Chapter 14 Regulation of Transcription
|
|
|
- Osborne Cameron
- 5 years ago
- Views:
Transcription

1 Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors


 site surface to recognize")

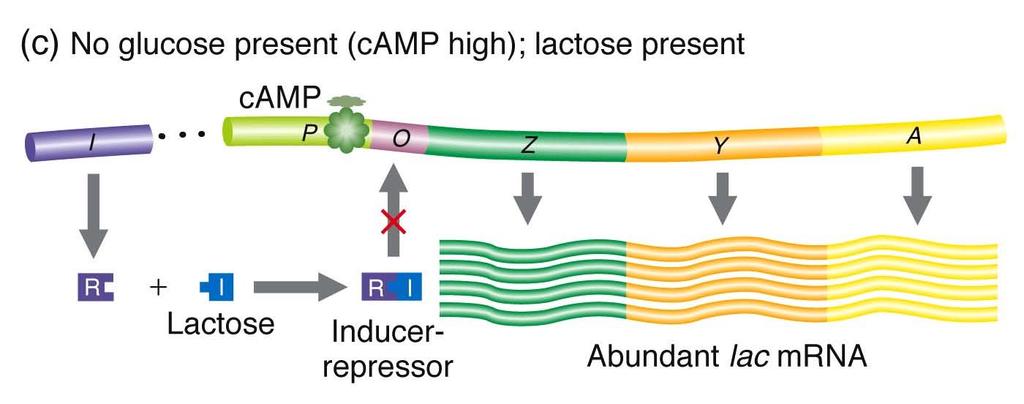
")

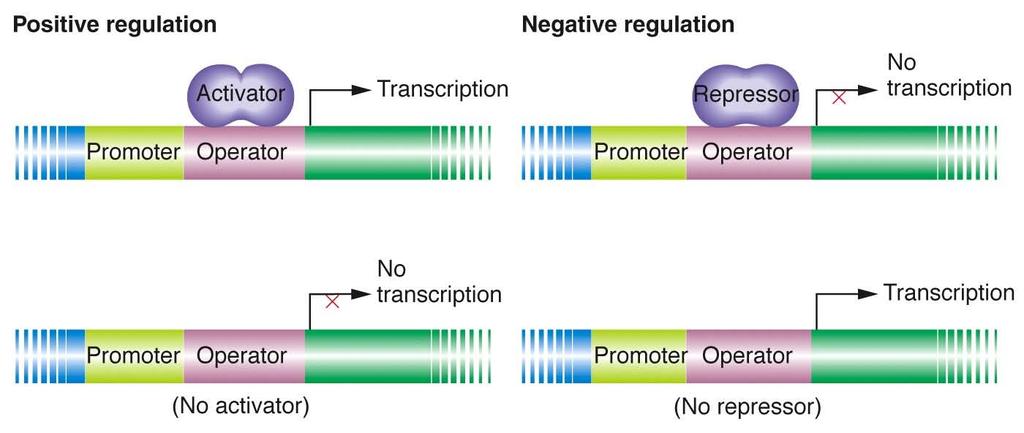
2 Overview transcriptional regulation Transcription factors Several regulatory domains sequence-specific DNA-binding surface to recognize DNA docking (target) site surface to recognize basal transcription apparatus surface to interact with proteins binding to nearby docking sites (may or may not be present) surface to act as sensor of environmental conditions (may or may not be present) Many variations and interactions
3
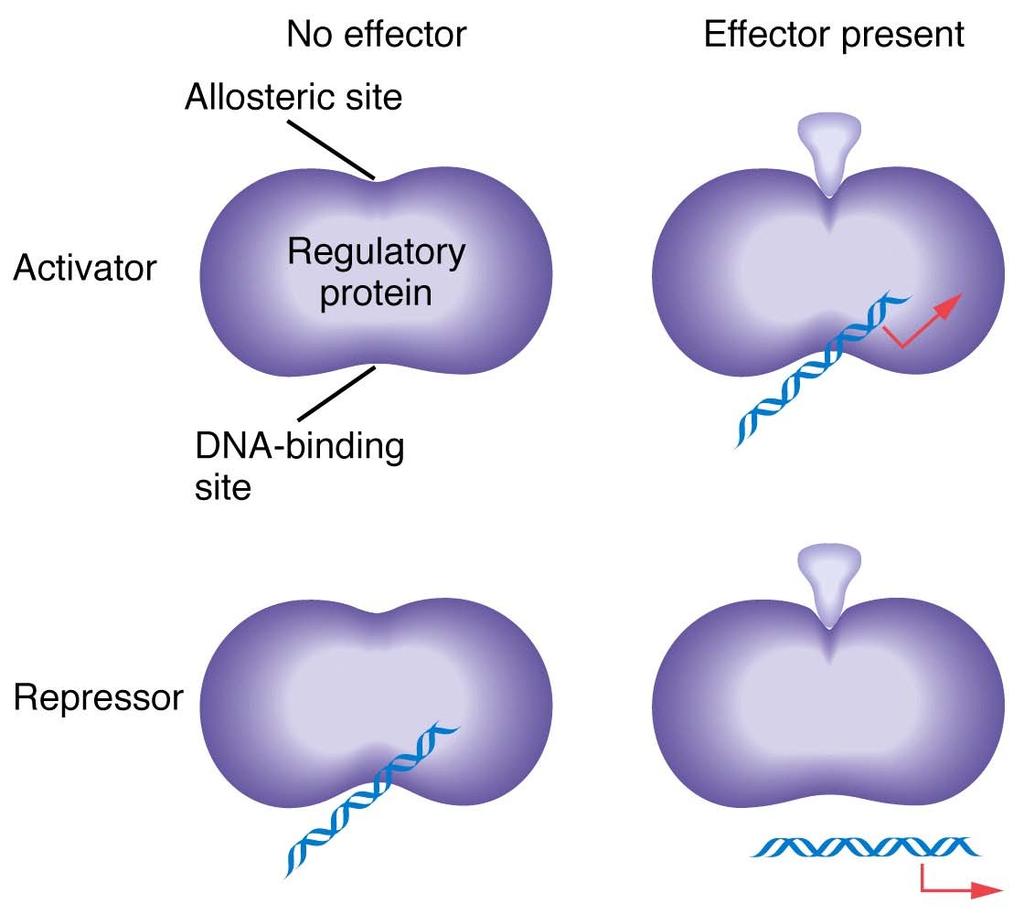
4 Allosteric effectors
5 The lac operon François Jacob and Jacques Monod
6 ß-galactosidase hydrolyses lactose, permease is required for import of lactose into the bacterial cell.
7 Regulation of the lac operon
8 Lac repressor has two recognition sites exists as tetramer of identical subunits single DNA-binding site to dock with O binds to O site with high affinity only one O site in genome prevents transcription when bound to O four allosteric sites that bind lactose and its analogs in absence of lactose or analog, repressor is tightly bound to O, repressing transcription of Z, Y, and A Binding of lactose or analog to allosteric sites reduces affinity of repressor for O, resulting in its release from O and permitting transcription
9 Catabolite repression
10 Catabolite repression is superimposed on the lac and related operons glucose is preferred carbon source induction occurs only in absence of glucose Two components camp (cyclic AMP) when glucose level is low, camp concentration rises because AMP is not converted to ATP catabolite activator protein (CAP) CAP is activated allosterically by camp active CAP binds to CAP site of lac operon, facilitating binding of RNA polymerase

11

12 Genetic lessons from the lac operon Heterozygotes (0 + / 0C) demonstrate that operators are cis-acting. Because a repressor cannot bind to 0 C operators, the lac structural genes linked to an 0 C operator are expressed even in the absence of an inducer. However, the lac genes adjacent to an 0 + operator are still subject to repression.
13
 gene provides a functional repressor that binds to both operators in a")
14 The recessive nature of I - mutations demonstrates that the repressor is transacting. Although no active repressor is synthesized from the I - gene, the wild-type (I + ) gene provides a functional repressor that binds to both operators in a diploid cell and blocks lac operon expression (in the absence of an inducer).
15

16 The dominance of I S mutations is due to the inactivation of the allosteric site on the lac repressor. In an I S / I + diploid cell, none of the lac structural genes are transcribed, even in the presence of an inducer. In contrast with the wild-type repressor, the I S repressor lacks a functional lactose-binding site (the allosteric site) and thus is not inactivated by an inducer. Thus, even in the presence of an inducer, the I S repressor binds irreversibly to all operators in a cell, thereby blocking transcription of the lac operon.
17 Summary
18
 and CAP")
19 Binding sites for lac repressor (a) and CAP (b)

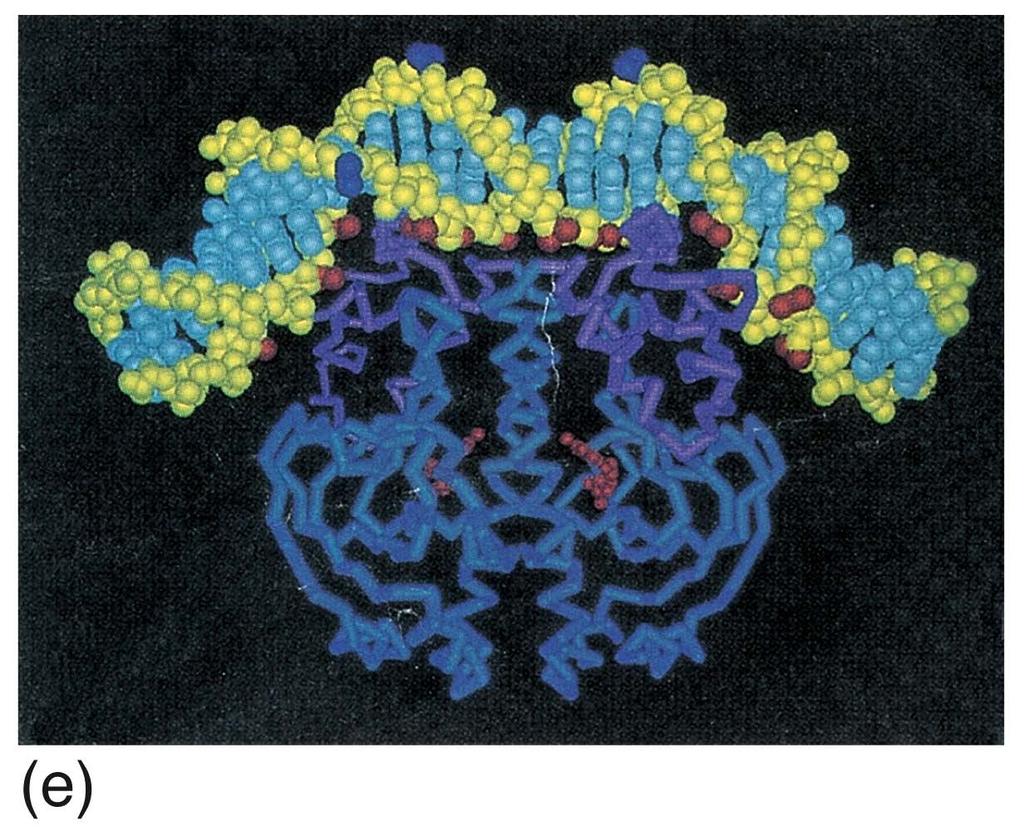
20 Binding of CAP to DNA
21 Comparison of repression and activation.
22 Cis-acting sequences, structure of promoters of eukaryotes
23 The gene control region of a typical eucaryotic gene. The promoter is the DNA sequence where the general transcription factors and the polymerase assemble. The most important feature of the promoter is the TATA box, a short sequence of T-A and A-T base pairs that is recognized by the general transcription factor TFIID. The start point of transcription is typically located about 25 nucleotide pairs downstream from the TATA box. The regulatory sequences serve as binding sites for gene regulatory proteins, whose presence on the DNA affects the rate of transcription initiation. These sequences can be located adjacent to the promoter, far upstream of it, or even downstream of the gene. DNA looping is thought to allow gene regulatory proteins bound at any of these positions to interact with the proteins that assemble at the promoter. Whereas the general transcription factors that assemble at the promoter are similar for all polymerase II transcribed genes, the gene regulatory proteins and the locations of their binding sites relative to the promoter are different for each gene.
24 The modular structure of a transcription factor (activator)
 A 480-nucleotide-pair piece of the eve regulatory region was removed and inserted upstream of a test promoter that directs the synthesis of the enzyme β galactosidase (the product of the E.")
25 The cis-acting binding motifs in promoters are organized in modules Experiment demonstrating the modular construction of the eve gene regulatory region. (A) A 480-nucleotide-pair piece of the eve regulatory region was removed and inserted upstream of a test promoter that directs the synthesis of the enzyme β galactosidase (the product of the E. coli lacz gene). (B) When this artificial construct was reintroduced into the genome of Drosophila embryos, the embryos expressed β galactosidase (detectable by histo-chemical staining) precisely in the position of the second of the seven eve stripes (C).

26 Characterization of promoter sequences by fusion of promotors to reporter genes, followed by transformation.
27 A molecular map of the dpp gene of Drosophila The transcribed region of the gene is indicated below the map. Several tissue specific enhancers are shown.

28 Control of eukaryotic transcription
29 Specific DNA sequence recognition occurs primarily via the major groove. How the different base pairs in DNA can be recognized from their edges without the need to open the double helix. The four possible configurations of base pairs are shown, with hydrogen bond donors indicated in blue, hydrogen bond acceptors in red, and hydrogen bonds themselves as a series of short parallel redlines. Methyl groups, which form hydrophobic protuberances, are shown in yellow, and hydrogen atoms that are attached to carbons, and are therefore unavailable for hydrogen bonding, are white.

30 α helices of proteins are often involved in recognition of DNA sequences
31 Some examples for structural classes of regulatory transcription factors 1. helix-turn-helix protein 2. zinc finger proteins 3. leucin zipper proteins
 is called the recognition helix because it participates in sequence-specific recognition of DNA.")
32 The DNA-binding helix-turn-helix motif. The motif is shown in (A), where each white circle denotes the central carbon of an amino acid. The carboxyl-terminal α helix (red) is called the recognition helix because it participates in sequence-specific recognition of DNA. As shown in (B), this helix fits into the major groove of DNA, where it contacts the edges of the base pairs.
.")
33 Homeodomain proteins are a special class of helix-turn-helix proteins. A homeodomain bound to its specific DNA sequence. The homeodomain is folded into three α helices, which are packed tightly together by hydrophobic interactions (A). The part containing helix 2 and 3 closely resembles the helix-turn-helix motif, with the recognition helix (red) making important contacts with the major groove (B).
34 One type of zinc finger protein. This protein belongs to the Cys-Cys-His-His family of zinc finger proteins, named after the amino acids that grasp the zinc. This zinc finger is from a frog protein of unknown function. (A) Schematic drawing of the amino acid sequence of the zinc finger. (B) The threedimensional structure of the zinc finger is constructed from an antiparallel Œ sheet (amino acids 1 to 10) followed by an α helix (amino acids 12 to 24). The four amino acids that bind the zinc (Cys 3, Cys 6, His 19, and His 23) hold one end of the α helix firmly to one end of the β sheet.
 dimerize through their α helical leucine zipper region (top) to form an inverted Y-shaped structure.")
35 The Leucine Zipper Motif Mediates Both DNA Binding and Protein Dimerization A leucine zipper dimer bound to DNA. Two α helical DNA-binding domains (bottom) dimerize through their α helical leucine zipper region (top) to form an inverted Y-shaped structure. Each arm of the Y is formed by a single α helix, one from each monomer, that mediates binding to a specific DNA sequence in the major groove of DNA. Each α helix binds to one-half of a symmetric DNA structure.

36 Transcription factors often bind as dimers or oligomers Heterodimerization of leucine zipper proteins can alter their DNA-binding specificity.

37 Regulatory transcription factors often act as dimers, trimers or even higher order complexes. This combinatorial mode provides the flexibility of transcription regulation that is needed for life.

38 HLH dimers
39

40 The gel-mobility shift assay. The principle of the assay is shown schematically in (A). In this example an extract of an antibody-producing cell line is mixed with a radioactive DNA fragment containing about 160 nucleotides of a regulatory DNA sequence from a gene encoding the light chain of the antibody made by the cell line. The effect of the proteins in the extract on the mobility of the DNA fragment is analyzed by polyacrylamide-gel electrophoresis followed by autoradiography. The free DNA fragments run rapidly to the bottom of the gel, while those fragments bound to proteins are retarded; the finding of six retarded bands suggests that the extract contains six different sequence-specific DNA-binding proteins (indicated as C1-C6) that bind to this DNA sequence. (For simplicity, any DNA fragments with more than one protein bound have been omitted from the figure.) In (B) the extract was fractionated by a standard chromatographic technique (top), and each fraction was mixed with the radio-active DNA fragment, applied to one lane of a polyacrylamide gel, and analyzed as in (A).
41 The modular structure of a transcription factor (activator)
42 The yeast one-hybrid system: a method for isolation of transcription factors that bind to given DNA cis-motifs. reporter construct: yeast is transformed with this construct. cis-motifs cis-motifs cis-motifs minimal promoter reporter gene activation construct: yeast cells containing the reporter construct are transformed with cdna library. The cdna fragments are fused to an activation domain. yeast promoter activation domain cdna inserts If one cdna encodes a DNA-binding domain that binds to the cis-motif, then the reporter gene is activated. The cdna in these yeast cells encodes the transcript
43 Yeast one-hybrid: inside the cell, selection of a candidate yeast promoter activation domain cdna insert a recombinant transcription factor RNAP cis-motifs cis-motifs cis-motifs minimal promoter reporter gene mrna
44

45 Yeast two-hybrid system for detecting proteins that interact.
46 Yeast two-hybrid system for detecting proteins that interact.
47 Yeast two-hybrid system for detecting proteins that interact. (a) Recombinant DNA techniques can be used to prepare genes that encode hybrid (chimeric) proteins consisting of the DNA-binding domain (purple) or activation domain (orange) of a transcription factor fused to one of two interacting proteins, referred to as the "bait" domain (pink) and "fish" domain (green). (b) If yeast cells are transfected with genes encoding both hybrids, the bait and fish portions of the chimeric proteins interact to produce a functional transcriptional activator. One end of this protein complex binds to the upstream activating sequence (UAS) of a test gene (in this example, the HIS3 gene); the other end, consisting of the activation domain, stimulates assembly of the transcription-initiation complex (gray) at the promoter (yellow). (c) This strategy can be used to screen a cdna library for clones expressing proteins that interact with a protein of interest, in this case Ras. This approach requires two types of plasmids: The bait plasmid includes a DNA sequence encoding the DNA-binding domain of a transcription factor (purple) connected to the coding sequence for Ras (pink). The fish plasmids contain individual cdnas (green) from a library connected to the coding sequence for the activation domain (orange). Each type of plasmid also contains a wild-type selection gene (e.g., TRP1 or LEU2). Both types of plasmids are transfected into yeast cells with mutations in genes required for tryptophan, leucine, and histidine biosynthesis (trp1, leu2, his3 cells) and then grown in the absence of tryptophan and leucine. Only cells that contain the bait plasmid and at least one fish plasmid survive under these selection conditions. The cells that survive then are plated on medium lacking histidine; only cells that contain the bait plasmid and a fish plasmid encoding a protein that binds to Ras are able to grow, thus identifying cdnas encoding Ras-binding proteins.