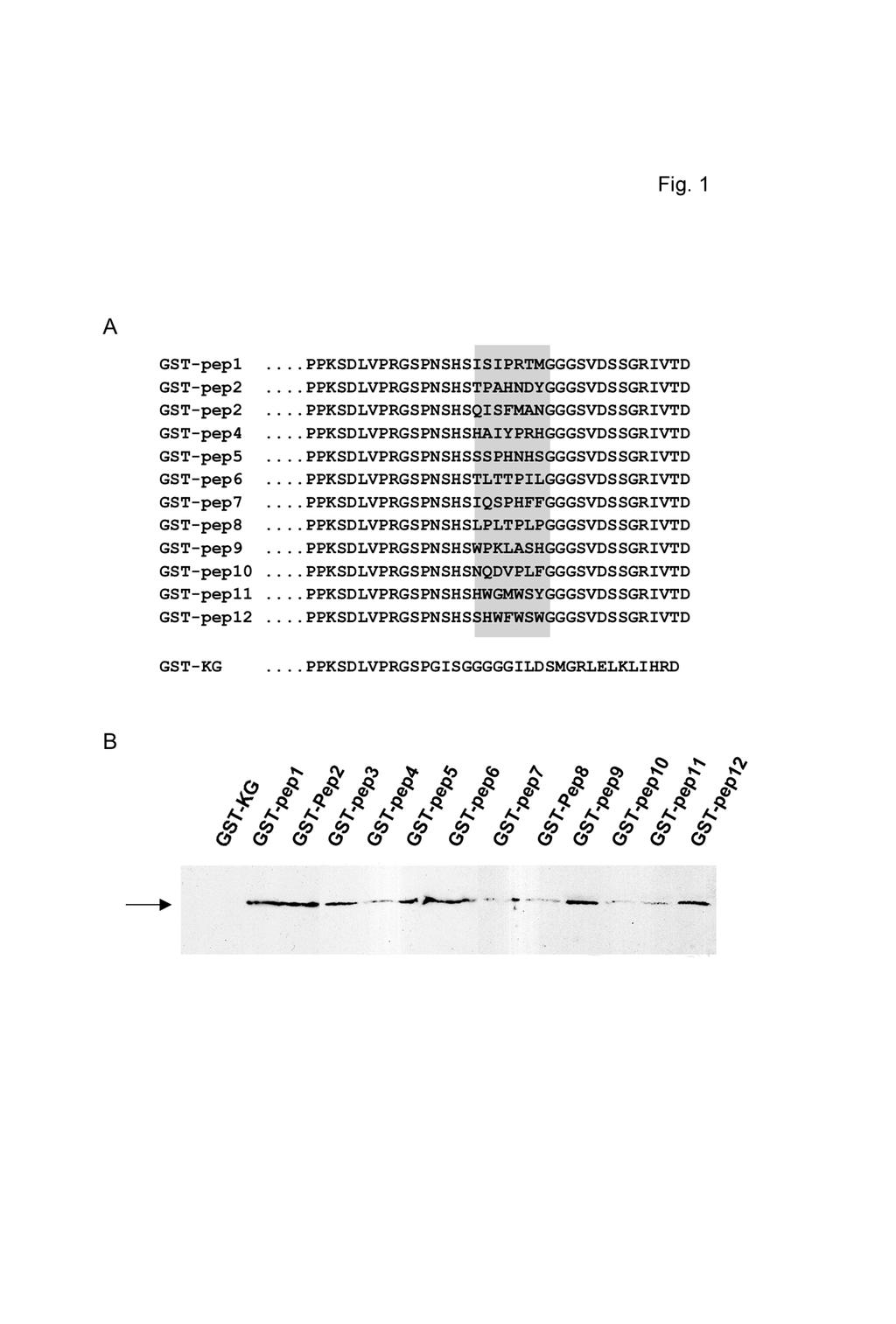
9 HWGMWSY appeared also in experiments 1 and 2. A minimal consensus WxxW motif could be a potential site for interaction. In the case of proline-enriched peptides it was more difficult to define a consensus given the diversity of these sequences. Because of the high complexity in the peptide sequences retrieved in the phage display experiments we were concerned about the potential existence of false positives in the selected set of peptides. As a second approach to confirm the bona fide association of HMGB1 with those peptides, we perfomed in vitro assays in which peptide sequences were fused to GST. The peptides used were twelve among those selected with HMGB1 boxes A and B and were representative of the different kinds of peptides obtained. They were fused to the C-terminus of GST to facilitate their expression and purification in E. coli. As a control a GST with an extended, unrelated and never selected peptide was included (GST-KG). Since direct GST pull-down assays did not work likely because of steric hindrance of the GST moeity to the very small peptide (not shown), Far-Western experiments were performed (Fig. 1). In contrast to the negative control, all the other peptides showed interaction with HMGB1 box B (Fig. 1B). The relative intensity of each band varied from experiment to experiment indicating that the results were not quantitative. Nevertheless, these interactions were always detectable whereas interaction with the control was never detected (Fig1B, lane GST-KG). Moreover, the same assays were also done using either HMGB1 box A or HMGB1 full-length and results were the same (not shown). We noted that none of the peptides selected was represented in the sequence of HMGB1. Since HMGB1 can only very inefficiently interact with itself forming a homodimer (27) and HMG-box domains do not interact with each other inside the HMGB1 molecule (28) this was also suggesting that peptides selected were representative of reasonable rather than very weak interactions. Discovery of new partners for HMGB1 Once our results were confirmed it became interesting to make a survey in order to look for proteins containing regions of homology to the peptides selected in the phage display assay. This point was addressed by searching the available databases with the Blast program for nuclear proteins. HWGMWSY was a peptide that generated interesting candidates and among them appeared ER, a factor previously identified to interact with HMGB1 (15). We noted a potential WxxW consensus motif (Table IIB) for the factors belonging to this group. These factors
10 were not studied in detail because in comparison another peptide, LPLTPLP, generated the most interesting homologies and allowed the identification of a new set of nuclear proteins that could potentially interact with HMGB1. These putative factors were summarized in Table IIA. Once again, two other factors - p53 and PR - already described to interact with HMGB1 appeared (10,14) and interestingly, the region homologous to our peptide in p53 was highly conserved in mammals. Remarkably, all the proteins listed presented a potential PxxPxP consensus motif, and among them components for many transcription factor complexes can be found. In this list we had only included the proteins containing the sequence motifs homologous to the peptide that were conserved among mammalian species. Note that in some cases two conserved motifs could be found (e.g. Grg1, p53). Also, the two orientations of the motif were considered because similar proline-rich motifs were reported to be recognized by SH3 domains independently of their orientation ((29) and references therein). To confirm that HMGB1 could physically interact with the surveyed proteins we used a pull-down approach employing GST-p53 as a positive control and some other proteins fused to GST that were selected as representative of the several classes described above: the RNA binding protein and positive regulator hnrnp K, the negative regulator MeCP2, and co-repressors prb, Grg1 and Grg5. Since HMGB1 is a protein that presents many posttranslational modifications - at least including acetylation, ADP-ribosylation and methylation (reviewed in (30)) - we have taken this fact into account and have analyzed in parallel the interaction with recombinant and native calf thymus purified HMGB1 (i.e. presenting many posttranslational modifications, not shown). Fig. 2 shows that HMGB1 can effectively interact with all the proteins tested with some notable differences between the recombinant and the purified forms. Whereas either recombinant or purified HMGB1 interacted with Grg1 to a similar extent, recombinant HMGB1 showed some preference for Grg5, p53, hnrnp K, and prb. Purified HMGB1, in contrast, interacted with MeCP2 more efficiently than recombinant HMGB1. Neither recombinant nor purified HMGB1 could interact with GST. We wanted to emphasize, however, that since those results were semiquantitative only large differences should be taken into consideration. Thus, it was tempting to suggest that whereas posttranslational modification did not seem to have a major effect on HMGB1 interaction with some of them (mainly Grg1, but also Grg5 to some extent) it looks like negatively affecting the interaction with some others (especially prb and p53) and in one case being slightly preferred (MeCP2). In this assay the
11 efficiency of the interaction was dependent on the fusion protein. Preliminary data obtained using recombinant HMG-boxes A and B indicated the same results (not shown). Mapping of the interacting regions on Grg1 We further studied the potential of the PxxPxP motif by using Grg1 as a model target. Grg1 is structured in five domains each of them having a particular function (Fig. 3A). A highly conserved N-ter glutamine-rich domain (Q) is involved in protein oligomerization, a C-ter WD-repeat domain is used for interaction with other proteins, and a central domain encompassing the CcN motif allows the nuclear localization of Grg1. The two other domains, GP and SP, are poorly conserved and may play a direct role in repression of transcription (31). By using different deletions of Grg1 the involvement of the PxxPxP motif in the interaction with HMGB1 was tested (Fig. 3A). As shown in Fig. 3B splitting of Grg1 molecule in two moeities, N- and C-terminal, showed that HMGB1 was able to interact with both, the one containing domains Q-GP-CcN (lane 4) and the other containing domains SP-WD (lane 8) respectively. The binding site on the N-terminal moeity was located at the N-terminal region of the GP domain since deletion encompassing residues 131-to-155 (Fig. 3B, compare lanes 6, and 7, GST-GP-CcN and GST- GP-CcN, respectively) completely abolished the interaction. Additionally, the Q domain clearly showed that it could not interact with HMGB1 either (lane 5). The sequence that upon deletion abolished binding of HMGB1 on this side of the protein precisely corresponded to the region of homology to the LPLTPLP peptide (Fig. 3C, shadowed). Note that in the closely related Grg5 factor this region is also present and is likely used for the interaction with HMGB1 as well (Fig. 2). The site of interaction in the C-terminal moeity is likely located at the SP domain since deletion of WD domain does not abolish it. Nevertheless, the existence of still another site on the WD domain cannot be rouled out. A careful examination of the amino acid sequence of the SP domain revealed another region of weaker homology (vpfppmp) to the LPLTPLP peptide (Fig. 3D). In this case, this site is absent in the related Grg5 factor. Discussion On the HMGB1 side
12 HMGB1 was first isolated almost thirty years ago and since then much effort has been put to characterize this protein and find its functional role (30). The latter point has shown to be difficult because of controversial results and it is still a matter of discussion. Since HMGB1 does not present any enzymatical activity on its own or any sequence specific DNA or RNA binding, it is clear that whatever the functions in which it could be involved it would require the assistance of other factors, likely proteins. Several reports have clearly shown that HMGB1 interacts with a set of different factors that present very little, if any, sequence or structure in common. The question of how HMGB1 can recognize its partners is the one we have addressed here. So far, the rules that govern the interactions between two given proteins are fairly unknown and in any case interactions are impossible to predict. An hypothesis to reconcile all the data on HMGB1 interactions could be that the interaction surface recognized by HMGB1 on these proteins might be very short, so that only few amino acids would be really important. Eventually, not only one of such motifs could be recognized by HMGB1. This idea is based on recent findings showing that many different proteins can recognize motifs as short as four to five residues long as for example, LxCxE for prb (32) or WRPY/W for Groucho in Drosophila (33). As an approach that sometimes helped in determining the residues that might be required for the interactions to take place, the screening of peptide repertoires has been successfully used by several groups to shed some light on the mechanisms of ligand recognition of macromolecular complexes. The use of a peptide phage display approach allowed us to begin understanding some of the previously reported interactions, to predict for some others and to begin testing them. In our hands, HMGB1 can recognize several short motifs through the HMGboxes. However, the lack of a unique sequence motif or a single consensus sequence derived from them complicated the interpretation of the results and clearly suggested that HMGB1 is a protein that does not have a highly preferred sequence to interact but can establish interactions with many peptides having rather different sequences. Therefore, no strong enrichment is observed and this heterogeneity makes difficult the interpretation of the results. As far as we know, this seems to be a particular behaviour of HMGB1. Some of the peptides were isolated with both HMG-boxes A and B (HWGMWSY and HAIYPRH) suggesting that either the recognition with these peptides is due to the three-dimensional structure of the HMG-box or uses some of the most highly conserved regions
13 between the two domains. The other peptides seem to be different for each HMGbox. However, the complexity of the patterns obtained suggests that the relative affinities for the peptides are similar and weak. This may in turn enrich a particular experiment in a certain set of peptides because of slight changes (in temperature for instance). But it could also be attributable to the differences in sequence of the two HMG-boxes since their three-dimensional structures are rather similar (6,8). On the other hand, taken into account that sequence identity in HMG-boxes A and B is really high when compared to the corresponding domains in HMGB2 it is also very likely that many if not all of the peptide motifs will also be recognized by HMGB2 as well. In fact, the apparent redundancy of HMGB1 and HMGB2 is a general observation for all of the interactions described so far for these proteins including Oct factors (11,12), steroid receptors (15), htbp (our unpublished results), and RAG1 (9) among others. Our results do not discard the highly acidic C-terminal domain as potentially interacting with other factors. In fact, the C-ter domain is the one involved in the interaction with histone H1 (34) and with the histone dimer H2A.H2B as well (24), and recently it has been claimed to interact with the glutamine-rich N-ter domain of human and Drosophila TBP (35). However, the highly acidic nature of this domain makes its interactions highly electrostatic and in general weakly specific. New HMGB1-interacting proteins come to light The use of peptides in our assay is likely limiting the set of potential partners for interaction with HMGB1 to those that are recognized only by sequence because little structure can be expected from heptapeptides. Therefore, it is reasonable to expect some of the interactions previously described not to appear in this assay. This may be the case, for instance, of the interaction to the H2 α-helix of htbp previously described (17). Moreover, the fact that a particular protein contains one of the motifs in its sequence does not necessarily imply that this will be the site used for interaction or that another motif can be used. In addition, the contribution of peptidic sequences around the motifs identified here can be determinant, as in some cases have been shown to play a major role either in modulating or even in affecting the specificity the interactions. Additionally, the accessibility of the motifs embedded in some protein contexts might be rather limited. Finally, there is always a potential for HMGB1 to be actively recognized by the other partner (see below) and then motifs can be completely different.
14 When searching protein databases, a point that must be taken into account is that the relative representation of the different peptides and motifs is not homogeneous. Thus, some peptide motifs produce long lists of proteins, like for PxxPxP, whereas other motifs do not. Nevertheless, some motifs not studied here in detail also generated very interesting candidates. For instance, a sequence closely related to the potential WxxW motif - and in particular to peptide HWGMWS - can be found in conserved sequences of the α- and β-er of mouse, rat and human, of ETS-1 and also in a conserved sequence of the mouse and human RNA-binding protein nucleolysin TIAR among others (Table IIB). Remarkably, the interaction with α-er was previously described. Moreover, the proposed HMGB1-binding sequence lies very close to the Zn-finger of the ER that interacts with DNA and is clearly accessible in the co-crystal structure (15,36). These features might help to explain how can HMGB1 stabilize the interaction of ER with DNA. The PxxPxP consensus motif has predicted the most interesting partners upon Blast search and for that reason has been studied in detail. These include prb, MeCP2, hnrnp K, Grg1, Grg5 and p53. Remarkably, p53, which was previously described to interact with HMGB1, appeared giving credit to this linear sequence as a potential HMGB1 recognition motif. In fact, all the PxxPxP containing proteins tested here showed to interact with HMGB1. The reason for this clear result is likely due to the high proline content of this motif, that difficults the adoption of a defined secondary structure even if embedded in a protein sequence. Thus, it is very likely that other proteins containing this motif and listed in Table IIA will interact with HMGB1 as well. HMGB1 was already described to interact with p53 and be a unique activator of this factor (10). The PxxPxP motif is always found at the N-terminus of p53 around position in the different species and forming part of a prolinerich domain that negatively affects p53 interaction with DNA (37). The interaction of HMGB1 at this domain could explain the stimulatory effects observed on DNA binding and transactivation (10). Since the proline-rich domain is dispensable for transactivation (38) the role of HMGB1 might be restricted to stabilize the p53- DNA complex. We must mention, however, that residues 363-to-376 in the basic domain of p53 have been recently reported to recognize HMG-box A on HMGB1 and that p53 conversely enhanced HMGB1 interaction with cisplatin-modified DNA (22). In this region of p53 no homology to any of the peptides identified in this work can be found. This might be in apparent contradiction to the data discussed
15 above but they could be reconciled if there is mutual recognition between p53 and HMGB1. If so, HMG-box A would be recognized by the C-terminal region of p53 and the proline-rich domain of p53 would be recognized by the HMG-boxes of HMGB1. Depending on the experimental conditions one or the other interaction would prevail and be selected. This may also be the case of prb because in addition of having a sequence showing homology to the PxxPxP motif, the "pocket" region of prb can recognize the LxCxE motif in other proteins (32). This motif (as the sequence LFCSE) occurs to be present in the HMG-box B of both HMGB1 and HMGB2 and is absolutely conserved in vertebrates. We have not analyzed here in detail whether recognition is via the LxCxE motif (i.e. prb recognizes HMGB1) or the PxxPxP motif in prb (i.e. HMGB1 recognizes prb) and simply described this interaction. Although it is possible that the interaction may work in both directions we reason that it looks difficult since the LFCSE sequence is structured into an α-helix facing the concave side of the L-shaped domain and is partially buried in HMG-box B (6). The interaction between HMGB1 and Grg1 has been studied in more detail because of the high homology to the peptide and also because this is the first interaction of HMGB1 with a co-repressor. In Grg1 two regions of interaction with HMGB1 have been uncovered. The one corresponding to the GP domain fits nicely with high homology to the LPLTPLP peptide isolated in the phage display analysis and very likely accounts for that interaction as predicted. In the second domain interacting with HMGB1, the SP domain, also a region with the PxxPxP motif is present albeit homology to the original peptide sequence is weaker. These results support the PxxPxP motif as a good recognition sequence for HMGB1. Brantjes et al. have recently described that all Tcf HMG-box transcription factors interact with Groucho-related co-repressors and even more, all the long members containing five domains (here represented by Grg1) mediated repression of the Tcfs, whereas the short member (Grg5) mediated de-repression. All Tcfs are HMG-box transcription factors but so far the region that interacts with the Grgs has never included the HMG-box domain. At a difference HMGB1 uses the HMGbox domains to interact with Grg proteins suggesting that recognition of the PxxPxP motif is not a feature common to all HMG-box domains. On the other hand, transcription mediated by AR is inhibited by Grg5 (39). Since the PxxPxP motif is also present in the AR sequence (Table IIA) and the interaction of HMGB1 with AR stabilizes the AR-DNA complex as shown by others, the interaction of HMGB1 with Grg5 could cooperate by displacing it from AR and
16 helping to relieve inhibition (15). MeCP2 is a protein involved in the recognition of methylated DNA at CpG islands and is closely related to gene silencing (see (40) for a review). The HMGB1 binding motif is located at the C-terminus of this protein (residues ) in a region where no other factor has been shown to interact. Therefore, it is possible that HMGB1 interaction does not interfere the binding of the many factors known to interact with MeCP2, (among other msin3a, NcoR and c-ski) and remains to be analyzed whether in this case this interaction could also stabilize MeCP2-DNA complexes as done by HMGB1 in other complexes. hnrnp K presents a remarkable variety of protein interactions with some factors involved in signal transduction, and others involved in several aspects of gene expression (reviewed in (41,42)). This diversity suggests that hnrnp K protein may act as a docking platform or as a scaffold protein within multiple functional modules. Then, HMGB1-hnRNP K interaction may connect two systems which have a large protein-protein interaction potential to extend their respective domains of action even further. For example, hnrnp K has shown to be a transcription activating factor for the c-myc promoter (43,44). Although we have not been able to establish a direct connection, it is remarkable that c-myc mrna levels in HMGB1 -/- mouse cells are about 5-to-10 fold lower than those in wt cells (unpublished results). We have focus so far on the nuclear environment where HMGB1 is expected to play a role in gene expression. However, HMGB1 is an extraordinary protein and besides of being a nuclear protein in most cases it is also true that for more than ten years HMGB1, under the name of amphoterin was found on the outer cell membrane of some cell types (45). It is now clear that HMGB1 could be actively released outside the cell in response to tumor necrosis factor and interleukin 1 (46), and passively by necrosis or cell damage in a variety of cell types, mainly immature and transformed cells (47). Release of HMGB1 induces some pathological processes as a potent late mediator of endotoxin lethality and inflammation. Many of these phenomena require the interaction of HMGB1 with the receptor for advanced glycation products (RAGE) (reviewed in (48)). Examination of the mouse RAGE amino acid sequence shows a region highly homologous to the LPLTPLP motif in the second Ig-like C2-type domain that is highly conserved among several species. Similarly, HMGB1 was also shown to interact with Syndecan-1, a cell surface heparan sulfate-rich proteoglycan (49) in which sequence a PxxPxP motif can also be found. These and other data suggest
17 that HMGB1 might use the same motifs to interact with cell surface proteins and with nuclear factors. Final considerations Prior to this work, HMGB1 already had a long list of proteins to which it interacts. Along with the new candidates reported here it may give the impression that HMGB1 interacts with almost every protein in the cell. Despite HMGB1 is in fact a "sticky" protein (48), that is by no means true. For example, we have been trying to extend the initial interaction of HMGB1 with human TBP to other factors of the general transcription machinery and we failed to observe any interaction of HMGB1 with TFIIA, TFIIB, TFIIF, the CTD of RNA pol II, TFIIE (despite a clear interaction can be observed with the p56 subunit no interaction at all can be observed with the native tetramer), and a TBP-related factor among others (unpublished results), suggesting that the list of factors interacting with HMGB1 may be long but by no means indiscriminate. This particular behaviour makes difficult to attribute a defined role to HMGB1 in the organism but may explain the general weakness observed in the HMGB1 knock-out mice that lead them to death 24 hours after birth (2). HMGB1 seems to have developped a high potential for protein-protein interaction with multiple partners, always taking part of a macromolecular complex and either assisting in the assembly or stabilizing the assembled factors. This feature along with the remarkable abundance of HMGB1 in the nucleus and the many posttranslational modifications that undergoes could explain its general involvement in nuclear processes and its modulation. On one hand, HMGB1 is a protein that can interact with angled DNA on its own. On the other hand, a general observation is that upon interaction HMGB1 can help stabilizing many proteins previously bound to DNA. These functions may be compatible but whereas the concave region of the L-shaped HMG-box domain clearly is the DNA interaction site, there is no data about the region of the HMGbox domain involved in the interaction with other protein factors. Some evidence at the enhanceosome of BHLF-1 suggests that HMGB1 can at the same time interact with the narrow groove of DNA and with protein factors (50). However, more detailed work is required to know whether HMGB1 could simultaneously interact with proteins and DNA, the stoichiometry of the interactions and how its binding can stabilize protein-dna complexes. A potential role of co-factor is emerging for HMGB1 in many different processes.
18 Acknowledgements We are very grateful to Dr C. Lobe (Molecular Biology Institute, McMaster University, Ontario) for generously provinding cdnas for Grg1 and Grg5, Dr A. Bird (Institute of Cell and Molecular Biology, University of Edinburgh) for the pgex/mecp2 plasmids, Drs. S. K. Jang and J. H. Kim (Department of Life Science, Pohancy University of Science and Technology, Korea) for the pgex- KG/hnRNP K plasmid, and Dr M. A. Martínez-Balbás (Institut de Biologia Molecular de Barcelona, CSIC) for the pgex/p53 and pgex/prb plasmids. We also thank Mr J. Font for making Figure 3. This work was supported by a European Union Grant FMRX-CT and Comissió Interdepartamental de Recerca i Innovació Tecnològica (CIRIT) of the Generalitat de Catalunya Grant SGR This work was carried out in the context of the Centre de Referència en Biotecnologia of the CIRIT of the Generalitat de Catalunya. References 1. Bustin, M. (2001) Trends Biochem. Sci. 26, Calogero, S., Grassi, F., Aguzzi, A., Voitländer, T., Ferrier, P., Ferrari, S. and Bianchi, M. E. (1999) Nat. Genet. 22, Bustin, M. and Reeves, R. (1996) Prog. Nucleic Acids Res. Mol. Biol. 54, Bustin, M. (1999) Mol. Cell. Biol. 19, Thomas, J. O. and Travers, A. A. (2001) Trends Biochem. Sci. 26, Read, C. M., Cary, P. D., Crane-Robinson, C., Driscoll, P. C. and Norman, D. G. (1993) Nucleic Acids Res. 21, Weir, H. M., Kraulis, P. J., Hill, C. S., Raine, A. R., Laue, E. D. and Thomas, J. O. (1993) EMBO J. 12, Hardman, C. H., Broadhurst, R. W., Raine, A. R. C., Grasser, K. D., Thomas, J. O. and Laue, E. D. (1995) Biochemistry 34, Aidinis, V., Bonaldi, T., Beltrame, M., Santgata, S., Bianchi, M. E. and Spanopoulou, E. (1999) Mol. Cell. Biol. 19, Jayaraman, L., Moorthy, N. C., Murthy, K. G. K., Manley, J. L., Bustin, M. and Prives, C. (1998) Genes Dev. 12, Zwilling, S., König, H. and Wirth, T. (1995) EMBO J. 14,
19 12. Butteroni, C., De Felici, M., Scholer, H. R. and Pesce, M. (2000) J. Mol. Biol. 304, Zappavigna, V., Faciola, L., Citterich, M. H., Mavilio, F. and Bianchi, M. E. (1996) EMBO J. 15, Oñate, S. A., Prendergast, P., Wagner, J. P., Nissen, M., Reeves, R., Pettijhon, D. E. and Edwards, D. P. (1994) Mol. Cell. Biol. 14, Boonyaratanakornkit, V., Melvin, V., Prendergast, P., Altmann, M., Ronfani, L., Bianchi, M. E., Taraseviciene, L., Nordeen, S. K., Allegretto, E. A. and Edwards, D. P. (1998) Mol. Cell. Biol. 18, Ge, H. and Roeder, R. G. (1994) J. Biol. Chem. 269, Sutrias-Grau, M., Bianchi, M. E. and Bernues, J. (1999) J. Biol. Chem. 274, Lu, W., Peterson, R. and Dasgupta, A., Scovell, W.M. (2000) J. Biol. Chem. 275, Costello, E., Saudan, P., Winocour, E., Pizer, L. and Beard, P. (1997) EMBO J. 16, Shykind, B. M., Kim, J. and Sharp, P. A. (1995) Genes Dev. 9, Van Gent, D. C., Hiom, K., Paull, T. T. and Gellert, M. (1997) EMBO J. 16, Imamura, T., Izumi, H., Nagatani, G., Ise, T., Nomoto, M., Iwamoto, Y. and Kohno, K. (2001) J. Biol. Chem. 276, Kay, B. K., Kasanov, J., Knight, S. and Kurakin, A. (2000) FEBS Lett. 480, Bernués, J., Espel, E. and Querol, E. (1986) Biochim. Biophys. Acta 866, Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W. and Lipman, D. J. (1997) Nucleic Acids Res. 25, Corpet, F. (1988) Nucleic Acids Res. 16, Ranatuga, W., Lebowitz, J., Axe, B., Pavlik, P., Kar, S. R. and Scovell, W. M. (1999) Biochim. Biophys. Acta 1432, Ramstein, J., Locker, D., Bianchi, M. E. and Leng, M. (1999) Eur. J. Biochem. 260, Kay, B. K., Williamson, M. P. and Sudol, M. (2000) FASEB J. 14, Johns, E. W. (1982) The HMG Chromosomal Proteins. Johns, E. W., Academic Press, London, 31. Chen, G. and Courey, A. J. (2000) Gene 249, 1-16
20 32. Dahiya, A., Gavin, M. R., Luo, R. X. and Dean, D. C. (2000) Mol. Cell. Biol. 20, Fisher, A. L. and Caudy, M. (1998) Genes Dev. 12, Carballo, M., Puigdomènech, P. and Palau, J. (1983) EMBO J. 2, Das, D. M. and Scovell, W. M. (2001) J. Biol. Chem. 276, Schwabe, J. W. R., Chapman, L., Finch, J. T. and Rhodes, D. (1993) Cell 75, Müller-Tiemann, B. F., Halazonetis, T. D. and Elting, J. J. (1998) Proc. Natl. Acad. Sci. USA 95, Walker, K. K. and Levine, A. J. (1996) Proc. Natl. Acad. Sci. USA 93, Yu, X., Li, P., Roeder, R. G. and Wang, Z. (2001) Mol. Cell. Biol. 21, Bird, A. and Wolffe, A. P. (1999) Cell 99, Siomi, H. and Dreyfuss, G. (1997) Curr. Opin. Genet. Dev. 7, Shyu, A.-B. and Wilkinson, M. F. (2000) Cell 102, Tomonaga, T. and Levens, D. (1995) J. Biol. Chem. 270, Takimoto, M., Tomonaga, T., Matunis, M., Avigan, M., Krutzsch, H., Dreyfuss, G. and Levens, D. (1993) J. Biol. Chem. 268, Parkkinen, J., Raulo, E., Merenmies, J., Nolo, R., Olavi Kajander, E., Baumann, M. and Rauvala, H. (1993) J. Biol. Chem. 268, Wang, H., Bloom, O., Zhang, M., Vishnubhakat, J. M., Ombrellino, M., Che, J., Frazier, A., Yang, H., Ivanova, S., Borovikova, L., Manogue, K. R., E., F., Abraham, E., Andersson, J., Andersson, U., Molina, P. E., Abumrad, N. N., Sama, A. and Tracey, K. J. (1999) Science 285, Degryse, B., Bonaldi, T., Scaffidi, P., Müller, S., Resnati, M., Sanvito, F., Arrigoni, G. and Bianchi, M. E. (2001) J. Cell. Biol. 152, Müller, S., Scaffidi, P., Degryse, B., Bonaldi, T., Ronfani, L., Agresti, A., Beltrame, M. and Bianchi, M. E. (2001) EMBO J. 20, Salvimirta, M., Rauvala, H., Helenius, K. and Jalkanen, M. (1992) Exp. Cell Res. 200, Elwood, K. B., Yen, Y.-M., Johnson, R. C. and Carey, M. (2000) Mol. Cell. Biol. 20,
21 Footnote 1 The abbreviations used are: HMGB1, High Mobility Group protein 1; prb, Retinoblastoma protein; hnrnp K, heterogeneous nuclear ribonucleoprotein K; MeCP2, Methyl-CpG binding protein 2; Grg1, Groucho-related gene protein 1; Grg5, Groucho-related gene protein 5; GST, Glutathione S-transferase. Figure legends Table I Amino acid sequences of the heptapeptides bound to HMG-boxes A and B of HMGB1. Heptapeptide amino acid sequences deduced from phage sequencing are recapitulated here for each independent experiment. a Peptides eluted unspecifically by low ph b Peptides eluted by affinity with the same HMG-box used for the biopanning c Number of peptides sequenced per experiment The number of clones encoding the same peptide for each experiment is indicated in brackets. Table II Some potential candidates for interaction with HMGB1. Blast search results of homology for the heptapeptides LPLTPLP (A) and HWGMWSY/HSWLWWP (B). The name of the candidate protein is indicated on the left column, the sequence of homology to the query peptide with indication of the residue numbers for each case is shown in the middle column, and the accession number is quoted on the right column. Figure 1 HMGB1 interacts with twelve different peptide sequences A. Scheme of the C-terminal amino acid sequences of the twelve GST-peptide fusions analyzed. Residues boxed in gray correspond to the heptapeptide sequences. Residues around them correspond to the sequence of piii protein of M13 in which they were included. GST-KG was the negative control used for the
22 experiments. B. Far-western assay of the twelve GST-peptide fusions described above and GST-KG as a negative control probed with HMGB1 box B. Figure 2 HMGB1 interacts with several transcriptional regulators Pull-down assay of recombinant (r) and purified (p, highly modified) HMGB1 with GST fusions of p53, prb, hnrnp K, MeCP2, Grg1 and Grg5 (see Experimental Procedures for details). GST alone was used as negative control. Lanes containing 10% of the input material and corresponding to recombinant (Input r) and purified (Input p) HMGB1 are shown on the left and the right, respectively. In order to analyze the effect of posttranslational modifications on the interactions of HMGB1, equal amounts of GST-fusions were used with the two HMGB1 preparations. Recombinant HMGB1 migrates slightly above purified HMGB1 because of the addition of a ~1kD histidine-tag. Figure 3 Determination of HMGB1 binding sites on Grg1 A. Scheme of the structure of Grg1 in five domains (Q, GP, CcN, Sp, and WDrepeat) and of the GST fusions used below. The black and white arrows indicate the positions of the potential HMGB1 binding sites. B. Mapping of the HMGB1 binding sites on Grg1 by pull-down analysis. GSTfusions described in A were used to map the interaction of recombinant HMGB1. Lane 1 shows 10% of the input used for the experiments and lane 2 GST as negative control. C. Alignment of the amino acid sequence homologous to the LPLTPLP and corresponding to the black arrow. D.Alignment of the amino acid sequence homologous to the LPLTPLP and corresponding to the white arrow.
23 Table I Exp. 1 box A a HWGMWSY NWGMWSY PHWTWVL HMSKPVQ SSGTHAK YNINIRP NYTQTVP c 7 Exp. 2 box A b 31c SSPHNHS (6) HAIYPRH (6) QISFMAN (4) TLTTPIL (2) HWGMWSY ISIPRTM TPAHNDY FHMGQPF APTPVKL HMALNxV MHSLSYR WHWWPxL GETRAPL VQASNSN GNSLRWD SAPQILL DTDPPGL Exp. 3 box B b 29c IQSPHFF (8) LPLTPLP (7) NQDVPLF (2) WPKLASH (2) SPAHAAK ASMSVAI AWLPWAK MPNRTAN NLPAYTS CSSVETH GTPTLxS VMPWVHK YAPRLRS APPTRNQ Exp. 4 box B b 20c HWGMWSY (5) HSWLWWP (5) LAMPQYE (2) KLWVIPQ SHWFWSW HAIYPRH SRPHTSD HYWWWPR SSSSHPT PGAQLTK STTLRYF
24 Table II A Putative Interactors Motifs of homology LPLTPLP Accession number Grg5 140 LPLTPLP 146 Q06195 Grg1/TL1 141 VPLTPHP 147 Q VPFPPMP 465 TLE2 160 VPLTPRP 166 NP p53 76 GPVAPAP 82 P APATPWP 89 hnrnp K 307 LPLPPPP 313 Q60577 MeCP2 380 MPLLPSP 386 NP prb 780 RPIHPIP 774 A33718 P PPLSPFP 962 Q64701 P PPLSPYP 1034 Q64700 DP2 19 NPYTPAP 25 Q64163 RelB 75 GPAAPPP 81 Q04863 PR 415 FPLAPAP 421 Q00175 AR 280 LPACPTP 274 P19091 HAIRLESS 179 WPLAPNP 185 Q61645 CDP 913 QPTTPLP 919 P53564 RBP-Jκ 375 APVTPVP 381 P31266 ERF 243 EPLSPFP 249 P70459 IRF2 315 APVTPTP 321 P23906 MNT 511 LPLYPQP 517 O08789 MCMT 1327 LPLFPEP 1333 P13864 RFX1 80 QPATPAP 86 P48377 Sox NPLSPPP 294 P43680 TTP 69 APLAPRP 75 P22893 Six5 535 LPVGPSP 541 P70178 FREAC 3 65 GPYTPQP 72 CAA11239 TFE3 312 LPVPPNP 318 Q64092 SREBP-1 93 APLSPPP 99 Q9WTN3 TGIF 154 RPVSPKP 160 AAH05724 CTCF 648 QPVAPAP 654 NP AF SPLSPMP 1109 AAB82427 HNF-3G 26 SPVNPVP 32 CAA52892 C-EBPβ 12 LPLPPPP 18 P17676 TAF GPYTPQP 1646 P21675 HES1 187 PPLVPIP 193 P35428 MATH1 29 PPLTPQP 35 P48985 MATH2 258 GPLSPPP 265 P48986 Consensus XPLTPXP VS AA YP IL TH GY F G N V C
25 Table II B Putative Interactors Motifs of homology HWGMWSY HSWLWWP Accession number ER α 200 HYGVWSC 206 P19785 ER β 160 HYGVWSC 166 O08537 TIAR 309 QWGQWSQ 315 P70318 ETS1 79 DWVMWAV 85 P27577 T2A 123 QWNMWIP 129 Q16514 prb 67 RAWLTWE 73 P13405 TOP1 203 QKWKWWE 209 BAA00950 ERGB 139 RQWLEWA 145 CAA42055
26
Analysing protein protein interactions using a GST-fusion protein to pull down the interacting target from the cell lysate Hong Wang and Xin Zeng
 Analysing protein protein interactions using a GST-fusion protein to pull down the interacting target from the cell lysate Hong Wang and Xin Zeng Department of Molecular Genetics, Biochemistry and Microbiology,
Analysing protein protein interactions using a GST-fusion protein to pull down the interacting target from the cell lysate Hong Wang and Xin Zeng Department of Molecular Genetics, Biochemistry and Microbiology,
Chapter 14 Regulation of Transcription
 Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
Complex Transcription Machinery
 Complex Transcription Machinery Subunits of the Basal Txn Apparatus Stepwise Assembly of the Pre-initiation Complex Interplay of Activators, Co-regulators and RNA Polymerase at the Promoter Divide and
Complex Transcription Machinery Subunits of the Basal Txn Apparatus Stepwise Assembly of the Pre-initiation Complex Interplay of Activators, Co-regulators and RNA Polymerase at the Promoter Divide and
Eukaryotic Transcription
 Eukaryotic Transcription I. Differences between eukaryotic versus prokaryotic transcription. II. (core vs holoenzyme): RNA polymerase II - Promotor elements. - General Pol II transcription factors (GTF).
Eukaryotic Transcription I. Differences between eukaryotic versus prokaryotic transcription. II. (core vs holoenzyme): RNA polymerase II - Promotor elements. - General Pol II transcription factors (GTF).
Differential Gene Expression
 Biology 4361 Developmental Biology Differential Gene Expression September 28, 2006 Chromatin Structure ~140 bp ~60 bp Transcriptional Regulation: 1. Packing prevents access CH 3 2. Acetylation ( C O )
Biology 4361 Developmental Biology Differential Gene Expression September 28, 2006 Chromatin Structure ~140 bp ~60 bp Transcriptional Regulation: 1. Packing prevents access CH 3 2. Acetylation ( C O )
Molecular Biology (BIOL 4320) Exam #1 March 12, 2002
 Exam #1 March 12, 2002") Molecular Biology (BIOL 4320) Exam #1 March 12, 2002 Name KEY SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses after the question number.
Molecular Biology (BIOL 4320) Exam #1 March 12, 2002 Name KEY SS# This exam is worth a total of 100 points. The number of points each question is worth is shown in parentheses after the question number.
Enhancers mutations that make the original mutant phenotype more extreme. Suppressors mutations that make the original mutant phenotype less extreme
 Interactomics and Proteomics 1. Interactomics The field of interactomics is concerned with interactions between genes or proteins. They can be genetic interactions, in which two genes are involved in the
Interactomics and Proteomics 1. Interactomics The field of interactomics is concerned with interactions between genes or proteins. They can be genetic interactions, in which two genes are involved in the
Technical tips Session 5
 Technical tips Session 5 Chromatine Immunoprecipitation (ChIP): This is a powerful in vivo method to quantitate interaction of proteins associated with specific regions of the genome. It involves the immunoprecipitation
Technical tips Session 5 Chromatine Immunoprecipitation (ChIP): This is a powerful in vivo method to quantitate interaction of proteins associated with specific regions of the genome. It involves the immunoprecipitation
Enhancers. Activators and repressors of transcription
 Enhancers Can be >50 kb away from the gene they regulate. Can be upstream from a promoter, downstream from a promoter, within an intron, or even downstream of the final exon of a gene. Are often cell type
Enhancers Can be >50 kb away from the gene they regulate. Can be upstream from a promoter, downstream from a promoter, within an intron, or even downstream of the final exon of a gene. Are often cell type
DNA Binding Domains: Structural Motifs. Effector Domain. Zinc Fingers. Zinc Fingers, continued. Zif268
 DNA Binding Domains: Structural Motifs Studies of known transcription factors have found several motifs of protein design to allow sequence-specific binding of DNA. We will cover only three of these motifs:
DNA Binding Domains: Structural Motifs Studies of known transcription factors have found several motifs of protein design to allow sequence-specific binding of DNA. We will cover only three of these motifs:
Motivation From Protein to Gene
 MOLECULAR BIOLOGY 2003-4 Topic B Recombinant DNA -principles and tools Construct a library - what for, how Major techniques +principles Bioinformatics - in brief Chapter 7 (MCB) 1 Motivation From Protein
MOLECULAR BIOLOGY 2003-4 Topic B Recombinant DNA -principles and tools Construct a library - what for, how Major techniques +principles Bioinformatics - in brief Chapter 7 (MCB) 1 Motivation From Protein
Chapter 25: Regulating Eukaryotic Transcription The Ligand Responsive Activators
 Chapter 25: Regulating Eukaryotic Transcription The Ligand Responsive Activators At least 5 potential gene expression control points Superfamily of Gene Regulators Activation of gene structure Initiation
Chapter 25: Regulating Eukaryotic Transcription The Ligand Responsive Activators At least 5 potential gene expression control points Superfamily of Gene Regulators Activation of gene structure Initiation
Chapter 6 - Molecular Genetic Techniques
 Chapter 6 - Molecular Genetic Techniques Two objects of molecular & genetic technologies For analysis For generation Molecular genetic technologies! For analysis DNA gel electrophoresis Southern blotting
Chapter 6 - Molecular Genetic Techniques Two objects of molecular & genetic technologies For analysis For generation Molecular genetic technologies! For analysis DNA gel electrophoresis Southern blotting
Differential Gene Expression
 Biology 4361 Developmental Biology Differential Gene Expression June 19, 2008 Differential Gene Expression Overview Chromatin structure Gene anatomy RNA processing and protein production Initiating transcription:
Biology 4361 Developmental Biology Differential Gene Expression June 19, 2008 Differential Gene Expression Overview Chromatin structure Gene anatomy RNA processing and protein production Initiating transcription:
Lecture 7: Affinity Chromatography-II
 Lecture 7: Affinity Chromatography-II We have studied basics of affinity purification during last lecture. The current lecture is continuation of last lecture and we will cover following: 1. Few specific
Lecture 7: Affinity Chromatography-II We have studied basics of affinity purification during last lecture. The current lecture is continuation of last lecture and we will cover following: 1. Few specific
Chapter 20 Recombinant DNA Technology. Copyright 2009 Pearson Education, Inc.
 Chapter 20 Recombinant DNA Technology Copyright 2009 Pearson Education, Inc. 20.1 Recombinant DNA Technology Began with Two Key Tools: Restriction Enzymes and DNA Cloning Vectors Recombinant DNA refers
Chapter 20 Recombinant DNA Technology Copyright 2009 Pearson Education, Inc. 20.1 Recombinant DNA Technology Began with Two Key Tools: Restriction Enzymes and DNA Cloning Vectors Recombinant DNA refers
The role of erna in chromatin looping
 The role of erna in chromatin looping Introduction Enhancers are cis-acting DNA regulatory elements that activate transcription of target genes 1,2. It is estimated that 400 000 to >1 million alleged enhancers
The role of erna in chromatin looping Introduction Enhancers are cis-acting DNA regulatory elements that activate transcription of target genes 1,2. It is estimated that 400 000 to >1 million alleged enhancers
AP Biology Gene Expression/Biotechnology REVIEW
 AP Biology Gene Expression/Biotechnology REVIEW Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Gene expression can be a. regulated before transcription.
AP Biology Gene Expression/Biotechnology REVIEW Multiple Choice Identify the choice that best completes the statement or answers the question. 1. Gene expression can be a. regulated before transcription.
Design. Construction. Characterization
 Design Construction Characterization DNA mrna (messenger) A C C transcription translation C A C protein His A T G C T A C G Plasmids replicon copy number incompatibility selection marker origin of replication
Design Construction Characterization DNA mrna (messenger) A C C transcription translation C A C protein His A T G C T A C G Plasmids replicon copy number incompatibility selection marker origin of replication
CONTENTS. LIST OF FIGURES... i. LIST OF TABLES... iii. SUMMARY OF THE WORK DONE... vi
 CONTENTS SECTION ~ LIST OF FIGURES... i LIST OF TABLES... iii LIST OF ABBREVIATIONS USED... ~... iv SUMMARY OF THE WORK DONE... vi 1. INTROD UCTION... :.................................... 1 1.1. AN OVERVIEW
CONTENTS SECTION ~ LIST OF FIGURES... i LIST OF TABLES... iii LIST OF ABBREVIATIONS USED... ~... iv SUMMARY OF THE WORK DONE... vi 1. INTROD UCTION... :.................................... 1 1.1. AN OVERVIEW
Supplemental Information
 Supplemental Information ATP-dependent unwinding of U4/U6 snrnas by the Brr2 helicase requires the C-terminus of Prp8 Corina Maeder 1,3, Alan K. Kutach 1,2,3, and Christine Guthrie 1 1 Department of Biochemistry
Supplemental Information ATP-dependent unwinding of U4/U6 snrnas by the Brr2 helicase requires the C-terminus of Prp8 Corina Maeder 1,3, Alan K. Kutach 1,2,3, and Christine Guthrie 1 1 Department of Biochemistry
Differential Gene Expression
 Developmental Biology Biology 4361 Differential Gene Expression October 13, 2005 core transcription initiation site 5 promoter 3 TATAT +1 upstream downstream Basal transcription factors (eukaryotes) TFIID
Developmental Biology Biology 4361 Differential Gene Expression October 13, 2005 core transcription initiation site 5 promoter 3 TATAT +1 upstream downstream Basal transcription factors (eukaryotes) TFIID
CONSTRUCTION OF GENOMIC LIBRARY
 MODULE 4-LECTURE 4 CONSTRUCTION OF GENOMIC LIBRARY 4-4.1. Introduction A genomic library is an organism specific collection of DNA covering the entire genome of an organism. It contains all DNA sequences
MODULE 4-LECTURE 4 CONSTRUCTION OF GENOMIC LIBRARY 4-4.1. Introduction A genomic library is an organism specific collection of DNA covering the entire genome of an organism. It contains all DNA sequences
Supplemental Data. Lee et al. Plant Cell. (2010) /tpc Supplemental Figure 1. Protein and Gene Structures of DWA1 and DWA2.
 /tpc Supplemental Figure 1. Protein and Gene Structures of DWA1 and DWA2.") Supplemental Figure 1. Protein and Gene Structures of DWA1 and DWA2. (A) Protein structures of DWA1 and DWA2. WD40 region was determined based on the NCBI conserved domain databases (B, C) Schematic representation
Supplemental Figure 1. Protein and Gene Structures of DWA1 and DWA2. (A) Protein structures of DWA1 and DWA2. WD40 region was determined based on the NCBI conserved domain databases (B, C) Schematic representation
B. Incorrect! Ligation is also a necessary step for cloning.
 Genetics - Problem Drill 15: The Techniques in Molecular Genetics No. 1 of 10 1. Which of the following is not part of the normal process of cloning recombinant DNA in bacteria? (A) Restriction endonuclease
Genetics - Problem Drill 15: The Techniques in Molecular Genetics No. 1 of 10 1. Which of the following is not part of the normal process of cloning recombinant DNA in bacteria? (A) Restriction endonuclease
IMMUNOPRECIPITATION (IP)
") 1 IMMUNOPRECIPITATION (IP) Overview and Technical Tips 2 CONTENTS 3 7 8 9 12 13 17 18 19 20 Introduction Factors Influencing IP General Protocol Modifications Of IP Protocols Troubleshooting Contact Us
1 IMMUNOPRECIPITATION (IP) Overview and Technical Tips 2 CONTENTS 3 7 8 9 12 13 17 18 19 20 Introduction Factors Influencing IP General Protocol Modifications Of IP Protocols Troubleshooting Contact Us
Molecular Genetics Techniques. BIT 220 Chapter 20
 Molecular Genetics Techniques BIT 220 Chapter 20 What is Cloning? Recombinant DNA technologies 1. Producing Recombinant DNA molecule Incorporate gene of interest into plasmid (cloning vector) 2. Recombinant
Molecular Genetics Techniques BIT 220 Chapter 20 What is Cloning? Recombinant DNA technologies 1. Producing Recombinant DNA molecule Incorporate gene of interest into plasmid (cloning vector) 2. Recombinant
EPIGENTEK. EpiQuik Tissue Chromatin Immunoprecipitation Kit. Base Catalog # P-2003 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE
 EpiQuik Tissue Chromatin Immunoprecipitation Kit Base Catalog # P-2003 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Tissue Chromatin Immunoprecipitation Kit is suitable for combining the specificity
EpiQuik Tissue Chromatin Immunoprecipitation Kit Base Catalog # P-2003 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Tissue Chromatin Immunoprecipitation Kit is suitable for combining the specificity
Transcriptional Regulation in Eukaryotes
 Transcriptional Regulation in Eukaryotes Concepts, Strategies, and Techniques Michael Carey Stephen T. Smale COLD SPRING HARBOR LABORATORY PRESS NEW YORK 2000 Cold Spring Harbor Laboratory Press, 0-87969-537-4
Transcriptional Regulation in Eukaryotes Concepts, Strategies, and Techniques Michael Carey Stephen T. Smale COLD SPRING HARBOR LABORATORY PRESS NEW YORK 2000 Cold Spring Harbor Laboratory Press, 0-87969-537-4
Yeast 2-Hybrid Kayla Nygaard
 Yeast 2-Hybrid 2.26.18 Kayla Nygaard Y2H - What is it? A method to screen for protein-protein interactions in yeast Capitalizes on GAL4 system in yeast. GAL4 has 2 domains DNA-Binding Domain (DB) Transcriptional
Yeast 2-Hybrid 2.26.18 Kayla Nygaard Y2H - What is it? A method to screen for protein-protein interactions in yeast Capitalizes on GAL4 system in yeast. GAL4 has 2 domains DNA-Binding Domain (DB) Transcriptional
fibrils, however, oligomeric structures and amorphous protein aggregates were
 Supplementary Figure 1: Effect of equimolar EGCG on αs and Aβ aggregate formation. A D B E αs C F Figure S1: (A-C) Analysis of EGCG treated αs (1 µm) aggregation reactions by EM. A 1:1 molar ratio of EGCG
Supplementary Figure 1: Effect of equimolar EGCG on αs and Aβ aggregate formation. A D B E αs C F Figure S1: (A-C) Analysis of EGCG treated αs (1 µm) aggregation reactions by EM. A 1:1 molar ratio of EGCG
Lecture Four. Molecular Approaches I: Nucleic Acids
 Lecture Four. Molecular Approaches I: Nucleic Acids I. Recombinant DNA and Gene Cloning Recombinant DNA is DNA that has been created artificially. DNA from two or more sources is incorporated into a single
Lecture Four. Molecular Approaches I: Nucleic Acids I. Recombinant DNA and Gene Cloning Recombinant DNA is DNA that has been created artificially. DNA from two or more sources is incorporated into a single
Applicazioni biotecnologiche
 Applicazioni biotecnologiche Analisi forense Sintesi di proteine ricombinanti Restriction Fragment Length Polymorphism (RFLP) Polymorphism (more fully genetic polymorphism) refers to the simultaneous occurrence
Applicazioni biotecnologiche Analisi forense Sintesi di proteine ricombinanti Restriction Fragment Length Polymorphism (RFLP) Polymorphism (more fully genetic polymorphism) refers to the simultaneous occurrence
Methods and Materials
 SUMMARY OF PH.D. THESIS Molecular biological and genetic study of the sumoylation of Drosophila melanogaster p53 Pardi Norbert Supervisor: Dr. Boros Imre Doctoral School of Biology University of Szeged
SUMMARY OF PH.D. THESIS Molecular biological and genetic study of the sumoylation of Drosophila melanogaster p53 Pardi Norbert Supervisor: Dr. Boros Imre Doctoral School of Biology University of Szeged
Recitation CHAPTER 9 DNA Technologies
 Recitation CHAPTER 9 DNA Technologies DNA Cloning: General Scheme A cloning vector and eukaryotic chromosomes are separately cleaved with the same restriction endonuclease. (A single chromosome is shown
Recitation CHAPTER 9 DNA Technologies DNA Cloning: General Scheme A cloning vector and eukaryotic chromosomes are separately cleaved with the same restriction endonuclease. (A single chromosome is shown
The Two-Hybrid System
 Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine The Two-Hybrid System Carolina Vollert & Peter Uetz Institut für Genetik Forschungszentrum Karlsruhe PO Box 3640 D-76021 Karlsruhe
Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine The Two-Hybrid System Carolina Vollert & Peter Uetz Institut für Genetik Forschungszentrum Karlsruhe PO Box 3640 D-76021 Karlsruhe
CELL BIOLOGY - CLUTCH CH. 7 - GENE EXPRESSION.
 !! www.clutchprep.com CONCEPT: CONTROL OF GENE EXPRESSION BASICS Gene expression is the process through which cells selectively to express some genes and not others Every cell in an organism is a clone
!! www.clutchprep.com CONCEPT: CONTROL OF GENE EXPRESSION BASICS Gene expression is the process through which cells selectively to express some genes and not others Every cell in an organism is a clone
BS1940 Course Topics Fall 2001 Drs. Hatfull and Arndt
 BS1940 Course Topics Fall 2001 Drs. Hatfull and Arndt Introduction to molecular biology Combining genetics, biochemistry, structural chemistry Information flow in biological systems: The Central Dogma
BS1940 Course Topics Fall 2001 Drs. Hatfull and Arndt Introduction to molecular biology Combining genetics, biochemistry, structural chemistry Information flow in biological systems: The Central Dogma
7 Gene Isolation and Analysis of Multiple
 Genetic Techniques for Biological Research Corinne A. Michels Copyright q 2002 John Wiley & Sons, Ltd ISBNs: 0-471-89921-6 (Hardback); 0-470-84662-3 (Electronic) 7 Gene Isolation and Analysis of Multiple
Genetic Techniques for Biological Research Corinne A. Michels Copyright q 2002 John Wiley & Sons, Ltd ISBNs: 0-471-89921-6 (Hardback); 0-470-84662-3 (Electronic) 7 Gene Isolation and Analysis of Multiple
This Technical Note describes these characterization studies in more detail.
 Technical Note Characterization of SOMAmer Reagents Binding Specificity in the SOMAscan 1.3k Assay Summary Slow Off-Rate Modified Aptamer (SOMAmer) reagents are identified via the SELEX process against
Technical Note Characterization of SOMAmer Reagents Binding Specificity in the SOMAscan 1.3k Assay Summary Slow Off-Rate Modified Aptamer (SOMAmer) reagents are identified via the SELEX process against
Plant Molecular and Cellular Biology Lecture 9: Nuclear Genome Organization: Chromosome Structure, Chromatin, DNA Packaging, Mitosis Gary Peter
 Plant Molecular and Cellular Biology Lecture 9: Nuclear Genome Organization: Chromosome Structure, Chromatin, DNA Packaging, Mitosis Gary Peter 9/16/2008 1 Learning Objectives 1. List and explain how DNA
Plant Molecular and Cellular Biology Lecture 9: Nuclear Genome Organization: Chromosome Structure, Chromatin, DNA Packaging, Mitosis Gary Peter 9/16/2008 1 Learning Objectives 1. List and explain how DNA
Shaping Epigenetics Discovery
 Shaping Epigenetics Discovery Kits, Assays, Controls, Inhibitors and Antibodies for DNA Methylation The life science business of Merck KGaA, Darmstadt, Germany operates as MilliporeSigma in the U.S. and
Shaping Epigenetics Discovery Kits, Assays, Controls, Inhibitors and Antibodies for DNA Methylation The life science business of Merck KGaA, Darmstadt, Germany operates as MilliporeSigma in the U.S. and
Chromatographic Separation of the three forms of RNA Polymerase II.
 Chromatographic Separation of the three forms of RNA Polymerase II. α-amanitin α-amanitin bound to Pol II Function of the three enzymes. Yeast Pol II. RNA Polymerase Subunit Structures 10-7 Subunit structure.
Chromatographic Separation of the three forms of RNA Polymerase II. α-amanitin α-amanitin bound to Pol II Function of the three enzymes. Yeast Pol II. RNA Polymerase Subunit Structures 10-7 Subunit structure.
However, only a fraction of these genes are transcribed in an individual cell at any given time.
 All cells in an organism contain the same set of genes. However, only a fraction of these genes are transcribed in an individual cell at any given time. It is the pattern of gene expression that determines
All cells in an organism contain the same set of genes. However, only a fraction of these genes are transcribed in an individual cell at any given time. It is the pattern of gene expression that determines
Eukaryotic & Prokaryotic Transcription. RNA polymerases
 Eukaryotic & Prokaryotic Transcription RNA polymerases RNA Polymerases A. E. coli RNA polymerase 1. core enzyme = ββ'(α)2 has catalytic activity but cannot recognize start site of transcription ~500,000
Eukaryotic & Prokaryotic Transcription RNA polymerases RNA Polymerases A. E. coli RNA polymerase 1. core enzyme = ββ'(α)2 has catalytic activity but cannot recognize start site of transcription ~500,000
GENETICS EXAM 3 FALL a) is a technique that allows you to separate nucleic acids (DNA or RNA) by size.
 is a technique that allows you to separate nucleic acids (DNA or RNA) by size.") Student Name: All questions are worth 5 pts. each. GENETICS EXAM 3 FALL 2004 1. a) is a technique that allows you to separate nucleic acids (DNA or RNA) by size. b) Name one of the materials (of the two
Student Name: All questions are worth 5 pts. each. GENETICS EXAM 3 FALL 2004 1. a) is a technique that allows you to separate nucleic acids (DNA or RNA) by size. b) Name one of the materials (of the two
RNA does not adopt the classic B-DNA helix conformation when it forms a self-complementary double helix
 Reason: RNA has ribose sugar ring, with a hydroxyl group (OH) If RNA in B-from conformation there would be unfavorable steric contact between the hydroxyl group, base, and phosphate backbone. RNA structure
Reason: RNA has ribose sugar ring, with a hydroxyl group (OH) If RNA in B-from conformation there would be unfavorable steric contact between the hydroxyl group, base, and phosphate backbone. RNA structure
2014 Pearson Education, Inc. CH 8: Recombinant DNA Technology
 CH 8: Recombinant DNA Technology Biotechnology the use of microorganisms to make practical products Recombinant DNA = DNA from 2 different sources What is Recombinant DNA Technology? modifying genomes
CH 8: Recombinant DNA Technology Biotechnology the use of microorganisms to make practical products Recombinant DNA = DNA from 2 different sources What is Recombinant DNA Technology? modifying genomes
Supplementary Note 1. Enzymatic properties of the purified Syn BVR
 Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Supplementary Note 1. Enzymatic properties of the purified Syn BVR The expression vector pet15b-syn bvr allowed us to routinely prepare 15 mg of electrophoretically homogenous Syn BVR from 2.5 L of TB-medium
Bi 8 Lecture 7. Ellen Rothenberg 26 January Reading: Ch. 3, pp ; panel 3-1
 Bi 8 Lecture 7 PROTEIN STRUCTURE, Functional analysis, and evolution Ellen Rothenberg 26 January 2016 Reading: Ch. 3, pp. 109-134; panel 3-1 (end with free amine) aromatic, hydrophobic small, hydrophilic
Bi 8 Lecture 7 PROTEIN STRUCTURE, Functional analysis, and evolution Ellen Rothenberg 26 January 2016 Reading: Ch. 3, pp. 109-134; panel 3-1 (end with free amine) aromatic, hydrophobic small, hydrophilic
Bi 8 Lecture 10. Ellen Rothenberg 4 February 2016
 Bi 8 Lecture 10 Bacterial regulation, II Ellen Rothenberg 4 February 2016 Not all bacterial promoters use the same σ factors, and this provides added regulation capability Most sigma factors are related
Bi 8 Lecture 10 Bacterial regulation, II Ellen Rothenberg 4 February 2016 Not all bacterial promoters use the same σ factors, and this provides added regulation capability Most sigma factors are related
Structure/function relationship in DNA-binding proteins
 PHRM 836 September 22, 2015 Structure/function relationship in DNA-binding proteins Devlin Chapter 8.8-9 u General description of transcription factors (TFs) u Sequence-specific interactions between DNA
PHRM 836 September 22, 2015 Structure/function relationship in DNA-binding proteins Devlin Chapter 8.8-9 u General description of transcription factors (TFs) u Sequence-specific interactions between DNA
Lecture 22 Eukaryotic Genes and Genomes III
 Lecture 22 Eukaryotic Genes and Genomes III In the last three lectures we have thought a lot about analyzing a regulatory system in S. cerevisiae, namely Gal regulation that involved a hand full of genes.
Lecture 22 Eukaryotic Genes and Genomes III In the last three lectures we have thought a lot about analyzing a regulatory system in S. cerevisiae, namely Gal regulation that involved a hand full of genes.
Newsletter Issue 7 One-STrEP Analysis of Protein:Protein-Interactions
 www.iba-biotagnology.com Newsletter Issue 7 One-STrEP Analysis of Protein:Protein-Interactions Strep-tag and One-STrEP-tag PPI Analysis with the co-precipitation/ mass spectrometry approach 3 Background
www.iba-biotagnology.com Newsletter Issue 7 One-STrEP Analysis of Protein:Protein-Interactions Strep-tag and One-STrEP-tag PPI Analysis with the co-precipitation/ mass spectrometry approach 3 Background
THE ANALYSIS OF CHROMATIN CONDENSATION STATE AND TRANSCRIPTIONAL ACTIVITY USING DNA MICROARRAYS 1. INTRODUCTION
 JOURNAL OF MEDICAL INFORMATICS & TECHNOLOGIES Vol.6/2003, ISSN 1642-6037 Piotr WIDŁAK *, Krzysztof FUJAREWICZ ** DNA microarrays, chromatin, transcription THE ANALYSIS OF CHROMATIN CONDENSATION STATE AND
JOURNAL OF MEDICAL INFORMATICS & TECHNOLOGIES Vol.6/2003, ISSN 1642-6037 Piotr WIDŁAK *, Krzysztof FUJAREWICZ ** DNA microarrays, chromatin, transcription THE ANALYSIS OF CHROMATIN CONDENSATION STATE AND
Schematic representation of the endogenous PALB2 locus and gene-disruption constructs
 Supplementary Figures Supplementary Figure 1. Generation of PALB2 -/- and BRCA2 -/- /PALB2 -/- DT40 cells. (A) Schematic representation of the endogenous PALB2 locus and gene-disruption constructs carrying
Supplementary Figures Supplementary Figure 1. Generation of PALB2 -/- and BRCA2 -/- /PALB2 -/- DT40 cells. (A) Schematic representation of the endogenous PALB2 locus and gene-disruption constructs carrying
Enzyme that uses RNA as a template to synthesize a complementary DNA
 Biology 105: Introduction to Genetics PRACTICE FINAL EXAM 2006 Part I: Definitions Homology: Comparison of two or more protein or DNA sequence to ascertain similarities in sequences. If two genes have
Biology 105: Introduction to Genetics PRACTICE FINAL EXAM 2006 Part I: Definitions Homology: Comparison of two or more protein or DNA sequence to ascertain similarities in sequences. If two genes have
Biochemistry 302, February 11, 2004 Exam 1 (100 points) 1. What form of DNA is shown on this Nature Genetics cover? Z-DNA or left-handed DNA
 1. What form of DNA is shown on this Nature Genetics cover? Z-DNA or left-handed DNA") 1 Biochemistry 302, February 11, 2004 Exam 1 (100 points) Name I. Structural recognition (very short answer, 2 points each) 1. What form of DNA is shown on this Nature Genetics cover? Z-DNA or left-handed
1 Biochemistry 302, February 11, 2004 Exam 1 (100 points) Name I. Structural recognition (very short answer, 2 points each) 1. What form of DNA is shown on this Nature Genetics cover? Z-DNA or left-handed
Introduction to Affinity Chromatography (AC) By: Sunsanee Yoojun Bang Trading 1992 Co., Ltd.
 By: Sunsanee Yoojun Bang Trading 1992 Co., Ltd.") Introduction to Affinity Chromatography (AC) By: Sunsanee Yoojun Bang Trading 1992 Co., Ltd. Content What is affinity chromatography (AC)? What is AC used for? Principles Experimental set up Summary What
Introduction to Affinity Chromatography (AC) By: Sunsanee Yoojun Bang Trading 1992 Co., Ltd. Content What is affinity chromatography (AC)? What is AC used for? Principles Experimental set up Summary What
30 Gene expression: Transcription
 30 Gene expression: Transcription Gene structure. o Exons coding region of DNA. o Introns non-coding region of DNA. o Introns are interspersed between exons of a single gene. o Promoter region helps enzymes
30 Gene expression: Transcription Gene structure. o Exons coding region of DNA. o Introns non-coding region of DNA. o Introns are interspersed between exons of a single gene. o Promoter region helps enzymes
Site directed mutagenesis, Insertional and Deletion Mutagenesis. Mitesh Shrestha
 Site directed mutagenesis, Insertional and Deletion Mutagenesis Mitesh Shrestha Mutagenesis Mutagenesis (the creation or formation of a mutation) can be used as a powerful genetic tool. By inducing mutations
Site directed mutagenesis, Insertional and Deletion Mutagenesis Mitesh Shrestha Mutagenesis Mutagenesis (the creation or formation of a mutation) can be used as a powerful genetic tool. By inducing mutations
Biology 201 (Genetics) Exam #3 120 points 20 November Read the question carefully before answering. Think before you write.
 Exam #3 120 points 20 November Read the question carefully before answering. Think before you write.") Name KEY Section Biology 201 (Genetics) Exam #3 120 points 20 November 2006 Read the question carefully before answering. Think before you write. You will have up to 50 minutes to take this exam. After
Name KEY Section Biology 201 (Genetics) Exam #3 120 points 20 November 2006 Read the question carefully before answering. Think before you write. You will have up to 50 minutes to take this exam. After
Practice Exam A. Briefly describe how IL-25 treatment might be able to help this responder subgroup of liver cancer patients.
 Practice Exam 2007 1. A special JAK-STAT signaling system (JAK5-STAT5) was recently identified in which a gene called TS5 becomes selectively transcribed and expressed in the liver upon induction by a
Practice Exam 2007 1. A special JAK-STAT signaling system (JAK5-STAT5) was recently identified in which a gene called TS5 becomes selectively transcribed and expressed in the liver upon induction by a
CH 8: Recombinant DNA Technology
 CH 8: Recombinant DNA Technology Biotechnology the use of microorganisms to make practical products Recombinant DNA = DNA from 2 different sources What is Recombinant DNA Technology? modifying genomes
CH 8: Recombinant DNA Technology Biotechnology the use of microorganisms to make practical products Recombinant DNA = DNA from 2 different sources What is Recombinant DNA Technology? modifying genomes
Understanding the Cellular Mechanism of the Excess Microsporocytes I (EMSI) Gene. Andrew ElBardissi, The Pennsylvania State University
 Gene. Andrew ElBardissi, The Pennsylvania State University") Understanding the Cellular Mechanism of the Excess Microsporocytes I (EMSI) Gene Andrew ElBardissi, The Pennsylvania State University Abstract: Hong Ma, The Pennsylvania State University The Excess Microsporocytes
Understanding the Cellular Mechanism of the Excess Microsporocytes I (EMSI) Gene Andrew ElBardissi, The Pennsylvania State University Abstract: Hong Ma, The Pennsylvania State University The Excess Microsporocytes
Genome annotation & EST
 Genome annotation & EST What is genome annotation? The process of taking the raw DNA sequence produced by the genome sequence projects and adding the layers of analysis and interpretation necessary
Genome annotation & EST What is genome annotation? The process of taking the raw DNA sequence produced by the genome sequence projects and adding the layers of analysis and interpretation necessary
Recombinant DNA Technology. The Role of Recombinant DNA Technology in Biotechnology. yeast. Biotechnology. Recombinant DNA technology.
 PowerPoint Lecture Presentations prepared by Mindy Miller-Kittrell, North Carolina State University C H A P T E R 8 Recombinant DNA Technology The Role of Recombinant DNA Technology in Biotechnology Biotechnology?
PowerPoint Lecture Presentations prepared by Mindy Miller-Kittrell, North Carolina State University C H A P T E R 8 Recombinant DNA Technology The Role of Recombinant DNA Technology in Biotechnology Biotechnology?
Gα 13 Activation Assay Kit
 A helping hand for your research Product Manual Configuration-specific Monoclonal Antibody Based Gα 13 Activation Assay Kit Catalog Number: 80401 20 assays NewEast Biosciences 1 Table of Content Product
A helping hand for your research Product Manual Configuration-specific Monoclonal Antibody Based Gα 13 Activation Assay Kit Catalog Number: 80401 20 assays NewEast Biosciences 1 Table of Content Product
Genetics and Genomics in Medicine Chapter 3. Questions & Answers
 Genetics and Genomics in Medicine Chapter 3 Multiple Choice Questions Questions & Answers Question 3.1 Which of the following statements, if any, is false? a) Amplifying DNA means making many identical
Genetics and Genomics in Medicine Chapter 3 Multiple Choice Questions Questions & Answers Question 3.1 Which of the following statements, if any, is false? a) Amplifying DNA means making many identical
The Biotechnology Toolbox
 Chapter 15 The Biotechnology Toolbox Cutting and Pasting DNA Cutting DNA Restriction endonuclease or restriction enzymes Cellular protection mechanism for infected foreign DNA Recognition and cutting specific
Chapter 15 The Biotechnology Toolbox Cutting and Pasting DNA Cutting DNA Restriction endonuclease or restriction enzymes Cellular protection mechanism for infected foreign DNA Recognition and cutting specific
CHAPTER 9 DNA Technologies
 CHAPTER 9 DNA Technologies Recombinant DNA Artificially created DNA that combines sequences that do not occur together in the nature Basis of much of the modern molecular biology Molecular cloning of genes
CHAPTER 9 DNA Technologies Recombinant DNA Artificially created DNA that combines sequences that do not occur together in the nature Basis of much of the modern molecular biology Molecular cloning of genes
A Modified Digestion-Circularization PCR (DC-PCR) Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks
 Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks") A Modified Digestion-Circularization PCR (DC-PCR) Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks SARAH K. DICKERSON AND F. NINA PAPAVASILIOU Laboratory of Lymphocyte Biology, The
A Modified Digestion-Circularization PCR (DC-PCR) Approach to Detect Hypermutation- Associated DNA Double-Strand Breaks SARAH K. DICKERSON AND F. NINA PAPAVASILIOU Laboratory of Lymphocyte Biology, The
2. Outline the levels of DNA packing in the eukaryotic nucleus below next to the diagram provided.
 AP Biology Reading Packet 6- Molecular Genetics Part 2 Name Chapter 19: Eukaryotic Genomes 1. Define the following terms: a. Euchromatin b. Heterochromatin c. Nucleosome 2. Outline the levels of DNA packing
AP Biology Reading Packet 6- Molecular Genetics Part 2 Name Chapter 19: Eukaryotic Genomes 1. Define the following terms: a. Euchromatin b. Heterochromatin c. Nucleosome 2. Outline the levels of DNA packing
XactEdit Cas9 Nuclease with NLS User Manual
 XactEdit Cas9 Nuclease with NLS User Manual An RNA-guided recombinant endonuclease for efficient targeted DNA cleavage Catalog Numbers CE1000-50K, CE1000-50, CE1000-250, CE1001-250, CE1001-1000 Table of
XactEdit Cas9 Nuclease with NLS User Manual An RNA-guided recombinant endonuclease for efficient targeted DNA cleavage Catalog Numbers CE1000-50K, CE1000-50, CE1000-250, CE1001-250, CE1001-1000 Table of
1. Cross-linking and cell harvesting
 ChIP is a powerful tool that allows the specific matching of proteins or histone modifications to regions of the genome. Chromatin is isolated and antibodies to the antigen of interest are used to determine
ChIP is a powerful tool that allows the specific matching of proteins or histone modifications to regions of the genome. Chromatin is isolated and antibodies to the antigen of interest are used to determine
What is an Aptamer? smallest unit of repeating structure
 What is an Aptamer? apto: mer: to fit smallest unit of repeating structure Aptamers are single stranded folded oligonucleotides that bind to molecular (protein) targets with high affinity and specificity
What is an Aptamer? apto: mer: to fit smallest unit of repeating structure Aptamers are single stranded folded oligonucleotides that bind to molecular (protein) targets with high affinity and specificity
Chapter 8 DNA Recognition in Prokaryotes by Helix-Turn-Helix Motifs
 Chapter 8 DNA Recognition in Prokaryotes by Helix-Turn-Helix Motifs 1. Helix-turn-helix proteins 2. Zinc finger proteins 3. Leucine zipper proteins 4. Beta-scaffold factors 5. Others λ-repressor AND CRO
Chapter 8 DNA Recognition in Prokaryotes by Helix-Turn-Helix Motifs 1. Helix-turn-helix proteins 2. Zinc finger proteins 3. Leucine zipper proteins 4. Beta-scaffold factors 5. Others λ-repressor AND CRO
EPIGENTEK. EpiQuik Chromatin Immunoprecipitation Kit. Base Catalog # P-2002 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE
 EpiQuik Chromatin Immunoprecipitation Kit Base Catalog # P-2002 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Chromatin Immunoprecipitation Kit is suitable for combining the specificity of
EpiQuik Chromatin Immunoprecipitation Kit Base Catalog # P-2002 PLEASE READ THIS ENTIRE USER GUIDE BEFORE USE The EpiQuik Chromatin Immunoprecipitation Kit is suitable for combining the specificity of
Gene expression DNA RNA. Protein DNA. Replication. Initiation Elongation Processing Export. DNA RNA Protein. Transcription. Degradation.
 Gene expression DNA RNA Protein DNA DNA Degradation RNA Degradation Protein Replication Transcription Translation Initiation Elongation Processing Export Initiation Elongation Processing Targeting Chapter
Gene expression DNA RNA Protein DNA DNA Degradation RNA Degradation Protein Replication Transcription Translation Initiation Elongation Processing Export Initiation Elongation Processing Targeting Chapter
WesternMAX Alkaline Phosphatase Chemiluminescent Detection Kits
 WesternMAX Alkaline Phosphatase Chemiluminescent Detection Kits Code N221-KIT N220-KIT Description WesternMAX Chemiluminescent AP Kit, Anti-Mouse Includes: Alkaline Phosphatase (AP) Conjugated Anti-Mouse
WesternMAX Alkaline Phosphatase Chemiluminescent Detection Kits Code N221-KIT N220-KIT Description WesternMAX Chemiluminescent AP Kit, Anti-Mouse Includes: Alkaline Phosphatase (AP) Conjugated Anti-Mouse
Polyclonal ARHGAP25 antibody was prepared from rabbit serum after intracutaneous
 Preparation and purification of polyclonal antibodies Polyclonal ARHGAP25 antibody was prepared from rabbit serum after intracutaneous injections of glutathione S-transferase-ARHGAP25-(509-619) (GST-coiled
Preparation and purification of polyclonal antibodies Polyclonal ARHGAP25 antibody was prepared from rabbit serum after intracutaneous injections of glutathione S-transferase-ARHGAP25-(509-619) (GST-coiled
Solutions to 7.02 Quiz II 10/27/05
 Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
Solutions to 7.02 Quiz II 10/27/05 Class Average = 83 Standard Deviation = 9 Range Grade % 87-100 A 43 74-86 B 39 55-73 C 17 > 54 D 1 Question 1 (56 points) While studying deep sea bacteria, you discover
SUPPLEMENTARY INFORMATION
 R2A MASNSEKNPLL-SDEKPKSTEENKSS-KPESASGSSTSSAMP---GLNFNAFDFSNMASIL 56 R2B MASSSEKTPLIPSDEKNDTKEESKSTTKPESGSGAPPSPS-PTDPGLDFNAFDFSGMAGIL 60 R2A NDPSIREMAEQIAKDPAFNQLAEQLQRSIPNAGQEGGFPNFDPQQYVNTMQQVMHNPEFK
R2A MASNSEKNPLL-SDEKPKSTEENKSS-KPESASGSSTSSAMP---GLNFNAFDFSNMASIL 56 R2B MASSSEKTPLIPSDEKNDTKEESKSTTKPESGSGAPPSPS-PTDPGLDFNAFDFSGMAGIL 60 R2A NDPSIREMAEQIAKDPAFNQLAEQLQRSIPNAGQEGGFPNFDPQQYVNTMQQVMHNPEFK
Cdc42 Activation Assay Kit
 A helping hand for your research Product Manual Configuration-specific Monoclonal Antibody Based Cdc42 Activation Assay Kit Catalog Number: 80701 20 assays 1 Table of Content Product Description 3 Assay
A helping hand for your research Product Manual Configuration-specific Monoclonal Antibody Based Cdc42 Activation Assay Kit Catalog Number: 80701 20 assays 1 Table of Content Product Description 3 Assay
HPV E6 oncoprotein targets histone methyltransferases for modulating specific. Chih-Hung Hsu, Kai-Lin Peng, Hua-Ci Jhang, Chia-Hui Lin, Shwu-Yuan Wu,
 1 HPV E oncoprotein targets histone methyltransferases for modulating specific gene transcription 3 5 Chih-Hung Hsu, Kai-Lin Peng, Hua-Ci Jhang, Chia-Hui Lin, Shwu-Yuan Wu, Cheng-Ming Chiang, Sheng-Chung
1 HPV E oncoprotein targets histone methyltransferases for modulating specific gene transcription 3 5 Chih-Hung Hsu, Kai-Lin Peng, Hua-Ci Jhang, Chia-Hui Lin, Shwu-Yuan Wu, Cheng-Ming Chiang, Sheng-Chung
Parallel analysis of translated ORF (PLATO)
") Parallel analysis of translated ORF (PLATO) Technical Journal Club 16.04.2013 Mary-Aude Rochat, PhD student Speck group Proteomics Rationale: Increase in the DNA sequence information Understand biological
Parallel analysis of translated ORF (PLATO) Technical Journal Club 16.04.2013 Mary-Aude Rochat, PhD student Speck group Proteomics Rationale: Increase in the DNA sequence information Understand biological
Molecular Cell Biology - Problem Drill 11: Recombinant DNA
 Molecular Cell Biology - Problem Drill 11: Recombinant DNA Question No. 1 of 10 1. Which of the following statements about the sources of DNA used for molecular cloning is correct? Question #1 (A) cdna
Molecular Cell Biology - Problem Drill 11: Recombinant DNA Question No. 1 of 10 1. Which of the following statements about the sources of DNA used for molecular cloning is correct? Question #1 (A) cdna
Problem Set 8. Answer Key
 MCB 102 University of California, Berkeley August 11, 2009 Isabelle Philipp Online Document Problem Set 8 Answer Key 1. The Genetic Code (a) Are all amino acids encoded by the same number of codons? no
MCB 102 University of California, Berkeley August 11, 2009 Isabelle Philipp Online Document Problem Set 8 Answer Key 1. The Genetic Code (a) Are all amino acids encoded by the same number of codons? no
(Supplementary Methods online)
") (Supplementary Methods online) Production and purification of either LC-antisense or control molecules Recombinant phagemids and the phagemid vector were transformed into XL-1 Blue competent bacterial
(Supplementary Methods online) Production and purification of either LC-antisense or control molecules Recombinant phagemids and the phagemid vector were transformed into XL-1 Blue competent bacterial
Comparison of different methods for purification analysis of a green fluorescent Strep-tag fusion protein. Application
 Comparison of different methods for purification analysis of a green fluorescent Strep-tag fusion protein Application Petra Sebastian Meike Kuschel Stefan Schmidt Abstract This Application Note describes
Comparison of different methods for purification analysis of a green fluorescent Strep-tag fusion protein Application Petra Sebastian Meike Kuschel Stefan Schmidt Abstract This Application Note describes
Module 2 overview SPRING BREAK
 1 Module 2 overview lecture lab 1. Introduction to the module 1. Start-up protein eng. 2. Rational protein design 2. Site-directed mutagenesis 3. Fluorescence and sensors 3. DNA amplification 4. Protein
1 Module 2 overview lecture lab 1. Introduction to the module 1. Start-up protein eng. 2. Rational protein design 2. Site-directed mutagenesis 3. Fluorescence and sensors 3. DNA amplification 4. Protein
Development of Immunogens to Protect Against Turkey Cellulitis. Douglas. N. Foster and Robyn Gangl. Department of Animal Science
 Development of Immunogens to Protect Against Turkey Cellulitis Douglas. N. Foster and Robyn Gangl Department of Animal Science University of Minnesota St. Paul, MN 55108 Introduction Clostridial dermatitis
Development of Immunogens to Protect Against Turkey Cellulitis Douglas. N. Foster and Robyn Gangl Department of Animal Science University of Minnesota St. Paul, MN 55108 Introduction Clostridial dermatitis
Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question in Section B and ONE question from Section C.
 UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2013-2014 MOLECULAR BIOLOGY BIO-2B02 Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question
UNIVERSITY OF EAST ANGLIA School of Biological Sciences Main Series UG Examination 2013-2014 MOLECULAR BIOLOGY BIO-2B02 Time allowed: 2 hours Answer ALL questions in Section A, ALL PARTS of the question
Intracellular receptors specify complex patterns of gene expression that are cell and gene
 SUPPLEMENTAL RESULTS AND DISCUSSION Some HPr-1AR ARE-containing Genes Are Unresponsive to Androgen Intracellular receptors specify complex patterns of gene expression that are cell and gene specific. For
SUPPLEMENTAL RESULTS AND DISCUSSION Some HPr-1AR ARE-containing Genes Are Unresponsive to Androgen Intracellular receptors specify complex patterns of gene expression that are cell and gene specific. For
Proteomics. Manickam Sugumaran. Department of Biology University of Massachusetts Boston, MA 02125
 Proteomics Manickam Sugumaran Department of Biology University of Massachusetts Boston, MA 02125 Genomic studies produced more than 75,000 potential gene sequence targets. (The number may be even higher
Proteomics Manickam Sugumaran Department of Biology University of Massachusetts Boston, MA 02125 Genomic studies produced more than 75,000 potential gene sequence targets. (The number may be even higher
Diagnosis Sanger. Interpreting and Troubleshooting Chromatograms. Volume 1: Help! No Data! GENEWIZ Technical Support
 Diagnosis Sanger Interpreting and Troubleshooting Chromatograms GENEWIZ Technical Support DNAseq@genewiz.com Troubleshooting This troubleshooting guide is based on common issues seen from samples within
Diagnosis Sanger Interpreting and Troubleshooting Chromatograms GENEWIZ Technical Support DNAseq@genewiz.com Troubleshooting This troubleshooting guide is based on common issues seen from samples within
Use of In-Fusion Cloning for Simple and Efficient Assembly of Gene Constructs No restriction enzymes or ligation reactions necessary
 No restriction enzymes or ligation reactions necessary Background The creation of genetic circuits and artificial biological systems typically involves the use of modular genetic components biological
No restriction enzymes or ligation reactions necessary Background The creation of genetic circuits and artificial biological systems typically involves the use of modular genetic components biological
CHAPTER 4 Cloning, expression, purification and preparation of site-directed mutants of NDUFS3 and NDUFS7
 CHAPTER 4 Cloning, expression, purification and preparation of site-directed mutants of NDUFS3 and NDUFS7 subunits of human mitochondrial Complex-I Q module N DUFS2, 3, 7 and 8 form the core subunits of
CHAPTER 4 Cloning, expression, purification and preparation of site-directed mutants of NDUFS3 and NDUFS7 subunits of human mitochondrial Complex-I Q module N DUFS2, 3, 7 and 8 form the core subunits of
Supplementary information Bi-specific Aptamers mediating Tumour Cell Lysis
 Supplementary information Bi-specific Aptamers mediating Tumour Cell Lysis Figure S1 SELEX scheme of different CD16 DNA selection approaches. Nine rounds of filter SELEX were conducted in parallel, only
Supplementary information Bi-specific Aptamers mediating Tumour Cell Lysis Figure S1 SELEX scheme of different CD16 DNA selection approaches. Nine rounds of filter SELEX were conducted in parallel, only
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Supplementary figures Supplementary Figure 1: Suv39h1, but not Suv39h2, promotes HP1α sumoylation in vivo. In vivo HP1α sumoylation assay. Top: experimental scheme. Middle: we
SUPPLEMENTARY INFORMATION Supplementary figures Supplementary Figure 1: Suv39h1, but not Suv39h2, promotes HP1α sumoylation in vivo. In vivo HP1α sumoylation assay. Top: experimental scheme. Middle: we