Käesolev leiutis on seotud FGF-21 polüpeptiididega, mida on valikuliselt modifitseeritud vähemalt ühe mittelooduslikult kodeeritava aminohappega.
|
|
|
- Meghan Boyd
- 6 years ago
- Views:
Transcription
1
2
3 RISTVIITED SARNASTELE PATENDITAOTLUSTELE Selle taotlusega taotletakse prioriteetsust ja eesõigust 30. märtsil 07. aastal esitatud USA eeltaotluse seerianumbriga 60/921,297 suhtes ja 14. novembril 07. aastal esitatud USA eeltaotluse seerianumbriga 60/988,060 suhtes. 5 LEIUTISE VALDKOND Käesolev leiutis on seotud FGF-21 polüpeptiididega, mida on valikuliselt modifitseeritud vähemalt ühe mittelooduslikult kodeeritava aminohappega. LEIUTISE TAUST Fibroblastide kasvufaktorid on suured polüpeptiidid, mida ekspresseeritakse kõikjal arenevates ja täiskasvanud kudedes (Baird et al., Cancer Cells, 3: , 1991) ja nad mängivad väga olulist rolli paljudes füsioloogilistes funktsioonides, sealhulgas angiogeneesis, mitogeneesis, mustrite tekkes, rakkude diferentseerumises, ainevahetuse regulatsioonis ja koekahjustuste parandamises (McKeehan et al., Prog. Nucleic Acid Res. Mol. Biol. 59: , 1998; Burgess, W. H. et al., Annu. Rev. Biochem. 58: (1989). Prototüüpsed fibroblastide kasvufaktorid (FGF-id) ehk FGF-1 ja FGF-2 isoleeriti kõigepealt fibroblastide mitogeenidena ajust ja ajuripatsist. Praeguseks on tehtud kindlaks, et FGF-3 on hiire rinnanäärmekasvaja viiruse poolt aktiveeritav tavapärane sihtmärk (Dickson et al., Ann. N.Y. Acad. Sci. 638:18-26 (1991); FGF-4 kuni FGF-6 on onkogeeni saadused (Yoshida et al., Ann. NY Acad. Sci. 638:27-37 (1991); Goldfarb et al., Ann. NY Acad. Sci 638:38-52 (1991); Coulier et al., Ann. NY Acad. Sci. 638:53-61 (1991)). FGF- tuvastati hiire kopsudes homoloogial põhineva polümeraasi ahelreaktsiooni (PCR) teel (Yamasaki et al., J. Biol. Chem. 271: (1996)). FGF-11 kuni FGF-14 (FGF homoloogilised faktorid (FHF-id) 1 kuni 4) tuvastati inimese võrkkestas juhusliku cdna sekveneerimise, andmebaasides tehtud otsingute ja homoloogial põhineval PCR-meetodil (Smallwood et al., Proc. Natl. Acad. Sci. USA 93: (1996)). Tehti kindlaks, et FGF- on kimäärse homeodomeeni onkovalgu allavoolu sihtmärk (McWhirter et al., Development 124: (1997)). FGF-16, FGF-17 ja FGF-18 tuvastati homoloogial põhineva PCR teel vastavalt roti südames ja embrüotes (Miyake et al., Biochem. Biophys. Res. Commun. 243:148-2 (1998); Hoshikawa et al. Biochem. Biophys. Res. Commun.
4 2 244: (1998); Ohbayashi et al., J. Biol. Chem. 273: (1998)). FGF-19 tuvastati andmebaasides tehtud otsingute abil inimloote ajus (Nishimura et al., Biochim. Biophys. Acta 1444:148-1 (1999)). Neil on konserveerunud, umbes 1 aminohappejäägist koosnev tuum, mille aminohapete identsus on -30 kuni 60% Looduslike mutatsioonide loommudelite, üleekspresseerimise ja analüüside põhjal on fibroblastide kasvufaktoreid ja nende retseptoreid seostatud paljude erinevate haigustega (nt Wilkie et al., Current Biology, (1995) 5: ; Pugh-Humphreys et al, In: The Cytokine Handbook, A. Thomson ed, 2nd edition, Academic Press, Harcourt Brace & co. publishers, London, lk 5-566), millest saab järeldada, et neid haigusi saab ravida fibroblastide kasvufaktorite aktiivsuse reguleerimise teel. Näiteks fibroblastide kasvufaktori 2 inhibeerimise teel ühendiga Suramin saab hiirtel ära hoida neovaskularisatsiooni ja kasvajate arengut (Pesenti et al., British Journal of Cancer, 66: ). Fibroblastide kasvufaktorid osalevad ka angiogeneesis (Lyons, M. K., et al., Brain Res. (1991) 558:3-3), haavade paranemises (Uhl, E., et al., Br. J. Surg. (1993) 80: , 1993), astroglioosis, gliaalrakkude proliferatsioonis ja diferentseerumises (Biagini, G. et al., Neurochem. Int. (1994) :17-24), aju veresoonte laienemises (Tanaka, R. et al., Stroke (1995) 26:24-29) ja neurotroofilistes/neuromodulatoorsetes protsessides. Fibroblastide kasvufaktoritel on ka mitmed positiivsed mõjud ehk nad muuhulgas parandavad ajuisheemia korral verevoolu ja kaitsevad kaltsiumi toksilise toime eest, parandades seega haiguse prognoosi (Mattson, M. P. et al., Semin. Neurosci. (1993) 5: ; Doetrocj. W. D. et al., J. Neurotrauma (1996) 13: ). Aluseline FGF ravi soodustab müokardi isheemia puhul neoangiogeneesi (Schumacher et al., Circulation (1998) 97: ). Aluseline FGF soodustab pärast fokaalset ajuinfarkti kahjustatud funktsioonide taastumist ja uute neuronite teket (Kawamata et al., Proc.Natl. Acad. Sci.(1997) 94 (): ). Erialakirjanduses avaldatud andmetel koosneb FGF perekond vähemalt 22 liikmest (Reuss et al., Cell Tissue Res. 313:139-7 (03)). Avaldatud andmetel ekspresseeritakse fibroblastide kasvufaktorit 21 (FGF-21) eelkõige maksas (Nishimura et al., Biochimica et Biophysica Acta, 1492:3-6 (00); WO 01/36640; ja WO 01/18172) ja seda on kirjeldatud vahendina, mida saab kasutada veresoonkonna isheemiliste haiguse, haavade paranemise ja kopsu-, bronhi- ja
5 alveoolirakkude funktsionaalsuse langusega seotud haiguste ja paljude teiste haiguste ravis. Faktorit FGF-21 ekspresseeritakse peamiselt maksas, neerudes ja lihaskoes (vt USA patendidokumendis nr sisalduvat näidet 2). FGF-21 geen koosneb kolmest eksonist ja paikneb 19. kromosoomil. Erinevalt teistest FGF-idest, faktoril FGF-21 puudub proliferatiivne ja tumorigeenne toime (Genome Biol. 01;2(3):REVIEWS3005). USA patendidokumendis nr kirjeldatakse inimese FGF-21 nukleotiidset ja valgulist järjestust (vt US patendidokumendis nr toodud järjestusi SEQ ID NO: 1 ja 2). Eespool nimetatud publikatsioonis toodud järjestus SEQ ID NO: 2 tähisega sbgfgf-19 on 9 aminohappe pikkune ja selle järjestuse N-otsas paikneb 28 aminohappest koosnev liiderjärjestus. Käesolevas dokumendis toodud inimese FGF-21 järjestus tähisega SEQ ID NO: 3 vastab USA patendidokumendis nr toodud järjestusele SEQ ID NO: 2. Sellel järjestusel on üksiku nukleotiidi polümorfism (SNP), millel on positsioonis 174 proliin (P) ja mida käesolevas dokumendis nimetatakse edaspidi FGF-21 9 aminohappest koosnevaks P-vormiks. USA patendis nr 6,716,626 käsitletakse inimese kasvufaktorit FGF-21 ja teiste loomade, eeskätt hiirte ja rottide organismis leiduvaid homoloogilisi valke. Hiire FGF-i, mis USA patendis nr 6,716,626 oli järjestusega SEQ ID NO: 1, ekspresseeriti intensiivselt maksas ja lisaks ka munandites ja tüümuses ning on jõutud järeldusele, et inimese FGF-21 võib olla seotud maksahaiguste ja/või munandite talitlushäirete või osaleda tüümusest pärinevate rakkude talitlushäirete tekkes ja nendest taastumisel. USA patendis nr toodud järjestus SEQ ID NO: 4 on 9 aminohappe pikkune ja selle N-otsas paikneb 28 aminohappest koosnev liiderjärjestus. Käesolevas dokumendis toodud inimese FGF-21 järjestus tähisega SEQ ID NO: 6 vastab USA patendis nr toodud järjestusele SEQ ID NO: 4. Selle järjestusel on üksiku nukleotiidi polümorfism (SNP), millel on positsioonis 174 leutsiin (L) ja mida käesolevas dokumendis edasipidi nimetatakse FGF aminohappest koosnevaks L-vormiks. USA patendidokumendis nr käsitletakse inimese faktorit FGF-21 ja avaldatakse järjestus, mis on 8 aminohappe pikkune (USA patendipublikatsioonis nr toodud järjestus SEQ ID NO: 2) ja mille N-otsas paikneb 27 aminohappest koosnev liiderjärjestus. Käesolevas dokumendis toodud inimese FGF-21 järjestus tähisega SEQ ID NO: 7 vastab USA patendidokumendis toodud järjestusele SEQ ID NO: 2. Selle
6 4 järjestusel on üksiku nukleotiidi polümorfism (SNP), millel on positsioonis 173 leutsiin (L) ja mida käesolevas dokumendis edasipidi nimetatakse FGF-21 8 aminohappest koosnevaks L-vormiks On näidatud, et FGF-21 stimuleerib insuliini manulusel ja puudumisel hiire 3T3-L1 rasvarakkudes glükoosi imendumist ning vähendab ob/ob ja db/db hiirtel ja 8-nädalastel ZDF rottidel annusest sõltuvalt nii söönud kui ka söömata olekus vere glükoosi-, triglütseriidide ja glükagoonisisaldust, mille põhjal saab faktorit FGF-21 kasutada diabeedi ja rasvtõve ravis (WO 03/ ja Kharitonenkov et al. J Clin Invest. 05 Jun;1(6): ). Kharitonenkov et al. J Clin Invest. 05 Jun;1(6): samuti näitasid, et inimese FGF-21 ekspresseerivad transgeensed hiired on hüpoglükeemilised, tundlikud insuliini suhtes ja neil ei teki toitumisest põhjustatud rasvtõbe. Kharitonenkov et al. Endocrinology (trükkimisel) on näidanud ka seda, et FGF-21 langetas diabeeti põdevate reesusahvide organismis glükoosi-, triglütseriidide, insuliini- ja glükagoonide taset. Lisaks on näidatud, et FGF-21 vähendab efektiivselt kriitiliselt haigete patsientide suremust ja haigestumist (WO 03/059270). USA patenditaotluses kirjeldatu järgi mõjutab FGF-21 üleüldiselt ainevahetuse seisundit ja võib neutraliseerida negatiivseid kõrvaltoimeid, mis võivad tekkida sepsise põhjustatud organismi stressireaktsiooni ajal ja ka mittenakkuslikest patoloogilistest põhjustest tingitud süsteemse põletikulise vastuse sündroomi (SIRS) ajal. Faktorit FGF-21 võib seega kasutada kriitiliselt haigete patsientide suremuse ja haigestumuse vähendamiseks. Kriitiliselt haigete patsientide hulka kuuluvad patsiendid, kes on füsioloogiliselt ebastabiilsed ja vajavad pidevat arsti järelevalvet, põetamist ja hingamisabi. Sellise hoolduse ehk pideva järelevalve ja ravi optimeerimise tagamiseks tuleb pöörata suurt tähelepanu üksikasjadele. Kriitiliselt haigete patsientide hulka loetakse ka patsiendid, kellel on füsioloogilise dekompensatsiooni tekke oht ja keda tuleb seetõttu pidevalt jälgida, et intensiivravi spetsialistid saaksid soovimatute tagajärgede ärahoidmiseks vajadusel kohe sekkuda. Spetsialistid peavad kriitiliselt haiged patsiendid vajavad jälgimist ja hingamisabi, mille peab tagama spetsialistid, kes oskavad ravi pidevalt optimeerida. PEGüleeritud FGF-21 polüpeptiide kirjeldatakse patendidokumendis WO 05/ Patendidokumendis WO 05/ kirjeldatav FGF-21 polüpeptiid on 181
7 aminohappest koosnev polüpeptiid. Küpsel, metsikut tüüpi või looduslikul inimese FGF- 21 järjestusel, mis patendidokumendis WO 05/ vastab järjestusele SEQ ID NO: 1, puudub liiderjärjestus. Selline inimese FGF-21 on suurel määral identne hiire FGF-21 ( 79% aminohappeline identsus) ja roti FGF-21 ( 80% aminohappeline identsus). Käesolevas dokumendis toodud inimese FGF-21 järjestus SEQ ID NO: 5 vastab patendidokumendis WO 05/ toodud järjestusele SEQ ID NO: 1. Sellel järjestusel on üksiku nukleotiidi polümorfism (SNP), millel on positsioonil 146 leutsiin (L). Asjatundjad oskavad käesolevas dokumendis kirjeldatavatel kasutusviisidel hõlpsasti kasutada ka teisi imetaja FGF-21 polüpeptiidide järjestusi või analooge, muteiine või derivaate, mis on piisavalt homoloogilised inimese FGF-21 järjestustega. Käesolevas dokumendis toodud inimese FGF-21 järjestusel SEQ ID NO: 1 on üksiku nukleotiidi polümorfism (SNP), millel on positsioonis 146 proliin (P). Järjestuse SEQ ID NO: 1 versioon, millel on N-otsas His märgis, vastab käesolevas dokumendis järjestusele SEQ ID NO: 2. Patendidokumendis WO 05/ kirjeldatakse ühe või mitme PEG molekuli kovalentset kinnitamist FGF-21 ühendi konkreetsete aminohappejääkide külge. Selle tulemusena saadi bioloogiliselt aktiivne PEGüleeritud FGF-21 ühend, millel on võrreldes loodusliku FGF-21-ga pikem eliminatsiooni poolväärtusaeg ja aeglasem kliirens. PEG-molekulid kinnitati kovalentselt tsüsteiini- või lüsiinijääkide külge. Asendused tehti tsüsteiiniga mitmetes positsioonides, et oleks võimalik kinnitada vähemalt üks PEGmolekul. Nimetatud dokumendis kirjeldati ühe või mitme lüsiinijäägi (56, 59, 69 ja 122) PEGüleerimist. PEGüleeritud FGF-21 ühenditega saab ravida patsiente, kellel on häired ehk näiteks, kuid mitte ainult, 2. tüübi diabeet, rasvtõbi, insuliiniresistentsus, hüperinsulineemia, glükoositalumatus, hüperglükeemia ja metaboolne sündroom. Eriti kasulikud oleksid PEGüleeritud FGF-21 ühendid, mis muudavad ravi efektiivsemaks ehk neil pikem poolväärtusaeg veres ja neid saab manustada väiksemate annustena, mis muudab ravi patsiendi jaoks mugavamaks ja suurendab manustamisrežiimist kinnipidamise tõenäosust. Metaboolne sündroom on vähemalt kolme järgmise ilmingu kombinatsioon: alakõhu rasvumine, millele on enamikul meestest viitab 40-tolline või suurem vööümbermõõt; kõrge veresuhkru tase ehk tühja kõhu korral vähemalt 1 mg /dl; kõrge triglütseriidide tase veres ehk vähemalt 0 mg/dl; madal HDL tase ehk madalam kui 40 mg/dl ja vererõhk 130/85 või kõrgem.
8 Hüdrofiilse polümeeri polü(etüleenglükooli) lühendiga PEG kovalentne kinnitamine on meetod, mille abil saab parandada paljude bioloogiliselt aktiivsete molekulide, sealhulgas valkude, peptiidide ja eeskätt hübrofoobsete molekulide vees lahustuvust ja biosaadavust, pikendada poolväärtusaega seerumis ja terapeutilist poolväärtusaega, muuta immunogeensust ja bioloogilist aktiivsust või pikendada vereringes püsimise aega. PEG-i on kasutatud laialdaselt ravimites, tehisimplantaatides ja teistes rakendustes, mille korral on oluline bioloogiline ühilduvus ning toksilisuse ja immunogeensuse puudumine. Selleks et maksimeerida PEG-i soovitud omadusi, peab bioloogiliselt aktiivse molekuli külge kinnitatava PEG polümeeri või polümeeride üldine molekulmass ja hüdratatsiooniaste olema piisavad suured, et saavutada PEG-molekuli lisamisest tulenevad kasulikud omadused nagu parem lahustuvus vees ja poolväärtusaeg vereringes, mõjutamata sealjuures soovimatul viisil vanemmolekuli bioaktiivsust. PEG derivaate sageli seotakse bioloogiliselt aktiivsete molekulidega reaktsioonivõimeliste funktsionaalrühmade nagu lüsiini-, tsüsteiini- ja histidiinijäägid, N-ots ja süsivesikfragmendid kaudu. Praeguseks on uuritud raviotstarbelise FGF-21 ühendi valmistusvõimalusi, kuid see on paljudel põhjustel problemaatiline: üheks neist on asjaolu, et valkudel ja teistel molekulidel on sageli piiratud arv reaktsioonivõimelisi kohti, kuhu saab kinnitada polümeeri. Kohad, mis sobivad kõige paremini polümeeri kinnitamise teel modifitseerimiseks, mängivad sageli olulist rolli retseptoriga seondumises ja on seega vajalikud molekuli bioloogilise aktiivsuse säilitamiseks. Polümeeriahelate valimatul kinnitamisel bioloogiliselt aktiivse molekuli sellistesse reaktsioonivõimelistesse kohtadesse väheneb polümeeriga modifitseeritud molekuli bioloogiline aktiivsus oluliselt või kaob täielikult (R. Clark et al., (1996), J. Biol. Chem., 271: ). Selleks et saada konjugaate, mille polümeeri molekulmassist piisab sihtmolekulile soovitud kasulike omaduste tagamiseks, on tehnika tasemes tuntud strateegiate korral üldjuhul kinnitatud selle molekuli külge juhuslikult hulk polümeerseid õlgu, mille tulemusena väheneb vanemmolekuli bioaktiivsuse vähenemise või isegi täieliku kadumise oht. Reaktsioonivõimelised kohad, mis moodustavad PEG derivaatide kinnitamiseks sobivad lookused, sõltuvad valgu struktuurist. Valgud, sealhulgas ensüümid, koosnevad alfaaminohapete mitmesugustest järjestustest üldstruktuuriga H 2 N--CHR COOH. Ühe aminohappe alfa-aminofragment (H 2 N--) on ühendatud sellega külgneva aminohappe karboksüülrühmaga (-COOH), moodustades amiidsideme tähisega --(NH--CHR-CO) n --,
9 7 milles n võib olla sadades või tuhandetes olev arv. Fragment tähisega R sisaldab reaktsioonivõimelisi alasid, mis on vajalikud valgu bioloogilise aktiivsuse tagamiseks ja PEG derivaatide kinnitamiseks Näiteks aminohappe lüsiin korral on --NH 2 fragment nii epsilon- kui ka alfa-asendis. Epsilon-asendis olev -NH 2 -rühm on alulise ph korral reaktsioonivõimeline. Paljude valkude PEG-iga derivatiseerimise meetodite korral püütakse luua PEG derivaadid, mis kinnitatakse valkudes sisalduvate lüsiinijääkide epsilon --NH 2 fragmendi külge. Polyethylene Glycol and Derivatives for Advanced PEGylation, Nektar Molecular Engineering Catalog, 03, lk Sellistel PEG derivaatidel on aga ühine puudus ehk neid ei saa kinnitada selektiivselt valkude pinnal olevate, sageli arvukate lüsiinijääkide hulka. See võib olla oluliseks puuduseks juhtudel, kui lüsiinijääk on valgu aktiivsuse seisukohalt oluline ehk näiteks kui see paikneb ensüümi aktiivsel saidil või kui lüsiinijääk osaleb valgu ja teiste bioloogiliste molekulide vahelise interaktsiooni vahendamises nagu retseptori sidumiskohtade puhul. Olemasolevate valgu PEGüleerimismeetodite teiseks ja sama oluliseks puuduseks on asjaolu, et PEG derivaadid võivad osaleda soovimatutes kõrvalreaktsioonides teiste, soovimatute aminohappejääkidega. Histidiin sisaldab reaktsioonivõimelist iminofragmenti struktuurivalemiga - N(H)-, kuid paljud keemiliselt reaktsioonivõimelised molekulid, mis reageerivad epsilon -NH 2 rühmaga, reageerivad ka - N(H)---fragmendiga. Aminohappe tsüsteiini kõrvalahelas on ka vaba sulfhüdrüülrühm struktuurivalemiga -SH. Mõnel juhul reageerivad lüsiini epsilon--nh 2 rühmale suunatud PEG derivaadid ka tsüsteiini-, histidiini- ja teiste aminohappejääkidega. Selle tulemusel võivad moodustuda PEGderivatiseeritud bioaktiivsete molekulide komplekssed heterogeensed segud ja tekkida sihitava bioaktiivse molekuli aktiivsuse kaotamise oht. Seega on soovitav töötada välja PEG-i derivaadid, mille puhul saab valgu ühele saidile sisestada keemiliselt funktsionaalse rühma, mille abil saab seejärel siduda bioaktiivse molekuliga selektiivselt ühe või mitu PEG polümeeri, kasutades selleks valgu pinnal paiknevaid spetsiifilisi kohti, mis on nii hästi määratletud kui ka ennustatavad. Selles valdkonnas on lisaks lüsiinijääkidega seotud uurimistööle tehtud ka palju katseid töötada välja aktiveeritud PEG reagente, mis on suunatud teistele aminohapetest koosnevatele kõrvalahelatele, sealhulgas tsüsteiinile, histidiinile ja N-otsale. Vt nt USA
10 patenti nr 6,6,281 ja allikat Polyethylene Glycol and Derivatives for Advanced PEGylation, Nektar Molecular Engineering Catalog, 03, lk Tsüsteiinijäägi saab sisestada valkude struktuuri saitide osas selektiivselt kohtsuunatud mutageneesi teel ja teistel tehnika tasemes tuntud meetoditel ja tekkinud vaba sulfhüdrüülfragmendi saab panna reageerima PEG derivaatidega, millel on tiooliga reageerivad funktsionaalrühmad. Sellise strateegia kasutamist raskendab aga asjaolu, et vaba sulfhüdrüülrühma sisestamine võib muuta saadava valgu ekspressiooni, kokkuvoltimise ja stabiilsuse tagamise keerukaks. Seega oleks vaja vahendit, mille abil saab FGF-21 polüpeptiidi sisestada keemilise funktsionaalrühma, mis võimaldab valguga selektiivselt siduda üks või mitu PEG polümeeri ja sobib samas (ehk ei osale soovimatutes kõrvalreaktsioonides) sulfhüdrüülfragmentide ja teiste tavaliselt valkudes leiduvate funktsionaalrühmadega. Nagu on näha paljudest sellest valdkonnast pärinevatest näidetest, on paljude selliste derivaatide, mis on välja töötatud valkude kõrvaahelate külge kinnitamiseks, eriti aminohappe lüsiini kõrvalahelas paikneva -- NH 2 fragmendi ja tsüsteiini kõrvalahelas paikneva SH fragmendi külge kinnitamiseks, sünteesimine ja kasutamine osutunud keerukaks. Mõned neist moodustavad valguga ebastabiilsed sidemed, mis vesikeskkonnas nagu vereringe hüdrolüüsuvad ja seega lõhustuvad, lagunevad või on muul viisil ebastabiilsed. Mõned neist küll moodustavad stabiilsemad sidemed, kuid võivad hüdrolüüsuda enne sideme moodustumist ehk PEG derivaadi reaktsioonivõimeline rühm võidakse enne valgu külge kinnitamist inaktiveerida. Mõned neist on mõnevõrra toksilised ja seega ei sobi nii hästi in vivo kasutamiseks. Mõned neist reageerivad liiga aeglaselt ja ei ole seetõttu praktikas kasulikud. Mõned neist võivad valgu aktiivsuse seisukohalt olulistesse kohtadesse kinnitamise korral kaotada valgu aktiivsuse. Mõned ei ole kinnituskohtades spetsiifilised, mille tulemusel võib soovitud aktiivsus kaduda ja tulemused ei ole reprodutseeritavad. Selleks et lahendada probleemid, mis kaasnevad valkude modifitseerimisega polü(etüleenglükool) fragmentidega, on välja töötatud PEG derivaadid, mis on senisest stabiilsemad (nt USA patent 6,602,498) või mis reageerivad selektiivselt molekulidel ja pindadel paiknevate tiooli molekulidega (nt USA patent 6,6,281). Selles valdkonnas vajatakse selgelt PEG derivaate, mis on füsioloogilistes keskkondades keemiliselt inertsed kuni nad peavad selektiivselt reageerima ja moodustama stabiilsed keemilised sidemed.
11 Hiljuti teatati täiesti uuest valkude keemia vallas väljatöötatud tehnoloogiast, mille abil peaks olema võimalik lahendada palju valkude kohtspetsiifilise modifitseerimisega seotud probleeme. Täpsemalt, prokarüoodi Escherichia coli (E. coli) (nt L. Wang, et al., (01), Science 292: ) ja eukarüoodi Sacchromyces cerevisiae (S. cerevisiae) (nt J. Chin et al., Science 301:964-7 (03)) valgu biosünteesi mehhanismile on lisatud uusi koostisosi, mis on võimaldanud valkudesse in vivo sisestada mittegeneetiliselt kodeeritud aminohappeid. Sellisel meetodil on vastusena amberkoodonile TAG sisestatud E.coli bakterites ja pärmis valkudesse efektiivselt ja suure usaldusväärsusega palju uute keemiliste, füüsikaliste või bioloogiliste omadustega uusi aminohappeid, sealhulgas fotoafiinsuse märgiseid ja fotoisomeriseeritavaid aminohappeid, fotoristseonduvaid aminohappeid (vt nt Chin, J. W., et al. (02) Proc. Natl. Acad. Sci. U. S. A. 99:1-124; ja Chin, J. W., et al., (02) J. Am. Chem. Soc. 124: ), ketoaminohappeid, raskeid aatomeid sisaldavaid aminohappeid ja glükosüülitud aminohappeid. Vt nt J. W. Chin et al., (02), Journal of the American Chemical Society 124: ; J. W. Chin, & P. G. Schultz, (02), ChemBioChem 3(11): ; J. W. Chin, et al., (02), PNAS United States of America 99:1-124; ja L. Wang, & P. G. Schultz, (02), Chem. Comm., 1:1-11. Need uuringud on näidanud, et sellisel viisil on võimalik selektiivselt ja rutiinselt sisestada keemiliselt funktsionaalseid rühmi nagu ketoon-, alküün- ja asiidrühmad, mida ei leidu valkudes ja mis on kõigi tavapärases geneetiliselt kodeeritud aminohappes leiduvate funktsionaalrühmade suhtes keemiliselt inertsed ja mille abil saab efektiivselt ja selektiivselt moodustada stabiilseid kovalentseid sidemeid. Tänu võimalusele sisestada valkudesse mittegeneetiliselt kodeeritud aminohappeid saab sisestada valkudesse keemiliselt funktsionaalseid rühmi, mis võivad olla väärtuslikeks alternatiivideks looduslikele funktsionaalrühmadele nagu lüsiini epsilon -NH 2, tsüsteiini sulfhüdrüül SH-rühm histidiini iminorühm, jne. Teatud keemiliselt funktsionaalsed rühmad on teadaolevalt inertsed tavapärases geneetiliselt kodeeritud aminohappes leiduvate funktsionaalrühmade suhtes, kuid nad reageerivad puhtalt ja efektiivselt ning moodustavad stabiilsed sidemed. Selles valdkonnas on näiteks teada, et asiid- ja atsetüleenrühmadega toimub vesikeskkonnas katalüütilise koguse vase manulusel Huisgeni [3+2] tsükloliitumise reaktsioon. Vt nt Tornoe, et al., (02) J. Org. Chem. 67: ; ja Rostovtsev, et al., (02) Angew. Chem. Int. Ed. 41:96-99.
12 5 Asiidfragmendi sisestamise teel valgustruktuuri näiteks on võimalik sisestada funktsionaalrühm, mis on keemiliselt inertne valkudes sisalduvate amiinide, sulfhüdrüülide, karboksüülhapete ja hüdroksüülrühmade suhtes ja mis samas reageerivad sujuvalt ja efektiivselt atsetüleenfragmendiga ja moodustava tsükloliitumise saaduse. Oluline on see, et atsetüleenfragmendi puudumise korral jääb asiidrühm teiste valgu kõrvalahelate juuresolekul ja füsioloogilistes tingimustes keemiliselt inertseks ja ei reageeri nendega. Allikates Nishimura et al. (00) BBA 1492, 3-6 ja WO01/36640 kirjeldatakse keemilist modifitseerimist nagu FGF-21 PEGüleerimine. Patendidokumendis WO05/ kirjeldatakse modifitseeritud või täiustatud omadustega inimese kasvuhormooni polüpeptiide. Patendidokumentides WO06/ ja WO05/ kirjeldatakse FGF-21 keemilist modifitseerimist eesmärgiga parandada selle omadusi. Patendidokumentides US 06/1946 ja US kirjeldatakse mittelooduslikke aminohappeid sisaldavaid modifitseeritud polüpeptiide ja nende valmistusmeetodeid. Käesoleva leiutisega lahendatakse muu hulgas FGF-21 polüpeptiidide aktiivsuse ja tootmisega seotud probleeme ja see leiutis on seotud ka täiustatud bioloogiliste või farmakoloogiliste omadustega, sealhulgas parema terapeutilise poolväärtusajaga FGFpolüpeptiidi tootmisega. LEIUTISE OLEMUS Käesoleva leiutisega pakutakse FGF-21 polüpeptiid järjestusega SEQ ID NO: 1-7, milles üks aminohape positsioonis 8 on asendatud mittelooduslikult kodeeritud aminohappega (SEQ ID NO:1 või vastavad aminohapped järjestustes SEQ ID NO: 2-7), kusjuures vees lahustuv polümeer seotakse nimetatud FGF-21 polüpeptiidis sisalduva mittelooduslikult kodeeritud aminohappega. Mõnes teostuses seotakse FGF-21 polüpeptiid linkeri, polümeeri või bioloogiliselt aktiivse molekuliga. Mõnes teostuses seotakse FGF-polüpeptiid bifunktsionaalse polümeeri, bifunktsionaalse linkeri või vähemalt ühe teise FGF-21 polüpeptiidiga.
13 5 11 Mittelooduslikult kodeeritav aminohape seotakse vees lahustuva polümeeriga. Mõnes teostuses sisaldab vees lahustuv polümeer polü(etüleenglükooli) fragmenti. Mõnes teostuses seotakse mittelooduslikult kodeeritud aminohape vees lahustuva polümeeriga linkeri kaudu või seotakse vees lahustuva polümeeriga. Mõnes teostuses on polü(etüleenglükooli) molekul bifunktsionaalne polümeer. Mõnes teostuses on bifunktsionaalne polümeer seotud teise polüpeptiidiga. Mõnes teostuses on nimetatud teine polüpeptiid FGF-21 polüpeptiid. Mõnes teostuses sisaldab FGF-21 polüpeptiid vähemalt kahte aminohapet, mis on seotud polü(etüleenglükooli) fragmenti sisaldava vees lahustuva polümeeriga. Mõnes teostuses on nimetatud vähemalt üks aminohape mittelooduslikult kodeeritud aminohape. Üks mittelooduslikult kodeeritud aminohape on sisestatud FGF-21 polüpeptiidis järgmisesse positsiooni (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). Mõnes teostuses kloonitakse FGF-21 sekretsiooni konstruktid vektorisse (Nde/Eco) koos liiderjärjestustega, mis on valitud järjestuste SEQ ID NOs: 39, 40, 41, 42, 43 ja 44 seast. Mõnes teostuses seotakse ühes neist positsioonidest paiknev aminohape vees lahustuva polümeeriga positsioonis 8 (SEQ ID NO: 1 või vastava aminohappe järjestustes SEQ ID NOs: 2-7). Leiutise mõnes teostuses on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis moduleerib FGF-21 polüpeptiidi afiinsust FGF-21 polüpeptiidi või seondumispartneri suhtes, sealhulgas, kuid mitte ainult, valgu, polüpeptiidi, väikese molekuli või nukleiinhappe suhtes. Leiutise teatud teostuste korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis suurendab FGF-21 polüpeptiidi stabiilsust võrreldes sama FGF-21 polüpeptiidiga, millel puudub selline asendus, aditsioon või deletsioon. Leiutise teatud teostuste korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis moduleerib FGF-21 polüpeptiidi immunogeensust võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi immunogeensusega. Leiutise teatud teostuste korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis moduleerib FGF-21 polüpeptiidi poolväärtusaega seerumis või vereringes püsimise aega
14 12 võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi poolväärtusajaga seerumis või vereringes püsimise ajaga Leiutise teatud teostuste korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis suurendab FGF polüpeptiidi lahustuvust vees võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi lahustuvusega vees. Leiutise mõne teostuse korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis suurendab peremeesrakus toodetud FGF-21 polüpeptiidi lahustuvust võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi lahustuvusega. Leiutise mõnes teostuses on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis suurendab FGF- 21 polüpeptiidi ekspressiooni peremeesrakus või sünteesi in vivo võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi ekspressiooni või sünteesiga. Sellise asendusega FGF-21 polüpeptiidil säilib agonistlik toime ja see säilitab või suurendab ekspressioonitaset peremeesrakus. Leiutise mõne teostuse korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis suurendab FGF-21 polüpeptiidi valgu vastupidavust võrreldes, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi vastupidavusega. USA patendis nr on näidatud, et potentsiaalsed kohad, mida võib proteaasiga lõhustamise muutmiseks asendada, on näiteks, kuid mitte ainult, proliini kahe aminohappejäägi kohta tulev ühealuseline sait Leiutise mõne teostuse korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis moduleerib FGF-21 retseptori signaali ülekande aktiivsust võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi interaktsioonis osaleva sama retseptori aktiivsusega. Leiutise mõne teostuse korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis moduleerib selle seondumist teise molekuliga nagu retseptor võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi seondumisega. Leiutise mõne teostuse korral on FGF-21 polüpeptiidis asendus, aditsioon või deletsioon, mis suurendab FGF-21 polüpeptiidi ühilduvust farmatseutiliste säilitusainetega (nt m- kresooli, fenooli, bensüülalkoholiga) võrreldes sama, kuid ilma asenduse, aditsiooni või deletsioonita FGF-21 polüpeptiidi ühilduvusega. Tänu sellisele paremale ühilduvusele saab valmistada säilitatava ravipreparaadi, millel säilivad säilitamise ajal valgu füüsikaliskeemilised omadused ja bioloogiline aktiivsus. Patendidokumendis WO 05/091944
15 käsitletakse näiteid FGF-21 muteiinidest, millel on suurem farmatseutiline stabiilsus: üks või mitu järgmistest aminohapetest on asendatud laetud ja/või polaarse, kuid laenguta aminohappega: patendidokumendis WO 05/ toodud järjestuses SEQ ID NO: 1 glütsiin 42, glutamiin 54, arginiin 77, alaniin 81, leutsiin 86, fenüülalaniin 88, lüsiin 122, histidiin 1, arginiin 126, proliin 130, arginiin 131, leutsiin 139, alaniin 145, leutsiin 146, isoleutsiin 2, alaniin 4, glutamiin 6, glütsiin 161, seriin 163, glütsiin 170 või seriin 172. Käesolevas leiutise kohases FGF-21 polüpeptiidis võib selle polüpeptiidi vastavas positsioonis olla üks või mitu sellist asendust, aditsiooni või deletsiooni. Leiutise mõnes teostuses tehakse üks või mitu asendust mitteloodusliku aminohappega ühes või mitmes järgmises positsioonis: glütsiin 42, glutamiin 54, arginiin 77, alaniin 81, leutsiin 86, fenüülalaniin 88, lüsiin 122, histidiin 1, arginiin 126, proliin 130, arginiin 131, leutsiin 139, alaniin 145, proliin/leutsiin 146, isoleutsiin 2, alaniin 4, glutamiin 6, glütsiin 161, seriin 163, glütsiin 170, seriin 172 (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). Leiutise mõnes teostuses tehakse üks või mitu asendust mitteloodusliku aminohappega ühes või mitmes järgmises positsioonis: glutamaat 91, arginiin 131, glutamiin 8, arginiin 77, arginiin 72, histidiin 87, leutsiin 86, arginiin 126, glutamaat 1, türosiin 83, proliin 146, arginiin 135, arginiin 96, arginiin 36, (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). Patendidokumendis WO 05/ kirjeldatakse FGF-21 teisi suurema farmatseutilise stabiilsusega muteiine. Sellistel muteiinidel on kaks või rohkem FGF-21 järgmist aminohapet asendatud tsüsteiiniga (vt SEQ ID NO: 1 patendidokumendis WO 05/091944):arginiin 19, türosiin, leutsiin 21, türosiin 22, treoniin 23, aspartaat 24, aspartaat, alaniin 26, glutamiin 27, glutamiin 28, alaniin 31, leutsiin 33, isoleutsiin 35, leutsiin 37, valiin 41, glütsiin 42, glütsiin 43, glutamaat 50, glutamiin 54, leutsiin 58, valiin 62, leutsiin 66, glütsiin 67, lüsiin 69, arginiin 72, fenüülalaniin 73, glutamiin 76, arginiin 77, aspartaat 79, glütsiin 80, alaniin 81, leutsiin 82, glütsiin 84, seriin 85, proliin 90, alaniin 92, seriin 94, fenüülalaniin 95, leutsiin 0, aspartaat 2, türosiin 4, türosiin 7, seriin 9, glutamaat 1, proliin 1, histidiin 117, leutsiin 118, proliin 119, asparagiin 121, lüsiin 122, seriin 123, proliin 124, histidiin 1, arginiin 126, aspartaat 127, alaniin 129, proliin 130, glütsiin 132, alaniin 134, arginiin 135, leutsiin 137, proliin 138 või leutsiin 139. Käesoleva leiutise kohastel FGF-21 polüpeptiididel võib olla polüpeptiidi vastavas positsioonis üks või mitu sellist asendust ja/või neis võib olla üks või
16 mitu teist asendust, aditsiooni või deletsiooni. Leiutise mõnes teostuses on üks või mitu mitteloodusliku aminohappe asendust ühes või mitmes järgmises positsioonis: arginiin 19, türosiin, leutsiin 21, türosiin 22, treoniin 23, aspartaat 24, aspartaat, alaniin 26, glutamiin 27, glutamiin 28, alaniin 31, leutsiin 33, isoleutsiin 35, leutsiin 37, valiin 41, glütsiin 42, glütsiin 43, glutamaat 50, glutamiin 54, leutsiin 58, valiin 62, leutsiin 66, glütsiin 67, lüsiin 69, arginiin 72, fenüülalaniin 73, glutamiin 76, arginiin 77, aspartaat 79, glütsiin 80, alaniin 81, leutsiin 82, glütsiin 84, seriin 85, proliin 90, alaniin 92, seriin 94, fenüülalaniin 95, leutsiin 0, aspartaat 2, türosiin 4, türosiin 7, seriin 9, glutamaat 1, proliin 1, histidiin 117, leutsiin 118, proliin 119, asparagiin 121, lüsiin 122, seriin 123, proliin 124, histidiin 1, arginiin 126, aspartaat 127, alaniin 129, proliin 130, glütsiin 132, alaniin 134, arginiin 135, leutsiin 137, proliin 138 või leutsiin 139 (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). Patendidokumendis WO 05/ kirjeldatakse sisseviidud disulfiidsidemetega FGF-21 spetsiifilisi muteiine (mille aminohapped on asendatud tsüsteiiniga), millel lisaks looduslikule disulfiidsidemele Cys75-Cys93 vahel on disulfiidsidemed Gln76Cys- Ser9Cys, Cys75-Ser85Cys, Cys75-Ala92Cys, Phe73Cys-Cys93, Ser123Cys-His1Cys, Asp2Cys-Tyr4Cys, Asp127Cys-Gly132Cys, Ser94Cys-Glu1Cys, Pro1Cys- His117Cys, Asn121Cys-Asp127Cys, Leu0CysAsp2Cys, Phe95Cys-Tyr7Cys, Arg19CysPro138Cys, TyrCys-Leu139Cys, Tyr22Cys-Leu137Cys, Arg77Cys- Asp79Cys, Pro90Cys-Ala92Cys, Glu50Cys-Lys69Cys, Thr23Cys-AspCys, Ala31Cys- Gly43Cys, Gln28Cys-Gly43Cys, Thr23Cys-Gln28Cys, Val41Cys-Leu82Cys, Leu58Cys- Val62Cys, Gln54Cys-Leu66Cys, Ile35Cys-Gly67Cys, Gly67Cys-Arg72Cys, Ile35Cys- Gly84Cys, Arg72Cys-Gly84Cys või Arg77Cys-Ala81Cys, vahel, milles kasutatud arvud vastavad patendidokumendis WO 05/ toodud järjestusele SEQ ID NO: 1. Teised töödeldud disulfiidsidemetega muteiinid on Tyr22Cys-Leu139Cys; Asp24Cys- Arg135Cys; Leu118Cys-Gly132Cys; His117Cys-Pro130Cys; His117Cys-Ala129Cys; Leu82Cys-Pro119Cys; Gly80Cys-Ala129Cys; Gly43Cys-Pro124Cys; Gly42Cys- Arg126Cys; Gly42Cys-Pro124Cys; Gln28Cys-Pro124Cys; Gln27Cys-Ser123Cys; Ala26Cys-Lys122Cys; or AspCys-Lys122Cys, milles kasutatud arvud vastavad patendidokumendis WO 05/ toodud järjestusele SEQ ID NO: 1. Teised töödeldud disulfiidsidemetega muteiinid on Leu118Cys-Ala134Cys; Leu21Cys-Leu33Cys; Ala26Cys-Lys122Cys; Leu21Cys-Leu33Cys/Leu118Cys-Ala134Cys, milles kasutatud
17 5 30 arvud vastavad patendidokumendis WO 05/ toodud järjestusele SEQ ID NO: 1. Käesoleva leiutise kohastes FGF-21 polüpeptiidides võib polüpeptiidi vastavas positsioonis/positsioonides olla üks või mitu sellist asendust või nendes võib olla üks või mitu teistsugust asendust, aditsiooni või deletsiooni. Leiutisekohastel FGF-21 polüpeptiidides võib olla polüpeptiidi vastavates positsioonides (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7) üks või mitu sellist asendust. Mõnes teostuses võib käesoleva leiutise kohastel FGF-21 polüpeptiididel olla vastavates positsioonides ehk järjestustes SEQ ID NOs: enne positsiooni 1 (ehk N-otsas) kuni C-otsani üks või mitu sellist asendust. Patendidokumendis WO 05/ kirjeldatakse FGF-21 teisi, PEGüleeritud muteiine. Sellistel muteiinidel oli üks järgmistest asendustest: DC, D38C, L58C, K59C, P60C, K69C, D79C, H87C, E91C, E1C, D2C L114C, L116C, K122C, R126C, P130C, P133C, P140C. Käesoleva leiutise kohastel FGF-21 polüpeptiididel võib olla polüpeptiidi vastavates positsioonides üks või mitu sellist asendust ja/või neis võib olla üks või mitu teist asendust, aditsiooni või deletsiooni. Leiutise teatud teostuste korral asendatakse ühe või mitme mitteloodusliku aminohappega üks või mitu aminohapet ühes või mitmes järgmises positsioonis:, 38, 58, 59, 60, 69, 79, 87, 91, 1, 2, 114, 116, 122, 126, 130, 133, 140 (SEQ ID NO: I või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). Leiutise teatud teostustes võib leiutisekohastel FGF-21 polüpeptiididel olla järjestustes SEQ ID NOs: vastavates positsioonides enne positsiooni 1 (ehk N-otsas) kuni C- otsani üks või mitu sellist asendust. Patendidokumendis WO 05/ kirjeldatakse tsüsteiini asendusi järgmistes positsioonides: 19, 21, 26, 28, 29, 30, 36, 39, 42, 50, 56, 6I, 64, 65, 68, 70, 71, 77, 81, 85, 86, 90, 92, 94, 98, 7, 8, 112, 113, 123 ja 124. Patendidokumendis WO 05/ kirjeldatakse tsüsteiini asendusi järgmistes positsioonides: 24, 27, 37, 40, 44, 46, 49, 57, 88, 89, 6, 1, 111, 1, 1 ja 139. Patendidokumendis WO 05/ samuti kirjeldatakse tsüsteiini asendusi järgmistes positsioonides: 18, 45, 47, 48, 78, 83, 99, 3, 1, 128, 131, 132 ja 138. Patendidokumendis WO 05/ samuti kirjeldatakse tsüsteiini asendusi järgmistes positsioonides:, 38, 58, 59, 60, 69, 79, 87, 91, 1, 2, 114, 116, 122, 126, 130, 133 ja 140.
18 5 16 Mõnes teostuses luuakse ühe või mitme mitteloodusliku aminohappega üks või mitu töödeldud sidet. Molekulisisese sideme võib luua paljudel viisidel, näiteks, kuid mitte ainult, valgus kahe aminohappe vahelise reaktsiooni teel sobivates tingimustes (aminohapetest üks või mõlemad võivad olla mittelooduslikud); kahe aminohappe, millest kumbki võib olla looduslikult kodeeritud või mittelooduslikult kodeeritud aminohappe) reaktsiooni teel linkeri, polümeeri või muu molekuliga sobivates tingimustes, jne. Leiutise teatud teostustes võib FGF-21 polüpeptiidis võib üks või mitu aminohapet olla asendatud ühe või mitme loodusliku või mitteloodusliku aminohappega. Leiutise mõnes teostuses võib FGF-21 polüpeptiidis võib üks või mitu aminohapet olla asendatud ühe või mitme loodusliku või mitteloodusliku aminohappega, tingimusel et vähemalt üks asendus on tehtud mittelooduslikult kodeeritud aminohappega. Leiutise mõnes teostuses võib FGF- 21 polüpeptiidis olla üks või mitu aminohapet asendatud ühe või mitme loodusliku aminohappega ja lisaks on vähemalt üks asendus tehtud mittelooduslikult kodeeritud aminohappega. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape karbonüül-, atsetüül-, aminooksü-, hüdrasiin-, hüdrasiid-, semikarbasiid-, asiid- või alküünrühma. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape karbonüülrühma. Mõnes teostuses on mittelooduslikult kodeeritud aminohappe struktuur: milles n on 0-; R 1 on alküül-, arüül- või asendatud arüülrühm; R 2 on H, alküül-, arüül-, asendatud alküül- ja asendatud arüülrühm; ja R 3 on H, aminohape, polüpeptiid või aminootsa modifitseerimisrühm ja R 4 on H, aminohape, polüpeptiid või karboksü-otsa modifitseerimisrühm. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape aminooksürühma. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape hüdrasiidrühma. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape hüdrasiinrühma. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape semikarbasiidrühma.
19 17 Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape asiidrühma. Mõnes teostuses on mittelooduslikult kodeeritud aminohappe struktuur: 5 milles n on 0-; R 1 on alküül-, arüül-, asendatud alküül-, asendatud arüülrühm või puudub; m on 0-; R 2 on H, aminohape, polüpeptiid või amino-otsa modifitseerimisrühm ja R 3 on H, aminohape, polüpeptiid või karboksü-otsa modifitseerimisrühm. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape alküünrühma. Mõnes teostuses on mittelooduslikult kodeeritud aminohappe struktuur: milles n on 0-; R 1 on alküül-, arüül-, asendatud alküül- või asendatud arüülrühm; X on O, N, S või puudub; m on 0-, R 2 on H, aminohape, polüpeptiid või amino-otsa modifitseerimisrühm ja R 3 on H, aminohape, polüpeptiid või karboksü-otsa modifitseerimisrühm. Leiutise mõnes teostuses on leiutisekohane polüpeptiid FGF-21 polüpeptiidide agonist, osaline agonist, antagonist, osaline antagonist või pöördagonist. Leiutise mõnedes teostustes sisaldab FGF-21 polüpeptiidide agonist, osaline agonist, antagonist, osaline antagonist või pöördagonist mittelooduslikult kodeeritud aminohapet, mis on seotud vees lahustuva polümeeriga. Mõnes teostuses sisaldab vees lahustuv polümeer polü(etüleenglükool)fragmenti. Leiutise mõnes teostuses sisaldab FGF-21 polüpeptiidide agonist, osaline agonist, antagonist, osaline antagonist või pöördagonist mittelooduslikult kodeeritud aminohapet ja ühte või mitut translatsioonijärgset modifikatsiooni, linkerit, polümeeri või bioloogiliselt aktiivset molekuli. Käesoleva leiutisega pakutakse ka isoleeritud nukleiinhapped, mis sisaldavad polünukleotiidi, mis hübridiseerub karmides tingimustes järjestustega SEQ ID NO: Käesoleva leiutisega pakutakse ka isoleeritud nukleiinhapped, mis sisaldavad polünukleotiidi, mis hübridiseerub karmides tingimustes järjestustega SEQ ID NO: 8-14,
20 kusjuures selline polünukleotiid sisaldab vähemalt üht selektorkoodonit. Käesoleva leiutisega pakutakse ka isoleeritud nukleiinhapped, mis sisaldavad polünukleotiidi, mis kodeerib polüpeptiide, mis on näidatud järjestustena SEQ ID NOs.: 1-7. Käesoleva leiutisega pakutakse ka isoleeritud nukleiinhapped, mis sisaldavad polünukleotiidi, mis kodeerib järjestustes SEQ ID NOs.: 1-7 näidatud polüpeptiide, milles on üks või mitu mittelooduslikult kodeeritud aminohapet. Asjatundjad mõistavad, et mis tahes leiutisekohane polüpeptiid võib kodeerida paljusid erinevaid polünukleotiide. Mõnes teostuses valitakse selektorkoodon rühmast, kuhu kuuluvad amberkoodon, ookerkoodon, opaalkoodon, unikaalne koodon, haruldane koodon, viiealuseline koodon ja neljaaluseline koodon. Käesoleva leiutisega pakutakse ka meetodid vees lahustuva polümeeriga seotud FGF-21 polüpeptiidi valmistamiseks. Mõnes teostuses viiakse leiutisekohase meetodi korral mittelooduslikult kodeeritud aminohapet sisaldav isoleeritud FGF-21 polüpeptiid kokkupuutesse vees lahustuva polümeeriga, mis sisaldab mittelooduslikult kodeeritud aminohappega reageerivat fragmenti. Mõnes teostuses reageerib FGF-21 polüpeptiidi sisestatud mittelooduslikult kodeeritud aminohape vees lahustuva polümeeriga, mis üldjuhul ei reageeri tavapärase aminohappega. Mõnes teostuses reageerib FGF-21 polüpeptiidi sisestatud mittelooduslikult kodeeritud aminohape linkeri, polümeeri või bioloogiliselt aktiivse molekuliga, mis üldjuhul ei reageeri tavapärase aminohappega. Mõnes teostuses valmistatakse vees lahustuva polümeeriga seotud FGF-21 polüpeptiid karbonüülrühma sisaldavat aminohapet sisaldava FGF-21 polüpeptiidi reaktsiooni teel polü(etüleenglükooli) molekuliga, mis sisaldab aminooksü-, hüdrasiin-, hüdrasiid- või semikarbasiidrühma. Mõnes teostuses seotakse aminooksü-, hüdrasiin-, hüdrasiid- või semikarbasiidrühm polü(etüleenglükooli) molekuliga amiidsideme kaudu. Mõnes teostuses valmistatakse vees lahustuva polümeeriga seotud FGF-21 polüpeptiid karbonüülrühma sisaldava polü(etüleenglükooli) molekuli reaktsiooni teel polüpeptiidiga, mis sisaldab mittelooduslikult kodeeritud aminohapet, milles on aminooksü-, hüdrasiin-, hüdrasiid- või semikarbasiidrühm. Mõnes teostuses valmistatakse vees lahustuva polümeeriga seotud FGF-21 polüpeptiid alküünrühma sisaldavat aminohapet sisaldava FGF-21 polüpeptiidi reaktsiooni teel asiidrühma sisaldava polü(etüleenglükooli) molekuliga.
21 Mõnes teostuses seotakse asiid- või alküünrühm polü(etüleenglükooli) molekuliga amiidsideme kaudu. Mõnes teostuses valmistatakse vees lahustuva polümeeriga seotud FGF-21 polüpeptiid asiidrühma sisaldavat aminohapet sisaldava FGF-21 polüpeptiidi reaktsiooni teel alküünrühma sisaldava polü(etüleenglükooli) molekuliga. Mõnes teostuses seotakse asiid- või alküünrühm polü(etüleenglükooli) molekuliga amiidsideme kaudu. Mõnes teostuses on polü(etüleen)glükooli molekuli molekulmass umbes 0,1 kda kuni umbes 0 kda. Mõnes teostuses on polü(etüleen)glükooli molekuli molekulmass umbes 0,1 kda kuni 50 kda. Mõnes teostuses on polü(etüleen)glükooli molekul hargnenud polümeer. Mõnes teostuses on polü(etüleen)glükooli molekul hargnenud polümeeri iga ahela molekulmass 1 kda kuni 0 kda või 1 kda kuni 50 kda. Mõnes teostuses sisaldab FGF-21 polüpeptiidiga seotud vees lahustuv polümeer polüalküleenglükooli fragmenti. Mõnes teostuses sisaldab FGF-21 polüpeptiidi sisestatud mittelooduslikult kodeeritud aminohappejääk karbonüül-, aminooksü-, hüdrasiid-, hüdrasiin-, semikarbasiid, asiid- või alküünrühma. Mõnes teostuses sisaldab FGF-21 polüpeptiidi sisestatud mittelooduslikult kodeeritud aminohappejääk karbonüülrühma ja vees lahustuv polümeer sisaldab aminooksü-, hüdrasiid-, hüdrasiin- või semikarbasiidrühma. Mõnes teostuses sisaldab FGF-21 polüpeptiidi sisestatud mittelooduslikult kodeeritud aminohappejääk alküünrühma ja vees lahustuv polümeer sisaldab asiidrühma. Mõnes teostuses sisaldab FGF-21 polüpeptiidi sisestatud mittelooduslikult kodeeritud aminohappejääk asiidrühma ja vees lahustuv polümeer sisaldab alküünrühma. Käesoleva leiutisega pakutakse ka kompositsioonid, mis sisaldavad mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi ja farmatseutiliselt sobivat tugiainet. Mõnes teostuses on mittelooduslikult kodeeritud aminohape seotud vees lahustuva polümeeriga. Käesoleva leiutisega pakutakse ka rakud, mis sisaldavad polünukleotiidi, mis kodeerib selektorkoodonit sisaldavat FGF-21 polüpeptiidi. Leiutise mõnede teostuste korral sisaldavad rakud ortogonaalse RNA süntetaasi ja/või ortogonaalset trna-d, mille abil saab FGF-21 polüpeptiidi sisestada mittelooduslikult kodeeritud aminohappe. Käesoleva leiutisega pakutakse ka meetodid mittelooduslikult kodeeritud aminohapet sisaldava FGF- 21 polüpeptiidi valmistamiseks.
22 5 30 Leiutise teatud teostuste korral hõlmab leiutisekohane meetod FGF-21 polüpeptiidi kodeerivat/kodeerivaid polünukleotiidi/polünukleotiide sisaldavate rakkude, ortogonaalse RNA süntetaasi ja/või ortogonaalse trna kasvatamist tingimustes, kus on võimalik ekspresseerida FGF-21 polüpeptiidi, ja FGF-21 polüpeptiidi eraldamist rakkudest ja/või rakukultuurisöötmest. Käesoleva leiutisega pakutakse ka meetodid FGF-21 polüpeptiidide terapeutilise poolväärtusaja, seerumi poolväärtusaja või vereringes püsimise aja pikendamiseks. Käesoleva leiutisega pakutakse ka meetodid FGF-21 polüpeptiidide immunogeensuse moduleerimiseks. Mõnes teostuses hõlmavad leiutisekohased meetodid looduslikes FGF- 21 polüpeptiidides positsioonis 8 ühe aminohappe asendamist mitteloodusliku aminohappega (SEQ ID No:1 või vastavad aminohapped järjestustes SEQ ID Nos: 2-7) ja/või FGF-21 sidumist linkeri, polümeeri, vees lahustuva polümeeri või bioloogiliselt aktiivse molekuliga. Käesoleva leiutisega pakutakse ravimeetodid, mille korral manustatakse sellist ravi vajavale patsiendile efektiivses koguses käesoleva leiutise kohast FGF-21 molekuli. Leiutisekohaste meetodite teatud teostuste korral manustatakse patsiendile terapeutiliselt efektiivses koguses farmatseutilist kompositsiooni, mis sisaldab mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi ja farmatseutiliselt sobivat tugiainet. Mittelooduslikult kodeeritud aminohape on seotud vees lahustuva polümeeriga. Käesoleva leiutisega pakutakse ka FGF-21 polüpeptiid, mis sisaldab järjestust SEQ ID NO: 1-7 või tahes muud FGF-21 järjestust, milles üks aminohape positsioonis 8 on asendatud mittelooduslikult kodeeritud aminohappega (SEQ ID No:1 või vastavad aminohapped järjestustes SEQ ID Nos: 2-7). Käesoleva leiutisega pakutakse ka FGF-21 polüpeptiidid, mis sisaldavad järjestust SEQ ID NO: 1, 2, 4 ja 5. Mõnes teostuses seotakse mittelooduslikult kodeeritud aminohape vees lahustuva polümeeriga. Mõnes teostuses sisaldab vees lahustuv polümeer polü(etüleenglükool) fragmenti. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape karbonüül-, aminooksü-, hüdrasiid-, hüdrasiin-, semikarbasiid-, asiid- või alküünrühma. Käesoleva leiutisega pakutakse ka farmatseutilised kompositsioonid, mis sisaldavad farmatseutiliselt sobivat tugiainet ja FGF-21 polüpeptiidi järjestusega SEQ ID NO: 1-7 või mis tahes muud FGF-21 polüpeptiidi järjestust, milles üks aminohape on asendatud positsioonis 8 mittelooduslikult kodeeritud aminohappega (SEQ ID No:1 või vastavad aminohapped
23 järjestustes SEQ ID Nos: 2-7). Käesoleva leiutisega pakutakse farmatseutilised kompositsioonid, mis sisaldavad farmatseutiliselt sobivat tugiainet ja FGF-21 polüpeptiidi järjestusega SEQ ID NO: 1-7 või mis tahes muud FGF-21 polüpeptiidi järjestust, milles üks aminohape positsioonis 8 (SEQ ID No:1 või vastavad aminohapped järjestustes SEQ ID Nos: 2-7) on asendatud mittelooduslikult kodeeritud aminohappega. Mõnes teostuses sisaldab mittelooduslikult kodeeritud aminohape sahhariidfragmenti. Mõnes teostuses seotakse vees lahustuv polümeer polüpeptiidiga sahhariidfragmendi kaudu. Mõnes teostuses seotakse linker, polümeer või bioloogiliselt aktiivne molekul FGF-21 polüpeptiidiga sahhariidfragmendi kaudu. Käesoleva leiutisega pakutakse ka FGF-21 polüpeptiid, mis sisaldab vees lahustuvat polümeeri, mis on seotud ühe aminohappe juures kovalentse sideme abil FGF-21 polüpeptiidiga. Mõnes teostuses sisaldab vees lahustuv polümeer polü(etüleenglükooli) fragmenti. Vees lahustuva polümeeriga kovalentselt seotud aminohape on polüpeptiidis sisalduv mittelooduslikult kodeeritud aminohape. Käesoleva leiutisega pakutakse FGF-21 polüpeptiid, mis sisaldab vähemalt ühte linkerit, polümeeri või bioloogiliselt aktiivset molekuli, kusjuures see linker, polümeer või bioloogiliselt aktiivne molekul kinnitatakse polümeeri külge polüpeptiidi ribosomaalselt sisestatud mittelooduslikult kodeeritud aminohappe funktsionaalrühma kaudu. Mõnes teostuses on polüpeptiid monopegüleeritud. Käesoleva leiutisega pakutakse ka FGF-21 polüpeptiid, mis sisaldab linkerit, polümeeri või bioloogiliselt aktiivset molekuli, mis kinnitub ühe mittelooduslikult kodeeritud aminohappe külge, kusjuures see mittelooduslikult kodeeritud aminohape sisestatakse polüpeptiidi eelselekteeritud kohtadesse ribosomaalselt. Leiutise ulatusse kuulub FGF-21 liider- või signaaljärjestus, mis ühendatakse FGF-21 kodeeriva piirkonnaga, ja ka heteroloogiline signaaljärjestus, mis seotakse FGF-21 kodeeriva piirkonnaga. Selline heteroloogiline järjestus peaks olema järjestus, mida peremeesraku sekretsioonisüsteem suudab ära tunda ja töödelda, et seda oleks võimalik peremeesrakus sekreteerida ja signaalpeptidaasiga lõhustada. Käesoleva leiutise kohased liiderjärjestused võib valida järgmiste järjestuste seast: kolme leutsiini sisalduv liiderjärjestus järjestustest SEQ ID NO: 3 ja SEQ ID NO: 6 (aminohapete positsioonid 1-28), kahte leutsiini sisalduv liiderjärjestus järjestustest SEQ ID NO: 4 ja SEQ ID NO: 7 (aminohapete positsioonid 1-27), His märgis vastavalt järjestusele SEQ ID NO: 2 (aminohapete positsioonid 1-), SEQ ID NO: 39, SEQ ID NO: 40, SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43, SEQ ID NO: 44. Haigusseisundi või häire
24 5 22 ravimeetodil, mille korral kasutatakse leiutisekohast FGF-21 polüpeptiidi, kasutatakse ravimiseks FGF-21 koos signaal- või liiderpolüpeptiidiga või ilma selleta. Käesoleva leiutisega pakutakse ka meetodid glükoosi haarde suurendamise indutseerimiseks rasvarakkudes, kusjuures selle meetodi puhul manustatakse FGF-21 nimetatud rakkudesse koguses, mille abil saab efektiivselt indutseerida glükoosi haarde suurenemise. Selline glükoosi haarde suurenemine võib glükoosi kiirema ja efektiivsema kasutamise toel suurendab energiakulu. Teises teostuses saadakse ühte mittelooduslikku aminohapet sisaldava FGF-21 konjugeerimisel teise molekuliga, näiteks, kuid mitte ainult PEG-iga, põhimõtteliselt puhastatud FGF-21, mille tagab mitteloodusliku aminohappe konjugeerimiseks kasutatav unikaalne keemiline reaktsioon. Ühte mittelooduslikult kodeeritud aminohapet sisaldava FGF-21 võib konjugeerida teise molekuliga nagu PEG enne või pärast konjugeerimisetappi kasutatavate teiste puhastusmeetodite raames, et saada üldjoontes puhas FGF-21. JOONISTE LÜHIKIRJELDUS FIG 1 Sellel joonisel on kujutatud amber-mutatsiooni FGF-21 polüpeptiidis ja vastavaid saite FGF-19 polüpeptiidis. FIG 2 Sellel joonisel on kujutatud inimese FGF-19 struktuuri. FIG 3 Sellel joonisel on kujutatud amber-mutatsiooni FGF-21 polüpeptiidis ja vastavaid saite FGF-2 polüpeptiidis. FIG 4 Sellel joonisel on kujutatud inimese FGF-19 struktuuri. FIG 5 Sellel joonisel on kujutatud FGF-21, mille N-otsas on His märgis, ekspressiooni ja supressiooni 7 ambersaitidel. FIG 6 Sellel joonisel on kujutatud N-terminaalse His märgisega FGF-21 ekspressiooni ja supressiooni 7 ambersaitide BPER supernatantide proove. FIG 7a SigmaPlot, mille abil arvutatakse FGF21 variantide 30K PEG-391, 30K PEG- 477, 30K PEG-R131, 30K PEG-Q8, HIS-FGF21 (His-märgisega metsikut tüüpi) seerialahjenduste EC50 väärtused.
25 23 FIG 7b Joonisel olevas tabelis on toodud loetletud PEGüleeritud FGF21 variantide aktiivsuse keskmine langus kordades. FIG 8 E.coli bakterites ekspresseeritud His-märgiseta FGF-21 SDS-PAGE analüüs. 5 FIG 9 - (A) FGF-21-Y83pAF elueeritud fraktsioonide SDS-PAGE analüüs. (B) Märgiseta FGF-21-Y83pAF Q HP elueerimise kromatogramm. (C) FGF-21-Y832pAFQ HP koondatud eluaatide SDS-PAGE analüüs FIG - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 11 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 12 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 13 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 14 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 16 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 17 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 18 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 19 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis.
26 24 FIG - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 21 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. 5 FIG 22 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 23 - Näitest 28 pärinevad andmed, FGF-21 ühendite farmakoloogilised omadused rottide organismis. FIG 24 - pvk-fgf21 vektori kaart. FIG - pvk-fgf21 vektori järjestus. FIG 26a N-6His WT FGF21 kolme annuse seerumikontsentratsiooni ja aja profiilid rottide organismis. Rottidele manustati üks kord subkutaanselt katseühendit. N= 4 looma rühmas. Sümbolitega on tähistatud mõõdetud seerumikontsentratsioonid, veatulbad vastavad standardhälbele. FIG 26b Seerumikontsentratsiooni ja aja profiilid subkutaanselt või intravenoosselt kontsentratsioonis 0, mg/kg manustatud N-6His WT FGF21 korral. Rottidele manustati subkutaanselt üks annus katseühendit. N= 4 looma rühmas. Sümbolitega on tähistatud mõõdetud seerumikontsentratsioonid, veatulbad vastavad standardhälbele. Üldine biosaadavus on 87%. FIG 27a Katseühendi annuse ja seerumikontsentratsiooni suhe Cmax korral. Cmax väärtused on täheldatud väärtused, mitte teoreetilised väärtused. Lineaarse regressiooni väärtus on 0,59 tõusuga 348,5 ± 91,22. FIG 27b Katseühendi annuse ja terminal poolväärtusaja suhe. N=4 looma katserühmas. Lineaarse regressiooni väärtust ei saanud kliirensi ilmse küllastatuse tõttu kontsentratsioonidel üle 0, mg/kg arvutada.
27 FIG 27c Annuse ja seerumikontsentratsiooni AUC (kõveraalune pindala) suhe. AUC väärtused on esitatud originaalkujul arvutatuna lõpmatuse suhtes. N=4 looma ravirühmas. Lineaarse regressiooni väärtus on 0,75 tõusuga 79 ± 194,1 5 FIG 28a PP WT FGF21 kolme annuse seerumikontsentratsiooni ja aja profiilid rottidel. Rottidele manustati subkutaanselt üks annus katseühendit. N= 4 looma rühmas. Sümbolitega on tähistatud mõõdetud seerumikontsentratsioonid, veatulbad vastavad standardhälbele. FIG 28b Seerumikontsentratsiooni ja aja profiilid subkutaanselt või intravenoosselt kontsentratsioonis 0, mg/kg manustatud PP WT FGF21 korral. Rottidele manustati subkutaanselt üks annus katseühendit. N= 4 looma rühmas. Sümbolitega on tähistatud mõõdetud seerumikontsentratsioonid, veatulbad vastavad standardhälbele. Üldine biosaadavus on 65%. FIG 29a Katseühendi annuse ja seerumikontsentratsiooni suhe Cmax korral. Cmax väärtused on täheldatud väärtused, mitte teoreetilised väärtused. N=4 looma ravirühmas. Lineaarse regressiooni väärtus on 0,92 tõusuga 454,2 ± 42,42. FIG 29b Katseühendi annuse ja terminal poolväärtusaja suhe. N=4 looma ravirühmas. Lineaarse regressiooni väärtust ei saanud kliirensi näiva küllastumise tõttu väärtustel üle 0, mg/kg arvutada. FIG 29c Annuse ja seerumi kontsentratsiooni AUC (kõveraalune pindala) suhe. AUC väärtused on esitatud originaalkujul arvutatuna lõpmatuse suhtes. N=4 looma ravirühmas. Lineaarse regressiooni väärtus on 0,93 tõusuga 85 ± 137,1. FIG 30a Rottidele subkutaanselt 0,5 mg/kg suuruse annusena manustatud PP versus N6- His WT FGF21 ühendite puhul arvutatud terminaalse poolväärtusaja võrdlus. Kahepoolse t-testi abil arvutati p väärtus on 0,77. N=3-4 looma rühmas. FIG 30b Cmax väärtuste võrdlus PP versus N6-His WT FGF21 ühendite korral, mida manustati rottidele subkutaanselt kontsentratsioonis 0,5 mg/kg. Kahepoolse t-testiga arvutatud p väärtus on 0,7652. N=3-4 looma rühmas.
28 26 FIG 30c AUCinf võrdlus PP versus N6-His WT FGF21 ühendite korral, mida manustati rottidele subkutaanselt kontsentratsioonis 0,5 mg/kg. Kahepoolse t-testi abil arvutatud p- väärtus on 0, FIG 31a Kümne PEGüleeritud, N6-His märgisega FGF21 isomeeri farmakokineetilised profiilid FIG 31b PEGüleeritud FGF21 isomeeride absorptsiooniprofiilid pärast 0, mg/kg subkutaanset süstimist. FIG 31c PEGüleeritud FGF21 isomeeride eliminatsiooniprofiilid pärast nende subkutaanset süstimist kontsentratsioonis 0, mg/kg. FIG 32 - ja 30 kda PEGüleerimise farmakokineetiline võrdlus. FIG 33 Plasmakontsentratsiooni ja aja kõverad rottidel, kellele manustati intravenoosselt või subkutaanselt 0, mg/kg KPEG-pAF91(N6-His)FGF21. Igale loomale manustati üks annus. N=4 looma rühmas. Sümbolitega on tähistatud mõõdetud plasmakontsentratsioonid, tulbad vastavad standardhälbele. Üldine biosaadavus on 30%. FIG 34 Kaks geeli, mis näitavad FGF-21 sekretsiooni E.coli bakterites ja asjaolu, et OmpA, MalE, and StII toimisid väga hästi nagu näitab periplasmast vabanenud lahustuv fraktsioon geelis number 2. MÕISTED Käesoleval juhul tuleb mõista, et see leiutis ei piirdu käesolevas dokumendis kirjeldatavate konkreetsete meetodite, protokollide, rakuliinide, konstruktide ja reagentidega ning et selles võib esineda variatsioone. Lisaks tuleb mõista, et käesolevas dokumendis kasutatav terminoloogia on mõeldud vaid konkreetsete teostuste kirjeldamiseks ja see ei piira käesoleva leiutise ulatust, mis on määratletud kaasasolevas patendinõudluses. Kui kontekst ei näita selgelt teisiti, peetakse käesolevas dokumendis ja sellega kaasasolevas patendinõudluses ainsuse vormide all silmas ka vastavate sõnade mitmuse vorme, Seega näiteks peetakse mõiste FGF-21 või FGF-21 polüpeptiid all silmas ühte või mitut sellist valku ja nende ekvivalente, mis on asjatundjatele teada, jne.
29 Kui ei ole määratletud teisiti, on kõik käesolevas tekstis kasutatavad tehnilised ja teaduslikud terminid selle leiutise valdkonna asjatundjale tuntud tavapärase tähendusega. Kuigi selle leiutise teostamisel või katsetamisel võib kasutada mis tahes meetodeid, vahendeid ja materjale, mis sarnanevad käesolevas dokumendis kirjeldatutele või on nendega samaväärsed, kirjeldatakse alljärgnevalt eelistatud meetodeid, vahendeid ja materjale. Kõik käesolevas dokumendis nimetatavad publikatsioonid ja patendid on mõeldud nendes publikatsioonides kirjeldatavate näiteks konstruktide ja meetodite kirjeldamiseks ja avaldamiseks, mida võib kasutada ka käesoleva leiutise korral. Käesolevas dokumendis käsitletavad publikatsioonid on esitatud vaid seetõttu, et nad on avaldatud enne käesoleva taotluse esitamise kuupäeva. Ühtegi käesolevas dokumendis esitatud fakti ei saa käsitleda tõdemusena, et käesoleva leiutise autoritel puudub eelneva leiutise tõttu või muul põhjusel õigus prioriteetsusele. Mõiste üldjoontes puhastatud viitab FGF-21 polüpeptiidile, mis ei sisalda põhimõtteliselt ega olulisel määral koostisosi, mis tavaliselt leiduvad koos või interakteeuvad valguga selle looduslikus keskkonnas ehk looduslikus rakus või rekombinantselt toodetud FGF-21 polüpeptiidide korral peremeesrakus. FGF-21 polüpeptiid, mis ei sisalda olulisel määral rakulist materjali, hõlmab valgupreparaate, mis sisaldavad vähem kui umbes 30%, vähem kui umbes %, vähem kui umbes %, vähem kui umbes %, vähem kui umbes %, vähem kui umbes 5%, vähem kui umbes 4%, vähem kui umbes 3%, vähem kui umbes 2% või vähem kui umbes 1% (kuivmassi järgi) saastavat valku. Kui FGF21 polüpeptiidi või selle varianti toodetakse rekombinantselt peremeesrakkudes, võib valgu sisaldus moodustada umbes 30%, umbes %, umbes %, umbes %, umbes %, umbes 5%, umbes 4%, umbes 3%, umbes 2% või umbes 1% või vähem rakkude kuivmassist. Kui FGF21 polüpeptiidi või selle varianti toodetakse rekombinantselt peremeesrakkudes, võib kasvusöötmes sisalduv valk moodustada umbes 5 g/l, umbes 4 g/l, umbes 3 g/l, umbes 2 g/l, umbes 1 g/l, umbes 750 mg/l, umbes 500 mg/l, umbes 0 mg/l, umbes 0 mg/l, umbes 50 mg/l, umbes mg/l, või umbes 1 mg/l või vähem rakkude kuivmassist. Käesoleva leiutise kohastel meetoditel saadud üldjoontes puhastatud FGF-21 polüpeptiidi puhtusaste võib olla vähemalt umbes 30%, vähemalt umbes 35%, vähemalt
30 umbes 40%, vähemalt umbes 45%, vähemalt umbes 50%, vähemalt umbes 55%, vähemalt umbes 60%, vähemalt umbes 65%, vähemalt umbes 70%, konkreetsemalt vähemalt umbes 75%, 80%, 85% ja veelgi konkreetsemalt vähemalt umbes 90%, vähemalt umbes 95%, vähemalt umbes 99% või suurem, mis on määratud asjakohastel meetoditel nagu SDS/PAGE analüüs, RP-HPLC, SEC ja kapillaarelektroforees. Rekombinantne peremeesrakk või peremeesrakk on rakk, mis sisaldab endogeenset polünukleotiidi, olenemata selle sisestusmeetodist ehk sellest, kas see sisestatud näiteks otsese sisestamise, transduktsiooni, f-mating või teiste tehnika tasemes tuntud rekombinantsete peremeesrakkude loomise meetoditel. Eksogeenset polünukleotiidi võib selles rakus säilitada mitteintegreeritud vektorina, näiteks plasmiidina, või integreerida selle peremeesraku genoomi. Käesolevas kontekstis on sööde või söötmed mis tahes kasvusööde, lahus, tahke, pooltahke või jäik tugistruktuur, mis võib toetada või sisaldada mis tahes peremeesrakku, sealhulgas, bakteriaalseid peremeesrakke, pärmi peremeesrakke, putuka peremeesrakke, taimseid peremeesrakke, eukarüootseid peremeesrakke, imetaja peremeesrakke, CHOrakke, prokarüootseid peremeesrakke, E. coli või Pseudomonas peremeesrakke ja rakkude sisu. See mõiste võib seega hõlmata söödet, milles peremeesrakku on kasvatatud, nt söödet, millesse FGF-21 polüpeptiidi sekreteeriti, sealhulgas söödet enne või pärast proliferatsioonietappi. See mõiste hõlmab ka puhvreid või reagente, mis sisaldavad peremeesraku lüsaate nagu juhul, kui FGF-21 polüpeptiidi toodetakse intratsellulaarselt ja FGF-21 polüpeptiidi vabastamiseks peremeesrakud lüüsitakse või lõhutakse. Redutseeriv aine on käesolevas dokumendis valkude kokkuvoltimise kontekstis mis tahes ühend või materjal, mis säilitab redutseeritud olekus sulfhüdrüülrühmad ja redutseerib molekulidesiseseid ja vahelisi disulfiidsidemeid. Sobivate redutseerivate ainete seas on näiteks, kuid mitte ainult, ditiotreitool (DTT), 2-merkaptoetanool, ditioerütritool, tsüsteiin, tsüsteamiin (2-aminoetaantiool) ja redutseeritud glutatioon. Asjatundjatele on ilmne, et käesoleva leiutise kohaste meetodite ja kompositsioonide puhul saab kasutada paljusid erinevaid redutseerivaid aineid. Oksüdeeriv aine on käesolevas dokumendis valkude kokkuvoltimise kontekstis mis tahes ühend või materjal, millel on võime eemaldada oksüdeeritavast ühendist elektron. Sobivate oksüdeerivate ainete seas on näiteks, kuid mitte ainult, oksüdeeritud glutatioon,
31 29 tsüstiin, tsüstamiin, oksüdeeritud ditiotreitool, oksüdeeritud erütreitool ja hapnik. Asjatundjatele on ilmne, et käesoleva leiutise kohaste meetodite puhul võib kasutada paljusid erinevaid oksüdeerivaid aineid Diabeedivastane raviaine on mis tahes ravim, mis on kasulik mis tahes glükoosi ainevahetuse häire või selle mis tahes komplikatsiooni, sealhulgas mis tahes käesolevas dokumendis kirjeldatava seisundi, haiguse või komplikatsiooni ravis, ennetamisel või selle ägeduse muul viisil vähendamisel. Diabeedivastaste raviainete seas on insuliin, tiasolidiindioonid, sulfonüüluuread, bensoehappe derivaadid, alfa-glükosidaasi inhibiitorid ja teised sarnased ühendid. Kõnealusesse kompositsiooni kuuluda võivate diabeedivastaste raviainete teiste üldkategooriate seas on (määratletud mõisted on toodud jutumärkides): ravimiartiklid (drug articles), mida tunnustatakse ametlikus Ameerika Ühendriikide Farmakopöas või ametlikus dokumendis National Formulary (või selle mis tahes lisas); uus ravim ja uus loomaravim (new animal drug), mille on heaks kiitnud USA Ravimija Toiduamet (FDA); neid mõisteid kasutatakse Ameerika Ühendriikide seadustikus alajaotises 21 (Title 21 of the United States Code); mis tahes ravim, mille puhul on vajalik USA või välisriigi valitsusüksuse heakskiit ( heakskiidetud ravim ); mis tahes ravim, mille puhul on dokumendiga 21 U.S.C. 355(a) kooskõla saavutamiseks vajalik ametlik heakskiit ( ametlikult heakskiidetud ravim ); mis aine, mille puhul taotletakse või on taotletud luba kasutamiseks humaanravimina vastavalt 21 U.S.C. 379(g) ( humaanravim"). (Kõik selle mõiste puhul toodud viited seadustikule kehtivad käesoleva taotluse esitamise kuupäeva seisuga). Teisi diabeedivastaseid raviaineid kirjeldatakse käesolevas dokumendis ja need on asjatundjatele tuntud. Leiutisekohaselt kasutatavad diabeedivastased raviained eelistatavalt langetavad HbA1c taset võrreldes baasväärtusega vähemalt % ja leiutisekohaselt kasutatavad diabeedivastased raviained eriti eelistatavalt vähendavad HbAlc taset vähemalt 50% võrreldes baasväärtusega. Diabeedivastaste raviainete hulka kuuluvad insuliini võimendajad, näiteks, kuid mitte ainult, väikesemolekulilised insuliinivõimendajad, tauriin, alfa-lipoehape, mooruspuu ekstrakt, kroom, glutamiin, Enicostemma littorale Blume ekstrakt, Scoparia dulcis ekstrakt, estragonpuju ekstrakt, Andrographis paniculata ekstrakt, isomaltoos, trehaloos või D-mannoos, mis võivad veelgi stimuleerida insuliinieritust või insuliini aktiivsust.
32 Denatureeriv aine või denaturant on käesolevas kontekstis mis tahes ühend või materjal, mis kutsub esile valgu pöörduva lahtivoltimise. Denatureeriva aine või denaturandi tugevus määratakse nii konkreetse denatureeriva aine või denaturandi omaduste kui ka kontsentratsiooni järgi. Sobivad denatureerivad ained või denaturandid võivad olla kaotroobid, detergendid, orgaanilised lahustid, vees segunevad lahustid, fosfolipiidid või kahe või rohkema aine kombinatsioon. Sobivate kaotroopide seas on näiteks, kuid mitte ainult, uurea, guanidiin ja naatriumtiotsüanaat. Kasulikud detergendid võivad olla näiteks, kuid mitte ainult, tugevad detergendid nagu naatriumdodetsüülsulfaat või polüoksüetüleeneetrid (nt Tween või Triton detergendid), sarkosüül, leebed mitteioonsed detergendid (nt digitoniin), leebed katioonsed detergendid nagu N->2,3- (dioleüoksü)-propüül-n,n,n-trimetüülammoonium, leebed ioonsed detergendid (nt naatriumkolaat või naatriumdeoksükolaat) või tsvitterioonsed detergendid, sealhulgas, kuid mitte ainult, sulfobetaiinid ((Zwittergent), 3-(3- koolamidopropüül)dimetüülammonio-1-propaansulfaat (CHAPS) ja 3-(3- chlolamidopropüül)dimetüülammonio-2-hüdroksü-1-propaansulfonaat (CHAPSO). Denaturantidena võib kasutada orgaanilisi vees segunevaid lahusteid nagu atsetonitriil, madalamad alkanoolid (eelkõige C 2 - C 4 alkanoolid nagu etanool või isopropanool) ja madalamad alkaandioolid (eelkõige C 2 - C 4 alkaandioolid nagu etüleenglükool). Käesoleva leiutise korral kasulikud fosfolipiidid võivad olla looduses esinevad fosfolipiidid nagu fosfatidüületanoolamiin, fosfatidüülkoliin, fosfatidüülseriin ja fosfatidüülinositool või sünteetilised fosfoliidi derivaadid või variandid nagu diheksanoüülfosfatidüülkoliin või diheptanoüülfosfatidüülkoliin. Ümbervoltimine on käesolevas kontekstis mis tahes protsess, reaktsioon või meetod, mille abil volditakse valesti kokkuvolditud või kokkuvoltimata, disulfiidsidemeid sisaldavad polüpeptiidid kokku, nii et selle tulemusena on neil disulfiidsidemete suhtes looduslik või õige konformatsioon. Mõiste koos kokkuvoltimine all peetakse käesolevas kontekstis silmas ümbervoltimise protsesse, reaktsioone või meetodeid, mille korral kasutatakse kahte üksteisega interakteeruvat polüpeptiidi ja mille abil volditakse valesti kokkuvolditud või voltimata polüpeptiidid kokku, nii et selle tulemusena on neil looduslik õige konformatsioon.
33 Käesolevas kontekstis hõlmavad mõisted FGF-21 polüpeptiid, fibroblastide kasvufaktor 21 või FGF-21 ja selliste mõistete sidekriipsuta vormid polüpeptiide ja valke, millel on vähemalt üks fibroblastide kasvufaktorile 21 omane bioloogiline aktiivsus/toime, ja ka FGF-21 analooge, FGF-21 isovorme, FGF-21 mimeetikuid, FGF-21 fragmente, hübriidseid FGF-21 valke, nende fusioonvalke, oligomeere ja multimeere, homolooge, glükosüülimismustri põhiseid variante, variante, splaissvarinate ja muteiine sõltumata nende bioloogilisest aktiivsusest ja ka nende sünteesi- või valmistusmeetodist ehk sellest, kas need on saadud näiteks, kuid mitte ainult, rekombinantselt (saadud cdnast, genoomsest DNA-st, sünteetilisest DNA-st või nukleiinhappe muust vormist), in vitro, in vivo, nukleiinhappe molekulide mikroinjektsiooni teel, sünteetilistel, transgeensetel ja geenaktiveeritud meetoditel. Mõisted FGF-21 polüpeptiid ja FGF-21 hõlmavad ühte või mitut aminohappe asendust, aditsiooni või deletsiooni. Asendusi, mida tehakse loodusliku FGF-21 polüpeptiidi paljudes erinevatest aminohapete positsioonides, on kirjeldatud. Asenduste seas on näiteks, kuid mitte ainult, asendused, mis moduleerivad polüpeptiidi farmatseutilist stabiilsust, suurendavad agonistlikku toimet ja vastupidavust proteaasi suhtes, muundavad polüpeptiidi antagonistiks, jne ja mõisted FGF-21 polüpeptiid ja FGF-21 hõlmavad selliseid asendusi. Ilma liiderjärjestuseta FGF-21 osas vt käesolevas dokumendis toodud järjestusi SEQ ID NO: 1, SEQ ID NO: 2 ja SEQ ID NO: 5. Liiderjärjestusega FGF-21 osas vt käesolevas dokumendis toodud järjestusi SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 6 ja SEQ ID NO: 7. Mõnes teostuses on FGF-21 polüpeptiidid üldjoontes identsed järjestustega SEQ ID NOs: 1-7, kuid neil on üks aminohape positsioonis 8 (SEQ ID NO: 1 või vastavad aminohapped järjestuses SEQ ID NOs: 2-7) asendatud mittelooduslikult kodeeritud aminohappega või FGF-21 polüpeptiidi mis tahes teise järjestusega. Lisaks on identifitseeritud FGF-21 mitmed polümorfismid. Leutsiini või proliini kirjeldatakse samas positsioonis USA patendipublikatsioonis nr ja USA patendis nr N-terminaalsed liider- või signaaljärjestused, mis erinevad 1 aminohappe (leutsiini) poolest, on toodud USA patendis nr ja USA patendipublikatsioonis nr Nukleiinhapped, mis kodeerivad FGF-21 ja FGF-21 polüpeptiide, sealhulgas mutante, ja FGF-21 polüpeptiidide ekspressiooni- ja puhastusmeetodid on hästi tuntud ja nende seas on näiteks, kuid mitte ainult, need, mida kirjeldatakse USA patendis nr ; USA patendipublikatsioonides 05/ , 05/ , 04/ , 04/09780,
34 32 02/ , and 01/ ; WO 01/36640; WO 03/011213; WO 03/059270; WO 04/1472; WO 05/061712; WO 05/072769; WO 05/091944; WO 05/113606; WO 06/028595; WO 06/028714; WO 06/050247; WO 06/065582; WO 06/ Mõiste FGF-21 polüpeptiid hõlmab ka farmatseutiliselt sobivaid sooli ja eelravimeid ja soolade eelravimeid, polümorfe, hüdraate, solvate, bioloogiliselt aktiivseid fragmente, loodusliku FGF-21 polüpeptiidi bioloogiliselt aktiivseid fragmente ja ka loodusliku FGF- 21 polüpeptiidi agonistlikke, mimeetilisi ja antagonistlikke variante ja selle polüpeptiidide fusioone. Mõiste FGF-polüpeptiid hõlmab ka fusioone, millel on amino-otsas, karboksüül-otsas või mõlemas otsas täiendavad aminohapped. Selliste fusioonvalkude seas on näiteks, kuid mitte ainult, metionüül-fgf-21, milles metioniin on seotud FGF-21 N-otsaga, mis tuleneb rekombinantselt ekspresseeritavast FGF-21 küpsest vormist, millel puudub liider- või signaaljärjestus või osa sellest (metioniin on seotud rekombinantselt ekspresseeritud FGF- 21 N-otsaga), puhastamiseks ette nähtud fusioone (sealhulgas, kuid mitte ainult, polühistidiini või afiinsusepitoopidega), fusioone seerumi albumiini siduvate valkudega ja fusioone seerumi valkudega nagu seerumi albumiin. USA patendis nr kirjeldatakse meetodit, mille abil saab selekteerida uudseid valke nagu kasvuhormoon ja antikeha fragmendi variandid, millel on muudetud omadused nende retseptori molekulide suhtes. Selle meetodi korral sulandatakse huvipakkuvat geeni kodeeriv valk filamentoosse faagi M13 geeni III kitse valgu karboksü-otsa domeeniga. Nii saab luua kimäärseid molekule, mis sisaldavad FGF-21 ja ühte või mitut teist molekuli, sealhulgas, kuid mitte ainult, keratinotsüütide kasvufaktorit (KGF) (Reich-Slotky, R. et al., J. Biol. Chem. 270: (1995)). Sellised kimäärsed molekulid võivad sisaldada ühe või mõlema FGF-21 ja KGF molekulide spetsiifilisi piirkondi või fragmente. Mis tahes selliseid fragmente saab valkudest valmistada standardsetel biokeemilistel meetoditel või sellist fragmenti kodeeriva polünukleotiidi ekspresseerimise teel. FGF-21 või selle fragmendi saab luua fusioonvalguna, mis sisaldab inimese seerumi albumiini (HAS) või osa sellest. Sellised fusioonkonstruktid sobivad FGF-21 või selle ekspressiooni stimuleerimiseks eukarüootses peremeesrakus. HAS osade seas on N-otsa polüpeptiid (aminohapped 1-369, ja vahepealsed pikkused, mis algavad aminohappega 1), mida kirjeldatakse USA patendis nr ja
35 33 PCT publikatsioonis WO 97/ Teiste kimäärsete polüpeptiidide seas on HAS valk koos FGF-21-ga või selle fragmentidega, mis on kinnitatud HAS C-otsa ja N-otsa külge. Selliseid HSA konstrukte kirjeldatakse USA patendis nr FGF-21 ekspressiooni imetajarakkudes kirjeldatakse patendidokumendis WO 05/ Erinevates viidatud allikates kirjeldatakse polüpeptiidide modifitseerimist polümeeriga konjugeerimise või glükosüülimise teel. Mõiste FGF-21 polüpeptiid hõlmab polüpeptiide, mis on konjugeeritud polümeeriga nagu PEG ja neis võib olla veel üks või mitu tsüsteiini, lüsiini või teiste aminohappejääkide derivatsiooni. FGF-21 polüpeptiid võib lisaks sisaldada linkerit või polümeeri, kusjuures aminohape, millega selline linker või polümeer on konjugeeritud, võib olla käesoleva leiutise kohane aminohape, või võib olla konjugeeritud tehnika tasemes tuntud meetoditel nagu lüsiini või tsüsteiiniga sidumise meetod looduslikult kodeeritud aminohappega. FGF-21 ja teiste polüpeptiidide konjugeerimist polümeeriga on kirjeldatud. Vt nt WO 05/ USA patendis nr kirjeldatakse PEGüleeritud ja eemaldatud lüsiiniga polüpeptiide, millest vähemalt üks lüsiinijääk on eemaldatud või asendatud mis tahes teise aminohappejäägiga. Patendidokumendis WO 99/67291 kirjeldatakse valgu PEG-iga konjugeerimise protsessi, milles korral deleteeritakse vähemalt üks valgus sisalduv aminohappejääk ja valk viiakse valgu konjugeerimiseks sobivates tingimustes kokkupuutesse PEG-iga. Patendidokumendis WO 99/03887 kirjeldatakse kasvuhormoonide ülemperekonda kuuluvate polüpeptiidide PEGüleeritud variante, milles tsüsteiinijääk on asendatud mitteessentsiaalse aminohappega, mis paikneb polüpeptiidi konkreetses piirkonnas. Patendidokumendis WO 00/26354 kirjeldatakse vähendatud allergeensusega glükosüülitud polüpeptiidi variandi valmistusmeetodit, mille abil saadakse polüpeptiid, millel on võrreldes vastava eellaspolüpeptiidiga vähemalt üks täiendav glükosüülimiskoht. USA patendis nr kirjeldatakse granulotsüütide kolooniaid stimuleeriva faktori (G-CSF) ja teiste polüpeptiidide modifitseerimist, mille korral sisestatakse võrreldes loodusliku polüpeptiidiga vähemalt üks täiendav süsivesikahel. Mõiste FGF-21 polüpeptiid hõlmab ka glükosüülitud FGF-21, näiteks, kuid mitte ainult polüpeptiide, mis on mis tahes aminohappe positsioonis glükosüülitud ja polüpeptiidi N- seoselisi või O-seoselisi glükosüülitud vorme. Variante, milles on ühe nukleotiidi muutused, peetakse samuti FGF-21 polüpeptiidi bioloogiliselt aktiivseteks variantideks. Lisaks hõlmab see mõiste ka splaissvariante. Mõiste FGF-21 polüpeptiid alla kuuluvad
36 ka FGF-21 polüpeptiidi heterodimeerid, homodimeerid, ja mis tahes ühe või mitme FGF- 21 polüpeptiidi või mis tahes teise polüpeptiidi, valgu, süsivesiku, polümeeri, väikese molekuli, linkeri, ligandi või mis tahes teist tüüpi bioloogiliselt aktiivse molekuli heteromultimeerid või homomultimeerid, mis on seotud keemiliselt või mida ekspresseeritakse fusioonvalguna, ja ka polüpeptiidi analooge, milles on näiteks spetsiifilised deletsioonid või teised modifikatsioonid, kuid neil on säilinud bioloogiline aktiivsus. Kui ei ole öeldud teisiti (kui näiteks on öeldud, et võrdluse aluseks on järjestus SEQ ID NO: 2, 3, 4, 5, 6, 7 või muu FGF-21 järjestus), viidatakse käesolevas dokumendis FGF-21 aminohappe positsioonidest rääkides järjestuses SEQ ID NO: 1 toodud positsioonidele. Näiteks järjestuses SEQ ID NO: 1 paikneb aminohappe positsioonis 77 arginiin ja järjestuses SEQ ID NO: 2 paikneb vastav arginiin positsioonis 87. Asjatundjad mõistavad, et järjestuse SEQ ID NO: 1 aminohapetele vastavaid aminohapete positsioone saab mis tahes teises FGF-21 molekulis nagu SEQ ID NO: 2, 3, 4, 5, 6 ja 7 kergesti määrata. Asjatundjad mõistavad, et järjestuse SEQ ID NO: 1, 2, 3, 4, 5, 6, 7 või mis tahes muu FGF-21 järjestuse aminohapete positsioonidele vastavaid aminohapete positsioone saab mis tahes teises FGF-21 molekulis nagu FGF-21 fusioonid, variandid, fragmendid, jne kergesti määrata. Näiteks järjestuste reastamise programmidega nagu BLAST saab reastada ja määrata valgus konkreetse positsiooni, mis vastab mingile positsioonile järjestuses SEQ ID NO: 1, 2, 3, 4, 5, 6, 7 või teises FGF-21 järjestuses. Kui käesolevas dokumendis kirjeldatakse aminohapete asendusi, deletsioone või aditsioone järjestustes SEQ ID NO: 1, 2, 3, 4, 5, 6, 7 või mis tahes teises FGF-21 järjestuses, viidatakse ühtlasi ka asendustele, deletsioonidele või aditsioonidele käesolevas dokumendis kirjeldatavate või tehnika tasemes tuntud FGF-21 fusioonide, variantide, fragmentide, jne vastavates positsioonides ja need kuuluvad samuti käesoleva leiutise ulatusse. Mõiste FGF-21 polüpeptiid või FGF-21 hõlmab FGF-21 polüpeptiide, milles on üks või mitu aminohappe asendust, aditsiooni või deletsiooni. Leiutisekohastes FGF-21 polüpeptiidid sisaldavad ühe või mitme loodusliku aminohappe modifikatsioone koos ühe mitteloodusliku aminohappe modifikatsiooniga. On kirjeldatud näiteid asendustest looduslike FGF-21 polüpeptiidide paljudes erinevates aminohappe positsioonides, sealhulgas, kuid mitte ainult asendusi, millega moduleeritakse FGF-21 polüpeptiidi farmatseutilist stabiilsust, millega moduleeritakse FGF-21 polüpeptiidi ühte või mitut bioloogilist aktiivsust, näiteks, kuid mitte ainult, suurendatakse agonistlikku toimet,
37 parandatakse polüpeptiidi lahustuvust, vähendatakse tundlikkust proteaasi suhtes, muundatakse polüpeptiid antagonistiks, jne ja kõik need asendused kuuluvad samuti mõiste FGF-21 polüpeptiid alla. Mõnes teostuses sisaldab FGF-antagonist mittelooduslikult kodeeritud aminohapet, mis on seotud vees lahustuva polümeeriga, mis sisaldub FGF-21 molekuli retseptorit siduvas piirkonnas. Leiutise mõnes teostuses on FGF-21 polüpeptiidides ka aditsioon, asendus või deletsioon, millega moduleeritakse FGF-21 polüpeptiidi bioloogilist aktiivsust. Selliste aditsioonide, asenduste või deletsioonidega saab moduleerida FGF-21 ühte või mitut omadust või aktiivsust. Selliste aditsioonide, asenduste või deletsioonidega näiteks saab moduleerida afiinsust FGF-21 polüpeptiidi retsetori suhtes, poolväärtusaega vereringes, terapeutilist poolväärtusaega, polüpeptiidi stabiilsust, lõhustumist proteaaside toimel, annust, vabanemist või biosaadavust, hõlbustada puhastamist või parandada või muuta konkreetsel viisil manustamist, FGF-21 polüpeptiidid võivad sisaldada ka proteaasiga lõhustamise järjestusi, reaktsioonivõimelisi rühmi antikeha siduvaid domeene (sealhulgas, kuid mitte ainult, FLAG või poly-his) või teisi afiinsusel põhinevaid järjestusi (sealhulgas, kuid mitte ainult, FLAG, poly-his, GST, jne) või seotud molekule (sealhulgas, kuid mitte ainult, biotiini), mis parandavad detekteerimist (sealhulgas, kuid mitte ainult, GF), puhastamist ja polüpeptiidi teisi omadusi. Mõiste FGF-21 polüpeptiid hõlmab ka homodimeere, heterodimeere, homomultimeere ja heteromultimeere, mis on seotud, sealhulgas, kuid mitte ainult, mittelooduslikult kodeeritud aminohappe kõrvalahelate kaudu looduslikult kodeeritud aminohappe kõrvalahelatega või kaudselt linkeri kaudu. Sobivate linkerite seas on näiteks, kuid mitte ainult, väikesed orgaanilised ühendid, erineva pikkusega vees lahustuvad polümeerid nagu erineva pikkusega polü(etüleenglükool) või polüdekstraan või polüpeptiidid. Mittelooduslikult kodeeritud aminohape on aminohape, mis ei kuulu tavapärase aminohappe hulka ega ole pürrolüsiin või selenotsüsteiin. Teised samaväärsed mõisted, mida võib kasutada mõiste mittelooduslikult kodeeritud aminohape sünonüümina, on mittelooduslik aminohape, looduslikult mitteesinev aminohape, looduses mitte esinev aminohape. Mõiste mittelooduslikult kodeeritud aminohape hõlmab ka näiteks, kuid mitte ainult aminohappeid, mis saadakse loodusliku aminohappe (sealhulgas, kuid mitte ainult, tavapärase aminohappe või pürrolüsiini ja selenotsüsteiini)
38 36 modifitseerimise (nt translatsioonijärgse modifitseerimise) tulemusena ja mida translatsioonkompleks ei lisa looduslikult kasvavasse polüpeptiidi ahelasse. Sellised mittelooduslikud aminohapped on näiteks, kuid mitte ainult, N-atsetüülglükosaminüül-Lseriin, N-atsetüülglükosaminüül-L-treoniin ja O-fosfotürosiin. 5 Amino-otsa modifitseerimisrühm on mis tahes molekul, mida saab kinnitada polüpeptiidi amino-otsa külge. Karboksü-otsa modifitseerimisrühm on analoogselt mis tahes molekul, mida saab kinnitada polüpeptiidi karboksü-otsa külge. Otste modifitseerimisrühmade seas on näiteks, kuid mitte ainult, mitmesugused vees lahustuvad polümeerid, peptiidid või valgus nagu seerumi albumiin või teised fragmendid, mis pikendavad peptiidide poolväärtusega seerumis. Mõistetega funktsionaalne rühm, aktiivne fragment, aktiveeriv rühm, lahkuv rühm, reaktsioonivõimeline koht/sait, keemiliselt reaktsioonivõimeline rühm ja keemiliselt reaktsioonivõimeline fragment tähistatakse tehnika tasemes ja ka käesolevas dokumendis molekuli selgesti eristuvaid konkreetseid osi või ühikuid. Need mõisted on keemias osaliselt sünonüümid ja nende all peetakse käesolevas dokumendis silmas molekulide osi, millel on teatud funktsioon või aktiivsus ja mis reageerivad teiste molekulidega. Käesolevas kontekstis peetakse mõiste side või linker all silmas rühmi või sidemeid, mis tavaliselt moodustuvad keemilise reaktsiooni tulemusena ja on üldjuhul kovalentsed sidemed. Hüdrolüütiliselt stabiilsed sidemed on sidemed, mis on vees üldjoontes stabiilsed ja ei reageeri kasulike ph väärtuste korral, näiteks, kuid mitte ainult, füsioloogilistes tingimustes pikema aja jooksul ja võimalik et määramatult pika aja jooksul veega. Hüdrolüütiliselt ebastabiilsed või lagunevad sidemed on sidemed, mis lagunevad vees või vesilahustes, sealhulgas näiteks veres. Ensümaatiliselt ebastabiilse või laguneva/lõhustatava sideme all peetakse silmas sidet, mis võib ühe või mitme ensüümi toimel laguneda. Nagu on tehnika tasemes teada, PEG-il ja selle sarnastel polümeeridel võivad lõhustatavad sidemed paikneda polümeeri selgroos või polümeeri selgroo ja polümeeri molekuli ühe või mitme terminaalse funktsionaalrühma vahel.
39 37 Näiteks estersidemed, mis moodustuvad PEG karboksüülhapete või aktiveeritud PEG karboksüülhapete reageerimisel bioloogiliselt aktiivse aine alkoholrühmadega, füsioloogilistes tingimustes üldjuhul hüdrolüüsuvad ja vabastavad aine Teiste hüdrolüütiliselt lõhustatavate sidemete seas on näiteks, kuid mitte ainult, karbonaatsidemed, amiini ja aldehüüdi vahelise reaktsiooni tulemusel tekkinud imiinsidemed, alkoholi ja fosfaatrühma vahelise reaktsiooni tulemusel tekkinud fosfaatestri sidemed, hüdrasiidi ja aldehüüdi vahelise reaktsiooni tulemusel tekkinud hüdrasoonsidemed; aldehüüdi ja alkoholi vahelise reaktsiooni tulemusel tekkinud atsetaalsidemed; formaadi ja alkoholi vahelise reaktsiooni tulemusena tekkinud atsetaalsidemed; formaadi ja alkoholi vahelise reaktsiooni tulemusel tekkinud ortoestersidemed; aminorühma moodustatud peptiidsidemed, sealhulgas, kuid mitte ainult, polümeeri nagu PEG otsas ja peptiidi karboksüülrühm ajuures; ja fosforamidiitrühma moodustatud oligonukleotiidsidemed, sealhulgas, kuid mitte ainult, polümeeri otsas ja oligonukleotiidi 5' hüdroksüülrühma juures. Bioloogiliselt aktiivne molekul, bioloogiliselt aktiivne fragment või bioloogiliselt aktiivne aine on käesolevas kontekstis mis tahes aine, mis suudab mõjutada organismiga, sealhulgas, kuid mitte ainult, viiruste, bakterite, bakteriofaagi, transposoni, priooni, putukate, seente, taimede, loomade ja inimeste organismiga seotud bioloogilise süsteemi, raja, molekuli või interaktsiooni omadusi. Käesolevas dokumendis peetakse bioloogiliselt aktiivsete molekulide all silmas eelkõige, kuid mitte ainult, mis tahes ainet, mis on ette nähtud inimestel ja teistel loomadel haiguse diagnoosimiseks, väljaravimiseks, leevendamiseks või ennetamiseks või inimeste või loomade füüsilise või vaimse tervise muul viisil parandamiseks. Bioloogiliselt aktiivsed molekulid on näiteks, kuid mitte ainult, peptiidid, valgud, ensüümid, väikesemolekulilised ravimid, vaktsiinid, immunogeenid, tugevad ravimid, pehmed ravimid, süsivesikud, anorgaanilised aatomid ja molekulid, värvained, lipiidid, nukleosiidid, radionukliidid, oligonukleotiidid, toksoidid, toksiinid, prokarüootsed ja eukarüootsed rakud, viirused, polüsahhariidid, nukleiinhapped ja nende osad, mis on saadud või deriveeritud viirustest, bakteritest, putukatest, loomarakkudest või mis tahes teistest rakkudest või rakutüübist, liposoomid, mikroosakesed ja mitsellid. Käesoleva leiutise puhul sobivate bioloogiliselt aktiivsete ainete klasside seas on näiteks, kuid mitte ainult, ravimid, eelravimid, radionukliidid, pildindusained, polümeerid, antibiootikumid, fungitsiidid, viirusevastased ained, põletikuvastased ained,
40 38 kasvajavastased ained, südameveresoonkonna ained, ärevusvastased ained, hormoonid, kasvufaktorid, steroidsed ained, mikroobidest deriveeritud toksiinid ja teised sarnased ained. 5 Bifunktsionaalne polümeer on polümeer, mis sisaldab kahte eraldiseisvat funktsionaalrühma, millel on võime reageerida spetsiifiliselt teiste fragmentidega (sealhulgas, kuid mitte ainult, aminohapete kõrvalrühmadega) ja moodustada kovalentseid või mittekovalentseid sidemeid. Bifunktsionaalse linkeriga, millel on üks funktsionaalne rühm, mis reageerib konkreetse bioloogiliselt aktiivse koostisosa rühmaga, ja teine rühm, mis reageerib teise bioloogiliselt aktiivse koostisosa rühmaga, saab moodustada konjugaadi, mis sisaldab esimest bioloogiliselt aktiivset koostisosa, bifunktsionaalset linkerit ja teist bioloogiliselt aktiivset koostisosa. Paljud protseduurid ja linkermolekulid, mida kasutatakse erinevate ühendite kinnitamiseks peptiidide külge, on tuntud. Vt nt Euroopa patendipublikatsiooni nr 188,6; USA patente nr , , , , ja Multifunktsionaalne polümeer on polümeer, mis sisaldab kahte või rohkemat eraldiseisvat funktsionaalrühma, millel on võime reageerida spetsiifiliselt teiste fragmentidega (sealhulgas, kuid mitte ainult, aminohapete kõrvalrühmadega) ja moodustada kovalentseid või mittekovalentseid sidemeid. Bifunktsionaalne või multifunktsionaalne polümeer võib olla mis tahes soovitud pikkuse või molekulmassiga ja selle võib valida nii, et FGF-21ga seotud ühe või mitme molekuli ja selle retseptori või FGF-21 vahel oleks soovitud vahemaa või konformatsioon. Kui asendusrühmad on tähistatud nende tavapäraste keemiliste valemitega, mis on toodud vasakult paremale, hõlmavad nad ka keemiliselt identseid asendajaid, mille puhul on valem esitatud suunaga paremalt vasakule ehk näiteks struktuur -CH 2 O- on samaväärne struktuuriga -OCH 2 -. Mõiste asendajad hõlmab ka näiteks, kuid mitte ainult, mittehäirivaid asendajaid. Mittehäirivad asendajad on sellised asendajad, mille kasutamisel saadakse stabiilsed ühendid. Sobivate mittehäirivate asendajate või radikaalide seas on näiteks, kuid mitte 30 ainult, halo-, C 1 -C alküül-, C 2 -C alkenüül-, C 2 -C alkünüül-, C 1 -C alkoksü-, C 1 -C 12 aralküül-, C 1 -C 12 alkarüül-, C 3 -C 12 tsükloalküül-, C 3 -C 12 tsükloalkenüül-, fenüül-, asendatud fenüül-, toluoüül-, ksülenüül-, bifenüül-, C 2 -C 12 alkoksüalküül-, C 2 -C 12
41 39 5 alkoksüarüül-, C 7 -C 12 arüüloksüalküül-, C 7 -C 12 oksüarüül-, C 1 -C 6 alküülsulfinüül-, C 1 -C alküülsulfonüül-, --(CH 2 ) m --O--(C 1 -C alküül), milles m on 1 kuni 8, arüül-, asendatud arüül-, asendatud alkoksü-, fluoroalküül-, heterotsükliline radikaal, asendatud heterotsükliline radikaal, nitroalküül-,--no 2, --CN, --NRC(O)--(C 1 -C alküül), --C(O)-- (C 1 -C alküül), C 2 -C alküül tioalküül, --C(O)O--( C 1 -C alküül), --OH, --SO 2, =S, --COOH, --NR 2, karbonüül, --C(O)--(C 1 -C alküül)-cf3,-c(o)-- CF3, --C(O)NR2, --(C 1 -C arüül)-s--(c 6 -C arüül), --C(O)--(C 1 -C arüül), --(CH 2 ) m -- O-(--(CH 2 ) m --O--(C 1 -C alküül)fragment, milles iga m on 1 kuni 8, --C(O)NR 2, --C(S)NR 2, -- SO 2 NR 2, --NRC(O) NR 2, --NRC(S) NR 2, nende soolad ja muud sarnased üksused. Iga R on selles käesolevas kontekstis H, alküül- või asendatud alküül-, arüül- või asendatud arüül-, aralküül- või alkarüülrühm. Mõiste halogeen all peetakse silmas fluori, kloori, joodi ja broomi. Kui ei ole öeldud teisiti, on mõiste alküül- omaette või teise asendaja osana sirge või hargnenud ahelaga või tsükliline süsivesinikradikaal või nende kombinatsioon, mis võib olla täielikult küllastunud, mono- või polüküllastumata ja võib sisaldada di- ja multivalentseid radikaale, milles on näidatud arv süsiniku aatomeid (ehk C 1 -C tähendab üks kuni kümme süsinikuaatomit). Küllastunud süsivesinikradikaalid on näiteks, kuid mitte ainult, metüül-, etüül-, n-propüül-, isopropüül-, n-butüül-, t-butüül-, isobutüül-, secbutüül-, tsükloheksüül-, (tsükloheksüül)metüül-, tsüklopropüülmetüül-, näiteks n-pentüül-, n-heksüül-, n-heptüül-, n-oktüülrühma homoloogid ja isomeerid ja teised sarnased radikaalid. Küllastamata alküülrühm on alküülrühm, milles on üks või mitu kaksik- või kolmiksidet. Küllastamata alküülrühmade seas on näiteks, kuid mitte ainult, vinüül-, 2-propenüül-, krotüül-, 2-isopentenüül-, 2-(butadienüül)-, 2,4-pentadienüül-, 3-(1,4-pentadienüül)-, etünüül-, 1- ja 3-propünüül-, 3-butünüülrühm ja kõrgemad homoloogid ja isomeerid. Kui ei ole öeldud teisiti, hõlmab mõiste alküülrühm ka alküülfragmendi derivaate, mida määratletakse täpsemalt allpool, nagu heteroalküülrühm. Alküülrühmi, mis on piiratud süsivesinikrühmadega, nimetatakse homoalküülrühmadeks.
42 40 Mõiste alküleen- on omaette või teise asendaja osana alkaanist deriveeritud divalentne radikaal ja nende seas on näiteks, kuid mitte ainult struktuurid nagu -CH 2 CH 2 - ja --CH 2 CH 2 CH 2 CH 2 - ning ka allpool kirjeldatavad heteroalküleenrühmad. 5 Alküül-(või alküleen)rühmas on üldjuhul 1 kuni 24 süsinikuaatomit, kusjuures käesolevas dokumendis kirjeldavate meetodite ja kompositsioonide puhul on üheks konkreetseks teostuseks rühmad, milles on või vähem süsinikuaatomit. Madalam alküülrühm või madalam alküleenrühm on lühema ahelaga alküül- või alküleenrühm, milles on üldjuhul 8 või vähem süsinikuaatomit. 30 Mõisteid alkoksü-, alküülamino- and "alküültiorühm (või tioalkoksürühm ) kasutatakse nende tavapärases tähenduses ja nende all peetakse silmas alküülrühmi, mis kinnituvad ülejäänud molekuli külge vastavalt hapnikuaatomi, aminorühma või väävliaatomi kaudu. Kui ei ole öeldud teisiti, on heteroarüülrühm nii omaette kui ka teise mõiste osana stabiilne sirge või hargnenud ahel või tsükliline süsivesinikradikaal või nende kombinatsioon, mis koosneb näidatud arvust süsinikuaatomitest ja vähemalt ühest heteroatomist, mis on valitud rühmast, kuhu kuuluvad O, N, Si ja S, ja milles lämmastikuja väävliaatomid võivad valikuliselt olla oksüdeeritud ja lämmastiku heteroaatom võib olla valikuliselt kvaterneeritud. Heteroaatom(id) O, N ja S ja Si võivad paikneda heteroarüülrühma mis tahes sisemises positsioonis või positsioonis, milles alküülrühm kinnitub ülejäänud molekuli külge. Asjakohased rühmad on näiteks, kuid mitte ainult, -CH 2 -CH 2 -O-CH 3, -CH 2 -CH 2 -NH-CH 3,-CH 2 -CH 2 -N(CH 3 )-CH 3, -CH 2 -S-CH 2 -CH 3, -CH 2 -CH 2,-S(O)-CH 3, -CH 2 -CH 2 -S(O) 2 -CH 3,-CH=H-O-CH 3, -Si(CH 3 ) 3, -CH 2 -CH=N- OCH 3 ja -CH=CH-N(CH 3 )-CH 3. Kuni kaks heteroaatomit võivad paikneda järjestikku nagu näiteks -CH 2 -NH-OCH 3 ja -CH2-O-Si(CH 3 ) 3. Heteroalküleenrühm on analoogselt kas omaette või osana teisest asendajast divalentne radikaal, mis on deriveeritud heteroalküülrühmast, näiteks, kuid mitte ainult, -CH 2 -CH 2 -S- CH 2 -CH 2 - ja -CH 2 -S-CH 2 -CH 2 -NH-CH 2 -. Heteroalküleenrühmades võivad ahela ühes või mõlemas otsas olla samasugused või erinevad heteroaatomid (sealhulgas, kuid mittes ainult, alküleenoksü-, alküleendioksü-, alküleenamino-, alküleendiamino-, aminooksüalküleen- ja teised sarnased üksused). Alküleen- ja heteroalküleen-
43 5 41 sidumisrühmade puhul ei näita sidumisrühma valemi esitussuund sidumisrühma orientatsiooni. Näiteks valemiga C(O) 2 R'- tähistatakse nii -C(O) 2 R'- ja -R'C(O) 2 - rühma. Kui ei ole öeldud teisiti, on mõisted tsükloalküül- ja heterotsükloalküül- omaette või kombinatsioonis teiste mõistetega vastavalt alküülrühma ja heteroalküülrühma tsüklilised versioonid. Tsükloalküül- ja heterotsükloalküülrühmad sisaldavad seega küllastunud, osaliselt küllastumata ja täielikult küllastumata tsükli sidemeid. Heterotsükloalküülrühmades võib heteroaatom paikneda positsioonis, milles heterotsükkel kinnitub ülejäänud molekuli külge. Tsükloalküülrühmad on näiteks, kuid mitte ainult, tsüklopentüül-, tsükloheksüül-, 1- tsükloheksenüül-, 3-tsükloheksenüül-, tsükloheptüül- ja teised sarnased rühmad. Heterotsükloalküülrühmade seas on näiteks, kuid mitte ainult, 1-(1,2,5,6- tetrahüdropüridüül-), 1-piperidinüül-, 2-piperidinüül-, 3-piperidinüül-, 4-morfolinüül-, 3- morfolinüül-, tetrahüdrofuraan-2-üül-, tetrahüdrofuraan-3-üül-, tetrahüdrotieen-2-üül-, tetrahüdrotieen-3-üül-, 1-piperasinüül-, 2-piperasinüülrühm ja teised sarnased rühmad. See mõiste hõlmab lisaks ka kahetsüklilisi ja kolmetsüklilisi süsteeme. Heterotsükloalküleen on analoogselt kas omaette või teise asendaja osana divalentne radikaal, mis on deriveeritud heterotsükloalküülrühmast, ja tsükloalküleen on omaette või osana teisest asendajast divalentne radikaal, mis on deriveeritud tsükloalküülrühmast. 30 Käesolevas kontekstis on vees lahustuv polümeer mis tahes polümeer, mis lahustub veepõhises lahustis. FGF-21 polüpeptiidide sidumisel vees lahustuvate polümeeridega saab kutsuda esile teatud muutusi, näiteks, kuid mitte ainult, pikendada või moduleerida seerumi poolväärtusaega, pikendada või moduleerida terapeutilist poolväärtusega võrreldes modifitseerimata vormiga, moduleerida füüsikalise seotuse omadusi nagu agregeerumist ja multimeeride moodustumist, muuta retseptoriga seondumist, muuta seondumist ühe või mitme seondumispartneriga ja muuta retseptori dimerisatsiooni ja multimerisatsiooni. Vees lahustuv polümeer võib olla või mitte olla bioloogiliselt aktiivne ja seda võib kasutada linkerina FGF-21 kinnitamisel teiste ainete, näiteks, kuid mitte ainult, ühe või mitme FGF-21 polüpeptiidi või ühe või mitme bioloogiliselt aktiivse molekuli külge. Sobivate polümeerides seas on näiteks, kuid mitte ainult,
44 42 polüetüleenglükool, polüetüleenglükoolpropioaldehüüd, selle mono-c1-c alkoksü- või arüüloksüderivaadid (mida kirjeldatakse USA patendis nr ), monometoksü-polüetüleenglükool, polüvinüülpürrolidoon, polüvinüülalkohol, polüaminohapped, divinüüleetri maleiinhappe anhüdriid, N-(2-hüdroksüpropüül)- methakrüülamiid, dekstraan, dekstraani derivaadid, sealhulgas dekstraansulfaat, polüpropüleenglükool, polüpropüleenoksiidi/etüleenoksiidi kopolümeer, polüoksüetüülitud polüool, hepariin, hepariini fragmendid, polüsahhariidid, oligosahhariidid, glükaanid, tselluloos ja tselluloosi derivaadid, sealhulgas, kuid mitte ainult, metüültselluloos ja karboksümetüültselluloos, tärklis ja tärklise derivaadid, polüpeptiidid, polüalküleenglükool ja selle derivaadid, polüalküleenglükoolide kopolümeerid ja nende derivaadid, polüvinüületüüleetrid ja alfa-beeta-polü [(2- hüdroksüetüül)-dl-aspartamiid ja teised sarnased ühendid või nende segud. Selliste vees lahustuvate polüpeptiidide seas on näiteks, kuid mitte ainult polüetüleenglükool ja seerumi albumiin. Käesolevas kontekstis on polüalküleenglükool või polü(alkeenglükool) polüetüleenglükool (polü(etüleenglükool)), polüpropüleenglükool, polübutüleenglükool ja nende derivaadid. Mõiste polüalküleenglükool all peetakse silmas nii lineaarseid kui ka hargnenud polümeere keskmise molekulmassiga 0,1 kda kuni 0 kda. Teised näited on loetletud näiteks kaubanduslikes kataloogides nagu Shearwater Corporation' kataloog Polyethylene Glycol and Derivatives for Biomedical Applications" (01). Kui ei ole öeldud teisiti, on arüülrühm polüküllastumata aromaatne süsivesinikasendaja, mis võib koosneda ühest või mitmest tsüklist (näiteks, kuid mitte ainult, 1 kuni 3 tsüklist), mis on kokku sulandatud või seotud kovalentselt). Heteroarüülrühmad on arüülrühmad (või tsüklid), mis sisaldavad ühte kuni nelja heteroaatomit, mis on valitud N, O ja S seast, kusjuures lämmastiku- ja väävliaatomid valikuliselt oksüdeeritakse ja lämmastikuaatom(id) valikuliselt kvaterneeritakse. Heteroarüülrühma saab kinnitada ülejäänud molekuli külge heteroaatomi kaudu. Arüül- ja heteroarüülrühmad on näiteks, kuid mitte ainult, fenüül-, 1-naftüül-, 2-naftüül-, 4-bifenüül-, 1-pürrolüül-, 2-pürrolüül-, 3- pürrolüül-, 3-pürasolüül, 2-imidasolüül-, 4-imidasolüül-, pürasinüül-, 2-oksasolüül-, 4- oksasolüül-, 2-fenüül-4-oksasolüül-, 5-oksasolüül-, 3-isoksasolüül-, 4-isoksasolüül-, 5-
45 isoksasolüül-, 2-tiasolüül-, 4-tiasolüül-, 5-tiasolüül-, 2-furüül-, 3-furüül-, 2-tienüül-, 3- tienüül-, 2-püridüül-, 3-püridüül-, 4-püridüül-, 2-pürimidüül-, 4-pürimidüül-, 5- bensotiasolüül-, purinüül-, 2-bensimidasolüül-, 5-indolüül-, 1-isokvinolüül-, 5- isokvinolüül-, 2-kvinoksalinüül-, 5-kvinoksalinüül-, 3-kvinolüül-, ja 6-kvinolüülrühm. Iga eespool nimetatud arüül- ja heteroarüültsüklite süsteemide asendajad valitakse allpool toodud sobivate asendajate seast. Lühiduse mõttes hõlmab mõiste arüülrühm, kui seda kasutatakse kombinatsioonis teiste mõistetega (sealhulgas, kuid mitte ainult, arüüloksü-, arüültioksü-, arüülalküülrühm), nii eespool määratletud arüül- kui ka heteroarüülrühmi. Mõiste arüülalküülrühm hõlmab seega ka radikaale, milles arüülrühm kinnitub alküülrühma külge (sealhulgas, nesnüül-, fenetüül-, püridüülmetüülrühm ja teised sarnased rühmad), sealhulgas selliste alküülrühmade külge, milles süsinikuaatom (näiteks, kuid mitte ainult metüleenrühm) on asendatud näiteks hapnikuaatomiga (sealhulgas, kuid mitte ainult, fenoksümetüül-, 2- püridüüloksümetüül-, 3-(1-naftüüloksü)propüülrühm ja teised sarnased rühmad). Kõik eespool toodud mõisted (sealhulgas, kuid mitte ainult, alküül-, heteroalküül-, arüül- ja heteroarüülrühm ) hõlmavad nii näidatud radikaali asendatud kui ka asendamata vorme. Iga radikaali tüübi asendajate näited on toodud allpool. Alküül- ja heteroalküülradikaalide asendajad (sealhulgas rühmad, mida sageli nimetatakse alküleen-, alkenüül-, heteroalküleen-, heteroalkenüül-, alkünüül, tsükloalküül, heterotsükloalküül-, tsükloalkenüül- ja heterotsükloalkenüülrühmadeks) võivad olla üks või mitu rühma, mis on valitud näiteks, kuid mitte ainult, järgmiste üksuste seast: -OR', =O, =NR', =N- OR', -NR'R", -SR', -halogeen, -SiR'R"R"', -OC(O)R', -C(O)R', -CO 2 R', -CONR'R", -OC(O)NR'R", -NR"C(O)R', -NR'-C(O)NR"R"', -NR"C(O) 2 R', -NR-C(NR'R"R"')=NR"", -NR-C(NR'R")=NR"', -S(O)R', -S(O) 2 R', -S(O) 2 NR'R", -NRSO 2 R', -CN ja -NO 2, mille arv võib olla null kuni (2m'+1), milles m on süsinikuaatomite üldarv sellises radikaalis. R', R", R"' ja R"" on igaüks sõltumatult vesinik, asendatud või asendamata heteroalküülrühm, asendatud või asendamata arüülrühm, sealhulgas, kuid mitte ainult, arüülrühm, mis on asendatud 1-3 halogeeniga, asendatud või asendamata alküül-, alkoksüvõi tioalkoksürühmad või arüülalküülrühmad. Kui leiutisekohases ühendis on näiteks rohkem kui üks R rühm, valitakse iga R rühm sõltumatult, ja kui leiutisekohases ühendis on rohkem kui üks R', R", R'" ja R'"' rühm, valitakse ka need rühmad sõltumatult. Kui R'
46 44 ja R" kinnituvad sama lämmastikuaatomi külge, võivad nad moodustada koos lämmastikuaatomiga 5-, 6- või 7-lülilise tsükli Näiteks -NR'R" hõlmab muuhulgas, kuid mitte ainult, 1-pürrolidinüül- ja 4- morfolinüülrühma. Asjatundjad mõistavad eespool käsitletud asendajate põhjal, et mõiste alküülrühm hõlmab ka rühmi, milles olevad süsinikuaatomid on seotud rühmadega, mis ei ole vesinikurühmad, nagu haloalküülrühm (näiteks, kuid mitte ainult, -CF 3 ja -CH 2 CF 3 ) ja atsüülrühm (näiteks, kuid mitte ainult, -C(O)CH 3, -C(O)CF 3, - C(O)CH 2 OCH 3 ja teised sarnased rühmad). Sarnaselt alküülradikaaliga seoses kirjeldatud rühmadele võib arüül- ja heteroarüülrühmadel olla erinevaid asendajaid ja need valitakse näiteks, kuid mitte ainult, rühmast, kuhu kuuluvad: halogeen, -OR', =O, =NR', =N-OR', -NR'R", -SR', -halogeen, -SiR'R"R"', -OC(O)R', -C(O)R', -CO 2 R',-CONR'R", -OC(O)NR'R", -NR"C(O)R', -NR'-C(O)NR"R"', -NR"C(O) 2 R', -NR-C(NR'R"R'")=NR"", -NR- C(NR'R")=NR'", -S(O)R', -S(O) 2 R', -S(O) 2 NR'R", -NRSO 2 R', -CN ja -NO 2, -R', -N 3, -CH(Ph) 2, fluoro(c 1 -C 4 )alkoksü- ja fluoro(c 1 -C 4 )alküülrühm ja nende arv võib olla vahemikus null kuni aromaatses tsükkelsüsteemis olevate vabade valentside üldarv; ja milles R', R", R"' ja R"" on sõltumatult asendatud asendajaga, mis on valitud vesiniku, alküül-, heteroalküül-, arüül- ja heteroarüülrühma seast. Kui leiutisekohases ühendis on rohkem kui üks R rühm, valitakse iga R rühm sõltumatult, ja sama tehakse ka siis, kui ühend sisaldab rohkem kui ühte R', R", R'" ja R"" rühma. Käesolevas kontekstis viitab mõiste moduleeritud seerumi poolväärtusaeg positiivsele või negatiivsele muutusele modifitseeritud FGF-21 poolväärtusajas vereringes võrreldes selle modifitseerimata vormiga. Seerumi poolväärtusaja mõõtmiseks võetakse katsealustelt erinevatel ajahetkedel pärast FGF-21 manustamist vereproovid ja määratakse igas vereproovis vaatlusaluse molekuli kontsentratsioon. Seerumikontsentratsiooni ja aja korrelatsiooni põhjal saab arvutada seerumi poolväärtusaja. Seerumi poolväärtusaeg pikeneb eelistatavalt vähemalt kaks korda, kuid kasu võib olla ka väiksemast pikenemisest, näiteks siis, kui see võimaldab kasutada rahuldavat manustamisrežiimi ja sellega ei kaasne toksilist toimet. Mõnes teostuses pikeneb poolväärtusaeg vähemalt umbes kolm korda, vähemalt umbes viis korda või vähemalt umbes kümme korda.
47 Käesolevas kontekstis viitab mõiste moduleeritud terapeutiline poolväärtusaeg positiivsele või negatiivsele muutusele FGF-21 terapeutiliselt efektiivse koguse poolväärtusajas võrreldes selle modifitseerimata vormiga. Terapeutilist poolväärtusega mõõdetakse vaatlusaluse molekuli farmakokineetiliste ja/või farmakodünaamiliste omaduste mõõtmise teel erinevatel ajahetkedel pärast manustamist. Pikem terapeutiline poolväärtusaeg võimaldab rakendada kasulikku manustamisrežiimi, konkreetset kasulikku üldannust või hoiab ära soovimatu toime. Mõnes teostuses tuleneb pikem terapeutiline poolväärtusaeg suuremast mõjususest, modifitseeritud molekuli suuremast või väiksemast võimest seonduda oma sihtmärgiga, molekulide suurenenud või vähenenud lagunemisest ensüümide nagu proteaasid toimel või modifitseerimata molekuli teise parameetri või toimemehhanismi suurenemisest või vähenemisest või molekuli retseptori vahendatud kliirensi aeglustumisest. Isoleeritud tähendab nukleiinhappe või valgu puhul seda, et see nukleiinhape või valk ei sisalda vähemalt osasid raku koostisosi, millega see on looduslikus olekus seotud, või seda, et nukleiinhapet või valku on kontsentreeritud võrreldes selle kontsentratsiooniga in vivo või in vitro tootmisel. See võib olla homogeenne. Isoleeritud ained võivad olla kuivad või poolkuivad või lahused, sealhulgas, kuid mitte ainult, vesilahused. See võib olla osa farmatseutilisest kompositsioonist, mis sisaldab ka farmatseutiliselt sobivaid tugiaineid ja/või abiaineid. Puhtuse ja homogeensuse määramiseks kasutatakse tavaliselt analüütilise keemia meetodeid nagu polüakrüülamiidgeel-elektroforees või kõrgefektiivne vedelikkromatograafia. Valk, mis moodustab valdava osa preparaadist, on üldjoontes puhastatud. Isoleeritud geen eraldatakse avatud lugemisraamidest, mis paiknevad geeni külgedel ja kodeerivad huvipakkuvast geenist erinevat geeni. Puhastatud on selline nukleiinhape või valk, mille korral tekib elektroforeetilises geelis põhimõtteliselt üks bänd. See võib eeskätt tähendada seda, et nukleiinhappe või valgu puhtusaste on vähemalt 85%, vähemalt 90%, vähemalt 95%, vähemalt 99% või suurem. Mõiste nukleiinhapped all peetakse silmas deoksüribonukleotiide, deoksüribonukleosiide, ribonukleosiide või ribonukleotiide ja nende polümeere, mis on ühe- või kaheahelalised. Kui ei ole öeldud teisiti, hõlmab see mõiste ka nukleiinhappeid, mis sisaldavad looduslike nukleotiidide tuntud analooge, mis sarnanevad omaduste
48 poolest referents-nukleiinhappele ja mida metaboliseeritakse sarnaselt looduslikele nukleotiididele. Kui ei ole öeldud teisiti, hõlmab see mõiste ka oligonukleotiidide analooge, sealhulgas PNA-d (peptidonukleiinhapet), DNA analooge, mida kasutatakse antisenss-tehnoloogias (fosforotioaadid, fosforoamidaadid ja teised sarnased ühendid). Kui ei ole öeldud teisiti, hõlmab konkreetne nukleiinhappeline järjestus olemuslikult ka selle konservatiivselt modifitseeritud variante (sealhulgas, kuid mitte ainult, kõdunud koodoni asendusi) ja komplementaarseid järjestusi ja ka selgelt näidatud järjestusi. Kõdunud koodoni asenduste tegemiseks võib luua järjestused, milles ühe või mitme (või kõigi) valitud koodonite kolmas positsioon on asendatud segatud alusega ja/või deoksüionosiinijääkidega (Batzer et al., Nucleic Acid Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260: (1985); Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)). Mõisteid polüpeptiid, peptiid ja valk kasutatakse käesolevas dokumendis sünonüümidena aminohappejääkidest koosneva polümeeri tähenduses ehk polümeeriga seotud kirjeldus kehtib ühtlasi ka peptiidi ja valgu puhul ning vastupidi. Nende mõistete puhul peetakse silmas nii looduses esinevate aminohapete polümeere kui ka aminohapete polümeere, milles üks või mitu aminohappejääki on mittelooduslikud. Käesolevas kontekstis hõlmavad need mõisted mis tahes pikkusega aminohappeahelaid, sealhulgas täispikki valke, milles aminohappejäägid on seotud kovalentsete peptiidsidemetega. Mõiste aminohape all peetakse silmas nii looduslikke kui ka mittelooduslikke aminohappeid ja ka aminohapete analooge ja mimeetikuid, mis toimivad looduslikele aminohapetele sarnasel viisil. Looduslikult kodeeritud aminohapped on tavapärast aminohapet (alaniin, arginiin, asparagiin, asparagiinhape, tsüsteiin, glutamiin, glutamiinhape, glütsiin, histidiin, isoleutsiin, leutsiin, lüsiin, metioniin, fenüülalaniin, proliin, seriin, treoniin, trüptofaan, türosiin ja valiin) ja pürrolüsiin ja selenotsüsteiin. Aminohapete analoogide all peetakse silmas ühendeid, millel on sama keemiline struktuur mis looduslikul aminohappel ehk see sisaldab vesinikuga seotud süsinikku, karboksüülrühma, aminorühma ja R rühma nagu homoseriin, norleutsiin, metioniinsulfoksiid, metioniinmetüülsulfoonium. Sellistel analoogidel on modifitseeritud R rühmad (nagu norleutsiin) või modifitseeritud peptiidselgrood, kuid neil on sama keemiline põhistruktuur mis looduslikul aminohappel. Aminohapetest rääkides peetakse silmas ka näiteks looduslikke proteogeenseid L-aminohappeid, D-aminohappeid, keemiliselt modifitseeritud aminohappeid nagu aminohapete variandid ja derivaadid,
49 looduslikud mitteproteogeensed aminohapped nagu β-alaniin, ornitiin, jne ja keemiliselt sünteesitud ühendeid, millel on tehnika tasemes aminohapetega seostatavad omadused. Mittelooduslike aminohapete seas on näiteks, kuid mitte ainult, α-metüülaminohapped (nt α-metüülalaniin), D-aminohapped, histidiinilaadsed aminohapped (nt 2-aminohistidiin, β- hüdroksühistidiin, homohistidiin, α-fluorometüülhistidiin ja α-metüülhistidiin), aminohapped, millel on kõrvalahelas täiendav metüleenrühm ( homo - aminohapped), ja aminohapped, milles on karboksüülhappe funktsionaalrühm asendatud sulfoonhapperühmaga (nt tsüsteiinhappega). Käesolevas tekstis võidakse aminohappeid tähistada nende üldtuntud kolmetäheliste tähiste või IUPAC-IUB Biochemical Nomenclature Commission soovitatavate ühetäheliste tähistega. Nukleotiidide puhul kasutatakse samuti nende üldtunnustatud ühetähelisi koode. Mõiste konservatiivselt modifitseeritud variandid all peetakse silmas nii aminohappelisi kui ka nukleiinhappelisi järjestusi. Konkreetsete nukleiinhappeliste järjestuste korral peetakse mõiste konservatiivselt modifitseeritud variandid all silmas nukleiinhappeid, mis kodeerivad identseid või üldjoontes identseid aminohappelisi järjestusi, või kui nukleiinhape ei kodeeri aminohappelist järjestust, üldjoontes identseid järjestusi. Geneetilise koodi kõdumise tõttu kodeerivad suur hulk funktsionaalselt identseid nukleiinhappeid mis tahes konkreetset valku. Näiteks kõik koodonid GCA, GCC, GCG ja GCU kodeerivad aminohapet alaniin. Seega saab igas positsioonid, kus koodon määrab alaniini, muuta koodoni mis tahes vastavaks kirjeldatavaks koodoniks, muutmata sealjuures kodeeritud koodonit. Selliseid nukleiinhapete variatsioone nimetatakse vaikseteks variatsioonideks, mis moodustavad konservatiivselt modifitseeritud variatsioonide ühe tüübi. Iga käesolevas dokumendis toodud nukleiinhappe järjestusega, mis kodeerib polüpeptiidi, kirjeldatakse ka nukleiinhappe võimalikku vaikset variatsiooni. Asjatundjad teavad, et nukleiinhappes sisalduvat igat koodonit (välja arvatud AUG, mis on tavaliselt ainus metioniini koodon, ja TGG, mis on tavaliselt ainus trüptofaani koodon) saab modifitseerida, nii et saadakse funktsionaalselt identne molekul. Iga järjestus sisaldab seetõttu olemuslikult polüpeptiidi kodeeriva nukleiinhappe igat vaikset variatsiooni. Seoses aminohappeliste järjestustega asjatundjad teavad, et nukleiinhappe, peptiidi, polüpeptiidi või valgu järjestuse asendused, deletsioonid või aditsioonid, millega
50 5 48 muudetakse, lisatakse või deleteeritakse kodeeritud järjestuses üks aminohape või väike protsent aminohappeid, on konservatiivselt modifitseeritud variant, mille korral muutmise tulemusena deleteeritakse üks aminohape, lisatakse üks aminohape või asendatakse üks aminohape keemiliselt sarnase aminohappega. Konservatiivsete asenduste tabelid, mille abil on võimalik saada funktsionaalselt sarnaseid aminohappeid, on asjatundjatele tuntud. Sellised konservatiivselt modifitseeritud variandid on täienduseks leiutisekohastele polümorfsetele variantidele, liikidevahelistele homoloogidele ja alleelidele ja ei välista neid. Konservatiivsete asenduste tabelid, mille abil on võimalik saada funktsionaalselt sarnaseid aminohappeid, on asjatundjatele tuntud. Järgnevas kaheksas rühmas on aminohapped, millega saab üksteist konservatiivselt asendada 1) Alaniin (A), glütsiin (G); 2) Aspartic hape (D), glutamiinhape (E); 3) Asparagiin (N), glutamiin (Q); 4) Arginiin (R), lüsiin (K); 5) Isoleutsiin (I), leutsiin (L), metioniin (M), valiin (V); 6) Fenüülalaniin (F), türosiin (Y), trüptofaan (W); 7) Seriin (S), treoniin (T); ja 8) Tsüsteiin (C), metioniin (M) (vt nt Creighton, Proteins: Structures and Molecular Properties (W H Freeman & Co.; 2nd edition (December 1993) Mõistete identne või identsuse protsent peetakse kahe või rohkema nukleiinhappelise või polüpeptiidi järjestuse puhul silmas kahte või rohkemat järjestust või alamjärjestust, mis on samasugused. Järjestused on üldjoontes identsed, kuid neis sisaldub teatud hulk samasuguseid aminohappejääke või nukleotiide (ehk nende identsus on teatud kindlas piirkonnas umbes 60%, umbes 65%, umbes 70%, umbes 75%, umbes 80%, umbes 85%, umbes 90% või umbes 95%), kui võrrelda ja reastada neid maksimaalse vastavuse määramiseks võrdlusaknas, või mõõta teatud kindlat piirkonda ühe järgmise järjestuste
51 võrdlemise algoritmiga (või teiste asjatundjatele kättesaadavate algoritmidega) või käsitsi reastamise ja visuaalse vaatlemise teel. Selle määratluse all peetakse silmas ka testitava järjestuse komplementi. Identsus võib esineda vähemalt umbes 50 aminohappe või nukleotiidi pikkuse piirkonda ulatuses või 75 kuni 0 aminohappe või nukleotiidi pikkuse piirkonna ulatuses või määramata piirkondade korral terve polünukleotiidi või polüpeptiidi ulatuses. Leiutisekohast polüpeptiidi kodeeriva polünukleotiidi, sealhulgas homoloogid, mis pärinevad inimesest erinevatelt liikidelt, saab valmistada protsessiga, milles skriinitakse panka karmides hübridiseerimistingimustes märgistatud sondiga, mis sisaldab leiutisekohast polünukleotiidset järjestust või selle fragmenti, ja isoleeritakse seda polünukleotiidset järjestust sisaldav täispikk cdna ja genoomsed kloonid. Sellised hübridiseerimismeetodid on asjatundjatele tuntud. Järjestuste võrdlemisel on üks järjestus tavaliselt referentsjärjestus, millega võrreldakse teisi järjestusi. Järjestuse võrdlusalgoritmi kasutamisel sisestatakse katse- ja referentsjärjestused arvutisse, vajadusel määratakse alamjärjestuse koordinaadid ja seadistatakse järjestuste algoritmi programmi parameetrid. Tavaliselt kasutatakse vaikimisi programmi parameetreid, kuid vajadusel saab kasutada ka teisi parameetreid. Järjestuste võrdlusalgoritm arvutab seejärel programmi parameetrite järgi katsejärjestuste ja referentsjärjestuse vahelise identsuse protsendi. Võrdlusaken on käesolevas kontekstis mis tahes hulgast külgnevatest positsioonidest koosnev segment, mille pikkus on valitud rühmast, kuhu kuulub 2 kuni 600, tavaliselt umbes 50 kuni umbes 0, sagedamini umbes 0 kuni umbes 0 külgnevat positsiooni, mille korral järjestust võib pärast kahe järjestuse reastamist võrrelda samast hulgast külgnevatest positsioonidest koosneva referentsjärjestusega. Võrdlemisel kasutatavad järjestuste reastamise meetodid on asjatundjatele tuntud. Võrreldavaid järjestusi saab optimaalselt reastada näiteks, kuid mitte ainult, lokaalse homoloogia algoritmiga, mida kirjeldavad Smith and Waterman (1970) Adv. Appl. Math. 2:482c, homoloogilise reastamise algoritmiga, mida kirjeldavad Needleman and Wunsch (1970) J. Mol. Biol. 48:443, sarnasuste otsimise meetodiga, mida kirjeldavad Pearson and Lipman (1988) Proc. Nat'l. Acad. Sci. USA 85:2444, algoritmide arvutipõhiste (GAP, BESTFIT, FASTA, and TFASTA in the Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI) rakendamise teel, käsitsi reastamise ja visuaalse vaatlemise teel (vt nt Ausubel et al., Current Protocols in Molecular Biology (1995 supplement)).
52 5 50 Üheks näiteks algoritmist, mis sobib järjestuste vahelise identsuse ja sarnasuse määramiseks on BLAST ja BLAST 2.0 algoritmid, mida kirjeldatakse vastavalt allikates Altschul et al. (1997) Nuc. Acids Res. : ja Altschul et al. (1990) J. Mol. Biol. 2: BLAST analüüside tegemiseks vajalikku tarkvara on võimalik saada asutuselt National Center for Biotechnology Information aadressil BLASTalgoritmi paramaatri W, T ja X määravad reastamise tundlikkuse ja kiiruse. BLASTN programmis (nukleotiidsete järjestuste puhul) kasutatakse vaikeväärtustena sõnapikkust/wordlength (W) = 11, ootus/ expectation (E) =, M=5, N=-4 ja mõlema ahela võrdlust. Aminohappeliste järjestuste puhul kasutatakse BLASTP programmis vaikeväärtustena sõnapikkust 3 ja ootust ja BLOSUM62 punktimaatriksi (vt Henikoff and Henikoff (1992) Proc. Natl. Acad Sci. USA 89:9) reastusi (B) 50, ootust (E), M=5, N=-4 ja mõlema ahela võrdlust. BLAST algoritmi kasutamisel on low complexity filter tavaliselt desaktiveeritud. BLAST algoritm teeb tavaliselt ka kahe järjestuse sarnasuse statistilise analüüsi (vt nt Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90: ). Üks BLAST algoritmi puhul kasutatav sarnasuse näitaja on smallest sum probability, mis näitab kahe nukleotiidse või aminohappelise järjestuse juhusliku kokkulangevuse tõenäosust. Nukleiinhapet näiteks peetakse referentsjärjestuse sarnaseks, kui testitava nukleiinhappe ja referentsnukleiinhappe võrdlemisel on the smallest sum probability väiksem kui 0,2 või väiksem kui umbes 0,01 või väiksem kui umbes 0,001. Väljend hübridiseerub selektiivselt (või spetsiifiliselt) tähendab seda, et antud molekul seondub, moodustab dupleksi või hübridiseerub karmides hübridiseerimistingimustes ainult konkreetse nukleotiidse järjestusega, kui see järjestus sisaldub kompleksses segus (sealhulgas, kuid mitte ainult, kogu raku või panga DNA-s või RNA-s). 30 Väljendi karmid hübridiseerimistingimused all peetakse silmas DNA, RNA, PNA või teiste nukleiinhapete mimeetikute järjestuste või nende kombinatsioonide hübridiseerimist madala ioonjõuga kõrgel temperatuuril, nagu on tehnika tasemes tuntud. Karmides tingimustes üldjuhul sond hübridiseerub nukleiinhappe kompleksses segus (sealhulgas, kuid mitte ainult, kogu raku või panga DNA-s või RNA-s) sihitava alamjärjestusega, kuid ei hübridiseeru kompleksses segus sisalduvate teiste järjestustega. Karmid tingimused sõltuvad järjestusest ja on erinevate asjaolude korral erinevad. Pikemad järjestused
53 hübridiseeruvad spetsiifiliselt kõrgemal temperatuuril. Põhjalikud juhised nukleiinhapete hübridiseerimiseks on toodud allikas Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Probes, Overview of principles of hybridization and the strategy of nucleic acid assays" (1993). Üldjuhul järgitakse karmide tingimuste valimisel põhimõttest, et temperatuur peab olema umbes 5- C madalam kui konkreetse järjestuse sulamispunkt (T m ) määratletud ioontugevuse ja ph juures. T m on temperatuur (määratletud ioontugevuse, ph ja nukleiinhappe kontsentratsiooni korral), millel 50% sondidest, mis on sihtmärgi suhtes komplementaarsed, hübridiseeruvad sihtjärjestusega tasakaaluolekus (kuna sihtjärjestusi kasutatakse liiaga, on tasakaaluolekus temperatuuril T m, 50% sondidest on tasakaaluolekus hõivatud). Karmides tingimustes võib soola kontsentratsioon olla väiksem kui umbes 1,0 M naatriumiiooni ja tavaliselt on see umbes 0,01 kuni 1,0M naatriumiooni (teisi soolasid) ph 7,0 kuni 8,3 juures ja temperatuur on lühikeste (näiteks, kuid mitte ainult, kuni 50 nukleotiidi pikkuste) sondide korral vähemalt umbes 30 C ja pikkade (näiteks, kuid mitte ainult, üle 50 nukleotiidi pikkuste) sondide korral vähemalt umbes 60 C. Karmide tingimuste loomiseks võib lisada ka destabiliseerivaid aineid nagu formamiid. Selektiivse või spetsiifilise hübridiseerumise korral võib positiivne signaal olla vähemalt kaks korda tugevam kui tausthübridiseerumine ja valikuliselt korda tugevam kui tausthübridiseerumine. Karmid hüdridiseerimistingimused näiteks võivad olla sellised: 50% formamiidi, 5X SSC, ja 1% SDS, inkubeeritakse temperatuuril 42 C või 5X SSC, 1% SDS, inkubeeritakse temperatuuril 65 C, pestakse 0,2X SSC ja 0,1% SDS-is temperatuuril 65 C. Selline pesemine võib kesta 5,, 30, 60, 1 või rohkem minutit. Käesolevas kontekstis on peetakse mõiste eukarüoot all silmas organisme, mis kuuluvad fülogeneetilisse domeeni Eucarya, nagu loomad (näiteks, kuid mitte ainult, imetajad, putukad, roomajad, linnud, jne), ripsloomad, taimed (näiteks, kuid mitte ainult, üheidulehelised, kaheidulehelised, vetikad, jne), seened, pärmid, viburilased, pisieosloomad, protistid, jne. Käesolevas kontekstis peetakse mõiste mitteeukarüoot all silmas mitteeukarüootseid organisme. Mitteeukarüootne organism võib kuuluda Eubacteria (näiteks, kuid mitte ainult,
54 5 52 Escherichia coli, Thermus thermophilus, Bacillus stearothermophilus, Pseudomonas fluorescens, Pseudomonas aeruginosa, Pseudomonas putida, jne) fülogeneetilise domeeni või Archaea (näiteks, kuid mitte ainult, Methanococcus jannaschii, Methanobacterium thermoautotrophicum, Halobacterium nagu Haloferax volcanii ja Halobacterium liigid NRC-1, Archaeoglobus fulgidus, Pyrococcus furiosus, Pyrococcus horikoshii, Aeuropyrum pernix, jne) fülogeneetilisse domeeni. Indiviid/patsient/katsealune on käesolevas kontekstis loom, leiutise mõnes teostuses imetaja ja teistes teostustes inimene, keda ravitakse, jälgitakse või kellel tehakse katseid. Loom võib olla lemmikloom (nt koer, kass ja teised sarnased loomad), taluloom (nt lehm, lammas, siga, hobune ja teised sarnased loomad) või laboratooriumiloom (nr rott, hiir, merisiga ja teised sarnased loomad). Käesolevas kontekstis on efektiivne kogus manustatava modifitseeritud mittelooduslikke aminohappeid sisaldava polüpeptiidi kogus, mis vähendab mingil määral ravitava haiguse, seinsundi või häire ühte või mitut sümptomit. Käesolevas dokumendis kirjeldatavaid kompositsioone, mis sisaldavad modifitseeritud mittelooduslikke aminohappeid sisaldavat polüpeptiidi saab manustada profülaktilises, võimendavad ja/või terapeutilises ravis. Mõistete võimendama või võimendamine/võimendav all peetakse silmas mõjususe suurendamist või soovitud toime kestvuse pikendamist. Raviainete toime võimendamise puhul peetakse mõiste võimendav all seega silmas võimet suurendada või pikendada teiste raviainete mõjusust või nende toime kestvust süsteemis. Käesolevas kontekstis on võimendamiseks efektiivne kogus kogus, millest piisab teise raviaine toime võimendamiseks soovitud süsteemis. Patsiendil kasutamise korral sõltuvad sellisel juhul kasulikud kogused haiguse, häire või seisundi raskuastmest ja kulust, eelnevast ravist, patsiendi tervislikust seisundist ja vastusest ravimitele ning raviarsti äranägemisest. Käesolevas kontekstis peetakse mõiste modifitseeritud all silmas polüpeptiidis tehtud mis tahes muudatusi nagu muudatused polüpeptiidi pikkuses, aminohappelises järjestuses, keemilises struktuuris, polüpeptiidi kotranslatsioonilises modifikatsioonis või translatsioonijärgses modifikatsioonis.
55 53 Vorm (modifitseeritud) näitab, et kõnealuseid polüpeptiide modifitseeritakse valikuliselt ehk kõnealused polüpeptiidid võivad olla modifitseeritud või modiftseerimata. 5 Mõiste translatsioonijärgselt modifitseeritud all peetakse silmas loodusliku või mitteloodusliku aminohappe mis tahes modifikatsiooni, mis tekib selles aminohappes pärast selle kaasamist polüpeptiidi ahelasse. See mõiste hõlmab näiteks, kuid mitte ainult, kotranslatsioonilisi in vivo modifikatsioone, kotranslatsioonilisi in vivo modifikatsioone ja translatsioonijärgseid in vitro modifikatsioone. Profülaktilistes rakendustes manustatakse FGF-21 polüpeptiidi sisaldavaid kompositsioone patsiendile, kellel on konkreetse haiguse, häire või seisundi tekke oht. Sellisel juhul manustatavat kogust nimetatakse profülaktiliselt efektiivseks koguseks. Selliste annuste täpne suurus sõltub patsiendi tervislikust seisundist, kehakaalust ja teistest sarnastest näitajatest. Asjatundjad oskavad tavapärase katsetamise teel profülaktiliselt efektiivseid koguseid kergesti määrata (nt annuse eskaleerimisel põhineva kliinilise katsega). Mõiste kaitstud viitab sellele, et süsteemis on kaitserühm või fragment, mis hoiab teatud reaktsioonitingimustes keemiliselt reaktsioonivõimelise funktsionaalrühma reaktsioonid. Kaitserühmad võivad olla erinevad ja sõltuvad sellega kaitstava keemiliselt reaktsioonivõimelise rühma tüübist. Kui keemiliselt reaktsioonivõimeline rühm on amiinvõi hüdrasiidrühm, võib kaitserühma valida rühmast, kuhu kuuluvad tertbutüüloksükarbonüül (t-boc) ja 9-fluorenüülmetoksükarbonüülühm (Fmoc). Kui keemiliselt reaktsioonivõimeline rühm on tioolrühm, võib kaitserühm olla ortopüridüüldisulfiid. Kui keemiliselt reaktsioonivõimeline rühm on karboksüülhape nagu butaan- või propioonhape või hüdroksüülrühm, võib kaitserühm olla bensüül- või alküülrühm nagu metüül-, etüül- või tert-butüülrühm. Käesolevas dokumendis kirjeldatavatel meetodite ja kompositsioonide puhul võib kasutada ka teisi tehnika tasemes tuntud kaitserühmi, sealhulgas fotolabiilseid rühmi nagu Nvoc ja MeNvoc. Käesoleva leiutise kohastel meetoditel või kompositsioonides võib kasutada ka teisi tehnika tasemes tuntud kaitserühmi. Blokeerivad/kaitserühmad võib valida näiteks, kuid mitte ainult, järgmiste rühmade seast:
56 54 5 Teisi kaitserühmi kirjeldatakse näiteks allikas Greene and Wuts, Protective Groups in Organic Synthesis, 3rd Ed., John Wiley & Sons, New York, NY, Terapeutilistes rakendustes manustatakse modifitseeritud, mittelooduslikke aminohappeid sisaldavat polüpeptiidi sisaldavaid kompositsioone patsiendile, kellel on juba tekkinud haigus, seisund või häire, koguses, millest piisab haiguse, häire või seisundi väljaravimiseks või selle sümptomite vähemalt osaliseks peatamiseks. Sellist kogust nimetatakse terapeutiliselt aktiivseks koguseks ja see sõltub haiguse, häire või seisundi raskuastmest ja kulust, eelnevast ravist, patsiendi tervislikust seisundist ja vastusest ravimitele ning raviratsi äranägemisest. Asjatundjad oskavad tavapärase katsetamise teel profülaktiliselt efektiivseid koguseid kergesti määrata (nt annuse eskaleerimisel põhineva kliinilise katsega). Mõistet ravima kasutatakse profülaktilise ja/või terapeutilise ravi tähenduses. Leiutisekoased mittelooduslikult kodeeritud aminohappeid sisaldavad polüpeptiidid võivad sisaldada isotoobiga märgistatud ühendeid, milles üks või mitu aatomit on asendatud aatomiga, mille aatommass või massiarv erineb tavaliselt looduses leiduva aatomi aatommassist või massiarvust. Isotoobid, mida saab sisestada leiutisekohastesse ühenditesse, on näiteks vesiniku, süsiniku, lämmastiku, hapniku, fluori ja kloori isotoobid nagu vastavalt 2 H, 3 H, 13 C, 14 C, N, 18 O, 17 O, 35 S, 18 F, 36 Cl. Käesolevas dokumendis kirjeldatavad teatud isotoobiga märgistatud ühendid, näiteks ühendid, millesse on sisestatud radioaktiivsed isotoobid nagu 3 H ja 14 C, võiad olla kasulikud ravimi ja/või
57 55 substraadi kudedes jaotumise analüüsides. Isotoopidega nagu deuteerium ehk asendamisel tekivad teatud terapeutilised eelised, mis tulenevad suuremast metaboolsest stabiilsusest, nt pikenenud in vivo poolväärtusajast või manustamisvajaduse vähenemisest. 2 H 5 Leiutisekohaste kompositsioonide hulka kuuluvad ka kõik isomeerid, sealhulgas diastereomeerid, enantiomeerid ja nende segud. Teistes või lisateostustes mittelooduslikult kodeeritud aminohappeid sisaldavad polüpeptiidid metaboliseeritakse organism manustamisel, et tekiks metaboliit, mida seejärel kasutatakse soovitud toime, sealhulgas soovitud ravitoime esilekutsumiseks. Teistes või lisateostustes kasutatakse mittelooduslikult kodeeritud aminohapet sisaldavate polüpeptiidide aktiivseid metaboliite. Mõnel juhul võivad mittelooduslikult kodeeritud aminohapet sisaldavad polüpeptiidid olla tautomeeride kujul. Käesolevas dokumendis kirjeldatavad mittelooduslikult kodeeritud aminohapet sisaldavad polüpeptiidid võivad olla ka solvateerimata ja ka solvateeritud kujul koos farmatseutiliselt sobivate lahustitega nagu vesi, etanool ja teised sarnased ühendid. Sellised solvateeritud vormid kuuluvad samuti käesoleva leiutise ulatusse. Asjatundjad teavad, et mõned käesolevas dokumendis kirjeldatavad ühendid võivad olla mitme tautomeeri kujul. Kõik sellised tautomeersed vormid kuuluvad käesoleva leiutise kohaste ühendite hulka. Kui ei ole öeldud teisiti, kasutatakse seoses käesolevas leiutisega tavapäraseid massispektromeetria, NMR, HPLC, valkude keemia, biokeemia, rekombinantse DNA ja farmakoloogilisi meetodeid, mis on asjatundjatele teada. LEIUTISE ÜKSIKASJALIK KIRJELDUS I. Sissejuhatus Käesoleva leiutisega pakutakse FGF-21 molekulid, mis sisaldavad ühte mittelooduslikku aminohapet. Leiutise teatud teostustes on ühte mittelooduslikku aminohapet sisaldavas
58 FGF-21 polüpeptiidis vähemalt üks translatsioonijärgne modifikatsioon, milleks on polüpeptiidi külge kinnitatud vees lahustuva polümeeri molekul. Leiutise ühes teostuses kinnitatakse polüpeptiidi külge nimetatud vähemalt ühe translatsioonijärgse modifikatsioonina molekul, mis võib olla näiteks, kuid mitte ainult: märgis, värvaine, polümeer, vees lahustuv polümeer, polüetüleenglükooli derivaat, fotoristside, radionukliid, tsütotoksiline ühend, raviaine, afiinsusmärgis, fotoafiinsusmärgis, reaktsioonivõimeline ühend, vaik, teine valk või polüpeptiid või polüpeptiidi analoog, antikeha või antikeha fragment, metallkelaator, kofaktor, rasvhape, süsivesik, polünukleotiid, DNA, RNA, antisenss-polünukleotiid, sahhariid, vees lahustuv dendrimeer, tsüklodekstriin, inhibeeriv ribonukleiinhape, bioloogiline materjal, nanoosake, spin-märgis, fluorofoor, metalli sisaldav fragment, radioaktiivne fragment, uudne funktsionaalrühm, teiste molekulidega kovalentselt või mittekovalentselt interakteeruv rühm, valguse poolt lõhustatav (photocaged) fragment, aktiiniga kiiritamise teel ergastatav fragment, fotoisomeriseeritav fragment, biotiin, biotiini derivaat, biotiini analoog, rasket aatomit sisaldav rühm, keemiliselt lõhustatav rühm, valguse toimel lõhustatav rühm, pikendatud kõrvalahel, süsinikuga seotud suhkur, redoks-aktiivne aine, amino-tiohape, toksiline fragment, isotoobiga märgistatud fragment, biofüüsikaline sond, fosforestseeruv märgis, kemoluminetseeruv märgis, elektrontihe rühm, magnetiline rühm, interkaleeruv rühm, kromofoor, energia ülekande aine, bioloogiliselt aktiivne aine, tuvastatav märgis, väike molekul, kvantpunkt, nanotransmitter, radionukleotiid, radiotransmitter, neutronhaarde agent või eespool nimetatud ainete kombinatsioon või mis tahes teine soovikohane ühend või aine, mis sisaldab teist rühma, mis reageerib esimest reaktsioonivõimelist rühma sisaldava vähemalt ühe mitteloodusliku aminohappega, kusjuures selline reaktsioonivõimeline rühm on sisestatud keemilistel meetoditel, mis asjatundjatele teadaolevalt sobivad selliste reaktsioonivõimeliste rühmade sisestamiseks. Esimene reaktsioonivõimeline rühm näiteks on alkünüülrühm (sealhulgas, kuid mitte ainult, mittelooduslikus aminohappes p-propargüüloksüfenüülalaniinis, milles sisalduvat propargüülrühma nimetatakse vahel ka atsetüleenrühmaks) ja teine reaktsioonivõimeline rühm on asidorühm ning nende puhul kasutatakse keemilise [3+2] tsükloliitumise meetodeid. Teise näite kohaselt on esimene reaktsioonivõimeline rühm asidorühm (sealhulgas, kuid mitte ainult, mittelooduslikus aminohappes p-asido-l-fenüülalaniin) ja teine reaktsioonivõimeline rühm on alkünüülrühm. Leiutisekohase modifitseeritud FGF- 21 polüpeptiidi teatud teostuste korral kasutatakse vähemalt ühte mittelooduslikku
59 aminohapet (sealhulgas, kuid mitte ainult, mittelooduslikku aminohapet, mis sisaldab funktsionaalset ketorühma), milles on vähemalt üks translatsioonijärgne modifikatsioon, kusjuures selline vähemalt üks translatsioonijärgne modifikatsioon hõlmab sahhariidfragmenti. Teatud teostustes tehakse translatsioonijärgne modifikatsioon in vivo eukarüootses rakus või mitteeukarüootses rakus. Sellise molekuli võib kinnitada polüpeptiidi külge linkeri, polümeeri, vees lahustuva polümeeri või muu molekuli kaudu. Sellise molekuli võib siduda ka otse polüpeptiidiga. Teatud teostustes on valgus vähemalt üks translatsioonijärgne modifikatsioon, mis on tehtud in vivo ühes peremeesrakus, kusjuures sellist translatsioonijärgset modifikatsiooni ei tehta tavaliselt teist tüüpi peremeesrakus. Teatud teostustes on valgus vähemalt üks translatsioonijärgne modifikatsioon, mis on tehtud in vivo eukarüootses rakus, kusjuures sellist translatsioonijärgset modifikatsiooni ei tehta tavaliselt mitteeukarüootses rakus. Translatsioonijärgsete modifikatsioonide seas on näiteks, kuid mitte ainult, glükosüülimine, atsetüülimine, atsüülimine, lipiidiga modifitseerimine, palmitüülimine, palmitaadi lisamine, fosforüüliline, glükolipiidse sideme modifitseerimine modifitseerimine ja teised sarnased modikatsioonid. Ühes teostuses seisneb translatsioonijärgne modifitseerimine oligosahhariidi kinnitamises GlcNAc-asparagiini sideme kaudu asparagiini külge (sealhulgas, kuid mitte ainult, juhul kui oligosahhariid sisaldab (GlcNAc-Man) 2 -Man-GlcNAc-GlcNAc, ja teisi sarnaseid üksusi). Teises teostuses seisneb translatsioonijärgne modifitseerimine oligosahhariidi kinnitamises GalNAc-seriini, GalNAc-treoniini, GlcNAc-seriini või GlcNAc-treoniini sideme kaudu seriini või treoniini külge. Teatud teostustes võib leiutisekohane valk või polüpeptiid sisaldada sekretsiooni- või lokalisatsioonijärjestust, epitoobi märgist, FLAG märgist, polühistidiini märgist, GST fusiooni ja/või muud sarnast üksust. Sekretsiooni signaaljärjestused on näiteks, kuid mitte ainult, prokarüootne sekretsiooni signaaljärjestus, eukarüootne sekretsiooni signaaljärjestus, eukarüootne sekretsiooni signaaljärjestus, mille 5 -otsa on bakteris ekspresseerimise jaoks optimeeritud, uudne sekretsiooni signaaljärjestus, pektaatlüaasi sekretsiooni signaaljärjestus, Omp A sekretsiooni signaaljärjestus ja faagi sekretsiooni signaaljärjestus. Sekretsiooni signaaljärjestused on näiteks, kuid mitte ainult, STII (prokarüootne), Fd GIII ja M13 (faag), Bgl2 (pärm) ja transposonist pärinev signaaljärjestus bla. Igat sellist järjestust võib vastavalt polüpeptiidi soovitud kasutusviisile modifitseerida ehk näiteks, kuid mitte ainult, asendada ühe signaaljärjestuse teistsuguse signaaljärjestusega, asendada liiderjärjestuse teistsuguse
60 5 58 liidejärjestusega, jne. Huvipakkuv valk või polüpeptiid sisaldab ühte mittelooduslikku aminohapet. Üks looduslikus valgus sisalduv konkreetne aminohape asendatakse mitteloodusliku aminohappega. Käesoleva leiutisega pakutakse meetodid ja kompositsioonid, mis põhinevad ühte mittelooduslikult kodeeritud aminohapet sisaldaval FGF-21 polüpeptiidil. Ühe mittelooduslikult kodeeritud aminohappe sisestamisel FGF-21 saab kasutada konjugatsioonireaktsioone, mis hõlmavad spetsiifilisi keemilisi reaktsioone, sealhulgas, kuid mitte ainult, ühe mittelooduslikult kodeeritud aminohappega, kuid mitte tavapärase aminohappega. Mõnes teostuses seotakse mittelooduslikult kodeeritud aminohapet sisaldav FGF vees lahustuva polümeeriga nagu polüetüleenglükool (PEG) mittelooduslikult kodeeritud aminohappe kõrvaahela kaudu. Leiutisega pakutakse väga efektiivne meetod valkude selektiivseks modifitseerimiseks PEG derivaatidega, kusjuures see meetod hõlmab mittegeneetiliselt kodeeritud aminohapete, sealhulgas, kuid mitte ainult, aminohapete, mis sisaldavad funktsionaalseid rühmi või asendajaid, mida ei leidu looduslikus aminohappes, sealhulgas ketoon-, asiid- või atsetüleenfragmenti, valkudesse vastusena selektorkoodonile ja sellele järgnevat sisestatud aminohapete modifitseerimist sobiva reaktsioonivõimelise PEG derivaadiga. Sisestatud aminohappe kõrvalahelaid saab seejärel modifitseerida keemilistel meetoditel, mis asjatundjatele teadaolevalt sobivad kasutamiseks mittelooduslikult kodeeritud aminohappes sisalduvate konkreetsete funktsionaalsete rühmade või asendajate korral. Käesoleva leiutise korral saab vees lahustuva polümeeri sisestada valku mitmesugustel tuntud keemilistel meetoditel. Selliste meetodite seas on näiteks, kuid mitte ainult, Huisgeni [3+2] tsükloliitumise reaktsioon (vt nt Padwa, A. Comprehensive Organic Synthesis, Vol. 4, (1991) Ed. Trost, B. M., Pergamon, Oxford, p ; ja Huisgen, R. 1,3-Dipolar Cycloaddition Chemistry, (1984) Ed. Padwa, A., Wiley, New York, p ) näiteks, kuid mitte ainult, vastavalt atsetüleen- või asiidderivaatidega. Kuna Huisgeni [3+2] tsükloliitumise meetod hõlmab tsükloliitumist ja mitte nukleofiilset asendusreaktsiooni, saab valke väga selektiivselt modifitseerida. Selle reaktsiooni saab läbi viia toatemperatuuril vees väga hea regioselektiivsusega (1,4 > 1,5), lisades reaktsioonisegule katalüütilistes kogustes Cu(I) sooli. Vt nt Tornoe, et al., (02) J. Org. Chem. 67: ; ja Rostovtsev, et al., (02) Angew. Chem. Int. Ed. 41:96-99; ja WO 03/1972. Molekul, mille saab [3+2] tsükloliitumise teel lisada leiutisekohasele valgule, võib olla põhimõtteliselt mis
61 tahes molekul, millel on sobiv funktsionaalrühm või asendaja, sealhulgas, kuid mitte ainult asido- või atsetüleenderivaat. Sellised molekulid saab lisada mittelooduslikule aminohappele, millel on atsetüleenrühm, sealhulgas, kuid mitte ainult, p- propargüüloksüfenüülalaniin- või asidorühm, sealhulgas, kuid mitte ainult, p-asidofenüülalaniinrühm. Huisgeni [3+2] tsükloliitumise puhul tekkiv viielüliline tsükkel ei ole redutseerivates tingimustes üldjuhul tagasipööratav ja see ei hüdrolüüsu vesikeskkonnas ka pikema aja jooksul. Leiutisekohaste aktiivsete PEG derivaatidega saab seega modifitseerida paljude erinevate ainete füüsikalisi ja keemilisi omadusi nõudlikes vesitingimustes. Veelgi olulisem on asjaolu, et kuna asiid- ja atsetüleenrühmad on üksteise suhtes spetsiifilised (ja ei reageeri näiteks mitte ühegi aminohappega, mis kuulub tavapärase geneetiliselt kodeeritud aminohappe hulka, saab valke väga suure selektiivsusega ühes või mitmes spetsiifilises kohas modifitseerida. Leiutisega pakutakse ka PEG derivaatide ja nendega seotud hüdrofiilsete polümeeride vees lahustuvad ja hüdrolüütiliselt stabiilsed derivaadid, millel on üks või mitu atsetüleen- või asiidfragmenti. Atsetüleenfragmente sisaldavad PEG polümeeri derivaadid on väga selektiivsed sidumiseks asiidfragmentidega, mis on sisestatud selektiivselt valkudesse vastusena selektorkoodonile. Asiidfragmente sisaldavad PEG polümeeri derivaadid on samuti väga selektiivsed sidumiseks atsetüleenfragmentidega, mis on sisestatud selektiivselt valkudesse vastusena selektorkoodonile. Asiidfragmendid on täpsemalt näiteks, kuid mitte ainult, alküülasiidid, arüülasiidid ja nende asiidide derivaadid. Alküül- ja arüülasiidide derivaadid võivad sisaldada ka teisi asendajaid, tingimusel et derivaadid säilitavad võime reageerida spetsiifiliselt atsetüleeniga. Atsetüleenfragmendite hulka kuuluvad alküül- ja arüülatsetüleenid ja nende derivaadid. Alküül- ja arüülatsetüleenide derivaadid võivad sisaldada teisi asendajaid, tingimusel et derivaadid säilitavad võime reageerida spetsiifiliselt asiidiga. Käesoleva leiutise pakutakse paljude erinevate funktsionaalrühmade, asendajate või fragmentidega ainete konjugaadid teiste ainetega, sealhulgas vees lahustuva polümeeriga. Lisaks kirjeldatakse konjugaate näiteks, kuid mitte ainult märgise, värvaine, polümeeri, polüetüleenglükooli derivaadi, fotoristsideme, radionukliidi, tsütotoksilise ühendi, raviaine, afiinsusmärgise, fotoafiinsusmärgise, reaktsioonivõimelise ühendi, vaigu, teise valgu või polüpeptiidi või polüpeptiidi analoogi, antikeha või antikeha fragmendi, metallkelaatori, kofaktori, rasvhappe, süsivesiku, polünukleotiidi, DNA, RNA, antisensspolünukleotiidi, sahhariidi, vees lahustuva dendrimeeri, tsüklodekstriini, inhibeeriva ribonukleiinhappe, bioloogilise materjali, nanoosakese, spin-märgise, fluorofoori, metalli
62 sisaldava fragmendi, radioaktiivse fragmendi, uudse funktsionaalrühma, teiste molekulidega kovalentselt või mittekovalentselt interakteeruva rühma, valgusega poolt lõhustatava fragmendi, aktiiniga kiiritamise teel ergastatava fragmendi, fotoisomeriseeritava fragmendi, biotiini, biotiini derivaadi, biotiini analoogi, rasket aatomit sisaldava rühma, keemiliselt lõhustatava rühma, valguse toimel lõhustatava rühma, pikendatud kõrvalahela, süsinikuga seotud suhkru, redoks-aktiivse aine, amino-tiohappe, toksiline fragmendi, isotoobiga märgistatud fragmendi, biofüüsikalise sondi, fosforestseeruva märgise, kemoluminestseeruva märgise, elektrontiheda rühma, magnetilise rühma, interkaleeruva rühma, kromofoori, energia ülekande agendi, bioloogiliselt aktiivse aine, tuvastatava märgise, väikese molekuli, kvantpunkti, nanotransmitteri, radionukleotiidi, radiotransmitteri, neutronhaarde agendi või eespool nimetatud ainete mis tahes kombinatsiooniga või mis tahes muu soovikohase ühendi või ainega. Käesolev leiutis hõlmab ka asiid- või atsetüleenfragmentiga ainete konjugaate OEG polümeeri derivaatidega, milles on vastavad atsetüleen- või asiidfragmendid. Näiteks asiidfragmenti sisaldava PEG polümeeri saab siduda atsetüleenrühma sisaldavat mittegeneetiliselt kodeeritud aminohapet sisaldavas valgus teatud positsioonis bioloogiliselt aktiivse molekuliga. Side, mille kaudu PEG ja bioloogiliselt aktiivne molekul seotakse, võib olla näiteks, kuid mitte ainult Huisgeni [3+2] tsükloliitumise reaktsiooni saadus. Tehnika tasemes on teada, et PEG-iga saab modifitseerida bioloogiliste materjalide pindu (vt nt USA patenti ; Mehvar, R., J. Pharm Pharm Sci., 3(1):1-136 (00)). Käesolev leiutis hõlmab ka bioloogilisi materjale, mille pinnal on üks või mitu reaktsioonivõimelist asiid- või atsetüleensaiti ja üks või mitu leiutisekohast asiid- või atsetüleenfragmenti sisaldavat polümeeri, mis on seotud selle pinnaga Huisgeni [3+2] tsükloliitumise teel loodud sideme kaudu. Bioloogilised materjalid ja teised ained saab liita asiid- või atsetüleen-aktiveeritud polümeeri derivaatidega ka asiid- või atsetüleensidemest erineva sideme kaudu ehk näiteks üksusega, mis sisaldab karboksüülhappe, amiini, alkoholi või tioolfragmenti, jättes asiidvõi atsetüleenfragmendi vabaks järgnevate reaktsioonide läbiviimiseks. Käesolev leiutis hõlmab ka leiutisekohaste asiid- ja atsetüleenrühma sisaldavate polümeeride sünteesimeetodit. Asiidfragmenti sisaldava PEG derivaadi korral saab asiidfragmendi siduda otse polümeeri süsinikuaatomiga. Asiidfragmenti sisaldava PEG derivaadi saab valmistada ka sidumisaine, mille ühes otsas on asiidfragment, tavapärase
63 aktiveeritud polümeeri külge kinnitamise teel, mille tulemusena saadud polümeeri otsas on asiidfragment. Atsetüleenfragmenti sisaldava PEG derivaadi korral saab asiidfragmendi siduda otse polümeeri süsinikuaatomiga. Atsetüleenfragmenti sisaldava PEG derivaadi saab valmistada ka sidumisaine, mille ühes otsas on atsetüleenfragment tavapärase aktiveeritud polümeeri külge kinnitamise teel, mille tulemusena saadud polümeeri otsas on atsetüleenfragment. Asiidfragmenti sisaldava PEG derivaadi korral toimub vähemalt ühe aktiivse hüdroksüülfragmendiga vees lahustuva polümeeriga reaktsioon, mille tulemusel tekib asendatud polümeer, millel on reaktsioonivõimelisem fragment nagu lahkuv mesülaat-, tresülaat-, tosülaat- või halogeenrühm. Sulfonüülhappe halogeniide, halogeeniaatomeid ja teisi lahkuvaid rühmi sisaldavate PEG derivaatide valmistus- ja kasutusmeetodid on asjatundjatele teada. Saadud asendatud polümeeriga viiakse seejärel läbi reaktsioon, mille tulemusena asendatakse polümeeri otsas paiknev asiidfragment reaktsioonivõimelisema fragmendiga. Selle asemel võib vees lahustuva polümeeri, millel on vähemalt üks aktiivne nukleofiilne või elektrofiilne rühm, panna reageerima sidumisainega, millel on ühes otsas asiidfragment, mille tulemusena moodustub PEG polümeeri ja sidumisaine vahele kovalentne side ja polümeeri otsa kinnitatakse asiidfragment. Nukleofiilsed ja elektrofiilsed fragmendid, sealhulgas amiinid, tioolid, hüdrasiidid, alkoholid, karboksülaadid, aldehüüdid, ketoonid, tioestrid ja teised sarnased ühendid on asjatundjatele teada. Atsetüleenfragmenti sisaldava PEG derivaadi korral viiakse vees lahustuva polümeeriga, millel on vähemalt üks aktiivne hüdroksüülfragment, läbi reaktsioon, mille käigus eemaldatakse atsetüleenfragmenti sisaldavast eelkäijast halogeenrühm või muu aktiveeritud lahkuv rühm. Selle asemel võib vees lahustuva polümeeri, millel on vähemalt üks aktiivne nukleofiilne või elektrofiilne fragment, panna reageerima sidumisainega, mille ühes otsas on atsetüleenfragment, nii et PEG polümeeri ja sidumisaine vahele moodustub kovalentne side ja polümeeri otsa kinnitatakse atsetüleenfragment. Halogeenifragmentide, aktiveeritud lahkuva rühma ning nukleofiilsete ja elektrofiilsete fragmentide kasutamine orgaanilises sünteesis ja PEG derivaatide kasutamine on asjatundjatele hästi teada. Käesoleva leiutisega pakutakse ka meetod valkude selektiivseks modifitseerimiseks, mille korral lisatakse modifitseeritud valgule veel teisi aineid, näiteks, kuid mitte ainult, vees lahustuvaid polümeere nagu PEG ja PEG derivaadid, mis sisaldavad asiid- või atsetüleenfragmenti. Kui on oluline tagada bioloogiline ühilduvus,
64 62 stabiilsus, lahustuvus ja immunogeensuse puudumine, saab asiid- ja atsetüleenfragmenti sisaldavate PEG derivaatidega saab modifitseerida pindade ja molekulide omadusi, luues ühtlasi tehnika tasemes tuntud meetodist selektiivsema vahendi PEG derivaatide kinnitamiseks valkude külge. 5 II. Fibroblastide kasvufaktorid 30 Kuna fibroblastide kasvufaktorid on mõjusad paljude erinevate raku- ja koetüüpide kasvu, proliferatsiooni, ellujäämise ja diferentseerumise soodustamisel, kasutatakse neid raviainetena paljude erinevate näidustuste korral, sealhulgas haavade paranemise nagu luu- ja lihaskonna häired, nt luumurrud, ligamentide ja kudede taastamine, kõõlusepõletik, jne; nahaga seotud seisundid, nt põletused, lõikehaavad, rebendid, lamatised, aeglaselt paranevad haavandid, jne; kudede kaitsmine, taastamine ja angiogeneesi indutseerimine müokardi infarkti ja isheemia puhul, neuroloogiliste häirete nagu näiteks neurodegeneratiivse haiguse ja rabanduse ravis ning silmahaiguste, sealhulgas maakuli degeneratsiooni ravis. Seni identifitseeritud fibroblastide kasvufaktori (FGF) valgud kuuluvad signaliseerivate molekulide perekonda, mis reguleerivad mitmesuguste rakutüüpide kasvu ja diferentseerumist. FGF valkude tähtsus inimese füsioloogias ja patoloogias on osaliselt seotud nende olulise rolliga embrüogeneesis, veresoonte tekkes ja arengus ning luude arengus. In vitro katsed on näidanud, et FGF osaleb endoteliaalsete rakkude, veresoonte silelihaste rakkude, fibroblastide ning südame ja luustiku müotsüütide kasvus ja pooldumises. Teisi FGF perekonna liikmeid ja nende bioloogilisi rolle kirjeldatakse allikates Crossley et al., Development 121: (1995); Ohuchi et al., Development 124: (1997); Gemel et al., Genomics 35:3-7 (1996); ja Ghosh et al., Cell Growth and Differentiation 7: (1996). FGF valgud on inimeste tervise ja haiguste seisukohalt olulised ka nende rolli tõttu vähirakkude arengus. Näiteks FGF-8 identifitseeriti androgeeni indutseeriva kasvufaktorina rinna- ja eesnäärmevähi rakkudes (Tanaka et al., FEBS Lett. 363: (1995) ja P.N.A.S. 89: (1992)). FGF roll normaalses arengus on osaliselt välja selgitatud FGF retseptorite uurimise teel. Wilke, T. et al., Dev. Dynam. 2:41-52 (1997) avastasid, et kanadel paiknesid FGFR1,
65 63 FGFR2 ja FGFR3 transkriptid embüronaalses arengujärgus pea spetsiifilistes piirkondades. Nende ekspressioonimuster oli korrelatsioonis Crouzoni sündroomi ehk seisundi, millele on iseloomulik luude ebanormaalne membraanidevaheline moodustumine Belluardo, N. et al., Jour. Comp. Neur. 379: (1997) uurisid FGFR 1, 2 ja 3 mrna-de lokaliseerumist rottide ajus ja avastasid rakkude spetsiifilisuse mitmes ajupiirkonnas. FGFR1 ja FGF-2 mrna-sid ekspresseeriti ka astrogliia reaktsioonivõimelistes rakkudes (astroglial reactive cells) pärast ajukahjustuse teket, mis toetas teatud FGF-de rolli ajuhaigustes ja kahjustustes. Ozawa, K. et al., Mol. Brain Res. 41: (1996) avaldatud andmetel suurenes FGF-1 ja FGF-5 ekspressioon pärast sündi, FGF-3, FGF-6, FGF-7 ja FGF-8 geene aga ekspresseeriti intensiivsemalt embrüonaalse arengu viimastes staadiumites ja mitte pärast sündi. FGF-21 ja selle retseptori signaalide edasikandes või osaleda ka kofaktor Klotho beta (K1b). Avaldatud andmetel suurendab Klb FGFR1 ja FGFR-4 võimet seonduda FGF-21-ga. Klb on membraani üks kord läbiv valk ja kuigi selle täielikult transmembraanse vormi roll ei ole teada, on seoses FGF-23-ga näidatud, et Klotho suurendab FGF-23 seondumist ja FGF retseptori fosforüülimist, Klotho beeta aga suurendab FGF-21 seondumist (H. Kurosu, Y. Ogawa, M. Miyoshi, M. Yamamoto, A. Nandi, K. P. Rosenblatt, M. G. Baum, S. Schiavi, M.-C. Hu, O. W. Moe, M. Kuro-o, Regulation of fibroblast growth factor-23 signaling by Klotho. J. Biol. Chem. 281, (06)). Katoh et al. (International Journal of Oncology (06) 29: ) kirjeldavad FGF perekonda ja selle perekonna liikmete fülogeneetilisi analüüse. Katoh et al. käsitlevad ka signaalideradade võrgustikku seedetraktis. Plotnikov et al. (Cell (1999) 98: ) kirjeldavad FGF2 kristallstruktuuri koos FGF retseptoriga 1 (FGFR1) ja 2-kordse sümmeetrilise dimeeriga, mis tekib kahe sellise kompleksi vahele. Plotnikov et al. pakuvad mudeli retseptori dimeriseerimiseks ja dimeriseerimise esilekutsumiseks FGF ja hepariiniga. Tulevikus avastatakse tõenäoliselt uusi FGF perekonna liikmeid. FGF perekonna uusi liikmeid saab identifitseerida ennustatavate valgujärjestuste sekundaarse ja tertsiaarse struktuuri kompuuteranalüüside abil ja selekteerimismeetoditel, mis on ette nähtud konkreetse sihtmärgiga seonduvate molekulide identifitseerimiseks. FGF perekonna kirjeldus on seega toodud vaid illustreerimiseks ja näitena ning see ei piira käesolevas dokumendis kirjeldatavate meetodite, kompositsioonide, strateegiate ja tehnikate ulatust. Lisaks kasutatakse käesolevas taotluses mõistet FGF-21 üldise mõistena
66 64 ja selle all peetakse näitlikult silmas selle perekonna mis tahes liikmeid. Seega tuleb mõista, et selles dokumendis seoses FGF-21 polüpeptiididega kirjeldatavaid modifikatsioone ja keemilisi reaktsioone võib kasutada ka FGF perekonna mis tahes teiste liikmete, sealhulgas käesolevas dokumendis konkreetselt loetletud liikmete puhul. 5 III. Käesoleva leiutise puhul kasutatavad üldised rekombinantsed nukleiinhapetel põhinevad meetodid Käesoleva leiutise paljude teostuste korral isoleeritakse, kloonitakse ja sageli modifitseeritakse huvipakkuvat FGF-21 polüpeptiidi kodeerivaid nukleiinhappeid rekombinantsetel meetoditel. Leiutise selliseid teostusi kasutatakse näiteks, kuid mitte ainult, valgu ekspresseerimiseks või FGF-21 polüpeptiidi variantide, derivaatide, ekspressioonikassettide või sellest deriveeritud teiste järjestuste valmistamiseks. Leiutise mõnes teostuses seotakse leiutisekohased polüpeptiidid funktsionaalselt heteroloogilise promootoriga. Mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi kodeeriva nukleotiidse järjestuse võib sünteesida vanempolüpeptiidi aminohappelise järjestuse põhjal, sealhulgas aminohappelisest järjestusest SEQ ID NO: 1-7, muutes seejärel nukleotiidset järjestust, nii et oleks võimalik sisestada (ehk inkorporeerida või asendada) või eemaldada (ehk deleteerida või asendada) asjakohast aminohapet/aminohappeid. Nukleotiidset järjestust saab käepäraselt modifitseerida tavapärastel meetoditel kohtsuunatud mutageneesi teel. Nukleotiidse järjestuse võib ka keemiliselt sünteesida näiteks, kuid mitte ainult, oligonukleotiidide sünteesijat, milles luuakse oligonukleotiide soovikohase polüpeptiidi aminohappelise järjestuse põhjal ja eelistatavalt valitakse sellised koodonid, mis on eelistatud peremeesrakus, milles hakatakse tootma rekombinantset polüpeptiidi. Näiteks on võimalik sünteesid ja PCR meetodil, ligeerimise teel või ligeerimise ahelreaktsiooni teel koostada mitu väikest oligonukleotiidi, mis kodeerivad soovitud polüpeptiidi segmente. Vt nt Barany, et al., Proc. Natl. Acad Sci. 88: (1991); USA patent Käesoleva leiutise korral kasutatakse rekombinantse geenitehnoloogia (recombinant genetics) valdkonnas tavapäraseid meetodeid. Peamised allikad, milles käsitletakse käesoleva leiutise korral kasutatavaid üldmeetodeid, on näiteks Sambrook et al.,
67 Molecular Cloning, A Laboratory Manual (3rd ed. 01); Kriegler, Gene Transfer and Expression: A Laboratory Manual (1990); ja Current Protocols in Molecular Biology (Ausubel et al., eds., 1994)). Üldised allikad, milles kirjeldatakse molekulaarbioloogia meetodeid, on näiteks Berger and Kimmel, Guide to Molecular Cloning Techniques, Methods in Enzymology volume 2 Academic Press, Inc., San Diego, CA (Berger); Sambrook et al., Molecular Cloning - A Laboratory Manual (2nd Ed.), Vol. 1-3, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1989 ( Sambrook ) and Current Protocols in Molecular Biology, F.M. Ausubel et al., eds., Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (täiendatud 1999) ( Ausubel )). Nendes allikates kirjeldatakse mutageneesi, vektorite ja promootorite kasutamist ja käsitletakse paljusid teisi olulisi teemasid, mis on seotud näiteks, kuid mitte ainult, mittelooduslikke aminohappeid, ortogonaalseid trna-sid, ortogonaalseid süntetaase ja nende paare sisalduvate valkude tootmiseks vajalikke selektorkoodoneid sisaldavate geenide või polünukleotiidide loomisega. Leiutise korral kasutatakse mutageneesi erinevaid tüüpe erinevatel eesmärkidel, sealhulgas, kuid mitte ainult, uute süntetaaside või trnade tootmiseks, trna molekulide muteerimiseks, süntetaase kodeerivate polünukleotiidide muteerimiseks, trnade pankade loomiseks, süntetaaside pankade loomiseks, selektorkoodonite loomiseks, huvipakkuvas valgus või polüpeptiidis mittelooduslikke aminohappeid kodeerivate selektorkoodonite sisestamiseks. Selliste meetodite seas on näiteks, kuid mitte ainult kohtsuunatud, juhuslik punktmutagenees, homoloogiline rekombinatsioon, DNA lohistamine (shuffling) või teised rekursiivsed mutageneesi meetodid, kimääri konstrueerimine, uratsiili sisaldavatel matriitsidel põhineva mutagenees, oligonukleotiid-suunatud mutagenees, fosforotioaatmodifitseeritud DNA mutagenees, gapped dupleks-dna-l põhinev mutagenees või teised sellised meetodid, PCT-vahendatud mutagenees või nende mis tahes kombinatsioon. Teiste sobivate meetodite seas on point mismatch repair, repair-deficient peremeestüvede kasutamisel põhinev mutagenees, restriktsioonselektsioon ja restriktsioonpuhastamine, deletsioon-mutagenees, kogu geeni sünteesil põhinev mutagenees, double-strand break repair ja teised sarnased meetodid. Käesoleva leiutise puhul võib kasutada ka mutageneesi, mille puhul kasutatakse näiteks, kuid mitte ainult kimäärseid konstrukte. Ühes teostuses saab mutageneesi läbiviimisel lähtuda olemasolevast teabest loodusliku molekuli või modifitseeritud või muteeritud loodusliku molekuli, sealhulgas, kuid mitte ainult,
68 järjestuse, järjestuste võrdluse, füüsikaliste omaduste, sekundaarse, tertsiaarse või kvaternaarse struktuuri, kristallstruktuuri ja muude sarnaste omaduste kohta. Neid protseduure kirjeldatakse käesolevas dokumendis toodud tekstides ja näites. Lisateavet leidub järgmistes publikatsioonides ja neis viidatud allikates: Ling et al., Approaches to DNA mutagenesis: an overview, Anal Biochem. 4(2): (1997); Dale et al., Oligonucleotide-directed random mutagenesis using the phosphorothioate method, Methods Mol. Biol. 57: (1996); Smith, In vitro mutagenesis, Ann. Rev. Genet. 19: (1985); Botstein & Shortle, Strategies and applications of in vitro mutagenesis, Science 229: (1985); Carter, Site-directed mutagenesis, Biochem. J. 237:1-7 (1986); Kunkel, The efficiency of oligonucleotide directed mutagenesis, in Nucleic Acids & Molecular Biology (Eckstein, F. and Lilley, D.M.J. eds., Springer Verlag, Berlin) (1987); Kunkel, Rapid and efficient site-specific mutagenesis without phenotypic selection, Proc. Natl. Acad. Sci. USA 82: (1985); Kunkel et al., Rapid and efficient sitespecific mutagenesis without phenotypic selection, Methods in Enzymol. 4, (1987); Bass et al., Mutant Trp repressors with new DNA-binding specificities, Science 242: (1988); Zoller & Smith, Oligonucleotide-directed mutagenesis using M13- derived vectors: an efficient and general procedure for the production of point mutations in any DNA fragment, Nucleic Acids Res. : (1982); Zoller & Smith, Oligonucleotide-directed mutagenesis of DNA fragments cloned into M13 vectors, Methods in Enzymol. 0: (1983); Zoller & Smith, Oligonucleotide-directed mutagenesis: a simple method using two oligonucleotide primers and a single-stranded DNA template, Methods in Enzymol. 4: (1987); Taylor et al., The use of phosphorothioate-modified DNA in restriction enzyme reactions to prepare nicked DNA, Nucl. Acids Res. 13: (1985); Taylor et al., The rapid generation of oligonucleotide-directed mutations at high frequency using phosphorothioate-modified DNA, Nucl. Acids Res. 13: (1985); Nakamaye & Eckstein, Inhibition of restriction endonuclease Nci I cleavage by phosphorothioate groups and its application to oligonucleotide-directed mutagenesis, Nucl. Acids Res. 14: (1986); Sayers et al., 5 '-3' Exonucleases in phosphorothioate-based oligonucleotide-directed mutagenesis, Nucl. Acids Res. 16: (1988); Sayers et al., Strand specific cleavage of phosphorothioate-containing DNA by reaction with restriction endonucleases in the presence of ethidium bromide, (1988) Nucl. Acids Res. 16: ; Kramer et al., The gapped duplex DNA approach to oligonucleotide-directed mutation construction, Nucl.
69 Acids Res. 12: (1984); Kramer & Fritz Oligonucleotide-directed construction of mutations via gapped duplex DNA, Methods in Enzymol. 4: (1987); Kramer et al., Improved enzymatic in vitro reactions in the gapped duplex DNA approach to oligonucleotide-directed construction of mutations, Nucl. Acids Res. 16: 77 (1988); Fritz et al., Oligonucleotide-directed construction of mutations: a gapped duplex DNA procedure without enzymatic reactions in vitro, Nucl. Acids Res. 16: (1988); Kramer et al., Different base/base mismatches are corrected with different efficiencies by the methyl-directed DNA mismatch-repair system of E. coli, Cell 38: (1984); Carter et al., Improved oligonucleotide site-directed mutagenesis using M13 vectors, Nucl. Acids Res. 13: (1985); Carter, Improved oligonucleotide-directed mutagenesis using M13 vectors, Methods in Enzymol. 4: (1987); Eghtedarzadeh & Henikoff, Use of oligonucleotides to generate large deletions, Nucl. Acids Res. 14: 51 (1986); Wells et al., Importance of hydrogen-bond formation in stabilizing the transition state of subtilisin, Phil. Trans. R. Soc. Lond. A 317: (1986); Nambiar et al., Total synthesis and cloning of a gene coding for the ribonuclease S protein, Science 223: (1984); Sakmar and Khorana, Total synthesis and expression of a gene for the alphasubunit of bovine rod outer segment guanine nucleotide-binding protein (transducin), Nucl. Acids Res. 14: (1988); Wells et al., Cassette mutagenesis: an efficient method for generation of multiple mutations at defined sites, Gene 34:3-323 (1985); Grundström et al., Oligonucleotide-directed mutagenesis by microscale 'shot-gun' gene synthesis, Nucl. Acids Res. 13: (1985); Mandecki, Oligonucleotide-directed double-strand break repair in plasmids of Escherichia coli: a method for site-specific mutagenesis, Proc. Natl. Acad. Sci. USA, 83: (1986); Arnold, Protein engineering for unusual environments, Current Opinion in Biotechnology 4: (1993); Sieber, et al., Nature Biotechnology, 19: (01); W. P. C. Stemmer, Nature 370, (1994); and, I. A. Lorimer, I. Pastan, Nucleic Acids Res. 23, (1995). Lisateavet paljude eespool nimetatud meetodite kohta leidub allikas Methods in Enzymology Volume 4, milles kirjeldatakse ka kontrollvahendeid, mis kasulikud mitmesuguste mutageneesi meetodite korral veaotsingul. Oligonukleotiide ehk näiteks oligonukleotiide, mida kasutatakse käesoleva leiutise kohases mutageneesis ehk näiteks süntetaaside pankade muteerimisel või trna-de modifitseerimisel, sünteesitakse tavaliselt keemiliselt tahke faasi fosforamidiittriestri meetodil, mida kirjeldavad Beaucage and Caruthers, Tetrahedron Letts. 22(): , (1981), näiteks automaatse sünteesijaga,
70 5 68 mida kirjeldatakse allikas Needham-VanDevanter et al., Nucleic Acids Res., 12: (1984). Käesolev leiutis on seotud ka eukarüootsete peremeesrakkude, mitteeukarüootsete peremeesrakkude ja organismidega, mida kasutatakse mitteloodusliku aminohappe in vivo inkorporeerimiseks ortogonaalse trna/rs paaride abil. Peremeesrakke modifitseeritakse geneetiliselt (näiteks, kuid mitte ainult, transformeeritakse, modifitseeritakse tranduktsiooni teel või transfekteeritakse) leiutisekohaste polünukleotiididega või leiutisekohast polüpeptiidi sisaldavate konstruktidega, sealhulgas, kuid mitte ainult, leiutisekohase vektoriga, mis võib olla näiteks kloonimis- või ekspressioonivektor. Näiteks ortogonaalse trna, ortogonaalse trna süntetaasi ja derivatiseeritava valgu kodeerivad piirkonnad seotakse funktsionaalselt geenekspressiooni kontrollivate elementidega, mis on soovitud peremeesrakus funktsionaalsed. Vektor võib olla näiteks plasmiidi, kosmiidi, faagi, bakteri, viiruse, palja polünukleotiidi või konjugeeritud polünukleotiidi kujul. Vektorid sisestatakse rakkudesse ja/või mikroorganismidesse standardsetel meetoditel, sealhulgas elektroporeerimise teel (Fromm et al., Proc. Natl. Acad. Sci. USA 82, 5824 (1985)), viirusvektoriga nakatamise teel, suure kiirusega ballistilise penetratsiooni abil nukleiinhapet sisaldavate väikeste osakestega kas väikeste helmeste või osakestega matriitsis või pinnal (Klein et al., Nature 327, (1987)), ja/või sarnastel meetoditel. 30 Selliselt modifitseeritud peremeesrakke võib kasvatada tavapärastes söötmetes, mida on modifitseeritud vastavalt vajalikule tegevusele ehk skriinimiseks, promootorite aktiveerimiseks või transformantide selekteerimiseks. Selliseid rakke võib valikuliselt kasvatada transgeensetes organismides. Teised kasulikud allikad, sealhulgas, kuid mitte ainult, rakkude isoleerimise ja kasvatamise seisukohalt (nt järgnevaks nukleiinhapete isoleerimiseks) on näiteks Freshney (1994) Culture of Animal Cells, a Manual of Basic Technique, third edition, Wiley- Liss, New York ja neis viidatud allikad; Payne et al. (1992) Plant Cell and Tissue Culture in liquid Systems John Wiley & Sons, Inc. New York, NY; Gamborg and Phillips (eds.) (1995) Plant Cell, Tissue and Organ Culture; Fundamental Methods Springer Lab Manual, Springer-Verlag (Berlin Heidelberg New York) ja Atlas and Parks (eds.) The Handbook of Microbiological Media (1993) CRC Press, Boca Raton, FL. Praegu on tuntud mitu meetodit sihtnukleiinhapete sisestamiseks rakkudesse ja käesoleva leiutise korral võib kasutada mis tahes sellist meetodit. Selliste meetodite seas on: retsipientrakkude sulandamine DNA-d sisaldavate bakteriaalsete
71 protoplastidega, elektroporeerimine, mürskpommitamine ja viirusvektoritega nakatamine (mida käsitletakse põhjalikumalt allpool), jne. Leiutisekohaseid DNA konstrukte sisaldavate plasmiidide arvu saab kordistada bakterirakkudes. Baktereid kasvatatakse logaritmilise faasini ja bakterites olevad plasmiidid saab isoleerida erinevatel tehnika tasemes tuntud meetoditel (vt nt Sambrook). Lisaks on olemas kaubanduslikult kättesaadavad komplektid plasmiidide eraldamiseks bakteritest (vt nt EasyPrep, FlexiPrep, mõlemaid valmistab ettevõte Pharmacia Biotech; StrataClean, mida valmistab ettevõte Stratagene; ja QIAprep, mida valmistab ettevõte Qiagen). Isoleeritud ja puhastatud plasmiididest valmistatakse töötlemise teel teised plasmiidid, nendega transfekteeritakse rakke või need inkorporeeritakse organismide nakatamiseks sarnastesse vektoritesse. Tüüpilised vektorid sisaldavad transkriptsiooni ja translatsiooni terminaatoreid, transkriptsiooni ja translatsiooni initsieerimisjärjestusi ja promootoreid, mis on kasulikud konkreetse sihtnukleiinhappe ekspressiooni reguleerimisel. Vektorid sisaldavad valikuliselt tavapäraseid ekspressioonikassette, mis sisaldavad vähemalt ühte sõltumatut terminaatorjärjestust, järjestusi, mis võimaldavad kassetti replitseerida eukarüootides või prokarüootides või mõlemas (sealhulgas, kuid mitte ainult, süstikvektorid) ja selektsioonimarkereid nii prokarüootsete kui ka eukarüootsete süsteemide jaoks. Kasseti replitseerimiseks ja integreerimiseks prokarüootides, eukarüootides või mõlemas sobivad vektorid. Vt Gillam & Smith, Gene 8:81 (1979); Roberts, et al., Nature 328:731 (1987); Schneider, E., et al., Protein Expr. Purif. 6(1):- 14 (1995); Ausubel, Sambrook, Berger (kõik supra). Kloonimiseks sobivate bakterite ja bakteriofaagide kataloogi on andnud välja näiteks ATCC, nt ATCC välja antud kataloog nimega The ATCC Catalogue of Bacteria and Bacteriophage (1992) Gherna et al. (eds). Teisi tavapäraseid sekveneerimis- ja kloonimisprotseduure ning teisi molekulaarbioloogia aspekte ja nende aluseks olevaid teoreetilisi kaalutlusi käsitletakse ka allikas Watson et al. (1992) Recombinant DNA Second Edition Scientific American Books, NY. Lisaks saab peaaegu kõiki nukleiinhappeid (ja põhimõtteliselt kõiki märgistatud standardseid ja ka mittestandardseid nukleiinhappeid) tellida või lasta eritellimusel valmistada mitmesugustel ettevõtetel nagu the Midland Certified Reagent Company (Midland, TX, The Great American Gene Company (Ramona, CA, ExpressGen Inc. (Chicago, IL, Operon Technologies Inc. (Alameda, CA) ja paljudel teistel ettevõtetel.
72 SELEKTORKOODONID Leiutisekohased selektorkoodonid laiendavad valgu biosünteetilise mehhanismi geneetiliste koodonite raamistikku. Selektorkoodonite seas on näiteks, kuid mitte ainult, unikaalne kolmealuseline koodon, nonsenss-koodon nagu stoppkoodon, sealhulgas, kuid mitte ainult, amber-koodon (UAG), ookerkoodon või opaalkoodon (UGA), mittelooduslik koodon, neljaaluseline või rohkemaaluseline koodon, haruldane koodon või muu sarnane koodon. Asjatundjatele on ilmne, et soovitud geeni või polünukleotiidi võib sisestada erineva hulga selektorkoodoneid ehk ühte polüpeptiidi, mis kodeerib vähemalt osa FGF- 21 polüpeptiidist, võib sisestada näiteks, kuid mitte ainult, ühe või rohke, kaks või rohkem, kolm või rohkem, 4, 5, 6, 7, 8, 9, või rohkem selektorkoodonit. Ühes teostuses kasutatakse leiutisekohase meetodi korral ühe või mitme mitteloodusliku aminohappe in vivo sisestamiseks selektorkoodonit, mis on stoppkoodon. Näiteks luuakse O-tRNA, mis tunneb ära stoppkoodoni, sealhulgas, kuid mitte ainult, UAG ja aminoatsüülitakse O-RS abil soovitud mitteloodusliku aminohappega. Sellist O-tRNA-d ei tunne ära looduslikus peremeesrakus sisalduvad aminoatsüül-trna süntetaasid. Stoppkoodoni, näiteks, kuid mitte ainult, TAG-i saab sisestada huvipakkuvas polüpeptiidis huvipakkuvasse kohta tavapärase kohtsuunatud mutageneesi teel. Vt nt Sayers, J.R., et al. (1988), 5'-3' Exonucleases in phosphorothioate-based oligonucleotide-directed mutagenesis. Nucleic Acids Res, 16: Kui O-RS, O-tRNA ja huvipakkuvat polüpeptiidi kodeeriv polüpeptiid kombineeritakse in vivo, sisestatakse mittelooduslik aminohape vastusena UAG koodonile ja saadakse polüpeptiid, milles on konkreetses positsioonis mittelooduslik aminohape. Mittelooduslikke aminohappeid saab in vivo inkorporeerida nii, et selle selle tegemisel ei häirita oluliselt eukarüootset peremeesrakku. Näiteks kuna UAG koodoni supresseerimise ekeftiivsus sõltub O-tRNA, näiteks, kuid mitte ainult, amber suppressor trna, ja eukarüootse vabanemisfaktori (näiteks, kuid mitte ainult, erf) (mis seondub stoppkoodoniga ja initsieerib kasvava peptiidi vabanemise ribosoomist) vahelisest konkureerimisest, saab supresseerimise efektiivsust moduleerida näiteks, kuid mitte ainult, O-tRNA ja/või supressor-trna ekspressioonitaseme tõstmise teel. Mittelooduslikke aminohappeid saab kodeerida ka haruldaste koodonitega. Näiteks kui in vitro valkude sünteesi reaktsioonil langetada arginiini kontsentratsiooni, siis on haruldane arginiini koodon AGG osutunud efektiivseks Ala sisestamisel
73 71 Vt nt Ma et al., Biochemistry, 32:7939 (1993) Sellisel juhul konkureerib sünteetiline trna loodusliku trnaarg-ga, mida leidub väiksemal määral Escherichia coli bakterites. Mõned organismid ei kasuta kõiki triplettkoodoneid. Aminohapete sisestamiseks in vitro transkriptsiooni/translatsiooni ekstrakti on kasutatud määramata koodonit AGA Micrococcus luteus bakteris. Vt nt Kowal and Oliver, Nucl. Acid. Res., :4685 (1997). Leiutisekohaseid koostisosi saab luua nende kahe haruldase koodoniga in vivo. Selektorkoodonid hõlmavad ka laiendatud koodoneid, sealhulgas, kuid mitte ainult, nelja- või rohkemaaluselist koodonit nagu nelja-, viie-, kuueja rohkemaaluseline koodon. Neljaaluseliste koodonite seas on näiteks, kuid mitte ainult, AGGA, CUAG, UAGA, CCCU ja teised sarnased koodonid. Viiealuseliste koodonite seas on näiteks, kuid mitte ainult, AGGAC, CCCCU, CCCUC, CUAGA, CUACU, UAGGC ja teised sarnased koodonid. Leiutise üheks tunnuseks on raaminihke allasurumisel põhinevate laiendatud koodonite kasutamine. Nelja- või rohkemaaluselise koodoniga saab lisada samasse valku näiteks, kuid mitte ainult, ühe või mitu looduslikku aminohapet. Näiteks muteeritud O-tRNA-de, sealhulgas, kuid mitte ainult raaminihke supressortrna antikoodon silmuste juuresolekul. Teistes teostustes võivad antikoodoni silmused dekodeerida näiteks, kuid mitte ainult välja neljaaluselist koodonit, vähemalt viiealuselist koodonit või vähemalt kuue- või rohkemaaluselist koodonit. Kuna võib olla 6 neljaaluselist koodonit, saab nelja- või rohkemaaluselise koodoniga kodeerida samas rakus mitut mittelooduslikku aminohapet. Vt Anderson et al., (02) Exploring the Limits of Codon and Anticodon Size, Chemistry and Biology, 9: ; Magliery, (01) Expanding the Genetic Code: Selection of Efficient Suppressors of Four-base Codons and Identification of "Shifty" Four-base Codons with a Library Approach in Escherichia coli, J. Mol. Biol. 307: Mittelooduslike aminohapete sisestamisel in vitro biosünteesi meetoditel valkudesse on näiteks kasutatud neljaaluselisi koodoneid. Vt nt Ma et al., (1993) Biochemistry, 32:7939; and Hohsaka et al., (1999) J. Am. Chem. Soc., 121:34. 2-naftüülalaniini ja lüsiini NBD derivaadi samaaegseks lisamiseks streptavidiini koos kahe keemiliselt atsüülitud raaminihke supressor-trna-ga kasutati koodoneid CGGG ja AGGU. Vt nt Hohsaka et al., (1999) J. Am. Chem. Soc., 121: Ühes in vivo uuringus uurisid Moore et al. NCUA antikoodonitega trnaleu derivaatide võimet supresseerida UAGN koodoneid (N võib
74 72 olla U, A, G või C) ja avastasid, et nelik-uaga-d võib dekodeerida UCUA antikoodoniga trnaleu efektiivsusega 13 kuni 26% vähese dekodeerimisega 0 või -1 raamis Vt Moore et al., (00) J. Mol. Biol., 298:195. Leiutise ühes teostuses võib kasutada pikendatud koodoneid, mis põhinevad haruldastel koodonitel või nonsenss-koodonitel ja mis suudavad vähendada missense läbilugemist ja raaminihke supressiooni teistes mittesoovitavates kohtades. Antud süsteemi puhul võib selektorkoodon hõlmata ka ühte looduslikest kolmealuselistest koodonitest, kui endogeenne süsteem ei kasuta (või kasutab harva) looduslikku base koodonit. Siia kuulub näiteks süsteem, milles puudub trna, mis tunneb ära loodusliku kolmealuselise koodoni, ja/või süsteem, milles sisalduv kolmealuseline koodon on haruldane koodon. Selektorkoodonid sisaldavad valikuliselt mittelooduslikke aluspaare. Sellised mittelooduslikud aluspaarid laiendavad veelgi olemasolevat geneetilist teavet. Üks lisapaar suurendab triplett-koodonite arvu 64-lt 1-ni. Sellistele koodonitele on omane stabiilne ja selektiivne aluste paardumine, efektiivne ja väga usaldusväärne ensümaatiline lisamine polümeraasiga ja efektiivne jätkuv praimeri pikendamine pärast uue mitteloodusliku aluspaari sünteesi. Mittelooduslike aluspaare, mida saab kohandada leiutisekohaste meetodite ja kompositsioonide jaoks, kirjeldatakse näiteks allikas Hirao, et al., (02) An unnatural base pair for incorporating amino acid analogues into protein, Nature Biotechnology, : Vt Wu, Y., et al., (02) J. Am. Chem. Soc. 124: Teised asjakohased publikatsioonid on loetletud allpool. In vivo kasutamise korral on mitteloodusliku nukleosiidi membraan läbitav ja selle fosforüülimisel moodustub vastav trifosfaat. Lisandunud geneetiline teave on stabiilne ja ei hävi rakuliste ensüümide toimel. Varem püüdsid Brenner ja teised oma ära kasutada vesiniksidemete moodustumise mustrit, mis erineb mustritest, mis esinevad kanoonilistes Watson-Crick paarides, mille kõige märkimisväärsemaks näiteks on iso-c:iso-g paar. Vt nt Switzer et al., (1989) J. Am. Chem. Soc., 111:8322; and Piccirilli et al., (1990) Nature, 343:33; Kool, (00) Curr. Opin. Chem. Biol., 4:602. Sellised alused üldjuhul paarduvad mingil määral valesti ehk looduslike paaridega ja neid ei saa ensümaatiliselt replitseerida. Kool ja tema kolleegid näitasid, et alustevahelised hüdrofoobse pakkimise interaktsioonid võivad vesiniksidemete moodustumise asemel juhtida aluspaari moodustumist. Vt
75 Kool, (00) Curr. Opin. Chem. Biol., 4:602; and Guckian and Kool, (1998) Angew. Chem. Int. Ed. Engl., 36, 28. Selleks et töötada välja mittelooduslik aluspaar, mis vastab kõigile eespool toodud nõuetele, on Schultz, Romesberg ja nende kolleegid süstemaatiliselt sünteesinud ja uurinud seeriatena mittelooduslikke hüdrofoobseid aluseid. Praeguseks on tehtud kindlaks, et PICS:PICS paar on stabiilsem kui looduslikud aluspaarid ja seda saab efektiivselt sisestada DNA-sse Escherichia coli DNA polümeraasi I Klenow fragmendiga (KF). Vt nt McMinn et al., (1999) J. Am. Chem. Soc., 121:185-6; and Ogawa et al., (00) J. Am. Chem. Soc., 122:3274. KF abil saab efektiivselt ja bioloogilise funktsiooni tagamiseks piisavalt selektiivselt sünteesida 3MN:3MN paari. Vt nt Ogawa et al., (00) J. Am. Chem. Soc., 122:8803. Mõlemad alused aga toimivad edasise replikatsiooni korral ahela terminaatorina. Hiljuti töötati välja mutantne/mutantse DNA polümeraas, mida saab kasutada PICS paari replikatsioonil. Lisaks saab replitseerida 7AI paari. Vt nt Tae et al., (01) J. Am. Chem. Soc., 123:7439. Lisaks on loodud uudne metallaluspaar Dipic:Py, mis moodustab Cu(II) seondudes stabiilse paari. Vt Meggers et al., (00) J. Am. Chem. Soc., 122:714. Kuna pikendatud koodonid ja mittelooduslikud koodonid on olemuselt looduslike koodonite suhtes ortogonaalsed, saab leiutisekohaste meetodite korral selle omaduse varal luua nende ortogonaalseid trna-sid. Mitteloodusliku aminohappe saab sisestada soovitud polüpeptiidi ka translatsioonilise bypassing süsteemi abil. Translatsioonilise bypassing süsteemis lisatakse geeni suur järjestus, kuid seda ei transleerita valguks. Selles järjestuses on struktuur, mille ajel ribosoom hüppab üle selle järjestuse ja translatsioon jätkub insertsioonist allavoolu. Teatud teostustes kodeerib käesoleva leiutise kohaste meetodite ja/või kompositsioonide puhul huvi pakkuvat valku või polüpeptiidi (või osa sellest) nukleiinhape. Selline nukleiinhape sisaldab üldjuhul vähemalt ühte, vähemalt kahte, vähemalt kolme, vähemalt nelja, vähemalt viit, vähemalt kuut, vähemalt seitset, vähemalt kaheksat, vähemalt üheksat, kümmet või rohkemat selektorkoodonit. Huvipakkuvaid valke või polüpeptiide kodeerivaid geene saab muteerida asjatundjatele tuntud ja käesolevas dokumendis kirjeldatavatel meetoditel, mille abil sisestatakse mittelooduslikku aminohappesse näiteks üks või mitu selektorkoodonit. Huvipakkuva valgu nukleiinhape näiteks muteerimiseks lisatakse sellele üks või mitu selektorkoodonit, mille abil saab sisestada ühe või mitu mittelooduslikku aminohapet. Käesolev leiutis hõlmab kõiki selliseid variante, sealhulgas, kuid mitte ainult, variante, milles on vähemalt üks mittelooduslik aminohape. Käesolev leiutis hõlmab ka vastavaid nukleiinhappeid ehk mis tahes nukleiinhapet koos ühe või
76 5 74 mitme selektorkoodoniga, mis kodeerib ühte või mitut mittelooduslikku aminohapet. Huvipakkuvat valku nagu FGF-21 polüpeptiidi kodeerivaid nukleiinhappe molekule saab kergesti muteerida tsüsteiini sisestamiseks polüpeptiidi mis tahes soovikohasesse positsiooni. Tsüsteiini kasutatakse laialdaselt reaktsioonivõimeliste molekulide, vees lahustuvate polümeeride, valkude või mitmesuguste teiste molekulide sisestamiseks huvipakkuvasse valku. Meetodid, mille abil saab sisestada tsüsteiini polüpeptiidi soovikohasesse positsiooni, on asjatundjatele tuntud ja nende hulka kuuluvad USA patendis nr kirjeldatavad meetodid ja standardsed mutageneesi meetodid. IV. Mittelooduslikult kodeeritud aminohapped Käesoleva leiutise korral võib kasutada väga paljusid erinevaid mittelooduslikult kodeeritud aminohappeid. FGF-21 polüpeptiidi võib sisestada mis tahes hulga mittelooduslikult kodeeritud aminohappeid. Sisestatud mittelooduslikud aminohapped on üldjuhul tavapärase geneetiliselt kodeeritud aminohappe (ehk alaniini, arginiini, asparagiini, asparagiinhappe, tsüsteiini, glutamiini, glutamiinhappe, glütsiini, histidiini, isoleutsiini, leutsiini, lüsiini, metioniini, fenüülalaniini, proliini, seriini, treoniini, trüptofaani, türosiini ja valiini) suhtes põhimõtteliselt keemiliselt inertsed. Mõnes teostuses on mittelooduslikult kodeeritud aminohapetel kõrvalahelates funktsionaalrühmad, mis reageerivad efektiivselt ja selektiivselt funktsionaalrühmadega, mida ei leidu tavapärases aminohappes (sealhulgas, kuid mitte ainult, asido-, ketoon-, aldehüüd- ja aminooksürühmadega), ja moodustab stabiilsed konjugaadid. Näiteks FGF-21 polüpeptiidi, milles on funktsionaalset asidorühma sisaldav mittelooduslikult kodeeritud aminohape, saab panna reageerima polümeeriga (sealhulgas, kuid mitte ainult polü(etüleenglükooliga) või teise, alküünfragmenti sisaldava polüpeptiidiga, ja moodustada stabiilse konjugaadi Huisgeni [3+2] tsükloliitumise saaduse moodustamiseks funktsionaalsete asiid- ja alküünrühmade selektiivse reaktsiooni teel. Alfa-aminohappe üldstruktuur on järgmine (valem I):
77 Mittelooduslik aminohape on üldjuhul mis tahes struktuuriga, mis vastab eespool toodud valemile, milles R rühm on mis tahes asendaja, mis erineb looduslikus aminohappes kasutatavatest asendajatest ja sobib käesoleva leiutise puhul kasutamiseks. Kuna leiutisekohased mittelooduslikult kodeeritud aminohapped erinevad üldjuhul looduslikest aminohapetest ainult kõrvalahela struktuuri poolest, moodustavad mittelooduslikult kodeeritud aminohapped teiste aminohapetega, sealhulgas, kuid mitte ainult looduslike või mittelooduslikult kodeeritud aminohapetega amiidsidemed samal viisil nagu need moodustuvad looduslikes polüpeptiidides. Mittelooduslikult kodeeritud aminohapetel on aga kõrvalahelates rühmad, mis eristavad neid looduslikest aminohapetest. R rühm näiteks hõlmab valikuliselt alküül-, arüül-, atsüül-, keto-, asido-, hüdroksüül-, hüdrasiin, tsüano-, halo-, hüdrasiid-, alkenüül-, alkünüül-, eeter, tiool-, seleno-, sulfonüül-, boraat-, boronaat-, fosfo-, fosfono-, fosfiin-, heterotsüklilist, enoon-, imiin-, aldehüüd-, ester-, tiohappe, hüdroksüülamiin-, amino- või muud sarnast rühma või nende rühmade mis tahes kombinatsiooni. Teised huvipakkuvad mittelooduslikult kodeeritud aminohapped, mis võivad sobida käesoleva leiutise puhul kasutamiseks, on näiteks, kuid mitte ainult, aminohapped, mis sisaldavad valguse toimel aktiveeritavat ristsidet, spin-märgisega aminohapped, fluorestseeruvad aminohapped, metalli siduvad aminohapped, metalli sisaldavad aminohapped, radioaktiivsed aminohapped, uudsete funktsionaalrühmadega aminohapped, teiste molekulidega kovalentselt või mittekovalentselt interkateeruvad aminohapped, valguse poolt lõhustatavad/fotoisomeriseeritavad aminohapped, biotiini või biotiini analoogi sisaldavad aminohapped, glükosüülitud aminohapped nagu suhkruga asendatud seriin, teiste süsivesikutega modifitseeritud aminohapped, ketorühma sisaldavad aminohapped, polüetüleenglükooli või polüeetrit sisaldavad aminohapped, raske aatomiga asendatud aminohapped, keemiliselt lõhustatavad ja/või valguse toimel lõhustatavad aminohapped, aminohapped, millel on võrreldes looduslike aminohapetega pikemad kõrvalahelad, sealhulgas, kuid mitte ainult polüeetrid või pika ahelaga süsivesinikud, sealhulgas, kuid mitte ainult, rohkema umbes 5 või rohkema kui umbes süsinikuaatomiga süsivesinikud, süsinikuaatomiga seotud suhkur sisaldavad aminohapped, redoksaktiivsed aminohapped, amino-tiohapet sisaldavad aminohapped ja aminohapped,
78 5 76 mis sisaldavad ühte või mitut toksilist fragmenti. Mittelooduslikult kodeeritud aminohapped, mis võivad sobida käesoleva leiutise korral kasutamiseks ja on kasulikud vees lahustuvate polümeeridega läbiviidavate reaktsioonide korral, on näiteks, kuid mitte ainult, mittelooduslikult kodeeritud aminohapped, millel on reaktsioonivõimelised karbonüül-, aminooksü-, hüdrasiin-, hüdrasiid-, semikarbasiid-, asiid- ja alküünrühmi. Mõnes teostuses sisaldavad mittelooduslikult kodeeritud aminohapped sahhariidfragmenti. Selliste aminohapete seas on näiteks N-atsetüül-L-glükosaminüül-L-seriin, N-atsetüül-Lgalaktosaminüül-L-seriin, N-atsetüül-L-glükosaminüül-L-treoniin, N-atsetüül-Lglükosaminüül-L-asparagiin ja O-mannosaminüül-L-seriin. Selliste aminohapete näidete seas on aminohapped, milles aminohappe ja sahhariidi vaheline looduslik N- või O-side on asendatud kovalentse sidemega, mida tavaliselt ei leidu looduses, sealhulgas, kuid mitte ainult, alkeen-, oksiim-, tioeeter-, amiid- ja sarnase sidemega. Selliste aminohapete näited sisaldavad ka sahhariide, mida tavaliselt ei leidu looduslikes valkudes, nagu 2-deoksü-glükoos, 2-deoksügalaktoos ja teised sarnased ühendid. Paljud mittelooduslikult kodeeritud aminohapped on ka kaubanduslikult kättesaadavad näiteks ettevõtetelt Sigma-Aldrich (St. Louis, MO, USA), Novabiochem (ettevõtte EMD Biosciences allüksus Saksamaal Darmstadtis) või Peptech (Burlington, MA, USA). Aminohappeid, mis ei ole kaubanduslikult kättesaadavad, saab valikuliselt sünteesida käesolevas dokumendis kirjeldatavatel meetoditel või asjatundjatele tuntud standardsetel meetoditel. Teavet orgaanilise sünteesi meetodite kohta leidub näiteks allikates (1982, Second Edition, Willard Grant Press, Boston Mass.); Advanced Organic Chemistry by March (Third Edition, 1985, Wiley and Sons, New York); ja Advanced Organic Chemistry by Carey and Sundberg (Third Edition, Parts A and B, 1990, Plenum Press, New York). Vt ka USA patente nr ja Lisaks uudseid kõrvalahelaid sisaldavatele mittelooduslikele aminohapetele võib käesoleva leiutise puhul kasutada ka mittelooduslikke aminohappeid, millel on valikuliselt modifitseeritud selgroo struktuurid, sealhulgas, kuid mitte ainult, struktuurid valemiga II ja III:
79 milles Z üldjuhul hõlmab OH, NH 2, SH, NH-R' või S-R'; X ja Y, mis võivad olla samasugused või erinevad, üldjuhul on S või O, ning R ja R', mis on samasugused või erinevad, valitakse üldjuhul R rühma asendajate, mida loetletakse eespool seoses mittelooduslike aminohapetega, mille valem on I, või vesiniku seast. Leiutisekohastes mittelooduslikes aminohapetes on valikuliselt amino- või karboksüülrühmas asendused, nagu on näidatud valemites II ja III. Sellist tüüpi mittelooduslike aminohapete hulka kuuluvad näiteks, kuid mitte ainult, α-hüdroksühapped, α-tiohapped, α- aminotiokarboksülaadid, sealhulgas, kuid mitte ainult, kõrvalahelatega, mis vastavad tavapärasele aminohappele, või mittelooduslike kõrvalahelatega. α-süsiniku juures olevate asenduste seas on näiteks, kuid mitte ainult, L, D või α-α-diasendatud aminohapped nagu D-glutamaat, D-alaniin, D-metüül-O-türosiin, aminovõihape ja teised sarnased happed. Teiste struktuursete alternatiivide seas on tsüklilised aminohapped nagu proliini analoogid ja ka 3-, 4-,6-, 7-, 8- ja 9-lülilise tsükliga proliini analoogid, β ja γ aminohapped nagu asendatud β-alaniin- ja γ-aminovõihape. Paljud mittelooduslikud aminohapped põhinevad looduslikel aminohapetel nagu türosiin, glutamiin, fenüülalaniin ja teised sarnased aminohapped ja need sobivad käesoleva leiutise korral kasutamiseks. Türosiini analoogide seas on näiteks, kuid mitte ainult, para-asendatud türosiinid, orto-asendatud türosiinid ja meta-asendatud türosiinid, mille korral asendatud türosiin sisaldab näiteks, kuid mitte ainult keto- (sealhulgas, kuid mitte ainult, atsetüülrühma), bensoüül-, amino-, hüdrasiin-, hüdroksüamiin-, tiool-, karboksü-, isopropüül-, metüülrühma, C 6 - C sirge ahelaga või hargnenud ahelaga süsivesinik-, küülastundu või küllastumata süsivesinik-, O-metüül-, polüeeter-, nitro-, alkünüülrühma või muid sarnaseid üksusi. Lisaks on võimalikud ka mitme asendajaga arüültsüklid. Käesoleva leiutise puhul sobivate glutamiini analoogide seas on näiteks, kuid mitte ainult, α-hüdroksü derivaadid, γ-asendatud derivaadid, tsüklilised derivaadid ja amiid-asendatud glutamiini derivaadid. Käesoleva leiutise puhul sobivate fenüülalaniini analoogide seas on näiteks, kuid mitte ainult, para-asendatud, ortoasendatud ja meta-asendatud fenüülalaniinid, mille asendaja sisaldab näiteks, kuid mitte ainult, hüdroksü-, metoksü-, metüül-, allüül-, aldehüüd-, asido-, jodo-, bromo-, keto- (näiteks, kuid mitte ainult, atsetüülrühm), bensoüül-, alkünüül- või muud sarnast rühma.
80 Käesoleva leiutise puhul sobivate mittelooduslike aminohapete konkreetsete näidete seas on näiteks, kuid mitte ainult, ap-atsetüül-l- fenüülalaniin, O-metüül-L-türosiin, L-3-(2- naftüül)alaniin, 3-metüül-fenüülalaniin, O-4-allüül-L-türosiin, 4-propüül-L-türosiin, tri-oatsetüül-glcnacβ-seriin, L-Dopa, fluoritud fenüülalaniin, isopropüül-l-fenüülalaniin, p- asido-l-fenüülalaniin, p-atsüül-l-fenüülalaniin, p-bensoüül-l-fenüülalaniin, L-fosfoseriin, fosfonoseriin, fosfonotürosiin, p-jodo-fenüülalaniin, p-bromofenüülalaniin, p-amino-lfenüülalaniin, isopropüül-l-fenüülalaniin ja p-propargüüloksü-fenüülalaniin ja teised sarnased ühendid. Näiteid mitmesuguste käesoleva leiutise puhul sobivate mittelooduslike aminohapete struktuuridest leidub näiteks patendidokumendis WO 02/ pealkirjaga In vivo incorporation of unnatural amino acids. Vt ka Kiick et al., (02) Incorporation of azides into recombinant proteins for chemoselective modification by the Staudinger ligation, PNAS 99:19-24 for additional methionine analogs. Rahvusvahelises patenditaotluses PCT/US06/47822 pealkirjaga "Compositions Containing, Methods Involving, and Uses of Non-natural Amino Acids and Polypeptides," kirjeldatakse aromaatsete amiingragmentide, sealhulgas, kuid mitte ainult, p-aminofenüülalaniini redusteerivat alküülimist ja redutseerivat amiinimist. Ühe teostusega pakutakse FGF-21 polüpeptiidi kompositsioonid, mis sisaldavad mittelooduslikku aminohapet (nagu p- (propargüüloksü)-fenüülalaniin). Lisaks pakutakse ka mitmesugused kompositsioonid, mis sisaldavad p-(propargüüloksü)-fenüülalaniini ja näiteks, kuid mitte ainult, valke ja/või rakke. Ühe aspekti kohaselt sisaldab mittelooduslikku aminohapet p-(propargüüloksü)- fenüülalaniini sisaldav kompositsioon lisaks ka ortogonaalset trna-d. Mitteloodusliku aminohappe saab siduda (sealhulgas, kuid mitte ainult, kovalentselt) ortogonaalse trnaga, näiteks, kuid mitte ainult, siduda kovalentselt ortogonaalse trna-ga aminoatsüülsideme kaudu, siduda kovalentselt ortogonaalse trna terminaalse riboosi suhkru 3'OH või 2'OH-fragmendiga, jne. Keemilised fragmendid, mille kaudu saab valkudesse sisestada mittelooduslikke aminohappeid, pakuvad rea eeliseid ja manipulatsioone valkudega. Näiteks funktsionaalse ketorühma unikaalne reaktsioonivõime võimaldab valke selektiivselt modifitseerida in vitro ja in vivo mis tahes hulga hüdrasiini ja hüdroksüülamiini sisaldavate reagentidega. Raske aatomiga mittelooduslik aminohape näiteks võib olla kasulik for phasing X-ray structure data. Raskete aatomite kohtspetsiifiline sisestamine mittelooduslike aminohapete abil muudab rasketele
81 aatomitele positsioonide valimise selektiivseks ja paindlikuks. Fotoreaktiivsed mittelooduslikud aminohapped (sealhulgas, kuid mitte ainult, bensofenoon- ja arüülasiid (sealhulgas, kuid mitte ainult, fenüülasiid) kõrvalahelaga aminohapped võimaldavad valku efektiivselt in vivo ja in vitro fotoristsiduda. Fotoreaktiivsed mittelooduslikud aminohapped on näiteks, kuid mitte ainult, p-asido-fenüülalaniin ja p-bensoüülfenüülalaniin. Fotoreaktiivseid mittelooduslikke aminohappeid sisaldava valgu saab seejärel soovi korral ristsiduda ergastamisega fotoreaktiivset rühma pakkuva ajalise kontrolliga. Ühes näites saab mitteloodusliku aminohappe metüülrühma asendada lokaalse struktuuri ja dünaamika sondina isotüübiga märgitud, näiteks kuid mitte ainult, metüülrühmaga, kasutades selleks näiteks, kuid mitte ainult, tuumamagnetresonantsi ja vibratsioonspektroskoopiat. Funktsionaalsed alkünüül- või asidorühmad näiteks võimaldavad valke molekulidega selektiivselt modifitseerida [3+2] tsükloliitumise reaktsiooni teel. Polüpeptiidi amino-otsa lisatav mittelooduslik aminohape võib koosneda rühmast ehk mis tahes asendajast, mida ei leidu looduslikus aminohappes, ja teisest reaktsioonivõimelisest rühmast, mis ei ole tavaliselt α-aminohapetes sisalduv NH 2 rühm (vt valemit I). Sarnase mitteloodusliku aminohappe saab sisestada karboksüül-otsa teise reaktsioonivõimelise rühmaga, mis ei ole tavaliselt α-aminohapetes sisalduv COOH-rühm (vt valemit I). Leiutisekohaseid aminohappeid võib valida või disainida selliselt, et neil oleksid lisaomadused, mis puuduvad looduslikul aminohappel. Näiteks mittelooduslikke aminohappeid võib valikuliselt disainida või selekteerida, et modifitseerida näiteks valgu, millesse neid sisestatakse, bioloogilisi omadusi. Mitteloodusliku aminohappe sisestamise teel valku näiteks saab valikuliselt modifitseerida järgmisi omadusi: toksilisust, bioloogilist jaotumist, lahustuvust, stabiilsust, nt termilist, hüdrolüütilist, oksüdatiivset stabiilsust, vastupidavust ensümaatilise lagunemise suhtes ja teisi sarnaseid omadusi, puhastamise ja töötlemise käepärasust, stuktuuriomadusi, spektrokoopilisi omadusi, keemilisi ja/või fotokeemilisi omadusi, katalüütilist aktiivsust, redoks-potentsiaali, poolväärtusaega, võimet reageerida teiste molekulidega, nt kovalentselt või mittekovalentselt ja teisi sarnaseid omadusi. MITTELOODUSLIKE AMINOHAPETE STRUKTUUR JA SÜNTEESIMINE: KARBONÜÜL-, KARBONÜÜLISARNASED-, MASKEERITUD KARBONÜÜL- JA KAITSTUD KARBONÜÜLRÜHMAD NING HÜDROKSÜÜLAMIINRÜHMAD
82 80 Käesoleva leiutise mõne teostuse korral on FGF-21 seotud vees lahustuva polümeeri, nt PEG-ga, oksiimsideme kaudu. 5 Oksiinsidemete moodustamiseks sobivad mitmed mittelooduslikult kodeeritud aminohapete tüübid. Need hõlmavad muuhulgas, kuid mitte ainult, mittelooduslikult kodeeritud aminohappeid, mis sisaldavad karbonüül-, dikarbonüül- või hüdroksüülamiinrühma. Vastavaid aminohappeid on kirjeldatud USA patendipublikatsioonides nr US 06/01946, US 06/ ja US 06/ ning WO 06/ pealkirjaga Compositions containing, methods involving, and uses of non-natural amino acids and polypeptides. Mittelooduslikult kodeeritud aminohappeid on kirjeldatud ka USA patendis nr ja USA patendis nr , mis lisatakse viitena terviklikul kujul käesolevale dokumendile. Leiutise mõne teostuse korral kasutatakse FGF-21 polüpeptiide, mis on asendatud ühes positsioonis para-atsetüülfenüülalaniinaminohappega. p-atsetüül-(+/-)-fenüülalaniini ja m- atsetüül-(+/-)-fenüülalaniini sünteesimist on kirjeldatud allikas Zhang, Z., et al., Biochemistry 42: (03). Asjatundjad oskavad sarnasel viisil valmistada muid karbonüül- või dikarbonüülrühma sisaldavaid aminohappeid. Lisaks kirjeldatakse siin käsitletavate mittelooduslike aminohapete sünteesi mittepiiravaid näiteid patendi nr US 7,083,970 joonistel FIG 4, FIG ja FIG Elektrofiilse reaktisoonivõimelise rühmaga aminohapped võimaldavad mitmesuguseid reaktsioone molekulide sidumiseks mh. nukleofiilsete liitumisreaktsioonide kaudu. Sellised elektrofiilsed reaktsioonivõimelilised rühmad on muuhulgas karbonüülrühm (sealhulgas ketorühm ja dikarbonüülrühm), karbonüülisarnane rühm (mille reaktiivsus sarnaneb karbonüülrühma (sealhulgas ketorühma ja dikarbonüülrühma) omale ning mis sarnaneb struktuuri poolest karbonüülrühmale), maskeeritud karbonüülrühm (mida saab hõlpsasti muundada karbonüülrühmaks (sealhulgas ketorühmaks ja dikarbonüülrühmaks)), või kaitstud karbonüülrühm (mille reaktiivsus sarnaneb karbonüülrühma (sealhulgas ketorühma ja dikarbonüülrühma) omale kaitsva rühma eemaldamisel). Taolised aminohapped hõlmavad aminohappeid, mille struktuur vastab valemile (IV).
83 81 milles: 5 A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen- või asendatud alküleen)-, -S-, -S-(alküleen- või asendatud alküleen)-, -S(O) k - rühm, milles k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)-(alküleen- või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen- või asendatud alküleen)-, -N(R')-, -NR'-(alküleen- või asendatud alküleen)-, -C(O)N(R')-, -CON(R')- (alküleen- või asendatud alküleen)-, -CSN(R')-, -CSN(R')-(alküleen- või asendatud alküleen)-, -N(R')CO-(alküleen- või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N-N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')-N(R')-, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm; J on või
84 82 R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; iga R" on sõltumatult H, alküül-, asendatud alküül- või kaitserühm, või enam kui ühe R" rühma olemasolu korral moodustavad kaks R" rühma valikuliselt heterotsükloalküülrühma; 5 R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; R 3 ja R 4 on sõltumatult H, halogeen-, madalam alküül- või asendatud madalam alküülrühm, või R 3 ja R 4 või kaks R 3 rühma moodustavad valikuliselt tsükloalküül- või heterotsükloalküülrühma; või -A-B-J-R rühmad moodustavad koos bitsüklilise või tritsüklilise tsükloalküül- või heterotsükloalküülrühma, mille koosseisu kuulub vähemalt üks karbonüülrühm, sealhulgas dikarbonüülrühm, kaitstud karbonüülrühm, sealhulgas kaitstud dikarbonüülrühm, või maskeeritud karbonüülrühm, sealhulgas maskeeritud dikarbonüülrühm; või -J-R rühm moodustab koos monotsüklilise või bitsüklilise tsükloalküül- või heterotsükloalküülrühma, mille koosseisu kuulub vähemalt üks karbonüülrühm, sealhulgas dikarbonüülrühm, kaitstud karbonüülrühm, sealhulgas kaitstud dikarbonüülrühm, või maskeeritud karbonüülrühm, sealhulgas maskeeritud dikarbonüülrühm; tingimusel, et kui A on fenüleenrühm ja iga R 3 on H, on B olemas; ning et kui A on -(CH 2 ) 4 - ja iga R 3 on H, ei ole B -NHC(O)(CH 2 CH 2 )-; ning et kui A ja B puuduvad ning iga R 3 on H, ei ole R metüülrühm. Lisaks on kaasatud valemi (V) struktuuriga:
85 milles: 83 5 A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen või asendatud alküleen)-, -S-, -S-(alküleen või asendatud alküleen)-, -S(O) k - kus k on 1, 2, või 3, -S(O) k (alküleen või asendatud alküleen)-, -C(O)-, -C(O)-(alküleen või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen või asendatud alküleen)-, -N(R')-, -NR'- (alküleen või asendatud alküleen)-, -C(O)N(R')-, -CON(R')-(alküleen või asendatud alküleen)-, -CSN(R')-, -CSN(R')-(alküleen või asendatud alküleen)-, -N(R')CO-(alküleen või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N- N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')-N(R')-, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; tingimusel, et kui A on fenüleen, on B olemas; ning et kui A on -(CH 2 ) 4 -, ei ole B -NHC(O)(CH 2 CH 2 )-; ning et kui A ja B puuduvad, ei ole R metüülrühm. Lisaks on kaasatud aminohapped valemi (VI) struktuuriga:
86 84 5 milles: B on linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen, asendatud madalam alküleen, madalam alkenüleen, asendatud madalam alkenüleen, madalam heteroalküleen, asendatud madalam heteroalküleen, -O-, -O-(alküleen või asendatud alküleen)-, -S-, -S-(alküleen või asendatud alküleen -S(O) k - kus k on 1, 2, või 3, -S(O) k (alküleen või asendatud alküleen)-, -C(O)-, -C(O)-(alküleen või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen või asendatud alküleen)-, -N(R')-, -NR'-(alküleen või asendatud alküleen)-, -C(O)N(R')-, -CON(R')-(alküleen või asendatud alküleen)-, -CSN(R')-, -CSN(R')-(alküleen või asendatud alküleen)-, -N(R')CO-(alküleen või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N-N=, -C(R') 2 -N=Nja -C(R') 2 -N(R')-N(R')-, kus iga R' on sõltumatult H, alküül- või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; iga R a valitakse sõltumatult rühmast, kuhu kuuluvad H, halogeen-, alküül-, asendatud alküül-, -N(R') 2, -C(O) k R'-rühm, milles k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k R', milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm. Lisaks on kaasatud järgnevad aminohapped:

87 85 kusjuures taolised ühendid on valikuliselt aminokaitserühm, karboksüülkaitse või nende sool. Lisaks on võimalik ükskõik millise järgneva mitteloodusliku aminohappe kaasamine mittelooduslikku aminohappe polüpeptiidi. 5 Lisaks on kaasatud järgnevad aminohapped valemi (VII) struktuuriga: milles: B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen- või asendatud alküleen-, -S-, -S-(alküleen- või asendatud alküleen)-, -S(O) k - kus k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)-(alküleenvõi asendatud alküleen)-, -C(S)-, -C(S)-(alküleen- või asendatud alküleen)-, -N(R')-, -NR'- (alküleen- või asendatud alküleen)-, -C(O)N(R')-, -CON(R')-(alküleen- või asendatud alküleen-, -CSN(R')-, -CSN(R')-(alküleen- või asendatud alküleen)-, -N(R')CO-(alküleenvõi asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N- N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')-N(R')-, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning
 2, -C(O) k R'-rühm, milles k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k")
-.")

88 86 R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; 5 iga R a valitakse sõltumatult rühmast, kuhu kuuluvad H, halogeen-, alküül-, asendatud alküül-, -N(R') 2, -C(O) k R'-rühm, milles k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k R', milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm ning n on 0 kuni 8; tingimusel, et kui A on -(CH 2 ) 4 -, ei ole B -NHC(O)(CH 2 CH 2 )-. Lisaks on kaasatud järgnevad aminohapped: kusjuures taolised ühendid on valikuliselt aminokaitstud, valikuliselt karboksüülkaitstud, valikuliselt aminokaitstud ja karboksüülkaitstud, või nende sool. Lisaks võivad need mittelooduslikud aminohapped ja mis tahes järgnevatest mittelooduslikest aminohapetest olla kaasatud mittelooduslikku aminohappe polüpeptiidi. Lisaks on kaasatud järgnevad aminohapped valemi (VIII) struktuuriga:
89 milles: 87 5 A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen- või asendatud alküleen-, -S-, -S-(alküleen- või asendatud alküleen)-, -S(O) k - rühm, milles k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)- (alküleen- või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen- või asendatud alküleen)-, -N(R')-, -NR'-(alküleen- või asendatud alküleen)-, -C(O)N(R')-, -CON(R')-(alküleen või asendatud alküleen)-, -CSN(R')-, -CSN(R')-(alküleen või asendatud alküleen)-, -N(R')CO- (alküleen või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N- N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')-N(R')-, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (IX):
90 88 5 B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen või asendatud alküleen)-, -S-, -S-(alküleen või asendatud alküleen)-, -S(O) k - rühm, milles k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)- (alküleen- või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen- või asendatud alküleen)-, -N(R')-, -NR'-(alküleen- või asendatud alküleen)-, -C(O)N(R')-, -CON(R')-(alküleen- või asendatud alküleen)-, -CSN(R')-, -CSN(R')-(alküleen- või asendatud alküleen)-, -N(R')CO-(alküleen või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N-N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')-N(R')-rühm, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; kusjuures iga R a valitakse sõltumatult rühmast, kuhu kuuluvad H, halogeen-, alküül-, asendatud alküül-, -N(R') 2, -C(O) k R' kus k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k R' rühm, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm. Lisaks on kaasatud järgnevad aminohapped:

91 89 5 kusjuures sellised ühendid on valikuliselt aminokaitstud, valikuliselt karboksüülkaitstud, valikuliselt aminokaitstud ja karboksüülkaitstud, või nende sool. Lisaks võivad need mittelooduslikud aminohapped ja mis tahes järgnevatest mittelooduslikest aminohapetest olla kaasatud mittelooduslikku aminohappe polüpeptiidi. Lisaks on kaasatud järgnevad aminohapped struktuuriga, mis vastab valemile (X) : kusjuures B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen- või asendatud alküleen-, -S-, -S-(alküleen- või asendatud alküleen-, -S(O) k - rühm, milles k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)-(alküleen või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen- või asendatud alküleen)-, -N(R')-, -NR'-(alküleen- või asendatud alküleen, -C(O)N(R')-, -CON(R')-(alküleen- või asendatud alküleen)-, -CSN(R')-, -CSN(R')-(alküleen- või asendatud alküleen)-, -N(R')CO-(alküleen- või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N-N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')-N(R')-rühm, milles iga R' on sõltumatult H, alküül või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning
92 90 R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; 5 iga R a valitakse sõltumatult rühmast, kuhu kuuluvad H, halogeen-, alküül-, asendatud alküül-, -N(R') 2, -C(O) k R'-rühm, milles k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k R', milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm ning n on 0 kuni 8. Lisaks on kaasatud järgnevad aminohapped: ja kusjuures taolised ühendid on valikuliselt aminokaitstud, valikuliselt karboksüülkaitstud, valikuliselt aminokaitstud ja karboksüülkaitstud, või nende sool. Lisaks võivad need mittelooduslikud aminohapped ja mis tahes järgnevatest mittelooduslikest aminohapetest olla kaasatud mittelooduslikku aminohappe polüpeptiidi. Lisaks monokarbonüülstruktuuridele võivad siinkirjeldatud mittelooduslikud aminohapped hõlmata rühmi nagu dikarbonüül-, dikarbonüülisarnased, maskeeritud dikarbonüül- ja kaitstud dikarbonüülrühmad. Näiteks on kaasatud järgnevad aminohapped valemi (XI) struktuuriga:
93 5 91 kusjuures A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen- või asendatud alküleen)-, -S-, -S-(alküleen- või asendatud alküleen)-, -S(O) k - rühm, milles k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)-(alküleen- või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen- või asendatud alküleen)-, -N(R')-, -NR'-(alküleen- või asendatud alküleen)-, -C(O)N(R')-, -CON(R')- (alküleen- või asendatud alküleen, -CSN(R')-, -CSN(R')-(alküleen- või asendatud alküleen)-, -N(R')CO-(alküleen- või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N-N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')-N(R')-, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XII):
94 5 92 B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen- või asendatud alküleen)-, -S-, -S-(alküleen- või asendatud alküleen, -S(O) k - rühm, milles k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)- (alküleen- või asendatud alküleen)-, -C(S)-, -C(S)-(alküleen- või asendatud alküleen)-, -N(R')-, -NR'-(alküleen- või asendatud alküleen)-, -C(O)N(R')-, -CON(R')-(alküleen- või asendatud alküleen)-, -CSN(R')-, -CSN(R')-(alküleen- või asendatud alküleen)-, -N(R')CO-(alküleen- või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R')-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N-N=, -C(R') 2 -M=N- ja -C(R') 2 -N(R')-N(R')- rühm, milles iga R' on sõltumatult kas H, alküül- või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; kusjuures iga R a valitakse sõltumatult rühmast, kuhu kuuluvad H, halogeen-, alküül-, asendatud alküül-, -N(R') 2, -C(O) k R' rühm, milles k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k R'-rühm, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm. Lisaks on kaasatud järgnevad aminohapped: kusjuures sellised ühendid on valikuliselt aminokaitstud, valikuliselt karboksüülkaitstud, valikuliselt aminokaitstud ja karboksüülkaitstud, või nende sool. Lisaks võivad need mittelooduslikud aminohapped ja mis tahes järgnevatest mittelooduslikest aminohapetest olla kaasatud mittelooduslikku aminohappe polüpeptiidi.
95 93 Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XIII): 5 kusjuures B on valikuline, olles olemasolu korral linker, mis on valitud rühmast, kuhu kuuluvad madalam alküleen-, asendatud madalam alküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, madalam heteroalküleen-, asendatud madalam heteroalküleen-, -O-, -O-(alküleen- või asendatud alküleen)-, -S-, -S-(alküleen- või asendatud alküleen)-, -S(O) k rühm, milles k on 1, 2, või 3, -S(O) k (alküleen- või asendatud alküleen)-, -C(O)-, -C(O)-(alküleen- või asendatud alküleen)-, -C(S)-, -C(S)- (alküleen- või asendatud alküleen)-, -N(R')-, -NR'-(alküleen- või asendatud alküleen, -C(O)N(R')-, -CON(R')-(alküleen- või asendatud alküleen)-, -CSN(R')-, -CSN(R')- (alküleen või asendatud alküleen)-, -N(R')CO-(alküleen või asendatud alküleen)-, -N(R')C(O)O-, -S(O) k N(R')-, -N(R')C(O)N(R')-, -N(R')C(S)N(R')-, -N(R')S(O) k N(R)-, -N(R')-N=, -C(R')=N-, -C(R')=N-N(R')-, -C(R')=N-N=, -C(R') 2 -N=N- ja -C(R') 2 -N(R')- N(R')-rühm, milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; iga R a valitakse sõltumatult rühmast, kuhu kuuluvad H, halogeen-, alküül-, asendatud alküül-, -N(R') 2, -C(O) k R'-rühm, milles k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k R', milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm ning n on 0 kuni 8. Lisaks on kaasatud järgnevad aminohapped:



96 94 5 kusjuures sellised ühendid on valikuliselt aminokaitstud, valikuliselt karboksüülkaitstud, valikuliselt aminokaitstud ja karboksüülkaitstud, või nende sool. Lisaks võivad need mittelooduslikud aminohapped ja mis tahes järgnevatest mittelooduslikest aminohapetest olla kaasatud mittelooduslikku aminohappe polüpeptiidi. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XIV): milles: A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm;
97 95 R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning 5 R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; X 1 on C, S, või S(O) ja L on alküleen-, asendatud alküleen-, N(R')(alküleen)- või N(R')(asendatud alküleen)rühm, milles R' on H, alküül-, asendatud alküül-, tsükloalküülvõi asendatud tsükloalküülrühm. Lisaks on kaasatud järgnevad aminohapped, mille struktuur on valemiga (XIV-A): kus: A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid;
98 96 L on alküleen-, asendatud alküleen-, N(R')(alküleen) või N(R')(asendatud alküleen)rühm, milles R' on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm. Lisaks on kaasatud järgnevad aminohapped valemi (XIV-B) struktuuriga: 5 milles: A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; L on alküleen-, asendatud alküleen-, N(R')(alküleen) või N(R')(asendatud alküleen)rühm, milles R' on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XV):
99 97 milles: 5 A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; X 1 on C, S, või S(O); ja n on 0, 1, 2, 3, 4 või 5; ning iga R 8 ja R 9 igas CR 8 R 9 rühmas valitakse sõltumatult rühmast, kuhu kuuluvad H, alkoksü-, alküülamiin-, halogeen-, alküül-, arüülrühm või mis tahes R 8 ja R 9 võib koos moodustada =O või tsükloalküülrühma või mis tahes külgnevad R 8 rühmad võivad koos moodustada tsükloalküülrühma. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XV-A):
100 milles: A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; 2. R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; 3. R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning 4. R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; 5. n on 0, 1, 2, 3, 4 või 5; ning iga R 8 ja R 9 igal CR 8 R 9 rühmal valitakse sõltumatult rühmast, kuhu kuuluvad H, alkoksü-, alküülamiin-, halogeen-, alküül-, arüülvõi mis tahes R 8 ja R 9 võivad koos moodustada =O või tsükloalküülrühma või mis tahes külgnevad R 8 rühmad võivad koos moodustada tsükloalküülrühma. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XV-B): milles: A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud
101 99 heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; 5 R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; n on 0, 1, 2, 3, 4 või 5; ning iga R 8 ja R 9 igas CR 8 R 9 rühmas valitakse sõltumatult rühmast, kuhu kuuluvad H, alkoksü-, alküülamiin-, halogeen-, alküül-, arüül- või mis tahes R 8 ja R 9 võivad koos moodustada =O või tsükloalküülrühma või mis tahes külgnevad R 8 rühmad võivad koos moodustada tsükloalküülrühma. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XVI): milles: A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm;
102 0 R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; 5 X 1 on C, S, või S(O) ja L on alküleen-, asendatud alküleen-, N(R')(alküleen)- või N(R')(asendatud alküleen)rühm, milles R' on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XVI-A): milles: A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; L on alküleen-, asendatud alküleen-, N(R')(alküleen)- või N(R')(asendatud alküleen)rühm, milles R' on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm.
103 1 Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XVI-B): milles: 5 A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; R on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; L on alküleen-, asendatud alküleen-, N(R')(alküleen) või N(R')(asendatud alküleen)rühm, milles R' on H, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm. Lisaks on kaasatud aminohapped, mille struktuur vastab valemile (XVII): milles:
104 5 2 A on valikuline, olles olemasolu korral madalam alküleen-, asendatud madalam alküleen-, madalam tsükloalküleen-, asendatud madalam tsükloalküleen-, madalam alkenüleen-, asendatud madalam alkenüleen-, alkünüleen-, madalam heteroalküleen-, asendatud heteroalküleen-, madalam heterotsükloalküleen-, asendatud madalam heterotsükloalküleen-, arüleen-, asendatud arüleen-, heteroarüleen-, asendatud heteroarüleen-, alkarüleen-, asendatud alkarüleen-, aralküleen- või asendatud aralküleenrühm; M on -C(R 3 )-, milles (a) osutab sidumisele rühmaga A ja (b) sidumisele vastavate karbonüülrühmadega, R 3 ja R 4 valitakse sõltumatult järgnevate seast: H, halogeen-, alküül-, asendatud alküül-, või tsükloalküül- või asendatud tsükloalküülrühm, või R 3 ja R 4 või kaks R 3 rühma või kaks R 4 rühma moodustavad valikuliselt tsükloalküül- või heterotsükloalküülrühma; R on H, halogeen-, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; T 3 on side, C(R)(R), O, või S ja R on H, halogeen-, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid.
105 3 Lisaks on kaasatud aminohapped, mille struktuur vastab valemile (XVIII): kusjuures: M on -C(R 3 )-, 5 või milles (a) osutab sidumisele rühmaga A ja (b) sidumisele vastavate karbonüülrühmadega, R 3 ja R 4 valitakse sõltumatult järgnevate seast: H, halogeen-, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm, või R 3 ja R 4 või kaks R 3 rühma või kaks R 4 rühma moodustavad valikuliselt tsükloalküül- või heterotsükloalküülrühma; R on H, halogeen-, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; T 3 on side, C(R)(R), O, või S ja R on H, halogeen-, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; R 1 on valikuline, olles olemasolu korral H, aminokaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; ning
106 4 R 2 on valikuline, olles olemasolu korral OH, esterkaitserühm, vaik, aminohape, polüpeptiid või polünukleotiid; 5 iga R a valitakse sõltumatult rühmast, kuhu kuuluvad H, halogeen-, alküül-, asendatud alküül-, -N(R') 2, -C(O) k R'-rühm, milles k on 1, 2, või 3, -C(O)N(R') 2, -OR' ja -S(O) k R', milles iga R' on sõltumatult H, alküül- või asendatud alküülrühm. Lisaks on kaasatud aminohapped, mille struktuur vastab valemile (XIX): milles: R on H, halogeen-, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm; ning T 3 on O või S. Lisaks on kaasatud aminohapped, mille struktuur vastab valemile (XX): milles: R on H, halogeen-, alküül-, asendatud alküül-, tsükloalküül- või asendatud tsükloalküülrühm. Lisaks on kaasatud järgnevad aminohapped, mille struktuur vastab valemile (XXI):
107 5 5 Mõne teostuse korral modifitseeritakse mittelooduslikku aminohapet sisaldavat polüpeptiidi keemiliselt, et saada reaktsioonivõimeline karbonüül- või dikarbonüülfunktsionaalrühm. Näiteks saab konjugatsioonireaktsioonide jaoks kasulikku funktsionaalse aldehüüdrühma luua funktsionaalrühmast, millel on külgnevad amino- ja hüdroksüülrühmad. Kui bioaktiivne molekul on polüpeptiid, saab näiteks N-otsa seriini või treoniini (mis võib alaliselt kohal olla või avalduda keemilise või ensümaatilise lõhustumise tulemusena) kasutada funktsionaalse aldehüüdrühma tekitamiseks leebe oksüdatiivse lõhustumise tingimustes perjodaadi abil. Vt nt Gaertner, et. al., Bioconjug. Chem. 3: (1992); Geoghegan, K. & Stroh, J., Bioconjug. Chem. 3: (1992); Gaertner et al., J. Biol. Chem. 269: (1994). Tehnika tasemes tuntud meetodid piirduvad peptiidi või valgu N-otsas paikneva aminohappega. Käesoleva leiutise korral saab mittelooduslikku aminohapet, mis kannab külgnevaid hüdroksüül- ja aminorühmi, kaasata polüpeptiidi koosseisu maskeeritud funktsionaalse aldehüüdrühmana. Näiteks kannab 5-hüdroksülüsiin hüdroksüülrühma, mis külgneb epsilon-amiiniga. Aldehüüdi loomiseks tarvilikud reaktsioonitingimused hõlmavad tüüpiliselt naatriumperjodaadi molaarse ülejäägi lisamist pehmete tingimuste juures oksüdatsiooni vältimiseks muudel kohtadel polüpeptiidi piires. Oksüdatsioonireaktsiooni ph on tüüpiliselt ligikaudu 7.0. Tüüpiline reaktsioon hõlmab naatriummetaperjodaadi u 1,5 molaarse ülejäägi lisamist polüpeptiidi puhverdatud lahusele, millele järgneb ligikaudu -minutiline inkubeerimine pimeduses. Vt nt patenti USA patenti nr Funktsionaalse karbonüül- või dikarbonüülrühma võib panna leebetes tingimustes vesilahuses selektiivselt reageerima hüdroksüülamiini sisaldava reagendiga, mille tulemusel moodustub vastav oksiimside, mis on füsioloogilistes tingimustes stabiilne. Vt nt Jencks, W. P., J. Am. Chem. Soc. 81, (1959); Shao, J. and Tam, J. P., J. Am. Chem. Soc. 117: (1995). Lisaks võimaldab karbonüül- või dikarbonüülrühma unikaalne reaktsioonivõime selektiivset modifitseerimist muude aminohappe kõrvalahelate juuresolekul. Vt nt Cornish, V. W., et al., J. Am. Chem. Soc. 118:80-81 (1996);
108 6 Geoghegan, K. F. & Stroh, J. G., Bioconjug. Chem. 3: (1992); Mahal, L. K., et al., Science 276: (1997). Mittelooduslike aminohapete struktuur ja süntees: hüdroksüülamiini sisaldavad aminohapped 5 USA eelpatenditaotlus nr. US 60/638,418 on tervikuna kaasatud viitena. Seega kehtivad ajutise patenditaotluse nr. US 60/638,418 V jaotise (pealkirjaga Non-natural Amino Acids ) osas B (pealkirjaga Structure and Synthesis of Non-Natural Amino Acids: Hydroxylamine-Containing Amino Acids esitatud avastused täies ulatuses meetodite, kompositsioonide (sealhulgas valemid I-XXXV), tehnikate ja strateegiate suhtes siinkirjeldatud mittelooduslike aminohapete, mittelooduslike aminohapete polüpeptiidide ning modifitseeritud mittelooduslike aminohapete polüpeptiidide valmistamiseks, puhastamiseks, iseloomustamiseks ja kasutamiseks, võrdväärselt sellega, kui kõnealused leiutised oleksid siin täies mahus esitletud. USA patenditaotlused nr US 06/01946, US 06/ ja US 06/ ning WO 06/ pealkirjaga Compositions containing, methods involving, and uses of non-natural amino acids and polypeptides on samuti tervikuna kaasatud viitena. MITTELOODUSLIKE AMINOHAPETE KEEMILINE SÜNTEESIMINE 30 Paljud mittelooduslikud aminohapped, mis sobivad käesoleva leiutise raames kasutamiseks, on kaubanduslikult saadaval, neid turustavad näiteks ettevõtted Sigma (USA) või Aldrich (Milwaukee, WI, USA). Mittelooduslikke aminohappeid, mis pole kaubanduslikult saadaval, võib valikuliselt sünteesida käesolevas dokumendis või erinevates allikates kirjeldatud viisil või asjatundjatele teadaolevatel standardsetel meetoditel. Orgaanilise sünteesi tehnikaid käsitletakse näiteks allikates Organic Chemistry, Fessendon ja Fessendon, (1982, Second Edition, Willard Grant Press, Boston Mass.); Advanced Organic Chemistry, March (Third Edition, 1985, Wiley and Sons, New York); ning Advanced Organic Chemistry, Carey ja Sundberg (Third Edition, Parts A and B, 1990, Plenum Press, New York). Teised publikatsioonid, milles kirjeldatakse mittelooduslike aminohapete sünteesimist, on nt WO 02/ pealkirjaga In vivo incorporation of Unnatural Amino Acids Matsoukas et al., (1995) J. Med. Chem., 38, ; King, F.E. & Kidd, D.A.A. (1949) A New Synthesis of Glutamine and of γ- Dipeptides of Glutamic Acid from Phthylated Intermediates. J. Chem. Soc., ;
109 5 7 Friedman, O.M. & Chatterrji, R. (1959) Synthesis of Derivatives of Glutamine as Model Substrates for Anti-Tumor Agents. J. Am. Chem. Soc. 81, ; Craig, J.C. et al. (1988) Absolute Configuration of the Enantiomers of 7-Chloro-4 [[4-(diethylamino)-1- methylbutyl]amino]quinoline (Chloroquine). J. Org. Chem. 53, ; Azoulay, M., Vilmont, M. & Frappier, F. (1991) Glutamine analogues as Potential Antimalarials, Eur. J. Med. Chem. 26, 1-5; Koskinen, A.M.P. & Rapoport, H. (1989) Synthesis of 4- Substituted Prolines as Conformationally Constrained Amino Acid Analogues. J. Org. Chem. 54, ; Christie, B.D. & Rapoport, H. (1985) Synthesis of Optically Pure Pipecolates from L-Asparagine. Application to the Total Synthesis of (+)-Apovincamine through Amino Acid Decarbonylation and Iminium Ion Cyclization. J. Org. Chem. 50: ; Barton et al., (1987) Synthesis of Novel alpha-amino-acids and Derivatives Using Radical Chemistry: Synthesis of L- and D-alpha-Amino-Adipic Acids, L-alpha-aminopimelic Acid and Appropriate Unsaturated Derivatives. Tetrahedron 43: ; ning Subasinghe et al., (1992) Quisqualic acid analogues: synthesis of beta-heterocyclic 2-aminopropanoic acid derivatives and their activity at a novel quisqualate-sensitized site. J. Med. Chem. 35: Vt ka USA patenditaotlust nr 04/ pealkirjaga Protein Arrays. A. Reaktsioonivõimelised karbonüülrühmad Reaktsioonivõimelise karbonüülrühmaga aminohapped võimaldavad mitmesuguseid reaktsioone molekulide (muuhulgas, kuid mitte ainult, PEG või muud vees lahustuvad molekulid) sidumiseks muuhulgas nukleofiilse liitumise või aldooli kondensatsiooni reaktsioonide kaudu. Karbonüülrühma sisaldavate aminohapete näidete struktuurivalem võib olla: milles n on 0-; R 1 on alküül-, arüül-, asendatud alküül- või asendatud arüülrühm; R 2 on H, alküül-, arüül-, asendatud alküül- ja asendatud arüülrühm; ning R 3 on H, aminohape, polüpeptiid või aminootsa modifitseerimisrühm ja R 4 on H, aminohape, polüpeptiid või karboksüotsa modifitseerimisrühm. Mõne teostuse korral on n 1, R 1 on fenüül- ja R 2 lihtne alküülrühm (ehk metüül-, etüül- või propüülrühm) ning ketoonfragment paikneb alküül-
110 8 kõrvalahela suhtes para-positsioonis. Mõne teostuse korral on n 1, R 1 on fenüül- ja R 2 lihtne alküülrühm (ehk metüül-, etüül- või propüülrühm) ning ketoonfragment paikneb alküül-kõrvalahela suhtes meta-positsioonis. 5 p-atsetüül-(+/-)-fenüülalaniini ja m-atsetüül-(+/-)-fenüülalaniini sünteesimist kirjeldatakse allikas Zhang, Z., et al., Biochemistry 42: (03). Asjatundjad saavad analoogselt vamistada ka teisi karbonüülrühmi sisaldavaid aminohappeid. Mõne teostuse korral modifitseeritakse mittelooduslikku aminohapet sisaldavat polüpeptiidi keemiliselt, et saada reaktsioonivõimeline karbonüül-funktsionaalrühm. 30 Näiteks saab konjugatsioonireaktsioonide jaoks kasulikku funktsionaalset aldehüüdrühma luua funktsionaalrühmast, millel on külgnevad amino- ja hüdroksüülrühmad. Kui bioloogiliselt aktiivne molekul on polüpeptiid, saab näiteks N-otsa seriini või treoniini (mis võib alaliselt kohal olla või avalduda keemilise või ensümaatilise lõhustamise tulemusena) kasutada funktsionaalse aldehüüdrühma tekitamiseks leebe oksüdatiivse lõhustumise tingimustes perjodaadi abil. Vt nt Gaertner, et. al., Bioconjug. Chem. 3: (1992); Geoghegan, K. & Stroh, J., Bioconjug. Chem. 3: (1992); Gaertner et al., J. Biol. Chem. 269: (1994). Tehnika tasemes tuntud meetodid piirduvad peptiidi või valgu N-otsas paikneva aminohappega. Käesoleva leiutise korral saab mittelooduslikult kodeeritud aminohapet, mis kannab külgnevaid hüdroksüül- ja aminorühmi, kaasata polüpeptiidi koosseisu maskeeritud funktsionaalse aldehüüdrühmana. Näiteks kannab 5-hüdroksülüsiin hüdroksüülrühma, mis külgneb epsilon-amiiniga. Aldehüüdi loomiseks tarvilikud reaktsioonitingimused hõlmavad tüüpiliselt naatriumperjodaadi molaarse ülejäägi lisamist leebetes tingimustes, et vältida oksüdatsiooni muudes kohtades polüpeptiidi piires. Oksüdatsioonireaktsiooni ph on tüüpiliselt ligikaudu 7,0. Tüüpiline reaktsioon hõlmab naatriummetaperjodaadi u 1,5 molaarse ülejäägi lisamist polüpeptiidi puhverdatud lahusele, millele järgneb ligikaudu -minutiline inkubeerimine pimedas. Vt nt USA patenti nr Funktsionaalse karbonüülrühma saab panna selektiivselt reageerima hüdrasiin-, hüdrasiid-, hüdroksüülamiin- või semikarbasiidrühma sisaldava reagendiga leebetes tingimustes vesilahuses, et moodustada vastavaid hüdrasoon-, oksiim- või semikarbasoonsidemeid,
111 5 9 mis on füsioloogilistes tingimustes stabiilsed. Vt nt Jencks, W. P., J. Am. Chem. Soc. 81, (1959); Shao, J. and Tam, J. P., J. Am. Chem. Soc. 117: (1995). Lisaks võimaldab karbonüül- või dikarbonüülrühma unikaalne reaktsioonivõime selektiivset modifitseerimist muude aminohappe kõrvalahelate juuresolekul.vt nt Cornish, V. W., et al., J. Am. Chem. Soc. 118:80-81 (1996); Geoghegan, K. F. & Stroh, J. G., Bioconjug. Chem. 3: (1992); Mahal, L. K., et al., Science 276: (1997). B. Reaktsioonivõimelised hüdrasiin-, hüdrasiid- või semikarbasiidrühmad Mittelooduslikult kodeeritud aminohapped, mis sisaldavad nukleofiilset rühma nagu hüdrasiin-, hüdrasiid- või semikarbasiidrühm, reageerivad mitmesuguste elektrofiilsete rühmadega (muuhulgas, kuid mitte ainult, PEG või muude vees lahustuvate polümeeridega) ja moodustavad konjugaate. Hüdrasiin-, hüdrasiid- või semikarbasiidrühma sisaldavate aminohapete näited on järgnevad struktuuriga: kus n on 0-; R 1 on alküül-, arüül-, asendatud alküül- või asendatud arüülrühm või puudub; X on O, N või S või puudub; R 2 on H, aminohape, polüpeptiid või aminootsa modifitseerimisrühm ning R 3 on H, aminohape, polüpeptiid või karboksüotsa modifitseerimisrühm. Mõne teostuse korral on n 4, R 1 puudub ja X on N. Mõne teostuse korral on n 2, R 1 puudub ja X puudub. Mõne teostuse korral on n 1, R 1 on fenüülrühm, X on O ja hapnikuaatom paikneb arüültsüklis alifaatse rühma suhtes para-positsioonis. Hüdrasiid-, hüdrasiin- ja semikarbasiidrühma sisaldavad aminohapped on kaubanduslikult kättesaadavad. L-glutamaat-γ-hüdrasiidi näiteks turustab Sigma Chemical (St. Louis, MO). Muud, mitteturustatavad aminohapped on valmistatavad ala tavataseme asjatundja poolt. Vt nt patenti nr. US 6,281,211.
112 5 1 Polüpeptiide, mis sisaldavad mittelooduslikult kodeeritud aminohappeid, mis kannavad funktsionaalseid hüdrasiid-, hüdrasiin- või semikarbasiidrühmi, saab panna tõhusalt ja selektiivselt reageerima mitmesuguste molekulidega, mis sisaldavad aldehüüdrühma või muid sarnase keemilise reaktsioonivõimega funktsionaalrühmi. Vt nt Shao, J. and Tam, J., J. Am. Chem. Soc. 117: (1995). Hüdrasiid-, hüdrasiin- ja semikarbasiidfunktsionaalrühmade unikaalne reaktsioonivõime muudab need märksa reaktiivsemaks aldehüüdide, ketoonide ning muude elektrofiilsete rühmade suhtes võrreldes peamisel aminohappel leiduvate nukleofiilsete rühmadega (muuhulgas, kuid mitte ainult, seriini või treoniini hüdroksüülrühm või lüsiini ja N-otsa aminorühmad). C. Aminooksürühma sisaldavad aminohapped Mittelooduslikult kodeeritud aminohapped, mis sisaldavad aminooksü-(kasutatakse ka nimetust hüdroksüülamiin )-rühma, reageerivad mitmesuguste elektrofiilsete rühmadega ja moodustavad konjugaate (muuhulgas, kuid mitte ainult, PEG või muude vees lahustuvate polümeeridega). Sarnaselt hüdrasiin-, hüdrasiid- ja semikarbasiididrühmadele võimaldab aminooksürühma võimendatud nukleofiilsus panna seda tõhusalt ja selektiivselt reageerima mitmesuguste molekulidega, mis sisaldavad aldehüüdrühma või muid sarnase keemilise reaktsioonivõimega funktsionaalrühmi. Vt nt Shao, J. and Tam, J., J. Am. Chem. Soc. 117: (1995); H. Hang and C. Bertozzi, Acc. Chem. Res. 34: (01). Samas kui hüdrasiinrühmaga asetleidva reaktsiooni tulemusel tekib vastav hüdrasoon, tekib oksiim tavaliselt aminooksürühma ja karbonüülrühma sisaldava rühma nagu ketoonrühm reaktsiooni tulemusena. Aminooksürühmi sisaldavate aminohapete näidete struktuurivalem võib olla: milles n on 0-1.0; R 1 on alküül-, arüül-, asendatud alküül- või asendatud arüülrühm või puudub; X on O, N või S või puudub; m on 0-; Y = C(O) või puudub; R 2 on H, aminohape, polüpeptiid või aminootsa modifitseerimisrühm ning R 3 on H, aminohape, polüpeptiid või karboksüotsa modifitseerimisrühm. Mõne teostuse korral on n 1, R 1 on fenüülrühm, X on O, m on 1 ja Y on olemas. Mõne teostuse korral on n 2, R 1 ja X puuduvad, m on 0 ning Y puudub.
113 5 111 Aminooksürühma sisaldavaid aminohappeid saab hõlpsasti valmistada kättesaadavatest aminohappe prekursoritest (homoseriin, seriin ja treoniin). Vt nt M. Carrasco and R. Brown, J. Org. Chem. 68: (03). Teatud aminooksürühma sisaldavaid aminohappeid, näiteks L-2-amino-4-(aminooksü)võihapet, on isoleeritud looduslikest allikatest (Rosenthal, G., Life Sci. 60: (1997). Muud aminooksürühma sisaldavad aminohapped on asjatundjate poolt valmistatavad. D. Reaktsioonivõimelised asiid- ja alküünrühmad 30 Funktsionaalsete asiid- ja alküünrühmade unikaalne reaktiivsus muudab need äärmiselt kasulikeks polüpeptiidide ning muude bioloogiliste molekulide selektiivsel modifitseerimisel. Orgaanilised asiidid, eelkõige alifaatsed asiidid, ning alküünid on üldjuhul tavapärastes keemilistes reaktsioonitingimustes stabiilsed. Muuhulgas on nii asiid- kui ka alküünfunktsionaalrühmad inertsed looduslikult esinevates polüpeptiidides leiduva peamise aminohappe kõrvalahelate (ehk R-rühmade) suhtes. Samas avaldub asiid- ja alküünrühmade vedrulik olemus vahetusse lähedusse viimise korral ning nad reageerivad selektiivselt ja tõhusalt Huisgen [3+2] tsükloliitumisreaktsioonis ja moodustavad vastava triasooli. Vt nt Chin J., et al., Science 301:964-7 (03); Wang, Q., et al., J. Am. Chem. Soc. 1, (03); Chin, J. W., et al., J. Am. Chem. Soc. 124: (02). Kuivõrd Huisgeni tsükloliitumisreaktsioon hõlmab pigem selektiivset tsükloliitumisreaktsiooni (vt nt Padwa, A., COMPREHENSIVE ORGANIC SYNTHESIS, Vol. 4, (ed. Trost, B. M., 1991), lk ; Huisgen, R. 1,3-DIPOLAR CYCLOADDITION CHEMISTRY, (ed. Padwa, A., 1984), lk ) kui nukleofiilset asendamist, võimaldab asiidi ja alküüni sisaldavaid kõrvalahelaid kandvate mittelooduslikult kodeeritud aminohapete kaasamine saadavate polüpeptiidide selektiivset modifitseerimist mittelooduslikult kodeeritud aminohappe positsioonis. Asiidi või alküüni sisaldavat FGF-21 polüpeptiidi hõlmava tsükloliitumisreaktsiooni saab läbi viia toatemperatuuril vee juuresolekul Cu(II) lisamise teel (muuhulgas, kuid mitte ainult, CuSO 4 katalüütilise koguse kujul) redutseerija juuresolekul Cu(II) redutseerimiseks Cu(I)- ks kohapeal, katalüütilises koguses. Vt nt Wang, Q., et al., J. Am. Chem. Soc. 1, (03); Tornoe, C. W., et al., J. Org. Chem. 67: (02); Rostovtsev, et al., Angew. Chem. Int. Ed. 41:96-99 (02). Redutseerijate näideteks on
114 112 muuhulgas, kuid mitte ainult, askorbaat, metalliline vask, kiniin, hüdrokinoon, K-vitamiin, glutatioon, tsüsteiin, Fe 2+, Co 2+ ja elektrilise potentsiaali rakendamine. 5 Mõningatel juhtudel, kui soovitakse Huisgeni [3+2] tsükloliitumisreaktsiooni asiidi ja alküüni vahel, hõlmab FGF-21 polüpeptiid mittelooduslikult kodeeritud aminohapet, mille koosseisu kuulub alküünfragment, ning aminohappe külge kinnitatav vees lahustuv polümeer hõlmab asiidfragmenti. Alternatiivselt on võimalik ka vastupidise reaktsiooni (ehk asiidfragment aminohappel ja alküünfragment vees lahustuval polümeeril) läbiviimine. Samuti on võimalik asiidfunktsionaalrühma reaktsiooni selektiivne esilekutsumine arüülestrit sisaldava vees lahustuva polümeeriga ja selle kohane funktsionaliseerimine arüülfosfiinfragmendiga amiidsideme loomiseks. Arüülfosfiinrühm redutseerib asiidi in situ ja saadud amiin reageerib tõhusalt proksimaalse estersidemega, mille tulemusel tekib vastav amiid. Vt nt E. Saxon and C. Bertozzi, Science 287, 07- (00). Asiidi sisaldavaks aminohappeks võib olla kas alküülasiid (muuhulgas, kuid mitte ainult, 2- amino-6-asido-1-heksaanhape) või arüülasiid (p-asido-fenüülalaniin). Arüülestrit ja fosfiinfragmenti sisaldavate vees lahustuvate polümeeride näited on järgneva struktuuriga: milles X võib olla O, N, S või puududa, Ph on fenüülrühm, W on vees lahustuv polümeer ja R võib olla H, alküül-, arüül-, asendatud alküül- ning asendatud arüülrühm. R-rühmade näideteks on muuhulgas, kuid mitte ainult, -CH 2, -C(CH 3 ) 3, -OR', -NR'R", -SR', -halogeen, -C(O)R', -CONR'R", - S(O) 2 R', -S(O) 2 NR'R", -CN ja -NO 2. R', R", R'" ja R"" tähistavad eraldivõetuna vesinikku, asendatud või asendamata heteroalküül-, asendatud või asendamata arüülrühma, muuhulgas, kuid mitte ainult, 1-3 halogeeniga asendatud arüül-, asendatud või asendamata alküül-, alkoksü- või tioalkoksü- või arüülalküülrühmi. Kui leiutisekohane ühend sisaldab enam kui üht R rühma, valitakse näiteks iga selline R rühm sõltumatult, nagu ka iga R', R", R"' ja R"" rühm enam kui ühe vastava rühma olemasolu korral. Kui R' ja R" kinnituvad sama lämmastikuaatomi külge, saab neid selle lämmastikuaatomiga kombineerida 5-, 6- või 7-liikmelise rõnga moodustamiseks. Näiteks
115 5 113 on -NR'R" mõeldud hõlmama muuhulgas, kuid mitte ainult, 1-pürrolidinüüli ja 4- morfolinüüli. Asendajate eeltoodud käsitluse põhjal peab ala asjatundjale olema selge, et mõiste alküül on mõeldud hõlmama rühmi, sealhulgas süsinikuaatomeid, mis on seotud muude rühmadega peale vesinikurühmade, näiteks haloalküül (muuhulgas, kuid mitte ainult, -CF 3 ja -CH 2 CF 3 ) ning atsüül (muuhulgas, kuid mitte ainult, -C(O)CH 3, -C(O)CF 3, -C(O)CH 2 OCH 3 jms.). Samuti on võimalik asiidfunktsionaalrühma reaktsiooni selektiivne esilekutsumine tioestrit sisaldava vees lahustuva polümeeriga ja selle kohane funktsionaliseerimine arüülfosfiinfragmendiga amiidsideme loomiseks. Arüülfosfiinrühm redutseerib asiidi in situ ja saadud amiin reageerib tõhusalt tioestersidemega, luues vastava amiidi. Tioestrit ja fosfiinfragmenti sisaldavate vees lahustuvate polümeeride näited on järgneva valemiga: milles n on 1-; X võib olla O, N, S või puududa, Ph on fenüül ja W on vees lahustuv polümeer. Alküünrühma sisaldavate aminohapete näited on järgneva valemiga: milles n on 0-; R 1 on alküül-, arüül-, asendatud alküül- või asendatud arüülrühm või puudub; X on O, N või S või puudub; m on 0-, R 2 on H, aminohape, polüpeptiid või aminootsa modifitseerimisrühm, ning R 3 on H, aminohape, polüpeptiid või karboksüotsa modifitseerimisrühm. Mõne teostuse korral on n 1, R 1 on fenüülrühm, X puudub, m on 0 ning atsetüleenfragment paikneb alküül-kõrvalahela suhtes para-positsioonis. Mõne teostuse korral on n 1, R 1 on fenüülrühm, X on O, m on 1 ning propargüüloksürühm paikneb alküül-kõrvalahela suhtes para-positsioonis (ehk O-propargüül-türosiin). Mõne teostuse korral on n 1, R 1 ja X puuduvad ning m on 0 (ehk proparüülglütsiin). Alküünrühma sisaldavad aminohapped on kaubanduslikult kättesaadavad. Propargüülglütsiini näiteks turustab ettevõte Peptech (Burlington, MA). Alküünrühma sisaldavaid aminohappeid on võimalik valmistada ka standardsetel meetoditel. Näiteks
116 114 saab p-propargüüloksüfenüülalaniini sünteesida vastavalt kirjeldusele allikas Deiters, A., et al., J. Am. Chem. Soc. 1: (03) ning 4-alkünüül-L-fenüülalaniini sünteesida vastavalt kirjeldusele allikas Kayser, B., et al., Tetrahedron 53(7): (1997). Muud alküleenrühma sisaldavad aminohapped on asjatundjate poolt valmistatavad. 5 Asiidrühma sisaldavate aminohapete näited on järgmise valemiga: milles n on 0-; R 1 on alküül, arüül, asendatud alküül, asendatud arüül või puudub; X on O, N või S või puudub; m on 0-, R 2 on H, aminohape, polüpeptiid või aminootsa modifitseerimisrühm, ning R 3 on H, aminohape, polüpeptiid või karboksüotsa modifitseerimisrühm. Mõne teostuse korral on n 1, R 1 on fenüülrühm, X puudub, m on 0 ning asiidfragment paikneb alküül-kõrvalahela suhtes para-positsioonis. Mõne teostuse korral on n 0-4, R 1 ja X puuduvad ning m = 0. Mõne teostuse korral on n 1, R 1 on fenüülrühm, X on O, m on 2 ja β-asidoetoksüfragment paikneb alküül-kõrvalahela suhtes para-positsioonis. Asiidrühma sisaldavad aminohapped on kaubanduslikult kättesaadavad. Näiteks turustab 4-asidofenüülalaniini näiteks turustab Chem-Impex International, Inc. (Wood Dale, IL). Mis puutub asiidi sisaldavatesse aminohapetesse, mis pole kaubanduslikult saadaval, siis on asiidrühma suhteliselt lihtne valmistada ala tavataseme asjatundjatele teadaolevate standardsete meetodite abil, muuhulgas, kuid mitte ainult, sobiva lahkuva rühma (muuhulgas, kuid mitte ainult, halogeniid, mesülaat, tosülaat) ümberpaigutamise või sobivalt kaitstud laktooni avamise teel. Vt nt Advanced Organic Chemistry, March (Third Edition, 1985, Wiley and Sons, New York). E. Reaktsioonivõimelised aminotioolrühmad Beeta-asendatud aminotioolfunktsionaalrühmade unikaalne reaktiivsus muudab need äärmiselt kasulikeks polüpeptiidide ning muude aldehüüdrühmi sisaldavate bioloogiliste molekulide selektiivsel modifitseerimisel tiasolidiini moodustamise teel. Vt. nt. J. Shao and J. Tam, J. Am. Chem. Soc. 1995, 117 (14) Mõne teostuse korral saab beeta-asendatud aminotiool-aminohapped kaasata FGF-21 polüpeptiidide koosseisu ja
117 1 panna nad seejärel reageerima vees lahustuvate polümeeridega, mis sisaldavad funktsionaalset aldehüüdrühma. Mõne teostuse korral saab vees lahustuva polümeeri, ravimi konjugaadi või muu kasuliku lisandi siduda FGF-21 polüpeptiidiga, mis sisaldab beeta-asendatud aminotiool-aminohapet, tiasolidiini moodustamise kaudu. 5 F. Teised reaktsioonivõimelised rühmad Teisi reaktsioonivõimelisi rühmi ja mittelooduslikult kodeeritud aminohappeid, mida saab sisestada leiutisekohastesse FGF-21 polüpeptiididesse, kirjeldatakse järgnevates patenditaotlustes: USA patendipublikatsioon nr 06/01946, USA patendipublikatsioon nr 06/ , USA patendipublikatsioon nr 06/ , USA eelpatent nr US 60/755,338; USA eelpatent nr 60/755,711; ajutine patent nr. US 60/755,018; rahvusvaheline patenditaotlus nr PCT/US06/49397; WO 06/069246; USA eelpatent nr 60/743,041; USA eelpatent nr 60/743,040; rahvusvaheline patenditaotlus nr PCT/US06/47822; USA eelpatent nr 60/882,819; USA eelpatent nr 60/882,500; ning USA eelpatent nr 60/870,594. MITTELOODUSLIKE AMINOHAPETE SISENEMINE RAKKUDESSE Mittelooduslike aminohapete sisenemine rakkudesse on aspektiks, mida mittelooduslike aminohapete väljatöötamisel ja selekteerimisel reeglina arvesse võetakse, muuhulgas, kuid mitte ainult, valku sisestamisel. Näiteks viitab α-aminohapete kõrge laengutihedus sellele, et need ühendid ei ole tõenäoliselt raku poolt läbilastavad. Looduslikud aminohapped sisenevad eukarüootsesse rakku valgupõhiste transpordisüsteemide kogumi kaudu. Kiire skriinimise teel saab hinnata, milliseid mittelooduslikke aminohappeid, kui üldse, rakud suudavad endasse haarata. Vt nt toksilisuskatseid näiteks patendipublikatsioonis nr US 04/ pealkirjaga Protein Arrays ; ja Liu, D.R. & Schultz, P.G. (1999) Progress toward the evolution of an organism with an expanded genetic code. PNAS United States 96: Ehkki aminohapete sisenemist rakkudesse saab erinevate katsete abil hõlpsasti analüüsida, on rakutasandi sisenemisradadega kokkusobivate mittelooduslike aminohapete väljatöötamise alternatiiviks biosünteetiliste radade tagamine aminohapete loomiseks in vivo.
118 116 MITTELOODUSLIKE AMINOHAPETE BIOSÜNTEES 5 30 Rakkudes on juba olemas hulgaliselt biosünteesiradu aminohapete ja muude ühendite tootmiseks. Ehkki teatud konkreetse aminohappe biosünteesi meetodit ei pruugi eksisteerida looduses, muuhulgas, kuid mitte ainult, rakus, pakutakse käesoleva leiutisega sellised meetodid. Näiteks luuakse biosünteesirajad mittelooduslike aminohapete jaoks valikuliselt peremeesrakus uute ensüümide lisamise või peremeesraku olemasolevate radade modifitseerimise teel. Täiendavateks uuteks ensüümideks on valikuliselt kas looduslikult esinevad ensüümid või tehisensüümid. Näiteks tugineb p-aminofenüülalaniini biosüntees (mida näitena käsitletakse allikas WO 02/ pealkirjaga In vivo incorporation of unnatural amino acids ) muudest organismidest pärit teadaolevate ensüümide kombinatsiooni lisamisel. Nende ensüümide geene saab viia eukarüootsesse rakku selle transformeerimise teel vastavaid geene hõlmava plasmiidiga. Rakus ekspresseerimisel tagavad geenid ensümaatilise raja soovitud ühendi sünteesimiseks. Valikuliselt lisatavate ensüümide tüüpide näiteid on esitatud alltoodud näidetes. Täiendavaid ensüümijärjestusi võib leida näiteks GenBank ist. Samuti lisatakse rakule valikuliselt samal viisil tehislikult loodud ensüümid. Nõnda manipuleeritakse rakutasandi mehhanisme ja raku ressursse mittelooduslike aminohapete tootmiseks. Olemas on mitmesugused meetodid uudsete ensüümide tootmiseks biosünteesiradade koosseisus kasutamiseks või olemasolevate radade arendamiseks. Näiteks võib uudsete ensüümide ja radade väljatöötamiseks valikuliselt kasutada rekursiivset rekombinatsiooni, mida pakub muuhulgas, kuid mitte ainult, ettevõte Maxygen, Inc. (juurdepääsetav Internetis aadressil Vt nt Stemmer (1994), Rapid evolution of a protein in vitro by DNA shuffling, Nature 370(4): ; ja Stemmer, (1994), DNA shuffling by random fragmentation and reassembly: In vitro recombination for molecular evolution, Proc. Natl. Acad. Sci. USA., 91: Sarnasel viisil kasutatakse DesignPath -i, mille töötas välja ettevõte Genencor (juurdepääsetav Internetis aadressil valikuliselt metaboolse raja väljatöötamiseks, muuhulgas, kuid mitte ainult, raja väljatöötamiseks O-metüül-L-türosiini loomiseks rakus. Antud tehnoloogia puhul rekonstrueeritakse olemasolevad rajad peremeesorganismides muuhulgas, kuid mitte ainult, funktsionaalse genoomika ja molekulaarse evolutsiooni ning disaini kaudu tuvastatud uute geenide kombinatsiooni abil. Diversa Corporation (juurdepääsetav Internetis aadressil pakub samuti tehnoloogiat geenide ja geeniradade
119 117 pankade kiireks skriinimiseks, muuhulgas, kuid mitte ainult, uute radade loomise eesmärgil. 5 Tüüpiliselt toodetakse leiutisekohase tehisliku biosünteetilise rajaga valmistatud mittelooduslikku aminohapet kontsentratsioonis, millest piisab valgu tõhusaks biosünteesiks, muuhulgas, kuid mitte ainult, looduslikus rakutasandi koguses, kuid mitte niisugusel määral, et see mõjutaks muude aminohapete kontsentratsiooni või ammendaks rakutasandi ressursid. Tüüpilised niisugusel viisil in vivo toodetud kontsentratsioonid jäävad vahemikku alates u mm kuni u. 0,05 mm. Kui rakk on transformeeritud plasmiidiga, mis hõlmab geene, mida kasutatakse teatud konkreetse raja jaoks soovitud ensüümide tootmiseks ja saadakse mittelooduslik aminohape, kasutatakse in vivo selektsioone valikuliselt mitteloodusliku aminohappe tootmise edasiseks optimeerimiseks nii ribosomaalse valgusünteesi kui ka rakukasvu osas. MITTELOODUSLIKE AMINOHAPETEGA POLÜPEPTIIDID 30 Mitteloodusliku aminohappe kaasamine on võimalik mitmesugustel eesmärkidel, muuhulgas, kuid mitte ainult, muudatuste esilekutsumiseks valgu struktuuris ja/või talitluses, proteaasi sihtkohtade suuruse, happelisuse, nukleofiilsuse, vesiniku sidumise, hüdrofoobsuse, juurdepääsetavuse muutmiseks, fragmendi sihtmärgistamiseks (muuhulgas, kuid mitte ainult, valgukiibi jaoks), bioloogiliselt aktiivse molekuli lisamiseks, polümeeri kinnitamiseks, radionukliidi kinnitamiseks, seerumi poolväärtusaja moduleerimiseks, koe läbistamise moduleerimiseks (nt kasvajad), aktiivse transpordi moduleerimiseks, koe, raku või organi spetsiifilisuse või jaotuse moduleerimiseks, immunogeensuse moduleerimiseks, proteaasi resistentsuse moduleerimiseks jne. Mittelooduslikku aminohapet hõlmavatel valkudel võivad olla võimendatud või isegi sootuks uued katalüütilised või biofüüsilised omadused. Näiteks on võimalik järgnevate omaduste valikuline modifitseerimine mitteloodusliku aminohappe kaasamise teel valku: toksilisus, biojaotuvus, struktuuriomadused, spektroskoopilised omadused, keemilised ja/või fotokeemilised omadused, katalüüsivõime, poolväärtusaeg (muuhulgas, kuid mitte ainult, seerumi poolväärtusaeg), võime regaeerida teiste molekulidega, muuhulgas, kuid mitte ainult, kovalentselt või mittekovalentselt jne. Kompositsioonidest, sealhulgas valkudest, mis sisaldavad vähemalt ühte mittelooduslikku aminohapet, on kasu muuhulgas, kuid mitte ainult, järgnevates valdkondades: uudisravimid, diagnostika, katalüütilised ensüümid,
120 118 tööstuslikud ensüümid, sidumisvalgud (muuhulgas, kuid mitte ainult, antikehad), ning muuhulgas, kuid mitte ainult, valkude struktuuri ja talitluse uurimine. Vt nt Dougherty, (00) Unnatural Amino Acids as Probes of Protein Structure and Function, Current Opinion in Chemical Biology, 4: Leiutise ühe aspekti kohaselt sisaldab kompositsioon vähemalt ühte valku, mis sisaldab ühte mittelooduslikku aminohapet. Leiutise üheks tunnusjooneks on huvipakkuvad valgud või polüpeptiidid, mis sisaldavad ühte mittelooduslikku aminohapet. Leiutis hõlmab ka polüpeptiide või valke ühe mitteloodusliku aminohappega, mis on toodetud leiutisekohaseid kompositsioone ja meetodeid kasutades. Valgus võib sisalduda ka abiaine (muuhulgas, kuid mitte ainult, farmatseutiliselt sobiv abiaine). Huvipakkuvate valkude või polüpeptiidide valmistamisel ühe mitteloodusliku aminohappega eukarüootsetes rakkudes hõlmavad need valgud või polüpeptiidid tüüpiliselt eukarüootseid translatsioonijärgseid modifikatsioone. Teatud teostuste korral sisaldab valk üht mittelooduslikku aminohapet ja vähemalt üht translatsioonijärgset modifikatsiooni, mis on valmistatud in vivo eukarüootse raku poolt, kusjuures translatsioonijärgne modifikatsioon ei ole valmistatud prokarüootse raku poolt. Näiteks hõlmab translatsioonijärgne modifikatsioon muuhulgas, kuid mitte ainult, atsetüülimist, atsüülimist, lipiidmodifikatsiooni, palmitoüülimist, palmitaadi liitmist, fosforüülimist, glükolipiidsideme modifikatsiooni, glükosüülimist jms. Ühe aspekti kohaselt hõlmab translatsioonijärgne modifitseerimine oligosahhariidi (muuhulgas, kuid mitte ainult, (GlcNAc-Man) 2 -Man-GlcNAc-GlcNAc)) kinnitamist asparagiini külge GlcNAcasparagiinsidemega. Vt tabelit 1, kus tuuakse mõningaid näiteid eukarüootsete valkude N- seotud oligosahhariidide kohta (võimalik on muudegi jääkide juuresolek, mida ei kujutata). Teise aspekti kohaselt hõlmab translatsioonijärgne modifikatsioon oligosahhariidi (muuhulgas, kuid mitte ainult, Gal-GalNAc, Gal-GlcNAc jne.) kinnitamist seriini või treoniini külge GalNAc-seriin või GalNAc-treoniinsidemega, või GIcNAc-seriin või GlcNAc-treoniinsidemega. TABEL 1: OLIGOSAHHARIIDIDE NÄITED GlcNAc-SIDEME KAUDU
121 119 Tüüp Alusstruktuur Kõrge mannoos Hübriid Kompleks Ksüloos 5 Veel ühe aspekti kohaselt hõlmab translatsioonijärgne modifitseerimine prekursorite (muuhulgas, kuid mitte ainult, kaltsitoniini prekursor, kaltsitoniini geeniga seotud peptiidi prekursor, preproparatüroidhormoon, preproinsuliin, proinsuliin, preproopiomelanokortiin, pro-opiomelanokortiin jms) proteolüütilist töötlemist, kokkupanekut mitmest allüksusest koosnevaks valguks või makromolekulaarseks koostuks, translatsiooni teise kohta rakus (muuhulgas, kuid mitte ainult, organellid, näiteks endoplasmaatiline retiikulum, Golgi kompleks, tuum, lüsosoomid, peroksisoomid, mitokondrid, kloroplastid, vakuoolid jne. või sekretoorse raja kaudu). Teatud teostuste korral hõlmab valk sekretsiooni- või lokalisatsioonijärjestust, epitoopmärgist, FLAG märgist, polühistidiinmärgist, GST sulandit vms.
122 Mitteloodusliku aminohappe üheks eeliseks on asjaolu, et see pakub täiendavaid keemilisi fragmente, mida saab kasutada täiendavate molekulide lisamiseks. Vastavad modifikatsioonid on teostatavad in vivo eukarüootses või mitteeukarüootses rakus, või in vitro. Nõnda leiab translatsioonijärgne modifikatsioon teatud teostuste korral aset mitteloodusliku aminohappe kaudu. Näiteks võib translatsioonijärgne modifikatsioon toimuda nukleofiilse-elektrofiilse reaktsiooni kaudu. Suurem osa reaktsioonidest, mida tänapäeval kasutatakse valkude selektiivseks modifitseerimiseks, hõlmab kovalentse sideme moodustamist nukleofiilse ja elektrofiilse reaktsioonipartneri vahele, muuhulgas, kuid mitte ainult, α-haloketoonide reaktsiooni histidiini või tsüsteiini kõrvalahelatega. Selektiivsuse määrab vastavatel juhtudel ära nukleofiilsete jääkide arv ning juurdepääsetavus valgus. Leiutisekohastes valkudes saab kasutada muid, selektiivsemaid reaktsioone, näiteks mitteloodusliku ketoaminohappe reaktsiooni hüdrasiidide või aminooksüühenditega in vitro ja in vivo. Vt. nt. Cornish, et al., (1996) J. Am. Chem. Soc., 118:80-81; Mahal, et al., (1997) Science 276: ; Wang, et al., (01) Science 292: ; Chin, et al., (02) J. Am. Chem. Soc. 124: ; Chin, et al., (02) Proc. Natl. Acad. Sci., 99:1-124; Wang, et al., (03) Proc. Natl. Acad. Sci., 0:56-61; Zhang, et al., (03) Biochemistry, 42: ; ning Chin, et al., (03) Science, 301: See võimaldab praktiliselt mis tahes valgu selektiivset märgistamist suure arvu reaktiividega, sealhulgas fluorofoorid, ristsidujad, sahhariidi derivaadid ning tsütotoksilised molekulid. Vt. ka patent nr. US 6,927,042 pealkirjaga Glycoprotein synthesis. Translatsioonijärgsed modifikatsioonid, muuhulgas, kuid mitte ainult, asidoaminohappe kaudu, on teostatavad ka Staudingeri ligeerimise kaudu (muuhulgas, kuid mitte ainult, triarüülfosfiinreaktiividega). Vt nt Kiick et al., (02) Incorporation of azides into recombinant proteins for chemoselective modification by the Staudinger ligation, PNAS 99: Leiutisega pakutakse veel üks ülitõhus meetod valkude selektiivseks modifitseerimiseks, mis hõlmab muuhulgas, kuid mitte ainult, asiid- või alkünüülfragmenti sisaldavate mittelooduslike aminohapete geneetilist sisestamist valkudesse vastusena selektorkoodonile. Neid aminohappe kõrvalahelaid saab seejärel modifitseerida muuhulgas, kuid mitte ainult, Huisgeni [3+2] tsükloliitmisreaktsiooni teel (vt. nt Padwa, A., Comprehensive Organic Synthesis, Vol. 4. (1991) Ed. Trost, B. M., Pergamon, Oxford, lk ; ja, Huisgen, R. 1.3-Dipolar Cycloaddition Chemistry, (1984) Ed. Padwa,
123 5 121 A., Wiley, New York, lk ) muuhulgas, kuid mitte ainult, vastavalt kas alkünüül- või asiidderivaatidega. Kuivõrd kõnealune meetod hõlmab tsükloliitmist nukleofiilse asendamise asemel, saab valke modifitseerida ülimalt kõrge selektiivsusega. Reaktsiooni saab läbi viia toatemperatuuril vee juuresolekul suurepärase regioselektiivsuse (1,4 > 1,5) juures Cu(I) soolade katalüütiliste koguste lisamise teel reaktsioonisegule. Vt. nt. Tornoe, et al., (02) J. Org. Chem. 67: ; ja Rostovtsev, et al., (02) Angew. Chem. Int. Ed. 41: Veel üheks kasutatavaks meetodiks on ligandivahetus bisarseeniühendil tetratsüsteiini motiiviga, vt. nt. Griffin, et al., (1998) Science 281: Molekulid, mida saab leiutisekohasele valgule lisada [3+2] tsükloliitmise kaudu, hõlmavad praktiliselt mis tahes molekule asiid- või alkünüülderivaadiga. Molekulid hõlmavad muuhulgas, kuid mitte ainult, värvaineid, fluorofoore, ristsidujaid, sahhariidi derivaate, polümeere (muuhulgas, kuid mitte ainult, polüetüleenglükooli derivaadid), fotoristsidujaid, tsütotoksilisi ühendeid, afiinsusmärgiseid, biotiini derivaate, vaike, helmeid, teist (või enamat) valku või polüpeptiidi, polünukleotiidi (polünukleotiide) (muuhulgas, kuid mitte ainult, DNA, RNA jne.), metalli kelaatoreid, kofaktoreid, rasvhappeid, süsivesikuid jne. Neid molekule saab lisada vastavalt mittelooduslikule aminohappele alkünüülrühmaga, muuhulgas, kuid mitte ainult, p- propargüloksüfenüülalaniin, või asidorühmaga, muuhulgas, kuid mitte ainult, p- asidofenüülalaniin. V. Mittelooduslikult kodeeritud aminohappeid sisaldavate FGF-21 polüpeptiidide tootmine in vivo 30 Leiutisekohaseid FGF-21 polüpeptiide saab valmistada in vivo modifitseeritud trna ja trna süntetaaside abil niisuguste aminohapete lisamiseks või asendamiseks, mida looduses esinevates süsteemides ei kodeerita. Meetodeid trna ja trna süntetaaside valmistamiseks, mis kasutavad aminohappeid, mida looduses esinevates süsteemides ei kodeerita, on kirjeldatud nt. patentides nr. US 7,045,337 ja US 7,083,970. Need meetodid hõlmavad translatsioonimehhanismi loomist, mis toimib sõltumatult süntetaasidest ja trna-dest, mis on translatsioonisüsteemi suhtes
124 endogeensed (ning millele seetõttu mõnikord viidatakse kui ortogonaalsetele ). Tüüpiliselt hõlmab translatsioonisüsteem ortogonaalset trna-d (O-tRNA) ja ortogonaalset aminoatsüül-trna-süntetaasi (O-RS). Tüüpiliselt aminoatsüülib O-RS eelistatavalt O-tRNA-d vähemalt ühe looduslikult mitteesineva aminohappega translatsioonisüsteemis ja O-tRNA tuvastab vähemalt ühe selektorkoodoni, mida muud trna-d süsteemis ei tuvasta. Nõnda sisestab translatsioonisüsteem mittelooduslikult kodeeritud aminohappe süsteemis toodetud valku vastuseks kodeeritud selektorkoodonile, asendades niisugusel viisil aminohappe kodeeritud polüpeptiidi kohale. Tehnika tasemes on kirjeldatud palju erinevaid ortogonaalseid trna-sid ja aminoatsüültrna-süntetaase konkreetsete tehislike aminohapete sisestamiseks polüpeptiididesse, olles üldiselt sobivad käesolevas leiutises kasutamiseks. Näiteks on ketospetsiifilisi O- trna/aminoatsüül-trna süntetaase kirjeldatud allikates Wang, L., et al., Proc. Natl. Acad. Sci. USA 0:56-61 (03) ning Zhang, Z. et al., Biochem. 42(22): (03). Näitlikke O-RS või nende osi kodeerivad polünukleotiidjärjestused ja need hõlmavad aminohappejärjestusi, mida kirjeldatakse patentides nr. US 7,045,337 ja US 7,083,970. Vastavaid O-tRNA molekule kasutamiseks koos O-RS-idega on samuti kirjeldatud patentides nr. US 7,045,337 ja US 7,083,970. O-tRNA/aminoatsüül-tRNAsüntetaasipaaride täiendavaid näiteid kirjeldavad WO 05/007870, WO 05/ ning WO 05/0194. Asiidspetsiifilise O-tRNA/aminoatsüül-tRNA süntetaasisüsteemi näidet on kirjeldatud allikas Chin, J. W., et al., J. Am. Chem. Soc. 124: (02). Näitlikud O-RS järjestused p-asido-l-phe jaoks hõlmavad muuhulgas, kuid mitte ainult, nukleotiidijärjestusi SEQ ID numbritega ja ning aminohappejärjestusi SEQ ID numbritega ja 61-64, nagu kirjeldatud patendis nr. US 7,083,970. Selle leiutise raames kasutamiseks sobivad näitlikud O-tRNA järjestused hõlmavad muuhulgas, kuid mitte ainult, nukleotiidijärjestusi SEQ ID numbritega 1-3, nagu käsitletud patendis nr. US 7,083,970). Muid O-tRNA/aminoatsüül-tRNA süntetaasi paaride näiteid, mis on spetsiifilised konkreetsetele mittelooduslikult kodeeritud aminohapetele, on kirjeldatud patendis nr. US 7,045,337. O-RS ja O-tRNA-d, mis hõlmavad nii keto- kui ka asiidisisaldusega aminohappeid S. cerevisiae s on kirjeldatud allikas Chin, J. W., et al., Science 301: (03).
125 Teada on antud mitmest muust ortogonaalsest paarist. Glutaminüül- (vt. nt. Liu, D. R., and Schultz, P. G. (1999) Proc. Natl. Acad. Sci. U. S. A. 96: ), aspartüül- (vt. nt. Pastrnak, M., et al., (00) Helv. Chim. Acta 83: ) ja türosüülsüsteeme (vt. nt. Ohno, S., et al., (1998) J. Biochem. (Tokyo, Jpn.) 124:65-68; ja Kowal, A. K., et al., (01) Proc. Natl. Acad. Sci. U. S. A. 98: ), mis on tuletatud S. cerevisiae trna-dest ja süntetaasidest, on kirjeldatud seoses mittelooduslike aminohapete potentsiaalse kaasamisega E. coli sse. E. coli glutaminüüli (vt. nt. Kowal, A. K., et al., (01) Proc. Natl. Acad. Sci. U. S. A. 98: ) ja türosüüli (vt. nt. Edwards, H., and Schimmel, P. (1990) Mol. Cell. Biol. : ) süntetaasidest tuletatud süsteeme on kirjeldatud seoses kasutamisega S. cerevisiae s. E. coli türosüülsüsteemi on kasutatud 3- jodo-l-türosiini kaasamiseks in vivo, imetajarakkudes. Vt. Sakamoto, K., et al., (02) Nucleic Acids Res. 30: O-tRNA/aminoatsüül-tRNA süntetaaside kasutamine hõlmab spetsiifilise koodoni selektsiooni, mis kodeerib mittelooduslikult kodeeritud aminohapet. Samas kui kasutada võib ükskõik millist koodonit, on üldjuhul soovitatav valida koodon, mida rakus, kus O- trna/aminoatsüül-trna süntetaas ekspresseeritakse, kasutatakse harva või ei kasutata kunagi. Vastavate koodonite näideteks on nonsenss-koodon nagu stoppkoodonid (merevaik, ooker ja opaal), nelja või enam alusega koodonid ning muud looduslikud kolme alusega koodonid, mida kasutatakse harva või mitte kunagi. Spetsiifilist selektorkoodonit (või -koodoneid) saab sisestada kohastele positsioonidele FGF-21 polünukleotiidi kodeerimisjärjestuses tehnika tasemest teadaolevaid mutageneesimeetodeid (muuhulgas, kuid mitte ainult, koht-suunatud mutagenees, kassettmutagenees, restriktsioon-selektsioon mutagenees jne.) kasutades. Meetodeid valgu biosünteesimehhanismi komponentide loomiseks nagu O-RS-id, O- trna-d ja ortogonaalsed O-tRNA/O-RS paarid, mida saab kasutada mittelooduslikult kodeeritud aminohappe kaasamiseks, on kirjeldatud allikates Wang, L., et al., Science 292: (01); Chin, J. W., et al., J. Am. Chem. Soc. 124: (02); Zhang, Z. et al., Biochemistry 42: (03). Meetodeid ja kompositsioone mittelooduslikult kodeeritud aminohapete kaasamiseks in vivo on kirjeldatud patendis nr. US 7,045,337. Meetodeid ortogonaalse trna-trna süntetaasi paari selekteerimiseks organismi in vivo translatsioonisüsteemis kasutamise eesmärgil on samuti kirjeldatud
126 patentides nr. US 7,045,337 ja US 7,083,970. PCT publikatsioon nr. WO 04/ pealkirjaga Site Specific Incorporation of Keto Amino Acids into Proteins (Ketoaminohapete kohaspetsiifiline kaasamine valkudesse) kirjeldab ortogonaalseid RS ja trna paare ketoaminohapete kaasamiseks. PCT publikatsioon nr. WO 04/ pealkirjaga Expanding the Eukaryotic Genetic Code (Eukarüootse geneetilise koodi laiendamine) kirjeldab ortogonaalseid RS ja trna paare mittelooduslikult kodeeritud aminohapete kaasamiseks eukarüootsetesse peremeesrakkudesse. Meetodid vähemalt ühe rekombinantse ortogonaalse aminoatsüül-trna süntetaasi (O-RS) valmistamiseks hõlmavad: (a) panga loomine (valikuliselt mutantsetest) RS-idest, mis on tuletatud vähemalt ühest aminoatsüül-trna süntetaasist (RS) esimeselt organismilt, muuhulgas, kuid mitte ainult, prokarüootselt organismilt nagu Methanococcus jannaschii, Methanobacterium thermoautotrophicum, Halobacterium, Escherichia coli, A. fulgidus, P. furiosus, P. horikoshii, A. pernix, T. thermophilus vms., või eukarüootselt organismilt; (b) RS-ide (valikuliselt mutantsete RS-ide) panga selekteerimine (ja/või sõelumine) liikmete leidmiseks, mis aminoatsüülivad ortogonaalse trna (O-tRNA) mittelooduslikult kodeeritud aminohappe ning loodusliku aminohappe juuresolekul, luues nõnda aktiivsete (valikuliselt mutantsete) RS-ide kogumi; ja/või (c) kogumi selekteerimine (valikuliselt negatiivse selektsiooni kaudu) aktiivsete RS-ide (muuhulgas, kuid mitte ainult, mutantsete RS-ide) leidmiseks, mis eelistuslikult aminoatsüülivad O-tRNA-d mittelooduslikult kodeeritud aminohappe puudumisel, andes sedakaudu tulemuseks vähemalt ühe rekombinantse O-RS-i; kusjuures mainitud vähemalt üks rekombinantne O-RS aminoatsüülib eelistatavalt O-tRNA-d mittelooduslikult kodeeritud aminohappega. Ühe teostuse kohaselt on RS-iks mitteaktiivne RS. Mitteaktiivset RS-i saab tekitada aktiivse RS-i muteerimise teel. Näiteks saab mitteaktiivset RS-i tekitada vähemalt u. 1, vähemalt u. 2, vähemalt u. 3, vähemalt u. 4, vähemalt u. 5, vähemalt u. 6 või vähemalt u. või enama aminohappe muteerimise teel erinevate aminohapetega, muuhulgas, kuid mitte ainult, alaniiniga. Mutantsete RS-ide panku saab moodustada mitmesuguste valdkonnas teadaolevate tehnikate abil, muuhulgas, kuid mitte ainult, ratsionaalse disaini teel, mis põhineb valgu kolmemõõtmelisel RS-struktuuril, või RS nukleotiidide mutageneesi teel juhusliku või
127 1 ratsionaalse disainitehnika abil. Näiteks saab mutantseid RS-e valmistada kohaspetsiifiliste mutatsioonide, juhuslike mutatsioonide, mitmekesisust põhjustavate rekombinatsioonmutatsioonide, kimäärsete konstruktide, ratsionaalse disaini ja muude siinkirjeldatud või tehnika tasemest teadaolevate meetodite abil Leiutise ühe teostuse kohaselt hõlmab RS-ide (valikuliselt mutantsete RS-ide) panga selekteerimine (ja/või sõelumine) aktiivsete liikmete leidmiseks, muuhulgas, kuid mitte ainult, mis aminoatsüülivad ortogonaalset trna-d (O-tRNA) mittelooduslikult kodeeritud aminohappe ja loodusliku aminohappe juuresolekul: positiivse selektsiooni või sõelumise markeri, muuhulgas, kuid mitte ainult, antibiootikumi resistentsusgeeni vms. ja (valikuliselt mutantsete) RS-ide panga sisseviimist mitmesse rakku, kusjuures positiivse selektsiooni ja/või sõelumise marker hõlmab vähemalt üht selektorkoodonit, muuhulgas, kuid mitte ainult, merevaik-, ooker- või opaalkoodonit; mitme raku kasvatamist selektsiooniagensi juuresolekul; rakkude tuvastamist, mis jäävad ellu (või näitavad üles spetsiifilist vastust) selektsiooni- ja/või sõelumisagensi juuresolekul vähemalt ühe selektorkoodoni pärssimise teel positiivse selektsiooni või sõelumise markeris, mille kaudu luuakse positiivselt selekteeritud rakkude alamkogum, mis sisaldab aktiivsete (valikuliselt mutantsete) RS-ide kogumit. Valikuliselt on võimalik selektsiooni- ja/või sõelumisagensi kontsentratsiooni varieerimine. Leiutise ühe aspekti kohaselt on positiivse selektsiooni markeriks klooramfenikooli atsetüültransferaasi (CAT) geen ja selektorkoodoniks merevaik-stoppkoodon CAT geenis. Valikuliselt on positiivse selektsiooni markeriks β-laktamaasi geen ja selektorkoodoniks merevaik-stoppkoodon β-laktamaasi geenis. Leiutise veel ühe aspekti kohaselt hõlmab positiivse sõelumise marker fluorestsents- või luminestsents-sõelumismarkerit või afiinsuspõhist sõelumismarkerit (muuhulgas, kuid mitte ainult, rakupinna marker). Leiutise ühe teostuse kohaselt hõlmab aktiivsete RS-ide (valikuliselt mutantide) kogumi negatiivne selekteerimine või sõelumine, mis eelistatavalt aminoatsüülivad O-tRNA-d mittelooduslikult kodeeritud aminohappe puudumisel: negatiivse selektsiooni või sõelumise markeri sisseviimist aktiivsete (valikuliselt mutantsete) RS-ide positiivsest selektsioonist või sõelumisest tulenevasse kogumisse, kusjuures negatiivne selektsiooni või sõelumise marker hõlmab vähemalt üht selektorkoodonit (muuhulgas, kuid mitte ainult, antibiootikumi resistentsusgeeni, muuhulgas, kuid mitte ainult, klooramfenikooli
128 atsetüültransferaasi (CAT) geeni); ning niisuguste rakkude tuvastamist, mis jäävad ellu või näitavad üles spetsiifilist vastust sõelumisele esimeses keskkonnas, mida on täiendatud mittelooduslikult kodeeritud aminohappe ja sõelumis- või selektsiooniagensiga, kuid ei jää ellu ega näita üles spetsiifilist vastust teises keskkonnas, mida pole täiendatud mittelooduslikult kodeeritud aminohappe ja selektsiooni- või sõelumisagensiga, mille tulemusena ellujäävad rakud või sõelutud rakud varustatakse vähemalt ühe rekombinantse O-RS-iga. Näiteks toimib CAT-tuvastusprotokoll valikuliselt positiivse selektsiooni ja/või negatiivse sõelumisena kohaste O-RS rekombinantide tuvastamisel. Näiteks replitseeritakse kloonide kogum valikuliselt CAT-d (mis hõlmab vähemalt üht selektorkoodonit) sisaldavatel kasvuplaatidel, koos või ilma ühe või enama mittelooduslikult kodeeritud aminohappega. Seega loetakse kolooniad, mis kasvavad ainult mittelooduslikult kodeeritud aminohappeid sisaldavatel plaatidel, rekombinantset O-RS-i sisaldavateks. Leiutise ühe aspekti kohaselt varieeritakse selektsiooniagensi (ja/või sõelumisagensi) kontsentratsiooni. Leiutise mõningate aspektide kohaselt on esimene ja teine organism erinevad. Nõnda hõlmab esimene ja/või teine organism valikuliselt järgnevaid: prokarüoot, eukarüoot, imetaja, Escherichia coli, seen, pärm, arhebakter, eubakter, taim, putukas, protist jne. Leiutise muude teostuste korral hõlmab sõelumismarker fluorestsents- või luminestsentssõelumismarkerit või afiinsuspõhist sõelumismarkerit. Leiutise veel ühe teostuse kohaselt hõlmab kogumi sõelumine või selekteerimine (muuhulgas, kuid mitte ainult, negatiivne selekteerimine) aktiivse (valikuliselt mutantse) RS-i leidmiseks: aktiivsete mutantsete RS-ide kogumi isoleerimist positiivsest selektsioonietapist (b); negatiivse selektsiooni või sõelumise markeri sisseviimist, kusjuures negatiivne selektsiooni või sõelumise marker hõlmab vähemalt üht selektorkoodonit (muuhulgas, kuid mitte ainult, toksilist markergeeni, muuhulgas, kuid mitte ainult, ribonukleaas-barnaasi geeni, mis hõlmab vähemalt üht selektorkoodonit); ja aktiivsete (valikuliselt mutantsete) RS-ide kogumi sisseviimist teise organismi mitmesse rakku; ning niisuguste rakkude tuvastamist, mis jäävad ellu või näitavad üles spetsiifilist vastust sõelumisele esimeses keskkonnas, mida pole täiendatud mittelooduslikult kodeeritud aminohappega, kuid ei jää ellu ega näita üles spetsiifilist vastust sõelumisele teises keskkonnas, mida on täiendatud mittelooduslikult kodeeritud aminohappega, mille tulemusena ellujäävad või sõelutud rakud varustatakse vähemalt ühe rekombinantse O-
129 RS-iga, kusjuures see vähemalt üks rekombinantne O-RS on spetsiifiline mittelooduslikult kodeeritud aminohappe suhtes. Leiutise ühe aspekti kohaselt hõlmab vähemalt üks selektorkoodon ligikaudu kaht või enamat selektorkoodonit. Niisugused teostused võivad valikuliselt hõlmata juhtusid, mil vähemalt üks selektorkoodon hõlmab kaht või enamat selektorkoodonit, ning mil esimene ja teine organism on erinevad (muuhulgas, kuid mitte ainult, on iga organism valikuliselt, muuhulgas, kuid mitte ainult, prokarüoot, eukarüoot, imetaja, Escherichia coli, seen, pärm, arhebakter, eubakter, taim, putukas, protist jne.). Samuti hõlmavad leiutise mõningad aspektid seda, kui negatiivse selektsiooni marker hõlmab ribonukleaas-barnaasi geeni (mis hõlmab vähemalt üht selektorkoodonit). Leiutise muud aspektid hõlmavad seda, kui sõelumismarker hõlmab valikuliselt fluorestsents- või luminestsents-sõelumismarkerit või afiinsuspõhist sõelumismarkerit. Leiutise siinkirjeldatud teostuste korral hõlmavad sõelumised ja/või selektsioonid valikuliselt sõelumis- ning/või selektsiooniranguse varieerimist. Leiutise ühe teostuse kohaselt võivad meetodid vähemalt ühe rekombinantse ortogonaalse aminoatsüül-trna süntetaasi tootmiseks (O-RS) täiendavalt hõlmata: (d) vähemalt ühe rekombinantse O-RS-i isoleerimist; (e) teise O-RS (valikuliselt muteeritud) komplekti loomist, mis on tuletatud vähemalt ühest rekombinantsest O-RS-ist; ning (f) etappide (b) ja (c) kordamist muteeritud O-RS-i saamiseni, mis hõlmab O-tRNA eelistusliku aminoatsüüülimise võimet. Etappe (d)-(f) korratakse valikuliselt, muuhulgas, kuid mitte ainult, vähemalt kaks korda. Leiutise ühe aspekti kohaselt saab muteeritud O-RS-i teist komplekti, mis on tuletatud vähemalt ühest rekombinantsest O-RS-ist, valmistada mutageneesi teel, muuhulgas, kuid mitte ainult, juhusliku mutageneesi, kohaspetsiifilise mutageneesi, selle rekombineerimise või kombineerimise teel. Ülalkirjeldatud meetodite korral hõlmab selektsiooni/sõelumisetappide rangus, muuhulgas, kuid mitte ainult, positiivse selektsiooni/sõelumise etapp (b), negatiivse selektsiooni/sõelumise etapp (c), või nii positiivse kui ka negatiivse sõelumise etapid (b) ja (c), valikuliselt selektsiooni/sõelumise ranguse varieerimist. Leiutise veel ühe teostuse kohaselt hõlmavad positiivse selektsiooni/sõelumise etapp (b), negatiivse selektsiooni/sõelumise etapp (c), või nii positiivse kui ka negatiivse sõelumise etapid (b) ja (c) reporteri kasutamist, kusjuures reporter detekteeritakse fluorestsents-aktiveeritud rakkude sorteerimise (FACS) või luminestsentsi abil. Valikuliselt kuvatakse reporter rakupinnal, faagmeetodil vms. ja valitakse afiinsuse või katalüütilise aktiivsuse põhjal, mis
130 128 hõlmab mittelooduslikult kodeeritud aminohapet või selle analoogi. Leiutise ühe teostuse kohaselt kuvatakse muteeritud süntetaas raku pinnal, faagmeetodil vms Meetodid rekombinantse ortogonaalse trna (O-tRNA) valmistamiseks hõlmavad: (a) panga loomist mutantsetest trna-dest, mis on tuletatud vähemalt ühest trna-st, muuhulgas, kuid mitte ainult, supressor-trna-st, esimeselt organismilt; (b) panga selekteerimist (muuhulgas, kuid mitte ainult, negatiivset selekteerimist) või sõelumist (valikuliselt mutantsete) trna-de leidmiseks, mis on aminoatsüülitud aminoatsüül-trna süntetaasi (RS) poolt teisest organismist esimesest organismist pärit RS-i puudumisel, saades nõnda (valikuliselt mutantsete) trna-de kogumi; ning (c) (valikuliselt mutantsete) trna-de kogumi selekteerimist või sõelumist liikmete leidmiseks, mis on aminoatsüülitud sisseviidud ortogonaalse RS-i (O-RS) poolt, saades tulemuseks vähemalt ühe rekombinantse O-tRNA; kusjuures vähemalt üks rekombinantne O-tRNA tuvastab selektorkoodoni ega ole tõhusalt tuvastatav teisest organismist pärit RS-i poolt ning on eelistatavalt aminoatsüülitav O-RS-i poolt. Leiutise Mõne teostuse korral on mainitud vähemalt üks trna supressor-trna ja/või hõlmab unikaalset kolme alusega koodonit looduslike ning/või mittelooduslike alustega või on nonsenss-koodon, haruldane koodon, mittelooduslik koodon, vähemalt nelja alust hõlmav koodon, merevaikkoodon, ookerkoodon, või opaal-stoppkoodon. Leiutise ühe teostuse kohaselt on rekombinantsele O-tRNA-le omane ortogonaalsuse täiustus. Arusaadavaks loetakse seda, et leiutise Mõne teostuse korral imporditakse O-tRNA valikuliselt esimesse organismi teisest organismist modifikatsioonivajaduseta. Leiutise erinevate teostuste korral on esimene ja teine organism kas samad või erinevad ning valitakse valikuliselt muuhulgas, kuid mitte ainult, järgnevate seast: prokarüoodid (muuhulgas, kuid mitte ainult, Methanococcus jannaschii, Methanobacterium thermoautotrophicum, Escherichia coli, Halobacterium jne.), eukarüoodid, imetajad, seened, pärmid, arhebakterid, eubakterid, taimed, putukad, protistid jne. Lisaks aminoatsüülitakse rekombinantne trna valikuliselt mittelooduslikult kodeeritud aminohappega, kusjuures mittelooduslikult kodeeritud aminohape biosünteesitakse in vivo kas looduslikult või geneetilise manipulatsiooni kaudu. Mittelooduslikult kodeeritud aminohape lisatakse valikuliselt söötmele vähemalt esimese või teise organismi jaoks. Leiutise ühe aspekti kohaselt hõlmab panga selekteerimine (muuhulgas, kuid mitte ainult, negatiivne selekteerimine) või sõelumine (valikuliselt mutantsete) trna-de leidmiseks,
131 mis aminoatsüülitakse aminoatsüül-trna süntetaasi poolt (etapp (b)): toksilise markergeeni, kusjuures toksiline markergeen hõlmab vähemalt üht selektorkoodonitest (või geeni, mis toob kaasa toksilise või staatilise agensi või organismi jaoks eluliselt tähtsa geeni tootmise, kui vastav markergeen hõlmab vähemalt üht selektorkoodonit), ning (valikuliselt mutantsete) trna-de panga sisseviimist teisest organismist pärit mitmesse rakku ning ellujäävate rakkude selekteerimist, kusjuures ellujäävad rakud sisaldavad (valikuliselt mutantsete) trna-de kogumit, mis hõlmavad vähemalt üht ortogonaalset trna-d või mittefunktsionaalset trna-d. Ellujäävaid rakke saab valida näiteks võrdlussuhtel põhineva rakutiheduskatse abil. Leiutise veel ühe aspekti kohaselt võib toksiline markergeen hõlmata kaht või enamat selektorkoodonit. Meetodite veel ühe teostuse kohaselt on toksiliseks markergeeniks ribonukleaas-barnaasi geen, kusjuures ribonukleaas-barnaasi geen hõlmab vähemalt üht merevaikkoodonit. Valikuliselt võib ribonukleaas-barnaasi geen sisaldada kaht või enamat merevaikkoodonit. Leiutise ühe teostuse kohaselt võib (valikuliselt mutantsete) trna-de kogumi selekteerimine või sõelumine liikmete leidmiseks, mis aminoatsüülitakse sisseviidud ortogonaalse RS-i (O-RS) poolt, hõlmata: positiivse selektsiooni või sõelumise markergeeni, kusjuures positiivne markergeen hõlmab mõnd ravimresistentsusgeeni (muuhulgas, kuid mitte ainult, β-laktamaasi geeni, mis hõlmab vähemalt üht selektorkoodonitest, näiteks vähemalt üht merevaik-stoppkoodonit) või organismi jaoks eluliselt tähtsat geeni, või geeni, mis toob kaasa toksilise aine detoksifikatsiooni, koos O- RS-iga, ning (valikuliselt mutantsete) trna-de kogumi sisseviimist mitmetesse teisest organismist pärit rakkudesse; ning ellujäävate või sõelutud rakkude tuvastamist, mis on kasvatatud selektsiooni- või sõelumisagensi, muuhulgas, kuid mitte ainult, antibiootikumi juuresolekul, andes seeläbi tulemuseks rakkude kogumi, millel on vähemalt üks rekombinantne trna, kusjuures see vähemalt üks rekombinantne trna aminoatsüülitakse O-RS-i poolt ja sisestab aminohappe positiivse markergeeni poolt kodeeritud translatsioonisaadusesse, vastusena vähemalt ühele selektorkoodonile. Leiutise veel ühe aspekti kohaselt varieeritakse selektsiooniagensi (ja/või sõelumisagensi) kontsentratsiooni.
132 Ette nähakse meetodid spetsiifiliste O-tRNA/O-RS paaride valmistamiseks. Meetodid hõlmavad: (a) panga loomist mutantsetest trna-dest, mis on tuletatud vähemalt ühest trna-st esimeselt organismilt; (b) panga negatiivset selekteerimist või sõelumist (valikuliselt mutantsete) trna-de leidmiseks, mis on aminoatsüülitud aminoatsüül-trna süntetaasi (RS) poolt teisest organismist esimesest organismist pärit RS-i puudumisel, saades nõnda (valikuliselt mutantsete) trna-de kogumi; (c) (valikuliselt mutantsete) trna-de kogumi selekteerimist või sõelumist liikmete leidmiseks, mis on aminoatsüülitud sisseviidud ortogonaalse RS-i (O-RS) poolt, saades tulemuseks vähemalt ühe rekombinantse O-tRNA. See vähemalt üks rekombinantne O-tRNA tuvastab selektorkoodoni ega ole tõhusalt tuvastatav teisest organismist pärit RS-i poolt ning on eelistatavalt aminoatsüülitav O-RS-i poolt. Meetod hõlmab samuti (d) panga loomist (valikuliselt mutantsetest) RS-idest, mis on tuletatud vähemalt ühest aminoatsüül-trna süntetaasist (RS) kolmandalt organismilt; (e) mutantsete RS-ide panga selekteerimist või sõelumist liikmete leidmiseks, mis eelistatavalt aminoatsüülivad vähemalt üht rekombinantset O-tRNA-d mittelooduslikult kodeeritud aminohappe ja loodusliku aminohappega, andes seeläbi tulemuseks aktiivsete (valikuliselt mutantsete) RS-ide kogumi; ning (f) selle kogumi negatiivset selekteerimist või sõelumist aktiivste (valikuliselt mutantsete) RS-ide leidmiseks, mis eelistatavalt aminoatsüülivad vähemalt üht rekombinantset O-tRNA-d mittelooduslikult kodeeritud aminohappe puudumisel, andes seeläbi tulemuseks vähemalt ühe spetsiifilise O-tRNA/O-RS paari, kusjuures see vähemalt üks spetsiifiline O-tRNA/O-RS paar hõlmab vähemalt üht rekombinantset O- RS-i, mis on spetsiifiline mittelooduslikult kodeeritud aminohappe ja vähemalt ühe rekombinantse O-tRNA suhtes. Meetodite abil toodetud spetsiifilised O-tRNA/O-RS paarid on kaasatud. Näiteks võib spetsiifiline O-tRNA/O-RS paar hõlmata muuhulgas, kuid mitte ainult, mutrnatyr-muttyrrs paari nagu mutrnatyr-ss12tyrrs paar, mutrnaleu-mutleurs paari, mutrnathr-mutthrrs paari, mutrnaglu-mutglurs paari vms. Lisaks kuuluvad taoliste meetodite alla juhud, mil esimene ja kolmas organism on samad (muuhulgas, kuid mitte ainult, Methanococcus jannaschii). Käesoleva leiutise alla kuuluvad ka meetodid ortogonaalse trna-trna süntetaasipaari selekteerimiseks teise organismi in vivo translatsioonisüsteemis kasutamise eesmärgil. Need meetodid hõlmavad: esimesest organismist isoleeritud või tuletatud aminoatsüültrna süntetaasi, markergeeni ja trna sisseviimist teiselt organismilt saadud esimesse
133 5 131 rakukomplekti; markergeeni ja trna sisseviimist dublikaat-rakukomplekti teiselt organismilt; ning ellujäävate rakkude selekteerimist esimeses komplektis, mis ei suuda ellu jääda dublikaat-rakukomplektis, või rakkude sõelumist, millel avaldub spetsiifiline sõelumisvastus, kuid millel ei avaldu taolist vastust dublikaat-rakukomplektis, kusjuures esimene komplekt ja dublikaat-rakukomplekt kasvatatakse selektsiooni- või sõelumisagensi juuresolekul, kusjuures ellujäävad või sõelutud rakud hõlmavad ortogonaalset trna-trna süntetaasipaari kasutamiseks teise organismi in vivo translatsioonisüsteemis. Leiutise ühe teostuse kohaselt hõlmab võrdlemine ja selekteerimine või sõelumine in vivo komplementatsioonikatset. Võimalik on selektsioonivõi sõelumisagensi kontsentratsiooni varieerimine. Käesoleva leiutise organismid hõlmavad mitmesuguseid organisme ja mitmesuguseid kombinatsioone. Näiteks võivad käesoleva leiutise meetodite esimene ja teine organism olla samad või erinevad. Leiutise ühe teostuse korral on organismideks valikuliselt prokarüootsed organismid, muuhulgas, kuid mitte ainult, Methanococcus jannaschii, Methanobacterium thermoautotrophicum, Halobacterium, Escherichia coli, A. fulgidus, P. furiosus, P. horikoshii, A. pernix, T. thermophilus vms. Alternatiivselt võivad organismid valikuliselt hõlmata eukarüootseid organisme, muuhulgas, kuid mitte ainult, taimi (muuhulgas, kuid mitte ainult, kõrgemad taimed nagu üheidulehelised või kaheidulehelised), vetikaid, protiste, seeni (muuhulgas, kuid mitte ainult, pärmid jne.), loomi (muuhulgas, kuid mitte ainult, imetajad, putukad, lülijalgsed jne.) vms. Leiutise veel ühe teostuse kohaselt on teiseks organismiks prokarüootne organism, muuhulgas, kuid mitte ainult, Methanococcus jannaschii, Methanobacterium thermoautotrophicum, Halobacterium, Escherichia coli, A. fulgidus, Halobacterium, P. furiosus, P. horikoshii, A. pernix, T. thermophilus vms. Alternatiivselt võib teiseks organismiks olla eukarüootne organism, muuhulgas, kuid mitte ainult, pärm, loomarakk, taimerakk, seen, imetajarakk vms. Leiutise mõningate aspektide kohaselt on esimene ja teine organism erinevad. VI. Mittelooduslike aminohapete asukoht FGF-21 polüpeptiidides 30 Käesolev leiutis näeb ette ühe või enama mitteloodusliku aminohappe kaasamist FGF-21 polüpeptiididesse. Üks mittelooduslikest aminohapetest võib olla kaasatud konkreetsele positsioonile, mis ei katkesta polüpeptiidi aktiivsust. See on saavutatav
134 konservatiivsete asenduste sooritamise teel, muuhulgas, kuid mitte ainult, hüdrofoobsete aminohapete asendamine hüdrofoobsete aminohapetega, kogukate aminohapete asendamine kogukate aminohapetega, hüdrofiilsete aminohapete asendamine hüdrofiilsete aminohapetega ja/või mittelooduslike aminohapete sisestamine kohta, mis pole aktiivsuse jaoks vajalik. Võimalik on mitmesuguste biokeemiliste ja struktuursete lähenemiste rakendamine soovitud kohtade valimiseks mitteloodusliku aminohappega asendamise eesmärgil FGF- 21 polüpeptiidis. Ala tavataseme asjatundjatele peaks olema ilmne, et mittelooduslikult kodeeritud aminohappe kaasamiseks sobib polüpeptiidahela mis tahes positsioon, ning selektsioon võib põhineda ratsionaalsel disainil või juhuslikul valikul mingil kindlal soovitud põhjusel või ilma niisuguse põhjuseta. Soovitud kohtade selektsiooni sihiks võib olla FGF-21 molekuli tootmine, millel on mingi soovitud omadus või aktiivsus, muuhulgas, kuid mitte ainult, agonistid, superagonistid, pöördagonistid, antagonistid, retseptori seondumise modulaatorid, retseptori aktiivsuse modulaatorid, dimeeride või multimeeride moodustumine, aktiivsuse või omaduse muudatuste puudumine algupärase molekuliga võrreldes, või polüpeptiidi ükskõik millise füüsikalise või keemilise omaduse, näiteks lahustuvuse, agregatsiooni või stabiilsuse manipuleerimine. Näiteks on kohad polüpeptiidis, mis on tarvilikud FGF-21 polüpeptiidide bioaktiivsuse jaoks, tuvastatavad punktmutatsioonanalüüsi, alaniiniga skaneerimise, küllastava mutageneesi ja sõelumise teel bioaktiivsuse leidmiseks, või tehnika tasemest teadaolevate homoloogiga skaneerimise meetodite abil. Jäägid, mis on FGF-21 bioaktiivsuse seisukohast kriitilise tähtsusega, jäägid, mis on seotud farmatseutilise stabiilsusega, antikeha epitoobid või retseptori või hepariini sidumisjäägid võivad olla muteerunud. Patendid nr. US 5,580,723; US 5,834,0; US 6,013,478; US 6,428,954; ja US 6,451,561 kirjeldavad meetodeid polüpeptiidide nagu FGF-21 struktuuri ning talitluse süstemaatiliseks analüüsiks aktiivsete domeenide tuvastamise teel, mis mõjutavad polüpeptiidi aktiivsust sihtaine suhtes. Jäägid peale nende, mille kriitiline tähtsus bioaktiivsuse jaoks on tuvastatud alaniini või homoloogiga skaneeriva mutageneesi teel, võivad olla headeks kandidaatideks mittelooduslikult kodeeritud aminohappega asendamise seisukohast, olenevalt polüpeptiidi taotletavast aktiivsusest. Alternatiivselt võivad kohad, mille kriitiline tähtsus bioaktiivsuse jaoks on tuvastatud, olla headeks kandidaatideks ka mittelooduslikult kodeeritud aminohappega asendamise seisukohast, jällegi olenevalt polüpeptiidi
135 5 133 taotletavast aktiivsusest. Veel üheks alternatiiviks oleks järjestikuste asendamiste sooritamine mitteloodusliku aminohappega polüpeptiidahela igal positsioonil ja polüpeptiidi aktiivsustele avaldatava mõju seire. Ala tavataseme asjatundjatele peaks olema ilmne, et käesolevas leiutises kasutamiseks sobib mis tahes vahend, tehnika või meetod positsiooni valimiseks mitteloodusliku aminohappega asendamise eesmärgil ükskõik millises polüpeptiidis. Käesolevas taotluses esitletud meetodite abil on juba kogutud hulgaliselt teavet ja potentsiaalsed ning soodsad mutatsioonikohad on leitud ja neid on edukalt katsetatud, nagu kirjeldatud selle spetsifikatsiooni edasistes osades toodud näidetes. Lisateabe leidmine mutantide struktuuri ja aktiivsuse kohta, isegi juhtudel, mis hõlmavad mõningaid üksikasju formuleerimise ning/või testimise kohta, mida pole konkreetselt kirjeldatud näidetes FGF-21 polüpeptiidide osas, mis sisaldavad deletsioone, saab samuti uurida valgu nende piirkondade kindlaksmääramiseks, mis on tõenäoliselt tolerantsed mittelooduslikult kodeeritud aminohappega asendamise suhtes. Sarnasel viisil saab proteaasi digestiooni ja monokloonseid antikehi kasutada FGF-21 nende piirkondade tuvastamiseks, mis vastutavad FGF-21 retseptori sidumise eest. Kui jäägid, mis pole tõenäoliselt tolerantsed mittelooduslikult kodeeritud aminohapetega asendamise suhtes, on elimineeritud, saab uurida väljapakutud asenduste mõju igaühel allesjäänud positsioonidest. Võimalik on mudelite loomine muude FGF perekonna liikmete ja FGF retseptorite kolmemõõtmelistest kristallstruktuuridest. Protein Data Bank (PDB, juurdepääsetav Internetis aadressil kujutab endast tsentraliseeritud andmebaasi, mis sisaldab andmeid nukleiinhapete ja valkude suurte molekulide kolmemõõtmelise struktuuri kohta. Võimalik on mudelite loomine polüpeptiidide sekundaarse ja tertsiaarse struktuuri uurimise kaudu, kui kolmemõõtmelise struktuuri andmed pole saadaval. Nõnda oskavad ala tavataseme asjatundjad hõlpsasti tuvastada aminohapete positsioonid, mida saab asendada mittelooduslikult kodeeritud aminohapetega. Mõne teostuse korral hõlmavad leiutisekohased FGF-21 polüpeptiidid üht mittelooduslikku aminohapet, mis paikneb valgu piirkonnas, mis ei häiri polüpeptiidi struktuuri. 30 Mittelooduslikult kodeeritud aminohapete kaasamist võimaldavate jääkide näited võivad hõlmata jääke, mis asuvad väljaspool potentsiaalseid retseptori sidumise piirkondi, mis
136 võivad olla täielikult või osaliselt hüdrofiilsed, mille vesiniku sidumise interaktsioonid naaberjääkidega on minimaalsed või puuduvad, mis võivad olla minimaalselt eksponeeritud reaktiivsete naaberjääkide suhtes, mis võivad paikneda ühel või enamal eksponeeritud pinnal, mis võivad olla kohaks või kohtadeks, mis paiknevad kõrvuti teise FGF-21-ga või muu molekuli või selle fragmendiga, mis võivad paikneda piirkondades, mis on äärmiselt paindlikud või struktuuri poolest jäigad, nagu ennustab FGF-21 kolmemõõtmeline, sekundaarne, tertsiaarne või kvaternaarne struktuur, seotud või sidumata selle retseptoriga või seotud/sidumata muu bioloogiliselt aktiivse molekuliga, või mis võivad moduleerida FGF-21 enda või üht või enamat FGF-21 sisaldava dimeeri või multimeeri konformatsiooni kogu struktuuri paindlikkuse või jäikuse muutmise teel. FGF valkude perekonnal on ühine β-ristikuleht- või β-lehtstruktuur, nagu näitab kristallograafia (Harmer et al., Biochemistry 43: (04)). Ala tavataseme asjatundjale peaks olema arusaadav, et FGF-21 niisugune analüüs võimaldab selle kindlaksmääramist, millised aminohappejäägid on pinnal eksponeeritud võrdluses aminohappejääkidega, mis paiknevad valgu tertsiaarse struktuuri sees. Seetõttu näeb üks käesoleva leiutise teostustest ette mittelooduslikult kodeeritud aminohappe asendamist aminohappega, mis kujutab endast pinnal eksponeeritud jääki. Üks mittelooduslikult kodeeritud aminohape kaasatakse järgnevale positsioonile FGF-21- s: 8 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mittelooduslik aminohape on FGF-21-s positsioonil 8 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Ühe teostuse kohaselt on mittelooduslik aminohape FGF-21-s positsioonil 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 ja muu mittelooduslik aminohape ühel järgnevatest positsioonidest: enne positsiooni 1 (s.t. N- otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133,
137 , 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 ja muu mittelooduslik aminohape ühel järgnevatest positsioonidest: 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 91 ja positsioonil 131 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 91 ja positsioonil 77 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 91 ja positsioonil 8 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 131 ja positsioonil 8 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 131 ja positsioonil 77 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 131 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 8 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 77 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 72 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 87 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 86 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7) seotuna. Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 126 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 1 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on mittelooduslik aminohape positsioonil 83 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7).
138 Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 ja üks või mitu muud mittelooduslikku aminohapet ühel või enamal järgnevatest positsioonidest: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 ja üks või mitu muud mittelooduslikku aminohapet ühel või enamal järgnevatest positsioonidest: 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 ja muu mittelooduslik aminohape ühel või enamal järgnevatest positsioonidest: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 ja muu mittelooduslik aminohape ühel või enamal järgnevatest positsioonidest: 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7).
139 Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 ja kaks või enam muud mittelooduslikku aminohapet ühel või enamal järgnevatest positsioonidest: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 ja kaks või enam muud mittelooduslikku aminohapet ühel või enamal järgnevatest positsioonidest: 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 77 ja muu mittelooduslik aminohape ühel või enamal järgnevatest positsioonidest: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174. FGF-21 või perekonna FGF liikme(te) kristallstruktuuri ja selle interaktsiooni FOP retseptoriga uurimine võib näidata, millistel konkreetsetel aminohapetel on kõrvalahelad, mis on solvendile täielikult või osaliselt juurdepääsetavad. Mittelooduslikult kodeeritud aminohappe kõrvalahel nendel positsioonidel võib olla suunatud valgu pinnalt eemale, solvendi sisse. Mittelooduslik aminohape ühel neist positsioonidest on seotud vees
140 lahustuva polümeeriga, sealhulgas positsioon 8 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mõne teostuse korral on mittelooduslik aminohape kahel või enamal neist positsioonidest seotud vees lahustuva polümeeriga, muuhulgas, kuid mitte ainult, positsioonid enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mõne teostuse korral on mittelooduslik aminohape ühel või enamal neist positsioonidest seotud vees lahustuva polümeeriga, muuhulgas, kuid mitte ainult, positsioonid, 52, 117, 126, 131, 162, 87, 77, 83, 72, 69, 79, 91, 96, 8 ja 1 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mõne teostuse korral on mittelooduslik aminohape ühel või enamal neist positsioonidest seotud vees lahustuva polümeeriga:, 52, 77, 117, 126, 131, 162, (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mõne teostuse korral on mittelooduslik aminohape ühel või enamal neist positsioonidest seotud vees lahustuva polümeeriga: 87, 77, 83, 72 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mõne teostuse korral on mittelooduslik aminohape ühel või enamal neist positsioonidest seotud vees lahustuva polümeeriga: 69, 79, 91, 96, 8 ja 1 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mõne teostuse korral on mittelooduslik aminohape ühel või enamal neist positsioonidest seotud vees lahustuva polümeeriga: 91, 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt, kui mittelooduslik aminohape asub aminohappe positsioonil 91 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7), on mittelooduslik aminohape seotud vees lahustuva polümeeriga.
141 Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 seotud vees lahustuva polümeeriga ja muu mittelooduslik aminohape on ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 ja üks või mitu muud mittelooduslikku aminohapet ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on seotud vees
142 lahustuva polümeeriga: 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 91 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 131, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 seotud vees lahustuva polümeeriga ja muu mittelooduslik aminohape on ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163,
143 , 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 ja üks või mitu muud mittelooduslikku aminohapet ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 91, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on seotud vees lahustuva polümeeriga: 91, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126,
144 , 128, 129, 130, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 131 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 91, 8, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 seotud vees lahustuva polümeeriga ja muu mittelooduslik aminohape on ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 ja üks või mitu muud mittelooduslikku aminohapet ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 91, 131, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47,
145 , 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on seotud vees lahustuva polümeeriga: 91, 131, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78,79, 80, 81,82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 8 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 91, 131, 77, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 77 seotud vees lahustuva polümeeriga ja muu mittelooduslik aminohape on ühel järgnevatest
146 positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 77 ja üks või mitu muud mittelooduslikku aminohapet ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 91, 131, 8, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 77 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on ühel järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 77 seotud vees lahustuva polümeeriga ja üks või mitu muud mittelooduslikku aminohapet on seotud vees lahustuva polümeeriga: 91, 131, 8, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt
147 on üks mittelooduslik aminohape positsioonil 77 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: enne positsiooni 1 (s.t. N-otsal), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (s.t. valgu karboksüülotsal) (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Veel ühe teostuse kohaselt on üks mittelooduslik aminohape positsioonil 77 seotud vees lahustuva polümeeriga ja kaks või enam muud mittelooduslikku aminohapet on kahel või enamal järgnevatest positsioonidest, ning need mittelooduslikud aminohapped on seotud vees lahustuva polümeeriga: 91, 131, 8, 72, 87, 86, 126, 1, 83, 146, 135, 96 ja 36 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7). Mittelooduslik aminohape positsioonil 8 (SEQ ID number 1 või vastavad aminohapped SEQ ID numbritega 2-7) on seotud vees lahustuva polümeeriga. Mitmesuguseid erinevaid mittelooduslikult kodeeritud aminohappeid saab asendada või kaasata etteantud positsioonile FGF-21 polüpeptiidis. Üldjuhul valitakse konkreetne mittelooduslikult kodeeritud aminohape kaasamiseks FGF-21 polüpeptiidi või muu FGF perekonna liikme (koos selle retseptoriga) kolmemõõtmelise kristallstruktuuri uurimise, konservatiivsete asenduste eelistamise (s.t. arüülipõhised mittelooduslikult kodeeritud aminohapped nagu p-atsetüülfenüülalaniin või O-propargüültürosiin asendavad Phe, Tyr või Trp), ning spetsiifilise konjugatsioonikeemia põhjal, mida soovitakse FGF-21 polüpeptiidi sisse viia (nt. 4-asidofenüülalaniini sisseviimine juhul, kui eesmärgiks on Huisgeni [3+2] tsükloliitumise saavutamine vees lahustuva polümeeriga, mis kannab alküünfragmenti, või amiidsideme moodustamine vees lahustuva polümeeriga, mis kannab arüülestrit, mis omakorda hõlmab fosfiinfragmenti).
148 Leiutise ühe teostuse kohaselt hõlmab meetod täiendavalt mitteloodusliku aminohappe kaasamist valku, kusjuures mittelooduslik aminohape hõlmab esimest reaktiivset rühma; ning valgu kokkupuutesse viimist molekuliga (muuhulgas, kuid mitte ainult, märgise, värvaine, polümeeri, vees lahustuva polümeeri, polüetüleenglükooli derivaadi, fotoristsiduja, radionukliidi, tsütotoksilise ühendi, ravimi, afiinsusmärgise, fotoafiinsusmärgise, reaktiivse ühendi, vaigu, teise valgu või polüpeptiidi või polüpeptiidi analoogi, antikeha või antikeha fragmendi, metalli kelaatori, kofaktori, rasvhappe, süsivesiku, polünukleotiidi, DNA, RNA, antisenss-polünukleotiidi, sahhariidi, vees lahustuva dendrimeeri, tsüklodekstriini, pärssiva ribonukleiinhappe, biomaterjali, nanoosakese, spin-märgise, fluorofoori, metalli sisaldava fragmendi, radioaktiivse fragmendi, uudse funktsionaalse rühma, teiste molekulidega kovalentselt või mittekovalentselt seotud rühma, fotolabiilse fragmendi, aktiiniumiga kiiritamise teel ergastatava fragmendi, fotoisomeeritava fragmendi, biotiini, biotiini derivaadi, biotiini analoogi, rasket aatomit hõlmava fragmendi, keemiliselt lõhustatava rühma, fotolõhustatava rühma, pikendatud kõrvalahela, süsinikuga seotud suhkru, redoks-aktiivse aine, amino-tiohappe, toksilise fragmendi, isotoobiga märgistatud fragmendi, biofüüsikalise sondi, fosforestsentsrühma, kemiluminestsentsrühma, elektrontiheda rühma, magnetilise rühma, interkaleeruva rühma, kromofoori, energiaülekandeaine, bioaktiivse aine, detekteeritava märgise, väikse molekuli, kvantpunkti, nanotransmitteri, radionukleotiidi, radiotransmitteri, neutronipüüduri või ülaltoodute ükskõik millise kombinatsiooni, või mis tahes muu soovitava ühendi või ainega), mis hõlmab teist rektiivset rühma. Esimene reaktiivne rühm reageerib teise reaktiivse rühmaga molekuli kinnitamiseks mitteloodusliku aminohappe külge [3+2] tsükloliitumise kaudu. Leiutise ühe teostuse kohaselt on esimene reaktiivne rühm alkünüül- või asidofragment ja teine reaktiivne rühm asido- või alkünüülfragment. Näiteks on esimeseks reaktiivseks rühmaks alkünüülfragment (muuhulgas, kuid mitte ainult, mittelooduslik aminohape p- propargüloksüfenüülalaniin) ja teiseks reaktiivseks rühmaks asidofragment. Veel ühe näite korral on esimeseks reaktiivseks rühmaks asidofragment (muuhulgas, kuid mitte ainult, mittelooduslik aminohape p-asido-l-fenüülalaniin) ja teiseks reaktiivseks rühmaks alkünüülfragment. Leiutise Mõne teostuse korral kombineeritakse mittelooduslikult kodeeritud aminohappe asendust (asendusi) muude liitmiste, asenduste või deletsioonidega FGF-21 polüpeptiidis
149 FGF-21 polüpeptiidi muude bioloogiliste tunnusjoonte mõjutamiseks. Leiutise Mõne teostuse korral võivad muud liitmised, asendused või deletsioonid suurendada FGF-21 polüpeptiidi stabiilsust (muuhulgas, kuid mitte ainult, vastupanu proteolüütilisele lagundamisele) või tõsta FGF-21 polüpeptiidi afiinsust oma retseptori suhtes. Leiutise Mõne teostuse korral võivad muud liitmised, asendused või deletsioonid suurendada FGF- 21 polüpeptiidi farmatseutilist stabiilsust. Leiutise Mõne teostuse korral võivad muud liitmised, asendused või deletsioonid tõsta FGF-21 polüpeptiidi lahustuvust (muuhulgas, kuid mitte ainult, ekspressiooni korral E. coli-s või muudes peremeesrakkudes). Leiutise Mõne teostuse korral võivad liitmised, asendused või deletsioonid tõsta polüpeptiidi lahustuvust ekspressiooni järel E. coli-s või muudes rekombinantsetes peremeesrakkudes. Leiutise Mõne teostuse korral valitakse kohad asendamiseks looduslikult kodeeritud või mitteloodusliku aminohappega lisaks veel ühele kohale mitteloodusliku aminohappe kaasamiseks, mis toob kaasa polüpeptiidi lahustuvuse tõuse ekspressiooni järel E. coli-s või muudes rekombinantsetes peremeesrakkudes. Leiutise Mõne teostuse korral hõlmavad FGF-21 polüpeptiidid veel üht liitmist, asendamist või deletsiooni, mis moduleerib afiinsust FGF-21 polüpeptiidi retseptori, seostusvalkude või seotud ligandi suhtes, moduleerib signaali transduktsiooni sidumise järel FGF-21 retseptoriga, moduleerib ringluse poolväärtusaega, moduleerib vabastamist või biosaadavust, lihtsustab puhastumist või täiustab/muudab konkreetset manustamisviisi. Leiutise Mõne teostuse korral hõlmavad FGF-21 polüpeptiidid liitmist, asendamist või deletsiooni, mis suurendab FGF-21 variandi afiinsust oma retseptori suhtes. Sarnasel moel võivad FGF-21 polüpeptiidid hõlmata keemilise või ensümaatilise lõhustamise järjestusi, proteaasi lõhustamise järjestusi, reaktiivseid rühmi, antikeha siduvaid domeene (muuhulgas, kuid mitte ainult, FLAG või poly-his) või muid afiinsuspõhiseid järjestusi (muuhulgas, kuid mitte ainult, FLAG, poly- His, GST jne.) või seotud molekule (muuhulgas, kuid mitte ainult, biotiin), mis täiustavad detekteerimist (muuhulgas, kuid mitte ainult, GFP), puhastumist, transporti läbi kudede või rakumembraanide, eelravimi vabastamist või aktiveerimist, FGF-21 suuruse vähendamist või polüpeptiidi muid tunnusjooni. Mõne teostuse korral annab mittelooduslikult kodeeritud aminohappe asendamine tulemuseks FGF-21 antagonisti. Mõne teostuse korral asendatakse või liidetakse mittelooduslikult kodeeritud aminohape retseptori sidumisse kaasatud piirkonnas. Mõne teostuse korral asendatakse või liidetakse mittelooduslikult kodeeritud aminohape
150 5 148 hepariini sidumisse kaasatud piirkonnas. Mõne teostuse korral hõlmavad FGF-21 antagonistid vähemalt üht asendust, mis kutsub esile FGF-21 toimimise antagonistina. Mõne teostuse korral hõlmab FGF-21 antagonist mittelooduslikult kodeeritud aminohapet, mis on seotud vees lahustuva polümeeriga, mis paikneb FGF-21 molekuli retseptorit siduvas piirkonnas. Üks aminohape asendatakse ühe mittelooduslikult kodeeritud aminohappega. Mõningatel juhtudel seotakse üks mittelooduslikult kodeeritud jääk ühe või mitme väiksema molekulmassiga lineaarse või hargnenud PEG-ga, võimendades seeläbi sidumisafiinsust ja võrreldavat seerumi poolväärtusaega võrreldes variandiga, mis kinnitub üksiku, suurema molekulmassiga PEG külge. VII. Ekspressioon mitteeukarüootides ja eukarüootides 30 Kloonitud FGF-21 polünukleotiidi kõrge ekspressioonitaseme saavutamiseks subkloonitakse leiutisekohast FGF-21 polüpeptiidi kodeerivaid polünukleotiide üldjuhul ekspressioonivektorisse, mis sisaldab tugevat promootorit, transkriptsiooni/translatsiooni terminaatorit ja (valku kodeeriva nukleiinhappe korral) ribosoomi sidumiskohta translatsiooni initsieerimise jaoks. Sobivad bakteriaalsed promootorid on ala tavataseme asjatundjatele teada ja neid kirjeldavad nt. Sambrook el al. ja Ausubel et al. Bakteriaalsed ekspressioonisüsteemid leiutisekohaste FGF-21 polüpeptiidide ekspresseerimiseks on saadaval muuhulgas, kuid mitte ainult, järgnevates: E. coli, Bacillus sp., Pseudomonas fluorescens, Pseudomonas aeruginosa, Pseudomonas putida ja Salmonella (Palva et al., Gene 22: (1983); Mosbach et al., Nature 302: (1983)). Komplektid taoliste ekspressioonisüsteemide jaoks on kaubanduslikult saadaval. Eukarüootsed ekspressioonisüsteemid imetajarakkude, pärmi ja putukarakkude jaoks on ala tavataseme asjatundjatele teada ning samuti kaubanduslikult saadaval. Juhtudel, mil ortogonaalseid trna-sid ja aminoatsüül-trna süntetaase (kirjeldatakse eespool) kasutatakse leiutisekohaste FGF-21 polüpeptiidide ekspresseerimiseks, valitakse peremeesrakud ekspressiooniks nende võime põhjal kasutada ortogonaalseid komponente. Näitlikud peremeesrakud hõlmavad grampositiivseid baktereid (muuhulgas, kuid mitte ainult, B. brevis, B. subtilis või Streptomyces) ja gramnegatiivseid baktereid (E. coli, Pseudomonas fluorescens, Pseudomonas aeruginosa, Pseudomonas putida), samuti pärmi
151 149 ning muid eukarüootseid rakke. O-tRNA/O-RS paare hõlmavaid rakke saab kasutada siinkirjeldatud viisil Käesoleva leiutise kohane eukarüootne peremeesrakk või mitteeukarüootne peremeesrakk pakub võimet valkude sünteesimiseks, mis hõlmavad mittelooduslikke aminohappeid suurtes kasulikes kogustes. Ühe aspekti kohaselt hõlmab kompositsioon valikuliselt, kuid mitte ainult, vähemalt mikrogrammi, vähemalt 50 mikrogrammi, vähemalt 75 mikrogrammi, vähemalt 0 mikrogrammi, vähemalt 0 mikrogrammi, vähemalt 0 mikrogrammi, vähemalt 500 mikrogrammi, vähemalt 1 milligrammi, vähemalt milligrammi, vähemalt 0 milligrammi, vähemalt 1 grammi või rohkem valku, mis hõlmab mittelooduslikku aminohapet, või kogust, mis on saavutatav in vivo valgutootmismeetodite abil (üksikasjalik teave rekombinantse valgu tootmise ja puhastamise kohta on leiutises esitatud). Teise aspekti kohaselt sisaldub valk valikuliselt kompositsioonis kontsentratsiooni juures, muuhulgas, kuid mitte ainult, vähemalt mikrogrammi valku liitri kohta, vähemalt 50 mikrogrammi valku liitri kohta, vähemalt 75 mikrogrammi valku liitri kohta, vähemalt 0 mikrogrammi valku liitri kohta, vähemalt 0 mikrogrammi valku liitri kohta, vähemalt 0 mikrogrammi valku liitri kohta, vähemalt 500 mikrogrammi valku liitri kohta, vähemalt 1 milligramm valku liitri kohta või vähemalt milligrammi valku liitri kohta või rohkem, muuhulgas, kuid mitte ainult, rakulüsaadis, puhvris, farmatseutilises puhvris või muus vedelas suspensioonis (muuhulgas, kuid mitte ainult, mahu juures, muuhulgas, kuid mitte ainult, vahemikus alates u. 1 nl kuni u. 0 l või üle selle). Suurte valgukoguste tootmine (muuhulgas, kuid mitte ainult, suuremate kui üldjuhul võimalik muude meetodite abil, muuhulgas, kuid mitte ainult, in vitro translatsioon) eukarüootses rakus koos vähemalt ühe mitteloodusliku aminohappega on leiutise üheks eripäraks. Käesoleva leiutise kohane eukarüootne peremeesrakk või mitteeukarüootne peremeesrakk pakub võimet valkude biosünteesimiseks, mis hõlmavad mittelooduslikke aminohappeid suurtes kasulikes kogustes. Näiteks on võimalik mittelooduslikku aminohapet hõlmavate valkude tootmine muuhulgas, kuid mitte ainult, järgnevate kontsentratsioonide juures: vähemalt µg/liiter, vähemalt 50 µg/liiter, vähemalt 75 µg/liiter, vähemalt 0 µg/liiter, vähemalt 0 µg/liiter, vähemalt 0 µg/liiter või vähemalt 500 µg/liiter, vähemalt 1 mg/liiter, vähemalt 2 mg/liiter, vähemalt 3 mg/liiter, vähemalt 4 mg/liiter, vähemalt 5 mg/liiter, vähemalt 6 mg/liiter, vähemalt 7 mg/liiter, vähemalt 8 mg/liiter, vähemalt 9
152 0 mg/liiter, vähemalt mg/liiter, vähemalt, 30, 40, 50, 60, 70, 80, 90, 0, 0, 300, 400, 500, 600, 700, 800, 900 mg/liiter, 1g/liiter, 5 g/liiter, g/liiter või rohkem valku rakuekstraktis, rakulüsaadis, kultiveerimissöötmes, puhvris jms./vms. 5 I. Ekspressioonisüsteemid. Kultuur ja isoleerimine FGF-21 polüpeptiidide ekspressioon on võimalik ükskõik millistes sobivates ekspressioonisüsteemides, muuhulgas nt. pärm, putukarakud, imetajarakud ning bakterid. Näitlike ekspressioonisüsteemide kirjeldus järgneb allpool. Pärm Siin kasutatuna hõlmab mõiste pärm ükskõik milliseid erinevatest pärmiseentest, mis on võimelised FGF-21 polüpeptiidi kodeeriva geeni ekspresseerimiseks. Niisugused pärmid hõlmavad muuhulgas, kuid mitte ainult, askosporogeenseid pärme (Endomycetales), basidiosporogeenseid pärme ja pärme, mis kuuluvad rühma Fungi imperfecti (Blastomycetes). Askosporogeensed pärmid jagunevad kahte sugukonda, Spermophthoraceae ja Saccharomycetaceae. Viimane neist koosneb neljast alamsugukonnast Schizosaccharomycoideae (nt. perekond Schizosaccharomyces), Nadsonioideae, Lipomycoideae ja Saccharomycoideae (nt. perekonnad Pichia, Kluyveromyces ning Saccharomyces). Basidiosporogeensed pärmid hõlmavad perekondi Leucosporidium, Rhodosporidium, Sporidiobolus, Filobasidium ja Filobasidiella. Rühma Fungi Imperfecti (Blastomycetes) kuuluvad pärmid jagunevad kahte sugukonda, Sporobolomycetaceae (nt. perekonnad Sporobolomyces ja Bullera) ning Cryptococcaceae (nt. perekond Candida). Käesoleva leiutise raames kasutamise seisukohast pakuvad erilist huvi liigid, mis kuuluvad perekondadesse Pichia, Kluyveromyces, Saccharomyces, Schizosaccharomyces, Hansenula, Torulopsis ja Candida, muuhulgas, kuid mitte ainult, P. pastoris, P. guillerimondii, S. cerevisiae, S. carlsbergensis, S. diastaticus, S. douglasii, S. kluyveri, S, norbensis, S. oviformis, K. lactis, K. fragilis, C. albicans, C. maltosa ning H. polymorpha. Sobiva pärmi valimine FGF-21 polüpeptiidide ekspresseerimiseks kuulub ala tavataseme asjatundja oskuste piiridesse. Peremeespärmide valimisel ekspressiooniks võivad sobivad peremehed hõlmata pärme, millele on omased näiteks hea sekretsioonivõime, madal
153 1 proteolüütiline aktiivsus, hea lahustuva valgu tootmine ja üldine vastupidavus. Pärmid on üldjuhul saadaval mitmesugustest allikatest, muuhulgas, kuid mitte ainult, Yeast Genetic Stock Center, Department of Biophysics and Medical Physics, University of California (Berkeley, CA) ning American Type Culture Collection ( ATCC ) (Manassas, VA) Mõiste peremeespärm või peremees-pärmirakk hõlmab pärme, mida saab kasutada või on kasutatud rekombinantsete vektorite või muu ülekande-dna retsipiendina. Mõiste hõlmab algupärase peremees-pärmiraku järglaskonda, mis on üle võtnud rekombinantsed vektorid või muu ülekande-dna. Arusaadavaks loetakse seda, et ühe vanemraku järglaskond ei pruugi olla morfoloogia, genoomika või üldise DNA komplemendi poolest täiesti identne algupärase vanemaga juhusliku või tahtliku mutatsiooni tõttu. Vanemraku järglaskond, mis on vanemale piisavalt sarnane asjakohase omaduse põhjal, näiteks FGF- 21 polüpeptiidi kodeeriva nukleotiidjärjestuse olemasolu, kuulub antud kontekstis järglaskonna määratluse alla. Välja on töötatud ekspressiooni- ja transformatsioonivektorid, sealhulgas kromosoomivälised replikonid või integratsioonivektorid, transformatsiooniks paljudesse peremeespärmidesse. Ekspressioonivektorid on välja töötatud näiteks järgnevate jaoks: S. cerevisiae (Sikorski et al., GENETICS (1989) 122:19; Ito et al., J. BACTERIOL. (1983) 3:163; Hinnen et al., PROC. NATL. ACAD. SCI. USA (1978) 75:1929); C. albicans (Kurtz et al., MOL. CELL. BIOL. (1986) 6:142); C. maltosa (Kunze et al., J. BASIC MICROBIOL. (1985) :141); H. polymorpha (Gleeson et al., J. GEN. MICROBIOL. (1986) 132:3459; Roggenkamp et al., MOL. GENETICS AND GENOMICS (1986) 2:302); K. fragilis (Das et al., J. BACTERIOL. (1984) 8:1165); K. lactis (De Louvencourt et al., J. BACTERIOL. (1983) 4:737; Van den Berg et al., BIOTECHNOLOGY (NY) (1990) 8:135); P. guillerimondii (Kunze et al., J. BASIC MICROBIOL. (1985) :141); P. pastoris (patendid nr. US 5,324,639, US 4,929,555 ja US 4,837,148; Cregg et al., MOL. CELL. BIOL. (1985) 5:3376); Schizosaccharomyces pombe (Beach et al., NATURE (1982) 300:706); ning Y. lipolytica; A. nidulans (Ballance et al., BIOCHEM. BIOPHYS. RES. COMMUN. (1983) 112:284-89; Tilbum et al., GENE (1983) 26:5-221; ja Yelton et al., PROC. NATL. ACAD. SCI. USA (1984) 81: ); A. niger (Kelly and Hynes, EMBO J. (1985) 4: ); T. reesia (EP ); ning filamentsed seened nagu näiteks Neurospora, Penicillium, Tolypocladium (WO 91/00357).
154 Pärmivektorite kontrolljärjestused on ala tavataseme asjatundjatele teada ja hõlmavad muuhulgas, kuid mitte ainult, promootorpiirkondi geenidelt nagu alkoholi dehüdrogenaas (ADH) (EP ); enolaas; glükokinaas; glükoos-6-fosfaadi isomeraas; glütseeraldehüüd-3-fosfaat-dehüdrogenaas (GAP või GAPDH); heksokinaas; fosfofruktokinaas; 3-fosfoglütseraadi mutaas ning püruvaadi kinaas (PyK) (EP ). Pärmi PHO5 geen, mis kodeerib happelist fosfataasi, võib samuti pakkuda kasulikke promootorjärjestusi (Miyanohara et al., PROC. NATAL. ACAD. SCI USA (1983) 80:1). Muud sobivad promootorjärjestused kasutamiseks koos peremeespärmidega võivad hõlmata promootoreid 3-fosfoglütseraadi kinaasi jaoks (Hitzeman et al., J. BIOL. CHEM. (1980) 5:173) ning muidki glükolüütilisi ensüüme nagu püruvaadi dekarboksülaas, triosefosfaadi isomeraas ning fosfoglükoosi isomeraas (Holland et al., BIOCHEMISTRY (1978) 17:4900; Hess et al., J. ADV. ENZYME REG. (1969) 7:149). Indutseeritavad pärmipromootorid, millel on kasvutingimustega kontrollitud transkriptsiooni lisaeelis, võivad hõlmata alkoholi dehüdrogenaasi 2; isotsütokroomi C; happelise fosfataasi; metallotioniini; glütseeraldehüüd-3-fosfaadi dehüdrogenaasi; lämmastiku ainevahetusega seotud lagundavate ensüümide; ning maltoosi ja galaktoosi kasutamise eest vastutavate ensüümide promootorpiirkondi. Pärmis ekspresseerimisega seoses kasutamiseks sobivaid vektoreid ja promootoreid on täiendavalt kirjeldatud allikas EP Koos pärmipromootoritega võib kasutada ka pärmivõimendajaid. Lisaks võivad pärmipromootoritena toimida sünteetilised promootorid. Näiteks on võimalik pärmipromootori ülesvoolu aktiveerimisjärjestuste (UAS) ühendamine mõne muu pärmipromootori transkriptsiooni aktiveerimispiirkonnaga, luues sünteetilise hübriidpromootori. Taoliste hübriidpromootorite näited hõlmavad ADH regulatoorset järjestust, mis on seotud GAP transkriptsiooni aktiveerimispiirkonnaga. Vt. patendid nr. US 4,880,734 ja US 4,876,197. Muud hübriidpromootorite näited hõlmavad promootoreid, mis koosnevad ADH2, GAL4, GAL või PHO5 geenide regulatoorsetest järjestustest, mis on kombineeritud glükolüütilise ensüümi geeni nagu GAP või PyK transkriptsioonilise aktiveerimise piirkonnaga. Vt. EP Lisaks võib pärmipromootor hõlmata mitte-pärmist pärinevaid looduslikke promootoreid, mis on võimelised pärmi RNA polümeraasi sidumiseks ja transkriptsiooni initsieerimiseks. Muudeks kontrollielementideks, mis võivad hõlmata osa pärmi ekspressioonivektoritest, on muuhulgas terminaatorid, näiteks GAPDH-st või enolaasi geenidest (Holland et al., J.
155 BIOL. CHEM. (1981) 6:1385). Lisaks on pärmi jaoks sobilikuks replikatsiooni allikaks 2 µ plasmiidallikas. Pärmis kasutamiseks sobivaks selektsioonigeeniks on trp1 geen, mis sisaldub pärmi plasmiidis. Vt. Tschumper et al., GENE (1980) :7; Kingsman et al., GENE (1979) 7:141. trp1 geen võimaldab selektsioonimarkeri mutantse pärmitüve jaoks, millel puudub võime kasvada trüptofaanis. Sarnaselt täiendatakse Leu2-puudulikke pärmitüvesid (ATCC,622 või 38,626) teadaolevate plasmiidide poolt, mis kannavad Leu2 geeni. Meetodid eksogeense DNA sisseviimiseks peremeespärmidesse on ala tavataseme asjatundjatele teada ning hõlmavad üldjuhul, kuid mitte ainult, kas sfäroplastide või terviklike peremees-pärmirakkude, mida on töödeldud leelisekatioonidega, transformatsiooni. Näiteks saab pärmi transformatsiooni teostada kooskõlas meetodiga, mida on kirjeldatud allikates Hsiao et al., PROC. NATL. ACAD. SCI. USA (1979) 76:3829 ja Van Solingen et al., J. BACT. (1977) 130:946. Samas võib kasutada muidki meetodeid DNA viimiseks rakkudesse, näiteks nukleaarne injektsioon, elektroporatsioon või protoplastide liitmine, nagu seda üldiselt kirjeldatakse allikas SAMBROOK ET AL., MOLECULAR CLONING: A LAB. MANUAL (01). Seejärel on võimalik peremeespärmirakkude kultiveerimine standardsete tehnikate abil, mis on teada ala tavataseme asjatundjatele. Ala tavataseme asjatundjatele on teada muudki meetodid heteroloogsete valkude ekspresseerimiseks peremees-pärmirakkudes. Vt. üldiselt patendipublikatsioon nr. US , patendid nr. US 6,361,969; US 6,312,923; US 6,183,985; US 6,083,723; US 6,017,731; US 5,674,706; US 5,629,3; US 5,602,034 ja US 5,089,398; uuesti läbivaadatud USA patendid nr. RE37,343 ja RE35,749; PCT avaldatud patenditaotlused WO 99/07862; WO 98/378 ja WO 98/26080; Euroopa patenditaotlused EP ; EP ; EP ; WO 90/277; EP ; EP ; EP ning EP Vt. ka Gellissen et al., ANTONIE VAN LEEUWENHOEK (1992) 62(1-2):79-93; Romanos et al., YEAST (1992) 8(6): ; Goeddel, METHODS IN ENZYMOLOGY (1990) 185:3-7. Peremees-pärmitüvesid saab kasvatada fermenterites võimendusetapi vältel, kasutades standardseid söötme lisamisega fermenteerimismeetodeid, mis on ala tavataseme asjatundjatele teada. Vastavaid fermentatsioonimeetodeid saab kohandada võtma arvesse
156 erinevusi konkreetse peremeespärmi süsiniku kasutamistee või ekspressiooni juhtimisviisi osas. Näiteks võib Saccharomyces-peremeespärmi fermentatsioon eeldada ühekordset glükoosi etteannet, kompleks-lämmastikuallikat (nt. kaseiini hüdrolüsaadid) ning mitme vitamiiniga rikastamist. Seevastu metülotroopne pärm P. pastoris võib vajada glütserooli, metanooli ja mikromineraalide etteannet, ent vaid lihtsaid ammoonium-(lämmastiku-)sooli optimaalseks kasvuks ning ekspressiooniks. Vt. nt. patent nr. US 5,324,639; Elliott et al., J. PROTEIN CHEM. (1990) 9:95; ja Fieschko et al., BIOTECH. BIOENG. (1987) 29:1113. Niisugustel fermentatsioonimeetoditel võib aga olla teatud ühiseid omadusi, mis ei sõltu rakendatavast peremeespärmi tüvest. Näiteks on võimalik kasvu piirava toitaine, üldjuhul süsiniku, lisamine fermenterisse võimendusfaasi kestel maksimaalse kasvu saavutamiseks. Lisaks rakendavad fermentatsioonimeetodid reeglina fermentatsioonisöödet, mis on kujundatud sisaldama süsiniku, lämmastiku, basaalsete soolade, fosfori ja muude mikroelementide (vitamiinid, mikromineraalid ning soolad jne.) piisavaid koguseid. Pichia korral kasutamiseks sobivate fermentatsioonisöötmete näiteid on kirjeldatud patentides nr. US 5,324,639 ja US 5,231,178. WO 05/ kirjeldab FGF-21 ekspressiooni pärmis. Bakuloviirusega nakatatud putukarakud Mõiste peremeesputukas või peremeesputukarakk viitab putukale, mida saab kasutada või on kasutatud rekombinantsete vektorite või muu ülekande-dna retsipiendina. Mõiste hõlmab algupärase peremeesputukaraku transfektsioonijärgset järglaskonda. Arusaadavaks loetakse seda, et ühe vanemraku järglaskond ei pruugi olla morfoloogia, genoomika või üldise DNA komplemendi poolest täiesti identne algupärase vanemaga juhusliku või tahtliku mutatsiooni tõttu. Vanemraku järglaskond, mis on vanemale piisavalt sarnane asjakohase omaduse põhjal, näiteks FGF-21 polüpeptiidi kodeeriva nukleotiidjärjestuse olemasolu, kuulub antud kontekstis järglaskonna määratluse alla. Sobivate putukarakkude valimine FGF-21 polüpeptiidide ekspresseerimiseks on ala tavataseme asjatundjatele teada. Tehnika tasemes on põhjalikult kirjeldatud ja kaubanduslikult saadaval mitmed putukaliigid, sealhulgas Aedes aegypti, Bombyx mori, Drosophila melanogaster, Spodoptera frugiperda ning Trichoplusia ni. Putukarakkude valimisel ekspressiooniks võivad sobivad peremehed hõlmata rakke, millele on muuhulgas omased hea sekretsioonivõime, madal proteolüütiline aktiivsus ja üldine vastupidavus.
157 5 Putukad on üldjuhul saadaval mitmesugustest allikatest, muuhulgas, kuid mitte ainult, Insect Genetic Stock Center, Department of Biophysics and Medical Physics, University of California (Berkeley, CA) ning American Type Culture Collection ( ATCC ) (Manassas, VA) Üldjuhul hõlmavad bakuloviirusega nakatatud putuka ekspressioonisüsteemi komponendid ülekandevektorit, tavaliselt bakteriaalset plasmiidi, mis sisaldab nii bakuloviiruse genoomi fragmenti kui ka sobivat restriktsioonikohta ekspresseeritava heteroloogse geeni sisestamiseks; metsik-tüüpi bakuloviirust järjestustega, mis on homoloogsed bakuloviirus-spetsiifilise fragmendiga ülekandevektoris (see võimaldab heteroloogse geeni homoloogset rekombinatsiooni bakuloviiruse genoomi); ning sobivaid peremees-putukarakke ja söötmeid. Materjalid, meetodid ja tehnikad, mida kasutatakse vektorite konstrueerimisel, rakkude transfekteerimisel, plaakide valimisel, rakkude kasvatamisel söötmes jms. on valdkonnas teada ning olemas on vastavaid tehnikaid kirjeldavad juhendid. Heteroloogse geeni sisestamise järel ülekandevektorisse transfekteeritakse vektor ja metsik-tüüpi viiruse genoom peremees-putukarakku, kus leiab aset vektori ning viiruse genoomi rekombinatsioon. Pakitud rekombinantne viirus ekspresseeritakse ja rekombinantsed plaagid tuvastatakse ning puhastatakse. Materjale ja meetodeid bakuloviiruse/putukaraku ekspressioonisüsteemide jaoks turustab komplekti kujul näiteks Invitrogen Corp. (Carlsbad, CA). Need tehnikad on ala tavataseme asjatundjatele üldiselt teada ja neid kirjeldatakse täies ulatuses allikas SUMMERS AND SMITH, TEXAS AGRICULTURAL EXPERIMENT STATION BULLETIN NO. 55 (1987). Vt. ka RICHARDSON, 39 METHODS IN MOLECULAR BIOLOGY: BACULOVIRUS EXPRESSION PROTOCOLS (1995); AUSUBEL ET AL., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (1994); KING AND POSSEE, THE BACULOVIRUS SYSTEM: A LABORATORY GUIDE (1992); ning O'REILLY ET AL., BACULOVIRUS EXPRESSION VECTORS: A LABORATORY MANUAL (1992). Mitmesuguste heteroloogsete valkude tootmine bakuloviiruse/putukaraku ekspressioonisüsteemide abil on ala tavataseme asjatundjatele teada. Vt. nt. patendid nr. US 6,368,8; US 6,342,216; US 6,338,846; US 6,261,805; US 6,245,528, US 6,2,060; US 6,183,987; US 6,168,932; US 6,126,944; US 6,096,304; US 6,013,433; US 5,965,393;
158 US 5,939,285; US 5,891,676; US 5,871,986; US 5,861,279; US 5,858,368; US 5,843,733; US 5,762,939; US 5,753,2; US 5,605,827; US 5,583,023; US 5,571,709; US 5,516,657; US 5,290,686; WO 02/06305; WO 01/90390; WO 01/27301; WO 01/05956; WO 00/55345; WO 00/032; WO 99/51721; WO 99/45130; WO 99/317; WO 99/5; WO 99/09193; WO 97/26332; WO 96/29400; WO 96/496; WO 96/06161; WO 95/672; WO 93/03173; WO 92/16619; WO 92/02628; WO 92/01801; WO 90/14428; WO 90/078; WO 90/066; WO 90/02186; WO 90/056; WO 89/038; WO 89/037; WO 88/ Bakuloviiruse/putukaraku ekspressioonisüsteemides kasulikud vektorid on tehnika tasemest teada ja hõlmavad näiteks putuka ekspressiooni- ning ülekandevektoreid, mis on tuletatud bakuloviiruse Autographa californica nukleaarse polühedroosi viirusest (AcNPV), mis kujutab endast abistajast sõltumatut viiruse ekspressioonivektorit. Antud süsteemist tuletatud viiruse ekspressioonivektorid kasutavad tavaliselt võimsat polühedriini geeni promootorit heteroloogsete geenide ekspressiooni käivitamiseks. Vt. üldiselt O Reilly ET AL., BACULOVIRUS EXPRESSION VECTORS: A LABORATORY MANUAL (1992). Võõrgeeni bakuloviiruse genoomi sisestamise eel koondatakse ülalkirjeldatud komponendid, sealhulgas promootor, liider (soovi korral), huvipakkuv kodeerimisjärjestus ning transkriptsiooni lõpetav järjestus üldjuhul vahepealsesse ümberpaigutuskonstrukti (ülekandevektorisse). Vahepealseid ümberpaigutuskonstrukte hoitakse sageli alal replikonis, näiteks täiendavas kromosoomelemendis (nt. plasmiidid), mis on võimeline stabiilseks püsimiseks peremehes, näiteks bakterites. Replikonil on replikatsioonisüsteem, mis võimaldab selle alalhoidmist sobivas peremehes kloonimiseks ja amplifitseerimiseks. Veelgi konkreetsemalt võib plasmiid sisaldada polühedriini polüadenüleerimissignaali (Miller, ANN. REV. MICROBIOL. (1988) 42:177) ja prokarüootset ampitsilliini resistentsuse (amp) geeni ning replikatsiooniallikat selektsiooniks ja paljundamiseks E. coli-s. Üheks üldkasutatavaks ülekandevektoriks võõrgeenide viimisel AcNPV-sse on pac373. Välja on töötatud palju muidki vektoreid, mis on ala asjatundjatele teada, sealhulgas näiteks pvl985, mis muudab polühedriini alguskoodoni ATG-st ATT-ks ja sisestab BamHI kloonimissaidi 32 aluspaari ATT-st allavoolu. Vt. Luckow and Summers, VIROLOGY 170:31 (1989). Muudeks kaubanduslikult saadaolevateks vektoriteks on näiteks PBlueBac4.5/V5-His; pbluebachis2; pmelbac; pbluebac4.5 (Invitrogen Corp., Carlsbad, CA). Heteroloogse
159 geeni sisestamise järel kaastransfekteeritakse ülekandevektor ja metsikut tüüpi bakuloviiruse genoom peremees-putukarakku. Meetodid heteroloogse DNA sisestamiseks soovitud kohale bakuloviiruses on tehnika tasemest teada. Vt. SUMMERS AND SMITH, TEXAS AGRICULTURAL EXPERIMENT STATION BULLETIN NO. 55 (1987); Smith et al., MOL. CELL. BIOL. (1983) 3:26; Luckow and Summers, VIROLOGY (1989) 170:31. Näiteks võib sisestamine aset leida geeni nagu polühedriini geen homoloogse kahekordse ristsiire-rekombinatsiooni teel; samuti võib sisestamine aset leida restriktsiooniensüümi saidile, mis on lülitatud soovitud bakuloviiruse geeni koosseisu. Vt. Miller et al., BIOESSAYS (1989) 11(4):91. Võimalik on transfektsiooni sooritamine elektroporatsiooni teel. Vt. TROTTER AND WOOD, 39 METHODS IN MOLECULAR BIOLOGY (1995); Mann and King, J. GEN. VIROL. (1989) 70:3501. Alternatiivselt võib liposoome kasutada putukarakkude transfekteerimiseks rekombinantse ekspressioonivektori ja bakuloviirusega. Vt. nt. Liebman et al., BIOTECHNIQUES (1999) 26(1):36; Graves et al., BIOCHEMISTRY (1998) 37:6050; Nomura et al., J. BIOL. CHEM. (1998) 273(22):13570; Schmidt et al., PROTEIN EXPRESSION AND PURIFICATION (1998) 12:323; Siffert et al., NATURE GENETICS (1998) 18:45; TILKINS ET AL., CELL BIOLOGY: A LABORATORY HANDBOOK (1998); Cai et al., PROTEIN EXPRESSION AND PURIFICATION (1997) :263; Dolphin et al., NATURE GENETICS (1997) 17:491; Kost et al., GENE (1997) 190:139; Jakobsson et al., J. BIOL. CHEM. (1996) 271:223; Rowles et al., J. BIOL. CHEM. (1996) 271(37):22376; Reverey et al., J. BIOL. CHEM. (1996) 271(39):23607-; Stanley et al., J. BIOL. CHEM. (1995) 270:4121; Sisk et al., J. VIROL. (1994) 68(2):766; ja Peng et al., BIOTECHNIQUES (1993) 14(2):274. Kaubanduslikult saadaolevateks liposoomideks on näiteks Cellfectin ja Lipofectin (Invitrogen, Corp., Carlsbad, CA). Lisaks on võimalik kaltsiumfosfaadi transfektsiooni kasutamine. Vt. TROTTER AND WOOD, 39 METHODS IN MOLECULAR BIOLOGY (1995); Kitts, NAR (1990) 18(19):5667; ning Mann and King, J. GEN. VIROL. (1989) 70:3501. Bakuloviiruse ekspressioonivektorid sisaldavad tavaliselt bakuloviiruse promootorit. Bakuloviiruse promootor on mis tahes DNA järjestus, mis on võimeline siduma bakuloviiruse RNA polümeraasi ja initsieerima kodeerimisjärjestuse (nt. struktuurse geeni) allavoolu (3') transkriptsiooni mrna-sse. Promootoril peab olema transkriptsiooni
160 initsiatsioonipiirkond, mis tavaliselt paikneb proksimaalselt kodeerimisjärjestuse 5' otsa suhtes. Transkriptsiooni initsiatsioonipiirkond hõlmab üldjuhul RNA polümeraasi sidumiskohta ja transkriptsiooni initsiatsioonikohta. Bakuloviiruse promootor võib samuti omada teist domeeni, mida nimetatakse võimendajaks (enhanseriks) ja mis paikneb olemasolu korral tavaliselt struktuurse geeni suhtes distaalselt. Ekspressioon võib olla kas reguleeritud või konstitutiivne. Struktuursed geenid, mida külluslikult transkribeeritakse infektsioonitsükli hilisemates järkudes, on promootorjärjestustena eriti kasulikud. Näited hõlmavad järjestusi, mis on tuletatud viiruse polühedroni valku kodeerivast geenist (FRIESEN ET AL., The Regulation of Baculovirus Gene Expression, THE MOLECULAR BIOLOGY OF BACULOVIRUSES (1986); EP ja ), ning geeni, mis kodeerib valku p (Vlak et al., J. GEN. VIROL. (1988) 69:765). Vastloodud bakuloviiruse ekspressioonivektor pakitakse nakkavasse rekombinantsesse bakuloviirusesse ning hiljem kasvanud plaake saab puhastada tehnikate abil, mis on ala tavataseme asjatundjatele teada. Vt. Miller et al., BIOESSAYS (1989) 11(4):91; SUMMERS AND SMITH, TEXAS AGRICULTURAL EXPERIMENT STATION BULLETIN NO. 55(1987). Välja on töötatud rekombinantse bakuloviiruse ekspressioonivektorid mitme putukaraku nakatamiseks. Rekombinantsed bakuloviirused on välja töötatud muuhulgas järgnevate jaoks: Aedes aegypti (ATCC No. CCL-1), Bombyx mori (ATCC No. CRL-89), Drosophila melanogaster (ATCC No. 1963), Spodoptera frugiperda ja Trichoplusia ni. Vt. Wright, NATURE (1986) 321:718; Carbonell et al., J. VIROL. (1985) 56:3; Smith et al., MOL. CELL. BIOL. (1983) 3:26. Vt. üldiselt Fraser et al., IN VITRO CELL. DEV. BIOL. (1989) :2. Konkreetsemalt hõlmavad bakuloviiruse ekspressioonivektorisüsteemide korral kasutatavad rakutüved üldjuhul muuhulgas, kuid mitte ainult, Sf9 (Spodoptera frugiperda) (ATCC No. CRL-1711), Sf21 (Spodoptera frugiperda) (Invitrogen Corp., Cat. No (Carlsbad, CA)), Tri-368 (Trichopulsia ni) ning High-Five BTI-TN-5B1-4 (Trichopulsia ni). Kaubanduslikult on saadaval rakud ja söötmed heteroloogsete polüpeptiidide nii vahetuks kui ka sulandekspressiooniks bakuloviiruses/ekspressioonis, ning rakkude kultiveerimise tehnoloogia on ala tavataseme asjatundjate jaoks üldtuntud.
161 E. Coli, Pseudomonase liigid ja muud prokarüoodid Bakteriaalse ekspressiooni tehnikad on ala tavataseme asjatundjatele teada. Saadaval on lai valik vektoreid kasutamiseks bakter-peremeestes. Vektoriteks võivad olla ühekoopialised või madalad või kõrged mitmekoopialised vektorid. Vektorid võivad olla kasutusel kloonimiseks ja/või ekspresseerimiseks. Võttes arvesse rohket vektoreid käsitlevat kirjandust, paljude vektorite kaubanduslikku saadavalolekut ja isegi käsiraamatute olemasolu, kus kirjeldatakse vektoreid ning nende restriktsioonikaarte ja karakteristikuid, puudub vajadus antud teemat siin põhjalikumalt käsitleda. On hästi teada, et vektorid hõlmavad tavaliselt selektsiooni võimaldavaid markereid, millised markerid võivad soodustada resistentsust tsütotoksiliste ainete suhtes, prototroofsust või immuunsust. Sageli on olemas mitu markerit, mis võimaldavad erinevaid karakteristikuid. Bakteriaalne promootor on mis tahes DNA järjestus, mis on võimeline siduma bakteriaalse RNA polümeraasi ja initsieerima kodeerimisjärjestuse (nt. struktuurse geeni) allavoolu (3') transkriptsiooni mrna-sse. Promootoril peab olema transkriptsiooni initsieerimispiirkond, mis tavaliselt paikneb proksimaalselt kodeerimisjärjestuse 5' otsa suhtes. Transkriptsiooni initsiatsioonipiirkond hõlmab üldjuhul RNA polümeraasi sidumiskohta ja transkriptsiooni initsiatsioonikohta. Bakteriaalsel promootoril võib olla ka teine, operaatoriks nimetatav domeen, mis võib kattuda külgneva RNA polümeraasi sidumiskohaga, kus algab RNA süntees. Operaator võimaldab negatiivset reguleeritud (indutseeritavat) transkriptsiooni, sest geeni repressorvalk võib seostuda operaatoriga ja sedakaudu pärssida konkreetse geeni transkriptsiooni. Konstitutiivne ekspressioon võib aset leida negatiivsete regulatoorsete elementide (nagu operaator) puudumisel. Lisaks on võimalik positiivse regulatsiooni saavutamine geeni aktivaatorvalgu sidumisjärjestuse poolt, mis olemasolu korral on tavaliselt proksimaalne (5') RNA polümeraasi sidumisjärjestusega. Geeni aktivaatorvalgu näiteks on kataboliit-aktivaatorvalk (CAP), mis aitab algatada lac operoni transkriptsiooni Escherichia coli-s (E. coli) [Raibaud et al., ANNU. REV. GENET. (1984) 18:173]. Eeltoodust tulenevalt võib reguleeritud ekspressioon olla kas positiivne või negatiivne, võimendades või piirates transkriptsiooni. Eriti kasulikeks promootorjärjestusteks on järjestused, mis kodeerivad metaboolse raja ensüüme. Näited hõlmavad promootorjärjestusi, mis on tuletatud suhkrut metaboliseerivatest ensüümidest nagu galaktoos, laktoos (lac) [Chang et al., NATURE (1977) 198:56] ja maltoos. Täiendavad näited hõlmavad promootorjärjestusi, mis on
162 tuletatud biosünteetilistest ensüümidest nagu trüptofaan (trp) [Goeddel et al., NUC. ACIDS RES. (1980) 8:4057; Yelverton et al., NUCL. ACIDS RES. (1981) 9:731; patent nr. US 4,738,921; EP publikatsioonid nr ja ]. β-galaktosidaasi (bla) promootorsüsteem [Weissmann (1981) The cloning of interferon and other mistakes. Interferon 3 (Ed. I. Gresser)], bakteriofaag lambda PL [Shimatake et al., NATURE (1981) 292:128] ja T5 [patent nr. US 4,689,406] promootorsüsteemid võimaldavad samuti kasulikke promootorjärjestusi. Käesoleva leiutise eelistatud meetodid rakendavad võimsaid promootoreid nagu T7 promootor FGF-21 polüpeptiidide indutseerimiseks kõrgetel tasemetel. Niisuguste vektorite näited on ala tavataseme asjatundjatele teada ja hõlmavad pet29 seeriat Novagenilt ning ppop vektoreid, mida kirjeldab WO99/ Taolised ekspressioonisüsteemid toodavad peremehes kõrgeid FGF-21 polüpeptiidide tasemeid, kahjustamata sealjuures peremeesraku elumust või kasvuparameetreid. Veel üheks tehnika tasemest teadaolevaks vektoriks on pet19 (Novagen). Lisaks toimivad bakteriaalsete promootoritena sünteetilised promootorid, milliseid looduses ei leidu. Näiteks on võimalik ühe bakteriaalse või bakteriofaagi promootori transkriptsiooni aktiveerimisjärjestuste ühendamine teise bakteriaalse või bakteriofaagi promootori operon-järjestustega, luues sünteetilise hübriidpromootori [patent nr. US 4,551,433]. Näiteks tac promootor on hübriidne trp-lac promootor, mis hõlmab nii trp promootori kui ka lac operoni järjestusi, mida reguleerib lac repressor [Amann et al., GENE (1983) :167; de Boer et al., PROC. NATL. ACAD. SCI. (1983) 80:21]. Lisaks võib bakteriaalne promootor hõlmata mittebakteriaalse päritoluga looduslikke promootoreid, mis on võimelised bakteriaalse RNA polümeraasi sidumiseks ja transkriptsiooni initsieerimiseks. Samuti on võimalik mittebakteriaalse päritoluga loodusliku promootori ühendamine ühilduva RNA polümeraasiga mõningate geenide kõrgete ekspressioonitasemete saavutamiseks prokarüootides. Ühendatud promootorisüsteemi üheks näiteks on bakteriofaagi T7 RNA polümeraas/promootorsüsteem [Studier et al., J. MOL. BIOL. (1986) 189:113; Tabor et al., Proc Natl. Acad. Sci. (1985) 82:74]. Lisaks võib hübriidne promootor koosneda bakteriofaagi promootorist ja E. coli operaatorpiirkonnast (EP publikatsioon nr ). Lisaks toimivale promootorjärjestusele tuleb võõrgeenide ekspresseerimisel prokarüootides kasuks tõhus ribosoomi sidumiskoht. E. coli korral nimetatakse ribosoomi
163 sidumiskohta Shine-Dalgarno (SD) järjestuseks, see hõlmab initsiatsioonikoodonit (ATG) ja 3-9 nukleotiidi pikkust järjestust, mis asub initsiatsioonikoodonist 3-11 nukleotiidi ülesvoolu [Shine et al., NATURE (1975) 4:34]. Eeldatavalt soodustab SD järjestus mrna sidumist ribosoomiga aluste paardumise teel SD järjestuse ja 3' ning E. coli 16S rrna vahel [Steitz et al. Genetic signals and nucleotide sequences in messenger RNA, Biological Regulation and Development: Gene Expression (Ed. R. F. Goldberger, 1979)]. Ekspresseerimaks eukarüootseid geene ja prokarüootseid geene nõrga ribosoomi sidumiskohaga [Sambrook et al. Expression of cloned genes in Escherichia coli, Molecular Cloning: A Laboratory Manual, 1989]. Mõiste peremeesbakter või peremees-bakterirakk hõlmab baktereid, mida saab kasutada või on kasutatud rekombinantsete vektorite või muu ülekande-dna retsipiendina. Mõiste hõlmab algupärase peremees-bakteriraku transfektsioonijärgset järglaskonda. Arusaadavaks loetakse seda, et ühe vanemraku järglaskond ei pruugi olla morfoloogia, genoomika või üldise DNA komplemendi poolest täiesti identne algupärase vanemaga juhusliku või tahtliku mutatsiooni tõttu. Vanemraku järglaskond, mis on vanemale piisavalt sarnane asjakohase omaduse põhjal, näiteks FGF-21 polüpeptiidi kodeeriva nukleotiidjärjestuse olemasolu, kuulub antud kontekstis järglaskonna määratluse alla. Sobiva peremeesbakteri valimine FGF-21 polüpeptiidide ekspresseerimiseks on ala tavataseme asjatundjatele teada. Peremeesbakterite valimisel ekspressiooniks võivad sobivad peremehed hõlmata baktereid, millele on muuhulgas omased hea inklusioonikeha moodustamise võime, madal proteolüütiline aktiivsus ja üldine vastupidavus. Peremeesbakterid on üldjuhul saadaval mitmesugustest allikatest, muuhulgas, kuid mitte ainult, Bacterial Genetic Stock Center, Department of Biophysics and Medical Physics, University of California (Berkeley, CA) ning American Type Culture Collection ( ATCC ) (Manassas, VA). Tööstusliku/farmatseutilise fermentatsiooni raames kasutatakse üldjuhul K-tüvedest (nt. W31) või B-tüvedest (e.g. BL21) tuletatud peremeesbaktereid. Need tüved on eriti kasulikud seoses sellega, et nende kasvuparameetrid on ülihästi teada ja vastupidavad. Lisaks on kõnealused tüved mittepatogeensed, mis on ohutuse ja keskkonnakaitsega seotud põhjustel kaubanduslikult tähtis. Muudeks sobivate E. coli peremeeste näideteks on muuhulgas, kuid mitte ainult, BL21, DHB tüved või nende derivaadid. Käesoleva leiutise meetodite veel ühe teostuse kohaselt on E. coli peremees proteaas-miinus-tüvi, muuhulgas, kuid mitte ainult, OMP- ja
164 LON-. Peremeesrakutüveks võib olla mõni Pseudomonas liik, muuhulgas, kuid mitte ainult, Pseudomonas fluorescens, Pseudomonas aeruginosa ja Pseudomonas putida. Pseudomonas fluorescens biovar 1, määratud tüvi MB1, on teadaolevalt kasulik rekombinantse tootmise seisukohalt ja saadaval terapeutiliste valkude tootmisprotsesside jaoks. Pseudomonas e ekspressioonisüsteemide näited hõlmavad süsteemi, mida peremeestüvena pakub The Dow Chemical Company (Midland, MI, juurdepääsetav Internetis aadressil Kui rekombinantne peremeesrakutüvi on tuvastatud (ehk ekspressioonikonstrukt on peremeesrakku viidud ja kohase ekpressioonikonstruktiga peremeesrakud on isoleeritud), kultiveeritakse rekombinantset peremeesrakutüve tingimuste juures, mis sobivad FGF-21 polüpeptiidide tootmiseks. Nagu peaks olema ilmne ala asjatundja jaoks, sõltub rekombinantse peremeesrakutüve kultiveerimismeetod kasutatava ekspressioonikonstrukti olemusest ja peremeesraku identiteedist. Rekombinantseid peremeestüvesid kultiveeritakse tavaliselt meetodite abil, mis on ala tavataseme asjatundjatele teada. Rekombinantseid peremeesrakke kultiveeritakse üldjuhul vedelas söötmes, mis sisaldab süsiniku, lämmastiku ja anorgaaniliste soolade omastatavaid allikaid ning sisaldab valikuliselt vitamiine, aminohappeid, kasvufaktoreid ja muid valgulisi kultiveerimislisandeid, mis on ala tavataseme asjatundjatele teada. Vedel sööde peremeesrakkude kasvatamiseks võib valikuliselt sisaldada antibiootikume või seenevastaseid aineid soovimatute mikroorganismide ja/või ühendite kasvu ärahoidmiseks, muuhulgas, kuid mitte ainult, ekspressioonivektorit sisaldavate peremeesrakkude jaoks valitavaid antibiootikume. Võimalik on rekombinantsete peremeesrakkude kultiveerimine partiidena või pidevalt, kas rakkude kogumise (juhul kui FGF-21 polüpeptiid akumuleerub rakusiseselt) või kultuuri supernatandi kogumise teel kas partiidena või pidevalt. Prokarüootsetes peremeesrakkudes tootmise seisukohalt on eelistatavad partiikaupa kultiveerimine ja rakkude kogumine. Käesoleva leiutise kohased FGF-21 polüpeptiidid puhastatakse tavaliselt ekspressiooni järel rekombinantsetes süsteemides. FGF-21 polüpeptiidi saab puhastada peremeesrakkudest või söötmest mitmesuguste tehnika tasemest teadaolevate meetodite abil. Bakter-peremeesrakkudes toodetud FGF-21 polüpeptiidid võivad olla raskesti lahustuvad või lahustumatud (inklusioonikehade kujul). Käesoleva leiutise ühe teostuse
165 kohaselt on FGF-21 polüpeptiidis hõlpsasti teostatavad aminohappeasendused, mis valitakse rekombinantselt toodetud valgu lahustuvuse suurendamise eesmärgil, kasutades ära siinkirjeldatud ja tehnika tasemest teadaolevaid meetodeid. Lahustumatu valgu korral on võimalik valgu kogumine peremeesraku lüsaatidest tsentrifuugimise teel, millele võib järgneda rakkude homogeenimine. Raskestilahustuva valgu korral on võimalik ühendite, muuhulgas, kuid mitte ainult, polüetüleenimiini (PEI) lisamine osaliselt lahustuva valgu sadestumise esilekutsumiseks. Seejärel saab sadestunud valku tsentrifuugimise teel mugavalt koguda. Võimalik on rekombinantsete peremeesrakkude lõhustamine või homogeenimine inklusioonikehade vabastamiseks rakkude seest, kasutades mitmesuguseid ala tavataseme asjatundjatele teadaolevaid meetodeid. Peremeesraku lõhustamiseks või homogeenimiseks võib rakendada hästituntud tehnikaid, muuhulgas, kuid mitte ainult, ensümaatilist raku lõhustamist, ultrahelitöötlust, homogeenimist Dounce i homogenisaatori abil või kõrgsurvega lõhustamist. Leiutisekohase meetodi ühe teostuse kohaselt rakendatakse kõrgsurvega lõhustamise tehnikat E. coli peremeesrakkude lõhustamiseks FGF-21 polüpeptiidide inklusioonikehade vabastamiseks. FGF-21 polüpeptiidide inklusioonikehade käsitsemisel võib eelistatav olla homogeenimisaja minimeerimine korduste raames inklusioonikehade saagise maksimeerimiseks ilma kadudeta tegurite tõttu nagu solubilisatsioon, mehhaaniline fragmentatsioon või proteolüüs. Seejärel on võimalik lahustumatu või sadestunud FGF-21 polüpeptiidi solubiliseerimine, kasutades ükskõik millist arvukatest tehnika tasemest teadaolevatest sobivatest solubiliseerimisagensitest. FGF-21 polüpeptiidi saab solubiliseerida uurea või guanidiinhüdrokloriidiga. Solubiliseeritud FGF-21 polüpeptiidi ruumala tuleks minimeerida, võimaldamaks suurte partiide tootmist mugavalt hallatavate partiisuuruste juures. See tegur võib osutuda tähtsaks suuremastaabilise tööstusliku tootmise korral, millega seoses on võimalik rekombinantse peremehe kasvatamine partiidena, mille maht ulatub tuhandete liitriteni. Lisaks tuleks FGF-21 polüpeptiidi suuremastaabilise kaubandusliku tootmise korral (eelkõige inimtervishoiu kontekstis) võimalusel vältida kangeid kemikaale, mis võivad kahjustada seadmestikku ja mahutit või valgusaadust ennast. Käesoleva leiutise meetodi raames on demonstreeritud, et FGF-21 polüpeptiidi inklusioonikehade solubiliseerimisel võib kangema denatureeriva vahendi (guanidiin-hüdrokloriidi) asemel kasutada pehmemat denatureerivat vahendit (uureat). Uurea kasutamine vähendab oluliselt FGF-21 polüpeptiidi tootmis- ja puhastusprotsessi käigus kasutatava roostevabast terasest
166 164 seadmestiku kahjustamise ohtu, tagades samas FGF-21 polüpeptiidi inklusioonikehade tõhusa solubiliseerimise Lahustuva FGF-21 valgu korral on võimalik FGF-21 sekretsioon periplasmaatilisse ruumi või söötmesse. Näiteks sekreteeriti FGF-21 W31-B2 rakkude periplasmaatilisse ruumi konstrukte kodeerivate plasmiidide kasutamise teel, sealhulgas kaheksa erinevat liiderjärjestust, muuhulgas SEQ ID numbritega 39-44, ning nende transformeerimise teel W31-B2 rakkudesse, rakke kasvatati seejärel 37 C juures kuni OD saavutas u. 0.8, misjärel ekspressioon indutseeriti 0,01% arabinoosiga. Viie tunni pärast valmistati kultuuridest periplasmaatilise vabastuse proovid ja rakendati neid geelides (FIG 33) üldise ekspressiooni (lüsaadid kokku) ning periplasmaatiliste sekretsioonide (lahustuv fraktsioon) tuvastamiseks. Lisaks võib peremeesrakkude tsütoplasmas sisalduda lahustuv FGF-21. Soovitatav võib olla lahustuva FGF-21 kontsentreerimine enne puhastusetappide sooritamist. Lahustuva FGF-21 kontsentreerimiseks näiteks rakulüsaatidest või söötmest võib kasutada ala tavataseme asjatundjatele teadaolevaid standardseid tehnikaid. Lisaks võib ala tavataseme asjatundjatele teadaolevaid standardseid tehnikaid kasutada peremeesrakkude lõhustamiseks ja lahustuva FGF-21 vabastamiseks peremeesrakkude periplasmaatilisest ruumist või tsütoplasmast. FGF-21 polüpeptiidi tootmisel sulandvalguna on võimalik sulandjärjestuse eemaldamine. Sulandjärjestuse eemaldamine on saavutatav ensümaatilise või keemilise lõhustamise teel. Sulandjärjestuste ensümaatiline eemaldamine on teostatav ala tavataseme asjatundjatele teadaolevate meetodite abil. Sulandjärjestuse eemaldamiseks rakendatava ensüümi valiku määrab ära sulandi olemus ja reaktsioonitingimused määratletakse lähtuvalt valitud ensüümist, ala tavataseme asjatundjatele ilmsel viisil. Keemiline lõhustamine on saavutatav ala tavataseme asjatundjatele teadaolevate reaktiivide, muuhulgas, kuid mitte ainult, tsüanogeenbromiidi, TEV proteaasi ja muude reaktiivide abil. Lõhustatud FGF-21 polüpeptiidi saab lõhustatud sulandjärjestusest puhastada ala tavataseme asjatundjatele teadaolevate meetodite abil. Vastavad meetodid määratakse kindlaks sulandjärjestuse ja FGF-21 polüpeptiidi olemusest ning omadustest lähtuvalt, ala tavataseme asjatundjale ilmsel viisil. Puhastusmeetodid võivad hõlmata muuhulgas, kuid mitte ainult,
167 165 suuruskromatograafiat, hüdrofoobse interaktsiooni kromatograafiat, ioonvahetuskromatograafiat või dialüüsi või eelnevate ükskõik millist kombinatsiooni Samuti on võimalik FGF-21 polüpeptiidi puhastamine DNA eemaldamiseks valgulahusest. Võimalik on DNA eemaldamine ükskõik millise tehnika tasemest teadaoleva sobiva meetodi abil, näiteks sadestamine või ioonvahetuskromatograafia, kuid selle eemaldamine võib aset leida sadestamise teel nukleiinhappe sadestiga, näiteks, kuid mitte ainult, protamiinsulfaadiga. FGF-21 polüpeptiidi võib sadestatud DNA-st eraldada standardsete hästituntud meetodite abil, muuhulgas, kuid mitte ainult, tsentrifuugimine või filtreerimine. Peremehe nukleiinhappe molekulide eemaldamine on tähtsaks teguriks olukorras, kus FGF-21 polüpeptiidi soovitakse kasutada inimeste ravimiseks, ning käesoleva leiutise kohased meetodid vähendavad peremeesraku DNA-d farmatseutiliselt vastuvõetavate tasemeteni. Väikese- või suuremastaabilise fermenteerimise meetodeid saab kasutada ka valgu ekspressiooniga seoses, muuhulgas, kuid mitte ainult, fermenterid, loksutatavad kolvid, keevkiht-bioreaktorid, õõneskiud-bioreaktorid, rullpudel-kultiveerimissüsteemid ja segupaak-bioreaktorsüsteemid. Igaüht neist meetoditest saab teostada partiikaupa, söötme lisamisega või jätkuva protsessina. Leiutisekohased inimese FGF-21 polüpeptiidid on üldjuhul kogutavad standardsete tehnika tasemest teadaolevate meetodite abil. Näiteks saab söödet või rakulüsaati tsentrifuugida või filtreerida raku lagunemisjääkide eemaldamiseks. Supernatanti saab kontsentreerida või lahjendada soovitud mahuni või diafiltreerida sobivasse puhvrisse preparaadi ettevalmistamiseks edasiseks puhastamiseks. Käesoleva leiutise kohase FGF- 21 polüpeptiidi edasine puhastamine hõlmab FGF-21 polüpeptiidi deamideeritud ja kärbitud vormide eraldamist terviklikust vormist. Leiutisekohaste FGF-21 polüpeptiidide puhastamiseks saab rakendada ükskõik milliseid järgnevatest näitlikest menetlustest: afiinsuskromatograafia; anioon- või katioonvahetuskromatograafia (kasutades muuhulgas, kuid mitte ainult, DEAE SEPHAROSE); silikageelkromatograafia; kõrgefektiivne vedelikkromatograafia (HPLC); pöördfaasiline HPLC; geelfiltratsioon (kasutades muuhulgas, kuid mitte ainult, SEPHADEX G-75); hüdrofoobse interaktsiooni kromatograafia; suuruskromatograafia; metallkelaatkromatograafia; ultrafiltreerimine/diafiltreerimine; etanooliga sadestamine;
168 166 ammooniumsulfaadiga sadestamine; kromatofokusseerimine; asenduskromatograafia; elektroforeetilised protseduurid (muuhulgas, kuid mitte ainult, preparatiivne isoelektriline fokusseerimine), diferentsiaalne lahustamine (muuhulgas, kuid mitte ainult, ammooniumsulfaadiga sadestamine), SDS-PAGE või ekstraheerimine Leiutisekohaseid valke, muuhulgas, kuid mitte ainult, mittelooduslikke aminohappeid hõlmavad valgud, mittelooduslikke aminohappeid hõlmavad peptiidid, antikehad valkudele, mis hõlmavad mittelooduslikke aminohappeid, sidumispartnerid valkudele, mis hõlmavad mittelooduslikke aminohappeid jne. saab puhastada kas osalise või olulise homogeensuseni kooskõlas standardsete menetlustega, mida ala asjatundjad teavad ning kasutavad. Vastavalt saab leiutisekohaseid polüpeptiide koguda ja puhastada ükskõik millise abil mitmest ala tavataseme asjatundjatele teadaolevast meetodist, muuhulgas, kuid mitte ainult, sadestamine ammooniumsulfaadi või etanooliga, ekstraheerimine happe või alusega, kolonnkromatograafia, afiinsus-kolonnkromatograafia, anioon- või katioonvahetuskromatograafia, fosfotselluloos-kromatograafia, hüdrofoobse interaktsiooni kromatograafia, hüdroksülapatiit-kromatograafia, lektiinkromatograafia, geelelektroforees jne. Valkude ümbervoltimisetappe saab soovi korral kasutada õigesti volditud küpsete valkude valmistamiseks. Kui soovitakse kõrget puhtusastet, võib kõrgefektiivset vedelikkromatograafiat (HPLC), afiinsuskromatograafiat või muid sobivaid meetodeid rakendada lõplike puhastusetappide raames. Ühe teostuse korral kasutatakse mittelooduslike aminohapete (või mittelooduslikke aminohappeid sisaldavate valkude või peptiidide) vastu valmistatud antikehi puhastusreaktiividena, muuhulgas, kuid mitte ainult, üht või enamat mittelooduslikku aminohapet hõlmavate valkude või peptiidide afiinsuspõhiseks puhastamiseks. Puhastamise (osalise või homogeensuseni, soovi kohaselt) kasutatakse polüpeptiide valikuliselt mitmesugustel erinevatel otstarvetel, muuhulgas, kuid mitte ainult, katsekomponentide, terapeutikumide, profülaktikumide, diagnostikumide, uuringureaktiivide ja/või immunogeenidena antikehade tootmiseks. Käesoleva leiutise kohaste polüpeptiidide vastu loodud antikehad on valmistatavad polüpeptiidide või epitoopi kandvate fragmentide või rakkude manustamise teel loomale, eelistatavalt mitteinimloomale, rutiinseid menetlusi järgides. Ala tavataseme asjatundja on võimeline antikehade loomiseks mitmesuguseid teadaolevaid tehnikaid kasutades. Samuti võib humaniseeritud antikehade ekspresseerimiseks kasutada transgeenseid hiiri või muid organisme, sealhulgas muid imetajaid. Võimalik on ülalkirjeldatud antikehade
169 167 rakendamine polüpeptiidi ekspresseerivate kloonide isoleerimiseks või tuvastamiseks või polüpeptiidide puhastamiseks. Käesoleva leiutise kohaste polüpeptiidide vastaseid antikehi võib samuti kasutada haiguste ravimiseks Lisaks võib käesoleva leiutise kohaseid polüpeptiide ja polünukleotiide kasutada vaktsiinidena. Eeltoodust tulenevalt seondub käesolev leiutis täiendavalt meetodiga immunoloogilise vastuse esilekutsumiseks imetajas, mis hõlmab imetaja vaktsineerimist käesoleva leiutise polüpeptiidiga, mis võimaldab kutsuda esile antikeha tootmist ja/või T- raku immuunsusvastust, sealhulgas näiteks tsütokiini tootvad T-rakud või tsütotoksilised T-rakud, vastava looma kaitsmiseks haiguse eest, olgu see haigus tal juba välja kujunenud või mitte. Immunoloogilist vastust imetajal saab esile kutsuda ka meetodi teel, mis hõlmab käesoleva leiutise kohase polüpeptiidi kohaletoimetamist vektori kaudu, mis suunab polünukleotiidi ekspressiooni ja kodeerimist polüpeptiidi jaoks in vivo immunoloogilise vastuse esilekutsumiseks antikeha tootmiseks kõnealuse looma kaitsmiseks leiutises osutatud haiguste eest. Üheks vektori manustamisviisiks on selle kiirendamine soovitud rakkudesse kattekihina osakestel või muul viisil. Niisugune nukleiinhappe vektor võib hõlmata DNA-d, RNA-d, modifitseeritud nukleiinhapet või DNA/RNA hübriidi. Vaktsiinina kasutamiseks teostatakse polüpeptiid või nukleiinhappe vektor üldjuhul vaktsiini formulatsioonina (kompositsioonina). Formulatsioon võib täiendavalt hõlmata sobivat kandjat. Kuivõrd polüpeptiid on maos lagundatav, võib seda manustada parenteraalselt (näiteks subkutaanse, intramuskulaarse, intravenoosse või intradermaalse injektsiooni teel). Parenteraalseks manustamiseks sobivad formulatsioonid hõlmavad steriilseid süstelahuseid (veega ja ilma), mis võivad sisaldada antioksüdante, puhvreid, bakteriostaate ning soluute, mis muudavad formulatsiooni saaja verega kokkusobivaks, ning steriilseid suspensioone (veega ja ilma), mis võivad sisaldada suspendeerivaid agenseid või paksendeid. Vaktsiinformulatsioon võib samuti hõlmata adjuvandisüsteeme formulatsiooni immunogeensuse võimendamiseks, millised on ala tavataseme asjatundjatele teada. Annus oleneb vaktsiooni konkreetsest toimest ja on rutiinse eksperimenteerimise teel hõlpsasti kindlaksmääratav. Lisaks muudele siin äramärgitud viidetele on ala tavataseme asjatundjatele teada mitmesugused puhastus/valguvoltimismeetodid, muuhulgas, kuid mitte ainult, allikatest R. Scopes, Protein Purification, Springer-Verlag, N.Y. (1982); Deutscher, Methods in Enzymology Vol. 182: Guide to Protein Purification, Academic Press, Inc. N.Y. (1990);
170 Sandana, (1997) Bioseparation of Proteins, Academic Press, Inc.; Bollag et al. (1996) Protein Methods, 2nd Edition Wiley-Liss, NY; Walker, (1996) The Protein Protocols Handbook Humana Press, NJ, Harris and Angal, (1990) Protein Purification Applications: A Practical Approach IRL Press at Oxford, Oxford, England; Harris and Angal, Protein Purification Methods: A Practical Approach IRL Press at Oxford, Oxford, England; Scopes, (1993) Protein Purification: Principles and Practice 3rd Edition Springer Verlag, NY; Janson and Ryden, (1998) Protein Purification: Principles High Resolution Methods and Applications Second Edition Wiley-VCH, NY; ning Walker (1998), Protein Protocols on CD-ROM Humana Press, NJ; sh. eeltoodud allikates toodud viited. Üheks eeliseks mittelooduslikku aminohapet sisaldava huvipakkuva valgu või polüpeptiidi valmistamisel eukarüootses peremeesrakus või mitteeukarüootses peremeesrakus on asjaolu, et valgud või polüpeptiidid volditakse üldjuhul nende natiivsetes konformatsioonides. Küll aga peaks ala asjatundjatele leiutise teatud teostuste korral olema selge, et sünteesimise, ekspressiooni ja/või purifikatsiooni järel võib valkude või peptiidide konformatsioon erineda asjaomaste polüpeptiidide soovitud konformatsioonidest. Leiutise ühe aspekti kohaselt on võimalik ekspresseeritud valgu või polüpeptiidi valikuline denatureerimine ja renatureerimine. See saavutatakse valdkonnas teadaolevate meetodite abil, muuhulgas, kuid mitte ainult, tšaperoniini lisamine huvipakkuvale valgule või polüpeptiidile, valkude solubiliseerimine kaotroopses aines nagu guanidiin-hcl, valgu disulfiidisomeraasi ärakasutamine jne. Üldjuhul on aeg-ajalt soovitatav ekspresseeritud polüpeptiide denatureerida ja redutseerida, kutsudes seejärel esile polüpeptiidide ümbervoltumise eelistatud konformatsiooni. Näiteks võib huvipakkuvale translatsioonisaadusele lisada guanidiini, uureat, DTT-d, DTE-d ja/või tšaperoniini. Meetodid valkude redutseerimiseks, denatureerimiseks ja renatureerimiseks on ala tavataseme asjatundjatele teada (vt. eeltoodud viited ja Debinski, et al. (1993) J. Biol. Chem., 268: ; Kreitman and Pastan (1993) Bioconjug. Chem., 4: ; ning Buchner, et al., (1992) Anal. Biochem., 5: ). Näiteks kirjeldavad Debinski jt. inklusioonikeha-valkude denatureerimist ning redutseerimist guanidiin-dte-s. Valke saab ümber voltida redoks-puhvrisse, mis sisaldab muuhulgas, kuid mitte ainult, oksüdeeritud glutatiooni ja L-arginiini. Ümbervoltimisreaktiive saab vooluse teel või muul viisil viia kokkupuutesse ühe või enama polüpeptiidi või muu ekspressioonisaadusega, või vastupidi.
171 5 169 FGF-21 polüpeptiidi prokarüootse tootmise korral on võimalik niisugusel viisil saadud FGF-21 polüpeptiidi väärvoltimine, millega kaasneb puuduv või piiratud bioaktiivsus. Valgu bioaktiivsust saab taastada ümbervoltimise teel. Üldjuhul volditakse väärvolditud FGF-21 polüpeptiid ümber polüpeptiidahela solubiliseerimise (kui FGF-21 polüpeptiid on samuti lahustumatu), lahtivoltimise ja redutseerimise teel, kasutades näiteks üht või enamat kaotroopset ainet (nt. uurea ja/või guanidiin) ning disulfiidsidemete redutseerimist võimaldavat redutseerijat (nt. ditiotreitool, DTT või 2-merkaptoetanool, 2-ME). Kaotroopse aine mõõduka kontsentratsiooni juures lisatakse oksüdeerija (nt. hapnik, tsüstiin või tsüstamiin), mis võimaldab disulfiidsidemeid reformida. FGF-21 polüpeptiidi saab ümber voltida tehnika tasemest teadaolevate standardsete meetodite abil, mida on kirjeldatud näiteks patentides nr. US 4,511,502, US 4,511,503 ja US 4,512,922. Samuti on võimalik FGF-21 polüpeptiidi kaasvoltimine muude valkudega heterodimeeride või heteromultimeeride moodustamiseks. Ümbervoltimise järel saab FGF-21-t täiendavalt puhastada. FGF-21 puhastamine on saavutatav mitmesuguste ala tavataseme asjatundjatele teadaolevate tehnikate abil, sealhulgas hüdrofoobse interaktsiooni kromatograafia, suuruskromatograafia, ioonvahetuskromatograafia, pöörfaasiline kõrgefektiivne vedelikkromatograafia, afiinsuskromatograafia vms. või nende mis tahes kombinatsioon. Täiendav puhastamine võib samuti hõlmata puhastatud valgu kuivatamise või sadestamise etappi. Puhastamise järel on võimalik FGF-21 vahetamine erinevatesse puhvritesse ja/või kontsentreerimine ükskõik millise tehnika tasemest teadaoleva meetodi, muuhulgas, kuid mitte ainult, diafiltreerimise ning dialüüsi teel. Üksiku puhastatud valguna teostatud FGF- 21 suhtes võib rakendada agregatsiooni ja sadestamist. Puhastatud FGF-21 võib olla vähemalt 90% puhas (mõõdetuna pöördfaasilise kõrgefektiivse vedelikkromatograafia (RP-HPLC) või naatriumdodetsüülsulfaatpolüakrüülamiid geelelektroforeesi (SDS-PAGE) teel) või vähemalt 95% puhas või vähemalt 98% puhas või vähemalt 99% puhas või puhtam. Olenemata FGF-21 puhtuse täpsest numbrilisest väärtusest on FGF-21 piisavalt puhas kasutamiseks ravimtootena või täiendavaks töötlemiseks, näiteks konjugeerimiseks vees lahustuva polümeeriga nagu PEG.
172 170 Teatud FGF-21 molekulid on kasutatavad terapeutikumidena muude aktiivsete osiste või valkude puudumisel (peale abiainete, kandjate ja stabilisaatorite, seerumi albumiini jms.) või sobivad kompleksi moodustamiseks muu valgu või polümeeriga. 5 Üldised puhastusmeetodid Mis tahes paljudest isolatsioonietappidest on kohaldatav rakulüsaadi, ekstrakti, söötme, inklusioonikehade, peremeesrakkude periplasmaatilise ruumi, peremeesrakkude tsütoplasma või muu ainese suhtes, mis hõlmab FGF-21 polüpeptiidi, või ükskõik milliste FGF-21 polüpeptiidi segude suhtes, mis tulenevad ükskõik millistest isolatsioonietappidest, muuhulgas, kuid mitte ainult, afiinsuskromatograafia, ioonvahetuskromatograafia, hüdrofoobse interaktsiooni kromatograafia, geelfiltratsioonkromatograafia, kõrgefektiivne vedelikkromatograafia ( HPLC ), pöördfaasiline HPLC ( RP-HPLC ), paisutatud täidisega kromatograafia või nende mis tahes kombinatsioon ja/või kordus mis tahes sobivas järjekorras. Siinkirjeldatud tehnikate teostamiseks kasutatav seadmestik ja muud tarvilikud materjalid on kaubanduslikult saadaval. Pumpasid, fraktsioonikogureid, seireseadmeid, salvesteid ja terviklikke süsteeme pakuvad näiteks Applied Biosystems (Foster City, CA), BioRad Laboratories, Inc. (Hercules, CA) ning Amersham Biosciences, Inc. (Piscataway, NJ). Vastavad ettevõtted turustavad ka kromatograafilisi materjale, muuhulgas, kuid mitte ainult, vahetusmaatriksi materjale, söötmeid ja puhvreid. Tasakaalustamine ja muud kolonnkromatograafia protsesside siinkirjeldatud etapid, näiteks pesemine ning elueerimine, on kiiremini sooritatavad eriseadmestiku (nagu pump) abil. Kaubanduslikult saadaolevateks pumpadeks on muuhulgas, kuid mitte ainult, HILOAD Pump P-50, Peristaltic Pump P-1, Pump P-901 ja Pump P-903 (Amersham Biosciences, Piscataway, NJ). Fraktsioonikogurite näideteks on muuhulgas RediFrac Fraction Collector, fraktsioonikogurid FRAC-0 ja FRAC-0 ning SUPERFRAC Fraction Collector (Amersham Biosciences, Piscataway, NJ). Saadaval on ka segurid ph ja lineaarsete kontsentratsioonigradientide moodustamiseks. Kaubanduslikult saadaolevateks seguriteks on muuhulgas Gradient Mixer GM-1 ja In-Line Mixers (Amersham Biosciences, Piscataway, NJ).
173 Kromatograafilise protsessi seiret võib teostada ükskõik millise kaubanduslikult saadaoleva seireseadmega. Niisuguseid seireseadmeid võib kasutada teabe (nt. UV, ph ning juhtivuse kohta) kogumiseks. Detektorite näideteks on muuhulgas Monitor UV-1, UVICORDO S II, Monitor UV-M II, Monitor UV-900, Monitor UPC-900, Monitor ph/c- 900 ja Conductivity Monitor (Amersham Biosciences, Piscataway, NJ). Kaubanduslikult on saadaval ka terviklikud süsteemid, sealhulgas mitmesugused AKTA süsteemid ettevõttelt Amersham Biosciences (Piscataway, NJ). Käesoleva leiutise ühe teostuse kohaselt on näiteks võimalik FGF-21 polüpeptiidi redutseerimine ja denatureerimine esmalt saadud puhastatud FGF-21 polüpeptiidi denatureerimise teel uureas, millele järgneb lahjendus TRIS puhvris, mis sisaldab redutseerijat (nagu DTT) sobiva ph juures. Leiutise teise teostuse kohaselt denatureeritakse FGF-21 polüpeptiid uureas kontsentratsioonivahemikus u. 2 M kuni u. 9 M, millele järgneb lahjendus TRIS puhvris ph vahemiku u. 5.0 kuni u. 8.0 juures. Seejärel on võimalik teostuse kohase ümbervoltimissegu inkubeerimine. Leiutise ühe teostuse kohaselt inkubeeritakse ümbervoltimissegu toatemperatuuril neli kuni kakskümmend neli tundi. Seejärel võib redutseeritud ja denatureeritud FGF-21 polüpeptiidi segu täiendavalt isoleerida või puhastada. Nagu siin märgitud, võib esimese FGF-21 polüpeptiidisegu ph-d enne ükskõik milliste järgnevate isolatsioonietappide sooritamist kohandada. Lisaks võib esimest FGF-21 polüpeptiidisegu või selle ükskõik millist järgnevat segu tehnika tasemest teadaolevate tehnikate abil kontsentreerida. Samuti võib elutsioonipuhvri, mis hõlmab esimest FGF-21 polüpeptiidisegu või selle ükskõik millist järgnevat segu, asendada järgmise isolatsioonietapi läbiviimiseks sobiva puhvriga ala tavataseme asjatundjatele teadaolevate tehnikate abil. Ioonvahetuskromatograafia. Leiutise ühe teostuse korral võib esimese FGF-21 polüpeptiidi seguga valikuliselt viia läbi lisaetapina ioonvahetuskromatograafia. Vt üldteavet allikast ION EXCHANGE CHROMATOGRAPHY: PRINCIPLES AND METHODS (Cat. No , Amersham Biosciences (Piscataway, NJ)). Kaubanduslikult kättesaadavate ioonvahetuskolonnide seas on HITRAP, HIPREP ja HILOAD kolonnid (Amersham Biosciences, Piscataway, NJ). Selliste kolonnide puhul kasutatakse anioonvahetajaid nagu Q SEPHAROSE Fast Flow,
174 172 Q SEPHAROSE High Performance ja Q SEPHAROSE XL; tugevaid katioonvahetajaid nagu 5 30 SP SEPHAROSE High Performance, SP SEPHAROSE Fast Flow ja SP SEPHAROSE XL; nõrku anioonvahetajaid nagu DEAE SEPHAROSE Fast Flow; ja nõrku katioonivahetajaid nagu CM SEPHAROSE Fast Flow (Amersham Biosciences, Piscataway, NJ). Selleks et isoleerida üldjoontes puhastatud FGF-21 polüpeptiid, võib FGF-21 polüpeptiidiga läbi viia puhastusprotsessi mis tahes järgus viia läbi anioon- või katioonvahetuskolonnkromatograafia. Katioonvahetuskromatograafia etapi võib läbi viia mis tahes sobiva katioonvahetusmaatriksiga. Kasulike katioonvahetusmaatriksite seas on näiteks, kuid mitte ainult, kiuline, poorne, mittepoorne, mikrograanulitest koosnev, helmestest koosnev või ristseotud katioonvahetusmaatriksi materjal. Selliste katioonvahetusmaatriksi materjalide seas on näiteks, kuid mitte ainult, tselluloos, agaroos, dekstraan, polüakrülaat, polüvinüül, polüstüreen, ränidioksiid, polüeeter või mis tahes eespool nimetatud materjalide komposiidid. Katioonvahetusmaatriks võib olla mis tahes sobiv katioonvahetaja, sealhulgas tugev või nõrk katioonvahetaja. Tugevad katioonvahetajad jäävad ioniseerituks laias ph vahemikus ja seega suudavad siduda FGF- 21 laias ph vahemikus. Nõrgad katioonvahetajad aga võivad oma ioniseerituse ph funktsioonina kaotada. Nõrk katioonvahetaja näiteks võib kaotada laengu, kui ph langeb alla väärtuse 4 või 5. Sobivate tugevate katioonvahetajate seas on näiteks, kuid mitte ainult, laetud funktsionaalrühmad nagu sulfopropüül- (SP), metüülsulfonaat- (S) või sulfoetüül (SE) rühm. Katioonvahetusmaatriks võib olla tugev ioonvahetaja, mis seondub faktoriga FGF-21 eelistatavalt ph vahemikus umbes 2,5 kuni umbes 6,0. Tugev katioonvahetaja võib seonduda faktoriga FGF-21 ka ph vahemikus umbes 2,5 kuni umbes 5,5. Katioonvahetusmaatriks võib olla tugev katioonvahetusmaatriks, mis seondub faktoriga FGF-21 ph väärtusel umbes 3,0. Katioonvahetusmaatriks võib olla ka tugev katioonvahetaja, mis seondub faktoriga FGF-21 ph vahemikus umbes 6,0 kuni umbes 8,0. Katioonvahetusmaatriks võib olla tugev katioonvahetaja, mis seondub faktoriga FGF-21 ph vahemikus umbes 8,0 kuni umbes 12,5. Katioonvahetusmaatriks võib olla ka tugev katioonvahetaja, mis seondub faktoriga FGF-21 ph vahemikus umbes 8,0 kuni umbes 12,0. Enne FGF-21 laadimist võib katioonvahetusmaatriksit tasakaalustada näiteks mitme kolonni ruumalale vastava koguse lahjendatud nõrga happega, nt nelja kolonni ruumala
175 mm äädikhappega, ph 3. Pärast tasakaalustamist võib lisada FGF-21 ja kolonni pesta enne üldjoontes puhastatud FGF-21 elueerimist samuti nõrga happe lahusega nagu nõrk äädikhappe või fosforhappe lahus. Kolonni näiteks võib pesta umbes 2-4 kolonni ruumala mm äädikhappega, ph 3. Lisaks võib kolonni pesta näiteks 2-4 kolonni ruumala 0,05 M naatriumatsetaadiga, ph 5,5, või 0,05 M naatriumatsetaadiga, mis on segatud 0,1 M naatriumkloriidiga, ph 5,5. Selle asemel võib katioonvahetusmaatriksit tasakaalustada tehnika tasemes tuntud meetoditel mitme kolonni ruumala lahjendatud nõrga alusega. Selle asemel võib üldjoontes puhastatud FGF-21 elueerimiseks viia katioonvahetusmaatriksi kokkupuutesse puhvriga, millel on piisavalt madal ph või ioonjõud, et eemaldada FGF-21 maatriksist. Elueerimispuhvri ph võib olla vahemikus umbes 2,5 kuni 6,0. Elueerimispuhvri ph võib täpsemalt olla vahemikus umbes 2,5 kuni umbes 5,5, umbes 2,5 kuni umbes 5,0. Elueerimispuhvri ph võib olla umbes 3,0. Lisaks võib ka elueerimispuhvri kogus olla väga erinev ja tavaliselt on see umbes 2 kolonni ruumala kuni umbes kolonni ruumala. Pärast FGF-21 polüpeptiidi adsorbeerimist katioonvahetusmaatriksile võib üldjoontes puhastatud FGF-21 polüpeptiidi elueerumiseks viia maatriksi kokkupuutesse puhvriga, millel on piisavalt madal ph või ioontugevus, et eemaldada FGF-21 maatriksist. Üldjoontes puhastatud FGF-21 polüpeptiidi elueerimisel ph kõrgel väärtusel kasutatavate puhvrite seas on näiteks, kuid mitte ainult, tsitraat-, fosfaat-, formiaat-, atsetaat-, HEPES ja MES puhvrid kontsentratsiooniga vähemalt umbes 5 mm kuni vähemalt umbes 0 mm. Pöördfaaskromatograafia Valke võib puhastada RP-HPLC kromatograafiaga asjatundjatele tuntud sobivate protokollide järgi. Vt nt Pearson et al., ANAL BIOCHEM. (1982) 124: (1982); Rivier et al., J. CHROM. (1983) 268: ; Kunitani et al., J. CHROM. (1986) 359: FGF-21 polüpeptiidid korral saab RP-HPLC meetodil isoleerida üldjoontes puhastatud FGF-21 polüpeptiidi. Selle tegemisel võib kasutada erineva pikkusega, ränidioksiidiga derivatiseeritud vaike, millel on funktsionaalsed alküülrühmad, kusjuures võib kasutada vaike, mille pikkus on näiteks, kuid mitte ainult, vähemalt umbes C 3 kuni vähemalt umbes C 30, vähemalt umbes C 3 kuni vähemalt umbes C või vähemalt C 3 kuni vähemalt umbes C 18. Selle asemel võib kasutada polümeerset vaiku. Näiteks võib kasutada TosoHaas Amberchrome CG00sd vaiku, mis on stüreenpolümeeri vaik. Samuti võib kasutada väga erineva pikkusega alküülahelaga tsüano- või polümeerseid vaike. RP-HPLC kolonni võib pesta ka lahustiga nagu etanool.
176 RP-HPLC kolonni veel üheks näiteks on RP-HPLC kolonn. FGF-21 polüpeptiidi elueerimiseks RP-HPLC kolonnist võib kasutada sobivat elueerimispuhvrit, mis sisaldab ioonpaaride moodustamise ainet (ion pairing agent) ja orgaanilist modifitseerijat nagu metanool. isopropanool, tetrahüdrofuraan, atsetonitriil või etanool. Kõige sagedamini kasutatavate ioonpaare moodustavate ainete seas on näiteks, kuid mitte ainult, äädikhape, sipelghape, perkloorhape, fosforhape, trifluoroäädikhape, heptafluorovõihape, trietüülamiin, tetrametüülammoonium-, tetrabutüülammoonium- ja trietüülammooniumatsetaat. Elueerimisel võib kasutada ühte või mitut gradienti või isokraatilisi tingimusi, kusjuures eelistatud on gradiendid, et lühendada lahutusaega ja vähendada piigi laiust. Teise meetodi korral kasutatakse kahte gradientlahustit erineva kontsentratsioonivahemikuga. Käesoleva leiutise puhul sobivate elueerimispuhvrite seas on näiteks, kuid mitte ainult, ammooniumatsetaadi ja atsetonitriili lahused. Hüdrofoobse interaktsiooni kromatograafial põhinevad puhastusmeetodid. FGF-21 polüpeptiidi võib puhastada hüdrofoobse interaktsiooni kromatograafia (HIC) meetodil. Vt üldteavet allikast HYDROPHOBIC INTERACTION CHROMATOGRAPHY HANDBOOK: PRINCIPLES AND METHODS (Cat. No , Amersham Biosciences (Piscataway, NJ). Sobivate HIC maatriksite seas on näiteks, kuid mitte ainult, alküül- ja arüülasendatud maatriksid nagu butüül-, heksüül-, oktüül- või fenüülasendatud maatriksid, sealhulgas agaroos-, ristseotud agaroosi, sefaroos-, tselluloos-, ränidioksiid-, dekstraan-, polüstüreen-, polü(metakrülaat)maatriksid ja mixed mode vaigud, sealhulgas, kuid mitte ainult, polüetüleenamiinvaik või butüül- või fenüülasendatud polü(metakrülaat) maatriks. Kaubanduslikult kättesaadavate hüdrofoobse interaktsiooni kromatograafia kolonnide seas on näiteks, kuid mitte ainult, HITRAP, HIPREP ja HILOAD kolonnid (Amersham Biosciences, Piscataway, NJ). Enne laadimist võib HIC kolonni tasakaalustada asjatundjatele tuntud standardsete puhvritega nagu äädikhappe/naatriumkloriidi lahus või HEPES, mis sisaldab ammooniumsulfaati. Ammooniumsulfaati võib kasutada puhvrina HIC kolonni laadimisel. Pärast FGF-21 polüpeptiidi laadimist võib kolonni pesta standardsetes tingimustes standardsete puhvritega, et eemaldada soovimatud materjalid, kuid jätta FGF-21 polüpeptiid HIV kolonnile. FGF-21 polüpeptiidi võib elueerida muu hulgas umbes 3 kuni
177 175 umbes kolonni ruumala standardse puhvriga nagu HEPES, mis sisaldab EDTA-d ja madalamas kontsentratsioonis ammooniumsulfaati kui tasakaalustamispuhver, või äädikhappe/naatriumkloriidi puhvriga. 5 FGF-21 molekulide elueerimiseks võib kasutada ka lineaarset alanevat soola gradienti, näiteks kaaliumfosfaadi gradienti. Seejärel võib eluenti kontsentreerida näiteks filtreerimise nagu diafiltreerimine või ultrafiltreerimine teel. Diafiltreerimise teel võib eemdaldada FGF-21 polüpeptiidi elueerimisel kasutatud soola. Teised puhastusmeetodid 30 Selleks et eemaldada mis tahes üleliigsed soolad ja asendada puhvri järgmiseks isoleerimisetapiks sobiva puhvriga või isegi valmistada lõplik ravim, võib esimese FGF- 21 polüpeptiidi seguga või mis tahes järgneva FGF-21 polüpeptiidi seguga viia läbi veel ühe isoleerimisetapi, kasutades selleks geelfiltratsiooni (GEL FILTRATION: PRINCIPLES AND METHODS (Cat. No , Amersham Biosciences, Piscataway, NJ), hüdroksüapatiit-kromatograafiat (selleks sobivate maatriksite seas on näiteks, kuid mitte ainult, HA-Ultrogel, High Resolution (Calbiochem), CHT Ceramic Hydroxyapatite (BioRad), Bio - Gel HTP Hydroxyapatite (BioRad)), HPLC, expanded bed adsorption, ultrafiltreerimist, diafiltreerimist, lüofiliseerimist ja teisi sarnaseid meetodeid. FGF-21 polüpeptiidi, sealhulgas üldjoontes puhastatud FGF-21 polüpeptiidi saagist saab kõigis käesolevas dokumendis kirjeldatavates etappides jälgida tehnika tasemes tuntud metoditel. Sellistel meetoditel saab ka hinnata üldjoontes puhastatud FGF- 21 polüpeptiidi saagist pärast viimast isoleerimisetappi. FGF-21 polüpeptiidi saagist võib näiteks hinnata mis tahes pöördfaasi kõrgsurve vedelikkromatograafia kolonniga, millel on erineva pikkusega alküülahelad, nagu tsüano-rp-hplc, C 18 RP-HPLC, ja ka katioonvahetus HPLC ja geefiltratsiooni HPLC-ga. Käesoleva leiutise konkreetsetes teostustes võib FGF-21 polüpeptiidi saagis pärast igat puhastusetappi moodustada vähemalt umbes 30%, vähemalt umbes 35%, vähemalt umbes 40%, vähemalt umbes 45%, vähemalt umbes 50%, vähemalt umbes 55%, vähemalt umbes 60%, vähemalt umbes 65%, vähemalt umbes 70%, vähemalt umbes 75%, vähemalt umbes 80%, vähemalt umbes 85%, vähemalt umbes 90%, vähemalt umbes 91%, vähemalt umbes 92%, vähemalt umbes 93%, vähemalt umbes 94%, vähemalt umbes 95%, vähemalt umbes 96%, vähemalt umbes 97%,
178 vähemalt umbes 98%, vähemalt umbes 99%, vähemalt umbes 99.9% või vähemalt umbes 99.99% FGF-21 kogusest lähtematerjalis. Puhtuse võib määrata standardsetel meetoditel nagu SDS-PAGE või FGF-21 polüpeptiidi mõõtmise teel Western blot ja ELISAanalüüsiga. Näiteks võib luua negatiivse kontrolli pärmi fermentatsioonil ja katioonvahetusel isoleeritud valkude vastased polüklonaalsed antikehad. RP-HPLC materjal Vydac C4 (Vydac) koosneb ränidioksiidi osakestest, mille pinnal on C4- alküülahelad. FGF-21 polüpeptiidi eraldamine valgulistest lisanditest põhineb erinevustel hüdrofoobsete interaktsioonide tugevuses. Elueerimine viiakse läbi atsetonitriili gradiendiga lahejndatud trifluoroäädikhappes. Preparatiivne HPLC viiakse läbi roostevabast terasest kolonnis (mis on täidetud 2,8 kuni 3,2 liitri Vydac C4 silikageeliga). Hydroxyapatite Ultrogel eluaadile lisatakse hapestamiseks trifluoroäädikhapet ja see laaditakse Vydac C4 kolonni. Loputamiseks ja elueerimiseks kasutatakse atsetonitriili gradienti lahjendatud trifluroäädikhappes. Fraktsioonid kogutakse kokku ja neutraliseeritakse kohe fosfaatpuhvriga. IPC piiridesse jäävad FGF-21 polüpeptiidi fraktsioonid koondatakse. DEAE Sepharose (Pharmacia) materjal koosneb dietüülaminoetüül (DEAE)-rühmadest, mis on seotud kovalentselt Sepharose helmeste pinnaga. FGF-21 polüpeptiidi ja DEAE rühmade seondumine põhineb ioonide interaktsioonidel. Atsetonitriil ja trifluoroäädikhape läbivad kolonni. Pärast nende ainete eemaldamist pestakse kolonni sinna jäänud lisandite eemaldamiseks atsetaatpuhvriga, millel on madal ph. Seejärel loputatakse kolonni neutraalse fosfaatpuhvriga ja FGF-21 polüpeptiid elueeritakse puhvriga, millel on suurem ioontugevus. Kolonn täidetakse tootega DEAE Sepharose Fast Flow. Kolonni ruumala reguleeritakse selliselt, et FGF-21 polüpeptiidi sisaldus oleks 3- mg FGF-21 polypeptide/ml geeli. Kolonni loputatakse vee ja tasakaalustamispuhvriga (naatrium/kaaliumfosfaat). HPLC eluaadi koondatud fraktsioonid laaditakse kolonni ja kolonni pestakse tasakaalustamispuhvriga. Seejärel pesti kolonni pesupuhvriga (naatriumatsetaatpuhver) ja siis tasakaalustamispuhvriga. Seejärel elueeriti FGF-21 polüpeptiid elueerimispuhvriga (naatriumkloriid, naatrium/kaaliumfosfaat) kolonnist ja koguti master elution profile järgi ühe fraktsioonina kokku. DEAE Sepharose kolonni eluaati reguleeriti täpselt määratud juhtivuse järgi. Saadud raviaine pakendati steriilse filtreerimise teel Teflon
179 5 177 pudelitesse ja neid hoiti temperatuuril -70 C. Teised sobivad meetodid hõlmavad näiteks, kuid mitte ainult etappe, milles eemaldatakse endotoksiinid. Endotoksiinid on lipopolüsahhariidid (LPS), mis paiknevad gramnegatiivsete peremeesrakkude, näiteks Escherichia coli rakkude välismembraanil. Endotoksiinide taseme alandamise meetodid on asjatundjatele teada ning nende seas on näiteks, kuid mitte ainult, puhastusmeetodid, mille korral kasutatakse ränidioksiidist tugistruktuure või klaaspulbrit, hüdroksüapatiit-, pöördfaasi, afiinsus, suuruseraldus, anioonvahetus-, hüdrofoobse interaktsiooni kromatograafia, nimetatud meetodite kombinatsioonid ja muud sarnased meetodid. Huvipakkuva polüpeptiidi puhastamiseks komigreeruvatest valkudest võib olla vaja teha modifikatsioone või kasutada lisameetodeid. Endotoksiinide taseme mõõtmismeetodid on asjatundjatele teada ja nende seas on näiteks, kuid mitte ainult, Limulus Amebocyte Lysate (LAL) analüüsid. Endosafe -PTS analüüs on kolorimeetriline ühetoruline süsteem, milles kasutatakse kassette, mis on eelnevalt täidetud LAL reagendi, kromogeense substraadi ja kontrollina kasutatava standardse endotoksiiniga, ja mille juurde kuulub käes hoitav spektrofotomeeter. Teiste asjakohaste meetodite on näiteks, kuid mitte ainult, turbidimeetriline Kinetic LAL meetod, mille korral kasutatakse 96 süvendiga plaate. Ühte või mitut mittelooduslikult kodeeritud aminohapet sisaldava FGF-21 valgu saagist ja puhtust saab hinnata paljude erinevate meetodite ja protseduuridega, sealhulgas, kuid mitte ainult, Bradfordi analüüsi, SDS-PAGE, hõbeda värvainega SDS-PAGE, Coomassie sinise värvainega SDS-PAGE, massispektromeetria (näiteks, kuid mitte ainult, MALDI-TOF) ja teistel asjatundjatele tuntud valkude iseloomustamise meetoditel. Teised asjakohased meetodid on näiteks, kuid mitte ainult: SDS-PAGE koos valkude värvimise meetoditega, immunoblotanalüüsid, MALDI-MS (matrix assisted laser desorption/ionization-mass spectrometry), vedelikkromatograafia/massispektromeetria, isoelektriline fokusseerimine, analüütiline anioonvahetus, kromatofokusseerimine ja tsirkulaardikroism. VIII. Ekspresseerimine alternatiivsetes süsteemides 30 Mittelooduslikke aminohappeid on sisestatud mitterekombinantsetesse peremeesrakkudesse, muteeritud peremeesrakkudesse või rakuvabadesse süsteemidesse mitme strateegia abil. Sellised süsteemid sobivad ka käesoleva leiutise kohaste FGF-21 polüpeptiidide valmistamiseks. Reaktsioonivõimeliste kõrvalahelatega aminohapete nagu
180 Lys, Cys ja Tyr derivatiseerimisel muundus lüsiin N 2 -atsetüül-lüsiiniks. Keemiline süntees on samuti lineaarne meetod, mis sobib mittelooduslike aminohapete sisestamiseks. Tänu hiljutistele edusammudele peptiidifragmentide ensümaatilise ligeerimise ja natiivse keemilise ligeerimise vallas on võimalik valmistada suuremaid valke. Vt nt P. E. Dawson and S. B. H. Kent, Annu. Rev. Biochem, 69:923 (00). Keemilist peptiidide ligeerimist ja natiivset keemilist ligeerimist kirjeldatakse USA patendis nr , USA patendipublikatsioonis nr 04/ , USA patendipublikatsioonis 03/08046, WO 02/ ja WO 03/ Üldisel in vitro biosünteesi meetodil, mille korral supressortrna, mis on keemiliselt atsüülitud soovitud mitteloodusliku aminohappega lisatakse in vitro ekstraktile, millel on võime toetada valgu biosüsnteesi, on kohtspetsiifiliselt sisestatud peaaegu mis tahes suurusega erinevatesse valkudesse üle 0 mitteloodusliku aminohappe. Vt nt V. W. Cornish, D. Mendel and P. G. Schultz, Angew. Chem. Int. Ed. Engl., 1995, 34:621 (1995); C.J. Noren, S.J. Anthony-Cahill, M.C. Griffith, P.G. Schultz, A general method for site-specific incorporation of unnatural amino acids into proteins, Science 244: (1989); and, J.D. Bain, C.G. Glabe, T.A. Dix, A.R. Chamberlin, E.S. Diala, Biosynthetic site-specific incorporation of a non-natural amino acid into a polypeptide, J. Am. Chem. Soc. 111: (1989). Valkude stabiilsuse, kokkuvoltimise, ensüümi toimemehhanismi ja signaalide ülekande uurimiseks on valkudesse sisestatud paljusid erinevaid funktsionaalrühmi. Metsikut tüüpi süntetaaside mitmekesisuse ärakasutamiseks on loodud in vivo meetod nimega selective pressure incorporation. Vt nt N. Budisa, C. Minks, S. Alefelder, W. Wenger, F. M. Dong, L. Moroder and R. Huber, FASEB J., 13:41 (1999). Auksotroofset tüve, milles rakku konkreetse aminohappega varustav oluline ainevahetusrada on välja lülitatud, kasvatatakse minimaalses söötmes, mis sisaldab piiratud koguses looduslikku aminohapet ja sihtgeeni transkriptsioon surutakse alla. Statsionaarse kasvufaasi alguses looduslik aminohape eemaldatakse ja asendatakse mittelooduslikust aminohappest analoogiga. Rekombinantse valgu ekspressiooni indutseerimisel hakkab kuhjuma mittelooduslikku analoogi sisaldav valk. Selle strateegia abil on näiteks sisestatud valkudesse o, m and p-fluorofenüülalaniine ja UV-spektril tekib kaks iseloomulikku ja kergesti identifitseeritavat õlga, vt nt C. Minks, R Huber, L. Moroder and N. Budisa, Anal. Biochem., 284:29 (00); bakteriofaagi T4 lüsosoomis on metioniin asendatud trifluorometioniiniga, et uurida 19 F NMR meetodil selle interkatsioon kitoolisahhariidi
181 179 ligandidega, vt nt H. Duewel, E. Daub, V. Robinson and J. F. Honek, Biochemistry, 36:3404 (1997); ja leutsiin on asendatud trifluoroleutsiiniga, mille tulemusel suurenes leutsiini tõmblukk valgu termiline ja keemiline stabiilsus. Vt nt 5 30 Y. Tang, G. Ghirlanda, W. A. Petka, T. Nakajima, W. F. DeGrado and D. A. Tirrell, Angew. Chem. Int. Ed. Engl., 40:1494 (01). Lisaks sisestatakse mitmesugustesse rekombinantsetesse valkudesse selenometioniin ja tellurometioniin, et hõlbustada faaside lahutamist röntgenkristallograafias. Vt nt W. A. Hendrickson, J. R. Horton and D. M. Lemaster, EMBO J., 9:1665 (1990); J. O. Boles, K. Lewinski, M. Kunkle, J. D. Odom, B. Dunlap, L. Lebioda and M. Hatada, Nat. Struct. Biol., 1:283 (1994); N. Budisa, B. Steipe, P. Demange, C. Eckerskom, J. Kellermann and R. Huber, Eur. J. Biochem., 230:788 (1995); and, N. Budisa, W. Karnbrock, S. Steinbacher, A. Humm, L. Prade, T. Neuefeind, L. Moroder and R. Huber, J. Mol. Biol., 270:616 (1997). Lisaks on efektiivselt sisestatud ka funktsionaalsete alkeenvõi alküünrühmadega metioniini analooge, mis võimaldab valke keemiliselt veelgi rohkem modifitseerida. Vt nt J. C. van Hest and D. A. Tirrell, FEBS Lett., 428:68 (1998); J. C.. van Hest, K. L. Kiick and D. A. Tirrell, J. Am. Chem. Soc., 122:1282 (00); ja K. L. Kiick and D. A. Tirrell, Tetrahedron, 56:9487 (00); USA patent nr ; USA patendipublikatsioon 02/ Selle meetodi efektiivsus sõltub mittelooduslike aminohapete analoogide äratundmisest aminoatsüül-trna süntetaaside poolt, mille korral on valgu translatsiooni tagamiseks vaja suuremat selektiivsust. Üks võimalus selle meetodi laiendamiseks on vähendada aminoatsüül-trna süntetaaside substraadispetsiifilisust, mida on õnnestunud saavutada vähestel juhtudel. Näiteks Ala 294 asendamisel Escherichia coli fenüülalanüül-trna süntetaasis (PheRS) Gly-ga suurenenb substraadi sidumistasku ja toimub trnaphe atsüülimine p-cl-fenüülalaniiniga (p-cl-phe). Vt M. Ibba, P. Kast and H. Hennecke, Biochemistry, 33:77 (1994). Sellist mutantset PheRS-i sisaldav Escherichia coli tüvi võimaldab sisestada fenüülalaniini asemele p-clfenüülalaniini või p-br-fenüülalaniini. Vt nt M. Ibba and H. Hennecke, FEBS Lett., 364:272 (1995); ja N. Sharma, R. Furter, P. Kast and D. A. Tirrell, FEBS Lett., 467:37 (00). Lisaks tehti kindlaks, et Escherichia coli türosüül-trna süntetaasi aminohappe sidumiskoha lähedal paiknev punktmutatsioon võimaldab asatürosiini sisestada
182 efektiivsemalt kui türosiini. Vt F. Hamano-Takaku, T. Iwama, S. Saito-Yano, K. Takaku, Y. Monden, M. Kitabatake, D. Soll and S. Nishimura, J. Biol. Chem., 275:40324 (00). Teine strateegia, mille abil saab sisestada in vivo valkudesse mittelooduslikke aminohappeid, seisneb korrigeerimismehhanisme omavate süntetaaside modifitseerimises. Sellistel süntetaasidel puudub eristusvõime ja seetõttu aktiveerivad nad aminohappeid, mis sarnanevad struktuuri poolest kognaatsetele looduslikele aminohapetele. See viga parandatakse eraldi saidil, kus deatsüülitalse trna-st ekslikult vabastatud aminohape, et tagada valgu korrektne translatsioon. Kui süntetaasi korrigeerimismehhanism ei toimi, võivad valesti aktiveeritud struktuursed analoogid jääda korrigeerimata ja sattuda valku. Sellist strateegiat on hiljuti kasutatud valüül-trna süntetaasiga (VaIRS). Vt V. Doring, H. D. Mootz, L. A. Nangle, T. L. Hendrickson, V. de Crecy-Lagard, P. Schimmel and P. Marliere, Science, 292:501 (01). ValRS võib ekslikult aminoatsüülida trnaval-i Cys, Thr või aminobutüraadiga (Abu); need mittekognaatsed aminohapped hüdrolüüsib seejärel korrigeeriv domeen. Pärast Escherichia coli kromosoomi juhuslikku mutageneesi valiti mutantne Escherichia coli tüvi, millel oli ValRS korrigeerimisaidil mutatsioon. Selline puuduliku korresktsiooniga ValRS laeb trnaval-i vääralt Cys iga. Kuna Abu sarnaneb steeriliselt Cys-ile (Cys -SH rühm asendatakse Abu's -CH3-ga), sisestab mutantne ValRS valkudesse Abu, kui sellist mutantset Escherichia coli tüve kasvatatakse Abu juuresolekul. Massspektromeetriline analüüs näitab, et umbes 24% valiinidest asendatakse looduslikus valgus igas valiini positsioonis Abu'ga. Paljusid valke, mis sisaldavad uudseid aminohappeid, saab sünteesida ka tahke faasi sünteesi ja poolsünteetilistel meetoditel. Vt nt järgmisi publikatsioone ja neis toodud viiteid: Crick, F.H.C., Barrett, L. Brenner, S. Watts-Tobin, R. General nature of the genetic code for proteins. Nature, 192: (1961); Hofmann, K., Bohn, H. Studies on polypeptides. XXXVI. The effect of pyrazoleimidazole replacements on the S-protein activating potency of an S-peptide fragment, J. Am chem, 88(24): (1966); Kaiser, E.T. Synthetic approaches to biologically active peptides and proteins including enyzmes, Acc Chem Res, 22:47-54 (1989); Nakatsuka, T., Sasaki, T., Kaiser, E.T. Peptide segment coupling catalyzed by the semisynthetic enzyme thiosubtilisin, J Am Chem Soc, 9: (1987); Schnolzer, M., Kent, S B H. Constructing proteins by dovetailing unprotected synthetic peptides: backbone-engineered HIV protease, Science, 6(5054):221-2 (1992); Chaiken, I.M. Semisynthetic peptides and proteins, CRC Crit Rev Biochem, 11(3):5-301 (1981); Offord, R.E. Protein engineering by chemical means? Protein Eng., 1(3):1-7 (1987);
183 and, Jackson, D.Y., Burnier, J., Quan, C., Stanley, M., Tom, J., Wells, J.A. A Designed Peptide Ligase for Total Synthesis of Ribonuclease A with Unnatural Catalytic Residues, Science, 266(5183):243 (1994). Keemilise modifitseerimise teel on sisestatud valkudesse in vivo erinevaid mittelooduslikke kõrvalahelaid, sealhulgas kofaktoreid, spin-märgiseid ja oligonukleotiide. Vt nt Corey, D.R., Schultz, P.G. Generation of a hybrid sequencespecific single-stranded deoxyribonuclease, Science, 238(4832): (1987); Kaiser, E.T., Lawrence D.S., Rokita, S.E. The chemical modification of enzymatic specificity, Annu Rev Biochem, 54: (1985); Kaiser, E.T., Lawrence, D.S. Chemical mutation of enyzme active sites, Science, 226(4674): (1984); Neet, K.E., Nanci A, Koshland, D.E. Properties of thiol-subtilisin, J Biol. Chem, 243(24): (1968); Polgar, L. et M.L. Bender. A new enzyme containing a synthetically formed active site. Thiol-subtilisin. J. Am Chem Soc, 88:33-34 (1966); and, Pollack, S.J., Nakayama, G. Schultz, P.G. Introduction of nucleophiles and spectroscopic probes into antibody combining sites, Science, 242(4881):38-40 (1988). In vitro sünteesitud valkudesse on sisestatud mitu biofüüsikalist sondi ka biosünteetilistel meetditel, mille korral kasutatakse keemiliselt modifitseeritud aminoatsüül-trna-sid. Vt järgmisi publikatsioone ja neis toodud viiteid: Brunner, J. New Photolabeling and crosslinking methods, Annu. Rev Biochem, 62: (1993); and, Krieg, U.C., Walter, P., Hohnson, A.E. Photocrosslinking of the signal sequence of nascent preprolactin of the 54-kilodalton polypeptide of the signal recognition particle, Proc. Natl. Acad. Sci, 83(22): (1986). Varem on näidatud, et mittelooduslikke aminohappeid saab valkudesse in vitro kohtspetsiifiliselt sisestada keemiliselt aminoatsüülitud supressor-trna-de lisamise teel valgu sünteesi reaktsioonidele, milles kasutatakse soovitud amber nonsenss-mutatsiooni sisaldavat geeni. Selliste strateegiate abil saab asendada paljud tavapäraste aminohapete hulka kuuluvad aminohapped lähedaste struktuursete homoloogidega ehk näiteks fenüülalaniini fluorofenüülalaniiniga, kasutades selleks konkreetse aminohappe suhtes auksotroofseid tüvesid. Vt nt Noren, C.J., Anthony-Cahill, Griffith, M.C., Schultz, P.G. A general method for site-specific incorporation of unnatural (amino acids into proteins, Science, 244: (1989); M.W. Nowak, et al., Science 268: (1995); Bain, J.D., Glabe, C.G., Dix,
184 T.A., Chamberlin, A.R., Diala, E.S. Biosynthetic site-specific Incorporation of a nonnatural amino acid into a polypeptide, J. Am Chem Soc, 111: (1989); N. Budisa et al., FASEB J. 13:41-51 (1999); Ellman, J.A., Mendel, D., Anthony-Cahill, S., Noren, C.J., Schultz, P.G. Biosynthetic method for introducing unnatural amino acids sitespecifically into proteins, Methods in Enz., vol. 2, (1992); ja Mendel, D., Cornish, V.W. & Schultz, P.G. Site-Directed Mutagenesis with an Expanded Genetic Code, Annu Rev Biophys. Biomol Struct. 24, (1995). Näiteks valmistati supressor-trna, mis tundis ära stoppkoodoni UAG, ja see aminoatsüüliti keemiliselt mitteloodusliku aminohappega. Valgu huvipakkuvasse saiti sisestati tavapärase kohtsuunatud mutageneesi teel stoppkoodon TAG. Vt nt Sayers, J.R., Schmidt, W. Eckstein, F. 5'-3' Exonucleases in phosphorothioate-based oligonucleotidedirected mutagenesis, Nucleic Acids Res, 16(3): (1988). Atsüülitud supressortrna ja mutantse geeni kombineerimisel in vitro transkriptsiooni/translatsiooni süsteemi sisestati mittelooduslik aminohape vastusena UAG koodonile, mille tulemusena saadi valk, mis sisaldas kindlas positsioonis sisestatud aminohapet. [ 3 H]-Phe-ga tehtud katsed ja α- hüdroksühapetega tehtud katsed näitasid, et UAG koodoni poolt määratud kohta lisati vaid soovitud aminohape ja seda aminohapet ei lisatud valgus teistesse positsioonidesse. Vt nt Noren, et al, supra; Kobayashi et al., (03) Nature Structural Biology (6):4-432; and, Ellman, J.A., Mendel, D., Schultz, P.G. Site-specific incorporation of novel backbone structures into proteins, Science, 5(5041):197-0 (1992). trna võib aminoatsüülida soovitud aminohappega mis tahes meetodil või tehnika abil, sealhulgas, kuid mitte ainult, keemilise või ensümaatilise aminoatsüülimise teel. Aminoatsüülimiseks võib kasutada aminoatsüül trna süntetaase või teisi ensüümimolekule, sealhulgas, kuid mitte ainult, ribosüüme. Mõistet ribosüüm kasutatakse mõiste katalüütiline RNA sünonüümina. Cech ja kolleegid (Cech, 1987, Science, 236:32-39; McCorkle et al., 1987, Concepts Biochem. 64: ) näitasid katalüsaatorite (ribosüümide) ülesannet täitvate looduslike RNA-de olemasolu. Kuigi on selgunud, et need looduslikud RNA katalüsaatorid toimivad ribonukleiinhappe substraatidele vaid lõhustamisel ja splaissimisel, on hiljuti välja töötatud tehisribosüümide varal laiendatud katalüüsi mitmesugustele keemilistele reaktsioonidele.
185 Uuringutes on identifitseeritud RNA molekulid, millel on võime katalüüsida nende oma otstes paiknevaid aminoatsüül-rna sidemeid (Illangakekare et al., 1995 Science 267: ), ja RNA molekul, mis suudab viia aminohappe ühest RNA molekulist teise (Lohse et al., 1996, Nature 381: ). USA patendipublikatsioonis 03/ kirjeldatakse ribosüümide konstrueerimismeetodeid ja nende kasutamist trna-de aminoatsüülimisel looduslikult ja mittelooduslikult kodeeritud aminohapete aminoatsüülimisel. Ensümaatiliste molekulide substraadil immobiliseeritud vormidega, millel on võime aminoatsüülida trna-sid, sealhulgas, kuid mitte ainult, ribosüüme, võib olla võimalik aminoatsüülitud saadusi efektiivselt afiinsuspuhastada. Sobivate substraatide näidete seas on agaroos, sefaroos ja magnethelmed. Ribosüümi substraadil immobiliseeritud vormi valmistamist ja kasutamist aminoatsüülimiseks kirjeldatakse allikas Chemistry and Biology 03, :77-84 ja USA patenditaotluse publikatsioonis 03/ Keemiliste aminoatsüülimise meetodite seas on näiteks, kuid mitte ainult, meetodid, mida kirjeldavad Hecht ja tema kolleegid (Hecht, S. M. Acc. Chem. Res. 1992,, 545; Heckler, T. G.; Roesser, J. R.; Xu, C.; Chang, P.; Hecht, S. M. Biochemistry 1988, 27, 74; Hecht, S. M.; Alford, B. L.; Kuroda, Y.; Kitano, S. J. Biol. Chem. 1978, 3, 4517) ning Schultz, Chamberlin, Dougherty ja teised (Cornish, V. W.; Mendel, D.; Schultz, P. G. Angew. Chem. Int. Ed. Engl. 1995, 34, 621; Robertson, S. A.; Ellman, J. A.; Schultz, P. G. J. Am. Chem. Soc. 1991, 113, 2722; Noren, C. J.; Anthony-Cahill, S. J.; Griffith, M. C.; Schultz, P. G. Science 1989, 244, 182; Bain, J. D.; Glabe, C. G.; Dix, T. A.; Chamberlin, A. R. J. Am. Chem. Soc. 1989, 111, 8013; Bain, J. D. et al. Nature 1992, 356, 537; Gallivan, J. P.; Lester, H. A.; Dougherty, D. A. Chem. Biol. 1997, 4, 740; Turcatti, et al. J. Biol. Chem. 1996, 271, 19991; Nowak, M. W. et al. Science, 1995, 268, 439; Saks, M. E. et al. J. Biol. Chem. 1996, 271, 23169; Hohsaka, T. et al. J. Am. Chem. Soc. 1999, 121, 34), et oleks võimalik vältida aminoatsüülimisel süntetaaside kasutamist. trna molekulide aminoatsüülimisel võib kasutada selliseid ja teisi keemilisi aminoatsüülimise meetodeid. Katalüütilise RNA valmistusmeetodid võivad hõlmata randomiseeritud/juhuslike ribosüümi järjestuste eraldi kogumite loomist, selliste kogumite suunatud kasvatamist, kogumite skriinimist soovitud aminoatsüülimise aktiivsuse suhtes ja soovitud
186 aminoatsüülimise aktiivsusega ribosüümide järjestuste valimist. Ribosüümid võivad sisaldada motiive ja/või piirkondi, mis soodustavad atsüülimist, nagu GGU motiiv ja U- rohke piirkond. Näiteks on avaldatud andmed selle kohta, et U-rohked piirkonnad võivad soodustada aminohappe substraadi äratundmist ja et GGU-motiiv suudab moodustada trna 3 otstega aluspaare. GGU-motiiv ja U-rohke piirkond koos soodustavad nii aminohappe kui ka trna samaaegset äratundmist ja seeläbi soodustavad trna 3 otsa aminoatsüülimist. Ribosüüme saab luua in vitro selekteerimise teel osaliselt randomiseeritud r24mini-ga, mis on konjugeeritud trna Asn CCCG-ga, ja sellele järgneva aktiivsetes kloonides sisalduva konsensusjärjestuse süstemaatilise konstrueerimise teel. Sellisel meetodil loodud ribosüümi nimetatakse Fx3 ribosüümiks ja seda kirjeldatakse USA patenditaotluse publikatsioonis nr 03/ : see ribosüüm on kognaatseid mittelooduslikke aminohappeid sisaldavate mitmesuguste aminoatsüül-trna-de sünteesimisel mitmekülgne katalüsaator. Aminoatsüülitud trna-de efektiivseks afiinsuspuhastamiseks võib need immobiliseerida substraadil. Sobivad substraadid on näiteks, kuid mitte ainult, agaroos, sefaroos ja magnethelmed. Ribosüümid võib RNA keemilist struktuuri ära kasutades immobiliseerida vaikudel: RNA riboosil paikneva 3'-cisdiooli oksüdeerimisel periodaadiga tekib vastav dialdehüüd, mis hõlbustab RNA immobiliseerimist vaigul. Selleks võib kasutada erinavat tüüpi vaike, sealhulgas odavaid hüdrasiinvaike, mille korral tekib amiinimisel vaigu ja ribsosüümi vahelise interkatsioni tulemusena pöördumatu side. Aminoatsüül-tRNA-de sünteesimist saab oluliselt hõlbustada aminoatsüülimise teel kolonnil. Allikas Kourouklis et al. Methods 05; 36:239-4 kirjeldatakse kolonnil põhinevat aminoatsüülimismeetodit. Aminoatsüülitud trna-sid saab isoleerida erinevatel viisidel. Ühe sobiva meetodi korral elueeritakse aminoatsüülitud trna-d kolonnist puhvriga nagu naatriumatsetaadi lahus, mis sisaldab mm EDTA-d, puhver, mis sisaldab 50 mm N-(2-hüdroksüetüül)piperasiin-N'-(3- propaansulfoonhapet), 12,5 mm KCI, ph 7,0, mm EDTA, või lihtsalt EDTA-ga puhverdatud veega (ph 7,0). Aminoatsüülitud trna-d võib lisada translatsioonireaktsioonidele, et sisestada translatsioonireaktsioonil saadavasse valku soovitud positsioonile aminohape, millega trna aminoatsüüliti. Translatsioonisüsteemid, milles võib kasutada käesoleva leiutise kohaseid aminoatsüülitud trna-sid, on näiteks, kuid mitte ainult, rakulüsaadid. Rakulüsaatidega tagatakse reaktsioonil kasutatavad koostisosad, mis on vajalikud sisend-
187 mrna-st valgu transleerimiseks. Sellised reaktsioonil kasutatavad koostisosad on näiteks, kuid mitte ainult, ribosomaalsed valgud, rrna, aminohapped, trna-d, translatsiooni initsiatsiooni ja pikendamise faktorid ja teised translatsiooniga seotud faktorid. Translatsioonisüsteemid võivad olla batch translatsioonisüsteemid või kompartmentaliseeritud translatsioonisüsteemid. Batch translatsioonisüsteemides kombineeritakse reaktsioonil kasutatavad koostisosad ühes kompartmendis, kompartmentaliseeritud translatsioonisüsteemide puhul aga eraldatakse translatsioonil kasutatavad koostisosad reaktsioonisaadustest, mis võivad vähendada translatsiooni efektiivsust. Sellised translatsioonisüsteemid on kaubanduslikult kättesaadavad. Peale selle võib kasutada seotud transkriptsiooni/translatsiooni süsteemi. Seotud transkriptsiooni/translatsioonisüsteemid võimaldavad transkribeerida sisend-dna vastavaks mrna-ks ja seda reaktsiooni koostisosade toimel omakorda transleerida. Kaubanduslikult kättesaadava seotud transkriptsiooni/translatsiooni süsteemi üheks näiteks on Rapid Translation System (RTS, Roche Inc.). Selles süsteemis on segu, mis sisaldab E.coli lüsaati, millest saadakse translatsiooniks vajalikud koostisosad nagu ribsoomid ja translatsioonifaktorid. Süsteem sisaldab ka RNA polümeraasi, mis võimaldab transkribeerida sisend-dna translatsioonil kasutatavaks mrna matriitsiks. RTS süsteemis saab reaktsiooni koostisosi kompartmentaliseerida kompartmentide, sealhulgas supply/waste compartment ja supply/waste compartment vaheliste membraanidega. trnad võib aminoatsüülida teiste ainetega ehk näiteks, kuid mitte ainult, transferaaside, polümeraaside, katalüütiliste antikehade, multifunktsionaalsete valkude ja teiste sarnaste ainetega. Stephan kirjaldab allikas Scientist 05 Oct ; lk 30-33, teisi meetodeid, mille abil saab valkudesse sisestada mittelooduslikult kodeeritud aminohappeid. Lu et al. in Mol Cell. 01 Owt;8(4): kirjeldavad meetodit, mille korral valk ligeeritakse keemiliselt sünteetilise peptiidiga, mis sisaldab mittelooduslikke aminohappeid (ekspresseeritud valgu ligeerimine). Aminohappeid on valkudesse sisestatud ka mikroinjektsiooni meetoditel. Vt nt M. W. Nowak, P. C. Kearney, J. R. Sampson, M. E. Saks, C. G. Labarca, S. K. Silverman, W. G. Zhong, J. Thorson, J. N. Abelson, N. Davidson, P. G. Schultz, D. A. Dougherty and H. A. Lester, Science, 268:439 (1995); and, D. A. Dougherty, Curr. Opin. Chem. Biol., 4:645 (00). Konna Xenopus ootsüüti koinjekteeriti kaks in vitro valmistatud RNA liiki/molekuli (species): mrna, mis kodeeris sihtvalku, millel oli huvipakkuvas aminohappe positsioonis stoppkoodon UAG, ja amber-supressor-trna-d, mis oli aminoatsüülitud soovikohase mitteloodusliku aminohappega. Ootsüüdi
188 translatsioonimehhanism lisab seejärel UAG poolt määratud positsioonile mitteloodusliku aminohappe. See meetod on võimaldanud uurida terviklike membraanivalkude ehk valkude, mida üldjuhul ei saa uurida in vitro ekspressioonisüsteemides, struktuuri ja funktsiooni in vivo. Asjakohaste näidete seas on fluorestseeruva aminohappe sisestamine tahhükiini neurokiniin-2 retseptorisse, et mõõta fluorestsentsi resonantsenergia ülekande järgi kaugusi, vt nt G. Turcatti, K. Nemeth, M. D. Edgerton, U. Meseth, F. Talabot, M. Peitsch, J. Knowles, H. Vogel and A. Chollet, L Biol. Chem., 271:19991 (1996); biotinüleeritud aminohapete sisestamine ioonkanalite pinnal eksponeerinud jääkide identifitseerimiseks, vt. näiteks., J. P. Gallivan, H. A. Lester and D. A. Dougherty, Chem. Biol., 4:739 (1997); türosiini puuristatud (caged) analoogide kasutamine konformatsiooniliste muutuste jälgimiseks ioonkanalis reaalajas, vt. näiteks J. C. Miller, S. K. Silverman, P. M. England, D. A. Dougherty and H. A. Lester, Neuron, :619 (1998); ja alfa-hüdroksüaminohapete kasutamine ioonkanalite selgroogude muutmiseks nende puuristamise mehhanismide uurimiseks. Vt nt P. M. England, Y. Zhang, D. A. Dougherty and H. A. Lester, Cell, 96:89 (1999); and, T. Lu, A. Y. Ting, J. Mainland, L. Y. Jan, P. G. Schultz and J. Yang, Nat. Neurosci., 4:239 (01). Võimalus sisestada mittelooduslikke aminohappeid otse valkudesse in vivo annab mitmesuguseid eeliseid, sealhulgas, kuid mitte ainult, mutantsete valkude kõrge saagise, tehnilise lihtsuse, võimaluse uurida mutantseid valke rakkudes ja võimalik et ka elusorganismides ja kasutatada neid valke terapeutilises ravis ja diagnostikas. Võimalus sisestada valkudesse erineva suuruse, happelisuse, nukleofiilsuse, hüdrofoobsuse ja teiste omadustega mittelooduslikke aminohappeid võib oluliselt suurendada meie võimalusi valkude struktuure ratsionaalselt ja süstemaatiliselt töödelda nii valgu kasutamiseks sondina kui ka uudsete omadustega uute valkude või organismide loomiseks. Ühes para-f-phe kohtspetsiifilise sisestamise katses kasutati p-f-phe resistentses, Phe auksotroofses Escherichia coli tüves pärmi amber-supressori trnaphecua /fenüülalanüül-trna süntetaasi paari. Vt nt R. Furter, Protein Sci., 7:419 (1998). Käesoleva leiutise kohast FGF-21 polünukleotiidi võib olla võimalik ekspresseerida ka rakuvaba (in vitro) translatsioonisüsteemi abil. Translatsioonisüsteemid võivad olla rakupõhised või rakuvabad ja prokarüootsed või eukarüootsed. Rakupõhiste translatsioonisüsteemide seas on näiteks, kuid mitte ainult, terviklike rakkude preparaadid nagu permeabiliseeritud rakud või rakukultuurid, milles saab soovitud nukleiinhappelise
189 järjestuse transkribeerida mrna-ks ja saadud mrna transleerida. Rakuvabad translatsioonisüsteemid on kaubanduslikult kättesaadavad ja tuntud on paljud erinevad tüübid ja süsteemid. Rakuvabade süsteemide seas on näiteks, kuid mitte ainult, prokarüootsed lüsaadid nagu Escherichia coli lüsaadid ja eukarüootsed lüsaadid nagu nisuidu lüsaadid, putukarakkude lüsaadid, küüliku retikulotsüütide lüsaadid, küüliku ootsüüdi lüsaadid ja inimrakkude lüsaadid. Kui saadav valk peab olema glükosüülitud, fosforüülitud või muul viisil modifitseeritud, on eelistatud eukarüootsed ekstraktid või lüsaadid, kuna paljusid selliseid modifikatsioone on võimalik teha ainult eukarüootsetes süsteemides. Mõned sellised ekstraktid ja lüsaadid on kaubanduslikult kättesaadavad (Promega; Madison, Wis.; Stratagene; La Jolla, Calif.; Amersham; Arlington Heights, III.; GIBCO/BRL; Grand Island, N.Y.). Saadaval on ka membraanide ekstraktid nagu mikrosomaalseid membraane sisaldavad koera pankrease ekstraktid, mis on kasulikud sekretoorsete valkude transleerimisel. Sellistes süsteemides, mis võivad sisaldada matriitsina mrna-d (in vitro translatsioon) või DNA-d (kombineeritud in vitro transkriptsioon ja translatsioon), juhivad in vitro sünteesi ribosoomid. Praeguseks on tehtud olulisi jõupingutusi rakuvabade valgu ekspressioonisüsteemide väljatöötamiseks. Vt nt Kim, D.M. and J.R. Swartz, Biotechnology and Bioengineering, 74 : (01); Kim, D.M. and J.R. Swartz, Biotechnology Letters, 22, 37-42, (00); Kim, D.M., and J.R. Swartz, Biotechnology Progress, 16, , (00); Kim, D.M., and J.R. Swartz, Biotechnology and Bioengineering, 66, , (1999); ja Patnaik, R. and J.R. Swartz, Biotechniques 24, , (1998); USA patent nr ; USA patendipublikatsioon nr 02/ ; WO 00/55353; WO 90/ Teine strateegia, mida võib rakendada mittelooduslikult kodeeritud FGF-21 polüpeptiidide ekspresseerimisel, hõlmab mrna peptiidide fusioonimeetodit. Vt nt R. Roberts and J. Szostak, Proc. Natl Acad. Sci. (USA) 94: (1997); A. Frankel, et al., Chemistry & Biology :43-50 (03). Selle strateegia korral transleeritakse püromütsiiniga seotud mrna matriits peptiidiks ribosoomil. Kui üks või mitu trna molekuli on modifitseeritud, saab peptiidi sisestada ka mittelooduslikud aminohapped. Pärast viimase mrna koodoni lugemist haakub püromütsiin peptiidi C-otsaga. Kui in vitro analüüsis selgub, et saadud valgul on huvitavad omadused, saab selle identiteedi kergesti määrata mrna järjestuse järgi. Sellisel viisil saab ühte või mitut mittelooduslikult kodeeritud aminohapet sisaldavate FGF-21 polüpeptiidide pankade
190 5 188 skriinimise teel identifitseerida soovitud omadustega polüpeptiidid. Hiljem on avaldatud ka in vitro ribosoomide translatsioonid puhastatud koostisosadega, mis võimaldavad sünteesida peptiide, milles on tehtud asendusi mittelooduslike aminohapetega. Vt nt A. Forster et al., Proc. Natl Acad Sci. (USA) 0:6353 (03). Lisaks võib kasutada taastatud translatsioonisüsteeme. mrna-d on õnnestunud transleerida valguks ka puhastatud translatsioonifaktorite segudega ja ka lüsaatide kombinatsioonidega või lüsaatidega, mis sisaldavad puhastatud translatsioonifaktoreid nagu initsiatsioonifaktor 1 (IF-1), IF-2, IF-3 (a või β), elongatsioonifaktor T (EF-Tu) või terminatsioonifaktorid. Rakuvabu süsteeme võib kombineerida ka transkriptsiooni/translatsioonisüsteemidega, mille korral sisestatakse süsteemi DNA, transkribeeritakse see mrna-ks ja mrna transleeritakse viisil, mida kirjeldatakse allikas Current Protocols in Molecular Biology (F. M. Ausubel et al. editors, Wiley Interscience, 1993). Eukarüootses transkriptsioonisüsteemis transkribeeritud RNA võib olla heteronukleaarse RNA (hnrna) või 5'-otsa cap-struktuurid (7-metüülguanosiin) ja 3'-otsa poly A sabaga küpse mrna kujul, mis võib olla teatud translatsioonisüsteemides eeliseks. Mütsistatud (capped) mrna-sid näiteks transleeritakse väga efektiivselt retikulotsüütide lüsaadil põhinevas süsteemis. IX. FGF-21 polüpeptiidide sidumine makromolekulaarsete polümeeridega 30 Käesolevas dokumendis kirjeldatavate kompositsioonide, meetodite, tehnikate ja strateegiate abil saab teha mittelooduslikke aminohappeid sisaldavates polüpeptiidides mitmesuguseid modifikatsioone. Selliste modifikatsioonide hulka kuulub veel ühe funktsionaalse üksuse lisamine polüpeptiidi mittelooduslikust aminohappest koostisosa külge, kusjuures selline funktsionaalne üksus võib olla näiteks, kuid mitte ainult, märgis, värvaine, polümeer, vees lahustuv polümeer, polüetüleenglükooli derivaat, fotoristside, radionukliid, tsütotoksiline ühend, raviaine, afiinsusmärgis, fotoafiinsusmärgis, reaktsioonivõimeline ühend, vaik, teine valk või polüpeptiid või polüpeptiidi analoog, antikeha või antikeha fragment, metallkelaator, kofaktor, rasvhape, süsivesik, polünukleotiid, DNA, RNA, antisenss-polünukleotiid, sahhariid, vees lahustuv dendrimeer, tsüklodekstriin, inhibeeriv ribonukleiinhape, bioloogiline materjal, nanoosake, spin-märgis, fluorofoor, metalli sisaldav fragment, radioaktiivne fragment, uudne funktsionaalrühm,
191 teiste molekulidega kovalentselt või mittekovalentselt interakteeruv rühm, valguse pioolt lõhustatav fragment, aktiiniga kiiritamise teel ergastatav fragment, fotoisomeriseeritav fragment, biotiin, biotiini derivaat, biotiini analoog, rasket aatomit sisaldav rühm, keemiliselt lõhustatav rühm, valguse toimel lõhustatav rühm, pikendatud kõrvalahel, süsinikuga seotud suhkur, redoks-aktiivne aine, amino-tiohape, toksiline fragment, isotoobiga märgistatud fragment, biofüüsikaline sond, fosforestseeruv märgis, kemoluminetseeruv märgis, elektrontihe rühm, magnetiline rühm, interkaleeruv rühm, kromofoor, energia ülekande aine (energy transfer agent), bioloogiliselt aktiivne aine, tuvastatav märgis, väike molekul, kvantpunkt, nanotransmitter, radionukleotiid, radiotransmitter, neutronhaarde agent või eespool nimetatud ainete kombinatsioon või mis tahes teine soovikohane ühend või aine. Käesoleva leiutise kohaste kompositsioonide, meetodite, tehnikate ja strateegiate illustreeriva mittepiirava näitena kirjeldatakse järgnevalt makromolekulaarsete polümeeride lisamist mittelooduslikke aminohappeid sisaldavale polüpeptiidile põhimõttel, et selliste kompositsioonide, meetodite, tehnikate ja strateegiate abil (tehes neis vajaduse korral asjakohaseid modifikatsioone, mida asjatundjad suudavad teha käesolevas dokumendid kirjeldatu põhjal) saab lisada ka teisi funktsionaalseid üksusi, sealhulgas, kuid mitte ainult, neid, mis on loetletud eespool. Selleks et moduleerida FGF-21 polüpeptiidi bioloogilisi omadusi ja/või tagada FGF-21 polüpeptiidile uusi bioloogilisi omadusi, saab leiutisekohasele FGF-21 polüpeptiidile lisada mitmesuguseid makromolekulaarseid polümeere ja teisi molekule. Sellised makromolekulaarsed polümeerid saab siduda FGF-21 polüpeptiidiga looduslikult kodeeritud aminohappe kaudu, mittelooduslikult kodeeritud aminohappe kaudu või loodusliku või mitteloodusliku aminohappe mis tahes funktsionaalse asendaja kaudu või looduslikule või mittelooduslikule aminohappele lisatud mis tahes asendaja või funktsionaalse rühma kaudu. Polümeeri molekulmass võib jääda laia vahemikku ehk olla näiteks, kuid mitte ainult, umbes 0 Da kuni umbes Da või suurem. Polümeeri molekulmass võib olla umbes 0 Da kuni umbes Da, sealhulgas, kuid mitte ainult, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, 000 Da, 000 Da, 000 Da, 000 Da, 9000 Da, 8000 Da, 000 Da, 6000 Da, 5000 Da, 4000 Da, 3000 Da, 00 Da, 1,000 Da, 900 Da, 800 Da, 700 Da, 600 Da, 500 Da, 400 Da, 300 Da, 0 Da ja 0 Da. Mõnes teostuses on polümeeri molekulmass umbes 0 Da kuni umbes Da. Mõnes teostuses on polümeeri molekulmass umbes
192 Da kuni umbes Da. Mõnes teostuses on polümeeri molekulmass umbes 00 Da kuni umbes Da. Mõnes teostuses on polümeeri molekulmass umbes 5000 Da kuni umbes Da. Mõnes teostuses on polümeeri molekulmass umbes 000 Da kuni umbes Da. Käesoleva leiutisega pakutakse polümeeri:valgu konjugaatide üldjoontes homogeensed preparaadid. Käesolevas kontekstis tähendab üldjoontes homogeenne seda, et polümeeri:valgu konjugaadi molekulid moodustavad rohkem kui poole valgu üldmassist. Polümeeri:valgu konjugaat on bioloogiliselt aktiivne ja käesolevas dokumendis pakutavad üldjoontes homogeensed PEGüleeritud FGF-21 polüpeptiidid on piisavalt homogeensed, et neil oleksid homogeensele preparaadile omased eelised ehk nende farmakokineetikat saab kliinilistes rakendustes partiist partiisse kergesti ennustada. Lisaks võib valmistada polümeeri:valgu konjugaadi molekulide segu, mille eeliseks on see, et segus sisalduva monopolümeeri: valgu konjugaadi proportsiooni saab valida. Mitmesugustest valkudest, mille külge on kinnitatud erineval hulgal polümeerifragmente (st di-, tri-, tetra-, jne.), saab seega soovi korral valmistada segu ja kombineerida nimetatud konjugaate leiutisekohastel meetoditel valmistatud monopolümeeri: valgu konjugaadiga, saades tulemusena segu, milles on monopolümeeri: valgu konjugaatide etteantud osakaal. Kasutatav polümeer võib olla vees lahustuv, nii et valk, mille külge see kinnitatakse, ei sadestu vesikeskkonnas nagu füsioloogiline keskkond. Polümeer võib olla hargnenud või hargnemata. Kui lõppsaadust kasutatakse raviotstarbeliselt, peab selles sisalduv polümeer olema farmatseutiliselt sobiv. Sobivate polümeeride seas on näiteks, kuid mitte ainult, polüalküüleetrid ja nende alkoksü- mütsistatud analoogid (nt polüoksüetüleenglükool, polüoksüetüleen/propüleenglükool ja nende metoksü- või etoksü- mütsistatud analoogid, eeskätt polüoksüetüleenglükool ehk polüetüleenglükool ehk PEG); polüvinüülpürrolidoonid; polüvinüülalküüleetrid; polüoksasoliinid, polüalküüloksasoliinid ja polühüdroksüalküüloksasoliinid; polüakrüülamiidid, polüalküülakrüülmiidid ja polühüdroksüalküülakrüülamiidid (nt polühüdroksüpropüülmetakrüülamiid ja selle derivaadid); polühüdroksüalküülakrülaadid; polüsiaalhapped ja nende analoogid; hüdrofiilsete peptiidide järjestused; polüsahhariidid ja nende derivaadid, sealhulgas dekstraan ja dekstraani derivaadid, nt karboksümetüüldekstraan, dekstraansulfaadid, aminodekstraan; tselluloos ja selle derivaadid, nt karbokspmetüültselluloos, hüdroksüalküültselluloosid; kitiin ja selle derivaadid, nt kitosaan, suktsinüülkitosaan, karboksüraetüülkitiin, karboksümetüülkitosaan; hüaluroonhape ja selle derivaadid; tärklised; alginaadid;
193 kondroitiinsulfaat; albumiin; pullulaan ja karboksümetüülpullulaan; polüaminohapped ja nende derivaadid, nt polüglutamiinhapped, polülüsiinid, polüaspartaamhapped, polüaspartamiidid; maleiinhappe anhüdriidi kopolümeerid nagu: stüreeni maleiinhappe anhüdriidi kopolümeer, divinüületüüleetri maleiinhappe anhüdriidi kopolümeer; polüvinüülalkoholid; nende kopolümeerid; nende terpolümeerid; nende segud; ja eespool nimetatud ühendite derivaadid. Polüetüleenglükooli ja valgumolekulide suhe ja ka nende kontsentratsioon reaktsioonisegus on erinevad. Üldjuhul määratakse nende optimaalne suhe (reaktsiooni efektiivsuse mõistes ehk minimaalse reageerimata valgu või polümeeri koguse tagamiseks) valitud polüetüleenglükooli molekulmassi ja olemasolevate reaktsioonivõimeliste rühmade arvu järgi. Molekulmassi osas kehtib üldiselt põhimõte, et mida suurem on polümeeri molekulmass, seda vähem polümeeri molekule võib valgu külge kinnitada. Selliste parameetrite optimeerimisel tuleb võtta arvesse ka polümeeri hargnemist. Üldiselt kehtib põhimõte, et mida suurem on molekulmass (või mida suurem on ahelate arv), seda suurem on polümeeri:valgu suhe. Käesolevas kontekstis ja PEG:FGF-21 polüpeptiidi konjugaatide puhul on terapeutiliselt efektiivne kogus mis tahes kogus, mis tekitab patsiendi organismis soovitud kasuliku efekti. See kogus on indiviiditi erinev ja sõltub paljudest faktoritest, sealhulgas patsiendi üldisest füüsilisest seisundist ja ravitava seisundi põhjustest. Ravis kasutatava FGF-21 polüpeptiidi kogus tagab sobiva muundumise kiiruse ja säilitab soovitud vastuse organismile kasulikul tasemel. Asjatundjad oskavad avalikkusele kättesaadavate materjalide ja protseduuride abil kergesti määrata leiutisekohaste kompositsioonide terapeutiliselt efektiivse koguse. Vees lahustuv polümeer võib olla mis tahes struktuuriga, sealhulgas, kuid mitte ainult, lineaarse, kahvlikujulise või hargnenud struktuuriga. Vees lahustuv polümeer on üldjuhul polü(alküleenglükool) nagu polü(etüleenglükool) (PEG), kuid kasutada võib ka teisi vees lahustuvaid polümeere. Leiutise teatud teoste kirjeldamisel kasutatakse PEG-i. PEG on hästi tuntud vees lahustuv polümeer, mis on kaubanduslikult kättesaadavav ja mida saab asjatundjatele tuntud meetoditel valmistada etüleenglükooli tsükli avamise polümerisatsiooni teel
194 192 (Sandler and Karo, Polymer Synthesis, Academic Press, New York, Vol. 3, lk ). Mõistet PEG kasutatakse laias tähenduses ja see hõlmab mis tahes polüetüleenglükooli molekuli olenemata selle suurusest või PEG otsa modifikatsioonist ning seda saab FGF- 21 polüpeptiidiga seotuna tähistada valemiga: 5 XO-(CH 2 CH 2 O) n -CH 2 CH 2 -Y milles n on 2 kuni 000 ja X on H või otsa (terminaalne) modifikatsioon, sealhulgas, kuid mitte ainult, C 1-4 alküülrühm, kaitserühm või terminaalne funktsionaalrühm. Mõnel juhul on leiutise korral kasutatava PEG-i ühes otsas hüdroksü- või metoksürühm ehk X on H või CH 3 ( metoksü-peg ). Selle asemel võib PEG-i ühes otsas olla reaktsioonivõimeline rühm, mille tulemusena moodustub bifunktsionaalne polümeer. Tüüpiliste reaktsioonivõimeliste rühmade seas on reaktsioonivõimelised rühmad, mida tavaliselt kasutatakse reaktsioonides funktsionaalrühmadega, mida leidub tavapärases aminohappes (sealhulgas, kuid mitte ainult, maleimiidrühmade, aktiveeritud karbonaatide (sealhulgas, kuid mitte ainult, p-nitrofenüülestriga), aktiveeritud estrites (sealhulgas, kuid mitte ainult, N-hüdroksüsuktsinimiidi, p-nitrofenüülestriga) ja aldehüüdidega), ja ka funktsionaalsete rühmadega, mis on tavapärase aminohappe suhtes inertsed, kuid reageerivad spetsiifiliselt komplementaarsete aminohapetga, mida sisaldavad mittelooduslikult kodeeritud aminohapped (sealhulgas, kuid mitte ainult, asiidrühmade ja alküünrühmadega). PEG teine ots, mis on eespool toodud valemis tähistatud tähega Y, kinnitub FGF-21 polüpeptiidi külge otse või kaudselt loodusliku või mittelooduslikult kodeeritud aminohappe kaudu. X võib näiteks olla amiid-, karbamaat- või uureaside polüpeptiidi amiinrühmaga (sealhulgas, kuid mitte ainult, lüsiini või N-otsa epsilon-amiiniga. Selle asemel võib Y olla tioolrühma külge kinnituv maleimiidrühm (sealhulgas, kuid mitte ainult, tsüsteiini tioolrühm). Y võib olla ka side aminohappejäägiga, millele ei pääse tavapärase aminohappe korral üldjuhul ligi. Näiteks PEG-i asiidrühma saab panna reageerima FGF- 21 alküünrühmaga, mille tulemusel moodustub Huisgen [3+2] tsükloliitumise saadus.
195 Sarnase saaduse valmistamiseks saab PEG-i alkünüülrühma panna reageerima mittelooduslikult kodeeritud aminohappes sisalduva asiidrühmaga. Mõnes teostuses saab tugeva nukleofiili (sealhulgas, kuid mitte ainult, hüdrasiini, hüdrasiidi, hüdroksüülamiini, semikarbamiidi) panna reageerima mittelooduslikus aminohappes sisalduva aldehüüd- või ketoonrühmaga, saades tulemusena hüdrasooni, oksiimi või semikarbasooni, mida saab mõnel juhul sobiva redutseeriva ainega veelgi redutseerida. Tugeva nukleofiili saab sisestada FGF-21 polüpeptiidi ka mittelooduslikult kodeeritud aminohappega ja panna seal reageerima eelkõige vees lahustuvas polümeeris sisalduva ketoon- või aldehüüdrühmaga. Kasutatav PEG võib põhimõtteliselt olla mis tahes soovitud molekulmassiga, sealhulgas, kuid mitte ainult molekulmassiga umbes 0 daltonit (Da) kuni Da või soovi korral suurema molekulmassiga (näiteks, kuid mitte ainult, vahel 0,1-50 kda või -40 kda). PEG-i molekulmass võib jääda laia vahemikku ja olla näiteks, kuid mitte ainult, umbes 0 Da kuni umbes Da või suurem. PEG-i molekulmass võib olla umbes 0 Da kuni umbes Da, näiteks, kuid mitte ainult, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, 000 Da, 000 Da, 000 Da, 000 Da, 9000 Da, 8000 Da, 7000 Da, 6000 Da, 5000 Da, 4000 Da, 3000 Da, 00 Da, 00 Da, 900 Da, 800 Da, 700 Da, 600 Da, 500 Da, 400 Da, 300 Da, 0 Da ja 0 Da. Mõnes teostuses on PEG-i molekulmass umbes 0 Da kuni umbes Da. Mõnes teostuses on PEG-I molekulmass umbes 0 Da kuni umbes Da. Mõnes teostuses on PEG-i molekulmass umbes 00 Da kuni umbes Da. Mõnes teostuses on PEG-i molekulmass umbes 5000 Da kuni umbes Da. Mõnes teostuses on PEG-i molekulmass umbes 000 Da kuni umbes Da. Leiutise korral võib kasutada ka hargnenud ahelaga PEG-id, sealhulgas, kuid mitte ainult, PEG molekule, mille iga ahela molekulmass on 1-0 kda (sealhulgas, kuid mitte ainult, 1-50 kda või 5- kda). Hargnenud ahelaga PEG-i iga ahela molekulmass võib olla näiteks, kuid mitte ainult, umbes 00 Da kuni umbes Da või suurem. Hargnenud ahelaga PEG-i iga ahela molekulmass võib olla umbes 00 Da kuni umbes Da, näiteks, kuid mitte ainult, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, 000 Da, 000 Da, 000 Da, 000 Da, 9000 Da, 8000 Da, 7000 Da, 6000 Da, 5000 Da, 4000 Da, 3000 Da, 00 Da ja 00 Da. Mõnes teostuses on hargnenud ahelaga PEG-
196 5 194 i iga ahela molekulmass umbes 00 Da kuni umbes Da. Mõnes teostuses on hargnenud ahelaga PEG-i iga ahela molekulmass umbes 00 Da kuni umbes Da. Mõnes teostuses on hargnenud ahelaga PEG-i iga ahela molekulmass umbes 5000 Da kuni umbes Da. Mõnes teostuses on hargnenud ahelaga PEG-i iga ahela molekulmass umbes 5000 Da kuni umbes 000 Da. Paljusid erinevaid PEG molekule kirjeldatakse näiteks, kuid mitte ainult, ettevõtte Shearwater Polymers, Inc. kataloogis Nektar Therapeutics Catalog. Üldiselt on PEG molekuli ainult üks ots vaba mitteloodulikult kodeeritud aminohappega reageerimiseks. PEG-i kinnitamiseks käesolevas dokumendis kirjeldataval viisil mittelooduslikult kodeeritud aminohapete külge võib kasutada PEG-i derivaate, millel on alküün- ja asiidfragmendid, mis reageerivad aminohapete kõrvalahelatega. Kui mittelooduslikult kodeeritud aminohape sisaldab asiidrühma, sisaldab PEG üldjuhul alküünfragmenti, mille abil saab moodustada [3+2] tsükloliitumise saaduse või aktiveeritud PEGi derivaati (st estrit, karbonaati), mis sisaldab amiidsideme moodustamiseks vajalikku fosfiinrühma. Kui mittelooduslikult kodeeritud aminohape sisaldab alkünüülrühma, sisaldab PEG üldjuhul asiidfragmenti, mille abil saab moodustada [3+2] Huisgeni tsükloliitumise saaduse. Kui mittelooduslikult kodeeritud aminohape sisaldab karbonüülrühma, sisaldab PEG üldjuhul tugevatoimelist nukleofiili (sealhulgas, kuid mitte ainult, funktsionaalset hüdrasiid-, hüdrasiin-, hüdroksüülamiin- või semikarbasiidfragmenti), et moodustuks vastav hüdrasoon-, oksiim- ja semikarbasoonside. Teiste alternatiivsete võimaluste korral saab kasutada eespool kirjeldatavate reaktsioonivõimeliste rühmade vastupidist orientatsiooni ehk mittelooduslikult kodeeritud asiidfragmendi saab panna reageerima alküünrühma sisaldava PEG derivaadiga. 30 Mõnes teostuses sisaldab PEG derivaadiga seotud FGF-21 polüpeptiidi variant keemilist funktsionaalrühma, mis reageerib mittelooduslikult kodeeritud aminohappe kõrvalahelas paikneva keemilise funktsionaalrühmaga. Leiutise mõne teostusega pakutakse asiid- ja atsetüleenrühma sisaldavad polümeeri derivaadid, millel on veees lahustuva polümeeri selgroog keskmise molekulmassiga umbes 800 Da kuni umbes Da. Vees lahustuva polümeeri polümeer-selgroog võib olla polü(etüleenglükool). Samas tuleb silmas pidada seda, et käesolevas leiutise korral
197 195 võib kasutada paljusid erinevaid veeslahustuvaid polümeere, näiteks, kuid mitte ainult, polü(etüleen)glükooli ja teisi sarnaseid polümeere, sealhulgas polü(dekstraani) ja polü(propüleenglükooli) ja mõiste PEG ehk polü(etüleenglükool) all peetakse silmas kõiki selliseid molekule Mõiste PEG hõlmab näiteks, kuid mitte ainult, polü(etüleenglükooli) kõiki vorme, sealhulgas bifunktsionaalset PEG-i, mitmeõlgset PEG-i, derivatiseeritud PEG-i, kahvlikujulist PEG-i, hargnenud PEG-i, pendent PEG-i (ehk PEG-i või selle sarnaseid polümeere, mille polümeer-selgroo küljes ripub üks või mitu funktsionaalrühma) või PEG-i, milles on lõhustatavad sidemed. PEG on üldjuhul läbipaistev, värvitu, lõhnatu, vees lahustuv, termiliselt stabiilne, paljude keemiliste ainete suhtes inertne ja üldiselt mittetoksiline aine, mis ei hüdrolüüsu ega lagune. Polü(etüleenglükooli) peetakse bioloogiliselt ühilduvaks ehk PEG ei kahjusta neis olles elusaid kudesid või organisme. PEG on täpsemalt mitteimmunogeenne ehk PEG ei kutsu organismis esile immuunvastust. Kui PEG on kinnitatud organismis soovitud funktsiooni omava molekuli nagu bioloogiliselt aktiivne molekul külge, PEG üldjuhul maskeerib seda ainet ja suudab vähendada või kaotada mis tahes immuunvastust, mille tulemusena organism talub seda ainet. PEG-i konjugaadid ei tekita üldjuhul märkimisväärset immuunvastust ega põhjusta klompide teket (clotting) ega teisi soovimatuid kõrvalnähte. Käesoleva leiutise korral võib kasutada PEG-i valemiga -- CH 2 CH 2 O--(CH 2 CH 2 O) n - CH 2 CH 2 --, milles n on umbes 3 kuni umbes 4000, üldjuhul umbes kuni umbes 00. Käesoleva leiutise mõnes teostuses on eriti kasulik PEG, mille molekulmass on umbes 800 Da kuni umbes Da. PEG-I molekulmass võib jääda laia vahemikku ehk olla näiteks, kuid mitte ainult, umbes 0 Da kuni umbes Da või suurem. PEG-i molekulmass võib olla umbes 0 Da kuni umbes Da, sealhulgas, kuid mitte ainult, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, 000 Da, 000 Da, 000 Da, 000 Da, 9000 Da, 8000 Da, 7000 Da, 6000 Da, 5000 Da, 4000 Da, 3000 Da, 00 Da, 00 Da, 900 Da, 800 Da, 700 Da, 600 Da, 500 Da, 400 Da, 300 Da, 0 Da ja 0 Da. Mõnes teostuses on PEG-i molekulmass umbes 0 Da kuni umbes Da. Mõnes teostuses on PEG-i molekulmass umbes 0 Da kuni umbes Da. Mõnes teostuses on PEG-i molekulmass umbes 00 Da kuni umbes Da. Mõnes teostuses on PEG-i
198 5 196 molekulmass umbes 5000 Da kuni umbes Da. Mõnes teostuses on PEG-i molekulmass umbes 000 Da kuni umbes Da. Polümeeri selgroog võib olla lineaarne või hargenud. Hargnenud polümeeri selgroog on tehnika tasemes tuntud. Hargnenud polümeeril on üldjuhul keskne ahelate põhifragment ja keskse ahelate põhifragmendiga seotud mitu lineaarset polümeeriahelat. PEG-i puhul kasutatakse tavaliselt selle hargnenud vorme, mida saab valmistada etüleenoksiidi lisamise teel mitmesugustele polüoolidele nagu glütserool, glütserooli oligomeerid, pentaerütritool ja sorbitool. Keskse ahelate põhifragmendi võib deriveerida ka mitmest aminohappest nagu lüsiin. Hargnenud polü(etüleenglükooli) üldvormi võib tähistada valemiga R(-PEG-OH) m, milles R pärineb põhifragmendist nagu glütserool glütserooli oligomeerid või pentaerütritool ja m on õlgade arv. Polümeeri selgroona võib kasutada mitmeõlgseid PEGi molekule nagu molekulid, mida kirjeldatakse USA patentides nr , , , , USA patenditaotluses 03/ ning patendidokumentides WO 96/21469 ja WO 93/219. Hargnenud PEG võib olla ka kahvlikujuline PEG valemiga PEG(-YCHZ 2 ) n, milles Y on siduv rühm ja Z on aktiveeritud terminaalne rühm, mis on aatomitest koosneva määratletud pikkusega ahela kaudu seotud CH-fragmendiga. Veel ühel hargnenud vormil ehk pendant PEG-il ei paikne reaktsioonivõimelised rühmad nagu karbonüülrühmad mitte PEG ahelate otstes, vaid pigem piki ahelat. Lisaks nimetatud PEG-i vormidele saab valmistada ka polümeeri, mille selgroos on nõrgad või lõhustatavad sidemed. Näiteks saab valmistada PEG-i, mille polümeeri selgroos on estersidemed, mis hüdrolüüsuvad. Nagu on näidatud allpool, tekivad polümeeri hüdrolüüsi tulemusena väiksema molekulmassiga fragmendid: -PEG-CO 2 -PEG-+H 2 O PEG-CO 2 H+HO-PEG- 30 Asjatundjad mõistavad, et mõiste polü(etüleenglükool) ehk PEG hõlmab selle kõiki tehnika tasemes tuntud vorme, sealhulgas, kuid mitte ainult, käesolevas dokumendis käsitletavaid vorme. Käesoleva leiutise korral võib kasutada ka paljusid teisi polümeere. Leiutise mõne teostuse korral on eriti kasulikud vees lahustuvad polümeeri selgrood, millel on 2 kuni umbes 300 otsa. Sobivad polümeerid on näiteks, kuid mitte ainult, teised polü(alküleenglükoolid) nagu polü(propüleenglükool) (PPG), nende kopolümeerid (sealhulgas, kuid mitte ainullt, etüleenglükooli ja propüleenglükooli kopolümeerid), nende terpolümeerid, nende segud ja muud sarnased polümeerid. Kuigi polümeeri selgroo iga
199 5 197 ahela molekulmass võib olla erinev, on see tavaliselt umbes 800 Da kuni umbes Da, sageli umbes 6000 Da kuni umbes Da. Polümeeri selgroo iga ahela molekulmass võib olla umbes 0 Da kuni umbes Da, näiteks, kuid mitte ainult, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, Da, 000 Da, 000 Da, 000 Da, 000 Da, 9000 Da, 8000 Da, 7000 Da, 6000 Da, 5000 Da, 4000 Da, 3000 Da, 00 Da, 00 Da, 900 Da, 800 Da, 700 Da, 600 Da, 500 Da, 400 Da, 300 Da, 0 Da ja 0 Da. Mõnes teostuses on polümeeri iga ahela molekulmass umbes 0 Da kuni umbes Da. Mõnes teostuses on polümeeri iga ahela molekulmass umbes 0 Da kuni umbes Da. Mõnes teostuses on polümeeri iga ahela molekulmass umbes 00 Da kuni umbes Da. Mõnes teostuses on polümeeri iga ahela molekulmass umbes 5000 Da kuni umbes Da. Mõnes teostuses on polümeeri iga ahela molekulmass umbes 000 Da kuni umbes Da. Mõnes teostuses on polümeeri iga ahela molekulmass umbes 000 Da kuni umbes Da. Asjatundjad mõistavad, et eespool toodud üldjoontes vees lahustuvate selgroogude loend ei ole mitte mingil juhul täielik, vaid on illustreeriv ja et kõik polümeersed materjalid, millel on eespool kirjeldatavad omadused, sobivad käesoleva leiutise korral kasutamiseks. Leiutise mõnes teostuses on polümeeri derivaadid multifunktsionaalsed ehk polümeeri selgrool on vähemalt kaks otsa ja võimalik et kuni umbes 300 otsa, mis on funktsionaalrühmaga funktsionaliseeritud või aktiveeritud. Multifunktsionaalsete polümeeri derivaatide seas on näiteks, kuid mitte ainult, kahe otsaga lineaarsed polümeerid, kusjuures iga ots on seotud sama või erineva funktsionaalse rühmaga. Ühe teostuse korral on polümeeri derivaat struktuuriga: X-A-POLY-B-N=N=N milles: N=N=N on asiidfragment; B on siduv fragment, mis võib olla olemas või puududa;
200 198 POLY on vees lahustuv mitteantigeenne polümeer; A on siduv fragment, mis võib olla olemas või puududa ja mis võib olla sama mis B või sellest erineda ning X on teine funktsionaalrühm Siduva fragmendi A ja B näidete seas on näiteks, kuid mitte ainult, mitme üksusega funktsionaliseeritud alküülrühm, milles on kuni 18 ja võib olla 1- süsinikuaatomit. Alküülrühm võib sisaldada heteroaatomit nagu lämmastiku-, hapniku- või väävliaatom. Alküülrühm võib olla heteroaatomi juures hargnenud. Siduva fragmendi A ja B teiste näidete seas on näiteks, kuid mitte ainult, mitme üksusega funktsionaliseeritud arüülrühm, milles on kuni ja võib olla 5-6 süsinikuaatomit. Arüülrühm võib olla asendatud veel ühe süsiniku-, lämmastiku-, hapniku- või väävliaatomiga. Sobivate siduvate rühmade teiste näidete seas on siduvad rühmad, mida kirjeldatakse USA patentides nr , ja USA patenditaotluse publikatsioonis 03/ Asjatundjad mõistavad, et eespool toodud siduvate fragmentide loend ei ole mitte mingil juhul täielik, vaid on illustreeriv ja et kõik siduvad fragmendid, millel on eespool kirjeldatavad omadused, sobivad käesoleva leiutise korral kasutamiseks. Üksusena X kasutamiseks sobivate funktsionaalsete rühmade seas on näiteks, kuid mitte ainult, hüdroksüül-, kaitstud hüdroksüül-, alkoksüülrühm, aktiivne ester nagu N- hüdroksüsuktsinimidüülestrid ja 1-bensotriasoolestrid, aktiivne karbonaat nagu N- hüdrokspsuktsinimidüülkarbonaadid ja 1-bensotriasoolkarbonaadid, atsetaal-, aldehüüdrühm, aldehüüdhüdraadid, alkenüül-, akrülaat-, metakrülaat-, akrüülamiid-, aktiivne sulfoon-, amiin-, aminooksü-, kaitstud amiin-, hüdrasiid-, kaitstud hüdrasiid-, kaitstud tiool-, karboksüülhappe-, kaitstud karboksüülhappe, isotsüanaat-, isotiotsüanaat-, maleimiid-, vinüülsulfoon-, ditiopüridiin-, vinüülpüridiin-, jodoatsetamiid-, epoksiid-, glüooksaal-, dioon-, mesülaat-, tosülaat-, tresülaat-, alkeen-, ketoon- ja asiidrühmad. Asjatundjad teavad, et X fragmendi valimisel tuleb arvestada asiidrühmaga, nii et ei toimuks reaktsiooni asiidrühmaga. Asiidrühma sisaldavad polümeeri derivaadid võivad olla homobifunktsionaalsed ehk nende teine funktsionaalrühm (ehk X) on samuti asiidfragment või heterobifunktsionaalsed ehk nende teine funktsionaalrühm on teistsugune rühm. Mõiste kaitstud viitab sellele, et ühendis on kaitserühm või fragment, mis hoiab teatud reaktsioonitingimustes ära keemiliselt reaktsioonivõimelise rühma reaktsiooni. Kaitserühma olemus sõltub sellega kaitstava keemiliselt
201 reaktsioonivõimelise rühma tüübist. Näiteks kui keemiliselt reaktsioonivõimeline rühm on amino- või hüdrasiidrühm, võib kaitserühm olla tert-butüüloksükarbonüül- (t-boc) või 9- fluorenüülmetoksükarbonüülrühm (Fmoc). Kui keemiliselt reaktsioonivõimeline rühm on tioolrühm, võib kaitserühm olla ortopüridüülsulfiidrühm. Kui keemiliselt reaktsioonivõimeline rühm on karboksüülhape nagu butaan- või propioonhape või hüdroksüülrühm, võib kaitserühm olla bensüül- või alküülrühm nagu metüül-, etüül- või tert-butüülrühm. Käesoleva leiutise puhul võib kasutada ka teisi tehnika tasemes tuntud kaitserühmi. Erialakirjanduses avaldatud terminaalsete funktsionaalrühmade konkreetsete näidete seas on muu hulgas, kuid mitte ainult, N-suktsinimidüülkarbonaatrühm (vt nt USA patente nr , ), aminorühm (vt nt Buckmann et al. Makromol. Chem. 182:1379 (1981), Zalipsky et al. Eur. Polym. J. 19:1177 (1983)), hüdrasiidrühm (vt nt Andresz et al. Makromol. Chem. 179:301 (1978)), suktsinimidüülpropionaat- ja suktsinimidüülbutanoaat (vt nt Olson et al. in Poly(ethylene glycol) Chemistry & Biological Applications, lk , Harris & Zalipsky Eds., ACS, Washington, D.C., 1997; vt ka USA patenti nr ), suktsinimidüülsuktsinaatrühm (vt nt Abuchowski et al. Cancer Biochem. Biophys. 7:175 (1984) ja Joppich et al. Makromol. Chem. 180:1381 (1979), suktsinimidüülester (vt nt USA patenti nr ), bensotriasoolkarbonaatrühm (vt nt USA patenti ), glütsidüüleeterrühm (vt nt Pitha et al. Eur. J Biochem. 94:11 (1979), Elling et al., Biotech. Appl. Biochem. 13:354 (1991), oksükarbonüülimidasoolrühm (vt nt Beauchamp, et al., Anal. Biochem. 131: (1983), Tondelli et al. J. Controlled Release 1:1 (1985)), p-nitrofenüülkarbonaatrühm (vt nt Veronese, et al., Appl. Biochem. Biotech., 11: 141 (1985); ja Sartore et al., Appl. Biochem. Biotech., 27:45 (1991)), aldehüüdrühm (vt nt Harris et al. J. Polym. Sci. Chem. Ed. 22:341 (1984), USA patenti nr , USA patenti nr ), maleimiidrühm (vt nt Goodson et al. Biotechnology (NY) 8:343 (1990), Romani et al. in Chemistry of Peptides and Proteins 2:29 (1984)), and Kogan, Synthetic Comm. 22:2417 (1992)), ortopüridüül-disulfiidrühm (vt nt Woghiren, et al. Bioconj. Chem. 4:314(1993)), akrüloolrühm (vt nt Sawhney et al., Macromolecules, 26:581 (1993)), vinüülsulfoonrühm (Vt nt USA patenti nr ). Käesoleva leiutise teatud teostustes sisaldavad leiutisekohased polümeeri derivaadid polümeeri selgroogu struktuuriga: X-CH 2 CH 2 O--(CH 2 CH 2 O) n --CH 2 CH 2 -N=N=N
202 milles: 0 X on eespool kirjeldatav funktsionaalrühm; ja n on umbes kuni umbes Käesoleva leiutise teises teostuses sisaldavad leiutisekohased polümeeri derivaadid polümeeri selgroogu struktuuriga: X-CH 2 CH 2 O--(CH 2 CH 2 O) n --CH 2 CH 2 - O-(CH 2 ) m -W-N=N=N milles: W on alifaatne või aromaatne linkerfragment, mis sisaldab 1 kuni süsinikuaatomit; n on umbes kuni umbes 4000; ja X on eespool kirjeldatav funktsionaalrühm ja m on 1 kuni. Leiutisekohaseid asiidrühma sisaldavaid PEGi derivaate saab valmistada mitmesugustel tuntud ja/või käesolevas dokumendis kirjeldatavatel meetoditel. Ühe allpool kirjeldatava meetodi korral pannakse vees lahustuva polümeeri selgroog keskmise molekulmassiga umbes 800 Da kuni umbes 000 Da, mille esimene ots on seotud esimese funktsionaalrühmaga ja mille teine ots on seotud sobiva lahkuva rühmaga, reageerima asiidi aniooniga (mis võib olla paardunud mis tahes hulga sobivate vastasioonidega, sealhulgas naatrium-, kaalium, tert-butüülammoonium-, jne ioonidega). Lahkuva rühmaga toimub nukleofiilne asendamine ja see asendatakse asiidfragmendiga, mille tulemusel tekib soovitud asiidrühma sisaldav PEG polümeer. X-PEG-L + N - 3 X-PEG- N 3 Nagu on näidatud, käesoleva leiutise puhul sobiv polümeeri selgroog vastab valemile X- PEG-L, milles PEG on polü(etüleenglükool) ja X on funktsionaalrühm, mis ei reageeri asiidrühmadega, ja L on sobiv lahkuv rühm.
203 5 1 Sobivate funktsionaalrühmade seas on näiteks, kuid mitte ainult, hüdroksüül-, kaitstud hüdroksüül-, atsetaal-, alkenüül-, amino-, aminooksü-, kaitstud amino-, kaitstud hüdrasiid-, kaitstud tiool-, karboksüülhappe-, kaitstud karboksüülhappe-, maleimiid-, ditiopüridiin- ja vinüülpüridiin- ja ketoonrühmad. Sobivate lahkuvate rühmade hulka kuuluvad näiteks, kuid mitte ainult, kloriid-, bromiid-, jodiid-, mesülaat-, tresülaat- ja tosülaatrühmad. Leiutisekohaste asiidrühma sisaldavate polümeeri derivaatide teiste valmistusmeetodi korral viiakse funktsionaalse asiidrühmaga siduv aine kokkupuutesse vees lahustuva polümeeri selgrooga, mille keskmine molekulmass on umbes 800 Da kuni umbes Da, kusjuures siduval ainel on keemiliselt funktsionaalne rühm, mis reageerib selektiivselt PEG polümeeril paikneva keemiliselt funktsionaalse rühmaga, mille tulemusel moodustub asiidrühma sisaldav polümeeri derivaadi saadus, milles asiidrühma eraldab polümeeri selgroost siduv rühm. Näitlik reaktsiooniskeem on toodud allpool: X-PEG-M + N-linker-N=N=N PG-X-PEG-linker-N=N=N milles: PEG on polü(etüleenglükool) ja X on capping rühm nagu eespool kirjeldatav alkoksü- või funktsionaalrühm, ja M on funktsionaalrühm, mis ei reageeri funktsionaalse asiidrühmaga, kuid reageerib efektiivselt ja selektiivselt N funktsionaalrühmaga. Sobivate funktsionaalrühmade seas on näiteks, kuid mitte ainult, M on karboksüülhappe-, karbonaat- või aktiivne esterrühm, kui N on amiinrühm; M on ketoonrühm, kui N on hürdasiid- või aminoksüfragment; M on lahkuv rühm, kui N on nukleofiil. Toorsaadust saab vajadusel puhastada tuntud meetoditel, sealhulgas, kuid mitte ainult, saaduse sadestamise ja sellele järgneva kromatograafia abil. Üks konkreetsem näide on toodud allpool PEG diamiiniga, milles üks amiinfragment on kaitstud kaitserühma fragmendiga nagu tert-butüül-boc ja saadud mono-kaitstud PEG diamiin pannakse reageerima siduva fragmendiga, millel on funktsionaalne asiidrühm:
204 BocHN-PEG-NH 2 + HO 2 C-(CH 2 ) 3 -N=N=N 2 5 Sellisel juhul saab aminorühma siduda karboksüülhapperühmaga mitmesuguste aktiveerivate ainetega nagu tionüülkloriidi või karbodimimiidi põhised reagendid ja N- hüdroksüsuktsinimiid või N-hüdroksübensotriasool, mille tulemusel tekib monoamiini PEG derivaadi ja asiidrühma sisaldava linkerfragmendi vahele amiidside. Pärast korraliku amiidsideme moodustumist saab tekkinud N-tert-butüül-Boc-kaitstud asiidfragmenti sisaldava derivaadiga kohe modifitseerida bioaktiivseid molekule või sellele võib täienduseks lisada teisi funktsionaalrühmi. Näiteks N-t-Boc rühma hüdrolüüsimise teel tugeva happega töödeldes saab luua oomegaamino-peg-asiidi. Saadud amiinile saab sünteesimisel lisada teisi kasulikke funktsionaalrühmi nagu maleimiidrühmad, aktiveeritud disulfiidid, aktiveeritud estrid jne ja luua seeläbi väärtuslikke heterobifunktsionaalseid reagente. Heterobifunktsionaalsed derivaadid on eriti kasulikud siis, kui polümeeri mõlemasse otsa soovitakse kinnitada erinevaid molekule. Näiteks oomega-n-amino-n-asido-peg võimaldab kinnitada PEG-i ühte otsa molekuli, millel on aktiveeritud elektrofiilne rühm nagu aldehüüd-, ketoon-, aktiveeritud ester-, aktiveeritud karbonaat- või muu sarnane rühm, ja PEG-i teise otsa atsetüleenrühmaga molekuli. Leiutise teise teostuste korral on polümeeri derivaadil struktuur: X-A-POLY-B-C C-R milles: R võib olla H või alküül-, alkeen-, alkoksü- või arüül- või asendatud arüülrühm; B on siduv rühm, mis võib olla olemas või puududa; POLY on vees lahustuv mitteantigeenne polümeer; A on siduv fragment, mis võib olla olemas või puududa ja mis võib olla sama mis B või sellest erineda ning X on teine funktsionaalrühm.
205 5 3 Siduva fragmendi A ja B näidete seas on näiteks, kuid mitte ainult, mitme üksusega funktsionaliseeritud alküülrühm, milles on kuni 18 ja võib olla 1- süsinikuaatomit. Alküülrühm võib sisaldada heteroaatomit nagu lämmastiku-, hapniku- või väävliaatom. Alküülrühm võib olla heteroaatomi juures hargnenud. Siduva fragmendi A ja B teiste näidete seas on näiteks, kuid mitte ainult, mitme üksusega funktsionaliseeritud arüülrühm, milles on kuni ja võib olla 5-6 süsinikuaatomit. Arüülrühm võib olla asendatud veel ühe süsiniku-, lämmastiku-, hapniku- või väävliaatomiga. Sobivate siduvate rühmade teiste näidete seas on siduvad rühmad, mida kirjeldatakse USA patentides nr , ja USA patenditaotluse publikatsioonis 03/ Asjatundjad mõistavad, et eespool toodud siduvate fragmentide loend ei ole mitte mingil juhul täielik, vaid on illustreeriv ja et kõik siduvad fragmendid, millel on eespool kirjeldatavad omadused, sobivad käesoleva leiutise korral kasutamiseks. Üksusena X kasutamiseks sobivate funktsionaalsete rühmade seas on näiteks, kuid mitte ainult, hüdroksüül-, kaitstud hüdroksüül-, alkoksüülrühm, aktiivne ester nagu N- hüdroksüsuktsinimidüülestrid ja 1-bensotriasoolestrid, aktiivne karbonaat nagu N- hüdroksüsuktsinimidüülkarbonaadid ja 1-bensotriasoolkarbonaadid, atsetaal-, aldehüüdrühm, aldehüüdhüdraadid, alkenüül-, akrülaat-, metakrülaat-, akrüülamiid-, aktiivne sulfoon-, amino-, aminooksü-, kaitstud amino-, hüdrasiid-, kaitstud hüdrasiid-, kaitstud tiool-, karboksüülhappe-, kaitstud karboksüülhappe, isotsüanaat-, isotiotsüanaat-, maleimiid-, vinüülsulfoon-, ditiopüridiin-, vinüülpüridiin-, jodoatsetamiid-, epoksiid-, glüooksaal-, dioon-, mesülaat-, tosülaat-, tresülaat-, alkeen-, ketoon- ja atsetüleenrühmad. Asjatundjad teavad, et X fragmendi puhul tuleb lähtuda põhimõttest, et see ei tohi reageerida atsetüleenrühmaga. Atsetüleenfragmenti sisaldavad polümeeri derivaadid võivad olla homobifunktsionaalsed ehk nende teine funktsionaalrühm (ehk X) on samuti atsetüleenrühm või heterobifunktsionaalsed ehk nende teine funktsionaalrühm on teistsugune funktsionaalrühm. Leiutise teise teostuse korral sisaldavad polümeeri derivaadid polümeeri selgroogu struktuuriga: X-CH 2 CH 2 O-(CH 2 CH 2 O) n --CH 2 CH 2 - O-(CH 2 ) m -C CH milles: 30 X on eespool kirjeldatav funktsionaalrühm;
206 n on umbes kuni umbes 4000; ja 4 m on 1 kuni. Iga heterobifunktsionaalse PEG polümeeri konkreetsed näited on toodud allpool. 5 Leiutisekohaseid atsetüleenrühma sisaldavaid PEG derivaate saab valmistada mitmesugustel tuntud ja/või käesolevas dokumendis kirjeldatavatel meetoditel. Ühe allpool kirjeldatava meetodi korral pannakse vees lahustuva polümeeri selgroog keskmise molekulmassiga umbes 800 Da kuni umbes Da, mille esimene ots on seotud esimese funktsionaalrühmaga ja mille teine ots on seotud sobiva nukleofiilse rühmaga, reageerima ühendiga, millel on nii funktsionaalne atsetüleenrühm kui ka lahkuv rühm, mis sobib reaktsiooni läbiviimiseks PEG-il paikneva nuklofiilse rühmaga. Nukleofiilse rühmaga PEG-i ja lahkuva rühmaga molekuli kombineerimisel toimub lahkuva rühma nukleofiilne eemaldamine ja asendatakse nukleofiilse fragmendiga, mille tulemusel tekib soovitud atsetüleenfragmenti sisaldav polümeer. X-PEG-Nu + L-A-C X-PEG-Nu-A-C CR' Nagu on näidatud, kasutatakse sellise reaktsiooni läbiviimisel eelistatavalt polümeeri selgroogu valemiga X-PEG-Nu, milles PEG on polü(etüleenglükool), Nu on nukleofiilne fragment ja X on funktsionaalne rühm, mis ei reageeri Nu, L ega funktsionaalse atsetüülrühmaga. Nu näidete seas on muuhulgas, kuid mitte ainult, amino-, alkoksü-, arüüloksü-, sulfhüdrüül-, imino-, karboksülaat-, hüdrasiin- ja aminooksürühmad, mis reageerivad eeskätt SN2 tüüpi mehhanismi kaudu. Nu rühmade teiste näidete seas on funktsionaalrühmad, mis reageerivad eeskätt nukleofiilse liitumisreaktsiooni kaudu. L rühmade näidete seas on kloriid-, bromiid-, jodiid-, mesülaat-, tresülaat- ja tosülaatrühmad ja teised rühmad, millega eeldustekohaselt toimub nukleofiilne asendamine, ja ka ketoon-, aldehüüd., tioester-, olefiin-, alfa-beeta küllastumata karboksüülrühmad, karbonaatrühmad ja teised elektrofiilsed rühmad, millega võivad liituda nukleofiilid.
207 5 Käesoleva leiutise ühes teostuses on A alifaatne linker, milles on 1 kuni süsinikuaatomit, või asendatud arüültsükkel, milles on 6 kuni 14 süsinikuaatomit. X on funktsionaalne rühm, mis ei reageeri asiidrühmadega ja L on lahkuv rühm. 5 Leiutisekohaste atsetüleenrühma sisaldavate polümeeride teise valmistusmeetodi korral viiakse PEG polümeer keskmise molekulmassiga umbes 800 Da kuni umbes Da, mille ühes otsas on kaitstud funktsionaalrühm ja teises otsas sobiv lahkuv rühm, kokkupuutesse atsetüleenaniooniga Näitlik reaktsiooniskeem on toodud allpool: X-PEG-L + -C CR' X-PEG-C CR' milles: PEG on polü(etüleenglükool) ja X on capping rühm nagu eespool kirjeldatav alkoksürühm või funktsionaalrühm; ja R' on H, alküül-, alkoksü-, arüül- või arüüloksürühm või asendatud alküül-, alkoksü-, arüül- või arüüloksürühm. Eespool toodud näites peaks lahkuv rühm L olema piisavalt reaktsioonivõimeline, et piisavas kontsentratsioonis atsetüleenaniooniga kokkupuutesse viimisel toimuks SN2-tüüpi asendus. Lahkuvate rühmade SN2 asendamiseks atsetüleenanioonidega vajalikud reaktsioonitingumused on asjatundjatele tuntud. Toorsaadust saab üldjuhul puhastada tehnika tasemes tuntud meetoditel, sealhulgas, kuid mitte ainult, saaduse sadestamise ja vajadusel sellele järgneva kromatograafia abil. Leiutisekohaste FGF-21 polüpeptiididega saab siduda vees lahustuvad polümeerid. Vees lahustuvad polümeerid saab siduda FGF-21 polüpeptiidi sisestatud mittelooduslikult kodeeritud aminohapete kaudu või mittelooduslikult kodeeritud või looduslikult kodeeritud aminohappe mis tahes funktsionaalrühma või asendaja kaudu või mittelooduslikult kodeeritud või looduslikult kodeeritud aminohappele lisatud mis tahes funktsionaalrühma või asendaja kaudu. Vees lahustuvad polümeerid saab siduda FGF-21 polüpeptiidiga ka mittelooduslikult kodeeritud aminohappe sisestamise teel looduslikult kodeeritud aminohappe (näiteks, kuid mitte ainult, N-otsa aminohappejäägi tsüsteiin-, lüsiin- või aminorühma) kaudu. Leiutisekohased FGF-21 polüpeptiidid sisaldavad ühte
208 5 6 mittelooduslikku aminohapet, kusjuures see üks mittelooduslik aminohape on seotud vees lahustuva polümeeriga/polümeeridega (sealhulgas, kuid mitte ainult, PEG-i ja/või oligosahhariididega). Mõnel juhul sisaldavad leiutisekohased FGF-21 polüpeptiidid ka 1, 2, 3, 4, 5, 6, 7, 8, 9, või rohkemat vees lahustuvate polümeeridega seotud looduslikult kodeeritud aminohapet. Mõnel juhul sisaldavad leiutisekohased FGF-21 polüpeptiidid ühte või mitut vees lahustuvate polümeeridega seotud mittelooduslikult kodeeritud aminohapet ja ühte või mitut vees lahustuvate polümeeridega seotud looduslikult kodeeritud aminohapet. Mõnes teostuses pikendavad käesoleva leiutise puhul kasutatavad vees lahustuvad polümeerid FGF-21 polüpeptiidi poolväärtuseaega seerumis võrreldes selle konjugeerimata vormiga. Selleks et muuta (sealhulgas, kuid mitte ainult, vähendada või suurendada) FGF-21 polüpeptiidi farmakoloogilisi, farmakokineetilisi või farmakodünaamilisi omadusi nagu in vivo poolväärtusaeg, võib reguleerida leitutisekohase FGF-21 polüpeptiidiga seotavate vees lahustuvate polümeeride arvu. Mõnes teostuses pikeneb FGF-21 poolväärtusaeg võrreldes modifitseerimata polüpeptiidiga vähemalt umbes,, 30, 40, 50, 60, 70, 80, 90 protsenti, 2 korda, 5 korda, 6 korda, 7 korda, 8 korda, 9 korda, korda, 11 korda, 12 korda, 13 korda, 14 korda, korda, 16 korda, 17 korda, 18 korda, 19 korda, korda, korda, 30 korda, 35 korda, 40 korda, 50 korda või vähemalt umbes 0 korda. PEG derivaadid, mis sisaldavad tugevat nukleofiilset rühma (ehk hüdrasiid-, hüdrasiin-, hüdroksüülamiin- või semikarbasiidrühma) Käesoleva leiutise ühes teostuses modifitseeritakse karbonüülrühma sisaldavat mittelooduslikku aminohapet sisaldavat FGF-21 polüpeptiidi PEG-derivaadiga, milles on otse PEG selgrooga seotud terminaalne hüdrasiid-, hüdrasiin-, hüdroksüülamiin- või semikarbasiidrühm. Mõnes teostuses on terminaalse hüdroksüülrühmaga PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) m -O-NH 2 milles R on lihtne alküül (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00 (ehk keskmine molekulmass on 5 kuni 40 kda).
209 7 Mõnes teostuses on hüdrasiin- või hüdrasiidrühma sisaldaval PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) m -X-NH-NH 2 5 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00 ja X on valikuliselt karbonüülrühm (C=O), mis võib olla olemas või puududa. Mõnes teostuses on semikarbasiidrühma sisaldaval PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) m -NH-C(O)-NH-NH 2 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on Leiutise teises teostuses modifitseeritakse karbonüülrühma sisaldavat aminohapet sisaldavat FGF-21 polüpeptiidi PEG derivaadiga, milles on amiidsidemega PEG selgroo külge seotud terminaalne hüdroksüülamiin-, hüdrasiid-, hüdrasiin- või semikarbasiidrühm. Mõnes teostuses on terminaalset hüdroksüülamiinrühma sisaldavatel PEG derivaatidel järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 )-NH-C(O)(CH 2 ) m -O-NH 2 milles R on lihtne alküül(metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00 (ehk keskmine molekulmass on 5 kuni 40 kda). Mõnes teostuses on hüdrasiin- või hüdrasiidrühma sisaldaval PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) 2 -NH-C(O)(CH 2 ) m -X-NH-NH 2 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00 ja X on valikuliselt karbonüülrühm (C=O), mis võib olla olemas või puududa. Mõnes teostuses semikarbasiidrühma sisaldaval PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) 2 -NH-C(O)(CH 2 ) m -NH-C(O)-NH-NH 2 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00.
210 8 Leiutise ühes teostuses modifitseeritakse karbonüülrühma sisaldavat aminohapet sisaldavat FGF-21 polüpeptiidi hargnenud PEG derivaadiga, mis sisaldab terminaalset hüdrasiin-, hüdroksüülamiin-, hüdrasiid-, või semikarbasiidrühma, kusjuures hargnenud PEG-i iga ahela molekulmass on kuni 40 kda ja võib olla 5 kuni kda. 5 Leiutise ühes teostuses modifitseeritakse mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi PEG derivaadiga, millel on hargnenud struktuur. Näiteks mõnes teostuses on terminaalset hüdrasiin- või hüdrasiidrühma sisaldaval PEG derivaadil järgmine struktuur: [RO-(CH 2 CH 2 O) n -O-(CH 2 )-NH-C(O)] 2 CH(CH 2 )m-x-nh-nh 2 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00 ja X on valikuliselt karbonüülrühm (C=O), mis võib olla olemas või puududa. Mõnes teostuses on semikarbasiidrühma sisaldaval PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) 2 -C(O)-NH-CH 2 -CH 2 ] 2 CH-X-(CH 2 ) m -NH-C(O)-NH-NH 2 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), X on valikuliselt NH, O, S, C(O) või puudub, m on 2- ja n on Mõnes teostuses on terminaalset hüdroksüülamiinrühma sisaldavatel PEG derivaatidel järgmine struktuur: [RO-(CH 2 CH 2 O) n -O-(CH 2 ) 2 -C(O)-NH-CH 2 -CH 2 ] 2 CH-X-(CH 2 ) m -O-NH 2 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), X on valikuliselt NH, O, S, C(O) või puudub, m on 2- ja n on Vees lahustuva polümeeri(de) ja FGF-21 polüpeptiidi sidumise määra ja sidumiskohtade abil saab moduleerida FGF-21 polüpeptiidi võimet seonduda FGF-21 polüpeptiidi retseptoriga. Mõnes teostuses paigutatakse sidemed nii, et FGF-21 polüpeptiidi seondumisel FGF-21 polüpeptiidi retseptoriga on K d umbes 400 mm või madalam, 0 nm või madalam ja mõnel juhul 0 nm või madalam, mis on määratud seondumisanalüüsiga (equilibrium binding assay), mida kirjeldatakse seoses FGF-21 polüpeptiidiga allikas Spencer et al., J. Biol. Chem., 263: (1988).
211 Polümeeride aktiveerimismeetodeid ja sobivaid keemilisi reaktsioone ja ka peptiidide konjugeerimist kirjeldatakse erialakirjanduses ja need protseduurid on ka tehnika tasemes tuntud. Polümeeride tavapäraste aktiveerimismeetodite seas on näiteks, kuid mitte ainult, funktsionaalrühmade aktiveerimine tsüanogeenbromiidi, periodaadid, glutaraldehüüdid, biepoksiidide, epiklorohüdriini, divinüülsulfooni, karbodiimiidi, sulfonüülhalogeniidide, triklorotriasiiniga, jne (vt R. F. Taylor, (1991), PROTEIN IMMOBILIZATION. FUNDAMENTAL AND APPLICATIONS, Marcel Dekker, N.Y.; S. S. Wong, (1992), CHEMISTRY OF PROTEIN CONJUGATION AND CROSSLINKING, CRC Press, Boca Raton; G. T. Hermanson et al., (1993), IMMOBILIZED AFFINITY LIGAND TECHNIQUES, Academic Press, N.Y.; Dunn, R.L., et al., Eds. POLYMERIC DRUGS AND DRUG DELIVERY SYSTEMS, ACS Symposium Series Vol. 469, American Chemical Society, Washington, D.C. 1991). Saadaval on mitmed PEG-i funktsionaliseerimist ja konjugeerimist käsitlevad ülevaated ja monograafiad. Vt nt Harris, Macromol. Chem. Phys. C: (1985); Scouten, Methods in Enzymology 135: (1987); Wong et al., Enzyme Microb. Technol. 14: (1992); Delgado et al., Critical Reviews in Therapeutic Drug Carrier Systems 9: (1992); Zalipsky, Bioconjugate Chem. 6: (1995). Polümeeride aktiveerimismeetodeid käsitletakse ka patendidokumendis WO 94/17039, USA patendis nr , WO 94/18247, WO 94/04193, USA patendis nr , USA patendis nr , patendidokumendis WO 90/13540, USA patendis nr ja WO 93/189. Lisaks on kirjeldatud ka aktiveeritud polümeeride konjugeerimist ensüümidega, sealhulgas, kuid mitte ainult, koagulatsioonifaktoriga VIII (WO 94/6), hemoglobiiniga (WO 94/09027), hapnikku kandva molekuliga (USA patent nr ), ribonukleaasi ja superoksiidi dismutaasiga (Veronese at al., App. Biochem. Biotech. 11: (1985)). Mittelooduslikke aminohappeid nagu p-asido-l-fenüülalaniin sisaldavaid FGF-21 polüpeptiide PEGüleeritakse (ehk neile lisatakse mis tahes vees lahustuv polümeer) mis tahes käepärasel meetodil. FGF-21 polüpeptiidi näiteks PEGüleeritakse mpeg derivaadiga, millel on terminaalne alküünrühm. FGF-21 polüpeptiidi sisaldavale p-asido- L-Phe vesilahusele lisatakse toatemperatuuril segades liiaga tahket mpeg(5000)-o-ch 2 - C CH. Vesilahust üldjuhul puhverdatakse puhvriga, mille pk a on ligikaudu ph väärtusel, millel reaktsioon tuleb läbi viia (tavaliselt umbes ph 4-). Puhvrid, mis sobivad
212 PEGüleerimiseks ph 7,5 juures, on näiteks, kuid mitte ainult, HEPES, fosfaatpuhver, boraatpuhver, TRIS-HCl, EPPS ja TES. ph-d jälgitakse pidevalt ja vajadusel reguleeritakse. Reaktsioonil lastakse tavaliselt kulgeda umbes 1 kuni 48 tundi. Seejärel viiakse reaktsioonisaadustega läbi hüdrofoobse interaktsiooni kromatograafia, et eraldada PEGüleeritud FGF-21 polüpeptiidi variandid vabast mpeg(5000)-o-ch 2 -C CH-st ja PEGüleeritud FGF-21 polüpeptiidi mis tahes suure molekulmassiga kompleksidest, mis võivad moodustuda blokeerimata PEG-i molekuli mõlema otsa aktiveerimisel, mille tulemusel need ristseonduvad FGF-21 polüpeptiidi variantide molekulidega. Hüdrofoobse interaktsiooni kromatograafia läbiviimisel kasutatakse tingimusi, mille korral vaba mpeg(5000)-o-ch 2 -C CH voolab läbi kolonni ja mis tahes ristseondunud PEGüleeritud FGF-21 polüpeptiidi variantide kompleksid elueeruvad pärast soovitud vorme, mis sisaldavad ühte FGF-21 polüpeptiidi variandi molekuli, mis on konjugeeritud ühe või mitme PEG rühmaga. Sobivad tingimused on erinevad ja sõltuvad ristseotud komplekside ja soovitud konjugaatide suhtelisest suurusest ning asjatundjad oskavad selliseid tingimusi kergesti määrata. Soovitud konjugaate sisaldavat eluenti kontsentreeritakse ultrafiltreerimise teel ja soolastutatakse diafiltreerimise teel. Hüdrofoobse interaktsiooni kromatograafia abil saadud PEGüleeritud FGF-21 polüpeptiidi saab vajaduse korral veelgi puhastada ühe või mitme asjatundjatele tuntud protseduuriga, sealhulgas, kuid mitte ainult, afiinsuskromatograafia, anioon- või katioonvahetuskromatograafia (kasutades näiteks, kuid mitte ainult, toodet DEAE SEPHAROSE); ränidioksiidil põhineva kromatograafia, pöördfaasi kõrgefektiivse vedelikkromatograafia, geelfiltratsiooni (kasutades näiteks, kuid mitte ainult, toodet SEPHADEX G-75), hüdrofoobse interaktsiooni kromatograafia, suuruseralduskromatograafia, metallkelaadi kromatograafia, ultrafiltreerimise/diafiltreerimise, etanooliga sadestamise, ammooniumsulfaadiga sadestamise, kromatofokusseerimise, asenduskromatograafia (displacement chromatography), elektroforeetiliste protseduuride (sealhulgas, kuid mitte ainult, preparatiivse isoelektrilise fokusseerimise), diferentsiaalse lahustamise (sealhulgas, kuid mitte ainult, ammooniumsulfaadiga sadestamise) või ekstraheerimise teel. Näivat molekulmassi saab hinnata GPC abil globulaarsete valkude standarditega võrdlemise teel (Preneta, AZ in PROTEIN PURIFICATION METHODS, A PRACTICAL APPROACH (Harris & Angal, Eds.) IRL Press 1989, ).
213 211 FGF-21-PEG konjugaadi puhtust saab hinnata proteolüütilise lagunemise (sealhulgas, kuid mitte ainult, trüpsiiniga lõhustamise) ja sellele järgneva massispektromeetria analüüsiga. Pepinsky RB., et al., J. Pharmcol. & Exp. Ther. 297(3):59-66 (01). 5 Leiutisekohase FGF-21 polüpeptiidiga seotud vees lahustuvat polüpeptiidi saab piiranguteta täiendavalt derivatiseerida või asendada. Asiidrühma sisaldavad PEG derivaadid Leiutise teises teostuses modifitseeritakse FGF-21 polüpeptiidi PEG derivaadiga, mis sisaldab asiidfragmenti, mis reageerib mittelooduslikult kodeeritud aminohappe kõrvalahelas paikneva alküünfragmendiga. PEG derivaatide keskmine molekulmass on üldjuhul 1 kuni 0 kda ja mõnedes teostustes kuni 40 kda. Mõnes teostuses on terminaalset asiidrühma sisaldaval PEG derivaadil struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) m -N 3 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00 (ehk keskmine molekulmass on 5 kuni 40 kda). Teises teostuses on terminaalset asiidrühma sisaldaval PEG derivaadil struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) m -NH-C(O)-(CH 2 ) p -N 3 milles R on lihtne alküül(metüül-, etüül-, propüülrühm, jne), m on 2-, p on 2- ja n on 0-00 (ehk keskmine molekulmass on 5 kuni 40 kda). Leiutise ühes teostuses modifitseeritakse alküünrühma sisaldavat aminohapet sisaldavat FGF-21 polüpeptiidi terminaalset asiidrühma sisaldava hargnenud PEG derivaadiga, kusjuures hargnenud PEG-i iga ahela molekulmass on kuni 40 kda ja võib olla 5 kuni kda. Mõnes teostuses võib terminaalset asiidrühma sisaldaval PEG derivaadil olla järgmine struktuur: [RO-(CH 2 CH 2 O) n -O-(CH 2 ) 2 -NH-C(O)] 2 CH(CH 2 ) m -X-(CH 2 ) p N 3
214 212 milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2-, p on 2- ja n on 0-00 ja X on valikuliselt O, N, S või karbonüülrühm (C=O), mis võib olla olemas või puududa. 5 Alküünrühma sisaldavad PEG derivaadid Leiutise ühes teostuses modifitseeritakse FGF-21 polüpeptiidi PEG derivaadiga, mis sisaldab alküünfragmenti, mis reageerib mittelooduslikult kodeeritud aminohappe kõrvalahelas paikneva asiidfragmendiga. Mõnes teostuses on terminaalset alküünrühma sisaldaval PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) m -C CH milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2- ja n on 0-00 (ehk keskmine molekulmass on 5-40 kda). Leiutise teises teostuses modifitseeritakse alküünrühma sisaldavat mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi PEG derivaadiga, mis sisaldab amiidsideme kaudu PEG selgrooga seotud terminaalset asiid- või alküünfragmenti. Mõnes teostuses on terminaalset alküünrühma sisaldaval PEG derivaadil järgmine struktuur: RO-(CH 2 CH 2 O) n -O-(CH 2 ) m -NH-C(O)-(CH 2 ) p -C CH milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2-, p on 2- ja n on Leiutise ühes teostuses modifitseeritakse asiidrühma sisaldavat aminohapet sisaldavat FGF-21 polüpeptiidi terminaalset alküünrühma sisaldava hargnenud PEG derivaadiga, kusjuures hargnenud PEG-i iga ahela molekulmass on kuni 40 kda ja võib olla 5 kuni kda. Mõnes teostuses näiteks on terminaalset alküünrühma sisaldaval PEG derivaadil järgmine struktuur:
215 213 [RO-(CH 2 CH 2 O) n -O-(CH 2 ) 2 -NH-C(O) 2 CH(CH 2 ) m -X-(CH 2 ) p C CH milles R on lihtne alküülrühm (metüül-, etüül-, propüülrühm, jne), m on 2-, p on 2- ja n on 0-00 ja X on valikuliselt O, N, S või karbonüülrühm (C=O) või puudub. 5 Fosfiini sisaldavad PEG derivaadid Leiutise ühes teostuses modifitseeritakse FGF-21 polüpeptiidi PEG derivaadiga, mis sisaldab aktiveeritud funktsionaalrühma (sealhulgas, kuid mitte ainult, ester- või karbonaatrühma), mis sisaldab ka arüülfosfiinrühma, mis reageerib mittelooduslikult kodeeritud aminohappe kõrvalahelas paikneva asiidfragmendiga. PEG derivaatide keskmine molekulmass on üldiselt 1 kuni 0 kda ja mõnes teostuses -40 kda. Mõnes teostuses on PEG derivaadil järgmine struktuur: milles n on 1-; X võib olla O, N, S või puududa, Ph on fenüülrühm ja W on vees lahustuv polümeer. Mõnes teostuses on PEG derivaadil järgmine struktuur: milles X võib olla O, N, S või puududa, Ph on fenüülrühm, W on vees lahustuv polümeer ja R võib olla H, alküül-, arüül-, asendatud alküül- ja asendatud arüülrühm. R rühmade näidete seas on muuhulgas, kuid mitte ainult, -CH 2, -C(CH 3 ) 3, -OR', -NR'R", -SR', - halogeen, -C(O)R', -CONR'R", - S(O) 2 R', -S(O) 2 NR'R", -CN ja -NO 2. R', R", R"' ja R"" on igaüks sõltumatult vesinik, asendatud või asendamata heteroalküülrühm, asendatud või asendamata arüülrühm, sealhulgas, kuid mitte ainult, arüülrühm, mis on asendatud 1-3 halogeen-, asendatud või asendamata alküül-, alkoksü- või tioalkoksürühma või
216 5 214 arüülalküülrühmaga. Kui leiutisekohases ühendis on rohkem kui üks R rühm, valitakse iga R sõltumatult, ja kui leiutisekohases ühendis on rohkem kui üks R', R", R'" ja R"" rühm, valitakse ka need rühmad sõltumatult. Kui R' ja R" kinnituvad sama lämmastikuaatomi külge, võivad nad koos selle lämmastikuaatomiga moodustada 5-, 6- või 7-lülilise tsükli. Näiteks fragment -NR'R" võib hõlmata näiteks, kuid mitte ainult, 1-pürrolidinüül- ja 4- morfolinüülrühma. Asjatundjad mõistavad eespool käsitletavate asendajate põhjal, et mõiste alküülrühm hõlmab rühmi, mis sisaldavad süsinikuaatomeid, mis on seotud rühmadega, mis ei ole vesinikrühmad nagu haloalküül- (sealhulgas, kuid mitte ainult, -CF 3 ja -CH 2 CF 3 ) ja atsüülrühmad (sealhulgas, kuid mitte ainult, C(O)CH 3, -C(O)CF 3, - C(O)CH 2 OCH 3 ja teised sarnased rühmad).
217 2 Teised PEG derivaadid ja üldised PEGüleerimismeetodid 5 30 Teisi näiteid PEG molekulidest, mida võib siduda FGF-21 polüpeptiididega, ja PEGüleerimismeetodeid kirjeldatakse näiteks USA patendipublikatsioonis nr 04/ ; 02/00509; 03/ ; 04/ ; 03/ ; 03/02447; 03/08333; 03/ ; 03/ ; 03/05275; 03/05224; 03/ ; 02/06047; 02/ ; 02/ ; 02/ ; 02/00773; 02/ ; 02/ ; 02/ ; 02/00020; 01/ ; 01/ ; 01/ ; USA patentides nr ; ; ; ; ; ; ; ; ; ; ; ; ; ; 6 5 0; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; WO 97/32607, EP 229,8, EP 402,378, WO 92/16555, WO 94/04193, WO 94/14758, WO 94/17039, WO 94/18247, WO 94/28024, WO 95/00162, WO 95/11924, WO95/13090, WO 95/33490, WO 96/00080, WO 97/18832, WO 98/462, WO 98/48837, WO 99/32134, WO 99/32139, WO 99/32140, WO 96/40791, WO 98/32466, WO 95/06058, EP , WO 97/036, WO 96/21469, WO 95/13312, EP , WO 98/05363, EP , WO 96/41813, WO 96/07670, EP , EP 5 356, EP , EP ja EP Kõiki käesolevas dokumendis kirjeldatavaid PEG molekule võib kasutada mis tahes kujul, sealhulgas, kuid mitte ainult, ühe ahelaga, hargnenud ahelaga, mitmeõlgse ahelaga, monofunktsionaalsel, bifunktsionaalsel, multifunktsionaalsel või mis tahes nende kombinatsioonide kujul. Teisi polümeeride ja PEG derivaate kirjeldatakse järgmistes patenditaotlustes: USA patendipublikatsioon nr 06/01946, USA patendipublikatsioonis nr 06/ , USA patendipublikatsioonis nr 06/ , USA eelpatent nr 60/755,338; USA eelpatent nr 60/ ; USA eelpatent nr6 0/ ; rahvusvaheline patenditaotlus nr PCT/US06/49397; WO 06/069246; USA eelpatent nr 60/ ; USA eelpatent nr 60/ ; rahvusvaheline patenditaotlus nr PCT/US06/47822; USA eelpatent nr 60/ ; USA eelpatent nr 60/ ; ja USA eelpatent nr 60/
218 216 Afiinsuse suurendamine seerumi albumiini suhtes 5 Selleks et moduleerida FGF-21 polüpeptiidide poolväärtusaega seerumis, saab FGF-21 polüpeptiididega sulandada ka mitmesuguseid molekule. Mõnes teostuses seotakse või sulandatakse molekulid leiutisekohaste FGF21 polüpeptiididega, et suurendada looma organismis afiinsust endogeense seerumi albumiini suhtes. Näiteks mõnel juhul valmistatakse FGF-21 polüpeptiidi ja albumiini binding järjestuse rekombinantne fusioonvalk. Näitlike albumiini siduvate järjestuste seas on näiteks, kuid mitte ainult, streptokoki valgu G albumiini sidumisdomeen (vt nt Makrides et al., J. Pharmacol. Exp. Ther. 277: (1996) and Sjolander et al., J, Immunol. Methods 1:1-123 (1997)), või albumiini siduvad peptiidid nagu need, mida kirjeldatakse nt allikas Dennis, et al., J. Biol. Chem. 277: (02). Teistes teostustes atsüülitakse leiutisekohaseid FGF-21 polüpeptiide rasvhapetega. Mõnel juhul soodustavad rasvhapped seondumist seerumi albumiiniga. Vt nt Kurtzhals, et al., Biochem. J. 312:7-731 (1995). Teistes teostustes sulandatakse leiutisekohased FGF-21 polüpeptiidid otse seerumi albumiiniga (näiteks, kuid mitte ainult, inimese seerumi albumiiniga). Asjatundjad mõistavad, et leiutisekohase FGF-21 polüpeptiidiga saab siduda ka paljusid erinevaid molekule ja seeläbi moduleerida selle võimet seonduda seerumi albumiini või seerumi teiste koostisosadega. X. FGF-21 polüpeptiidide glükosüülimine Käesolev leiutis hõlmab FGF-21 polüpeptiide, mis sisaldavad ühte või mitut mittelooduslikult kodeeritud aminohapet, mis kannavad sahhariidijääke. Sahhariidijäägid võivad olla looduslikud (sealhulgas, kuid mitte ainult, N-atsetüülglükosamiin) või mittelooduslikud (sealhulgas, kuid mitte ainult 3-fluorogalaktoos). Sahhariidid võib siduda mittelooduslikult kodeeritud aminohapetega N- või O-seoselise glükosiidsideme kaudu (sealhulgas, kuid mitte ainult, N-atsetüülgalaktoos-L-seriini) või mitteloodusliku sideme (sealhulgas, kuid mitte ainult, oksiimsideme või vastava C- või S-seoselise glükosiidsideme kaudu). Sahhariidfragmente (sealhulgas, kuid mitte ainult, glükosüülfragmente saab FGF-21 polüpeptiididele lisada in vitro või in vivo. Leiutise
219 5 217 mõnes teostuses modifitseeritakse karbonüülrühma sisaldavat mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi aminooksürühmaga derivatiseeritud sahhariidiga, mille tulemusel saadakse vastav oksiimsidemega seotud glükosüülitud polüpeptiid. Mittelooduslikult kodeeritud aminohappe külge kinnitatud sahhariidi võib lisaks töödelda ka glükosüültransferaaside ja teiste ensüümidega, mille tulemusel saadakse FGF-21 polüpeptiidiga seotud oligosahhariid. Vt nt H. Liu, et al. J. Am. Chem. Soc. 1: (03). Leiutise mõnes teostuses modifitseeritakse mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi otse kindla struktuuriga glükaaniga, mis on valmistatud aminooksü-derivaadina. Asjatundjad mõistavad, et sahhariidi kinnitamiseks mittelooduslikult kodeeritud aminohappe külge saab kasutada ka teisi funktsionaalrühmi nagu asiid-, alküün-, hüdrasiid-, hüdrasiin- ja semikarbamiidrühmi. Leiutise mõnes teostuses saab asiid- või alkünüülrühma sisaldavat mittelooduslikult kodeeritud aminohapet sisaldavat FGF-21 polüpeptiidi modifitseerida näiteks, kuid mitte ainult, Huisgen [3+2] tsükloliitumise reaktsiooni teel näiteks, kuid mitte ainult, vastavalt alkünüül- või asiidderivaatidega. See meetod võimaldab valke väga selektiivselt modifitseerida. XI. FGF-21 dimeerid ja multimeerid 30 Käesoleva leiutisega pakutakse ka FGF-21 ja FGF-21 analoogide kombinatsioonid nagu homodimeerid, heterodimeerid, homomultimeerid või heteromultimeerid (ehk trimeerid, tetrameerid, jne), mille korral on ühte või mitut mittelooduslikult kodeeritud aminohapet sisaldav FGF-21 seotud otse polüpeptiidi selgroo või linkeri kaudu teise FGF-21 või selle FGF-21 variandiga või mis tahes teise polüpeptiidiga, mis ei ole FGF-21 või selle FGF-21 variant. Kuna FGF-21 dimeersetel või multimeersetel konjugaatidel on võrreldes monomeeridega suurem molekulmass, võivad neil olla uued või soovitud omadused, sealhulgas, kuid mitte ainult, võrreldes monomeerse FGF-21-ga erinevad farmakoloogilised, farmakokineetilised ja farmakodünaamilised omadused, moduleeritud terapeutiline poolväärtusaeg või moduleeritud poolväärtusaeg plasmas. Mõnes teostuses moduleerivad leiutisekohased FGF-21 dimeerid FGF-21 retseptori signaalide ülekannet. Teistes teostustes toimivad leiutisekohased FGF-21 dimeerid ja multimeerid FGF-21 retseptori antagonisti, agonisti või modulaatorina. Mõnes teostuses sisaldab FGF-i
220 sisaldavas dimeeris või multimeeris sisalduv üks või mitu FGF molekuli looduslikult mittekodeeritud aminohapet, mis on seotud vees lahustuva polümeeri külge. Mõnes teostuses seotakse FGF-21 polüpeptiidid otse ehk näiteks, kuid mitte ainult, Asn-Lys amiidsideme või Cys-Cys disulfiidsideme kaudu. Mõnes teostuses sisaldavad FGF-21 polüpeptiidid ja/või seotud mitte-fgf-21 molekul dimerisatsiooni hõlbustamiseks erinevaid looduslikult mittekodeeritud aminohappeid, sealhulgas, kuid mitte ainult, alküüni esimese FGF-21 polüpeptiidi mittelooduslikult kodeeritud aminohappes ja asiidi, mis on konjugeeritud Huisgeni [3+2] tsüklilise liitumise abil teise molekuli mittelooduslikult kodeeritud aminohappega. FGF-21 ja/või seotud mitte-fgf-21 molekuli, mis sisaldab ketoonrühma sisaldavat mittelooduslikult kodeeritud aminohapet, saab konjugeerida teise polüpeptiidiga, mis sisaldab hüdroksüülamiinrühma sisaldavat mittelooduslikult kodeeritud aminohapet ja polüpeptiidide reageerimisel tekib vastav oksiim. Kaks FGF-21 polüpeptiidi ja/või seotud mitte-fgf-21 molekuli seotakse linkeri kaudu. Kahe molekuli ja/või seotud mitte FGF-21 molekuli sidumiseks võib kasutada mis tahes hetero- või homobifunktsionaalset linkerit, millel võib olla kas sama või erinev primaarstruktuur. Mõnel juhul võib FGF-21 ja/või seotud mitte-fgf-21 molekulide kokkusidumiseks kasutatav linker olla bifunktsionaalne PEG reagent. Sellise linkeri molekulmass või selle molekuli pikkus võib jääda väga laia vahemikku. Vajadusel võib FGF-21 ja sellega seotud üksuse või FGF-21 ja selle retseptori või seotud üksuse ja selle seondumispartneri vahelise soovitud ruumilise suhte või konformatsiooni loomiseks kasutada ka suurema või väiksema molekuli pikkusega linkereid. Leiutise mõne teostusega pakutakse vees lahustuvad bifunktsionaalsed linkerid, millel on hantli struktuur, mis sisaldab: a) Polümeeri selgroo vähemalt esimeses otsas asiid-, alküün-, hüdrasiin-, hüdrasiid-, hüdroksüülamiin- või karbonüülrühma sisaldavat fragmenti ja b) polümeeri selgroo teises otsas vähemalt teist funktsionaalrühma. Nimetatud teine funktsionaalrühm võib olla sama mis esimene funktsionaalrühm või sellest erinev rühm. Mõnes teostuses ei reageeri teine funktsionaalrühm esimese funktsionaalrühmaga. Leiutise mõne teostusega pakutakse vees lahustuvad ühendid, mis sisaldavad hargnenud molekulaarse struktuuri vähemalt ühte õlga. Hargnenud molekulaarne struktuur võib olla dendriitne struktuur.
221 219 Leiutise mõne teostusega pakutakse multimeerid, mis sisaldavad ühte või mitut FGF-21 polüpeptiidi, mis on moodustatud reaktsioonide teel vees lahustuvate aktiveeritud polümeeridega, millel on järgmine struktuur: R-(CH 2 CH 2 O) n -O-(CH 2 ) m -X 5 milles n on umbes 5 kuni 3000, m on 2-, X võib olla asiid-, alküün-, hüdrasiin-, hüdrasiid-, aminooksü-, hüdroksüülamiin-, atsetüül- või karbonüülrühma sisaldav fragment ning R on capping rühm, funktsionaalrühm või lahkuv rühm, mis võib olla sama mis X või sellest erinev. R võib näiteks olla funktsionaalrühm, mis on on valitud rühmast, kuhu kuuluvad hüdroksüül-, kaitstud hüdroksüül-, alkoksüül-, N-hüdroksüsuktsinimidüülester, 1- bensotriasolüülester, N-hüdroksüsuktsinimidüülkarbonaat, 1-bensotriasolüülkarbonaat, atsetaal-, aldehüüd-, aldehüüdhüdraat-, alkenüül-, akrülaat-, metakrülaat-, akrüülamiid-, aktiivne sulfoon-, amiin-, aminooksü-, kaitstud amiin-, hüdarasiid-, kaitstud hüdrasiid-, kaitstud tiool-, karboksüülhappe-, kaitstud karboksüülhappe, isotüsanaat-, isotiotsüanaat-, maleimiid-, vinüülsulfoon-, ditiopüridiin-, vinüülpüridiin-, iodoatatsetamiid-, epoksiid-, glüooksaal-, dioon-, mesülaat-, tosülaat- ja tresülaat-, alkeen- ja ketoonrühmad. XII. FGF-21 polüpeptiidi aktiivsuse ja afiinsuse mõõtmine FGF-21 polüpeptiidi retseptori suhtes Rasvkoe ainevahetuse uurimiseks kasutatud in vitro mudelis, mida kirjeldatakse USA patendipublikatsiooni nr näites 3, on näidatud, et FGF-21 stimuleerib 3T3- L1 glükoosi imendumist rasvarakkudesse ja suurendab nende insuliinitundlikkust. 2 tüübi diabeedi üheks tunnuseks on glükoosi absorbtsiooni defitsiit erinevatesse koetüüpidesse, sealhulgas rasvkoesse. FGF-21 on seega kasulik 2. tüübi diabeedi ravis, kuna ta langetab veres glükoositaset. FGF-21 on kasulik ka rasvtõve ravis, kuna ta suurendab glükoosi kiirema ja efektiivsema kasutamise toel energiakulu. Kuna on näidatud ka seda, et FGF-21 stimuleerib insuliinist sõltumatul viisil glükoosi sisenemist 3T3-L1 rasvarakkudesse, on see kasulik ka 1. tüübi diabeedi ravis. Vt USA patendipublikatsiooni nr USA patendipublikatsioonis nr on näidatud, et FGF-21 stimuleerib
222 2 kontsentratsioonist sõltuval viisil insuliini suboptimaalse kontsentratsiooni korral (5 nm) ja insuliini puudumisel glükoosi sisenemist 3T3-L1 rasvarakkudesse. FGF-21 indutseerib glükoosi sisenemise ex vivo koemudelis, mida kirjeldatakse USA patenditaotluses nr Glükoosi sisenemist 3T3-1 rasvarakkudesse saab hinnata järgmisel meetodil. 3T3-L1 rakke on võimalik saada Ameerika Tüübikultuuri Kollektsioonist (ATCC, Rockville, Md.). Rakke kasvatatakse kasvusöötmes (GM), mis sisaldab % rauaga rikastatud veise loote seerumit Dulbecco modified Eagle söötmes. Rasvarakkude standardseks diferentseerimiseks viiakse rakud 2 päevas pärast konfluentsi saavutamist (ehk päeval 0) 48 tunniks kokkupuutesse diferentseerimissöötmega (DM), mis sisaldab % veise loote seerumit, µg/ml of insuliini, 1 µm deksametasooni ja 0,5 µm isobutüülmetüülksantiini. Rakke hoitakse seejärel diferentseerumisjärgses söötmes (post differentiation medium), mis sisaldab % veise loote seerumit ja µg/ml insuliini. In vitro mõjusust saab mõõta glükoosi sisenemise analüüsiga, mis on asjatundjatele tuntud. In vitro mõjususe saab defineerida FGF-21 ühendi mõjuna glükoosi sisenemisele rakupõhises analüüsis ja see on FGF ühendi bioloogilise mõjususe näitaja. Seda saab väljendada suurusena EC 50, mis on ühendi efektiivne kontsentratsioon, mille korral saavutatakse ühe annuse ja vastuse katses 50% aktiivsus. Glükoosi transpordi analüüs insuliinist sõltumatu--heksoosi absorptsioon, määratuna 0.1 mm 2-deoksü-D-[ 14 C]glükoosi akumuleerimise järgi, mõõdetakse järgmisel viisil: 12 süvendiga plaadile kantud 3T3-L1 adipotsüüte pestakse kaks korda KRP puhvriga (136 mm NaCl, 4,7 mm KCl, mm NaPO 4, 0,9 mm CaCl 2, 0,9 mm MgSO 4, ph 7,4), mis on soojendatud temperatuurini 37 C ja mis sisaldab 0,2% BSA-d, inkubeeritakse 2 tundi temperatuuril 37 C ümbritsevas õhus Leibovitz L- söötmes, mis sisaldab 0,2% BSA-d, pestakse veel kaks korda KRP-ga, mis sisaldab 0,2% BSA puhvrit, ja inkubeeritakse 2 tundi temperatuuril 37 C ümbritsevas õhus KRP-s, mis sisaldab 0,2 BSA puhvrit, ilma wortmanniinita (ainult Me 2 SO) ja wortmanniiniga. Seejärel lisatakse minutiks insuliini lõppkontsentratsioonini 0 nm ja viimase nelja minutit jooksul mõõdetakse 2-deoksü-D- [ 14 C]glükoosi sisenemist. Kõikidest väärtustest lahutatakse mittespetsiifiline haare, mida mõõdetakse µm tsütohalasiini B juuresolekul. Valgu kontsentratsioonid määratakse Pierce bitsinkoniinhappe (bicinchoninic acid) analüüsiga. Glükoosi haaret mõõdetakse iga katse puhul rutiinselt kolme või nelja paralleelprooviga. Antud juhul võib uurida 3T3-L1
223 rasvarakkude akuutset ja kroonilist eelravi FGF-21 ühendiga insuliini juuresolekul. Glükoosi transpordi analüüs insuliinist sõltumatu--3t3-l1 fibroblastid kantakse 96 süvendiga plaatidele ja diferentseeritakse 2 nädala jooksul rasvarakkudeks (adipotsüütideks). Pärast diferentseerimist näljutatakse rakke seerumivabas söötmes ja töödeldakse 24 tundi FGF-21-ga. Töötlemisel pestakse rakke kaks korda KRBH puhvriga, mis sisaldab 0,1% BSA-d. Glükoosi haaret uuritakse märgistatud glükoosi juuresolekul (ilma insuliinita) KPBH puhvris. USA patendipublikatsioonis nr on näidatud, et FGF-21 stimuleerib kontsentratsioonist sõltuval viisil insuliini suboptimaalse kontsentratsiooni (5 nm) ja insuliini puudumise korral glükoosi sisenemist 3T3-L1 rasvarakkudesse. Lisaks saab näidata, et leiutisekohased FGF-21 polüpeptiidid indutseerivad ex vivo koemudelis glükoosi haaret. Glükoosi transpordi ex vivo mudelit ehk glükoosi transpordi analüüsi kirjeldatakse järgmiselt: Krebs-Henseleit Buffer Stock Solutions--lähtelahus 1: NaCl (1.16 M); KCl (0.046 M); KH 2 PO 4 ( M); NaHCO 3 (0,03 M). Lähtelahus 2: CaCl 2 (0.0 M); MgSO 4 (2H 2 O) (0,0116 M). BSA: Kasutatakse toodet ICN Cohn Fraction V, rasvhappevaba BSA-d otse ilma dialüüsimiseta. Söötme valmistamine: 50 ml Krebs lähtelahust lisatakse 395 ml dh 2 O-le and gas with 95% O 2 /5% CO 2 for 1 hour. Seejärel lisatakse 50 ml lähtelahust 2 ja dh 2 O-d ruumalani 500 ml. Lisatakse 500 mg ICN rasvahappevaba BSA-s. Eelinkubeerimis- ja inkubeerimissöötmed: 32 mm mannitooli, 8 mm glükoosi. Pesulahus: 40 mm mannitooli, 2 mm püruvaati. Transpordilahus: 39 mm mannitooli, 1 mm 2-DG; 32 mm mannitooli, 8 mm 3-O-MG. Insuliinilahus: (sea insuliin [Lilly] µu/ml) lõppkontsentratsiooniga 00 µu/ml või 13,3 nm. Radioaktiivse märgise lahuse valmistamine: Spetsiifilised aktiivsused: 2DG=1,5 mci/ml; 3-0-G=437 µci/ml; või mannitool=8 µci/m. Rotte tuimestatakse nembutaaliga annuses 0,1 cc kehakaalu 0 g kohta. Rottidelt eemaldatakse lõikamise teel lihaskude, seda loputatakse 0,9% füsioloogilise soolalahusega ja asetatakse eelinkubeerimissöötmesse (2 ml), kus neid hoitakse 1 tund temperatuuril 29 C. Lihaskude asetatakse seejärel inkubeerimissöötmesse (2 ml, vastab muus osas eelinkubeerimissöötmele, kuid sisaldab insuliini või katseühendit) ja inkubeeritakse seal 30 minutit (sõltuvalt katsetingimustest). Seejärel asetatakse lihaskude pesulahusesse (2 ml), kus seda hoitakse minutit temperatuuril 29 C ja seejärel asetatakse minutiks (3-O-MG) või minutiks (2DG) märgise lahusesse (1,5 ml). Lihaskude trimmitakse, kaalutakse ja asetatakse
224 5 222 polüpropüleentuubidesse, mida hoitakse kuival jääl. Tuubidesse lisatakse 1 ml KOH-i ja tuubid asetatakse - minutiks vesivanni, mille temperatuur on 70 C, ja tuube töödeldakse iga paari minuti järel keerisvooluga. Tuube jahutatakse jääl, neisse lisatakse 1 ml 1N HCl-i ja segatakse korralikult. Seejärel pannakse 0 µl supernatanti kahe paralleelproovina stsintillatsiooniviaalidesse ja loetakse stsintillatsioonloenduril võrdluses tuntud radioaktiivsete standarditega. Kontraktsiooniks inkubeeritakse lihaseid kõigepealt 1 tund eelinkubeerimis/inkubeerimislahuses. 1 tunni pärast kinnitatakse igast paarist üks lihas (üks paar roti kohta) stimulatsiooniseadme külge ja teine lihas asetatakse teise kolbi, milles on inkubeerimislahus. Kokkutõmbunud lihast stimuleeritakse 0 msek kaupa sagedusel 70 Hz, kusjuures iga sellises jadas olev impulss kestab 0,1 msek. Stimuleerimine viiakse läbi sagedusel 1/sek pingel -V 2x minuti kaupa 1- minutilise vaheajaga. Stimuleerimise lõppedes eemaldatakse lihas stimulatsiooniseadme küljest ja asetatakse minutiks pesulahusesse ja seejärel eespool kirjeldavasse märgist sisaldavasse lahusesse. FGF retseptori saab valmistada asjatundjatele tuntud tehnikate ja meetodite abil. FGF-21 polüpeptiidi aktiivsuse saab määrata standardsete või tuntud in vitro või in vivo analüüside abil. Mittelooduslikku aminohapet sisaldava PEGüleeritud või PEGüleerimata FGF-21 polüpeptiidi korral saab FGF-21 afiinsust oma retseptori suhtes määrata biosensoriga BIAcore (Pharmacia). Sõltumata FGF-21 polüpeptiidi valmistamiseks kasutatud meetoditest saab analüüsida FGF-21 polüpeptiidide bioloogilist aktiivsust. Bioloogilise aktiivsuse katse tulemus peaks üldjuhul võimaldama analüüsida soovitud tulemust nagu bioloogilise aktiivsuse suurenemine või vähenemine (võrreldes modifitseeritud FGF-21-ga), erinev bioloogiline aktiivsus (võrreldes modifitseeritud FGF- 21-ga), retseptori või seondumispartneri suhtes avalduv afiinsus, FGF-21 või selle retseptori konformatsiooni või struktuuri muutused (võrreldes modifitseerimata FGF-21- ga) või seerumi poolväärtusaeg. Eespool toodud loend võimalikest analüüsidest ei ole täielik ja asjatundjad teavad teisi analüüse, mis võivad olla soovitud lõpptulemusel testimisel kasulikud.
225 223 XIII. Mõjususe, funktsionaalse in vivo poolväärtusaja ja farmakokineetiliste parameetrite mõõtmine 5 30 Käesoleva leiutise oluliseks aspektiks on polüpeptiidi FGF-21 pikem bioloogiline poolväärtusaeg, mis saavutatakse FGF-21 polüpeptiidi konjugeerimise teel vees lahustuva polümeeri fragmendiga. FGF-21 polüpeptiidi seerumikontsentratsiooni kiire langus pärast selle ühendi manustamist on tinginud vajaduse hinnata bioloogilisi vastuseid ravile konjugeeritud ja konjugeerimata FGF-21 polüpeptiidi ja selle variantidega. Käesoleva leiutise kohasel konjugeeritud ja konjugeerimata FGF-21 polüpeptiidil ja selle variantidel on pikem seerumi poolväärtusaeg ka näiteks pärast subkutaanset või intravenoosset manustamist, mida on võimalik mõõta näiteks ELISA meetodil või primaarse skriinimisanalüüsiga. Selleks võib kasutada kaubanduslikult kättesaadavaid ELISA või RIA komplekte. In vivo bioloogilist poolväärtusaega mõõdetakse käesolevas dokumendis kirjeldataval viisil. Mittelooduslikult kodeeritud aminohapet sisaldava FGF-21 polüpeptiidi mõjususe ja funktsionaalse in vivo poolväärtusaja saab määrata asjatundjatele tuntud protokollide järgi. Mittelooduslikult kodeeritud aminohapet sisaldava FGF-21 polüpeptiidi farmakoloogilisi parameetreid saab hinnata normaalsetel Sprague-Dawley isastel rottidel (N=5 looma igas katserühmas). Igale loomale manustatakse intravenoosselt ug või subkutaanselt 50 µg suurune annus ja neilt võetakse kindla ajalise graafiku järgi umbes 5-7 vereproovi ehk vees lahustuva polümeeriga konjugeerimata mittelooduslikku aminohapet sisaldava FGF- 21 polüpeptiidi korral umbes 6 tunni jooksul ja vees lahustuva polümeeriga konjugeeritud mittelooduslikku aminohapet sisaldava FGF-21 polüpeptiidi korral umbes 4 päeva jooksul. Mittelooduslikult koderitud aminohapet mittesisaldava FGF-21 farmakokineetilisi andmeid saab otse võrrelda mittelooduslikku aminohapet sisaldavate FGF-21 polüpeptiidide vastavate andmetega. Farmakokineetilisi parameetreid saab võrrelda ka primaatidel ehk näiteks Cynomolgus ahvidel. Tavaliselt manustatakse neile subkutaanselt või intravenoosselt üks süsteannus ja jälgitakse aja jooksul seerumi FGF-21 seerumitaset. Leiutisekohase FGF-21 polüpeptiidide spetsiifilise aktiivsuse saab määrata mitme tehnika tasemes tuntud analüüsiga. Leiutisekohaselt valmistatud ja puhastatud FGF-21 polüpeptiidide muteiinide või nende fragmentide bioloogilist aktiivsust saab testida
226 224 käesolevas dokumendis kirjeldatavatel või viidatavatel või asjatundjatele tuntud meetoditel Leiutisekohaste polüpeptiididega saab ravida imetajaid, kes põevad insuliinsõltumatut diabetes mellitus t (NIDDM: 2.tüüp), insuliinsõltumatut diabeeti (1.tüüp), ja ka rasvtõbe, ebapiisavat glükoosi kliirensit (inadequate glucose clearance), hüperglükeemiat, hüpereinsulineemiat ja teisi sarnaseid haigusi. Nagu on näidatud USA patendidokumendis nr , on FGF-21 efektiivne diabeedi ja rasvtõve loommudelites. Kuna rasvtõve ja diabeedi erinevates loommudelites esineb ainevahetusprofiilides erinevusi, on analüüsitud mitmeid mudeleid, et eristada hüperinsulineemia, hüperglükeemia ja rasvtõve toimet. Diabeeti (db/db) ja rasvtõbe (ob/ob) põdevatele hiirtele on iseloomulik raskekujuline rasvtõbi, hüperfaagia, varieeruv hüperglükeemia, insuliinitundlikkus, hüperinsulineemia ja halvenenud termogenees (Coleman, Diabetes 31:1, 1982; E. Shafrir, allikas Diabetes Mellitus; H. Rifkin and D. Porte, Jr. Eds. (Elsevier Science Publishing Co., Inc., New York, ed. 4, 1990), lk ). Diabeet on db/db mudelis oluliselt raskekujulisem (Coleman, Diabetes 31:1, 1982; E. Shafrir, in Diabetes Mellitus; H. Rifkin and D. Porte, Jr. Eds. (Elsevier Science Publishing Co., Inc., New York, ed. 4, 1990), lk ). Zucker (fa/fa) rottidel on raskekujuline rasvtõbi, hüperinsulineemia ja insuliiniresistentsus (Coleman, Diabetes 31:1, 1982; E. Shafrir, allikas Diabetes Mellitus; H. Rifkin and D. Porte, Jr. Eds. (Elsevier Science Publishing Co., Inc., New York, ed. 4, 1990), lk ) ja fa/fa mutatsioon võib olla hiire db mutatsiooni roti esinev vaste (Friedman et al., Cell 69:217-2, 1992; Truett et al., Proc. Natl. Acad. Sci. USA 88:7806, 1991). Tubby (tub/tub) hiirtele on iseloomulik rasvtõbi, mõõdukas insuliiniresistentsus ja hüperinsulineemia ilma olulise hüperglükeemiata (Coleman et al., J. Heredity 81:424, 1990). Lisaks võib uurida ka keemiliselt indutseeritud rasvtõve mononaatriumglutamaadi (MSG) mudelit, milles rasvtõbi on kergekujulisem kui geneetilistes mudelites ja tekib ilma hüperfaagia, hüperinsulineemia ja insuliiniresistentsuseta. Lõpuks võib hüperglükeemia mõju rasvtõve puudumisel uurida ka keemiliselt indutseeritud diabeedi streptosototsiini (STZ) mudeliga. STZ-ga töödeldud loomadel tekib insuliinipuudulikkus ja raskekujuline hüperglükeemia (Coleman, Diabetes 31:1, 1982; E. Shafrir, allikas Diabetes Mellitus; H. Rifkin and D. Porte, Jr. Eds. (Elsevier Science Publishing Co., Inc., New York, ed. 4, 1990), lk ).
227 2 Leiutisekohaseid FGF-21 polüpeptiide saab hinnata ob/ob hiirte in vivo septilise šoki mudelis. Vt USA patendipublikatsiooni nr XIV. Manustamine ja farmatseutilised kompositsioonid 5 Leiutisekohaseid polüpeptiide või valke (sealhulgas, kuid mitte ainult, FGF-21, süntetaase ja valke, mis sisaldavad ühte või mitut mittelooduslikku aminohapet, jne) kasutatakse valikuliselt raviotstarbeliselt, sealhulgas, kuid mitte ainult, kombinatsioonis sobiva farmatseutilise tugiainega. Sellised kompositsioonid näiteks sisaldavad terapeutiliselt aktiivses koguses kõnealust ühendit ja farmatseutiliselt sobivat tugi- või abiainet. Selliste tugi- või abiainete seas on näiteks, kuid mitte ainult, soolalahus, puhverdatud soolalahus, dekstroos, vesi, glütserool, etanool ja/või nende kombinatsioonid. Preparaat valmistatakse vastavalt selle manustamise viisile. Valkude manustamismeetodid on üldjuhul asjatundjatele teada ja neid saab rakendada ka leiutisekohaste polüpeptiidide manustamisel. Ühte või mitut leiutisekohast polüpeptiidi sisaldavat terapeutilist kompositsiooni saab valikuliselt testida ka ühes või mitmes asjakohases in vitro või in vivo haiguse loommudelis, et uurida asjatundjatele tuntud meetoditel efektiivsust ja kudede ainevahetust ning hinnata annuseid. Esialgsed annused saab määrata asjakohase analüüsi abil mittelooduslike ja looduslike aminohapete homoloogide aktiivsuse, stabiilsuse või teiste sobivate näitajate järgi (näiteks, kuid mitte ainult, modifitseerimise tulemusena ühte või kahte mittelooduslikku aminohapet sisaldava FGF-21 polüpeptiidi võrdlemise teel looduslikke aminohappeid sisaldava FGF-21 polüpeptiidiga). Kõnealuseid preparaate või manustada mis tahes viisil, mida tavaliselt kasutatakse molekuli viimiseks kokkupuutesse vere või koerakkudega. Leiutisekohased mittelooduslikke aminohappeid sisaldavad polüpeptiidid manustatakse mis tahes sobival viisil, valikuliselt koos ühe või mitme farmatseutiliselt sobiva tugiainega. Käesoleva leiutise kohaste polüpeptiidide manustamiseks sobivad meetodid on tuntud ja kuigi konkreetset kompositsiooni võib manustada rohkem kui ühel viisil, on alati mõni konkreetne manustamisviis otsesem ja efektiivsem kui mõni teine manustamisviis.
228 226 Farmatseutiliselt sobivad tugiained valitakse osaliselt konkreetse manustatava kompositsiooni järgi ja ka selle kompositsiooni manustamiseks kasutatava meetodi järgi. Käesoleva leiutise kohastest farmatseutilistest kompositsioonidest saab seega valmistada mitmesuguseid preparaate Leiutisekohaseid FGF-21 polüpeptiide saab manustada mis tahes tavapärasel valkude või peptiide manustamiseks sobival viisil, näiteks, kuid mitte ainult parenteraalselt, näiteks, kuid mitte ainult, subkutaanse või intravenoosse süstimise teel või mis tahes muul viisil tehtavate süstide või infusiooni teel. Polüpeptiidi kompositsioone saab manustada paljudel viisidel, sealhulgas, kuid mitte ainult, suu kaudu, intravenoosselt, intraperitoneaalselt, intramuskulaarselt, transdermaalselt, subkutaanselt, paikselt, sublinguaalselt või rektaalselt. Mittelooduslikke aminohappeid sisaldavaid modifitseeritud või modifitseerimata polüpeptiide võib manustada ka liposoomide abil. Sellised manustamisviisid ja sobivad preparaadid on asjatundjatele üldiselt tuntud. FGF-21 polüpeptiidi võib kasutada eraldi või kombinatsioonis teiste koostisosadega nagu farmatseutiline tugiaine. FGF-21 polüpeptiidi võib kasutada kombinatsioonis teiste ainetega, sealhulgas, kuid mitte ainult, suukaudse diabeedivastase raviainega. Diabeedivastane raviaine on mis tahes ravim, mis on kasulik mis tahes glükoosi ainevahetuse häire või selle mis tahes komplikatsiooni, sealhulgas mis tahes käesolevas dokumendis kirjeldatava seisundi, haiguse või komplikatsiooni ravis, ennetamisel või selle ägeduse vähendamisel. Diabeedivastaste raviainete seas on insuliin, tiasolidiindioonid, sulfonüüluuread, bensoehappe derivaadid, alfa-glükosidaasi derivaadid või muud sarnased raviained. Kõnealuse kompositsiooni koostisesse kuuluda võivate diabeedivastaste raviainete teiste üldkategooriate seas on (määratletud mõisted on toodud jutumärkides): ravimiartiklid (drug articles), mida tunnustatakse ametlikus Ameerika Ühendriikide Farmakopöas või ametlikus dokumendis National Formulary (või selle mis tahes lisas); uus ravim ja uus loomaravim (new animal drug), mille on heaks kiitnud USA Ravimija Toiduamet (FDA); neid mõisteid kasutatakse Ameerika Ühendriikide seadustikus alajaotises 21 (Title 21 of the United States Code); mis tahes ravim, mille puhul on vajalik USA või välisriigi valitsusüksuse heakskiit ( heakskiidetud ravim ); mis tahes ravim, mille puhul on dokumendiga 21 U.S.C. 355(a) kooskõla saavutamiseks vajalik ametlik heakskiit ( ametlikult heakskiidetud ravim ); mis aine, mille puhul taotletakse või on taotletud luba kasutamiseks humaanravimina vastavalt 21 U.S.C. 379(g)
229 ( humaanravim"). (Kõik selle mõiste puhul toodud viited seadustikule kehtivad käesoleva taotluse esitamise kuupäeva seisuga). Teisi diabeedivastaseid raviaineid kirjeldatakse käesolevas dokumendis ja need on asjatundjatele tuntud. Tehnika tasemed tuntud ravimid või diabeedivastased raviained, mida praegu kasutatakse 2. tüübi diabeedi kontrolli all hoidmisel, on näiteks järgmistesse kategooriatesse kuuluvad ained: biguaniidid, tiasolidiindioonid, sulfonüüluuread, bensoehappe derivaadid ja glükosidaasi inhibiitorid. Sellistel ravimitel on oma kindlad toimeviisid. Arvatakse, et biguaniidid, näiteks metformiin hoiavad ära liigse maksa glükoneogeneesi. Arvatakse, et tiasolidiindioonide toimemehhanism seisneb perifeerse glükoosi kõrvaldamise kiirendamises. Sulfonüüluuread, nt tolbutamiid ja glüburiis ning bensoehappe derivaadid, nt repagliniid langetavad insuliinisekretsiooni stimuleerimise teel plasmas glükoositaset. Alfaglükosidaasi inhibiitorid inhibeerivad konkureerivalt alfa-glükosidaasi, mis metaboliseerib süsivesikuid, lükates seeläbi süsivesinike imendumist edasi ja leevendab söömisjärgset hüperglükeemiat. Lisaks on pakutud välja veel terve rida diabeedi ravivõimalusi, mille kasutamist inimestel ei ole veel heaks kiidetud. Tehnika tasemes tuntud kaasaegsed ravimid või diabeedivastased ained, mida kasutatakse diabeedi ja sellele eelnevate sündroomide nagu insuliiniresistentsus kontrolli all hoidmisel, hõlmavad viit ühendiklassi: biguaniidid, nt metformiid; tiasolidiindioonid; sulfonüüluuread, nt tolbutamiid ja glüburiid; bensoehappe derivaadid, nt repagliniid; ja glükosidaasi inhibiitorid. Lisaks nimetatud ainetele võib glükoosi konrolli all hoidmiseks kasutada leiutisekohaseid FGF- 21 polüpeptiide paljude teiste ravivahenditega, sealhulgas, kuid mitte ainult, DPP-4 inhibiitoritega. Teatud sellised diabeedivastased raviained on kiidetud heaks inimestel kasutamiseks. Kliinilistes uuringutes kasutatud peamiste (lead) DPP-4 ühendite seas on vildagliptiin (Galvus) (LAF237), sitagliptiin (Januvia), saksagliptiin ja alogliptiin. USA-s kiideti 17. oktoobril 06. aastal heaks Januvia (Sitagliptin) kasutamine 2. tüübi diabeedi ravis monoteraapiana või kombineeritud teraapiana koos metformiini või tiasolidiindiooniga. Esimese põlvkonna Novartis ühendi 1-[[[2-[(5-tsüanipüridiin-2- üül)amino]etüül]amino]atsetüül]-2-tsüano-(s)- pürrolidiini (NVP DPP728) manustamisel 4-nädalase perioodi jooksul 93 2.tüübi diabeeti põdevale patsiendile (keskmine HbAlc 7,4%) langes 4-nädalase uuringuperioodi jooksul glükoosi, insuliini ja HbAlc plasmatase. Vt allikat Inhibition of Dipeptidyl Peptidase IV Improves Metabolic Control Over α 4- Week Study Period in Type 2 Diabetes. Diabetes Care. 02 May; (S) : Lisaks on tähele pandud, et metformiini saavatel patsientidel tekib pärast GLP-1 ravi alustamist
230 5 228 aditiivselt kasulik toime, mille tulemusel langeb glükoositase. Vt allikat Additive glucoselowering effects of glucagon-like peptide-1 and metformin in type 2 diabetes. Diabetes Care. 01 Apr;24(4):7-5. Uuringus, milles vaadeldi rasvunud meespatsienti, kellel ei olnud diabeeti, seostati metformiini manustamist pärast suu kaudu glükoosi manustamist tsirkuleeriva GLP-1 taseme tõusuga ja koondatud inimplasmaga läbiviidud katsetes inhibeeris metformiin oluliselt DPP-4 juuresolekul või puudumisel GLP-1(7-36)amiidi lagunemist pärast 30-minutilist inkubeerimist temperatuuril 37 C. Selle uuringu autorid pidasid võimalikuks, et metformiin inhibeerib nii in vitro kui ka in vivo GLP-1 ensümaatilist lagunemist. Vt allikat Effect of metformin on glucagon-like peptide I (GLP-1) and leptin levels in obese nondiabetic subjects. Diabetes Care. 01 Mar;24(3): Analysis of the relationship between DPP-4, and GLP-1 degradation using biochemical analyses in vitro. Demuth ja tema kolleegid, kes kasutasid inimese DPP-4 erinevaid allikaid, ei tuvastanud metformiini toimet GLP-1 DPP-4-vahendatud lagunemisele. Vt allikat Metformin Effects on Dipeptidylpeptidase IV Degradation of Glucagon-like Peptide-1. Biochem Biophys Res Commun. 02 Mar ;291(5): Diabeetiliste raviainetena kasulike biguaniididest on eriti häid tulemusi saavutatud metformiiniga. 30 Metformiin (N,N-dimetüülimidodikarbonimidikdiamiid; 1,1-dimetüülbiguaniid; N,Ndimetüülbiguaniid; N,N-dimetüüldiguaniid; N'-dimetüülguanüülguanidiin) on diabeedivastane raviaine, mis vähendab maksas glükoosi tootmist ja vähendab glükoosi imendumist soolestikust. Arvatakse, et see parandab ka mujal organismis kudede insuliinitundlikkust (suurendab perifeerset glükoosi haaret ja tarbimist). Metformiin suurendab vähenenud glükoositaluvusega (IGT) patsientidel ja 2. tüübi diabeeti põdevatel patsientidel nii söömiseelse kui ka järgse plasma glüositaseme langetamise kaudu glükoositaluvust. Metformiin ei ole tavaliselt glükoosi puudumisel efektiivne Bailey, Diabetes Care : (1992). Metformiini efektiivsust on tõestatud mitmes uuringus. Ühes mõõdukalt rasvunud 2. tüübi diabeeti põdevate katsealustega tehtud uuringus langes HbA1c tase pärast 29-ndalast ravi metformiiniga eraldi või kombinatsioonis sulfonüüluureaga 8,6%-lt 7,1%-ni
231 DeFronzo et al., New Engl. J. Med. 333: (1995). Metformiinil oli ka soodne toime seerumi lipiididele ehk see langetas söömata olekus seerumi keskmist triglütseriidide taset, kolesterooli üldtaset ja LDL kolesterooli taset, avaldamata seejuures soovimatut toimet teiste lipiidide tasemele. Teises uuringus parandas metformiin NIDDM patsientidel annusest sõltuval viisil glükeemia kontrolli. Metformiin, mida manustati 500 ja 00 mg ööpäevas, vähendas 14 päeva jooksul HbA1c taset vastavalt 0,9% ja 2,0% Garber et al., Am J. Med. 2: (1997). Metformiinil võib olla kasulik terapeutiline toime ka insuliiniresistentsusega patsientidel, kes ei põe diabeeti. Üks uuring näitas, et hüpertensiooniga mittediabeetilistel naistel metformiiniga läbiviidud ravi tulemusena langes vererõhk ja söömata oleku ja glükoos-stimuleeritud plasma insuliini fibrinogeenglükoos-stimuleeritud plasma insuliin ja fibrinogeen. Giugliano et al., Diabetes Care 16: (1993). Metformiini manustatakse tavaliselt metformiinvesinikkloriidina. Käesoleva leiutise kohaste polüpeptiidie korral võib kasutada nii sellist vormi kui ka teisi kasulikke vorme. Üldjuhul individualiseeritakse diabeedi korral hüperglükeemia kontrolli all hoidmiseks kindlat fikseeritud manustamisrežiimi metformiinvesinikkloriidi või mis tahes teise farmakoloogilise ainega. Annust individualiseeritakse nii efektiivsuse kui ka taluvuse põhjal, kuid üldiselt on maksimaalne soovitatud päevane annus 50 mg. Käesoleva leiutise puhul kasutamiseks sobivate tiasolidiindioonide seas on troglitasoon ja teised sarnased ühendid. Sellised ühendid on hästi tuntud ja neid kirjeldatakse näiteks USA patentides nr , , , , , , , , ja ; and Current Pharmaceutical Design 2:85-1 (1996). Troglitasoon on suukaudne hüperglükeemiline aine, mis stimuleerib glükoosi transporti peroksisoomi proliferaator-aktiveeritud retseptori -γ (PPARγ) aktiveerimise teel. Sellise aktiveerimise tulemusena võib troglitasoon stimuleerida GLU glükoosi transporterite ekspressiooni, mille tulemusena suureneb insuliini stimuleeritud glükoosi haare. Troglitasoon võib ka vähendada glükoneogeneesi ja/või glükolüüsi aktiveerimist. HbA1c on vereanalüüs, millega mõõdetakse glükosüülitud hemoglobiini kogust, mis on üldiselt suurem, kui patsiendil on vere glükoositase olnud mingitel perioodidel kõrgem. Selle analüüsiga saab anda hinnangu sellele, kuidas on diabeeti põdeval patsiendil
232 suudetud viimase 2-3 kuu jooksul haigust kontrolli all hoida. Kui hoida glükeemiat troglitasooniga kontrolli all, väheneb HbAlc umbes 1 kuni 2%. Mimura et al., Diabetes Med. 11: (1994); Kumar et al., Diabetologia 39: (1996). Selle toime võib avalduda alles paar nädalat pärast ravi alustamist. Ühes insuliinsõltumatut diabetes mellitus t põdevate patsientidega tehtud uuringus, milles manustati neile eksogeenset insuliini, vähenes keskmine HbAlc 0 and 600 mg suuruste troglitasooniannuste kasutamise korral vastavalt 0,8% ja 1,4%. Insuliinivajadus vähenes kuni 29% Schwartz et al., New Engl. J. Med. 338: (1998). Teises insuliinsõltumatut diabetes mellitus t põdevate patsientidega tehtud uuringus, milles katsealustele manustati 400 ja 600 mg troglitasooni, vähenes söömata olekus ja söömisjärgses olekus glükoositase ja hüperinsulineemiline euglükeemiline fiksatsioon näitas, et glükoosi kõrvaldamine ületas ravieelse taseme umbes 45% Maggs et al., Ann. Intern. Med. 128: (1998). Ühes uuringus suurendas 400 mg troglitasooni rasvunud patsientidel, kellel oli langenud või normaalne glükoositaluvus, glükoosi kõrvaldamise kiirust Nolan et al., New Eng. J. Med 331: (1994). Teises uuringus mis viidi läbi IGT-ga ja varem rasedusaegset diabeeti põdenud naistel, parandas troglitasoon annuses 600 mg insuliini homeostaasi, sealhulgas insuliinitundlikkust, ja langetas vereringes oleva insuliini kontsentratsiooni, kuid ei mõjutanud glükoositaluvust. Berkowitz et al., Diabetes 45: (1996). Tiasolidiindioone võib kasutada inimestel, kellel on risk haigestuda insuliinsõltumatusse diabetes mellitus se, nagu polütsüstiliste munasarjade sündroomi või GDM-ga naised, insuliinsõltumatut diabetes mellitus e ennetamiseks või selle tekke edasilükkamiseks. USA patent nr Kui troglitasooni kasutatakse eraldiseisvalt, on selle efektiivne kogus umbes mg kuni umbes 800 mg ööpäevas ja käesoleva leiutise korral kehtib sama vahemik. Lisaks metformiinile võib troglitasooni kasutada ka kombinatsioonis insuliini ja sulfonüüluureat sisaldava ainega. Vt nt USA patenti nr Sulfonüüluuread üldjuhul langetavad plasma glükoositaset pankreasest vabaneva insuliinikoguse suurendamise teel. Sulfonüüluuread täpsemalt blokeerivad ATP-tundlikud kaaliumikanalid. Sulfonooluurea nimega glimepiriid võib ka suurendada insuliinitundlikkust GLUT4 transporterite translokatsiooni stimuleerimise teel. Ravi sulfonüüluureatega määratakse üldjuhul siis, kui HbAlc on üle 8%. Vt ka USA patente nr ,
233 5 231 Sulfonüüluuread on ühendite klass, mis on tehnika tasemes hästi tuntud ehk neid näiteks kirjeldatakse USA patentides nr , , , , , , ja Käesoleva leiutise teatud teostuste korral sobivate sulfonüüluureate seas on näiteks (tüüpiline päevaannus on toodud sulgudes) atsetoheksamiid (umbes 0 kuni umbes 00 mg), kloorpropamiid (umbes 0 kuni umbes 500 mg), tolasamiid (umbes 0 kuni umbes 00 mg), tolbutamiid (500 kuni umbes 3000 mg), gliklasiid (umbes 80 kuni umbes 3 mg), glipisiid (umbes 5 kuni umbes 40 mg), glipisiid GITS (umbes 5 kuni umbes mg), glüburiid (umbes 1 kuni umbes mg), mikroniseeritud glüburiid (umbes 0,75 kuni umbes 12 mg), glimeperiid (umbes 1 kuni umbes 8 mg), AG-EE 623 ZW ja teised sarnased ühendid. Glimepiriid on esimene sellesse klassi kuuluv diabeedivastane raviaine, mis on kiidetud heaks kasutamiseks koos insuliiniga ja sellega kasutamisega võib kaasneda väiksem hüpoglükeemia tekke oht. Koos käesoleva leiutisega võib diabeedi ravis ja/või ennetamisel kasutada mitmesuguseid alfa-glükosidaasi inhibiitoreid. Sellised inhibiitorid inhibeerivad konkureerivalt alfaglükosidaasi, mis metaboliseerib süsivesikuid ja lükkab seeläbi edasi süsivesikute imendumist ja vähendab söömisjärgset hüperglükeemiat. Clissod et al., Drugs 35: (1988). Glükoositaseme langus võib väljenduda HbA1c taseme languses. Käesoleva leiutise korral sobivate alfa-glükosidaasi inhibiitorite seas on näiteks akarboos, miglitool ja teised sarnased ühendid. Nii akarboosi kui ka miglitooli efektiivsed annused on umbes kuni umbes 300 mg päevas. Alfa-glükosidaasi inhibiitoreid võib kasutada koos käesoleva leiutise kohaste polüpeptiididega kombinatsioonis sulfonüüluureatega. On näidatud, et alfa-glükosidaasi inhibiitorid kombinatsioonis sulfonüüluureatega vähendavad HbAlc taset üldjuhul umbes 0,5 kuni 1,0%. Lisaks on näidatud, et alfa-glükosidaasi inhibiitorid vähendavad efektiivselt söömisjärgset vere glükoositaseme tõusu Lefevre et al., Drugs 44:29-38 (1992). 30 Koos leiutisekohaste polüpeptiididega võib diabeedi ravis ja/või ennetamisel kasutada mitmesuguseid bensoehappe derivaate. Need ained ehk meglitiniidid on
234 mittesulfonüüluurea tüüpi hüpoglükeemilised ained, millel on võime stimuleerida insuliini sekretsiooni. Näiteks repagliniid seondub pankrease beeta-rakkudel ATP-tundlike kaltsiumikanalitega ja seeläbi suurendab insuliinieritust. Käesoleva leiutise korral sobivate bensoehappe derivaatide seas on näiteks repagliniid ja teised sarnased ühendid. Repagliniidi efektiivne annus võib olla umbes 0,5 mg kuni umbes 16 mg ja seda ühendit võib võtta enne igat söögikorda. Hetke uuritakse paljude ainete potentsiaalset kasutamist inimestel diabeedivastase raviainena. Kui selliseid aineid hakatakse raviotstarbeliselt kasutama, võib mis tahes sellist ainet kasutada koos käesoleva leiutise kohaste polüpeptiididega ainevahetushäirete ja eeskätt diabeedi ravis. Käesoleva leiutise ühe teostuse kohaselt võib vere glükoositaseme moduleerimiseks kasutada leiutisekohaseid FGF-21 polüpeptiide koos teise diabeedivastaste raviainete klassiga ehk karnitiinpalmitoüültransferaasi I (CPT-I) inhibiitoritega nagu etomoksiir. Etomoksiir inhibeerib pöördumatult karnitiinpalmitoüültransferaasi I, mis on vajalik rasvhapete oksüdatsiooniks. Sellise inhibeerimise tulemusena võib väheneda söömata olekus hüperglükeemia, kuna rasvhapete oksüdatsioonisaadused stimuleerivad maksas glükoneogeneesi. Etomoksiir võib suurendada 2. tüübi diabeeti põdevatel patsientidel insuliinitundlikkust Hubinger et al., Hormone Metab. Res. 24:1-18 (1992). Teiste tuntud diabeedivastaste raviainete seas on insuliinipreparaadid (nt preparaadid, mis sisaldavad loomset insuliini, mis pärineb veiste või sigade pankreasest, preparaadid, mis sisaldavad Escherichia coli või pärmi abil geneetiliselt sünteesida inimese insuliini, tsinkinsuliin, protamiin-tsink-insuliin, insuliini fragment või derivaat (nt INS-1), suukaudne insuliinipreparaat), insuliini suhtes sensibiliseerijad (nt pioglitasoon või selle sool (eelistatavalt vesinikkloriid), rosiglitasoon või selle sool (eelistatavalt maleaat), netoglitasoon, rivoglitasoon (CS-011), FK-614, ühend, mida kirjeldatakse patendidokumendis WO01/383, tesaglitasaar (AZ-242), ragaglitasaar (N,N-622), muraglitasaar (BMS ), edaglitasoon (BM-13-18), metaglidaseen (MBX-2); naveglitasaar (LY ), MX-6054, LY-5929, AMG-131(T-131), THR-0921), α- glükosaidaasi inhibiitorid (nt vogliboos, akarboos, miglitool, emiglitaat, jne), biguaniidid (nt fenformiin, metformiin, buformiin või nende sool (nt vesinikloriid, fumaraat, suktsinaat)), insuliini sekretagoogid [sulfonüüluurea (nt tolbutamiid, glibenklamiid, gliklasiid, kloorpropamiid, tolasamiid, atsetoheksamiid, glüklopüramiid, glimepiriid,
235 glipisiid, glübusool), repagliniid, nategliniid, mitigliniid või nende kaltsiumsoola hüdraat, dipeptidüülpeptidaasi IV inhibiitorid (nt vidagliptiin (LAF237), P32/98, sitagliptiin (MK- 431), P93/01, PT-0, saksagliptiin (BMS ), T-6666, TS-021),.beeta.3 agonistid (nt AJ-9677), GPR40 agonistid, glükagoonilaadsed polüpeptiidid (I) (glp 1), (glp2) või diabetogenic peptide hormoonid, GLP-1 retseptori agonistid [nt GLP-1, GLP-1MR aine, N,N-2211, AC-2993 (eksendiin-4), BIM-577, Aib(8,35)hGLP-1 (7,37)NH.sub.2, CJC- [131], amüliini agonistid (nt pramlintiid), fosfotürosiinfosfataasi inhibiitorid (nt naatriumvanadaat), glükoneogeneesi inhibiitorid (nt glükogeenfosforülaasi inhibiitorid, glükoos-6-fosfataasi inhibiitorid, glükagooni antagonistid), SGLUT (naatriumi ja glükoosi kotransporter) inhibiitorid (nt T-95), 11.beta.-hüdroksüsteroid dehüdrogenaasi inhibiitorid (nt BVT-3498), adiponektiin või selle agonistid, IKK inhibiitorid (nt AS- 2868), leptiini resistentsust parandavad ravimid, somatostatiini retseptori agonistid (ühendid, mida kirjeldatakse patendidokumentides WO01/228, WO03/424, WO98/44921, WO98/45285, WO99/2273.5, jne), glükokinaasi aktivaatorid (nt R.sup.o ), GIP (glükoosist sõltuv insulinotroopne peptiid) ja teised sarnased ühendid. Mittelooduslikke aminohappeid sisaldavatest FGF-21 polüpeptiididest võib eraldi või kombinatsioonis teiste sobivate koostisosadega valmistada ka inhaleerimise teel manustatavad aerosoolpreparaadid (ehk neid saab nebuliseerida ). Aeroolpreparaadid võib panna sobivatesse surve all olevatesse propellantidesse nagu diklorodifluorometaan, propaan, lämmastik ja teised sarnased ühendid. Preparaadid, mis sobivad parenteraalseks manustamiseks ehk näiteks intraartikulaarseks (liigestesse), intravenoosseks, intramuskulaarseks, intradermaalseks, intraperitoneaalseks ja subkutaanseks manustamiseks, on näiteks veepõhised ja mitteveepõhised isotoonilised steriilsed süstelahused, mis võivad sisaldada antioksüdante, puhvreid, bakteriostaate ja soluute, mis muudavad preparaadi saaja verega isotooniliseks, ja veepõhised ja mitteveepõhised steriilsed suspensioonid, mis võivad sisaldada suspendeerivaid aineid, lahustumist parandavaid aineid, paksendajaid, stabilisaatoreid ja säilitusaineid. FGF-21 prepraadid võivad olla ühte annust või mitut annust sisaldavatest suletud pakendites/mahutites nagu ampullid ja viaalid. Eelistatud manustamisviisideks on parenteraalne ja intravenoosne manustamine. Leiutisekohaste polüpeptiide preparaatide manustamiseks sobivad eelkõige
236 234 manustamisviisid, mida juba kasutatakse looduslike aminohapete homolooge sisaldavate ravimite (näiteks, kuid mitte ainult, EPO, GH, G-CSF, GM-CSF, IFN-ide, interleukiinide, antikehade, FGF-de ja/või mis tahes teiste farmatseutiliselt manustatavate valkude) ja praegu kasutatavate preparaatide manustamiseks. 5 Käesoleva leiutise korral peab patsiendile manustatav annus olema piisav patsiendi organismis aja jooksul kasuliku ravivastuse või muu sobiva toime tekitamiseks või muu sobiva, rakendusalast sõltuva toime avaldamiseks. Annus määratakse konkreetse vektori või preparaadi efektiivsuse järgi, kasutatava mittelooduslikke aminohappeid sisaldava polüpeptiidi stabiilsuse või seerumi poolväärtusaja järgi ning patsiendi seisundi ja ka kehakaalu või kehapindala järgi. Annuse suuruse valimisel lähtutakse ka konkreetsel patsiendil konkreetse vektori, preparaadi või muu sarnase manustamisega kaasnevate mis tahes kõrvaltoimete olemasolust, olemusest ja ulatusest. Selleks et määrata haiguse (näiteks, kuid mitte ainult, vähktõve, pärilike haiguste, diabeedi, AIDS-i või teiste sarnaste haiguste) ravis või profülaktikas manustatava vektori või preparaadi efektiivset kogust, hindab ühendi tsirkuleeriva plasma taset, preparaadi toksilisust, haiguse progresseerumist ja/või vajadusel ka mittelooduslikke aminohappeid sisaldavate polüpeptiidide vastaste antikehade tootmist. Näiteks 70 kg kaaluvale patsiendile manustatav annus on tavaliselt vahemikus, mida kasutatakse kaasaegsete raviotstarbeliste valkude manustamisel ja mida on reguleeritud vastavalt asjakohase kompositsiooni muutunud aktiivsusele või seerumi poolväärtusajale. Selle leiutise kohaseid vektoreid või ravipreparaate saab kasutada täiendusena mis tahes tuntud tavapärasele ravile, sealhulgas antikehade manustamisele, vaktsiini manustamisele, tsütotoksiliste ainete manustamisele, looduslikke aminohappeid sisaldavatele polüpeptiididele, nukleiinhapetele, nukleotiidide analoogidele, bioloogilise vastuse modifitseerijatele ja teistele sarnastele ravivahenditele. Leiutisekohaste preparaatide manustamise kiiruse saab määrata vastava preparaadi LD-50 või ED-50 järgi ja/või mittelooduslikke aminohappeid sisaldavate polüpeptiidide erinevate kontsentratsioonide korral täheldatud mis tahes kõrvaltoimete järgi ning lisaks ka näiteks, kuid mitte ainult, patsiendi kehamassi ja üldise tervisliku seisundi järgi. 30 Leiutisekohaseid ühendeid või manustada üksikannuste või osaannustena.
237 Kui preparaadi infusiooni saaval patsiendil tekib palavik, külmavärinad või lihasvalu, manustatakse talle sobivas annuses aspiriini, ibuprofeeni, atsetanofeeni või teist valuvaigistit või palavikuvastast ravimit. Patsientidele, kellel kaasnevad infusiooniga sümptomid nagu palavik, lihasvalu ja külmavärinad, manustatakse 30 minutit enne infusiooni algust aspiriini, atsetaminofeeni või näiteks, kuid mitte ainult, difenhüdramiidi. Meperidiini tugevamate külmavärinate ja lihasvalude puhul, mis ei reageeri kiiresti ravile antipüreetikumide ja antihistamiinidega. Infusiooni rakkudesse (cell infusion) aeglustatakse või see katkestatakse sõltuvalt reaktsiooni ägedusest. Leiutisekohased inimese FGF-21 polüpeptiidid võib manustada otse imetajast patsiendile. FGF-21 polüpeptiide võib manustada mis tahes viisil, mida tavaliselt kasutakse patsiendile FGF-21 polüpeptiidi manustamiseks. Käesoleva leiutise teostuste kohaste FGF-21 polüpeptiidi kompositsioonide seas on kompositsioonid, mis sobivad suu kaudu, rektaalseks, paikseks, inhaleerimise teel (sealhulgas, kuid mitte ainult, aerosoolina), bukaalseks (sealhulgas, kuid mitte ainult, sublinguaalseks), vaginaalseks, parenteraalseks (sealhulgas, kuid mitte ainult, subkutaanseks, intramuskulaarseks, intradermaalseks, intraartikulaarseks, intrapleuraalseks, intraperitoneaalseks, intratserebraalseks, intraarteriaalseks või intravenoosseks), paikseks (nii naha- kui limaskestadele pinnale, sealhulgas hingamisteede pinnale) ja transdermaalseks manustamiseks, kuigi sobivaim manustamisviis sõltub ravitava seisundi olemusest ja ägedusest. Manustamine võib olla lokaalne või süsteemne. Ühendite preparaadid võivad olla ühte või mitut annust sisaldavates suletud mahutites nagu ampullid või viaalid. Leiutisekohaseid FGF-21 polüpeptiide võib valmistada ühe süsteannuse kujul (näiteks, kuid mitte ainult, lahuse, suspensiooni või emulsiooni kujul) oleva seguna koos farmatseutiliselt sobiva tugiainega. Leiutisekohaseid FGF-21 polüpeptiide võib manustada ka pideva infusioona (kasutades selleks näiteks, kuid mitte ainult, minipumpasid nagu osmootsed pumbad, ühe booluse või toimeainet aeglaselt vabastavate depoopreparaatidena. Manustamiseks sobivate preparaatide seas on vesi- või mitteveepõhised lahused, isotoonilised steriilsed lahused, mis sisaldavad antioksüdante, puhvreid, bakteriostaate ja soluute, mis muudavad preparaadi isotooniliseks, ja vee- ja mittepõhised steriilsed suspensioonid, mis võivad sisaldada suspendeerivaid aineid, lahustuvuse parandajaid,
238 236 paksendajaid, stabilisaatoreid ja säilitusaineid. Lahused või suspensioonid võib valmistada steriilsetest pulbritest, graanulitest ja tablettidest, mida kirjeldatakse eespool Valgupreparaatide puhul kasutatakse sageli külmkuivatamist, mille korral eemaldatakse huvipakkuvast preparaadist vesi. Külmkuivatamine või lüofiliseerimine on protsess, milles kuivatatav materjal kõigepealt külmutatakse ja seejärel eemaldatakse vaakumis sublimeerimise teel jää või külmunud lahusti. Eellüofiliseeritud preparaatidele võib lisada abiainet, mis muudab need külmkuivatusprotsessi ajal stabiilsemaks ja/või muudab säilitatava lüofiliseeritud saaduse stabiilsemaks. Pikal, M. Biopharm. 3(9)26-30 (1990) and Arakawa et al. Pharm. Res. 8(3): (1991). Farmaatsiatoodete pihustuskuivatamine on samuti asjatundjatele tuntud. Vt nt Broadhead, J. et al., The Spray Drying of Pharmaceuticals, in Drug Dev. Ind. Pharm, 18 (11 & 12), (1992). Lisaks väikesemolekulilistele farmaatsiatoodetele on pihustuskuivatatud mitmesuguseid bioloogilisi materjale, mille seas on: ensüümid, seerum, plasma, mikroorganismid ja pärmid. Pihustuskuivatamine on kasulik tehnika, kuna see abil saab vedela ravipreparaadi muundada ühe etapiga peentest osakestest koosnevaks tolmuvabaks või aglomereerunud pulbriks. Kasutatav põhimeetod koosneb järgmisest neljast etapist: a) lähtelahuse atomiseerimine pihustusvedelikuks (spreiks); b) pihustusvedeliku viimine kokkupuutesse õhuga ja d) kuivatatud saaduse eraldamine kuivatusõhust. USA patentides nr ja kirjeldatakse rekombinantse erütropoietiini valmistamist pihustuskuivatamise teel. Leiutisekohased farmatseutilised kompositsioonid ja -preparaadid võivad sisaldada farmatseutiliselt sobivat tugiainet, abiainet või stabilisaatorit. Farmatseutiliselt sobivad kandjad määrab osaliselt konkreetne manustatav kompositsioon, kuid ka kompositsiooni manustamiseks kasutatav konkreetne meetod. Käesoleva leiutise farmatseutiliste kompositsioonide jaoks on kasutatavad mitmesugused sobivad preparaadid (mis sisaldavad vajadusel kasutatavaid farmatseutiliselt vastuvõetavaid kandjaid, abiaineid või stabilisaatoreid (vt nt Remington's Pharmaceutical Sciences, 17. tr, 1985)). Sobivate tugiainete hulka kuuluvad, kuid mitte ainult, puhvrid, mis sisaldavad suktsinaati, fosfaati, boraati, HEPESit, tsitraati, histidiini, imidasooli, atsetaati, bikarbonaati ja teisi orgaanilisi happeid; antioksüdandid, sealhulgas, kuid mitte ainult, askorbiinhape, väikese molekulmassiga polüpeptiidid, sealhulgas, kuid mitte ainul, need, mis sisaldavad vähem
239 kui ligikaudu aminohappejääki; valgud, sealhulgas, kuid mitte ainult, seerumi albumiin, želatiin või immunoglobuliinid; hüdrofiilsed polümeerid, sealhulgas, kuid mitte ainult, polüvinüülpürrolidoon; aminohapped, sealhulgas, kuid mitte ainult, glütsiin, glutamiin, asparaganiin, arginiin, histidiin või histidiini derivaadid, metioniin, glutamaat või lüsiin; monosahhariidid, disahhariidid ja teised süsivesikud, sealhulgas, kuid mitte ainult, trehaloos, sahharoos, glükoos, mannoos või dekstriinid; kelaativad ained, sealhulgas, kuid mitte ainult, EDTA ja edentaatdinaatrium; kahevalentsed metalliioonid, sealhulgas, kuid mitte ainult, tsink, koobalt või vask; suhkruralkoholid, sealhulgas, kuid mitte ainult, mannitool või sorbitool; soola moodustavad vastasioonid, sealhulgas, kuid mitte ainult, naatrium ja naatriumkloriid; ja/või mitteioonsed pindaktiivsed ained, sealhulgas, kuid mitte ainult, Tween (sealhulgas, kuid mitte ainult, Tween 80 (polüsorbaat 80) ja Tween (polüsorbaat ), Pluronics ja teised pluroonhapped, sealhulgas, kuid mitte ainult, pluroonhape F68 (poloksameer 188) või PEG. Sobivate pindaktiivsete ainete hulka kuuluvad näiteks, kuid mitte ainult, polüeetrid, mis põhinevad polü(etüleenoksiid)- polü(propüleenoksiid)-polü(etüleenoksiidil), näiteks (PEO-PPO-PEO) või polü(propüleenoksiid)-polü(etüleenoksiid)-polü(propüleenoksiidil), näiteks (PPO-PEO- PPO) või nende kombinatsioonil. PEO-PPO-PEO ja PPO-PEO-PPO on kaubanduslikult kättesaadavad kaubamärke Pluronics, R-Pluronics, Tetronics ja R-Tetronics kandvate toodetena (BASF Wyandotte Corp., Wyandotte, Michigan, Ameerika Ühendriigid) ja neid on täiendavalt kirjeldatud USA patendis nr Ka teised etüleeni-/polüpropüleeni blokkpolümeerid võivad olla sobivad pindaktiivseid ained. Pindaktiivset ainet või pindaktiivsete ainete kombinatsioone võib kasutada PEGüleeritud FGF-21 stabiliseerimiseks ühe või mitme välismõju vastu, mis ei ole piiratud liigutusest tingitud stressiga. Mõnda ülalnimetatud ainet võib nimetada ka täiteaineks. Mõnda ülalnimetatud ainet võib nimetada ka toonuse modifikaatoriks. Toote stabiilsuse ja antimikroobse efektiivsuse jaoks võib kasutada ka antimikroobseid säilitusaineid; sobivate säilitusainete hulka kuuluvad, kuid mitte ainult, bensüülalkohol, bensalkooniumkloriid, metakresool, metüül-/propüülparabeen, kresool ja fenool või nende kombinatsioonid. Leiutisekohaseid vesilahustuvate polümeeridega, näiteks PEG, ühendatud FGF-21 polüpeptiide võib manustada ka osana toimeainet püsivalt vabastavatest süsteemidest. Toimeainet püsivalt vabastavate kompositsioonide hulka kuuluvad, kuid mitte ainult, semipermeaabelsed polümeermaatriksid vormitud artiklite kujul, sealhulgas, kuid mitte
240 ainult, õhukesed polümeerikiled või mikrokapslid. Toimeainet püsivalt vabastavate maatriksite hulka kuuluvad bioühilduvad materjalid, näiteks polü(2- hüdroksüetüül)metakrülaat (Langer et al., J. Biomed Mater. Res., : (1981); Langer, Chem. Tech., 12: 98-5 (1982), etüleenvinüülatsetaat (Langer et al., supra) või polü-d-(-)-3-hüroksüvõihape (EP ), polülaktiidid (polüpiimhape) (USA patent nr ; EP ), polüglükoliid (glükoolhappe polümeer), polülaktiidkoglükoliid (piimhappe ja glükoolhappe kopolümeerid), polüanhüdriidid, L-glutaamhappe ja gammaetüül-l-glutarnaadi kopolümeerid (Sidman et al., Biopolymers, 22, (1983), polü(orto)estrid, polüpeptiidid, hüaluroonhape, kollageen, kondroitiinsulfaat, karboksüülhapped, rasvhapped, fosfolipiidid, polüsahhariidid, nukleiinhapped, polüaminohapped, aminohapped, näiteks fenüülalaniin, türosiin, isoleutsiin, polünukleotiidid, polüvinüülpropüleen, polüvinüülpürrolidoon ja silikoon. Toimeainelt püsivalt vabastavate kompositsioonide hulka kuulub ka liposoomidega ümbritsetud ühend. Ühendit sisaldavaid liposoome saab valmistada tuntud meetoditel: DE ; Eppstein et al., Proc. Natl. Acad Sci. U.S.A., 82: (1985); Hwang et al., Proc. Natl. Acad Sci. U.S.A., 77: (1980); EP ; EP ; USA patent nr ; EP ; USA patent nr ; Jaapani patent ; USA patendid nr ja ; ja EP Liposoomidesse sisestatud FGF-21 polüpeptiide saab valmistada meetoditel, mida kirjeldatakse järgmistes allikates, näiteks DE ; Eppstein et al., Proc. Natl. Acad Sci. U.S.A., 82: (1985); Hwang et al., Proc. Natl. Acad Sci. U.S.A., 77: (1980); EP ; EP ; USA patent nr ; EP ; USA patent nr ; Jaapani patent ; USA patendid ja ; ja EP Kompositsioonid ja liposoomide suurus on tehnika tasemes hästi tuntud või on need erialaasjatundja poolt empiiriliselt kergesti määratavad. Mõnesid näiteid liposoomide kohta kirjeldatakse näiteks allikas Park, J. W., et al., Proc. Natl. Acad. Sci. USA 92: (1995); Lasic, D, ja Papahadjopoulos, D. (toim-d): MEDICAL APPLICATIONS OF LIPOSOMES (1998); Drummond, D. C., et al., Liposomal drug delivery systems for cancer therapy, kogumikus: Teicher, B. (toim): CANCER DRUG DISCOVERY AND DEVELOPMENT (02); Park, J. W., et al., Clin. Cancer Res. 8: (02); Nielsen, U. B., et al., Biochim. Biophys. Acta 91(1-3):9-118 (02); Mamot, C., et al., Cancer Res. 63: (03).
241 Patsiendile käesoleva leiutise kontekstis manustatav annus peab olema piisav aja jooksul patsiendi organismis soodsa reaktsiooni tekitamiseks. Üldiselt on käesoleva leiutise kohase FGF-21 polüpeptiidi farmatseutiliselt efektiivne üldkogus parenteraalsel manustamisel annuse kohta vahemikus ligikaudu 0,01 µg/kg/ööpäevas kuni ligikaudu 0 µg/kg või ligikaudu 0,05 mg/kg kuni ligikaudu 1 mg/kg patsiendi kehamassi kohta, kuigi see sõltub konkreetsest raviotsusest. Annustamissagedus sõltub konkreetsest raviotsusest ja võib olla sagedam või harvem kui kaubanduslikult kättesaadavate inimkasutuseks heaks kiidetud FGF-21 polüpeptiidtoodete korral. Üldiselt võib leiutisekohaste pegüleeritud FGF-21 polüpeptiidi manustada kõigi ülakirjeldatud manustamisteede kaudu. Mõnes teostuses moduleerivad käesoleva leiutise kohased modifitseerivad FGF-21 polüpeptiidid diabeedivastase raviaine toimet. Käesoleva leiutise teises teostuses võib modifitseeritud FGF-21 polüpeptiide manustada koos diabeedivastase raviainega. Veel ühes käesoleva leiutise teostuses võib modifitseeritud FGF-21 polüpeptiide manustada enne ravi diabeedivastase raviainega. Veel ühes käesoleva leiutise teostuses võib modifitseeritud FGF-21 polüpeptiide manustada pärast ravi diabeedivastase ainega. Käesoleva leiutise veel ühes teostuses manustatakse modifitseeritud FGF-21 polüpeptiide koos metformiiniga. Käesoleva leiutise veel ühes teostuses suurendab ravi koos leiutisekohaste modifitseeritud FGF-21 polüpeptiididega ja metformiiniga metformiini võimet mõjutada plasma glükoosisisaldust kas insuliini juuresolekul või selle puudumisel. Kombineeritud ravis on metformiini kasutatud koos sulfonüüluurea preparaatidega, alfaglükosidaasi inhibiitoritega, troglitaseooniga ja insuliiniga. Metformiini ja sulfonüüluurea preparaadi kombinatsioon suurendab insuliinitundlikkust ja võib vähendada plasma glükoosisisaldust. Teise võimalusena võib metformiin koos repagliniidiga olla mitmete kuude jooksul glükeemilise kontrolli säilitamisel tõhusam kui glipisiid ja vähemalt sama efektiivne nagu gliburiid. Metformiin koos troglitasooniga parandab glükoosikontrolli paremini kui emb-kumb aine üksi. Inzucchi et al., New. Eng. J. Med. 338: (1998). Leiutise mõnes teostuses manustatakse käesoleva leiutise kohaseid FGF-21 polüpeptiide koos Klotho-betaga. Leiutise mõnes teostuses manustatakse käesoleva leiutise FGF-21 polüpeptiide koos Klotho-betaga, mis hõlmab ühte või mitut mittelooduslikult kodeeritud aminohapet. Leiutise mõnes teostuses manustatakse käesoleva leiutise kohaseid FGF-21 polüpeptiide koos Klotho-beta ja diabeedivastase ainega. Leiutise mõnes teostuses manustatakse käesoleva leiutise kohaseid FGF-21 polüpeptiide koos diabeedivastase
242 ainega. Leiutise mõnes teostuses kasutatakse käesoleva leiutise kohaseid FGF-21 polüpeptiide koos ühe või mitme järgneva ühendiga: tauriin, alfa-lipoehape, mooruspuuekstrakt, kroom, glutamiin, Enicostemma littorale õied, Scoparia dulcis, estragoni ja india kibelille ekstrakt. Leiutise mõnes teostuses kasutatakse käesoleva leiutise kohaseid FGF-21 polüpeptiide koos ühe või mitme järgneva ühendiga: insuliini preparaadid (nt loominsuliini preparaadid, mis on ekstraheeritud veise ja sea pankreasest; iniminsuliini preparaadid, mis on geneetiliselt sünteesitud Escherichia coli, pärmseene abil; tsinkinsuliin; protamiintsinkinsuliin; insuliinifragment või -derivaat (nt INS-1), suukaudne insuliinipreparaat), insuliini suhtes sensitiseerijad (nt pioglitasoon või selle sool (eelistatavalt hüdrokloriid), rosiglitasoon või selle sool (eelistatavalt maleaat), netoglitasoon, rivoglitasoon (CS-011), FK-614, patendis WO01/383 kirjeldatud ühend, tesaglitasaar (AZ-242), ragaglitasaar (N,N-622), muraglitasaar (BMS ), edaglitasoon (BM-13-18), metaglidaseen (MBX-2), naveglitasaar (LY ), MX- 6054, LY-5929, AMG-131(T-131), THR-0921), α-glükosidaasi inhibiitorid (nt vogliboos, akarboos, miglitool, emiglitaat jne), biguaniidid (nt fenformiin, metformiin, buformiin ja nende soolad (nt hüdrokloriidid, fumaraadid, suktsinaadid)), insuliini sekretagoogid [sulfonüüluurea preparaadid (nt tolbutamiid, glibenklamiid, gliklasiid, kloorpropamiid, tolasamiid, atseetoheksamiid, glüklopüramiid, glimepiriid, glipisiid, glibusool), repagliniid, nategliniid, mitiglineed või nende kaltsiumsoolade hüdraadid], dipeptidüülpeptidaas IV inhibiitorid (nt vidagliptiin (LAF237), P32/98, sitagliptiin (MK- 431), P93/01, PT-0, saksagliptiin (BMS ), T-6666, TS-021), beeta-3-agonistid (nt AJ-9677), GPR40 agonistid, glükagoonisarnased polüpeptiidid (I) (glp I), (glp2) või teised diabetogeensed peptiidhormoonid, GLP-1 retseptori agonistid [nt GLP-1, GLP- 1MR aine, N,N-2211, AC-2993 (eksendiin-4), BIM-577, Aib(8,35)hGLP-1 (7,37)NH.sub.2, CJC-[131], amüliini agonistid (nt pramlintiid), fosfotürosiini fosfataasi inhibiitorid (nt naatriumvanadaat), glükoneogeneesi inhibiitorid (nt glükogeeni fosforülaasi inhibiitorid, glükoos-6-fosfataasi inhibiitorid, glükagooni antagonistid), SGLUT (naatriumi-glükoosi kotransporter) inhibiitorid (nt T-95), 11- beetahüdroksüsteroidi dehüdrogenaasi inhibiitorid (nt BVT-3498), adiponektiin või selle agonistid, IKK inhibiitorid (nt AS-2868), leptiiniresistentsust parandavad ravimid, somatostatiini retseptori agonistid (ühendeid kirjeldatakse patentides WO01/228, WO03/424, WO98/44921, WO98/45285, WO99/ jne), glükokinaasi aktivaatorid (nt R.sup.o ), GIP (glükoossõltuv insulinotroopne peptiid).
243 Leiutise mõnes teostuses kasutatakse käesoleva leiutise polüpeptiide kombinatsioonis insuliini toime tugevdajatega, näiteks, kuid mitte ainult, tauriini, alfa-lipoehappe, mooruspuuekstrakti, kroomi, glutamiini, Enicostemma littorale õite, Scoparia dulcis'e, estragoni ja india kibelille ekstraktiga. Teises teostuses võib käesolev kirjeldus sisaldada ühte või mitut järgnevat komponenti: isomalt, trehaloos või D-mannoos, et veelgi tugevdada insuliini sekretsiooni või aktiivsust. Leiutise täiendavas teostuses kasutatakse insuliini toime tugevdajat ja käesoleva leiutise polüpeptiide lisaks muul diabeedivastasele raviravimile. Üks võimalus polüpeptiidide ja kombineeritud ravimeetodite, sealhulgas käesoleva leiutise kohaste polüpeptiidide, terapeutilise efektiivsuse määramiseks toimub patsiendi HbA1c sisalduse vähendamise kaudu. Ühes teostuses vähendavad käesoleva leiutise kohased polüpeptiidid HbA1c sisaldust vähemalt 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, %, 11 %, 12%, 13%, 14%, %, 16%, 17%, 18%, 19%, %, %, 30%, 35%, 40%, 45%, 50% või vähemalt 50% võrra võrreldes HbA1c sisaldusega kaks kuud enne ravi alustamist modifitseeritud FGF-21 polüpeptiididega, võrreldes HbA1c sisaldusega kolm kuud enne ravi alustamist modifitseeritud FGF-21 polüpeptiididega või algväärtustega võrreldes. Teises teostuses moduleerivad käesoleva leiutise kohased polüpeptiidid, mida manustatakse diabeedivastase ainega ravitavale patsiendile, diabeedivastase aine võimet vähendada vere glükoosisisaldust. Teises teostuses vähendavad diabeedivastast ainet saavale patsiendile manustatavad käesoleva leiutise kohased polüpeptiidid HbA1c sisaldust vähemalt 1 %, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, %, 11 %, 12%, 13%, 14%, %, 16%, 17%, 18%, 19%, %, %, 30%, 35%, 40%, 45%, 50% või vähemalt 50% võrra võrreldes HbA1c sisaldusega kaks kuud enne ravi alustamist modifitseeritud FGF- 21 polüpeptiididega, võrreldes HbA1c sisaldusega kolm kuud enne ravi alustamist modifitseeritud FGF-21 polüpeptiididega või algväärtustega võrreldes või ravieelse algväärtusega võrreldes. Teises teostuses moduleerivad käesoleva leiutise kohased modifitseeritud FGF-21 polüpeptiidid troglitasooni võimet vähendada insuliinivajadust. Veel ühes teostuses, täiendavas teostuses, vähendavad käesoleva leiutise kohased modifitseeritud FGF-21 polüpeptiidid manustatuna troglitasoooniga ravitavale patsiendile veelgi nimetatud patsiendi insuliinivajadust. Käesoleva leiutise teatud teostuses võib troglitasooni ja
244 242 käesoleva leiutise kombinatsiooni kasutada 2. tüüpi suhkurtõve tekke edasilükkamiseks või ennetamiseks. 5 Käesoleva leiutise ühes teostuses manustatakse modifitseeritud FGF-21 polüpeptiide koos sulfonüüluurea preparaadiga. Käesoleva leiutise veel ühes teostuses manustatakse modifitseeritud FGF-21 polüpeptiide enne ravi sulfonüüluurea preparaadiga. Käesoleva leiutise veel ühes teostuses manustatakse modifitseeritud FGF-21 polüpeptiide pärast ravi sulfonüüluurea preparaadiga. Käesoleva leiutise mõnes teostuses moduleerib ravi modifitseeritud FGF-21 polüpeptiidi terapeutilise annusega seerumi glükoosisisaldust. Leiutise veel ühes teostuses manustatakse käesoleva leiutise FGF-21 polüpeptiide koos Klotho-betaga, mis moduleerib peptiidide toimet vere glükoosisisaldusele. Veel ühes teostuses käesoleva leiutise kirjelduse kohaseid FGF-21 polüpeptiide manustatakse koos Klotho-betaga, mis vähendab vere glükoosisisaldust rohkem kui ainult modifitseeritud FGF-21 polüpeptiidide kasutamine. Leiutise teises teostuses mõõdetakse neid muutusi HbA1c analüüsidega. Leiutise teises teostuses manustatakse käesoleva leiutise kohaseid FGF-21 polüpeptiide ja Klotho-betat diabeedivastase raviainega ravitavale patsiendile, mis vähendab vere glükoosisisaldust rohkem kui ainult diabeedivastase raviaine kasutamine. XV. Leiutisekohaste FGF-21 polüpeptiidide terapeutilised kasutusalad Leiutisekohased FGF-21 polüpeptiidid on kasulikud mitmesuguste häirete ravis. 30 Leiutisekohaste FGF-21 polüpeptiididega võib ravida imetajaid, kes kannatavad mitteinsuliinisõltuva suhkurtõve (NIDDM, non-insulin dependent diabetes mellitus; 2. tüüpi suhkurtõbi), insuliinisõltuva suhkurtõve (1. tüüp), rasvumise, ebapiisava glükoosieemaldamise, hüperglükeemia, hüperinsulineemia ja mis tahes muu haiguse või seisundi all, mis võib olla vahendatud FGF-21 poolt. Glükoositalumatus on määratletud kui erandlik tundlikkus glükoosi suhtes. Hüperglükeemia on määratletud kui vere liigne suhkru- (glükoosi-) sisaldus. Hüperinsulineemia on määratletud kui normist kõrgem vere insuliinisisaldus. Insuliiniresistentsus on määratletud kui seisund, mille korral normaalse insuliinikogusega saadakse subnormaalne bioloogiline vastus. Inimestel määratletakse rasvumine kui kehamass, mis on üle % suurem antud populatsiooni ideaalsest kehamassist (R. H. Williams, Textbook of Endocrinology, 1974, lk ).
245 Suhkurtõbi on kliiniliste ilmingute alusel jagatud kaheks suureks rühmaks, milleks on mitteinsuliinisõltuv või küpses eas algav vorm, mida nimetatakse ka 2. tüüpi suhkurtõveks; ja insuliinisõltuv või juveniilses eas algav vorm, mida tuntakse ka 1. tüüpi suhkurtõve nime all. Kliiniliselt on enamik 2. tüüpi küpses eas algava vormiga diabeetikutest ülekaalulised, kusjuures haiguse kliiniliste sümptomite ilmingud ilmuvad tavaliselt üle 40aastasena. Vastupidiselt sellele ei ole 1. tüüpi juveniilses eas algava vormiga diabeetikud vanuse ja pikkuse suhtes ülekaalulised, haigus algab noores eas, sageli enne 30. eluaastat, kiiresti, kuigi 1. tüüpi suhkurtõbi võib tekkida ükskõik missuguses vanuses. Suhkurtõbi on inimeste ainevahetushäire, mille levimus on ligikaudu üks protsent üldpopulatsioonist, kusjuures neist neljandik on 1. tüüpi suhkurtõve juhtumid (Foster, D. W., Harrison's Principles of Internal Medicine, Chap. 114, lk ,. tr, McGraw- Hill, New York). Haigus avaldub hormoonindutseeritud ainevahetusanomaaliate seeriana, mis lõpuks põhjustavad raskeid pikaajalisi ja kurnavaid tüsistusi, mis haaravad mitmeid organsüsteeme, sealhulgas silmi, neerusid, närve ja veresooni. Patoloogiliselt iseloomustab haigust basaalmembraanide kahjustus, mis on tõestatav elektronmikroskoopial. Mitteinsuliinisõltuv suhkurtõbi (NIDDM, 2. tüüpi suhkurtõbi) on kurnav haigus, mida iseloomustab suur vere glükoosi, insuliini ja kortikosteroidide sisaldus. 2. tüüpi suhkurtõve esinemissagedus on suur ja tõusutendentsiga ja on muutumas juhtivaks surmade, haigestumise ja tervishoiukulude põhjuseks kogu maailmas (Amos et al., Diabetic Med. 14:S1-85, 1997). 2. tüüpi suhkurtõve põhjused ei ole hästi teada. 2. tüüpi suhkurtõve iseloomustab liigne glükoositootmine sõltumata insuliini olemasolust ja tsirkuleeriv glükoosisisaldus jääb ebapiisava glükoosi eemaldamise tõttu liiga suureks. Arvatakse, et tekib nii sihtkudede resistentsus insuliini toimete suhtes kui ka insuliinisekretsiooni vähenemine (β-raku puudulikkus). Peamised glükoosi homeostaasi puhul insuliinile reageerivad koed on maks, kus insuliin stimuleerib glükogeeni sünteesi ja pärsib glükoneogeneesi; lihas, kus insuliin stimuleerib glükoosi haaramist rakkudesse ja kus glükogeen stimuleerib glükoosi haaramist rakkudesse ja pärsib lipolüüsi. Diabeedi tagajärjel tekib glükoosisisalduse
246 244 suurenemine veres ja pikaajaline kõrgenenud veresuhkrusisaldus, mis näitab seisundi esinemist ja mis põhjustab veresoonte ja närvide kahjustusi Praegu on kasutusel mitmesugused farmakoloogilised lähenemised 2. tüüpi suhkurtõve ravis (Scheen et al., Diabetes Care, 22(9):68-77, 1999). Need toimivad erinevate toimemehhanismide kaudu: 1) sulfonüüluurea preparaadid stimuleerivad insuliinisekretsiooni; 2) biguaniidid (metformiin) toimivad glükoosi ärakasutamise soodustamise, maksa glükoosisünteesi vähendamise ja glükoosi soolest imendumise vähendamise kaudu; 3) α-glükosidaasi inhibiitorid (akarboos, miglitool) aeglustavad süsivesikute seedimist ja seega ka glükoosi imendumist soolest ning vähendavad söögijärgset hüperglükeemiat; 4) tiasolidiindioonid (troglitasoon) tugevdavad insuliini toimet, soodustades seega glükoosi ärakasutamist perifeersetes kudedes ja 5) insuliin stimuleerib glükoosi ärakasutamist kudedes ja pärsib glükoosi tootmist maksas. Ülalmainitud farmakoloogilisi lähenemisi võib kasutada eraldi või kombinatsioonis. Igal lähenemisel on piirangud ja kõrvaltoimed. Rasvumine on krooniline haigus, mille levimus kaasaegses ühiskonnas on suur ja mida seostatakse mitte ainult sotsiaalse stigma, vaid ka lühenenud eluea ja mitmesuguste meditsiiniliste probleemidega, sealhulgas negatiivne psühholoogiline areng, dermatoloogilised häired, näiteks infektsioonid, veenilaiendid, koormustaluvuse langus, suhkurtõbi, insuliiniresistentsus, hüpertensioon, hüperkolesteroleemia ja koronaartõbi. Rissanen et al., British Medical Journal, 301: (1990). Rasvumise olemasolevate ravimeetodite hulka kuuluvad standardsed dieedid ja füüsiline koormus, väga väikese kalorisisaldusega dieedid, käitumisteraapia, farmakoteraapia, sealhulgas isu vähendavad ained, termogeensed ravimid, söögi imendumise pärssijad, mehaanilised seadmed, näiteks lõualuu fikseerimine traatlinguga, vööpaelad ja balloonid ja operatsioonid. Jung ja Chong, Clinical Endocrinology, 35: 11- (1991); Bray, Am. J. Clin. Nutr., 55: 538S...544S (1992). Arvestades rasvumise suurt levimust meie ühiskonnas ja sellega seotud ülalkäsitletud raskeid tagajärgi, võib ükskõik missugusel terapeutilisel ravimil, mis on potentsiaalselt kasulik kehamassi vähendamiseks rasvunud isikutel, olla oluline kasulik toime nende isikute tervisele. Tehnika tasemes eksisteerib vajadus ravimi järele, mis vähendab
247 245 rasvunud isikute keha kogumassi ideaalse kehamassi suunas, omamata olulisi kõrvaltoimeid, ja mis aitab säilitada rasvunud isikutel vähendatud kehamassi Seega on soovitatav välja pakkuda raviskeem, mis on kasulik kehamassi langetamiseks rasvunud isikutel normaalse ideaalse kehamassi suunas. Lisaks on soovitatav välja pakkuda rasvumise ravi, millega saavutatakse alandatud kehamassi säilitamine pikema aja jooksul. Rasvumine korreleerub inimestel ja loomadel tehtud katsetes olulisel määral insuliiniresistentsus ja suhkurtõvega. Rasvumine ja insuliiniresistentsus, millest viimatinimetatuga kaasneb üldiselt hüperinsulineemia või hüperglükeemia või mõlemad, on mõlemad 2. tüüpi suhkurtõve iseloomulikud tunnused. Lisaks sellele on 2. tüüpi suhkurtõbi seotud kahe kuni neljakordse koronaartõve riskiga. Vaatamata aastakümneid kestnud uurimistööle nende raskete terviseprobleemide vallas on rasvumise ja insuliiniresistentsus etioloogia teadmata. 1. tüüpi suhkurtõbe iseloomustab tavaliselt väga väike või mittemõõdetav plasma insuliinisisaldus ja suurenenud glükagoonisisaldus. Sõltumata täpsest etioloogiast on enamikul 1. tüüpi suhkurtõvega patsientidel tsirkuleerivad antikehad, mis on suunatud nende enda pankreaserakkude vastu, sealhulgas antikehad insuliini, saarekeste Langerhansi rakkude tsütoplasma ja glutaamhappe dekarboksülaasi vastu. Spetsiifiliselt beeta-rakkude (insuliini sünteesivad rakud) vastu suunatud immuunvastus põhjustab 1. tüüpi suhkurtõve tekke. Seda spetsiifilisust toetab ülalkirjeldatud kliiniline leid, sest beetarakud sekreteerivad insuliini ja alfa-rakud glükagooni. 1. tüüpi suhkurtõve praegused raviskeemid sisaldavad dieedi modifikatsiooni kehaomase insuliini puudusest, mis omakorda on tingitud kahjustunud beeta-rakkudest, põhjustatud hüperglükeemia vähendamiseks. Dieeti modifitseeritakse ka seoses insuliini manustamisega hormooni hüpoglükeemiliste toimete neutraliseerimiseks. Sõltumata ravivormist on kõigil 1. tüüpi suhkurtõvega patsientidel vajalik insuliini parenteraalne manustamine, millest lähtub ka mõiste insuliinisõltuv suhkurtõbi. Seega esineb vajaduse 2. tüüpi suhkurtõve efektiivseks raviks, millel on vähem kõrvaltoimeid kui olemasolevatel farmatseutilistel lähenemistel. Lisaks võib efektiivne, insuliinile alternatiivne ravi olla kasulik ka 1. tüüpi suhkurtõve raviks. Käesolev kirjeldus
248 5 246 käsitleb farmakoloogilist ravi, mis stimuleerib glükoosi haaramist rakkudesse ja suurendab perifeersete kudede insuliinitundlikkust ning millel on vähem kõrvaltoimeid kui praegustel 2. tüüpi suhkurtõve raviks kasutavatel raviskeemidel. Lisaks sellele käsitleb käesolev kirjeldus alternatiivset ravi 1. tüüpi suhkurtõve korral. Lisaks on käesolev leiutis kasulik rasvumise ravis, suurendades energiakulu glükoosi kiirema ja tõhusama ärakasutamise kaudu. Käesolev kirjeldus käsitleb meetodit imetaja raviks, kellel esineb üks või mitu seisundit 1. tüüpi suhkurtõve, 2. tüüpi suhkurtõve, rasvumise, insuliiniresistentsus, hüperinsulineemia, glükoositalumatuse või hüperglükeemia hulgast, mis hõlmab seda ravi vajavale imetajale leiutisekohase FGF-21 polüpeptiidi terapeutiliselt efektiivse koguse manustamist. Ravimeetod võib olla küllaldane nimetatud imetajal vähemalt ühe alljärgneva modifikatsiooni saavutamiseks: keha rasvavarude vähendamine, insuliiniresisentsuse vähendamine, hüperinsulineemia vähendamine, glükoositaluvuse suurendamine ja hüperglükeemia vähendamine. 30 Teises toestuses käsitleb käesolev kirjeldus meetodit kodulooma ravimiseks, sealhulgas, kuid mitte ainult, veiste, sigade, lammaste, hobuste ja muude sarnaste ravimiseks, mis hõlmab FGF-21 või selle variandi tõhusa koguse manustamist keha rasvavarude vähendamiseks. Kodulooma keha rasvavarude pikaajaline või püsiv vähendamine on inimesele olulise majandusliku kasuga, sest loomad moodustavad suure osa inimdieedist ja loomne rasv võib inimesel põhjustada de novo rasvladestusi. Fibroblastide kasvufaktorit 21 (FGF-21) võib kasutada kriitiliselt haigete patsientidega seotud haigestumise ja suremuse vähendamiseks. Vt USA patendipublikatsiooni nr Pikema aja jooksul intensiivravi vajavatel kriitiliselt haigetel patsientidel on suurem surma ja olulise haigestumise risk. Üks sage põhjus patsientide hospitaliseerimiseks intensiivraviosakondadesse on süsteemne põletikuvastuse sündroom (SIRS, systemic inflammatory response syndrome), mida seostatakse infektsioonidega (sepsis), kuid ka mitteinfektsioossete patoloogiliste põhjuste, näiteks pankreatiidi, isheemia, hulgitrauma ja koekahjustusega, hemorraagilise šoki ja immuunvahendatud organkahjustusega. Käesolev kirjeldus hõlmab ka meetodit suremuse ja haigestumise vähendamiseks kriitiliselt haigetel patsientidel, kes kannatavad süsteemse põletikureaktsiooni sündroomi (SIRS) all, mis on seotud infektsioossete põhjuste, kuid ka
249 mitteinfektsioossete patoloogiliste põhjustega, mis hõlmab FGF-21 terapeutiliselt efektiivse koguse manustamist kriitiliselt haigele patsiendile. SIRSiga kulgevate seisundite näidete hulka kuuluvad sepsis, pankreatiit, isheemia, hulgitrauma ja koevigastus, hemorraagiline šokk, immuunvahendatud organkahjustus, äge respiratoorse distressi sündroom (ARDS, acute respiratory distress syndrome), šokk, neerupuudulikkus ja hulgiorganpuudulikkuse sündroom (MODS, multiple organ dysfunction syndrome). Käesolev kirjeldus hõlmab ka meetodit respiratoorse distressi all kannatavate kriitilisel haigete patsientide suremuse ja haigestumise vähendamiseks. SIRSi sage tüsistus on organsüsteemide puudulikkuse teke, sealhulgas äge respiratoorse distressi sündroom (ARDS), šokk, neerupuudulikkus ja hulgorganpuudulikkuse sündroom (MODS), mis kõik kordistavad negatiivse ravitulemuse riski. Hoolimata sellest, et paljud spetsialistid usuvad, et teatud tüüpi toitumistugi on kriitiliselt haigetele patsientidele kasulik metaboolse stabiilsuse taastamisel, on nende tugimeetodite kasulikud toimed ja spetsiifika vastuolulised, sest puuduvad hästi kontrollitud randomiseeritud kliinilised uuringud. Kuna hüperglükeemia ja insuliiniresistentsus on toitumistuge saavatel kriitiliselt haigetel patsientidel sagedased, manustatakse mõnes intensiivraviosakonnas liigse hüperglükeemia raviks toitu saavatele kriitiliselt haigetele patsientidele insuliini. Hiljutised uuringud näitavad, et eksogeense insuliini kasutamine vere glükoosisisalduse hoidmiseks mitte üle 1 mg/dl vähendas kirurgilistes intensiivraviosakondades kriitiliselt haigete patsientide haigestumist ja suremust sõltumata sellest, kas patsientidel varem oli suhkurtõbe esinenud või mitte (Van den Berghe, et al. N Engl J Med., 345(19):1359, 01). Käesolev kirjeldus hõlmab meetodit suremuse ja haigestumise vähendamiseks neil kriitiliselt haigetel patsientidel FGF-21 manustamise kaudu. Käesoleva leiutisega hõlmatud kriitiliselt haigetel patsientidel esineb tavaliselt ebastabiilne hüpermetaboolne seisund. See ebastabiilne metaboolne seisund on tingitud substraadi ainevahetuse muutustest, mis võib põhjustada teatud toitainete suhtelisi defitsiite. Üldiselt esineb nii rasvkoe kui ka lihaskoe suurem oksüdatsioon. Kriitiliselt haiged patsiendid, kellel FGF-21 manustamine võib vähendada suremuse ja haigestumise riski, on eelistatavalt patsiendid, kellel esineb süsteemne põletikuvastuse sündroom või respiratoorne distress. Haigestumise vähenemine tähendab tõenäosuse
250 248 vähenemist, et kriitiliselt haigel patsiendil tekivad täiendavad haigused, seisundid või sümptomid, või kaasuvate haiguste, seisundite või sümptomite raskusastme vähenemist. Näiteks võib haigestumise vähendamine vastata baktereemia või sepsise esinemissageduse või hulgiorganpuudulikkusega seotud tüsistuse vähenemisele Süsteemse põletikuvastuse sündroom (SIRS) kirjeldab põletikuprotsesse, mis on seotud paljude kliiniliste seisunditega ja hõlmab, kuid mitte ainult, rohkem kui ühte järgnevat kliinilist ilmingut: (1) kehatemperatuur üle 38 C või alla 36 C; (2) südame löögisagedus üle 90 korra minuti; (3) tahhüpnoe, mis väljendub hingamissageduses üle korra minutis, või hüpoventilatsioonis, mida näitab PaCo 2 alla 32 mm Hg; ja (4) vere valgeliblede arvu muutus, näiteks arv üle raku milliliitris või arv alla 4000 raku milliliitris või enam kui % ebaküpsete neutrofiilide esinemine. Need füsioloogilised muutused peaksid väljendama seisundi ägedat muutust algseisundiga võrreldes nende kõrvalekallete teiste teadaolevate põhjuste, näiteks keemiaravi, indutseeritud neutropeenia ja leukopeenia, puudumisel. Sepsis on määratletud kui infektsioonist tingitud SIRS. SIRSi mitteinfektsioossete patogeensete põhjuste hulka võivad kuuluda pankreatiit, isheemia, hulgitrauma ja koevigastus, sealhulgas, kuid mitte ainult, lömastavad vigastused või rasked põletused, hemorraagiline šokk, immuunvahendatud organkahjustus ja põletikuprotsesside arvatavate vahendajate, näiteks tuumori nekroosi faktor ja teised tsütokiinid, eksogeenne manustamine. Septiline šokk ja hulgiorganpuudulikkus on intensiivraviosakondades peamine haigestumise ja suremuse põhjus. Sepsis on seotud peremeesorganismi mitmesuguste kaitsemehhanismide, sealhulgas tsütokiinide võrgustik, leukotsüüdid ja komplemendi kaskaad, ja koagulatsiooni-/fibrinolüüsisüsteemide, sealhulgas endoteeli, aktivatsiooniga ning on nende poolt vahendatud. Dissemineeritud intravaskulaarne koagulatsioon (DIC, disseminated intravascular coagulation) ja muu raskusastmega kulutuskoagulopaatiad, mis on seotud fibriini ladestumisega mitmesuguste organite mikrovaskulatuuris, on sepsise / septilise šoki ilmingud. Peremeesorganismi kaitsemehhanismide otsesed toimed sihtorganitele on olulised mediaatorid hulgiorganpuudulikkuse sündroomi (MODS) tekkel, mis süvendavad sepsisega, raske sepsisega ja septilise šokiga patsiendi halba prognoosi.
251 Respiratoorne distress on seisundit, mille korral patsiendil on mis tahes tüüpi kopsuhäire tõttu raske hingata. Sageli esineb neil patsientidel erineva ulatusega hüpokseemia, mis võib olla refraktaarne ravile lisahapnikuga, kuid ei pruugi seda olla. Respiratoorne distress võib esineda patsientidel, kellel esineb kopsufunktsiooni häire otsese kopsukahjustuse tulemusena või kaudse kopsukahjustuse tulemusena, näiteks süsteemse protsessi tingimustes. Mitme soodustava häire olemasolu suurendab oluliselt riski, sarnaselt sekundaarsete faktorite, näiteks alkoholi krooniline kuritarvitamine, krooniline kopsuhaigus ja seerumi madal ph väärtus, olemasolule. Otsese kopsukahjustuse põhjuste hulka kuuluvad kopsupõletik, maosisu aspiratsioon, kopsukontusioon, rasvemboolia, peaaegu uppumine, inhalatsioonvigastus, kõrgmäestiku ja reperfusiooni kopsuturse pärast kopsusiirdamist või kopsuarteri embolektoomiat. Kaudse kopsukahjustuse põhjuste hulka kuuluvad sepsis, raske trauma koos šokiga ja mitmed transfusioonid; kehavälise kardiopulmonaalse ringe kasutamine, ravimi või uimasti üleannustamine, äge pankreatiit ja veretoodete transfusioonid. Üks respiratoorset distressi põhjustav kopsuhäirete klass on seotud kops-südame nime all tuntud sündroomiga. Need häired on seotud kroonilise hüpoksiaga, mille põhjuseks on rõhu tõus kopsuringes, mida nimetatakse pulmonaalhüpertensiooniks. Tekkinud pulmonaalhüpertensioon suurendab parema vatsakese töökoormust, põhjustades selle suurenemist või hüpertroofiat. Kops-süda avaldub tavaliselt parema südamepoole puudulikkusena, mida määratletakse parema vatsakese rõhkude püsiva suurenemise ja parema vatsakese venoosse naasu vähenemise kliiniliste tunnuste abil. Respiratoorset distressi põhjustavad ka krooniline obstruktiivne kopsuhaigus (COPD, chronic obstructive pulmonary disease), mis hõlmab emfüseemi ja kroonilist bronhiiti, mida iseloomustab õhuvoolu obstruktsioon. COPD on sageduselt neljas surmapõhjus ja selle tõttu sureb igal aastal üle inimese. Äge respiratoorse distressi sündroom (ARDS) on üldiselt progresseeruv haigus ja seda iseloomustavad eristatavad staadiumid. Sündroom avaldub tavaliselt kiiresti tekkiva hingamispuudulikkusena seisundi riskiteguriga patsiendil. Iseloomulik on lisahapnikuga ravile refraktoorse arteriaalse hüpokseemia esinemine. Allasetsevates kopsualades võib esineda alveolaarne täitumine, konsolidatsioon ja atelektaas; mitteallasetsevates alades võib esineda oluline põletik. See sündroom võib progresseeruda püsiva hüpokseemiaga
252 0 kulgevaks fibroseerivaks alveoliidiks, tekkida võib alveolaarse surnud ruumi suurenemine ja kopsu venitatavuse edasine vähenemine. Tekkida võib ka kopsu kapillaarsüsteemi kahjustusest tingitud pulmonaalhüpertensioon Kopsukahjustuse kliiniline raskusaste varieerub. Käesolev leiutis hõlmab patsiente, kellel arteriaalse hapniku osarõhu ja sissehingatava hapniku osakaalu vahelise suhte 300 või vähem kaudu määratletuna esineb vähem raske hüpokseemia, kui ka raskema hüpokseemiaga patsiente, kellel vastav suhe on 0. Üldiselt klassifitseeritakse patsiendid, kelle vastav suhe on 300 või väiksem, ägeda kopsukahjustusega patsientideks ja patsiendid, kelle vastav suhe on 0 või väiksem, klassifitseeritakse ägeda respiratoorse distressi sündroomiga patsientideks. Ägeda kopsukahjustuse ägedat faasi iseloomustab alveolaarse-kapillaarse barjääri vaskulaarse permeaabluse suurenemise tagajärjel valgurikka tursevedeliku sissevool õhuruumidesse. Epiteeliterviklikkuse kadumine permeaabluse muutusega kohtades võib põhjustada alveolaarid täitumist vedelikuga, häirida normaalset vedelikutransporti, mis mõjutab tursevedeliku eemaldamist alveolaarruumist, vähendab surfaktandi tootmist ja käitlemist, põhjustab bakteriaalse kopsupõletikuga patsientidel septilist šokki ja põhjustab fibroosi. Sepsis on seotud ägedaks kopsukahjustuseks progresseerumise suurima riskiga. Selliste seisundite korral nagu sepsis, mil esineb hüpermetabolism, esineb valkude kiirem lagunemine nii glükoneogeneesi tagamiseks kui ka suurema valgusünteesi jaoks vajalike aminohapete vabastamiseks. Esineda võib hüperglükeemia ja suur triglütseriidide ja teiste lipiidide suur kontsentratsioon seerumis. Respiratoorse funktsiooni häirega patsientidel võib hüpermetabolism mõjutada süsinikdioksiidi tootmise ja hapnikutarbimise vahelist suhet. See on tuntud kui respiratoorne koefitsient (R/Q) ja tervetel isikutel on see vahemikus ligikaudu 0,85 kuni ligikaudu 0,90. Liigsel rasvametabolismil on kalduvus langetada R/Q väärtust, samas kui liigne glükoosimetabolism suurendab R/Q väärtust. Respiratoorse distressiga patsientidel on sageli probleeme süsinikdioksiidi eemaldamisega ja neil on seega ebanormaalselt kõrged respiratoorsed koefitsiendid. Käesoleva leiutisega hõlmatud kriitiliselt haigetel patsientidel esineb tavaliselt ka spetsiifiline stressreaktsioon, mida iseloomustab enamike rakusaaduste ajutine
253 5 1 allaregulatsioon ja kuumašoki valkude ülesregulatsioon. Lisaks hõlmab see stressreaktsioon hormoonide, näiteks glükagooni, kasvuhormooni, kortisooli ja pro- ja antiinflammatoorsete tsütokiinide aktivatsiooni. Kuigi stressreaktsioonil näib olevat kaitsefunktsioon, tekitab vastus neil kriitiliselt haigetel patsientidel täiendava metaboolse ebastabiilsuse. Näiteks põhjustab nende spetsiifiliste hormoonide aktivatsioon seerumi glükoosisisalduse suurenemist, mis põhjustab hüperglükeemia tekke. Lisaks võib adrenergiline stimulatsioon süvendada südame ja teiste organite kahjustust. Esineda võivad ka kilpnäärmetöö muutused, millel on oluline mõju metaboolsele aktiivsusele. FGF-21 keskmine kogus võib varieeruda ja eelkõige peab see põhinema kvalifitseeritud arsti soovitustel ja retseptil. FGF-21 täpne kogus on eelistuse küsimus ja sõltub sellistest teguritest nagu ravitava seisundi täpne tüüp, ravitava patsiendi seisund ja kompositsiooni muud koostisained. Leiutis käsitleb ka teise toimeaine terapeutiliselt efektiivse koguse manustamist. Manustatava koguse suudab FGF-21 ravi alusel erialaasjatundja kergesti määrata. Leiutisekohaseid farmatseutilisi kompositsioone võib valmistada tavapärasel viisil. NÄITED Järgmised näited on mõeldud leiutise illustreerimiseks ja need ei piira taotletava leiutiskaitse ulatust Näide 1 Käesolevas näites kirjeldatakse osasid paljudest potentsiaalsetest kriteeriumitest, mille alusel valitakse FGF-21 saite, kuhu sisestatakse mittelooduslikke aminohappeid. Joonisel FIG 1 on näidatud FGF-21 (valgu registreerimisnumber BC018404) ja FGF-19 (valgu registreerimisnumber BAA75500) järjestuste homoloogilisus, mis on määratud vahendiga Vector NTI (Invitrogen; Carlsbad, CA). Kahe polüpeptiidi sarnased aminohapped on tähistatud tärniga. Loodusliku aminohappe asendamise teel mittelooduslikult kodeeritud aminohappega loodi seitse erinevat FGF-21 polüpeptiidi. Igas polüpeptiidis asendati üks aminohape, mis on joonisel FIG 1 tähistatud nelinurgaga, paraatsetüülfenüülalaniiniga. Polüpeptiididel üldiselt puudus joonistel FIG 1 ja FIG 3 näidatud liiderjärjestus ja neil oli
254 5 2 N-otsas 6 histidiinijäägiga. SEQ ID NO.: 1 on 181 aminohappest koosnev inimese FGF-21 järjestus (P vorm), millel puudub liiderjärjestus. SEQ ID NO.: 2 on inimese FGF-21 järjestus (P vorm), millel puudub liiderjärjestus ja millel on N-otsas His-märgis. Igal loodud polüpeptiidil oli järjestuse SEQ ID NO: 1 ühel positsioonidest, 52, 77, 117, 126, 131, ja 162 mittelooduslikult kodeeritud aminohappega tehtud asendus. Joonisel FIG 2 on näidatud Protein Data Bank (PDB) (Bernstein et al. J. Mol. Biol. 1997, 112, pp 535) (IPWA) saadud ja PyMOL tarkvara (DeLano Scientific; Palo Alto, CA) abil märgistatud inimese FGF-19 struktuur. FGF-19 V34, L79, G4, S144, K5, L160 ja S196-le vastavad aminohapped asendati leiutisekohastes FGF-21 polüpeptiidides paraatsetüülfenüülalaniiniga. Punktiirjoonega on näidatud piirkonnad, mida ei lahutatud originaalstruktuuris. Joonisel FIG 3 on näidatud FGF-21 (valgu registreerimisnumber BC018404) ja FGF-2 (valgu registreerimisnumber BAA75500) järjestuste vaheline homoloogia, mis on määratud vahendiga 30 Vector NTI (Invitrogen; Carlsbad, CA). Kahe molekuli sarnased aminohapped on tähistatud tärniga. Allajoonitud aminohapped on kahe polüpeptiidi korral identsed. Seitse aminohapet, mis on näidatud joonisel FIG 1 asendussaitidena, on paigutatud ka joonisel FIG 3 toodud nelinurka. Joonisel FIG 4 on näidatud inimese FGF-2 struktuur, mis saadi PDB-st (ICVS) ja märgistati PyMOL tarkvaraga (DeLano Scientific; Palo Alto, CA). Hallid struktuurid vastavad inimese FGF retseptorile 1 (FGFR1) ja mustad struktuurid inimese FGF2-le. Plotnikov, AN et al. Cell Sep 3;98(5): kirjeldavad FGF retseptoriga seotud FGF2 kristallstruktuuri. Leiutisekohastes aminohapetes asendati FGF- 2 aminohappejääkidele F21, K62, K86, V1, K134, T139 vastavad aminohapped paraatsetüülfenüülalaniiniga. Punktiirjoonega on näidatud piirkonnad, mida ei lahutatud originaalstruktuuris. Teised kriteeriumid, mille järgi valitakse mittelooduslike aminohapete sisestamiseks eelistatud saite, on järgmised. Valkude andmebaasist Protein Data Bank pärineva kümne kristallstruktuuri abil modelleeriti FGF-21: 1PWA (inimese FGF-19); 1IJT (inimese FGF-4); INUN (inimese FGF-FGF retseptori 2b kompleks); 1G82 (inimese FGF-9 dimeer FGF retseptori ja hepariiniga); 1IHK (inimese FGF-9); IBAR (veise FGF-1); 1QQK (roti FGF-7); 1K5U (inimese FGF-1); 1FQ9 (inimese FGF-2 FGF retseptoriga 1 ja hepariiniga); ja 2FDB (inimese FGF-8b koos FGF retseptoriga 2c) struktuur. Nende struktuuride koordinaate on võimalik saada valkude andmebaasist (PDB)
255 (Bernstein et al. J. Mol. Biol. 1997, 112, lk 535). Kristallstruktuuride võrdlus näitas, et kõigi nende põhistruktuur oli sarnane. Samas aga osutusid N- ja C-otsad nende FGF molekulide puhul väga erinevateks ja seetõttu ei olnud võimalik otsi modelleerida. Modelleerimisel identifitseeriti kaks aminohappejääki ehk Y22 ja Y4, mis olid väga konserveerunud ja osalesid retseptoriga seondumises. Lisaks identifitseeriti ka kaks potentsiaalset hepariini sidumiskohta, mis olid seotud aminohappejääkidega R36 ja E37. Retseptoriga seonduvate ja hepariiniga seonduvate aminohappejääkide positsioonid vastavad järjestusele SEQ ID NO: 1. Selle tulemusena identifitseeriti aminohappejäägid, mis 1) ei takista FGF retseptori või hepariiniga seondumist, 2) ei sisaldu valgu sisemuses ja 3) paiknevad piirkondades, mis on kristallstruktuuridel üsna sarnased. Üks mittelooduslik aminohape sisestatakse FGF-21: 8 SEQ ID NO: 1 järgmistesse positsioonidesse (või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). Selleks et sisestada mittelooduslikult kodeeritud aminohapet, hinnati FGF-21 kõiki positsioone: aminohappejääk (a) ei tohiks struktuurianalüüsi põhjal takistada FGF-21 retseptori seondumist, b) seda aminohappejääki ei tohiks mõjutada alaniin- või homoloogskaneeriv mutagenees (c) see peaks olema pinnal eksponeeritud ja omama minimaalselt van der Waalsi või vesiniku sidumise interaktsioone ümbritsevate aminohappejääkidega, (d) peaks olema FGF-21 variantides deleteeritud või varieeruma, (e) tagama mittelooduslikult kodeeritud aminohappega asendamisel konservatiivsed muutused ja (f) paiknema väga paindlikes piirkondades või struktuurselt jäikades piirkondades. Ühe või mitme mittelooduslikult kodeeritud aminohappe FGF-21 valku sisestamiseks sobivate saitide valimiseks võib kasutada ka teisi või erinevaid FGF perekonna liikmete kristallstruktuure nagu FGF-21 ja/või FGF-19 struktuure. Näiteks inimese FGF-19 (PDB ID 2P23) kristallstruktuurist ja/või inimese FGF-19 (PDB ID 2P23) kristallstruktuurist ja/või inimese FGF-23 (PDB ID 2P39) kristallstruktuurist võib saada lisateavet, mille põhjal saab valida FGF-21 valgu saite, mis sobivad ühe või mitme mitteloodusliku aminohappe sisestamiseks. Sellised saidi võivad paikneda valgu erinevates piirkondades, näiteks, kuid mitte ainult N- ja C-otsas, retseptori sidumispiirkonnas ja hepariini sidumispiirkonnas. Selleks et hinnata iga valguaatomi eendumist, saab lisaks FGF-21 1 molekuli hinnata ka Cx programmiga (Pintar et al. (02) Bioinformatics, 18, lk 980) tehtavate arvutuste põhjal.
256 4 Üks mittelooduslik aminohape sisestatakse positsioonidesse FGF-21: 8 (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). Näide 2 5 Selles näites antakse ülevaade mittelooduslikult kodeeritud aminohapet sisaldava FGF-21 polüpeptiidi kloonimisest ja ekspressioonist E. coli bakterites. Selles näites kirjeldatakse ka modifiseeritud FGF-21 polüpeptiidide bioloogilise aktiivsuse hindamismeetodit. FGF-21 kloonimismeetodid on asjatundjatele tuntud. FGF-21 polüpeptiidide ja polünukleotiidide järjestusi ja FGF-21 kloonimist peremeesrakkudesse kirjeldatakse USA patendis nr , USA patendipublikatsioonides nr 05/ , 05/ , 04/ , 04/09780, 02/ ja 01/ ; WO 01/36640; WO 03/011213; WO 03/059270; WO 04/1472; WO 05/061712; WO 05/072769; WO 05/091944; WO 05/113606; WO 06/028595; WO 06/028714; WO 06/050247; WO 06/065582; WO 06/ cdna, mis kodeerib liiderjärjestuseta FGF-21 P vormi, on näidatud järjestusena SEQ ID NO: 8. See polüpeptiid vastab järjestusele SEQ ID NO: 1. cdna, mis kodeerib liiderjärjestuseta FGF-21 His-märgisega P vormi, on näidatud järjestuses SEQ ID NO: 9. See polüpeptiid vastab järjestusele SEQ ID NO: 2. cdna, mis kodeerib FGF-21 P vormi koos 3 leutsiini sisaldava liiderjärjestusega, on näidatud järjestusena SEQ ID NO:. See polüpeptiid vastab järjestusele SEQ ID NO: 3. cdna, mis kodeerib FGF-21 P vormi koos 2 leutsiini sisaldava liiderjärjestusega, on näidatud järjestusena SEQ ID NO: 11. See polüpeptiid vastab järjestusele SEQ ID NO: 4. cdna, mis kodeerib liiderjärjestuseta FGF-21 L vormi, on näidatud järjestusena SEQ ID NO: 12. See polüpeptiid vastab järjestusele SEQ ID NO: 5. cdna, mis FGF-21 L vormi koos 3 leutsiini sisaldava liiderjärjestusega, on näidatud järjestusena SEQ ID NO: 13. See polüpeptiid vastab järjestusele SEQ ID NO: 6. cdna, mis FGF-21 L vormi koos 2 leutsiini sisaldava liiderjärjestusega, on näidatud järjestusena SEQ ID NO: 14. See polüpeptiid vastab järjestusele SEQ ID NO: 7.
257 5 5 Mittelooduslikku aminohapet sisaldava FGF-21 ekspresseerimiseks kasutatakse sisestatud translatsioonisüsteemi, mis sisaldab ortogonaalset trna-d (O-tRNA) ja ortogonaalset aminoatsüül-trna süntetaasi (O-RS). O-RS eelkõige aminoatsüülib O-tRNA, milles on mittelooduslikult kodeeritud aminohape. Translatsioonisüsteem omakorda sisestab vastusena kodeeritud selektorkoodonile FGF-21 polüpeptiidi mitteloodusliku aminohappe. Sobivaid O-RS ja O-tRNA järjestusi kirjeldatakse patendidokumendis WO 06/ pealkirjaga Compositions of Aminoacyl-tRNA Süntetaas and Uses Thereof (E9; SEQ ID NO: ) ja patendidokumendis WO 07/ pealkirjaga Compositions of trna and Uses Thereof (F13; SEQ ID NO: 16) Tabel 2: O-RS ja O-tRNA järjestused SEQ ID NO: 17 M. jannaschii mtrna trna SEQ ID NO: 18 HLAD03; optimeeritud amber-supressor-trna trna SEQ ID NO: 19 HL3A; optimeeritud AGGA raaminihke supressortrna trna SEQ ID NO: Aminoatsüül- trna süntetaas p-asido-l-fenüülalaniin p-az-phers(6)sisestamiseks RS SEQ ID NO:21 Aminoatsüül-tRNA süntetaas p-bensoüül-lfenüülalaniin p-bpars(1)sisestamiseks RS SEQ ID NO:22 Aminoatsüül-tRNA süntetaas propargüül-fenüülalaniin propargüül-phers sisestamiseks RS SEQ ID NO:23 Aminoatsüül-tRNA süntetaas propargüül-fenüülalaniin propargüül-phers sisestamiseks RS SEQ ID NO:24 Aminoatsüül-tRNA süntetaas propargüül-fenüülalaniin RS
258 6 propargüül-phers sisestamiseks SEQ ID NO: Aminoatsüül-tRNA süntetaas p-asido-fenüülalaniin p- Az-PheRS(1)sisestamiseks RS SEQ ID NO:26 Aminoatsüül-tRNA süntetaas p-asido-fenüülalaniin p- Az-PheRS(3)sisestamiseks RS SEQ ID NO:27 Aminoatsüül-tRNA süntetaas p-asido-fenüülalaniini p- Az-PheRS(4) sisestamiseks RS SEQ ID NO:28 Aminoatsüül-tRNA süntetaas p-asido-fenüülalaniini p- Az-PheRS(2) sisestamiseks RS SEQ ID NO:29 Aminoatsüül-tRNA süntetaas p-atsetüül-fenüülalaniini (LW1)sisestamiseks RS SEQ ID NO:30 Aminoatsüül-tRNA süntetaas for the incorporation of p- acetyl -fenüülalaniini (LW5) RS SEQ ID NO:31 Aminoatsüül-tRNA süntetaas p-atsetüüll-fenüülalaniini (LW6)sisestamiseks RS SEQ ID NO:32 Aminoatsüül-tRNA süntetaas p-asido-fenüülalaniini (AzPheRS-5)sisestamiseks RS SEQ ID NO:33 Aminoatsüül-tRNA süntetaas p-asido-fenüülalaniini (AzPheRS-6)sisestamiseks RS E.coli bakterite transformeerimine plasmiididega, mis sisaldavad modifitseeritud FGF-21 geeni ja ortogonaalse aminoatsüül-trna süntetaasi/trna paari (mis on spetsiifiline soovitud mittelooduslikult kodeeritud aminohappe suhtes) võimaldab sisestada mittelooduslikult kodeeritud aminohapet kohtspetsiifiliselt FGF-21 polüpeptiidi.
259 5 7 Metsikut tüüpi FGF-21 polüpeptiidi kordistati PCR-meetodil cdna sünteesireaktsiooni saadusest, mis saadi standardprotokollide abil terve inimese maksa polya+ mrna-st (Biochain) ja klooniti vektorisse pet30 (NcoI-BamHI). Pärast järjestuse kontrollimist subklooniti FGF-21, mis sisaldas N-terminaalset HHHHHHSGG järjestust, supressioonivektorisse, mis sisaldas Methanococcus jannaschii (Mj trna Tyr/CUA ) ambersupressor-türosüül TRNA Tyr/CUA d, mida kontrollis konstitutiivselt sünteetiline promootor, mis pärines E. coli lipovalgu promootorjärjestusest (Miller, J.H., Gene, 1986), ja ka ortogonaalset türosüül-trna-süntetaasi (MjTyrRS), mis on E. coli GlnRS promootori kontrolli all. FGF-21 ekspressiooni kontrollis T7 promootor. Amber-mutatsioonid tehti standardsete quick change mutation protokollide järgi (Stratagene; La Jolla, California). Kõikide konstruktide järjestust kontrolliti. Supresseerimine para-atsetüül-fenüülalaniiniga (pacf) 30 Plasmiidid (pvk3-hisfgf21) transformeeriti E. coli tüvesse W31 B2, milles T7 polümeraasi ekspressiooni kontrollis arabinoosi indutseeritav promootor. Öö läbi kasvatatud bakterikultuure lahjendati vahekorras 1:0 loksutuskolbides, mis sisaldasid 2X YT kasvusöödet, ja kasvatati temperatuuril 37 C optilise tiheduseni OD = ~ 0,8. Valgu ekspressioon kutsuti esile arabinoosi (lõppkontsentratsioon 0,2%) ja para-atsetüülfenüülalaniini (pacf) lisamise teel lõppkontsentratsioonini 4 mm. Rakukultuure inkubeeriti 4 tundi temperatuuril 37 C. Rakkudest valmistati pelletid ja suspendeeriti need uuesti B-PER lüüsipuhvris (Pierce) 0 ul/od/ml + ug/ml Dnaasis ja inkubeeriti 30 minutit temperatuuril 37 C. Rakumaterjal eraldati tsentrifuugimise teel ja supernatant eemaldati. Pellet suspendeeriti uuesti võrdses koguses SDS-PAGE Protein Loading puhvris. Kõik proovid kanti 4-12% PAGE geelile koos MES ja DTT-ga. FGF-21 puhastusmeetodid on asjatundjatele tuntud ja nende efektiivsust saab kontrollida SDS- PAGE-meetodil, Western blot analüüside või elektropihustusionisatsiooniga ioonlõksmassispektromeetria abil ja teistel sarnastel meetoditel. N-terminaalse His-märgisega FGF-21 ekspressiooni ja supressiooni 7 amber-saitidel on kujutatud joonisel FIG 5. FGF-21 polüpeptiid on tähistatud noolega. Joonisel FIG 5 on näidatud B-PER pelleti proovid-rada 1: marker; rada 2: VK3-FGF21 eelindutseerimine, supernatant; rada 3: VK3-FGF21 eelindutseerimine, pellet; rada 4: VK3-FGF21 0,2%
260 5 8 arabinoos, supernatant; rada 5: VK3-FGF21 0,2% arabinoos, pellet; rada 6: VK3-FGF21- pacf- L, 0,2% arabinoos; rada 7: VK3-FGF21- pacf- L52, 0,2% arabinoos; rada 8: VK3-FGF21- pacf- R77, 0,2% arabinoos; rada 9: VK3-FGF21- pacf- H117, 0,2% arabinoos; rada : VK3-FGF21- pacf- R126, 0,2% arabinoos; rada 11: VK3-FGF21- pacf- R131, 0,2% arabinoos; rada 12: VK3-FGF21- pacf-s 162, 0,2% arabinoos. Aminohappe asenduste puhul näidatud positsioonide numbrid vastavad järjestusele SEQ ID NO: 1. Joonisel FIG 6 on näidatud B-PER supernatandi proovid--rada 1: VK3-FGF21 eelindutseerimine, supernatant; Rada 2: VK3-FGF21 eelindutseerimine, pellet; rada 3: Marker; rada 4: VK3-FGF21 0,2% arabinoos, supernatant; rada 5: VK3-FGF21 0,2% arabinoos, pellet; rada 6: VK3-FGF21- pacf- L, 0,2% arabinoos; rada 7: VK3-FGF21- pacf- L52, 0,2% arabinoos; rada 8: VK3-FGF21- pacf- R77, 0,2% arabinoos; rada 9: VK3-FGF21- pacf- H117, 0,2% arabinoos; rada : VK3-FGF21- pacf- R126, 0,2% arabinoos; rada 11: VK3-FGF21- pacf-r131, 0,2% arabinoos; rada 12: VK3-FGF21- pacf- S162, 0,2% arabinoos. Aminohappe asenduste puhul näidatud positsioonide numbrid vastavad järjestusele SEQ ID NO: 1. His-märgisega mutantseid FGF-21 valke saab puhastada asjatundjatele tuntud meetoditel. Selleks sobib vaik ProBond Nickel-Chelating Resin (Invitrogen, Carlsbad, CA), mida kasutatakse tootja poolt kaasa antud standardsetel His-märgisega valgu puhastusprotseduuride abil. pvk (joonis FIG 24) töötati välja märgiseta FGF-21 valgule, millel on joonisel FIG näidatud järjestus. Selle vektori abil valmistati R36am ja Y83am mutandid ja rohkem teavet selliste His-märgiseta FGF-21 mutantsete valkude ja nende puhastamise kohta on toodud allpool näidetes ja joonistel. 3T3-L1 diferentseerumine rasvarakkudeks ja glükoosi haarde analüüs 30 Selleks et hinnata FGF-21 polüpeptiidide bioloogilist aktiivsust, võib teha järgmise analüüsi. Hiire 3T3-L1 fibroblastid (ATCC #CL-173) külvatakse cm suurusesse nõusse, milles on DMEM, mis sisaldab % vasikaseerumit. Enne rasvarakkude diferentseerumise algust lastakse rakkudel kasvada 0% konfluentsini; söödet tuleb vahetada iga päev. Rakke loendatakse ja külvatakse koguses 000 rakku süvendi kohta 96 süvendiga plaadile (rakud võib kanda ka Cytostar-T 96 süvendiga plaadile) ja neid inkubeeritakse veel 48 tundi. Diferentseerumise indutseerimiseks lisatakse pärast eelmise kasvusöötme lisamist järgmist söödet: DMEM, millele on lisatud % FBS (veise loote seerum), 1 µm
261 deksametasooni (DBX), 0,5 mm 3-isobutüül-1-metüülksantiini (IBMX) ja 5 µg/ml insuliini. Teine võimalus diferentseerumise indutseerimiseks on töödelda rakke 1 µm rosiglitasooniga ja inkubeerida 6 päeva enne söötme asendamist DMEM/% FBS-iga, kuna sellel viisil saab kiiremini indutseerida 3T3-L1 fibroblastide diferentseerumist rasvarakkudeks. Kolmandaks võimaluseks on kiirendada diferentseerumist nimetatud kahe protseduuri kombineerimise teel. Rakkudele lisatakse DBX/IBMX/insuliini sisaldavat söödet ja neid inkubeeritakse 48 tundi. Sööde asendatakse DMEM/% FBS/5 µg/ml insuliiniga ja rakke inkubeeritakse 48 tundi. Seejärel asendatakse sööde DMEM/% FBS-iga ja sööde asendatakse iga 2 päeva järel värske söötmega. Rakud diferentseeruvad 7 kuni 14 päeva. Diferentseerunud rakkudesse kogunevad lipiidide piisad. Rakke saab värvida tootega OIL RED O. Kui 95% rasvarakkudest sisaldavad lipiidide piiskasid, saab rakke kasutada glükoosi haarde analüüsi tegemiseks. Diferentseerunud 3T3-L1 rakke töödeldakse rakkude näljutamiseks 18 tundi FGF-21-ga (1 µg/ml) DMEM-is, millele on lisatud 0,1% rasvhappevaba BSA-d. Seejärel pestakse rakke Kreb's-Ringer HEPES puhvriga (KRH= 0,118M NaCl, 5 mm KCl, 2,54 mm CaCl 2, 1,19 mm KH 2 PO 4, 1,19 mm MgSO 4 ja mm HEPES), millele on lisatud 0,1% FAF-BSA-d. Märgistussegu valmistamiseks lisatakse KRH/0,1% FAF-BSA puhvrile 4 µci, 0,1 mm 2- deoksüd-[1-3 H]-glükoosi. Sellele lisatakse rakud ja neid inkubeeritakse 1 tund temperatuuril 37 C. Reaktsiooni lõpetamiseks pestakse rakke jääkülma PBS-iga, mis sisaldab µm tsütohalasiini B. Plaati kuivatatakse, et eemaldada mis tahes puhvrijäägid. Igasse süvendisse lisatakse stsintillatsioonivedelikku ja proove loendatakse seadmega TopCounter. Teine võimalus glükoosi haarde mõõtmiseks on kanda diferentseerunud 3T3-L1 rakud mitteradioaktiivse substraadiga 2-NBDG ja registreerida fluorestsents-laadilugejaga vastavad näitajad. Kaudseks võimaluseks mõõta glükoosi haaret on mõõta GLUT1 või GLUT4 ekspressiooni rakumembraani pinnal. GLUT1 and GLUT4 on glükoosi transporterid, mis insuliini või FGF-21 stimuleerimisel translokeeritakse sisemistest vesiikulitest rakumembraani pinnale. Elusrakke saab analüüsida ELISA-analüüsiga.
262 Näide Selles näites kirjeldatakse karbonüülrühma sisaldava aminohappe sisestamist ja sellele järgnevat reaktsiooni aminooksürühma sisaldava PEG-iga. 5 Selles näites demonstreeritakse meetodit, mille abil luuakse FGF-21 polüpeptiid, mis sisaldab ketoonrühma sisaldavat mittelooduslikult kodeeritud aminohapet, mis pannakse reageerima aminooksürühma sisaldava PEG-iga, mille molekulmass on umbes Iga jääk, mis eelneb positsioonile 1 (ehk paikneb N-otsas), positsioonis 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (ehk valgu karboksüülotsas) (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7) asendatakse eraldi mittelooduslikult kodeeritud aminohappega, millel on järgmine struktuur: Järjestused, mida kasutatakse p-atsetüül-fenüülalaniini kohtspetsiifiliseks sisestamiseks FGF-21 polüpeptiidi, on SEQ ID NO: 1 (FGF-21) ja SEQ ID NO: 16 või 17 (muttrna, M. jannaschii mtrna 2. ) ja, 29, 30 või 31 (TyrRS LW1, 5 või 6), mida kirjeldatakse eespool näites
263 5 261 Modifiseeritud FGF-21 polüpeptiidi variant, mis sisaldab karbonüülrühma sisaldavat aminohapet, pannakse reageerima aminooksürühma sisaldava PEG derivaadiga, mis vastab valemile R-PEG(N)-O-(CH 2 ) n -O-NH 2, milles R on metüülrühm, n on 3 ja N on molekulmassiga umbes p-atsetüülfenüülalaniini sisaldav puhastatud FGF-21, mis on lahustatud kontsentratsiooniga mg/ml mm MES-is (Sigma Chemical, St. Louis, MO), ph 6,0, mm Hepes (Sigma Chemical, St. Louis, MO) ph 7,0, või mm naatriumatsetaadis (Sigma Chemical, St. Louis, MO), ph 4,5, pannakse reageerima - kuni 0-kordse liiaga aminooksürühma sisaldava PEG-iga ja segatakse seejärel -16 tundi toatemperatuuril (Jencks, W. J. Am. Chem. Soc. 1959, 81, lk 475). PEG-FGF-21 lahjendatakse seejärel koheseks puhastamiseks ja analüüsimiseks sobivas puhvris. Näide 4 Konjugeerimine PEG-ga, mis sisaldab amiidsideme kaudu PEG-ga seotud hüdroksüülamiinrühma. Alljärgneva struktuuriga PEG reagent seotakse näites 3 kirjeldatava protseduuri abil ketoonrühma sisaldava mittelooduslikult kodeeritud aminohappega: R-PEG(N)-O-(CH 2 ) 2 -NH-C(O)(CH 2 ) n -O-NH 2 milles R = metüülrühm, n=4 ja N on molekulmassiga umbes 000. Reaktsiooni-, puhastus- ja analüüsitingimused vastavad näites 3 kirjeldatud tingimustele. Näide 5 Selles näites kirjeldatakse kahe eraldiseisva mittelooduslikult kodeeritava aminohappe sisestamist FGF-21 polüpeptiididesse. Selles näites demonstreeritakse meetodit, mille abil luuakse FGF-21 polüpetiid, mis sisaldab funktsionaalset ketoonrühma sisaldavat mittelooduslikku aminohapet kahes positsioonis, mis on valitud järgmiste aminohappejääkide seast: iga jääk, mis eelneb positsioonile 1 (ehk paikneb N-otsas), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118,
264 , 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (ehk paikneb valgu karboksüül-otsas) (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7). FGF-21 polüpeptiid valmistatakse näidetes 1 ja 2 kirjeldatud viisil, kuid selektorkoodon sisestatakse nukleiinhappes kahte eraldiseisvasse kohta. Näide 6 Selles näites kirjeldatakse FGF-21 polüpeptiidi konjugeerimist hüdrasiidrühma sisaldava PEG-iga ja sellele järgnevat in situ redutseerimist. Karbonüülrühma sisaldavat aminohapet sisaldav FGF-21 polüpeptiid valmistatakse näidetes 2 ja 3 kirjeldatava protseduuri abil. FGF-21 polüpeptiid konjugeeritakse modifitseeritud, hüdrasiidrühma sisaldava PEG-iga, millel on järgmine struktuur: R-PEG(N)-O-(CH 2 ) 2 -NH-C(O)(CH 2 ) n -X-NH-NH 2 milles R = metüülrühm, n=2 ja N = 000 MW ja X on karbonüül (C=O) rühm. p- atsetüülfenüülalaniini sisaldav puhastatud FGF-21 lahustatakse kontsentratsiooniga 0,1- mg/ml mm MES (Sigma Chemical, St. Louis, MO), ph 6,0, mm Hepes (Sigma Chemical, St. Louis, MO) ph 7,0, või mm naatriumatsetaadid (Sigma Chemical, St. Louis, MO) ph 4,5, pannakse reageerima 1- kuni 0-kordse liiaga hüdrasiinrühma sisaldava PEG-iga, ja vastava hüdrasooni redutseerimiseks in situ lisatakse lõppkontsentratsioonini -50 mm lähteainet 1M NaCNBH 3 (Sigma Chemical, St. Louis, MO), mis on lahustatud vees (H 2 O). Reaktsioonidel lastakse toimuda tundi pimedas temperatuuril 4 C. Reaktsioonide lõpetamiseks lisati 1M Tris puhvrit (Sigma Chemical, St. Louis, MO), ph umbes 7,6, Tris puhvri lõppkontsentratsioonini 50 mm või lahjendati koheseks puhastamiseks sobivasse puhvrisse. Näide 7
265 263 Selles näites kirjeldatakse alküünrühma sisaldava aminohappe sisestamist FGF-21 polüpeptiidi ja derivatiseerimist mpeg-asiidiga. 5 Järgmised aminohappejäägid: jäägid, mis eelnevad positsioonile 1 (ehk paiknevad N- otsas), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26,.27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (ehk paikneb valgu karboksüül-otsas) (SEQ ID NO: 1 või vastavad aminohappejäägid järjestustes SEQ ID NOs: 2-7) asendatakse kõik järgmise mittelooduslikult kodeeritud aminohappega : Järjestused, mida kasutatakse p-propargüültürosooni kohtspetsiifiliseks sisestamiseks FGF-21 polüpeptiidi, on SEQ ID NO: 1 (FGF-21), SEQ ID NO: 16 või 17 (muttrna, M. jannaschii mtrna ) ja 22, 23 või 24, mida kirjeldatakse eespool näites 2. Propargüültürosiini sisaldavat FGF-21 polüpeptiidid ekspresseeritakse E.coli bakterites ja puhastatakse näites 3 kirjeldatavates tingimustes. Propargüültürosiini sisaldav puhastatud FGF-21, mis on lahustatud kontsentratsiooniga 0,1- mg/ml PB puhvris (0 mm naatriumfosfaati, 0, M NaCl, ph = 8 ja reaktsioonisegule lisatakse - kuni 00-kordse liiaga PEG-i, mis sisaldab asiidfragmenti. Seejärel lisatakse reaktsioonisegule katalüütilises koguses CuSO 4 ja vasktraati. Reaktsioonisegu inkubeeriti (näiteks, kuid mitte ainult, umbes 4 tundi toatemperatuuril või temperatuuril 37 C või öö läbi temperatuuril 4 C), lisatakse H 2 O ja
266 264 segu filtreeritakse läbi dialüüsimembraani. Proovi saab lisamise osas analüüsida näiteks, kuid mitte ainult, protseduuridega, mida kirjeldatakse näites 3. Selles näites on PEG-il järgmine struktuur: R-PEG(N)-O-(CH 2 ) 2 -NH-C(OXCH 2 ) n -N 3 5 milles R on metüülrühm, n on 4 ja N on molekulmassiga 000. Näide 8 Selles näites kirjeldatakse FGF-21 polüpeptiidis suure hüdrofoobse aminohappe asendamist propargüültürosiiniga. Phe, Trp või Tyr jääk, mis paikneb ühes FGF-21 järgmistest piirkondadest: enne positsiooni 1 (ehk N-otsas), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (ehk valgu karboksüül-otsas) (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7) asendatakse järgmise mittelooduslikult kodeeritud aminohappega, nagu kirjeldatakse näites 7: Modifitseeritud PEG kinnitatakse alküülrühma sisaldavat aminohapet sisaldava FGF-21 polüpeptiidi variandi külge. PEG-il on järgmine struktuur: Me-PEG(N)-O-(CH 2 ) 2 -N 3
267 265 ja sobivaid sidumisprotseduure kirjeldatakse näites 7. Nende tulemusena saadakse FGF-21 polüpeptiidi variant, mis sisaldab mittelooduslikult kodeeritud aminohapet, mis on ligikaudu isosteeriline ühe suure loodusliku aminohappega ja mis on polüpeptiidis paiknevas konkreetses kohas modifitseeritud PEG derivaadiga. 5 Näide 9 Selles näites kirjeldatakse ühe või mitme PEG linkeriga seotud FGF-21 polüpeptiidi homodimeeri, heterodimeeri, homomultimeeri või heteromultimeeri loomist. Näites 7 valmistatud alküünrühma sisaldav FGF-21 polüpeptiid pannakse reageerima bifunktsionaalse PEG derivaadiga, mille valem on: N 3 -(CH 2 ) n -C(O)-NH-(CH 2 ) 2 -O-PEG(N)-O-(CH 2 ) 2 -NH-C(O)-(CH 2 ) n -N 3 milles n on 4 ja PEG-i keskmine molekulmass on umbes 5000, saades tulemusena vastava FGF-21 polüpeptiidi homodimeeri, milles kaks FGF-21 molekuli on füüsiliselt eraldatud PEG-iga. Analoogselt saab FGF-21 polüpeptiidi siduda ühe või mitme teise polüpeptiidiga ja moodustada heterodimeere, homomultimeere või heteromultimeere. Sidumisel, puhastamisel ja analüüsimisel lähtutakse näidetes 7 ja 3 kirjeldatud protseduuridest. Näide Selles näites kirjeldatakse FGF-21 polüpeptiidi sidumist sahhariidifragmendiga. Üks järgmistest aminohappejääkidest asendatakse mittelooduslikult kodeeritud aminohappega: jäägid, mis eelnevad positsioonile 1 (ehk paiknevad N-otsas), 1, 2, 3, 4, 5, 6, 7, 8, 9,, 11, 12, 13, 14,, 16, 17, 18, 19,, 21, 22, 23, 24,, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50,51, 52, 53, 54, 55, 56, 57, 58, 9, 60, 61,62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 1, 111, 112, 113, 114, 1, 116, 117, 118, 119, 1, 121, 122, 123, 124, 1, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 (ehk paiknevad valgu karboksüül-otsas) (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7), nagu kirjeldatakse näites 3.
268 266 5 Modifitseeritud FGF-21 variant, mis sisaldab karbonüülrühma sisaldavat aminohapet, pannakse reageerima N-atsetüülglükosamiini (GlcNAc) β-seoselise aminooksü-analoogiga. FGF-21 polüpeptiidi variant ( mg/ml) ja aminooksüsahhariid (21 mm) segatakse 0 mm naatriumatsetaadi puhvri vesilahuses (ph 5,5) ja inkubeeritakse 7 kuni 26 tundi temperatuuril 37 C. Selleks et siduda teine sahhariid ensümaatiliselt esimese sahhariidiga, inkubeeritakse sahhariidiga konjugeeritud FGF-21 polüpeptiidi (5 mg/ml) 48 tundi ümbritseva õhu temperatuuril UDP-galaktoosi (16 mm) ja β-1,4- galatsütosüültransferaasiga (0,4 ühikut/ml) 0 mm HEPES puhvris (ph 7,4) (Schanbacher et al. J. Biol. Chem. 1970, 245, ). Näide 11 Selles näites kirjeldatakse PEGüleeritud FGF-21 polüpeptiidi antagonisti valmistamist. Üks aminohappejääk, näiteks, kuid mitte ainult, jääk, mis osaleb FGF-21 retseptori seondumises, asendatakse järgmise mittelooduslikult kodeeritud aminohappega, nagu on kirjeldatud näites 3. Modifitseeritud FGF-21 polüpeptiidi variant, mis sisaldab karbonüülrühma sisaldavat aminohapet, pannakse reageerima aminooksürühma sisaldava PEG variandiga, mille valem on: R-PEG(N)-O-(CH 2 ) n -O-NH 2 milles R on metüülrühm, n on 4 ja N on molekulmassiga 000 MW, saades tulemusena FGF-21 polüpeptiidi antagonisti, mis sisaldab mittelooduslikult kodeeritud aminohapet, mida on polüpeptiidis ühes kohas modifitseeritud PEG derivaadiga. Sidumine, puhastamine ja analüüsimine viiakse läbi näites 3 kirjeldatud viisil.
269 Näide Otseselt seotud FGF-21 molekulidega FGF-21 polüpeptiidi homodimeeri, heterodimeeri, homomultimeeri või heteromultimeeri loomine 5 Alküünrühma sisaldava FGF-21 polüpeptiidi variandi saab siduda otse teise FGF-21 polüpeptiidi variandiga, mis sisaldab asidofragmenti. FGF-21 polüpeptiidi saab analoogselt siduda ühe või mitme polüpeptiidiga ning moodustada seeläbi heterodimeere, homomultimeere või heteromultimeere. Sidumine, puhastamine ja analüüsimine viiakse läbi näites 3, 6 ja 7 kirjeldatud viisil. Näide 13 Polüalküleenglükool (P-OH) pannakse reageerima alküülhalogeniidiga (A) ja selle tulemusel moodustub eeter (B). Sellistes ühendites on n täisarv vahemikus 1 kuni 5 ja R' võib olla sirge või hargnenud ahelaga, küllastunud või küllastumata C1 kuni C alküülvõi heteroalküülrühm. R' võib olla ka C3 kuni C7 küllastunud või küllastumata tsükliline alküül- või tsütkliline heteroalküülrühm, asendatud või asendamata arüül- või heteroarüülrühm või asendatud või asendamata alkarüül (alküülrühm on C1 kuni C küllastunud või küllastumata alküülrühm) või heteroalkarüülrühm. PEG-OH on tavaliselt polüetüleenglükool (PEG) või monometoksü-polüetüleenglükool (mpeg) molekulmassiga 800 kuni daltonit (Da). Näide 14 mpeg-oh + Br-CH 2 -C CH mpeg-o-ch 2 -C CH mpeg-oh-d molekulmassiga 000 Da (mpeg-oh kda; 2,0 g, 0,1 mmol, Sunbio) töödeldi NaH-ga (12 mg, 0,5 mmol) THF-is (35 ml). Lahusele lisati seejärel propargüülbromiid, millest oli valmistatud 80-massiprotsendiline lahus ksüleenis (0,56 ml, 5 mmol, 50 ekv., Aldrich) ja katalüütilises koguses KI-d ning saadud segu soojendati 2 tundi püstjahuti all. Siis lisati vett (1 ml) ja lahusti eemaldati vaakumiga. Jäägile lisati CH 2 Cl 2 ( ml), orgaaniline kiht eraldati, kuivatati veevabal Na 2 SO 4 ja ruumala vähendati
270 268 umbes 2 ml-ni. Selline CH 2 Cl 2 lahus lisati tilkhaaval dietüüleetrile (0 ml). Tekkinud sade koguti kokku, pesti mitme portsjoni külma dietüüleetriga ja kuivatati, saades tulemusena proargüül-o-peg-i. Näide 5 mpeg-oh + Br-(CH 2 ) 3 -C CH mpeg-o-(ch 2 ) 3 -C CH mpeg-oh-d molekulmassiga 000 Da (mpeg-oh kda; 2,0 g, 0,1 mmol, Sunbio) töödeldi NaH-ga (12 mg, 0,5 mmol) THF-is (35 ml) ja seejärel lisati segule katalüütiline kogus KI-d. Saadud segu soojendati 16 tundi püstjahuti all. Siis lisati vett (1 ml) ja lahusti eemaldati vaakumiga. Jäägile lisati CH 2 Cl 2 ( ml), orgaaniline kiht eraldati, kuivatati veevabal Na 2 SO 4 -l ja ruumala vähendati umbes 2 ml-ni. Selline CH 2 Cl 2 lahus lisati tilkhaaval dietüüleetrile (0 ml). Tekkinud sade koguti kokku, pesti mitme portsjoni külma dietüüleetriga ja kuivatati, saades tulemusena vastava alküüni. Sarnase reaktsiooni võib läbi viia ka 5-kloro-1-pentüüniga. Näide 16 (1) m-hoch 2 C 6 H 4 OH + NaOH + Br- CH2-C CH m-hoch 2 C 6 H 4 O-CH 2 -C CH (2) m-hoch 2 C 6 H 4 O-CH 2 -C CH + MsCl + N(Et) 3 m-msoch 2 C 6 H 4 O-CH 2 -C CH (3) m-msoch 2 C 6 H 4 O-CH 2 -C CH + LiBr m-br-ch 2 C 6 H 4 O-CH 2 -C CH (4) mpeg-oh + m-br-ch 2 C 6 H 4 O-CH 2 -C CH mpeg-o-ch 2 -C 6 H 4 O-CH 2 -C CH 3-hüdroksübensüülalkoholi (2,4 g, mmol) lahusele THF-is (50 ml) ja vees (2,5 ml) lisati kõigepealt pulbrilist naatriumhüdroksiidi (1,5 g, 37,5 mmol) ja seejärel propargüülbromiidi 80-massiprotsendilist lahust ksüleenis (3,36 ml, 30 mmol). Reaktsioonisegu soojendati 6 tundi püstjahuti all. Segule lisati % sidrunhapet (2,5 ml) ja lahusti eemaldati vaakumiga. Jääk ekstraheeriti etüülatsetaadiga (3 x ml) ja koondatud orgaanilisi faase pesti küllastunud NaCl lahusega ( ml), kuivatati MgSO 4 kohal ja kontsentreeriti, saades 3-propargüüloksübensüülalkoholi. Ühendi 3 lahusele (2,0 g, 11,0 mmol) CH 2 Cl 2 -es temperatuuril 0 C lisati metaansulfonüülkloriidi (2,5 g,,7 mmol) ja trietüülamiini (2,8 ml, mmol) ja
271 5 269 reaktsioonisegu asetati 16 tunniks külmkappi. Tavalise protseduuri tulemusena saadud mesülaat oli kahvatukollase õli kujul. Selline õli (2,4 g, 9,2 mmol) lahustati THF-is ( ml) ja lisati LiBr (2,0 g, 23,0 mmol). Reaktsioonisegu soojendati 1 tund püstjahuti all ja jahutati siis toatemperatuurile. Segule lisati vett (2,5 ml) ja lahusti eemaldati vaakumiga. Jääk ekstraheeriti etüülatsetaadiga (3 x ml) ja koondatud orgaanilisi faase pesti küllastunud NaCl lahusega ( ml), kuivatati veevabal Na 2 SO 4 -l ja kontsentreeriti, saades soovitud bromiidi. mpeg-oh kda (1,0 g, 0,05 mmol, Sunbio) lahustati THF-is ( ml) ja lahust jahutati jäävannis. Lahusele lisati intensiivselt segades mitme minuti jooksul NaH-d (6 mg, 0, mmol) ja seejärel varem saadud bromiidi (2,55 g, 11,4 mol) ja katalüütiline kogus KI-d. Vesivann eemaldati ja saadud segu soojendati 12 tundi püstjahuti all. Segule lisati vett (1 ml) ja lahusti eemaldati vaakumiga. Jäägile lisati CH 2 Cl 2 ( ml), orgaaniline kiht eraldati, kuivatati veevaba Na 2 SO 4 kohal ja ruumala vähendati umbes 2 ml-ni. Jääk lisati tilkhaaval eetri lahusesse (0 ml), mille tulemusel tekkis valge sade, mis koguti kokku ja saadi PEG derivaat. Näide 17 mpeg-nh 2 + X-C(O)-(CH 2 ) n -C CR mpeg-nh-c(o)-(ch 2 ) n -C CR' Terminaalset alküünrühma sisaldavaid polü(etüleenglükool) polümeere saab valmistada ka terminaalset funktsionaalrühma sisaldava polü(etüleenglükooli) polümeeri sidumise teel funktsionaalset alküünrühma sisaldava reaktsioonivõimelise molekuliga, nagu on näidatud eespool. N on 1 kuni. R' võib olla H või väike alküülrühm C1 kuni C4. Näide 18 (1) HO 2 C-(CH 2 ) 2 -C CH + NHS +DCC NHSO-C(O)-(CH 2 ) 2 -C CH (2) mpeg-nh 2 + NHSO-C(O)-(CH 2 ) 2 -C CH mpeg-nh-c(o)-(ch 2 ) 2 -C CH 4-pentüünhape (2,943 g, 3,0 mmol) lahustati CH 2 Cl 2 -es ( ml). Seejärel lisati N- hüdroksüsuktsinimiidi (3,80 g, 3,3 mmol) ja DCC-d (4,66 g, 3,0 mmol) ning lahust segati öö läbi toatemperatuuril. Saadud NHS estrit 7 kasutati töötlemata ja puhastamata kujul järgmises reaktsioonis.
272 5 270 mpeg-nh 2 molekulmassiga 5000 Da (mpeg-nh 2, 1 g, Sunbio) jahutati THF-is (50 ml) ja segu jahutati temperatuurile 4 C. Seejärel lisati intensiivselt segades osade kaupa NHS ester 7 (400 mg, 0,4 mmol). Segu segati 3 tundi ja lasti selle aja jooksul soojeneda toatemperatuurini. Segule lisati vett (2 ml) ja lahusti eemaldati vaakumiga. Jäägile lisati CH 2 Cl 2 (50 ml), orgaaniline kiht eraldati, kuivatati veevaba Na 2 SO 4 kohal ja ruumala vähendati umbes 2 ml-ni. Saadud CH 2 Cl 2 lahus lisati tilkhaaval eetrile (0 ml). Tekkinud sade koguti kokku ja kuivatati vaakumis. Näide 19 Selle näites kirjeldatakse polü(etüleenglükooli) metaansulfonüülestri ehk metaansulfonaadi ehk polü(etüleenglükool)mesülaadi valmistamist. Vastava tosülaadi ja halogeniidid saab valmistada sarnaste protseduuride abil. mpeg-oh + CH 3 SO 2 Cl + N(Et) 3 mpeg-o-so 2 CH 3 mpeg-n 3 mpeg-oh (MW = 3,400, g, mmol) 0 ml tolueenis destilleeriti aseotroopiliselt 2 tundi lämmastikus ja lahus lahutati toatemperatuurile. Lahusele lisati 40 ml kuiva CH 2 Cl 2 ja 2,1 ml kuiva trietüülamiini ( mmol). Lahust jahutati jäävannis ja sellele lisati tilkhaaval 1,2 ml destilleeritud metaansulfonüülkloriidi ( mmol). Lahust segati öö läbi toatemperatuuril lämmastikus ja reaktsiooni lõpetamiseks lisati 2 ml absoluutset etanooli. Segust eemaldati vaakumis aurustamise teel lahustid, eeskätt lahustid, mis ei ole tolueen, segu filtreeriti, kontsentreeriti uuesti vaakumis ja seejärel sadestati 0 ml dietüüleetris. Filtraati pesti mitme portsjoni dietüüleetriga ja kuivatati vaakumis, saades tulemusena mesülaadi. Mesülaat ( g, 8 mmol) lahustati 75 ml THF-is ja lahus jahutati temperatuurile 4 C. Jahutatud lahusele lisati naatriumasiidi (1,56 g, 24 mmol). Reaktsioonisegu soojendati 2 tundi lämmastikus püstjahuti all. Seejärel eemaldati aurustamise teel lahustid ja jääki lahjendati CH 2 Cl 2 -ega (50 ml). Orgaanilist faasi pesti NaCl lahusega ja kuivatati veevaba MgSO 4 kohal. Ruumala vähendati ml-ni ja lisati saaduse sadestamiseks 0 ml külma kuiva eetrit. Näide (1) N 3 -C 6 H 4 -CO 2 H N 3 -C 6 H 4 CH 2 OH
273 271 (2) N 3 -C 6 H 4 CH 2 OH Br-CH 2 -C 6 H 4 -N 3 (3) mpeg-oh + Br-CH 2 -C 6 H 4 -N 3 mpeg-o-ch 2 -C 6 H 4 -N asidobensüülalkoholi saab valmistada meetodil, mida kirjeldatakse USA patendis nr asidobensüülalkoholi (1,75 g, 11,0 mmol) lahusele CH 2 Cl 2 -es temperatuuril 0 C lisati metaansulfonüülkloriidi (2,5 g,,7 mmol) ja trietüülamiini (2,8 ml, mmol) ja reaktsioonisegu asetati 16 tunniks külmkappi. Tavalise work-up i tulemusena saadud mesülaat oli kahvatukollase õli kujul. See õli (9,2 mmol) lahustati THF-is ( ml) ja lisati LiBr (2,0 g, 23,0 mmol). Reaktsioonisegu soojebdati 1 tund püstjahuti all ja seejärel jahutati toatemperatuurile. Segule lisati vett (2,5 ml) ja lahusti eemaldati vaakumiga. Jääk ekstraheeriti etüülatsetaadiga (3 x ml) ja koondatud orgaanilisi faase pesti küllastunud NaCl lahusega ( ml), kuivatati veevaba Na 2 SO 4 kohal ja kontsentreeriti, saades soovitud bromiidi. mpeg-oh kda (2,0 g, 0,1 mmol, Sunbio) töödeldi NaH-ga (12 mg, 0,5 mmol) THF-is (35 ml) ja segule lisati bromiidi (3,32 g, mmol) ja katalüütilises koguses KI-d. Saadud segu soojendati 12 tundi püstjahuti all. Segule lisati vett ja lahusti eemaldati vaakumiga. Jäägile lisati CH 2 Cl 2 ( ml), orgaaniline faas eemaldati, kuivatati veevaba Na 2 SO 4 kohal ja ruumala vähendati umbes 2 ml-ni. Jääk lisati tilkhaaval eetri lahusesse (0 ml), mille tulemusel tekkis valge sade, mis koguti kokku ja saadi mpeg-o-ch 2 -C 6 H 4 -N 3. Näide 21 NH 2 -PEG-O-CH 2 CH 2 CO 2 H + N 3 -CH 2 CH 2 CO 2 -NHS N 3 -CH 2 CH 2 -C(O)NH-PEG-O- CH 2 CH 2 CO 2 H NH 2 -PEG-O-CH 2 CH 2 CO 2 H (molekulmass 3400 Da, 2,0 g) lahustati NaHCO 3 küllastunud vesilahuses ( ml) ja lahus jahutati temperatuurile 0 C. Seejärel lisati intensiivselt segades 3-asido-1-N-hüdroksüsuktsinimidopropionaati (5 ekv.) 3 tunni pärast lisati ml H 2 O-d ja segu segati veel 45 minutit toatemperatuuril. PH reguleeriti 0,5 N H 2 SO 4 -ga väärtusele 3 ja lisati NaCl-i kontsentratsioonini umbes massiprotsenti. Reaktsioonisegu ekstraheeriti CH 2 Cl 2 (0 ml x 3), kuivatati Na 2 SO 4 kohal ja kontsentreeriti. Pärast külmas dietüüleetris sadestamist koguti saadus filtreerimise teel kokku ja kuivatati vaakumis, saades tulemusena oomega-karboksü-asiid PEG derivaadi.
274 Näide mpeg-oms + HC CLi mpeg-o-ch 2 -CH 2 -C C-H 5 Liitiumtatsetüliidi lahusele (4 ekv), mis valmistati tehnika tasemes tuntud meetodil ja jahutati THF-is temperatuurile -78 C, lisati intensiivselt segades tilkhaaval mpeg-oms lahus THF-is. Kolme tunni pärast lasti lahusel soojeneda toatemperatuurile ja selle lisati reaktsiooni lõpetamiseks 1 ml butanooli. Siis lisati ml H 2 O-d ja segu segati veel 45 minutit toatemperatuuril. ph reguleeriti 0,5 N H 2 SO 4 lisamise teel väärtusele 3 ja lisati NaCl kontsentratsioonini umbes massiprotsenti. Reaktsioonisegu ekstraheeriti CH 2 Cl 2 - ga (0 ml x 3), kuivatati Na 2 SO 4 kohal ja kontsentreeriti. Pärast külmas dietüüleetris sadestamist koguti saadus filtreerimise teel kokku ja kuivatati vaakumis, saades tulemusena 1-(but-3-ünüüloksü)-metoksüpolüetüleenglükooli (mpeg). Näide 23 Asiid- ja atsetüleenrühma sisaldavaid aminohappeid saab valkudesse kohtselektiivselt sisestada meetoditel, mida kirjeldavad L. Wang, et al., (01), Science 292: , J.W. Chin et al., Science 301:964-7 (03)), J. W. Chin et al., (02), Journal of the American Chemical Society 124: ; J. W. Chin, & P. G. Schultz, (02), Chem Bio Chem 3(11): ; J. W. Chin, et al., (02), PNAS United States of America 99:1-124: and, L. Wang, & P. G. Schultz, (02), Chem. Comm., 1:1-11. Pärast aminohapete sisestamist viiakse 0,01 mm valguga fosfaatpuhvris (PB), ph 8, 2 mm PEG derivaadi, 1 mm CuSO 4 ja ~1 mg vasktraadi manulusel 4 tunni jooksul temperatuuril läbi 37 C tsükloliitumise reaktsioon. Näide 24 Selles näites kirjeldatakse p-atsetüül-d,l-fenüülalaniini (paf) ja m-peghüdroksüülamiini derivaatide sünteesi. Ratseemilist paf-i sünteesitakse protseduuriga, mida kirjeldavad Zhang, Z., Smith, B. A. C., Wang, L., Brock, A., Cho, C. & Schultz, P. G., Biochemistry, (03) 42, m-peg-hüdroksüülamiini derivaadi sünteesimiseks viiakse läbi järgnev protseduur. (N-t-Boc-aminooksü)äädikhappe (0,382 g, 2,0 mmol) ja 1,3-diisopropüülkarbodiimiidi (0,16 ml, 1,0 mmol) lahusele diklorometaanis (DCM, 70 ml), mida segatakse 1 tund toatemperatuuril, lisatakse metoksü-
275 273 polüetüleenglükoolamiini (m-peg-nh 2, 7,5 g, 0, mmol, Mt. 30 K, ettevõttelt BioVectra) ja diisopropüületüülamiini (0,1 ml, 0,5 mmol). Reaktsioonisegu segatakse 48 tundi toatemperatuuril ja seejärel kontsentreeritakse ruumalani umbes 0 ml. Segu lisatakse tilkhaaval külmale eetrile (800 ml). 5 t-boc-kaitstud saadus sadestatakse, kogutakse filtreerimise teel kokku ja pestakse 3 x 0 ml eetriga. Saaduse täiendavaks puhastamiseks lahustatakse see uuesti DCM-is (0 ml) ja sadestatakse kaks korda eetris (800 ml). Saadust kuivatatakse vaakumis ja saadakse 7,2 g (96%) saadust, mida kinnitavad NMR ja Nihydrin analüüsid. Eespool valmistatud kaitstud saaduse (7,0 g) deboc iga töötlemine viiakse läbi 1 tunni jooksul temperatuuril 0 C ja seejärel 1,5 jooksul toatemperatuuril 50% TFA/DCM-is (40 ml). Siis eemaldatakse vaakumiga enamik TFA-st ja jäägile lisatakse 4N HCl dioksaanis (1 ml), mille tulemusel muundub hüdroksüülamiini derivaat HCl soolaks. Sade lahustatakse DCM-is (50 ml) ja sadestatakse uuesti eetris (800 ml). Lõppsaadus (6,8 g, 97%) kogutakse filtreerimise teel kokku, seda pestakse 3x 0 ml eetriga, kuivatatakse vaakumis ja hoitakse lämmastikus. Teisi PEG-i (5K, K) hüdroksüülamiinderivaate sünteesitakse sama protseduuri abil. Näide FGF-21 WT ja 30K PEG analoogide indutseeritud ERK1/2 fosforüülimise analüüs: rakud, mida on stabiilselt transfekteeritud inimese Klotho beeta valguga, külvatakse koguses rakku süvendi kohta (DMEM+% FBS) poly-lys kaetud plaadile. Järgmisel päeval on rakud 0% konfluentsed, sööde eemaldatakse aspireerimise teel, rakkudele lisatakse värsket söödet ja neid inkubeeritakse öö läbi. 24 tunni pärast stimuleeritakse rakke sobivate 30K PEG FGF-21 analoogidega, kasutades selleks standardset FGF21WT. Iga eri ühendi ettevalmistamiseks lahjendatakse neid PBS/1%BSA-s. Rakke töödeldakse kolme paralleelproovina inkubaatoris temperatuuril 37 C. Pärast -minutilist inkubeerimist eemaldatakse aspireerimise teel igast süvendist hoolikalt sööde ja igasse süvendisse lisatakse rakulüsaatide valmistamiseks 1 x 40 ul külma puhvrit nimega Signaling Lysis Buffer, mis sisaldab proteaasi/fosfataasi inhibiitoreid (PI kokteil, Na3VN4 ja PMSF). 96 süvendiga plaat asetatakse minutiks jääle ja tsentrifuugitakse siis minutit kiirusel 4000 pööret minutis. Rakulüsaadid külmutatakse temperatuurini -80 C. Hiljem sulatatakse iga proov ja ul rakulüsaate
276 5 274 lisatakse MSD-ga töödeldud plaadile, mis on kaetud antikehaga, mis püüab nii ERK1/2 fosforüülimata kui ka fosforüülitud vorme. Plaati inkubeeritakse 2 tundi primaarse antikehaga ja pestakse siis mitu korda spetsiifilise puhvriga ja seejärel lisatakse sekundaarne antikeha. Pärast 1-tunnilist inkubeerimist pestakse plaati veel mitu korda. Igasse süvendisse lisatakse lugemiseks puhvrit. Plaat asetatakse MSD lugemisseadmesse. Tekkinud kõvera aluseks on fosforüülimata ERK1/2 numbrid ja EC50 arvutatakse Sigma Plot abil. Selleks et arvutada aktiivsuse vähenemine kordades jagatakse 30K PEGüleeritud spetsiifilise ühendi EC50 väärtus WT EC50 väärtusega. Näide 26: Rakkude ERK 1/2 fosforüülimisanalüüsi (perk) protokoll ja MSD analüüs 293 βklotho-4 rakke hoiti DMEM+% FBS+P/S+0,5 mg/ml genetitsiinis. Kui rakud saavutasid 50-90% konfluentsuse, töödeldi neid trüpsiiniga ja külvati koguses rakku süvendi kohta DMEM+%FBS+P/S-is poly-d-lys kaetud 96 süvendiga plaatidele. Järgmisel päeval ehk siis, kui rakud olid ~0% konfluentsed, kontrolliti neid, et olla kindel, et sööde on ikka punane, seejärel eemaldati aspireerimise teel sööde ja 96 süvendiga plaadile kanti pipetiga FGF-21 variantide (PBS+1%BSA-s) seerialahjendused koguses 0 ul süvendi kohta. Seejärel asendati 96 süvendiga plaat täpselt 9 minutiks 5% CO2 inkubaatorisse temperatuuril 37 C. Kõik FGF-21 töötlusained eemaldati seejärel aspireerimise teel ja igasse süvendisse lisati 40 ul äsja valmistatud 1xCell Signaling Lysis Buffer + 1x Sigma toodet Protease Inhibitor Cocktail + 2 mm naatriumortovanadaati + 1mM PMSF + 1x MSD fosfataasi inhibiitorit I +1x MSD fosfataasi inhibiitorit II + 1x MSD toodet Protease Inhibitor Cocktail + 2 mm MSD PMSF. Nõu asetati jääle ja hoiti seal minutit, mille jooksul töödeldi kõiki süvendeid lüsaatide segamiseks P pipettoriga. Pärast süvendite segamist asetati jää-ämber ja nõu ülejäänud ajaks loksutisse. minuti pärast tsentrifuugiti plaati minutit 4C loksutis ja supernatandid asetati külmal ümarapõhjalisel 96 süvendiga plaadil jääle. MSD analüüs See analüüs tehti süsteemiga Meso Scale Discovery MULTI-SPOT Assay System wholecell lysate kit Phospho (T/Y 2/4; 185/187)/Total - ERK1/2/ Assay.
277 5 275 Kõik MSD reagendid sulatati toatemperatuurile ja kõik vajalikud puhvrid valmistati vastava komplekti juhiste järgi. PhosphoERK-total ERK Duplex plaadi igasse süvendisse lisati 0 ul toodet Blocker A ja plaati blokeeriti 1 tund toatemperatuuril loksutis. Plaati pesti seejärel 4x with 160ul süvendi kohta 1x pesupuhvriga. Siis lisati igasse süvendisse 16 ul lüüsipuhvrit + proteaasi ja fosfataasi inhibiitoreid (mis valmistati varem rakkude stimuleerimiseks) ja ul materjali süvendi kohta kanti külmalt lüsaadi supernatandi plaadilt üle MSD plaatidele (üldruumala oli nüüd 26 ul süvendi kohta). MSD plaati inkubeeriti 3 tundi toatemperatuuril loksutis, plaati pesti 4x 160 ul süvendi kohta 1x pesupuhvriga. Seejärel lisati igasse süvendisse ul detekteerimisantikeha (mida oli lahjendatud 50 korda antikeha lahjenduspuhvris) ja inkubeeriti 1 tund toatemperatuuril loksutis. Plaati pesti uuesti 4x 160 ul/süvendi kohta 1x pesupuhvriga, igasse süvendisse lisati 0 ul 1x toodet Read Buffer T ja plaati loeti kohe seadmel MSD Sector Imager BCA kvantiteerimine Kvantiteerimiseks kasutati komplekti Pierce BCA Protein Assay Kit. BSA standard lahjendati 96 süvendiga plaadil 1xCell Signaling Lysis Buffer is maksimaalselt kontsentratsioonilt 2 mg/ml 2 korda lahjemaks (viimastes süvendites oli puhver, mitte BSA). BSA standardid kanti pipetiga koguses ul süvendi kohta kahe paralleelproovina kahele MaxiSorp 96 süvendiga plaadile. Lüsaate lahjendati kolm korda 1xCell Signaling Lysis Buffer puhvris ja kanti koguses ul süvendi kohta MaxiSorp plaatidele. Tööreagent valmistati Pierce Kit kasutusjuhendi järgi ja MaxiSorp plaatide igasse süvendisse kanti pipetiga 0 ul tööreagenti. Plaate inkubeeriti toatemperatuuril ja loeti plaadilugejaga väärtusel λ=562 nm. Andmete analüüsimine 30 Kõikide lüsaatide puhul arvutati kontsentratsioonid komplektiga BCA Kit. Kui kõikide lüsaatide kontsentratsioonid on sarnased, ei normaliseerita neid MSD analüüsiks. MSD analüüsi jaoks keskmistatakse replikatsiooni alguspunktid ja arvutatakse standardhälve ja CV väärtused. SigmaPlot abil arvutatakse FGF-21 variantide seerialahjenduste EC50 väärtused ja variandid seatakse Fold Above WT EC50 järgi pingeritta
278 276 Tulemused on toodud käesoleva taotlse joonistel FIG 7a ja FIG 7b Näide 27: Märgistamata FGF-21 allavooluprotsess Inklusioonikehade preparaadi solubiliseerimine 5 Rakupasta uuesti suspendeerimiseks segati see 4 C inklusioonikehade (IB) puhvris I (50mM Tris ph 8,0; 0 mm NaCl; 1 mm EDTA; 1% Triton X-0; 4 C) tahke lõpliku kontsentratsioonini %. Rakkude lüüsimiseks passeeriti uuesti suspendeeritud materjal kokku kaks korda läbi mikrofluidisaatori, seejärel tsentrifuugiti ( 000 g; min; 4 C) ja supernatant dekanteeriti. IB pelletit pesti uuesti suspendeerimise teel lisakoguses IB puhvris I (50 mm Tris ph 8,0; 0 mm NaCl; 1 mm EDTA; 1% Triton X-0; 4 C) ja suspendeeritud materjal passeeriti kokku kaks korda mikrofluidisaatoris, seejärel tsentrifuugiti ( 000g; min; 4 C) ja supernatant dekanteeriti. IB pellet suspendeeriti uuesti ühes ruumalas puhvris II (50 mm Tris ph 8,0; 0 mm NaCl; 1 mm EDTA; 4 C), seejärel tsentrifuugiti ( 000 g; min; 4 C) ja supernatant dekanteeriti. IB pellet suspendeeriti uuesti ½ ruumalas puhvris II (50 mm Tris ph 8,0; 0 mm NaCl; 1 mm EDTA; 4 C). IB kanti alikvootidena sobivatesse mahutitesse, tsentrifuugiti ( 000 g; min; 4 C) ja supernatant dekanteeriti. Inklusioonikehad lahustati (vastasel juhul oleks neid hoitud edasise kasutamiseni temperatuuril -80 C). Inklusioonikehade solubiliseerimine Inklusioonikehad solubiliseeriti solubiliseeerimispuhvris lõppkontsentratsioonini vahemikus - mg/ml ( mm Tris, ph 8,0; 8M uurea; mm β-me) ja solubiliseeritud inklusioonikehi inkubeeriti pidevalt segades 1 tund toatemperatuuril. Lahustumatu materjal eemaldati filtreerimise teel (0,45 µm PES filtriga) ja valgu kontsentratsiooni reguleeriti (see ei ole alati vajalik) teise solubiliseerimispuhvriga (kui valgu kontsentratsioon on suur) lahjendamise teel. Kokkuvoltimine Uuesti kokku voltimiseks lahjendatakse materjali mm Tris valgu lõpliku kontsentratsioonini
279 277 0,5 mg/ml, mm Tris, ph 8,0; 4 C. Valgul lastakse 18 kuni 24 tundi temperatuuril 4 C end kokku voltida. Puhastamine 5 Kokkuvolditav valguga reaktsioonisegu filtreeritakse läbi 0,45 µm PES filtri. Materjal kantakse Q HP kolonnile (GE Healthcare), mida on tasakaalustatud tootega Buffer A ( mm Tris, ph 7,5). FGF-21 elueeritakse lineaarse gradiendiga üle CV 0% Buffer B- ni (mmtris, ph 7.5; 0mM NaCl). Monomeerne FGF-21 kogutakse kokku. PEGüleerimine ja puhastamine Võetakse Q HP kogum ja puhver asendatakse puhvriga mm Tris, ph 8,0; 2M uureat; 1mM EDTA-t. ph langetamiseks väärtusele 4,0 listakse 50% jää-äädikhapet. Proov kontsentreeritakse tasemele 4,0 ± 1,0 mg/ml. Seejärel lisatakse proovile 12:1 molaarse liiaga PEG-i ja lõppkontsentratsiooniga 1% äädikhappe hüdrasiidi, ph 4,0. Materjali inkubeeritakse tundi temperatuuril 28 C. PEGi reaktsioonisegule lisatakse 50 mm Tris alust ja segu lahjendatakse korda RO veega. Juhtivus peab olema <1 ms/cm ja ph 8,0-9,0. Materjal kantakse Source 30Q kolonnile (GE Healthcare), mida on tasakaalustatud tootega Buffer A ( mm Tris, ph 8,0). PEG-FGF-21 elueeritakse lineaarse gradiendiga üle CV to 0% Puhver B-ni ( mm Tris, ph 8,0; 0 mm NaCl). PEG-FGF-21 kogutakse kokku ja puhver asendatakse mm Tris, ph 7,4; 0 mm NaCl. PEG materjali kontsentreeritakse kontsentratsioonini 1-2 mg/ml ja filtersteriliseeritakse 0,22 µm PES filtriga. Materjali hoitakse temperatuuril 4 C. Pikemaajalise säilitamise korral materjal kuivatakse kiirmeetodil ja seda hoitakse temperatuuril -80 C. Näide 28 FGF-21 farmakokineetilised omadused rottide organismis Selle protokolli abil saadi andmeid (mis on toodud joonistel FIG 11-23) ettevõtte Ambrx patenteeritud tehnoloogia abil valmistatud natiivsete ja PEG-iga modifitseeritud FGF-21 ühendite farmakokineetiliste omaduste kohta kateetriga rottidel. Katseobjektide
280 278 farmakokineetikat analüüsiti inimese FGF-21 suhtes spetsiifilise ELISA analüüsiga seerumiproovide, mis võeti kindlatel ajahetkedel pärast ravimi manustamist. Katseobjektid: 1. Ambrx ühend PEG-R77 FGF-21 kontsentratsiooniga 0. mg/ml, lahjendatud 1X PBS Ambrx ühend PEG-Y4 FGF-21 kontsentratsiooniga 0. mg/ml lahjendatud 1X PBS. 3. Ambrx ühend PEG-R126 FGF-21 kontsentratsiooniga 0. mg/ml lahjendatud 1X PBS. Katseobjekti kvaliteet / preparaat: Lähtelahuste kontsentratsioonid = 1,0 mg/ml PEG-R77 FGF-21 1,16 mg/ml PEG-Y4 FGF-21 Loomad: 1,08 mg/ml PEG-R126 FGF-21 Kaheteistkümnele (12) isasele Sprague-Dawley (SD) rotile, kelle kehakaal oli uuringu alguses grammi, paigaldati enne ettevõttesse Ambrx saabumist kägiveeni kirurgilise protseduuri abil kateetrid. Ettevõttest CRL saadud loomad on heas seisukorras ja neil lasti 3 päeva enne uuringu algust uuringukohas kohaneda. Rotte kaaluti katseobjekti manustamise päeval. Loomi hoiti standardsetes patogeenivabades tingimustes ja neile anti piiramatus koguses süüa ja juua. Loomarühmad: Kõiki ühendeid manustati subkutaanselt 1. rühm (n=4): PEG-R77 SC süstimine (0, mg/kg). 2. rühm (n=4): PEG-Y4 SC süstimine (0, mg/kg). 3. rühm (n=4): PEG-R126 SC süstimine (0, mg/kg).
281 5 279 Loomi kaaluti enne katseobjekti manustamist. Ühenditest valmistati preparaadid, mida sai manustada 1X BW, ühikutes µl. Katseobjektid süstiti subkutaanselt selja abaluupiirkonda. Loomadele süstiti katseobjekti üks kord (ajahetk =0). Loomadelt võetakse kindlatel ajahetkedel täisvereproovid ja koguti need SST Microtainer Collection tuubidesse. Seerumil lasti enne tsentrifuugimist 30 minutit koaguleeruda. Seerum pandi prolüpropüleenist tiitertuubidesse, need suleti mikroribadega ja hoiti ELISA-meetodil analüüsimiseni ja selle tulemusel FGF-21 seerumi kontsentratsiooni määramiseni temperatuuril -800 C Andmete kogumine / tulemusnäitaja: Iga looma kasutatakse kogu farmakokineetilise uuringu jooksul. Loomalt võetakse kägiveeni kateetrist 0, ml täisverd. Kohe pärast vere võtmist loputatakse kateetreid 0,1 ml soolalahusega. Katseobjektide eeldatava farmakokineetilise profiili põhjal tuleb loomadelt võtta vereproovid järgmistel ajahetkedel: Enne manustamist, 1, 2, 4, 8, 24, 32 48, 56, 72 ja 96 tundi pärast manustamist. Surmamine: Kõik loomad surmatakse pärast uuringu läbiviimist. Tulemused: Tulemused on toodud alljärgnevas tabelis ja käesoleva taotlusega kaasasolevatel joonistel FGF-21 PEG isomeeri PK omadused-iv PEG isomeer 30K Intravenoosselt 0, mg/kg R72 R77 H87 E91 Y4 E1 R126 P146 Lambda_z 1/h 0,057 0,043 0,044 0,052 Lambda_z_lower h
282 280 PEG isomeer 30K Intravenoosselt 0, mg/kg R72 R77 H87 E91 Y4 E1 R126 P146 Lambda_z_upper h HL_Lambda_z h 12,27 16,44,61 13,67 Tmax h 0, 0, 0, 0,31 Cmax ng/ml 5998,6 5802,3 7821,4 8655,8 C0 ng/ml 6861,4 6662,5 9280,1 9149,9 AUCINF_obs h*ng/ml 53714, , , ,6 Vz_obs ml/kg 82,46 113, 81,65 58,06 Cl_obs ml/h/kg 4,65 4,74 3,61 2,92 MRTINF_obs h 14,49 13,81 16,18 14,83 Vss_obs ml/kg 67,46 65,53 58,53 43,48 FGF21 PEG isomeeri farmakokineetilised omadused SC PEG 30K Isomeer Subkutaanselt 0,1 mp/kg R72 R77 H87 E91 Y4 E1 R126 P146
283 281 PEG 30K Isomeer Subkutaanselt 0,1 mp/kg R72 R77 H87 E91 Y4 E1 R126 P146 Lambda_z 1/h 0,043 0,035 0,0317 Lambda_z_alumine h Lambda_z_ülemine h HL_Lambda_z h 14,72 16,14 19,93 22,01 Tmax h Cmax ng/ml 4,5 174,3 229,7 321,3 AUCINF_obs h*ng/ml 86, ,2 908,4 Vz_obs ml/kg 458,9 731,1 606,2 503,4 Cl_obs ml/h/kg 21,92 31,67,91,86 MRTINF_obs h 36,52 36,31 40,35 40,07 PEGüleeritud ühendite suurenenud Tmax PEGüleeritud ühendite suurenenud T1/2 PEGüleeritud ühendite suurenenud AUC PEGüleerimiskohast sõltuvad farmakokineetilised näitajad
284 282 Näide 29 PEGüleeritud FGF-21 in vivo uuringud 5 PEG-FGF-21, modifitseerimata FGF-21 ja puhverlahus manustatakse hiirtele või rottidele. Tulemused näitavad, et leiutisekohasel PEGüleeritud FGF-21 polüpeptiidil on võrreldes modifitseerimata FGF-21 polüpeptiidiga suurem aktiivsus ja pikem poolväärtusaeg. Hiirtele või rottidele manustatakse ka modifitseeritud FGF-21, modifitseerimata FGF-21 ja puhverlahust. Farmakokineetiline analüüs Patendidokumendis WO 05/ kirjeldatakse farmakokineetilisi uuringuid, mida saab läbi viia käesoleva leiutise kohaste FGF-21 ühenditega. Leiutisekohast FGF-21 polüpeptiidi manustatakse intravenoosselt või subkutaanselt hiirtele. Hiirtelt võetakse enne manustamist ja kindlatel manustamisjärgsetel ajahetkedel vereproovid. Igast proovist eraldatakse plasma ja seda uuritakse radioimmunoanalüüsiga. Sellise analüüsi puhul saab arvutada leiutisekohaste mittelooduslikult kodeeritud aminohapet sisaldavate FGF-21 polüpeptiidide ja metsikut tüüpi FGF-21 polüpeptiidide või FGF-21 polüpeptiidide elimininatsiooni poolväärtusaja ja võrrelda arvutatud väärtusi omavahel. Leiutisekohaseid FGF-21 polüpeptiidi võib sarnaselt manustada ka Cynomolgus ahvidele. Ahvidelt võetakse enne manustamist ja kindlatel manustamisjärgsetel ajahetkedel vereproovid. Igast proovist eraldatakse plasma ja seda analüüsitakse radioimmunoanalüüsiga. Leiutisekohaseid polüpeptiide võib manustada ZDF isastele rottidele (diabeeti põdevad rasvunud rotid, kes on uuringu alguses 8-nädalased, Charles River-GMI). Rottidele antakse piiramatus koguses Purina 5008 loomatoitu. Moodustatakse järgmised katserühmad: soolalahust; insuliini 4U päevas; FGF-21; 500 ug päevas akuutne (akuutsele katserühmale manustatakse üks annus ja võetakse vereproov ajahetkedel T=0 ja 2, 4, 8 ja 24 tundi pärast manustamist); FGF-21, 0 ug päevas; FGF- 21, 0 ug päevas; FGF-21, 500 ug päevas; FGF-21 (üks kord päevas) 500 ug/ml; Lean Saline; Lean insuliin 4U päevas; Lean FGF ug päevas. Lean rühmades on saledad ZDF rotid, kes ei põe diabeeti. Ühendeid süstitakse subkutaanselt (b.i.d), välja arvatud teisele rühmale, kellele manustatakse 500 ug päevas: see kogus süstitakse neile
285 5 283 uuringuperioodi (7 päeva jooksul) üks kord päevas. Kontrollrottidele süstitakse vehiiklit (PBS; 0,1 ml). 7 päeva pärast manustamist tehakse loomadel suukaudse glükoosi taluvuse analüüs. Glükoosi ja triglütseriidide sisalduse määramiseks vajalik veri võetakse loomadelt sabast ilma anesteetikumita. FGF-21 polüpeptiididel on võime vähendada plasma glükoositaset annusest sõltuvalt. Saledatel ZDF rottidel ei tarvitse pärast kokkupuudet leiutisekohaste FGF-21 polüpeptiididega tekkida võrreldes insuliini saavate rottidega hüpoglükeemiat. ob/ob rasvtõve mudel ob/ob hiiremudel on hüperglükeemia, insuliiniresistentsuse ja rasvtõve loommudel. ob/ob hiirtel võib mõõta plasma glükoositaset pärast FGF-21 polüpeptiidi manustamist ja võrrelda seda vehiiklit ja insuliini saavates kontrollrühmades mõõdetud vastavate näitajatega. Selles rasvtõve mudelis süstitakse isastest ob/ob hiirtest (7-nädalased) koosnevatele katserühmadele 7 päeva subkutaanselt (0,1 ml, b.i.d) ainult vehiiklit (PBS), insuliini (4 ühikut päevas) või FGF-21 polüpeptiidi (5 µg päevas ja µg päevas). Loomadelt võetakse vereproovid saba kaudu päevadel 1, 3 ja 7 üks tund pärast esimese ühendi süstimist ja proovides mõõdetakse standardprotokolli abil plasma glükoositaset. Kui leiutisekohased FGF-21 polüpeptiidid langetavad võrreldes vehiiklit saava kontrollrühmaga plasma glükoositaset, stimuleerivad nad glükoosi haaret. Triglütseriidide taset võib võrrelda pärast leiutisekohaste FGF-21 polüpeptiidide manustamist võrreldes teiste molekulidega. Kõnealust polüpeptiidi võib manustada hiirtele mitme annusena, pideva infusioonina või ühe annusena, jne. Näide FGF21 analoogide farmakokineetika hindamine: Erinevate PEG-i konjugeerimiskohtadega 30KPEG-pAF(N6-His)FGF21 analoogide farmakokineetilisi omadusi hinnati roti organismis. Teisteks uuritavateks parameetriteks olid PEG-i molekulmass ja ka manustatava ühendi annus. Lisaks määrati mõne 30KPEG-pAF(N6- His)FGF21 variandi biosaadavuse protsent. Loomad: Kõikide loomkatsete tegemisel järgiti asutuse nimega Institutional Animal Care and Use Committee heakskiidetud protokolle. Isased Sprague-Dawley rotid ( g) saadi ettevõttelt Charles River Laboratories. Rotte hoiti ühekaupa puurides ruumides, kus
286 284 rakendati 12-tunnist valguse/pimeduse tsüklit ja neil lasti enne katse algust vähemalt 3 päeva Ambrx vivaariumis kohaneda. Loomadel oli võimalus süüa piiratamatus koguses sertifitseeritud Purina närilistetoitu ja juua vett. Manustamine ja seerumiproovide võtmine: 5 30 Ettevõttes CRL paigaldati loomadele enne tarnimist vereproovide võtmiseks kirurgiliselt kägiveeni kateetrid. Pärast kateetrite edukat avamist jagati loomad enne ühendite manustamist katserühmadesse. Ühe annusena manustatavat ühendit manustati intravenoosselt või subkutaanselt 1 ml/kg suuruse annusena. Ühendi annuste kontsentratsioonid saadi PBS-is lahjendamise teel, kautades lähtekontsentratsioonina dokumendis Certificate of Release määratud kontsentratsiooni. Loomadelt võeti erinevatel ajahetkedel püsikateetri kaudu vereproovid ja need pandi SST tuubidesse. Pärast tsentrifuugimist koguti seerum kokku ja seda hoiti analüüsimiseni temperatuuril -80 C. Farmakokineetiline analüüs: Analüüs, mille abil saab kvantiteerida ühendit PEG-FGF-21 Sprague- Dawley rottide seerumis, töötati välja ettevõttes Ambrx Inc., La Jolla, CA. Mikroplaadi süvendid kaetakse kitse inimesevastase FGF-21 IgG polüklonaalse antikehaga (PAb; RnD Systems, kloon AF39), mis täidab püüdmisreagendi (capture reagent) ülesannet. Standardi (STD) ja kvaliteedikontrolliks kasutatavaid (QC) proove, millest mõlemad on valmistatud 0% Sprague Dawley roti seerumi spaikimise teel PEG- FGF-21 analoogiga, ja uuritavaid proove töödeldakse vahekorras 1:0 I-Block puhvriga ja kantakse siis süvenditesse. STDs, QC ja uuritavates proovides sisalduv FGF-21 püütakse kinni immobiliseeritud polüklonaalse antikehaga. Seondumata materjal eemaldatakse süvendite pesemise teel. Seejärel lisatakse süvenditesse biotiini kitse inimesevastast FGF-21 IgG polüklonaalset antikeha (RnD Systems, kloon BAF39), seejärel pestakse süvendeid ja lisatakse kinnipüütud PEG-FGF-21 detekteerimiseks streptavidiini mädarõika peroksüdaasi (SA-HRP; RnD Systems, Catalog # DY998). Pärast veel ühte pesemisetappi lisatakse süvenditesse tetrametüülbensidiini (TMB, Kirkegaard Perry Laboratories) substraadi lahust. TMB reageerib HRP juuresolekul peroksiidiga ja selle tulemusel tekib kolorimeetriline signaal, mis on proportsionaalne esimeses etapis capture reagendiga seondunud PEG-FGF-21 analoogi kogusega. Värvi tekke peatamiseks lisatakse 2N väävelhapet ja mõõdetakse värvi intensiivsust (optiline tihedus, OD) lainepikkusel 450 nm. Uuritavate proovide ja QC proovide OD ühikud teisendatakse
287 285 kontsentratsiooniühikuteks arvutitarkvara abil tehtava võrdluse teel sama plaadi standardkõveraga, mida regresseeritakse SOFTmax Pro v5 Data Reduction Package abil 5 parameetriga logistilise regressiooni mudeli järgi. Tulemused esitatakse kontsentratsiooniühikutes ng/ml. 5 Kontsentratsiooni saab mõõta ka kihtehnikal põhineva kaudse ELISA meetodil (double antibody sandwich assay) või teistel asjatundjatele tuntud meetoditel. Kontsentratsioonid arvutati vastava manustatud ühendi puhul saadud standardkõvera järgi. Farmakokineetilisi parameetreid hinnati modelleerimisprogrammiga (Pharsight, version 4.1). Hindamisel kasutati eri loomade andmete mittekompartmentaliseeritud analüüsi linear-up/log-down trapetsintegreerimisega ja kontsentratsiooniandmed ühtlustati. Kokkuvõte: WT N6-His FGF21 farmakokineetilised omadused vastasid Kharitonenkov et al, 05 poolt avaldatud andmetele ja olid võrreldavad märgiseta WT FGF21 valgu omadustega. 30KPEG-pAF(N6-His)FGF21 isomeeride farmakokineetilised profiilid paranesid 30 kda PEG molekuli lisamisel võrreldes WT (PEGüleerimata) FGF21 puhul saadud tulemustega oluliselt. PEGüleeritud ühendite farmakineetilised profiilid olid märgatavalt teistsugused, kui neid ühendeid manustati subkutaanselt annuses 0, mg/kg. Ühenditel H87 ja L86 olid võrreldes teiste isomeeridega kehvemad farmakokineetilised näitajad. R131 ja Q8 PEG30 ühendite puhul saadi diferentsiaalselt paremad farmakokineetilised omadused. Nimetatud ühenditel oli täpsemalt parem AUC, Cmax ja terminaalne poolväärtusaeg. Struktuuri ja aktiivsuse süvaanalüüsi teel võib olla võimalik määrata struktuuriga seotud iseärasused, mille tõttu on igal isomeeril erinevad farmakokineetilised omadused. PEG-i molekulmasside võrdlus näitas, et 30KPEGpAF91(N6-His)FGF21 püsib vereringes mõnevõrra paremini kui KPEG-pAF91(N6- His)FGF21. Isomeeril molekulmassiga kda oli suurem Cmax väärtus, kuid kahe ühendi kogu AUCinf oli võrreldav. kda isovormi biosaadavus oli mõnevõrra parem kui 30 kda variandi biosaadavus (vastavalt 30% ja %)
288 286 Tabel 3. Rottidele 0, mg/kg suuruse annusena manustatud ühendite farmakokineetilised parameetrid 0, mg/kg SC Terminaalne t 1/2 Tmax Cmax AUC all AUC INF Vz/f Cl/f MRT INF h h ng/ml h*ng/ml h*ng/ml ml/kg ml/h /kg h N6His WT 1,26 0,9 92,1 261,6 297,3 1768,9 971,2 2,52 PP WT 1,19 1,5 96,6 9,8 294,7 76,2 9,4 2,53 R72 14, ,5 NE 11824,7 458,9 21,9 36,52 R77 32, , , ,8 655,8 14,4 59,18 L86 33, ,2 32,7 3928,8 33,3 66,0 59,27 H87 16, ,3 NE 86,7 731,1 31,7 36,31 E91 19, ,7 NE 12177,2 606,2,9 40,35 Y4 12, , , ,1 416,3 11,9 47,63 Q8 27, , , ,1 272,4 7,0 51,71 R126 16,48 248, ,3 033,4 643,0 16,8 49,57 R131 26, , , ,0 3,0 8,3 50,41 P146 22, ,3 NE 908,4 503,4,9 40,07 Seerumikontsentratsiooni versus ajakõveraid hinnati mittekompartmentaliseeritud analüüsi teel (Pharsight, versioon 4.1). N=3-4 rotti iga ühendi kohta. ND: ei tehtud; NE: ei hinnatud.
289 5 287 Tmax: Cmax saavutamiseks kuluv aeg; Cmax: maksimaalne kontsentratsioon; terminal t 1/2 : terminaalne poolväärtusaeg; AUC last : kontsentratsiooni ja aja kõvera alune pindala plasma viimase proovivõtu ajahetkeni AUC inf : kontsentratsiooni ja aja kõvera alune pindala, mis on ekstrapoleeritud lõpmatuse suhtes; MRT: keskmine viibimisaeg; Cl/f: näiv kogu plasmakliirens; Vz/f: näiv jaotumine terminaalse faasi ajal. Tabel 4. Rottidele 0, mg/kg suuruse annusena manustatud KPEG-pAF91(16- hhis)fgf21 farmakokineetilised parameetrid Rott #1 Rott #2 Rott #3 Rott #4 Terminaalne h 22,6 24,4 17,3 16,9 t 1/2 Tmax h Cmax ng/ml 163,0 182,1 5,7 88,8 AUC all h*ng/ml 7629,1 7659,1 6461,4 4375,4 AUC INF h*ng/ml 8232,0 8333,8 6661,2 4521,7 Vz/f ml/kg 989,8 54,2 937,3 1347,7 Cl/f ml/h/kg 30,4 30,0 37,5 55,3 MRT INF h 39,9 41,0 32,5 34,3 Seerumikontsentratsiooni versus ajakõveraid hinnati mittekompartmentaliseeritud analüüsi teel (Pharsight, versioon 4.1). ND: ei tehtud; NE: ei hinnatud. Tmax: Cmax saavutamiseks kuluv aeg; Cmax: maksimaalne kontsentratsioon; terminal t 1/2 : terminaalne poolväärtusaeg; AUC last : kontsentratsiooni ja aja kõvera alune pindala plasma viimase proovivõtu ajahetkeni AUC inf : kontsentratsiooni ja aja kõvera alune pindala, mis on ekstrapoleeritud lõpmatuse suhtes; MRT: keskmine viibimisaeg; Cl/f: näiv kogu plasmakliirens; Vz/f: näiv jaotumine terminaalse faasi ajal.
290 Näide Mittelooduslikult kodeeritud aminohapet sisaldava PEGüleeritud FGF-21 ohutuse ja/või efektiivsuse uuring inimpatsientidel 5 30 Eesmärk Jälgida subkutaanselt manustatava mittelooduslikult kodeeritud aminohapet sisaldava PEGüleeritud rekombinantse inimese FGF-21 ohutust ja farmakokineetikat. Patsiendid Sellesse uuringusse kaasatakse 18 tervet vabatahtlikku vanuses kuni 40 aastat ja kehakaaluga 60 kuni 90 kg. Katsealustel puuduvad kliiniliselt olulised ebanormaalsed laboratoorsete analüüside tulemused hematoloogia või seerumi keemiliste näitajate osas ning neil on negatiivne uriini toksikoloogiline analüüs, HIV analüüs ja B- hepatiidi pinnaantigeeni analüüs. Katsealustel ei tohi olla järgmisi seisundeid/asjaolusid: hüpertensioon; mis tahes varasem primaarne hematoloogiline haigus; varasem oluline maksa-, neeru-, südame-veresoonkonna-, seedetrakti-, kuse-suguelundkonna, ainevahetusvõi neuroloogiline haigus; varasem aneemia või epilepsia; teadaolev tundlikkus bakteriaalset või loomset päritolu saaduste. PEG-i või inimese seerumi albumiini suhtes; kofeiini sisaldavate jookide krooniline ja suurtes kogustes tarbimine; osalemine mis tahes teises kliinilises uuringus või vereülekanne või vere andmine 30 päeva jooksul enne uuringusse kaasamist; kokkupuude FGF-21 polüpeptiidiga kolme kuu jooksul enne uuringusse kaasamist; haigus seitsme päevas jooksul enne uuringusse kaasamist; ja olulised kõrvalekalded 14 päeva jooksul enne uuringusse kaasamist tehtud uuringueelses füüsilises läbivaatuses või kliinilistes laboratoorsetes analüüsides (hinnangutes). Kõiki katsealuseid hinnatakse ohutuse osas ja kõik farmakokineetilise analüüsi jaoks vajalikud vereproovid võetakse vastava ajakava järgi. Uuringudisain See on 1. faasi ühekeskuseline, randomiseeritud, kaheperioodiline ristuuring, mis viiakse läbi tervetel meessoost vabatahtlikel katsealustel. 18 katsealust randomiseeritakse kahte katserühma (mõlemas rühmas 9 katsealust). FGF-21 polüpeptiidi manustatakse kahel eraldi manustamisperioodil subkutaanselt reie ülaossa süstiva boolusena, kasutades samaväärseid mittelooduslikult kodeeritud aminohapet sisaldavat PEGüleeritud FGF-21 polüpeptiidi ja valitud kaubanduslikult kättesaadavat toodet. Kaubanduslikult kättesaadava toote annuse suurus ja manustamissagedus vastavad toote pakendil toodud ettekirjutustele. Teiste katserühmade kaasamise teel saab uuringus vaadelda ka kaubanduslikult kättesaadavate toodete abil ka lisaannuseid, teistsugust
291 5 289 manustamissagedust ja teisi parameetreid. Manustamisperioodi vahel on alati 14-päevane väljauhteperiood (wash-out period). Katsealuseid hoitakse vähemalt 12 tunni jooksul enne ja 72 tundi pärast manustamist mõlema kahe manustamisperioodi korral uuringukeskuses, kuid manustamisperioodide vahel on nad vabad. Teiste katserühmade kaasamise teel saab uuringus vaadelda ka PEGüleeritud FGF-21 ka lisaannuseid, teistsugust manustamissagedust ja teisi parameetreid. FGF-21 katsepreparaat sisaldab PEGüleeritud FGF-21 polüpeptiidi, mis sisaldab mittelooduslikult kodeeritud aminohapet. Vereproovide võtmine Vereproovid võetakse seeriana veenis enne ja pärast FGF-21 manustamist. Veenivereproovid (5 ml), mida kasutatakse FGF-21 seerumikontsentratsiooni määramiseks, võetakse umbes 30, ja minutit enne manustamist (3 baasproovi) ja järgmistel ligikaudsetel ajahetkedel pärast manustamist: 30 minutit ja 1, 2, 5, 8, 12,, 18, 24, 30, 36, 48, 60 ja 72 tunni möödudes. Iga seerumiproov jagatakse kaheks alikvoodiks. Kõiki seerumiproove hoitakse temperatuuril - C. Seerumiproove tarnitakse kuival jääl. Söömata olekus tehtavad kliinilised laboratoorsed analüüsid (hematoloogia, seerumi keemiline analüüs ja uriini analüüs) viiakse läbi vahetult enne esimese annuse manustamist 1. päeval, 4. päeva hommikul, vahetult enne manustamist 16. päeval ja 19. päeva hommikul. Bioanalüütilised meetodid FGF-21 seerumikontsentratsioonid määratakse ELISA komplektiga. Ohutuse määramine Elulised näitajad dokumenteeritakse enne igat manustamist (1. ja 16. päeval) ja 6, 24, 48 ja 72 tundi pärast igat manustamist. Ohutuse määramisel lähtutakse soovimatute kõrvanähtude esinemisest ja tüübist ning kliiniliste laboratoorsete analüüside tulemuste muutumisest võrreldes vastavate baasväärtustega. Lisaks hinnatakse muutusi võrreldes uuringueelsete eluliste näitajatega, sealhulgas vererõhu ja füüsilise läbivaatuse käigus dokumenteeritud näitajatega. Andmete analüüs Annustamisjärgseid seerumikontsentratsiooni väärtuste korrigeerimiseks manustamiseelsete FGF-21 algkontsentratsioonide järgi lahutatakse igast
292 5 290 manustamisjärgsest väärtusest 30, ja minutit enne manustamist võetud kolme proovi FGF-21 taseme keskmistamise teel saadud keskmise FGF-21 kontsentratsiooni baasväärtus. Manustamiseelseid FGF-21 kontsentratsioone ei kaasata keskmise väärtuse arvutamisse, kui nad on analüüsis kvantiteerimistasemest väiksemad. Farmakokineetilised parameetrid määratakse FGF-21 kontsentratsioonide baasväärtuste järgi korrigeeritud seerumikontsentratsiooni andmete põhjal. Farmakokineetilised parameetrid arvutatakse mudelist sõltumatutel meetoditel Digital Equipment Corporation VAX 8600 arvutisüsteemiga, kasutades BIOAVL tarkvara kõige uuemat versiooni. Määratakse järgmised farmakokineetilised parameetrid: maksimaalne seerumikontsentratsioon (C max ); maksimaalse seerumikontsentratsiooni tekkeks kuluv aeg (t max ); kontsentratsiooni ja aja kõvera alune pindala (AUC), mis arvutatakse lineaarse trapetsreegli abil ajahetkest null kuni viimase vereproovi võtmise ajani (AUC 0-72 ) ja terminaalne eliminatsiooni poolväärtusaeg (t 1/2 ), mille arvuti arvutab eliminatsiooni kiiruse konstandi järgi. Väljutuse kiiruskonstanti hinnatakse järjestikuste andmepunktide lineaarse regressiooni teel loglinear kontsentratsiooni ja aja graafiku terminaalses lineaarses piirkonnas. Iga ravivariandi puhul arvutatakse farmakokineetiliste parameetrite keskmine standardhälve (SD) ja variatsioonikoefitsent. Seejärel arvutatakse parameetrite keskmiste väärtuste suhe säilitusainega/säilitusaineta ravimvormid 30 Ohutustulemused Soovimatute kõrvanähtude esinemissagedus on kõikides katserühmades võrdne. Puuduvad kliiniliselt olulised muutused võrreldes baasväärtustega või uuringueelsete kliiniliste laboratoorsete analüüside või vererõhu näitajatega ning märkimisväärsed muutused võrreldes uuringueelsele füüsilise läbivaatuse tulemuste ja eluliselt oluliste näitajatega. Kahe katserühma ohutusprofiilid peaksid olema sarnased. Farmakokineetilised tulemused Kõigil 18 katsealuse puhul koostatakse pärast mittelooduslikult kodeeritud aminohapet sisaldava PEGüleeritud FGF-21 manustamist igal ettenähtud ajahetkel mõõdetud tulemuste põhjal FGF-21 keskmise seerumikontsentratsiooni ja aja profiilid (mis ei ole korrigeeritud FGF-21 baastaseme järgi). Kõigi katsealuste manustamiseelsed FGF-21 baaskontsentratsioonid peaksid jääma füsioloogiliselt normaalsesse vahemikku. Farmakokineetilised parameetrid määratakse FGF-21 manustamiseelsete keskmiste baaskontsentratsioonide järgi korrigeeritud seerumikontsentratsiooni andmete kohta ja määratakse C max ning t max.
293 291 Kõigi valitud võrdlusravimite keskmine t max on oluliselt lühem kui mittelooduslikult kodeeritud aminohapet sisaldava PEGüleeritud FGF-21 t max. Testitud eelkliinilise võrdlusravimi(te) terminaalsed poolväärtusajad on oluliselt lühemad kui mittelooduslikult kodeeritud aminohapet sisaldava PEGüleeritud FGF-21 terminaalne poolväärtusaeg. 5 Kuigi käesolev uuring viiakse läbi tervetel meessoost katsealustel, on teistel patsiendipopulatsioonidel nagu vähktõve või kroonilise neerupuudulikkusega mees- või naissoost patsiendid neerupuudulikkusega lapsed, autoloogsetes predeposiitsetes programmides osalevad täiskasvanud või plaanilist elektiivset kirurgilist ravi ootavad patsiendid eeldustekohaselt sarnased imendumisega seotud omadused ja ohutusprofiilid. Kokkuvõttes saab öelda, et mittelooduslikult kodeeritud aminohapet sisaldava PEGüleeritud FGF-21 üksikannused on tervetele meessoost katsealustele ohutud ja nimetatud katsealused taluvad neid annuseid hästi. Soovimatute kõrvalnähtude esinemise, kliiniliste laboratoorsete väärtuste, eluliste näitajate ja füüsilise läbivaatuse tulemuste põhjal on FGF-21 kaubanduslikult kättesaadavate vormide ja mittelooduslikku aminohapet sisaldava PEGüleeritud FGF-21 ohutusprofiilid samaväärsed. Mittelooduslikku aminohapet sisaldav PEGüleeritud FGF-21 on patsientidele ja tervishoiutöötajatele väga suure kliinilise kasulikkusega ühend. Käesolevas dokumendis kirjeldatavad näited ja teostused on vaid illustreerivad ning asjatundjad võivad teha selles erinevaid modifikatsioone ja muudatusi, mis jäävad samuti käesoleva taotluse ja sellega kaasasoleva patendinõudluse ulatusse. TABEL 5: Viidatud järjestused SEQ ID # Järjestuse nimi 1 FGF-21 aminohappeline järjestus ilma liiderjärjestuseta (P vorm)
294 292 2 FGF-21 aminohappeline järjestus ilma liiderjärjestuseta -His märgisega 3 Liiderjärjestusega FGF-21 (P vorm) aminohappeline järjestus liiderjärjestuses on 3 leutsiini (9 aminohappest koosnev P-vorm) 4 Liiderjärjestusega FGF-21 (P vorm) aminohappeline järjestus - liiderjärjestuses on 3 leutsiini 5 Ilma liiderjärjestuseta FGF-21 aminohappeline järjestus (L vorm)
295 293 6 Liiderjärjestusega FGF-21 (L vorm) aminohappeline järjestus liiderjärjestuses on 3 leutsiini (9 aminohappest koosnev L-vorm) 7 Liiderjärjestusega FGF-21 (L vorm) aminohappeline järjestus liiderjärjestuses on 2 leutsiini (8 aminohappest koosnev L-vorm) 8 Liiderjärjestuseta FGF-21 (P vorm) nukleotiidne järjestus
296 294 9 Liiderjärjestuseta FGF-21 (P vorm) nukleotiidne järjestus His märgisega Liiderjärjestusega FGF-21 (P vorm) nukleotiidne järjestus liiderjärjestuses on 3 leutsiini 11 Liiderjärjestusega FGF-21 (P vorm) nukleotiidne järjestus - liiderjärjestuses on
297 295 2 leutsiini 12 Ilma liiderjärjestuseta FGF-21 (L vorm) nukleotiidne järjestus 13 Liiderjärjestusega FGF-21 nukleotiidne järjestus (L vorm) liiderjärjestuses on 3 leutsiini
298 Nucleotide Sequence for FGF-21 with leader (L vorm) liiderjärjestuses on 2 leutsiini 34 FGF-21 aminohappeline järjestus (Rattus norvegicus - ref NP_ [ ])
299 FGF-21 aminohappeline järjestus (Mus musculus - ref NP_ [99218]) 36 FGF-21 aminohappeline järjestus (Danio rerio - ref NP_ [ ]) 37 Klotho beeta aminohappeline järjestus (Homo sapiens - ref NP_ [ ])
300 Klotho beeta aminohappeline järjestus (Mus musculus - refnp_ GI: )
301 OmpA nukleotiidi liiderjärjestus atgaaaaaaactgctatcgcgatcgctgtagctctggctggtttcgcgaccgtagctaacgct 40 OmpA aminohappe liidejärjestus M K K T A I A I A V A L A G F A T V A N A 41 MalE nukleotiidi liiderjärjestus atgaaaataaaaacaggtgcacgcatcctcgcattatccgcattaacgacgatgatgttttccgcctcggctctcgcc 42 MalE aminohappe liidejärjestus M K I K T G A R I L A L S A L T T M M F S A S A L A 43 StII nukleotiidi liiderjärjestus atgaaaaagaatatcgcatttcttcttgcatctatgttcgttttttctattgctacaaatgcctatgca 44 StII aminohappe liidejärjestus M K K N I A F L L A S M F V F S I A T N A Y A
302 Patendinõudlus FGF-21 polüpeptiid, mis sisaldab järjestust SEQ ID NO: 1-7, milles üks aminohape positsioonis 8 (SEQ ID NO: 1 või vastavad aminohapped järjestustes SEQ ID NOs: 2-7) on asendatud mittelooduslikult kodeeritud aminohappega, kusjuures nimetatud FGF-21 polüpeptiidis on selle mittelooduslikult kodeeritud aminohappega seotud vees lahustuv polümeer. 2. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral polüpeptiid on seotud linkeri, polümeeri või bioloogiliselt aktiivse molekuliga. 3. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral polüpeptiid on seotud bifunktsionaalse polümeeri, bifunktsionaalse linkeri või vähemalt veel ühe FGF-2 polüpeptiidiga. 4. Nõudluspunktile 3 vastav FGF-21 polüpeptiid, mille korral bifunktsionaalne linker on seotud teise polüpeptiidiga. 5. Nõudluspunktile 4 vastav FGF-21 polüpeptiid, mille korral teine polüpeptiid on FGF-21 polüpeptiid. 6. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral vees lahustuv polümeer sisaldab polü(etüleenglükool)fragmenti. 7. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral mittelooduslikult kodeeritud aminohape sisaldab karbonüül-, aminooksü-, hüdrasiin-, hüdrasiid-, semikarbasiid- või alküünrühma. 8. Nõudluspunktile 7 vastav FGF-21 polüpeptiid, mille korral mittelooduslikult kodeeritud aminohape sisaldab karbonüülrühma. 9. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral vees lahustuva polümeeri molekulmass on umbes 0,1 kda kuni umbes 0 kda.. Nõudluspunktile 9 vastav FGF-21 polüpeptiid, mille korral vees lahustuva polümeeri molekulmass on umbes 0,1 kda kuni umbes 50 kda.
303 Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mis on valmistatud karbonüülrühma sisaldavat aminohapet sisaldava FGF-21 polüpeptiidi ja aminooksü-, hüdrasiin-, hüdrasiidvõi semikarbasiidrühma sisaldava vees lahustuva polümeeri vahelise reaktsiooni teel Nõudluspunktile 11 vastav FGF-21 polüpeptiid, mille korral aminooksü-, hüdrasiin-, hüdrasiid- või semikarbasiidrühm seotakse vees lahustuva polümeeriga amiidsideme kaudu. 13. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mis on valmistatud karbonüülrühma sisaldava vees lahustuva polümeeri ja aminooksü-, hüdrasiin-, hüdrasiid- või semikarbasiidrühma sisaldavat mittelooduslikult kodeeritud aminohapet sisaldava polüpeptiidi vahelise reaktsiooni teel. 14. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral vees lahustuv polümeer on hargnenud või mitmeõlgne polümeer.. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral vees lahustuva polümeeri iga ahela molekulmass on umbes 1 kda kuni umbes 0 kda. 16. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral mittelooduslikult kodeeritud aminohape sisaldab sahhariidifragmenti. 17. Nõudluspunktile 2 vastav FGF-21 polüpeptiid, mille korral lonker, polümeer või bioloogiliselt aktiivne molekul on seotud polüpeptiidiga sahhariidifragmendi kaudu. 18. Kompositsioon, mis sisaldab nõudluspunktile 1 vastavat FGF-21 polüpeptiidi ja farmatseutiliselt sobivat tugiainet. 19. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mida kodeerib polünukleotiid järjestusega SEQ ID NO: 8, 9,, 11, 12, 13 või 14, kusjuures see polünukleotiid sisaldab selektorkoodonit.. Nõudluspunktile 19 vastav FGF-21 polüpeptiid, mille korral vees lahustuv polümeer sisaldab polü(etüleen)fragmenti.
304 Nõudluspunktile 19 vastav FGF-21 polüpeptiid, mille korral mittelooduslikult kodeeritud aminohape sisaldab karbonüül-, aminooksü-, hüdrasiid-, hüdrasiin-, semikarbasiid-, asiid- või alküünrühma Nõudluspunktile vastav FGF-21 polüpeptiid, mille korral polü(etüleenglükool)fragmendi molekulmass on umbes 0,1 kda kuni umbes 0 kda. 23. Nõudluspunktile vastav FGF-21 polüpeptiid, mille korral polü(etüleenglükool)fragment on hargnenud või mitmeõlgne polümeer. 24. Nõudluspunktile 23 vastav FGF-21 polüpeptiid, mille korral polü(etüleenglükool)fragmendi molekulmass on umbes 1 kda kuni umbes 0 kda.. Kompositsioon, mis sisaldab nõudluspunktile 19 vastavat FGF-21 polüpeptiidi ja farmatseutiliselt sobivat tugiainet. 26. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mis sisaldab vees lahustuvat polümeeri, mis on mitteloodusliku aminohappe juures seotud kovalentselt FGF-21 polüpeptiidiga. 27. Nõudluspunktile 26 vastav FGF-21 polüpeptiid, mille korral vees lahustuv polümeer sisaldab polü(etüleenglükool)fragmenti. 28. Nõudluspunktile 7 vastav FGF-21 polüpeptiid, mille korral mittelooduslikult kodeeritud aminohape on seotud polü(etüleenglükool) molekuliga. 29. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mis sisaldab vähemalt ühte linkerit, polümeeri või bioloogiliselt aktiivset molekuli, kusjuures see linker, polümeer või bioloogiliselt aktiivne molekul kinnitub polüpeptiidi külge polüpeptiidi ribosomaalselt sisestatud mitteloodusliku aminohappe funktsionaalrühma kaudu. 30. Nõudluspunktile 29 vastav FGF-21 polüpeptiid, kusjuures see FGF-21 polüpeptiid on monopegüleeritud.
305 Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mis sisaldab vähemalt ühte linkerit, polümeeri või bioloogiliselt aktiivset molekuli, mis kinnitub mittelooduslikult kodeeritud aminohappe külge, kusjuures see mittelooduslikult kodeeritud aminohape on sisestatud ribosomaalselt polüpeptiidi Nõudluspunktile 31 vastav FGF-21 polüpeptiid, kusjuures see FGF-21 polüpeptiid sisaldab linkerit, polümeeri või bioloogiliselt aktiivset molekuli. 33. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral mittelooduslikult kodeeritud aminohape sisaldab ketoonrühma. 34. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, kusjuures see FGF-21 polüpeptiid sisaldab järjestust SEQ ID NO: 1, kuid selles on üks aminohape asendatud positsioonis 8 mittelooduslikult kodeeritud aminohappega. 35. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral mittelooduslikult kodeeritud aminohape on para-atsetüül-fenüülalaniin 36. Nõudluspunktile 1 vastav FGF-21 polüpeptiid, mille korral vees lahustuv polümeer on polü(etüleenglükool)fragment molekulmassiga 30 kda.
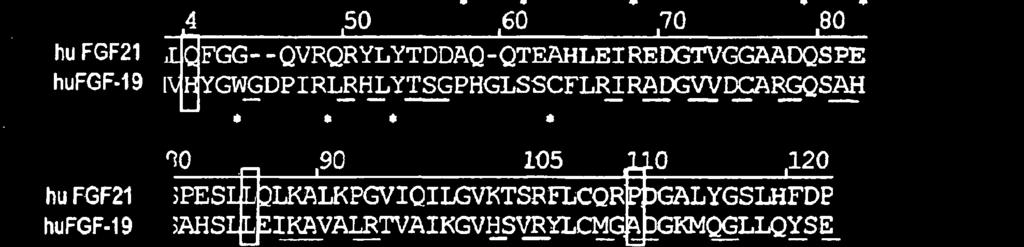

306 1/33

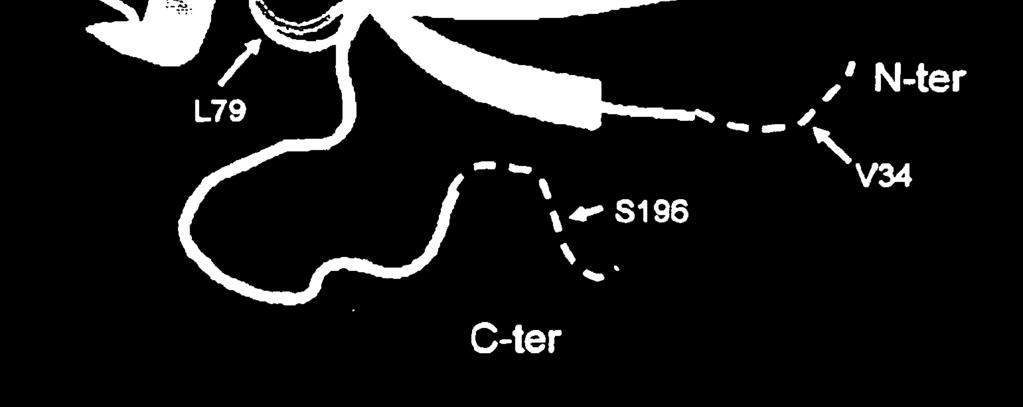
307 2/33


308 3/33



309 4/33
310 5/33
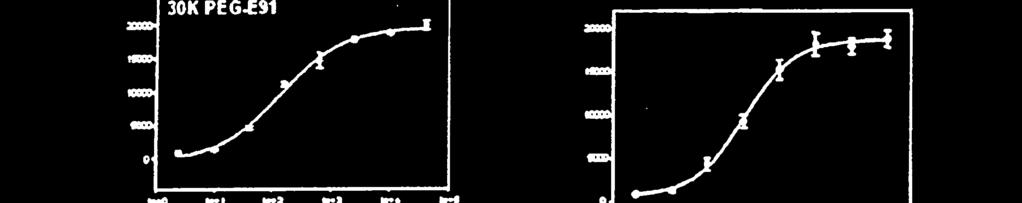
311 6/33


312 7/33
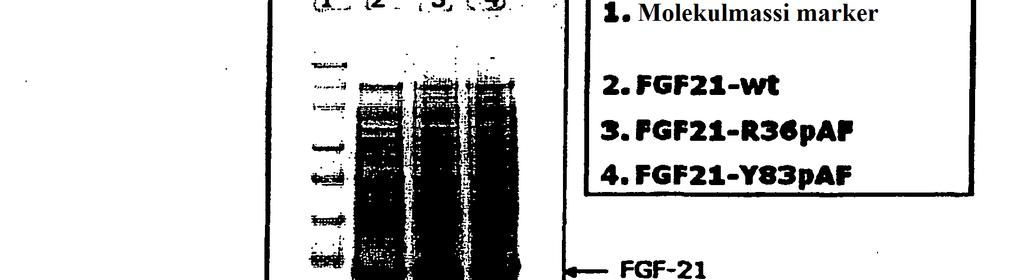
313 8/33


314 9/33
315 /33
316 11/33
317 12/33
318 13/33
319 14/33
320 /33
321 16/33
322 17/33

323 18/33



324 19/33
325 /33
326 21/33
327 22/33

328 23/33




329 24/33
330 /33
331 26/33
332 27/33
333 28/33
334 29/33
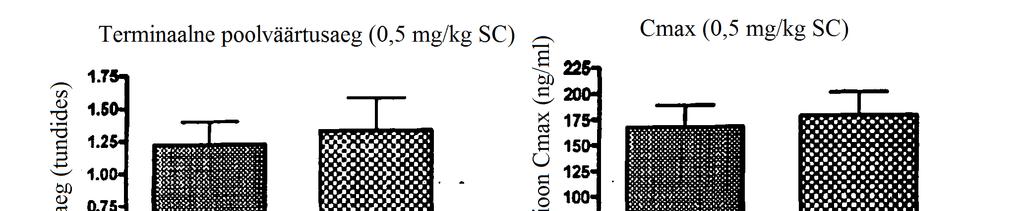


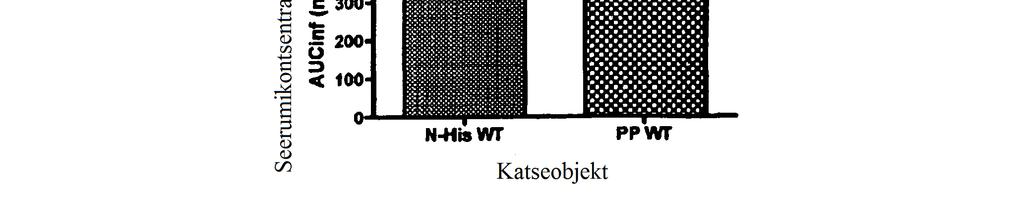
335 30/33

336 31/33 5
337 32/33


338 33/33
EE-EP B1. järjestus.
 1 2 LEIUTISE TAUST Paljuraksetes organismides reguleerivad rakkude paljunemist ja diferentseerumist hormoonid ja polüpeptiidsed kasvufaktorid. Sellised difundeeruvad molekulid võimaldavad rakkudel üksteisega
1 2 LEIUTISE TAUST Paljuraksetes organismides reguleerivad rakkude paljunemist ja diferentseerumist hormoonid ja polüpeptiidsed kasvufaktorid. Sellised difundeeruvad molekulid võimaldavad rakkudel üksteisega
Pikkade järjestuste koopiaarvu varieerumine inimese genoomis. Priit Palta
 Pikkade järjestuste koopiaarvu varieerumine inimese genoomis Priit Palta Varieerumine inimese genoomis SNP-d Väikesed insertsioon/deletsioon polümorfismid Kordusjärjestused Genoomi struktuursed muutused
Pikkade järjestuste koopiaarvu varieerumine inimese genoomis Priit Palta Varieerumine inimese genoomis SNP-d Väikesed insertsioon/deletsioon polümorfismid Kordusjärjestused Genoomi struktuursed muutused
ADVANCED SEQUENCING TECHNOLOGIES
 ADVANCED SEQUENCING TECHNOLOGIES Journal Club 24.11.2004 Miks sekveneerida? Evolutsioon ja ökoloogia globaalne biomass sisaldab 10 38 nukleotiidi, praeguseks sekveneeritud 10 11 nukleotiidi - liikide kirjeldamine
ADVANCED SEQUENCING TECHNOLOGIES Journal Club 24.11.2004 Miks sekveneerida? Evolutsioon ja ökoloogia globaalne biomass sisaldab 10 38 nukleotiidi, praeguseks sekveneeritud 10 11 nukleotiidi - liikide kirjeldamine
Bakteriaalselt saadud puutumata minirakud, mis sisaldavad plasmiidivaba funktsionaalset nukleiinhapet in vivo imetajarakkudesse transpordiks
 Bakteriaalselt saadud puutumata minirakud, mis sisaldavad plasmiidivaba funktsionaalset nukleiinhapet in vivo imetajarakkudesse transpordiks TEHNIKA TASE Välja on töötatud hulk nukleiinhappepõhiseid strateegiaid
Bakteriaalselt saadud puutumata minirakud, mis sisaldavad plasmiidivaba funktsionaalset nukleiinhapet in vivo imetajarakkudesse transpordiks TEHNIKA TASE Välja on töötatud hulk nukleiinhappepõhiseid strateegiaid
EESTI STANDARD EVS-ISO 4967:2007
 EESTI STANDARD TERAS Mittemetalsete lisandite sisalduse määramine Mikrograafiline meetod standardkaartide kasutamisega Steel Determination of content of nonmetallic inclusions Micrographic method using
EESTI STANDARD TERAS Mittemetalsete lisandite sisalduse määramine Mikrograafiline meetod standardkaartide kasutamisega Steel Determination of content of nonmetallic inclusions Micrographic method using
This document is a preview generated by EVS
 EESTI STANDARD EVS-ISO/IEC 15288:2009 Süsteemi- ja tarkvaratehnika Süsteemi elutsükli protsessid Systems and software engineering System life cycle processes EESTI STANDARDI EESSÕNA NATIONAL FOREWORD Käesolev
EESTI STANDARD EVS-ISO/IEC 15288:2009 Süsteemi- ja tarkvaratehnika Süsteemi elutsükli protsessid Systems and software engineering System life cycle processes EESTI STANDARDI EESSÕNA NATIONAL FOREWORD Käesolev
Enne kuimidagimuud tegemahakkate on tarvisinpa/ediabas instalidaomale arvutisse. M inaise kasutan INPA versiooni3.01,miskasutab EDIABAS v6.4.3.
 Enne kuimidagimuud tegemahakkate on tarvisinpa/ediabas instalidaomale arvutisse. M inaise kasutan INPA versiooni3.01,miskasutab EDIABAS v6.4.3. 1. Arvutiette valmistamine AvaControlPanel,ValiClassicView
Enne kuimidagimuud tegemahakkate on tarvisinpa/ediabas instalidaomale arvutisse. M inaise kasutan INPA versiooni3.01,miskasutab EDIABAS v6.4.3. 1. Arvutiette valmistamine AvaControlPanel,ValiClassicView
TTÜ ehituse ja arhitektuuri instituut MONTEERITAVATEST ELEMENTIDEST CON/SPAN SILLA JA INTEGRAALSILLA VÕRDLUS OARA SILLA (NR.
 TTÜ ehituse ja arhitektuuri instituut MONTEERITAVATEST ELEMENTIDEST CON/SPAN SILLA JA INTEGRAALSILLA VÕRDLUS OARA SILLA (NR. 715) NÄITEL COMPARISON OF PRECAST CON/SPAN BRIDGE SYSTEM AND INTEGRAL BRIDGE
TTÜ ehituse ja arhitektuuri instituut MONTEERITAVATEST ELEMENTIDEST CON/SPAN SILLA JA INTEGRAALSILLA VÕRDLUS OARA SILLA (NR. 715) NÄITEL COMPARISON OF PRECAST CON/SPAN BRIDGE SYSTEM AND INTEGRAL BRIDGE
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Hans Vellama PepFect14 in vivo transfektsiooniefektiivsuse hindamine hiire kopsus ja maksas laserkonfokaalmikrsoskoobiga.
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Hans Vellama PepFect14 in vivo transfektsiooniefektiivsuse hindamine hiire kopsus ja maksas laserkonfokaalmikrsoskoobiga.
Search for causality in ecological studies
 Search for causality in ecological studies Context1: estimating impact of the natural factors on the chemical and biological variables in Lake Peipsi Context2: preparing papers on the topic 7 Presented
Search for causality in ecological studies Context1: estimating impact of the natural factors on the chemical and biological variables in Lake Peipsi Context2: preparing papers on the topic 7 Presented
Thermal power plant cooperation with wind turbines
 Estonian Journal of Engineering, 2008, 14, 4, 317 324 doi: 10.3176/eng.2008.4.03 Thermal power plant cooperation with wind turbines Ivo Palu, Heiki Tammoja and Rein Oidram Department of Electrical Power
Estonian Journal of Engineering, 2008, 14, 4, 317 324 doi: 10.3176/eng.2008.4.03 Thermal power plant cooperation with wind turbines Ivo Palu, Heiki Tammoja and Rein Oidram Department of Electrical Power
This document is a preview generated by EVS
 EESTI STANDARD EVS-ISO 11799:2016 INFORMATSIOON JA DOKUMENTATSIOON Arhiivi- ja raamatukogumaterjalide hoiunõuded Information and documentation Document storage requirements for archive and library materials
EESTI STANDARD EVS-ISO 11799:2016 INFORMATSIOON JA DOKUMENTATSIOON Arhiivi- ja raamatukogumaterjalide hoiunõuded Information and documentation Document storage requirements for archive and library materials
EESTI STANDARD EVS-ISO :2009
 EESTI STANDARD EVS-ISO 21527-2:2009 TOIDU JA LOOMASÖÖTADE MIKROBIOLOOGIA Pärmide ja hallituste loendamise horisontaalmeetod Osa 2: Kolooniate loendamise tehnika toodetes, mille veeaktiivsus on väiksem
EESTI STANDARD EVS-ISO 21527-2:2009 TOIDU JA LOOMASÖÖTADE MIKROBIOLOOGIA Pärmide ja hallituste loendamise horisontaalmeetod Osa 2: Kolooniate loendamise tehnika toodetes, mille veeaktiivsus on väiksem
NIISKUSE JA UV-KIIRGUSE MÕJU PUITPLASTKOMPOSIITIDE MEHAANILISTELE OMADUSTELE
 TALLINNA TEHNIKAÜLIKOOL POLÜMEERMATERJALIDE INSTITUUT PUIDUTÖÖTLEMISE ÕPPETOOL NIISKUSE JA UV-KIIRGUSE MÕJU PUITPLASTKOMPOSIITIDE MEHAANILISTELE OMADUSTELE Magistritöö Heikko Kallakas Juhendaja: Triinu
TALLINNA TEHNIKAÜLIKOOL POLÜMEERMATERJALIDE INSTITUUT PUIDUTÖÖTLEMISE ÕPPETOOL NIISKUSE JA UV-KIIRGUSE MÕJU PUITPLASTKOMPOSIITIDE MEHAANILISTELE OMADUSTELE Magistritöö Heikko Kallakas Juhendaja: Triinu
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIA TEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL. Kristiina Hein
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIA TEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Kristiina Hein Kolmanda põlvkonna sekveneerimistehnoloogiad Bakalaureusetöö Juhendaja prof.
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIA TEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Kristiina Hein Kolmanda põlvkonna sekveneerimistehnoloogiad Bakalaureusetöö Juhendaja prof.
Procedures. Text Algorithm Projects. Automaton toolbox: RE >NFA, determinisation and minimization. Project
 Procedures Text Algorithm Projects Select topic (today) Prepare a project goal (hypothesis to test, programs to implement, experiments to run) 1 day Prepare a realistic project plan (3 hours) Aim at achieving
Procedures Text Algorithm Projects Select topic (today) Prepare a project goal (hypothesis to test, programs to implement, experiments to run) 1 day Prepare a realistic project plan (3 hours) Aim at achieving
Antikoagulandid, nende toimemehhanismid ja määramine TIIT SALUM
 Antikoagulandid, nende toimemehhanismid ja määramine TIIT SALUM Figure. Classification of established anticoagulants and new anticoagulants that were recently licensed for use or are in advanced stages
Antikoagulandid, nende toimemehhanismid ja määramine TIIT SALUM Figure. Classification of established anticoagulants and new anticoagulants that were recently licensed for use or are in advanced stages
This document is a preview generated by EVS
 EESTI STANDARD EVS-ISO 13528:2017 STATISTILISED MEETODID LABORITEVAHELISTE VÕRDLUSTE TASEMEKATSETES KASUTAMISEKS Statistical methods for use in proficiency testing by interlaboratory comparison (ISO 13528:2015)
EESTI STANDARD EVS-ISO 13528:2017 STATISTILISED MEETODID LABORITEVAHELISTE VÕRDLUSTE TASEMEKATSETES KASUTAMISEKS Statistical methods for use in proficiency testing by interlaboratory comparison (ISO 13528:2015)
Kätlin Karlson. Mg 2+ JA Mn 2+ MÕJU NUKLEOTIIDIDE HETEROTRIMEERSETE G-VALKUDEGA SEOSTUMISELE
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND Keemia Instituut Bioorgaanilise keemia õppetool Kätlin Karlson Mg JA Mn MÕJU NUKLEOTIIDIDE HETEROTRIMEERSETE G-VALKUDEGA SEOSTUMISELE Bakalaureusetöö (12
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND Keemia Instituut Bioorgaanilise keemia õppetool Kätlin Karlson Mg JA Mn MÕJU NUKLEOTIIDIDE HETEROTRIMEERSETE G-VALKUDEGA SEOSTUMISELE Bakalaureusetöö (12
This document is a preview generated by EVS
 EESTI STANDARD EVS-ISO 10005:2008 KVALITEEDIJUHTIMISSÜSTEEMID Juhised kvaliteediplaanidele Quality management systems Guidelines for quality plans EESTI STANDARDI EESSÕNA NATIONAL FOREWORD Käesolev Eesti
EESTI STANDARD EVS-ISO 10005:2008 KVALITEEDIJUHTIMISSÜSTEEMID Juhised kvaliteediplaanidele Quality management systems Guidelines for quality plans EESTI STANDARDI EESSÕNA NATIONAL FOREWORD Käesolev Eesti
Erinevate in silico meetodite võrdlus PCR praimerite kvaliteedi parandamiseks
 BIOLOOGIA-GEOGRAAFIA TEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Reidar Andreson Erinevate in silico meetodite võrdlus PCR praimerite kvaliteedi parandamiseks Magistritöö
BIOLOOGIA-GEOGRAAFIA TEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Reidar Andreson Erinevate in silico meetodite võrdlus PCR praimerite kvaliteedi parandamiseks Magistritöö
Ventilatsiooniõhu eelkütte lahendus ja tulemused Eesti kliimas. Jaanus Hallik Tartu Ülikool EETLabor
 Ventilatsiooniõhu eelkütte lahendus ja tulemused Eesti kliimas Jaanus Hallik Tartu Ülikool EETLabor Jaanus.hallik@ut.ee Ettekande sisu Ülevaade uuritud eelküttelahendusest maakontuuriga eelküttekalorifeer
Ventilatsiooniõhu eelkütte lahendus ja tulemused Eesti kliimas Jaanus Hallik Tartu Ülikool EETLabor Jaanus.hallik@ut.ee Ettekande sisu Ülevaade uuritud eelküttelahendusest maakontuuriga eelküttekalorifeer
Vasega joodetud plaatsoojusvahetite veekvaliteedi suunised
 Vasega joodetud plaatsoojusvahetite veekvaliteedi suunised 0 Kokkuvõte Danfoss District Heating on koostanud need vasega joodetud roostevabast terasest (1.4404, X2CrNiMo17-12-2 vastavalt standardile EN
Vasega joodetud plaatsoojusvahetite veekvaliteedi suunised 0 Kokkuvõte Danfoss District Heating on koostanud need vasega joodetud roostevabast terasest (1.4404, X2CrNiMo17-12-2 vastavalt standardile EN
VAJALIK, KUID MITTE PIISAV
 VAJALIK, KUID MITTE PIISAV Margus Püüa Riigi infosüsteemide osakond Majandus- ja Kommunikatsiooniministeerium kava avalik teenus mis on eesmärk? mis on probleem? mida muuta? milline on lahenduse suund?
VAJALIK, KUID MITTE PIISAV Margus Püüa Riigi infosüsteemide osakond Majandus- ja Kommunikatsiooniministeerium kava avalik teenus mis on eesmärk? mis on probleem? mida muuta? milline on lahenduse suund?
Helena Palmeos SEEMNED JA IDANDITE KASV
 Tartu Ülikool Loodus- ja tehnoloogiateaduskond Ökoloogia- ja Maateaduste Instituut Taimeökoloogia õppetool Helena Palmeos SEEMNED JA IDANDITE KASV Bakalaureusetöö (12 EAP) Juhendaja: PhD Elle Roosaluste
Tartu Ülikool Loodus- ja tehnoloogiateaduskond Ökoloogia- ja Maateaduste Instituut Taimeökoloogia õppetool Helena Palmeos SEEMNED JA IDANDITE KASV Bakalaureusetöö (12 EAP) Juhendaja: PhD Elle Roosaluste
EESTI AKREDITEERIMISKESKUS LISA I OÜ Tepso Labor akrediteerimistunnistusele K003 ANNEX 1 to accreditation certificate K003 of Tepso Labor Ltd
 Leht Page 1(3) EESTI AKREDITEERIMISKESKUS LISA I OÜ Tepso Labor akrediteerimistunnistusele K3 ANNEX 1 to accreditation certificate K3 of Tepso Labor Ltd 1. Akrediteerimisulatus kalibreerimiste osas on
Leht Page 1(3) EESTI AKREDITEERIMISKESKUS LISA I OÜ Tepso Labor akrediteerimistunnistusele K3 ANNEX 1 to accreditation certificate K3 of Tepso Labor Ltd 1. Akrediteerimisulatus kalibreerimiste osas on
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT. Vidrik Teder
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Vidrik Teder Vähidiagnostiliste peptiidide skriinimine in vivo faagidisplei meetodiga Magistritöö Juhendajad Kuldar
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Vidrik Teder Vähidiagnostiliste peptiidide skriinimine in vivo faagidisplei meetodiga Magistritöö Juhendajad Kuldar
CEVA-PHYLAXIA. Diarsanyl. Kiire ja usaldusväärne vahend koerte ja kasside kõhulahtisuse peatamiseks
 CEVA-PHYLAXIA Diarsanyl Kiire ja usaldusväärne vahend koerte ja kasside kõhulahtisuse peatamiseks PATOFÜSIOLOOGIA ÜKSKÕIK, mis kõhulahtisuse põhjustab, toimuvad ikka kokkuvõttes samad protsessid: muudatused
CEVA-PHYLAXIA Diarsanyl Kiire ja usaldusväärne vahend koerte ja kasside kõhulahtisuse peatamiseks PATOFÜSIOLOOGIA ÜKSKÕIK, mis kõhulahtisuse põhjustab, toimuvad ikka kokkuvõttes samad protsessid: muudatused
Horisont 2020 Ühiskonnaprobeem 1:
 Horisont 2020 Ühiskonnaprobeem 1: Tervishoid, demograafilised muutused ja heaolu Argo Soon konsultant Eesti Teadusagentuur Horisont 2020: Ühiskonnaprobleem 1 Märksõnad: personaalmeditsiin haruldased haigused
Horisont 2020 Ühiskonnaprobeem 1: Tervishoid, demograafilised muutused ja heaolu Argo Soon konsultant Eesti Teadusagentuur Horisont 2020: Ühiskonnaprobleem 1 Märksõnad: personaalmeditsiin haruldased haigused
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT TEHNOLOOGIA INSTITUUT
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT TEHNOLOOGIA INSTITUUT Kristjan Madissoo Inimese replikatiivse CMG helikaasi subühikuid ekspresseerivate vektorite konstrueerimine
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT TEHNOLOOGIA INSTITUUT Kristjan Madissoo Inimese replikatiivse CMG helikaasi subühikuid ekspresseerivate vektorite konstrueerimine
Äriprotsesside modelleerimine ja automatiseerimine Loeng 8 Äriprotsesside modelleerimise metoodika ja dokumenteerimine
 Äriprotsesside modelleerimine ja automatiseerimine Loeng 8 Äriprotsesside modelleerimise metoodika ja dokumenteerimine Enn Õunapuu enn.ounapuu@ttu.ee Feedback & Control Ronald E. Giachetti October 20,
Äriprotsesside modelleerimine ja automatiseerimine Loeng 8 Äriprotsesside modelleerimise metoodika ja dokumenteerimine Enn Õunapuu enn.ounapuu@ttu.ee Feedback & Control Ronald E. Giachetti October 20,
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT. Toomas-Julius Lill
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Toomas-Julius Lill Proteiinkinaasi CK2α ja punase fluorestseeruva valgu TagRFP aktiivse liitvalgu tootmine bakterirakkudes
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Toomas-Julius Lill Proteiinkinaasi CK2α ja punase fluorestseeruva valgu TagRFP aktiivse liitvalgu tootmine bakterirakkudes
pcmv-luc705 reportersüsteemi disain ja valideerimine
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIA TEADUSKOND TARTU ÜLIKOOLI MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Evelin Rammul pcmv-luc705 reportersüsteemi disain ja valideerimine Bakalaureusetöö Juhendajad MSc Kalle
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIA TEADUSKOND TARTU ÜLIKOOLI MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT Evelin Rammul pcmv-luc705 reportersüsteemi disain ja valideerimine Bakalaureusetöö Juhendajad MSc Kalle
The chipping cost of wood raw material for fuel in Estonian conditions
 Forestry Studies Metsanduslikud Uurimused, Vol. 66, Pages 65 74 The chipping cost of wood raw material for fuel in Estonian conditions Marek Irdla*, Allar Padari, Vahur Kurvits and Peeter Muiste Irdla,
Forestry Studies Metsanduslikud Uurimused, Vol. 66, Pages 65 74 The chipping cost of wood raw material for fuel in Estonian conditions Marek Irdla*, Allar Padari, Vahur Kurvits and Peeter Muiste Irdla,
PROJEKTI KLIENDISUHETE HALDAMISE TARKVARAD
 TARTU ÜLIKOOL Pärnu kolledž Ettevõtluse osakond Külli Kivioja-Ööpik, Mari-Liis Kukk, Valter Kolberg, Siret Ulp, Mihkel Ausmaa, Preedik Poopuu EP 3 PROJEKTI KLIENDISUHETE HALDAMISE TARKVARAD Referaat Juhendaja:
TARTU ÜLIKOOL Pärnu kolledž Ettevõtluse osakond Külli Kivioja-Ööpik, Mari-Liis Kukk, Valter Kolberg, Siret Ulp, Mihkel Ausmaa, Preedik Poopuu EP 3 PROJEKTI KLIENDISUHETE HALDAMISE TARKVARAD Referaat Juhendaja:
This document is a preview generated by EVS
 EESTI STANDARD EVS-ISO 15489-1:2017 INFORMATSIOON JA DOKUMENTATSIOON Dokumendihaldus Osa 1: Mõisted ja põhimõtted Information and documentation Records management Part 1: Concepts and principles (ISO 15489-1:2016)
EESTI STANDARD EVS-ISO 15489-1:2017 INFORMATSIOON JA DOKUMENTATSIOON Dokumendihaldus Osa 1: Mõisted ja põhimõtted Information and documentation Records management Part 1: Concepts and principles (ISO 15489-1:2016)
INDIVIDUAL VALUES AND PERCEPTION OF ORGANIZATIONAL CULTURE AMONG RUSSIAN-SPEAKERS IN ESTONIA AND LITHUANIA. Elina Tolmats University of Tartu
 INDIVIDUAL VALUES AND PERCEPTION OF ORGANIZATIONAL CULTURE AMONG RUSSIAN-SPEAKERS IN ESTONIA AND LITHUANIA Introduction Elina Tolmats University of Tartu Estonia and Lithuania have been independent for
INDIVIDUAL VALUES AND PERCEPTION OF ORGANIZATIONAL CULTURE AMONG RUSSIAN-SPEAKERS IN ESTONIA AND LITHUANIA Introduction Elina Tolmats University of Tartu Estonia and Lithuania have been independent for
EHITISINTEGREERITUD PÄIKESEENERGEETIKA. Andri Jagomägi, PhD Roofit.solar Tallinna Tehnikaülikool
 EHITISINTEGREERITUD PÄIKESEENERGEETIKA Andri Jagomägi, PhD Roofit.solar Tallinna Tehnikaülikool Andri Jagomägi, PhD andri.jagomagi@roofit.solar Tallinna Tehnikaülikool TEEMAD Mis on ehitisintegreeritud
EHITISINTEGREERITUD PÄIKESEENERGEETIKA Andri Jagomägi, PhD Roofit.solar Tallinna Tehnikaülikool Andri Jagomägi, PhD andri.jagomagi@roofit.solar Tallinna Tehnikaülikool TEEMAD Mis on ehitisintegreeritud
Mechanical properties of pinewood (Pinus Sylvestris) swollen in organic liquids
 swollen in organic liquids") Proc. Estonian Acad. Sci. Eng., 2006, 12, 2, 125 133 a b Mechanical properties of pinewood (Pinus Sylvestris) swollen in organic liquids Pille Meier a, Eve Stöör a, Tiit Kaps a and Urve Kallavus b Department
Proc. Estonian Acad. Sci. Eng., 2006, 12, 2, 125 133 a b Mechanical properties of pinewood (Pinus Sylvestris) swollen in organic liquids Pille Meier a, Eve Stöör a, Tiit Kaps a and Urve Kallavus b Department
TARKVARAARENDUSE KULU HINDAMISE METOODIKAD EESTIS: TEOORIA JA TEGELIKKUS
 TALLINNA TEHNIKAÜLIKOOL Infotehnoloogia teaduskond Informaatika instituut IDK40LT Ann-Claire Utt 134207IAPB TARKVARAARENDUSE KULU HINDAMISE METOODIKAD EESTIS: TEOORIA JA TEGELIKKUS Bakalaureusetöö Juhendaja:
TALLINNA TEHNIKAÜLIKOOL Infotehnoloogia teaduskond Informaatika instituut IDK40LT Ann-Claire Utt 134207IAPB TARKVARAARENDUSE KULU HINDAMISE METOODIKAD EESTIS: TEOORIA JA TEGELIKKUS Bakalaureusetöö Juhendaja:
SMS-TURUNDUSE KASUTAMINE EESTI ETTEVÕTETES
 TARTU ÜLIKOOL Majandusteaduskond Ettevõttemajanduse instituut Turunduse õppetool Jana Jukina SMS-TURUNDUSE KASUTAMINE EESTI ETTEVÕTETES Bakalaureusetöö Juhendaja: assistent Tanel Mehine Tartu 2012 Soovitan
TARTU ÜLIKOOL Majandusteaduskond Ettevõttemajanduse instituut Turunduse õppetool Jana Jukina SMS-TURUNDUSE KASUTAMINE EESTI ETTEVÕTETES Bakalaureusetöö Juhendaja: assistent Tanel Mehine Tartu 2012 Soovitan
Inimese luteiniseeriva hormooni ja kooriongonadotropiini beeta-subühiku geeniperekond: struktuur ja potentsiaalne seotus korduvate spontaanabortidega
 TARTU ÜLIKOOL BIOLOOGIA-GEOGRAAFIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL LIINA NAGIRNAJA Inimese luteiniseeriva hormooni ja kooriongonadotropiini beeta-subühiku geeniperekond:
TARTU ÜLIKOOL BIOLOOGIA-GEOGRAAFIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL LIINA NAGIRNAJA Inimese luteiniseeriva hormooni ja kooriongonadotropiini beeta-subühiku geeniperekond:
Calcium aluminothermal production of niobium and mineral composition of the slag
 Proc. Estonian Acad. Sci. Chem., 2007, 56, 3, 142 156 Calcium aluminothermal production of niobium and mineral composition of the slag Valeri Gorkunov and Rein Munter * Department of Chemical Engineering,
Proc. Estonian Acad. Sci. Chem., 2007, 56, 3, 142 156 Calcium aluminothermal production of niobium and mineral composition of the slag Valeri Gorkunov and Rein Munter * Department of Chemical Engineering,
CGGBP1 seondumissaidid inimese genoomis ja mõju rakutsüklile
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Kristine Kreitsman CGGBP1 seondumissaidid inimese genoomis ja mõju rakutsüklile Magistritöö
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Kristine Kreitsman CGGBP1 seondumissaidid inimese genoomis ja mõju rakutsüklile Magistritöö
This document is a preview generated by EVS
 EESTI STANDARD EVS-EN 1396:2007 Alumiinium ja alumiiniumisulamid. Rullis olevad pinnakattega lehed ja ribad üldotstarbeliseks kasutamiseks. Tehnilised nõuded Aluminium and aluminium alloys - Coil coated
EESTI STANDARD EVS-EN 1396:2007 Alumiinium ja alumiiniumisulamid. Rullis olevad pinnakattega lehed ja ribad üldotstarbeliseks kasutamiseks. Tehnilised nõuded Aluminium and aluminium alloys - Coil coated
PERSONAALBRÄNDING LINKEDIN-I JA ISIKLIKU KODULEHE KAUDU
 TARTU ÜLIKOOL Majandusteaduskond Silver Tilk PERSONAALBRÄNDING LINKEDIN-I JA ISIKLIKU KODULEHE KAUDU Bakalaureusetöö Juhendaja: Marge Täks Tartu 2016 Soovitan suunata kaitsmisele.. (juhendaja allkiri)
TARTU ÜLIKOOL Majandusteaduskond Silver Tilk PERSONAALBRÄNDING LINKEDIN-I JA ISIKLIKU KODULEHE KAUDU Bakalaureusetöö Juhendaja: Marge Täks Tartu 2016 Soovitan suunata kaitsmisele.. (juhendaja allkiri)
Microsoft Dynamics NAV (Microsoft Navision manufacturing introduction)
") Microsoft Dynamics NAV (Microsoft Navision manufacturing introduction) Microsoft Navision manufacturing What is the purpose of calculation in manufacturing? Today performed ACTIVITIES In future performed
Microsoft Dynamics NAV (Microsoft Navision manufacturing introduction) Microsoft Navision manufacturing What is the purpose of calculation in manufacturing? Today performed ACTIVITIES In future performed
Facebooki reklaamikampaania protsess
 Facebooki reklaamikampaania protsess Kasvata sihtgruppi, kontakte ja müüki Merlis Jusupov merlis@kodulehekoolitused.ee Probleem Milline on edukas FB reklaam? Sisu ja disain Sihitud õigele sihtgrupile FB
Facebooki reklaamikampaania protsess Kasvata sihtgruppi, kontakte ja müüki Merlis Jusupov merlis@kodulehekoolitused.ee Probleem Milline on edukas FB reklaam? Sisu ja disain Sihitud õigele sihtgrupile FB
Proceedings of the Estonian Academy of Sciences, 2017, 66, 4,
 Proceedings of the Estonian Academy of Sciences, 4017, 66, 2, Proceedings of the Estonian Academy of Sciences, 2017, 66, 4, 450 454 https://doi.org/10.3176/proc.2017.4.16 Available online at www.eap.ee/proceedings
Proceedings of the Estonian Academy of Sciences, 4017, 66, 2, Proceedings of the Estonian Academy of Sciences, 2017, 66, 4, 450 454 https://doi.org/10.3176/proc.2017.4.16 Available online at www.eap.ee/proceedings
MIKROVETIKATEST BIOGAASI TOOTMISE POTENTSIAAL EESTIS
 Tartu Ülikool Loodus- ja tehnoloogiateaduskond Ökoloogia ja Maateaduste Instituut Geograafia osakond Magistritöö keskkonnatehnoloogias ökotehnoloogia suunal MIKROVETIKATEST BIOGAASI TOOTMISE POTENTSIAAL
Tartu Ülikool Loodus- ja tehnoloogiateaduskond Ökoloogia ja Maateaduste Instituut Geograafia osakond Magistritöö keskkonnatehnoloogias ökotehnoloogia suunal MIKROVETIKATEST BIOGAASI TOOTMISE POTENTSIAAL
Kõrge olulisusega ravimi kõrvaltoimeid põhjustavate farmakogeneetiliste seoste replikatsioon Eesti Geenivaramu andmetel
 TARTU ÜLIKOOL LOODUS- JA TÄPPISTEADUSTE VALDKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Kõrge olulisusega ravimi kõrvaltoimeid põhjustavate farmakogeneetiliste seoste replikatsioon
TARTU ÜLIKOOL LOODUS- JA TÄPPISTEADUSTE VALDKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL Kõrge olulisusega ravimi kõrvaltoimeid põhjustavate farmakogeneetiliste seoste replikatsioon
Mis on sünteetiline bioloogia? Mart Loog Tehnoloogiainstituut
 Mis on sünteetiline bioloogia? Mart Loog Tehnoloogiainstituut Looduse ümberprogrammeerimine Reprogramming Life Sekvenaator Süntesaator Alused DNA sekveneerimise võimekuse kasv ja hinna langus
Mis on sünteetiline bioloogia? Mart Loog Tehnoloogiainstituut Looduse ümberprogrammeerimine Reprogramming Life Sekvenaator Süntesaator Alused DNA sekveneerimise võimekuse kasv ja hinna langus
AS CHEMI-PHARM TARNEAHELA JA SELLE JUHTMISE ARENDAMISE VÕIMALUSED
 TARTU ÜLIKOOL Majandusteaduskond Ettevõttemajanduse instituut Rahvusvahelise ettevõtluse ja innovatsiooni õppetool Teedu Pedaru AS CHEMI-PHARM TARNEAHELA JA SELLE JUHTMISE ARENDAMISE VÕIMALUSED Magistritöö
TARTU ÜLIKOOL Majandusteaduskond Ettevõttemajanduse instituut Rahvusvahelise ettevõtluse ja innovatsiooni õppetool Teedu Pedaru AS CHEMI-PHARM TARNEAHELA JA SELLE JUHTMISE ARENDAMISE VÕIMALUSED Magistritöö
Vea haldus ja logiraamat hajutatud süsteemides Enn Õunapuu.
 Vea haldus ja logiraamat hajutatud süsteemides Enn Õunapuu enn@cc.ttu.ee Millest tuleb jutt? Kuidas ma näen, millises sammus erinevad protsessid parasjagu on? Kuidas ma aru saan, kas protsess töötab või
Vea haldus ja logiraamat hajutatud süsteemides Enn Õunapuu enn@cc.ttu.ee Millest tuleb jutt? Kuidas ma näen, millises sammus erinevad protsessid parasjagu on? Kuidas ma aru saan, kas protsess töötab või
This document is a preview generated by EVS
 EESTI STANDARD TAHKEKÜTUSED Kloori määramine Eschka segu abil Solid mineral fuels Determination of chlorine using Eschka mixture (ISO 587:1997, modified) EESTI STANDARDI EESSÕNA NATIONAL FOREWORD See Eesti
EESTI STANDARD TAHKEKÜTUSED Kloori määramine Eschka segu abil Solid mineral fuels Determination of chlorine using Eschka mixture (ISO 587:1997, modified) EESTI STANDARDI EESSÕNA NATIONAL FOREWORD See Eesti
LÄMMASTIKVÄETISE MÕJUST VIKI-NISU SEGUKÜLVIDES
 260 LÄMMASTIKVÄETISE MÕJUST VIKI-NISU SEGUKÜLVIDES ABSTRACT. The effect of nitrogen fertilisation on vetch-wheat mixed crops. The study carried out on the experimental fields of the Department Field Crop
260 LÄMMASTIKVÄETISE MÕJUST VIKI-NISU SEGUKÜLVIDES ABSTRACT. The effect of nitrogen fertilisation on vetch-wheat mixed crops. The study carried out on the experimental fields of the Department Field Crop
(Candidate Gene Selection Protocol for Pig cdna Chip Manufacture Using TIGR Gene Indices)
") (Candidate Gene Selection Protocol for Pig Chip Manufacture Using TIGR Gene Indices) Chip Chip Chip Red Hat Linux 80 MySQL Perl Script TIGR(The Institute for Genome Research http://wwwtigrorg) SsGI (Sus
(Candidate Gene Selection Protocol for Pig Chip Manufacture Using TIGR Gene Indices) Chip Chip Chip Red Hat Linux 80 MySQL Perl Script TIGR(The Institute for Genome Research http://wwwtigrorg) SsGI (Sus
TARTU ÜLIKOOL BIOLOOGIA-GEOGRAAFIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL TARMO PUURAND
 TARTU ÜLIKOOL BIOLOOGIA-GEOGRAAFIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL TARMO PUURAND Rekombinatsioonipunktide jaotus inimese 19. ja 22. kromosoomis Magistritöö Juhendaja:
TARTU ÜLIKOOL BIOLOOGIA-GEOGRAAFIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT BIOTEHNOLOOGIA ÕPPETOOL TARMO PUURAND Rekombinatsioonipunktide jaotus inimese 19. ja 22. kromosoomis Magistritöö Juhendaja:
SAMAARIUMIGA AKTIVEERITUD TIO 2 MATERJALIDE LUMINESTSENTSOMADUSTE UURIMINE JA SOOL-GEEL VALMISTUSMEETODI OPTIMEERIMINE
 TARTU ÜLIKOOL Loodus- ja tehnoloogiateaduskond Materjaliteaduse instituut MARI ŠAVEL SAMAARIUMIGA AKTIVEERITUD TIO 2 MATERJALIDE LUMINESTSENTSOMADUSTE UURIMINE JA SOOL-GEEL VALMISTUSMEETODI OPTIMEERIMINE
TARTU ÜLIKOOL Loodus- ja tehnoloogiateaduskond Materjaliteaduse instituut MARI ŠAVEL SAMAARIUMIGA AKTIVEERITUD TIO 2 MATERJALIDE LUMINESTSENTSOMADUSTE UURIMINE JA SOOL-GEEL VALMISTUSMEETODI OPTIMEERIMINE
TARTU ÜLIKOOL ÕIGUSTEADUSKOND ERAÕIGUSE INSTITUUT AKTSIASELTSI JUHTIMISMUDELI ÕIGUSLIK REGULEERIMINE
 TARTU ÜLIKOOL ÕIGUSTEADUSKOND ERAÕIGUSE INSTITUUT Margit Vutt AKTSIASELTSI JUHTIMISMUDELI ÕIGUSLIK REGULEERIMINE Magistritöö Juhendaja professor Paul Varul Tartu 2006 Sissejuhatus...3 1. Ühingujuhtimise
TARTU ÜLIKOOL ÕIGUSTEADUSKOND ERAÕIGUSE INSTITUUT Margit Vutt AKTSIASELTSI JUHTIMISMUDELI ÕIGUSLIK REGULEERIMINE Magistritöö Juhendaja professor Paul Varul Tartu 2006 Sissejuhatus...3 1. Ühingujuhtimise
Annely Soosaar: Audentese finantsjuht. peakasutajast, kes tunneb ettevõtte tegevust ja tarkvara võimalusi. 5 soovitust
 Ärijuhtimistarkvara aitab pealinna kütta Kus peitub teeninduskvaliteedi parandamise võti 5 soovitust ERP hanke läbiviimiseks LK 8 LK 10 LK 26 ÄRIRAKENDUSTE IT AJAKIRI kevad 2012 Audentese finantsjuht Annely
Ärijuhtimistarkvara aitab pealinna kütta Kus peitub teeninduskvaliteedi parandamise võti 5 soovitust ERP hanke läbiviimiseks LK 8 LK 10 LK 26 ÄRIRAKENDUSTE IT AJAKIRI kevad 2012 Audentese finantsjuht Annely
EESTI STANDARD EVS-ISO :2001
 EESTI STANDARD EVS-ISO 8586-1:2001 SENSOORNE ANALÜÜS Üldine juhend assessorite valikuks, koolitamiseks ja jälgimiseks Osa 1: Valitud assessorid Sensory analysis General guidance for the selection, training
EESTI STANDARD EVS-ISO 8586-1:2001 SENSOORNE ANALÜÜS Üldine juhend assessorite valikuks, koolitamiseks ja jälgimiseks Osa 1: Valitud assessorid Sensory analysis General guidance for the selection, training
NEGFRY KARTULI-LEHEMÄDANIKU TÕRJEPROGRAMM
 96 NEGFRY KARTULI-LEHEMÄDANIKU TÕRJEPROGRAMM ABSTRACT. NEGFRY a program for control of potato late blight. One of the most serious diseases of the potato crop worldwide is late blight caused by Phytophthora
96 NEGFRY KARTULI-LEHEMÄDANIKU TÕRJEPROGRAMM ABSTRACT. NEGFRY a program for control of potato late blight. One of the most serious diseases of the potato crop worldwide is late blight caused by Phytophthora
EUROOPA PARLAMENT. Põllumajanduse ja maaelu arengu komisjon. Esitaja: põllumajanduse ja maaelu arengu komisjon
 EUROOPA PARLAMENT 2004 2009 Põllumajanduse ja maaelu arengu komisjon 2003/0210(COD) 2.2.2005 ARVAMUS Esitaja: põllumajanduse ja maaelu arengu komisjon Saaja: keskkonna-, rahvatervise- ja toiduohutuse komisjon
EUROOPA PARLAMENT 2004 2009 Põllumajanduse ja maaelu arengu komisjon 2003/0210(COD) 2.2.2005 ARVAMUS Esitaja: põllumajanduse ja maaelu arengu komisjon Saaja: keskkonna-, rahvatervise- ja toiduohutuse komisjon
PERSONALI TÖÖRAHULOLU JA MOTIVATSIOON HOTELL LONDON OÜ NÄITEL
 TARTU ÜLIKOOL Pärnu kolledž Turismiosakond Helena Murula PERSONALI TÖÖRAHULOLU JA MOTIVATSIOON HOTELL LONDON OÜ NÄITEL Lõputöö Juhendaja: Heli Tooman, PhD Pärnu 2016 Soovitan suunata kaitsmisele... (juhendaja
TARTU ÜLIKOOL Pärnu kolledž Turismiosakond Helena Murula PERSONALI TÖÖRAHULOLU JA MOTIVATSIOON HOTELL LONDON OÜ NÄITEL Lõputöö Juhendaja: Heli Tooman, PhD Pärnu 2016 Soovitan suunata kaitsmisele... (juhendaja
PEIPSI JÄRVE VEE OPTILISED OMADUSED
 TARTU ÜLIKOOL Füüsika-keemiateaduskond Keskkonnafüüsika instituut BIRGOT PAAVEL PEIPSI JÄRVE VEE OPTILISED OMADUSED Magistritöö keskkonnafüüsika erialal Juhendajad: ANU REINART, Ph.D Uppsala Ülikooli teadur
TARTU ÜLIKOOL Füüsika-keemiateaduskond Keskkonnafüüsika instituut BIRGOT PAAVEL PEIPSI JÄRVE VEE OPTILISED OMADUSED Magistritöö keskkonnafüüsika erialal Juhendajad: ANU REINART, Ph.D Uppsala Ülikooli teadur
Workplace performance analysis: methods and a system
 558 Proceedings of the Estonian Academy of Sciences, 2015, 64, 4S, 558 566 doi: 10.3176/proc.2015.4S.03 Available online at www.eap.ee/proceedings Workplace performance analysis: methods and a system Jaak
558 Proceedings of the Estonian Academy of Sciences, 2015, 64, 4S, 558 566 doi: 10.3176/proc.2015.4S.03 Available online at www.eap.ee/proceedings Workplace performance analysis: methods and a system Jaak
Some exploitation properties of wood plastic composites based on polypropylene and birch plywood sanding dust
 Proceedings of the Estonian Academy of Sciences, 4017, 66, 2, Proceedings of the Estonian Academy of Sciences, 2018, 67, 2, 117 123 https://doi.org/10.3176/proc.2018.2.01 Available online at www.eap.ee/proceedings
Proceedings of the Estonian Academy of Sciences, 4017, 66, 2, Proceedings of the Estonian Academy of Sciences, 2018, 67, 2, 117 123 https://doi.org/10.3176/proc.2018.2.01 Available online at www.eap.ee/proceedings
Materjalirühmade iseloomustus
 Materjalirühmade iseloomustus (uuendatud aprill 2010) Sisukord Keraamika...2 Klaas...3 Puit...4 Paber ja kartong...5 Regenereeritud tsellulooskile...6 Metallid...7 a) Alumiinium (Al)...8 b) Teras...9 c)
Materjalirühmade iseloomustus (uuendatud aprill 2010) Sisukord Keraamika...2 Klaas...3 Puit...4 Paber ja kartong...5 Regenereeritud tsellulooskile...6 Metallid...7 a) Alumiinium (Al)...8 b) Teras...9 c)
Testimise parendamise meetodid andmeaidas
 TALLINNA TEHNIKAÜLIKOOL Infotehnoloogia teaduskond Informaatikainstituut Tarkvaratehnika õppetool Testimise parendamise meetodid andmeaidas Bakalaureusetöö Üliõpilane: Üliõpilaskood: Juhendajad: Helen
TALLINNA TEHNIKAÜLIKOOL Infotehnoloogia teaduskond Informaatikainstituut Tarkvaratehnika õppetool Testimise parendamise meetodid andmeaidas Bakalaureusetöö Üliõpilane: Üliõpilaskood: Juhendajad: Helen
Energiajookide uuring
 Tervise Arengu Instituut Energiajookide uuring Raport Tagli Pitsi, Natalja Gluškova, Madli Martverk, Leila Oja, Krystiine Liiv Tallinn 2013 SISUKORD Sissejuhatus... 6 1. Taustinformatsioon energiajookidest...
Tervise Arengu Instituut Energiajookide uuring Raport Tagli Pitsi, Natalja Gluškova, Madli Martverk, Leila Oja, Krystiine Liiv Tallinn 2013 SISUKORD Sissejuhatus... 6 1. Taustinformatsioon energiajookidest...
UNDERSTANDING E-DEMOCRACY: KEY FEATURES TO INCREASE CITIZEN PARTICIPATION IN E-PARTICIPATORY BUDGETING PROJECTS
 TALLINN UNIVERSITY OF TECHNOLOGY School of Information Technologies Anders Røsten Mærøe 156411IVGM UNDERSTANDING E-DEMOCRACY: KEY FEATURES TO INCREASE CITIZEN PARTICIPATION IN E-PARTICIPATORY BUDGETING
TALLINN UNIVERSITY OF TECHNOLOGY School of Information Technologies Anders Røsten Mærøe 156411IVGM UNDERSTANDING E-DEMOCRACY: KEY FEATURES TO INCREASE CITIZEN PARTICIPATION IN E-PARTICIPATORY BUDGETING
Nitrogen removal from wastewater by heterotrophic denitrification with nitrite used as an electron acceptor
 Proc. Estonian Acad. Sci. Chem., 003, 5, 3, 10 19 Nitrogen removal from wastewater by heterotrophic denitrification with nitrite used as an electron acceptor Taimar Ala, Helen Sulg, Kaja Orupõld, and Toomas
Proc. Estonian Acad. Sci. Chem., 003, 5, 3, 10 19 Nitrogen removal from wastewater by heterotrophic denitrification with nitrite used as an electron acceptor Taimar Ala, Helen Sulg, Kaja Orupõld, and Toomas
COMPARISON OF ADVANCED OXIDATION PROCESSES FOR THE DESTRUCTION OF 2,4-DINITROPHENOL
 Proc. Estonian Acad. Sci. Chem., 21, 5, 1, 5 17 COMPARISON OF ADVANCED OXIDATION PROCESSES FOR THE DESTRUCTION OF 2,4-DINITROPHENOL Anna GOI and Marina TRAPIDO Department of Environmental Chemistry and
Proc. Estonian Acad. Sci. Chem., 21, 5, 1, 5 17 COMPARISON OF ADVANCED OXIDATION PROCESSES FOR THE DESTRUCTION OF 2,4-DINITROPHENOL Anna GOI and Marina TRAPIDO Department of Environmental Chemistry and
Trends in domestic hot water consumption in Estonian apartment buildings
 Proc. Estonian Acad. Sci. Eng., 26, 12, 1, 72 8 Trends in domestic ot water consumption in Estonian apartment buildings Teet-Andrus Kõiv and Alvar Toode Department of Environmental Engineering, Tallinn
Proc. Estonian Acad. Sci. Eng., 26, 12, 1, 72 8 Trends in domestic ot water consumption in Estonian apartment buildings Teet-Andrus Kõiv and Alvar Toode Department of Environmental Engineering, Tallinn
Ebola vaktsiin on saanud kättesaadavaks rekordkiirusel
 Ebola vaktsiin on saanud kättesaadavaks rekordkiirusel Rein Sikut Eesti Arst 206; 95(5):294 298 Saabunud toimetusse: 5.2.205 Avaldamiseks vastu võetud:.04.206 Avaldatud internetis: 27.05.206 GlaxoSmithKline
Ebola vaktsiin on saanud kättesaadavaks rekordkiirusel Rein Sikut Eesti Arst 206; 95(5):294 298 Saabunud toimetusse: 5.2.205 Avaldamiseks vastu võetud:.04.206 Avaldatud internetis: 27.05.206 GlaxoSmithKline
Ku i d a s h i n n a t a g m o d e m ŏ j u i n i m e s t e l e
 Ku i d a s h i n n a t a g m o d e m ŏ j u i n i m e s t e l e ja loodusele Väljaandjad Keskkonnaministeerium ja Tallinna Tehnikaülikool 2008 Kaasrahastanud ÜRO Keskkonnaprogramm Koostajad prof Erkki Truve,
Ku i d a s h i n n a t a g m o d e m ŏ j u i n i m e s t e l e ja loodusele Väljaandjad Keskkonnaministeerium ja Tallinna Tehnikaülikool 2008 Kaasrahastanud ÜRO Keskkonnaprogramm Koostajad prof Erkki Truve,
Transkriptsiooni ja transkriptsiooniga seotud DNA reparatsiooni mõju mutatsiooniprotsessidele bakteris Pseudomonas putida
 TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT GENEETIKA ÕPPETOOL Kärt Ukkivi Transkriptsiooni ja transkriptsiooniga seotud DNA reparatsiooni mõju mutatsiooniprotsessidele
TARTU ÜLIKOOL LOODUS- JA TEHNOLOOGIATEADUSKOND MOLEKULAAR- JA RAKUBIOLOOGIA INSTITUUT GENEETIKA ÕPPETOOL Kärt Ukkivi Transkriptsiooni ja transkriptsiooniga seotud DNA reparatsiooni mõju mutatsiooniprotsessidele
This document is a preview generated by EVS
 EESTI STANDARD EVS-EN ISO 9509:2006 Vee kvaliteet. Meetod kemikaalide ja heitvee pidurdava toime hindamiseks aktiivmudas mikroorganismidest põhjustatud nitrifitseerumisele Water quality - Toxicity test
EESTI STANDARD EVS-EN ISO 9509:2006 Vee kvaliteet. Meetod kemikaalide ja heitvee pidurdava toime hindamiseks aktiivmudas mikroorganismidest põhjustatud nitrifitseerumisele Water quality - Toxicity test
TÖÖSTUSKANEPI OMADUSTE UURIMINE JA KASUTUSVÕIMALUSED EHITUSES STUDYING THE PROPERTIES OF INDUSTRIAL HEMP AND ITS USABILITY IN CONSTRUCTION
 EESTI MAAÜLIKOOL Metsandus- ja maaehitusinstituut Maaehituse osakond Kristo Anslan TÖÖSTUSKANEPI OMADUSTE UURIMINE JA KASUTUSVÕIMALUSED EHITUSES STUDYING THE PROPERTIES OF INDUSTRIAL HEMP AND ITS USABILITY
EESTI MAAÜLIKOOL Metsandus- ja maaehitusinstituut Maaehituse osakond Kristo Anslan TÖÖSTUSKANEPI OMADUSTE UURIMINE JA KASUTUSVÕIMALUSED EHITUSES STUDYING THE PROPERTIES OF INDUSTRIAL HEMP AND ITS USABILITY
TTÜ Materjalitehnika instituut Metallide tehnoloogia õppetool. Deniss Tšernobajev
 TTÜ Materjalitehnika instituut Metallide tehnoloogia õppetool MTT70LT Deniss Tšernobajev Suurekiiruselise hapnik-kütuspihustuse teel saadud mehhanoaktiveeritud ja termosünteesitud Cr 3 C 2 -Ni pulberpinnete
TTÜ Materjalitehnika instituut Metallide tehnoloogia õppetool MTT70LT Deniss Tšernobajev Suurekiiruselise hapnik-kütuspihustuse teel saadud mehhanoaktiveeritud ja termosünteesitud Cr 3 C 2 -Ni pulberpinnete
TEETÖÖDE TEHNILINE KIRJELDUS
 TEETÖÖDE TEHNILINE KIRJELDUS Versioon: 12.02.2009-0102 1 Üldised 1 General Items 1.1 Lepingulised küsimused 1.1 Contractual items Sissejuhatus Käesolevad Tehnilised Töökirjeldused on ette nähtud Lepingu
TEETÖÖDE TEHNILINE KIRJELDUS Versioon: 12.02.2009-0102 1 Üldised 1 General Items 1.1 Lepingulised küsimused 1.1 Contractual items Sissejuhatus Käesolevad Tehnilised Töökirjeldused on ette nähtud Lepingu
METSARAIETE RUUMILISE PLANEERIMISE VÕIMALUSED SPATIAL PLANNING OPPORTUNITIES BY FOREST LOGGING
 EESTI MAAÜLIKOOL Metsandus- ja maaehitusinstituut Kalle Vislapuu METSARAIETE RUUMILISE PLANEERIMISE VÕIMALUSED SPATIAL PLANNING OPPORTUNITIES BY FOREST LOGGING Bakalaureusetöö Taastuvenergia ressursside
EESTI MAAÜLIKOOL Metsandus- ja maaehitusinstituut Kalle Vislapuu METSARAIETE RUUMILISE PLANEERIMISE VÕIMALUSED SPATIAL PLANNING OPPORTUNITIES BY FOREST LOGGING Bakalaureusetöö Taastuvenergia ressursside
MAJANDUSINFOSÜSTEEMIDE VAJADUSED EESTI MIKROETTEVÕTETES
 TARTU ÜLIKOOL Majandusteaduskond Ettevõttemajanduse instituut Majandusarvestuse õppetool Taavi Hõbejõgi MAJANDUSINFOSÜSTEEMIDE VAJADUSED EESTI MIKROETTEVÕTETES Magistritöö Juhendaja: lektor Sinaida Kalnin
TARTU ÜLIKOOL Majandusteaduskond Ettevõttemajanduse instituut Majandusarvestuse õppetool Taavi Hõbejõgi MAJANDUSINFOSÜSTEEMIDE VAJADUSED EESTI MIKROETTEVÕTETES Magistritöö Juhendaja: lektor Sinaida Kalnin
Species richness and range restricted species of liverworts in Europe and Macaronesia
 Folia Cryptog. Estonica, Fasc. 44: 143 149 (2008) Species richness and range restricted species of liverworts in Europe and Macaronesia Lars Söderström 1 & Ana Séneca 1,2 1 Department of Biology, Norwegian
Folia Cryptog. Estonica, Fasc. 44: 143 149 (2008) Species richness and range restricted species of liverworts in Europe and Macaronesia Lars Söderström 1 & Ana Séneca 1,2 1 Department of Biology, Norwegian
Mechanical and thermal properties of epoxy composite thermal insulators filled with silica aerogel and hollow glass microspheres
 Proceedings of the Estonian Academy of Sciences, 2017, 66, 2, Proceedings of the Estonian Academy of Sciences, 2017, 66, 4, 339 346 https://doi.org/10.3176/proc.2017.4.03 Available online at www.eap.ee/proceedings
Proceedings of the Estonian Academy of Sciences, 2017, 66, 2, Proceedings of the Estonian Academy of Sciences, 2017, 66, 4, 339 346 https://doi.org/10.3176/proc.2017.4.03 Available online at www.eap.ee/proceedings
PÕLENGUTE OSATÄHTSUS ÖKOSÜSTEEMIDES JA TAIMEDE KOHASTUMUSED TULEKAHJUDELE
 Tartu Ülikool Loodus- ja tehnoloogiateaduskond Ökoloogia ja Maateaduste Instituut Botaanika õppetool Heli Väljamets PÕLENGUTE OSATÄHTSUS ÖKOSÜSTEEMIDES JA TAIMEDE KOHASTUMUSED TULEKAHJUDELE Bakalaureusetöö
Tartu Ülikool Loodus- ja tehnoloogiateaduskond Ökoloogia ja Maateaduste Instituut Botaanika õppetool Heli Väljamets PÕLENGUTE OSATÄHTSUS ÖKOSÜSTEEMIDES JA TAIMEDE KOHASTUMUSED TULEKAHJUDELE Bakalaureusetöö
Effect of basalt and silica additives on erosive wear resistance of cast ceramics
 Proceedings of the Estonian Academy of Sciences, 2015, 64, 3, 1 9 Proceedings of the Estonian Academy of Sciences, 2016, 65, 2, 144 151 doi: 10.3176/proc.2016.2.05 Available online at www.eap.ee/proceedings
Proceedings of the Estonian Academy of Sciences, 2015, 64, 3, 1 9 Proceedings of the Estonian Academy of Sciences, 2016, 65, 2, 144 151 doi: 10.3176/proc.2016.2.05 Available online at www.eap.ee/proceedings
This document is a preview generated by EVS
 EESTI STANDARD EVS-EN ISO 8502-6:2006 Teraspindade ettevalmistamine enne värvide ja samalaadsete toodete pealekandmist. Pinna puhtuse hindamise katsed. Osa 6: Lahustuvate koostisosade ekstraheerimine analüüsimiseks.
EESTI STANDARD EVS-EN ISO 8502-6:2006 Teraspindade ettevalmistamine enne värvide ja samalaadsete toodete pealekandmist. Pinna puhtuse hindamise katsed. Osa 6: Lahustuvate koostisosade ekstraheerimine analüüsimiseks.
Predictive tools for the isothermal hardening of strip steel parts in molten salt
 Proceedings of the Estonian Academy of Sciences, 2015, 64, 3, 1 9 Proceedings of the Estonian Academy of Sciences, 2016, 65, 2, 152 158 doi: 10.3176/proc.2016.2.04 Available online at www.eap.ee/proceedings
Proceedings of the Estonian Academy of Sciences, 2015, 64, 3, 1 9 Proceedings of the Estonian Academy of Sciences, 2016, 65, 2, 152 158 doi: 10.3176/proc.2016.2.04 Available online at www.eap.ee/proceedings
This document is a preview generated by EVS
 EESTI STANDARD EVS-EN 1386:2007 Alumiinium ja alumiiniumisulamid. Astmeplaadid. Tehnilised nõuded Aluminium and aluminium alloys - Tread plate - Specifications EESTI STANDARDIKESKUS EESTI STANDARDI EESSÕNA
EESTI STANDARD EVS-EN 1386:2007 Alumiinium ja alumiiniumisulamid. Astmeplaadid. Tehnilised nõuded Aluminium and aluminium alloys - Tread plate - Specifications EESTI STANDARDIKESKUS EESTI STANDARDI EESSÕNA
TÖÖSTRESS JA TÖÖJÕU VOOLAVUS AS EESTI KESKKONNATEENUSED TARTU FILIAALI NÄITEL
 TARTU ÜLIKOOL Pärnu kolledž Ettevõtluse osakond Merili Tehvand TÖÖSTRESS JA TÖÖJÕU VOOLAVUS AS EESTI KESKKONNATEENUSED TARTU FILIAALI NÄITEL Lõputöö Juhendaja: assistent Liina Puusepp Pärnu 2014 SISUKORD
TARTU ÜLIKOOL Pärnu kolledž Ettevõtluse osakond Merili Tehvand TÖÖSTRESS JA TÖÖJÕU VOOLAVUS AS EESTI KESKKONNATEENUSED TARTU FILIAALI NÄITEL Lõputöö Juhendaja: assistent Liina Puusepp Pärnu 2014 SISUKORD
Sotsiaalmeedia turundus era- ja ärikliendi suunal Eesti ettevõtetes
 Tartu Ülikool Sotsiaal- ja haridusteaduskond Ajakirjanduse ja kommunikatsiooni instituut Sotsiaalmeedia turundus era- ja ärikliendi suunal Eesti ettevõtetes Bakalaureusetöö (6 EAP) Koostaja: Kristiina
Tartu Ülikool Sotsiaal- ja haridusteaduskond Ajakirjanduse ja kommunikatsiooni instituut Sotsiaalmeedia turundus era- ja ärikliendi suunal Eesti ettevõtetes Bakalaureusetöö (6 EAP) Koostaja: Kristiina
Influence of cellulose content on thermal properties of poly(lactic) acid/cellulose and low-density polyethylene/cellulose composites
 acid/cellulose and low-density polyethylene/cellulose composites") D. Šumigin et al.: Thermal properties of cellulose composites 237 Proceedings of the Estonian Academy of Sciences, 2012, 61, 3, 237 244 doi: 10.3176/proc.2012.3.14 Available online at www.eap.ee/proceedings
D. Šumigin et al.: Thermal properties of cellulose composites 237 Proceedings of the Estonian Academy of Sciences, 2012, 61, 3, 237 244 doi: 10.3176/proc.2012.3.14 Available online at www.eap.ee/proceedings
TEETÖÖDE TEHNILINE KIRJELDUS
 TEETÖÖDE TEHNILINE KIRJELDUS Versioon: 13.08.2009-0208 1 Üldised 1 General Items 1.1 Lepingulised küsimused 1.1 Contractual items Sissejuhatus Käesolevad Tehnilised Töökirjeldused on ette nähtud Lepingu
TEETÖÖDE TEHNILINE KIRJELDUS Versioon: 13.08.2009-0208 1 Üldised 1 General Items 1.1 Lepingulised küsimused 1.1 Contractual items Sissejuhatus Käesolevad Tehnilised Töökirjeldused on ette nähtud Lepingu
THE MORAL COMPETENCE OF AN EMERGING ORGANIZATION. Mari Meel and Maksim Saat Tallinn Tehnical University
 THE MORAL COMPETENCE OF AN EMERGING ORGANIZATION Mari Meel and Maksim Saat Tallinn Tehnical University The following paper is descriptive rather than normative: it mainly deals with factors shaping the
THE MORAL COMPETENCE OF AN EMERGING ORGANIZATION Mari Meel and Maksim Saat Tallinn Tehnical University The following paper is descriptive rather than normative: it mainly deals with factors shaping the
Tarkvara evolutsioon äriprotsesside modelleerimine ja automatiseerimine. Enn Õunapuu
 Tarkvara evolutsioon äriprotsesside modelleerimine ja automatiseerimine Enn Õunapuu enn@cc.ttu.ee Definitsioon Business Process Management (BPM) Gartner defines business process management (BPM) as a management
Tarkvara evolutsioon äriprotsesside modelleerimine ja automatiseerimine Enn Õunapuu enn@cc.ttu.ee Definitsioon Business Process Management (BPM) Gartner defines business process management (BPM) as a management
Implementation of robot welding cells using modular approach
 Estonian Journal of Engineering, 2010, 16, 4, 317 327 doi: 10.3176/eng.2010.4.07 Implementation of robot welding cells using modular approach Martinš Sarkans and Lembit Roosimölder Department of Machinery,
Estonian Journal of Engineering, 2010, 16, 4, 317 327 doi: 10.3176/eng.2010.4.07 Implementation of robot welding cells using modular approach Martinš Sarkans and Lembit Roosimölder Department of Machinery,
MULLA LIIKUVA MAGNEESIUMI SISALDUSE NING KALTSIUMI JA MAGNEESIUMI SUHTE MÕJU PÕLLUKULTUURIDE SAAGILE
 39 MULLA LIIKUVA MAGNEESIUMI SISALDUSE NING KALTSIUMI JA MAGNEESIUMI SUHTE MÕJU PÕLLUKULTUURIDE SAAGILE Taimed vajavad oma eluks suhteliselt suurtes kogustes kuut toiteelementi: lämmastikku, fosforit,
39 MULLA LIIKUVA MAGNEESIUMI SISALDUSE NING KALTSIUMI JA MAGNEESIUMI SUHTE MÕJU PÕLLUKULTUURIDE SAAGILE Taimed vajavad oma eluks suhteliselt suurtes kogustes kuut toiteelementi: lämmastikku, fosforit,
TALLINNA TEHNIKAÜLIKOOL Sotsiaalteaduskond Tööstuspsühholoogia instituut
 TALLINNA TEHNIKAÜLIKOOL Sotsiaalteaduskond Tööstuspsühholoogia instituut Mari-Ann Kruus PSÜHHOSOTSIAALSED OHUTEGURID TÖÖ KESKKONNAS JA NENDE MÕJU TÖÖTAJA PSÜHHOLOOGILISELE HEAOLULE Magistritöö Juhendaja:
TALLINNA TEHNIKAÜLIKOOL Sotsiaalteaduskond Tööstuspsühholoogia instituut Mari-Ann Kruus PSÜHHOSOTSIAALSED OHUTEGURID TÖÖ KESKKONNAS JA NENDE MÕJU TÖÖTAJA PSÜHHOLOOGILISELE HEAOLULE Magistritöö Juhendaja: