PLEKHM1 Regulates Autophagosome-Lysosome Fusion through HOPS Complex and LC3/GABARAP Proteins
|
|
|
- Benjamin Lloyd
- 6 years ago
- Views:
Transcription
1 Molecular Cell Article PLEKHM1 Regulates Autophagosome-Lysosome Fusion through HOPS Complex and LC3/GABARAP Proteins David G. McEwan, 1 Doris Popovic, 1 Andrea Gubas, 1,11 Seigo Terawaki, 2,3,4 Hironori Suzuki, 5 Daniela Stadel, 1 Fraser P. Coxon, 7 Diana Miranda de Stegmann, 7 Sagar Bhogaraju, 8 Karthik Maddi, 8 Anja Kirchof, 1 Evelina Gatti, 2,3,4 Miep H. Helfrich, 7 Soichi Wakatsuki, 6,10 Christian Behrends, 1 Philippe Pierre, 2,3,4 and Ivan Dikic 1,8,9, * 1 Institute of Biochemistry II, Goethe University School of Medicine, Theodor-Stern-Kai 7, D Frankfurt (Main), Germany 2 Centre d Immunologie de Marseille-Luminy, Aix-Marseille Université, UM2, Marseille, France 3 INSERM, U1104, Marseille, France 4 CNRS, UMR 7280, Marseille, France 5 Biomolecular Interaction Centre, School of Biological Sciences, University of Canterbury, Christchurch 8020, New Zealand 6 Structural Biology Research Center, Photon Factory, Institute of Materials Structure Science, High Energy Accelerator Research Organization (KEK), Tsukuba, Ibaraki , Japan 7 Musculoskeletal Research Programme, Division of Applied Medicine, Institute of Medical Sciences, University of Aberdeen, Foresterhill, Aberdeen AB25 2ZD, UK 8 Buchmann Institute for Molecular Life Sciences, Max-von-Laue-Str. 15, Goethe University Frankfurt am Main, Germany 9 University of Split, School of Medicine, Department of Immunology and Medical Genetics, Soltanska 2, Split, Croatia 10 Present address: SLAC National Accelerator Laboratory, Menlo Park, CA , USA 11 Present address: London Research Institute, Cancer Research UK, 44 Lincolns Inn Fields, London WC2A 3LY, UK *Correspondence: Ivan.Dikic@biochem2.de SUMMARY The lysosome is the final destination for degradation of endocytic cargo, plasma membrane constituents, and intracellular components sequestered by macroautophagy. Fusion of endosomes and autophagosomes with the lysosome depends on the GTPase Rab7 and the homotypic fusion and protein sorting (HOPS) complex, but adaptor proteins that link endocytic and autophagy pathways with lysosomes are poorly characterized. Herein, we show that Pleckstrin homology domain containing protein family member 1 (PLEKHM1) directly interacts with HOPS complex and contains a LC3-interacting region (LIR) that mediates its binding to autophagosomal membranes. Depletion of PLEKHM1 blocks lysosomal degradation of endocytic (EGFR) cargo and enhances presentation of MHC class I molecules. Moreover, genetic loss of PLEKHM1 impedes autophagy flux upon mtor inhibition and PLEKHM1 regulates clearance of protein aggregates in an autophagy- and LIR-dependent manner. PLEKHM1 is thus a multivalent endocytic adaptor involved in the lysosome fusion events controlling selective and nonselective autophagy pathways. INTRODUCTION Cellular membrane trafficking is regulated by small GTPases of the Rab, Arf, and Arl families, which cycle between inactive (GDP) and active (GTP) conformation states. In their active form, GTPases are anchored to intracellular vesicles and organelles, subsequently regulating recruitment of various effector proteins and tethering factors, thus mediating membrane fusion and movement of vesicles (reviewed in Zerial and McBride, 2001). Importantly, endocytosis and autophagy are both processes that regulate delivery of macromolecules to the lysosome for their degradation. A close relationship between autophagy and endocytic degradative pathways is evident in the dual role of numerous protein regulatory machineries, one of which is Rab7, the late endosomal/lysosomal Rab GTPase (Stenmark, 2009). Specifically, Rab7 is an essential GTPase that defines the integrity of the late endosomal/lysosomal compartment, regulating growth-factor receptor degradation (Ceresa and Bahr, 2006) and cation-independent Mannose-6-phosphate receptor (CI-M6PR) retrieval to the trans-golgi network, required for the sorting of acidic hydrolases and formation of functional lysosomes (Press et al., 1998). In conditions where lysosomal biogenesis or delivery is impaired, endocytic and autophagic cargo remains trapped in the late endosomes/autophagosomes and is inefficiently degraded, leading to various pathological conditions such as neurodegeneration or lysosomal storage diseases (Nixon, 2013). Multiple downstream effectors of Rab7 have been characterized, such as Rab7-interacting lysosomal protein (RILP) (Cantalupo et al., 2001) and FYVE and coiledcoil containing 1 (FYCO1) (Pankiv et al., 2010), which mediate fusion of late endosomes and lysosomes and their transport along microtubules, respectively. Fusion of endocytic vesicles is driven by tethering complexes that bring the vesicles together and position them into close proximity of the actual fusion machinery. Tethering complexes, such as homotypic fusion and vacuole protein sorting (HOPS) and class C core vacuole/endosome tethering (CORVET), directly recruit SNARE proteins to the late (HOPS) or early (CORVET) endosomes (Balderhaar and Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 39
2 Ungermann, 2013) and drive vesicle fusion. HOPS/CORVET consist of a core class C Vps complex including Vps11, Vps16, Vps18, and Vps33 and two variable subunits that are characteristic for HOPS (Vps39 and Vps41) or CORVET (Vps3 and Vps8) (Balderhaar and Ungermann, 2013). HOPS complex subunit Vps39 additionally functions as a GDP exchange factor for Rab7, thus increasing the pools of active Rab7 at the endosomal membrane (Wurmser et al., 2000). Notably, while RILP and HOPS are known to regulate endosome-lysosome fusion, Rab7 regulates the terminal autophagosome-lysosome fusion step (Gutierrez et al., 2004). Thus far, only a few Rab7 binding proteins have been described in the context of autophagy. RUN domain protein as Beclin1 interacting and cysteine-rich containing (Rubicon) and UV radiation resistance-associated gene (UVRAG) function together as components of Beclin1-PI3 kinase complex, regulating maturation of endosomes and autophagosomes (Matsunaga et al., 2009). UVRAG, on the other hand, associates with endosomes at both early (Rab5 positive) and late (Rab7 positive) stages (Liang et al., 2008) and is therefore implicated at earlier stages of endosomal maturation, characterized by Rab5 to Rab7 conversion and subsequent HOPS recruitment. The specific effectors of Rab7 that regulate the terminal stage of autolysosome formation remain unknown. Here, we characterize a Rab7 effector protein PLEKHM1 that we initially discovered as an LC3/GABARAP interacting protein. Using a Stable isotope labeling of amino acids in culture (SILAC)-based mass spectrometry approach, we found that PLEKHM1 interacted directly with HOPS complex via its RUN domain and colocalized at vesicle contact sites with Rab7 and HOPS. Loss of PLEKHM1 blocked epidermal growth factor receptor (EGFR) degradation, and Plekhm1 KO mouse embryonic fibroblasts (MEFs) displayed a defect in autophagosome to autolysosome maturation. Moreover, PLEKHM1, and specifically the LC3-interacting region (LIR), was required for the autophagymediated clearance of protein aggregates. Thus, we propose that PLEKHM1 is a specific effector for the terminal fusion of autophagosome and lysosome through simultaneous binding to the LC3, Rab7, and subsequent recruitment of HOPS to the vesicle contact sites. RESULTS PLEKHM1 Directly Interacts with LC3/GABARAP Proteins through a Conserved LIR By using a yeast two-hybrid system, we have identified several LC3/GABARAP-binding partners, including NBR1 (Kirkin et al., 2009a), Nix/BNiP3L (Novak et al., 2010), TBC1D5 (Popovic et al., 2012; Popovic and Dikic, 2014), and Optineurin (Wild et al., 2011). Additionally, PLEKHM1 was also present in these screens; therefore, we aimed to investigate the role of PLEKHM1 in autophagy pathways. Endogenous PLEKHM1 from HeLa lysate was readily precipitated with GST-tagged LC3B and GST-GABARAP-L1 but not with GST alone (Figure 1A). PLEKHM1 interacted with all mammalian LC3/GABARAP isoforms and yeast Atg8, but not other ubiquitin-like proteins (Figure 1B). Upon sequence analysis, we discovered a potential LC3 interaction region (LIR) nestled between the Pleckstrin homology (PH) domains 1 (PH1) and 2 (PH2) of PLEKHM1 (Figure 1C). Only regions of GST-tagged PLEKHM1 that contained the identified LIR sequence (amino acids [aa] ) precipitated Flag epitope-tagged LC3/GABARAPs (Flag-LC3A, -LC3B, -GABARAP, and -GABARAP-L1) from cells (Figures S1A and S1B available online). To analyze the interaction between PLEKHM1-LIR and autophagy modifiers, we obtained high-quality crystals of a chimeric protein: PLEKHM1- LIR peptide fused to the N terminus of LC3B molecule (PDB entry 3X0W, Table 1). The asymmetric unit of PLEKHM1- LIR-LC3B consists of two molecules, and each molecule interacts with the PLEKHM1 LIR region of symmetry-related molecule (molecule A and B bind to symmetry-related molecule A and B, respectively) (Figures S1C, S1D, and S1F). The PLEKHM1-LIR peptide from 634 to 638 (EWVNV) was assigned in molecule A (Figure 1D) and from 635 to 638 (WVNV) in molecule B (Figures S1F and S1G). PLEKHM1-LIR (EWVNV) showed a typical interaction with LC3B hydrophobic surfaces (Ile23, Lys49, Phe52, and Ile66 Phe108) with Lys51 and Leu53 forming hydrogen bonds with the main-chain of PLEKHM1-LIR (Figures 1D and S1E), as compared to p62/lc3b LIR (PDB 2ZJD;Figure 1D, right panel). Mutation of PLEKHM1-LIR abolished interaction with all LC3/GABARAP family members (Figure 1E). PLEKHM1 is therefore a direct interaction partner of mammalian Atg8 proteins through a highly conserved W-type LIR motif (Rogov et al., 2014). PLEKHM1 Is an Autophagy Adaptor that Localizes to The Limiting Membrane of Autolysosomes The direct interaction of PLEKHM1 with LC3/GABARAP proteins hinted at a potential role in autophagy pathways. LC3/GABARAP proteins localize to both the inner and outer autophagosomal membranes. Autophagy receptors (e.g., p62, NBR1, and OPTN) simultaneously bind LC3 proteins and autophagy cargo, whereas autophagy adaptor molecules (e.g., FYCO1 and TBC1D5) interact with surface-conjugated LC3 and, most notably, are not degraded in an autophagy-dependent manner (Stolz et al., 2014). Thus, we set out to identify whether PLEKHM1 was acting as an autophagy receptor or adaptor molecule. In autophagy-deficient MEFs (Atg5 KO, Atg5 / ; Kuma et al., 2004), we observed no difference in the total protein levels of endogenous Plekhm1 protein, in contrast to both p62 and Nbr1 autophagy receptor proteins (Figure 2A). Moreover, in HeLa cells endogenous (Figure S2A) or overexpressed PLEKHM1 (Figure S2B) protein levels were not affected by lysosomal inhibition (BafA1) or after prolonged autophagy induction (6 hr EBSS or 6 hr mtor inhibition; Ku ). Thus, PLEKHM1 is not degraded in an autophagy-dependent manner and is likely acting as an autophagy adaptor protein. Stable overexpression of PLEKHM1 resulted in colocalization with LAMP1 under full media (FM) conditions (Figure S2C), and subsequent treatment with Ku+BafA1 (Bafilomycin A1) caused the formation of enlarged vesicles, presumably autolysosomes, as PLEKHM1 and LAMP1 localized to the outside of LC3B-positive structures (Figures S2C and S2D; associated line graphs). In accordance, using electron microscopy and immunogold labeling, PLEKHM1-GFP localized to the limiting membrane of enlarged autolysosome-like structures after BafA1 treatment (Figures 2B and 2C). To verify that the majority of PLEKHM1 40 Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.
 PLEKHM1-WT-Flag was overexpressed in HEK293T cells and a GST-pulldown performed using purified GST-tagged ubiquitin and ubiquitin-like proteins.")
3 Figure 1. PLEKHM1 Interacts Directly with Atg8 Ubiquitin-Like Modifiers (A) GST-pulldown of endogenous PLEKHM1 from HeLa cell lysate using purified GST-alone, GST-LC3B, and GST-GABARAPL1. (B) PLEKHM1-WT-Flag was overexpressed in HEK293T cells and a GST-pulldown performed using purified GST-tagged ubiquitin and ubiquitin-like proteins. (C) Schematic image of PLEKHM1 domain composition and sequence alignment of known LIR motifs (NBR1 aa ; OPTN aa ; NIX/BNIP3L aa 27 42; scatg3 aa ; Calreticulin [CRT] aa ; p62/sqstm1 aa ) with predicted PLEKHM1-LIR sequence that is conserved among species. Alignment created using Jalview (Waterhouse et al., 2009). Amino acids colored according to Clustal X scheme. (D) Solvent-excluded surface rendering of X-ray crystal structure (2.7 Å) of LC3B in complex with PLEKHM1 LIR (EWVNV) peptide (left panel; PDB: 3X0W) and of LC3B in complex with p62 LIR (DDDWTHLK) peptide (PDB:2ZJD) (right panel). Surface of LC3B is colored gray. Orange and yellow colored amino acids represent the hydrophobic interaction surfaces. White labeling represents LIR residues and numbers and black lettering LC3B residues. See also Table 1. (E) Lysates of HEK293T cells expressing either GFP-tagged PLEKHM1-WT or PLEKHM1-DLIR (Daa ) were incubated with purified GST alone or GST- LC3/GABARAP proteins. See also Figure S1. is on the limiting membrane, rather than the luminal side of autolysosomal vesicles, we performed a protease protection assay, where vesicular luminal proteins (e.g., inside autophagosomes, endosomes, MVBs, etc.) remain protected from Proteinase-Kmediated degradation. HeLa cells were stimulated with either DMSO or Ku+CQ to induce the accumulation of autophagic cargo; permeabilized with digitonin; and the membrane fractions incubated with either buffer alone ( ), proteinase K (PK), or PK+Triton X-100 (PK+TX100). Endogenous PLEKHM1 was degraded by approximately 70% 75% in both DMSO and Ku+CQ-treated cells, with no apparent protection from PK (Figure 2D). In contrast, Ku+CQ treatment enhanced p62 PK protection by approximately 50% (Figure 2D), indicating the presence of p62 within the vesicular lumen (Figure 2D). Next, we confirmed that endogenous Plekhm1 colocalized with LAMP1-positive vesicles in wild-type (WT) MEFs (Figures S2E and S2F). Moreover, Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 41
4 Table 1. Data Collection and Refinement Statistics Parameter PLEKHM1 LIR-LC3B (PDB code 3X0W) Data collection Space group P Cell dimensions a, b, c ( A) 69.0, 69.0, a, b, g ( ) 90.0, 90.0, 90.0 Resolution ( A) ( ) R merge (0.661) Mean I/sI 13.5 (3.1) Completeness (%) 92.9 (99.8) Redundancy 6.9 (7.2) Refinement Resolution ( A) Number of reflections 7343 R work /R free 0.224/0.294 Number of atoms Protein 2043 Water 25 Average B factors (Å 2 ) Protein 54.2 Water 33.3 Ramachandran plot Favored (%) 95.9 Allowed (%) 4.1 Outlier (%) 0 RMSD Bond length (Å) Bond angles ( ) Values in parentheses are for the highest-resolution shell. Ku+CQ treatment resulted in enlarged Plekhm1/LAMP1-positive vesicles that contained endogenous LC3B (Figures 2E and 2F). Thus, PLEKHM1 is an autophagy adaptor that localizes with LAMP proteins to the limiting autolysosomal membrane upon activation of autophagy. PLEKHM1 Interacts Directly with HOPS Complex To gain further insight into PLEKHM1 function, we performed a SILAC-based proteomic analysis in doxycycline-inducible PLEKHM1-GFP HeLa cells. PLEKHM1-WT-GFP expression was induced only in HEAVY labeled cells and precipitated using anti-gfp beads. A total of 68 interactors were enriched in the PLEKHM1 fraction after processing using a p value of 0.01 as a cut-off (Figure 3A, red circles). All six components of the HOPS complex, four of which were highly enriched (hvps39 (Vam6), hvps11, hvp16, and hvps33a; blue circles) were found in the complex with PLEKHM1 (Figure 3A). No CORVET-specific components (Vps3 or Vps8) were detected. Moreover, PLEKHM1-GFP precipitated with endogenous HOPS complex components (Figures 3B and 3C) and with overexpressed epitope-tagged HOPS (Figures 3D and 3E). In addition, interaction between endogenous PLEKHM1 and endogenous hvps11 were increased upon serum starvation (Figure 3F), indicating that stimulation of autophagy may enhance their interactions. Next, we purified full-length maltose binding protein (MBP)- tagged PLEKHM1 and tested the direct interaction with N- (aa 1 310) and C-terminal (aa ) fragments of hvps39. PLEKHM1 readily interacted directly with the C-terminal region of hvps39 (Figure 3G). We then tested which regions of PLEKHM1 could coprecipitate HOPS components from cells. Both HOPS-specific subunits, hvps39 and hvps41, interacted with the RUN domain (aa 1 198) but not with the PH2 domain of PLEKHM1 (Figure 3H). Using bacterially purified GST-RUN (aa ) or GST-PH2 (aa ) we confirmed binding of hvps39 (Figure 3I) and hvps41 (Figure 3J) only to the RUN domain. Finally, the HOPS complex is required for the recruitment of SNARE proteins that are responsible for mediating vesicle fusion. We therefore tested whether we could coprecipitate PLEKHM1 with SNARE proteins. PLEKHM1-WT-Flag was expressed alone (Ø) or with various GFP-tagged SNAREs involved in the late endocytic/autophagic compartments, treated for 4 hr with either DMSO or Ku , and GFP-tagged SNAREs were immunoprecipitated. Surprisingly, PLEKHM1 coprecipitated predominantly with GFP-SNAP29 and GFP-STX17 (Figure 3K). However, upon Ku treatment, PLEKHM1 coprecipitated with both STX17, the autophagy-specific SNARE (Itakura et al., 2012; Jiang et al., 2014) and endogenous hvps41 (Figure 3K). Thus, upon autophagy induction, PLEKHM1 can be recruited to the autophagosome-lysosome fusion machinery. The RUN Domain of PLEKHM1 Tethers Rab7 and hvps41 at Lysosome Contact Zones The relationship between PLEKHM1 and HOPS was further analyzed in U2OS cells stably expressing PLEKHM1-WT-GFP. Stable overexpression of PLEKHM1-GFP caused the formation of enlarged PLEKHM1/LAMP2/Rab7-positive lysosomes (Figure 4A). PLEKHM1 was particularly enriched at the contact zones between vesicles (Figure 4A, inset arrows). In addition, endogenous hvps41-positive puncta were adjacent to, or present on, PLEKHM1 vesicles (Figures 4B, S3A, and S3B). Next, we monitored the behavior of PLEKHM1-GFP and mcherry-hvps41 in double stable U2OS cells. Both proteins strongly colocalized upon overexpression and formed tight clusters of vesicles, presumably late endosomes/lysosomes (Figures 4C and 4D, box i). The majority of PLEKHM1-hVps41-positive vesicles that clustered at the perinuclear region were relatively immobile (Movie S1). However, smaller PLEKHM1/hVps41-positive vesicles were also able to bud and fuse with other vesicles (Figures 4C, box ii, and 4D; Movie S2). To assess the effect of the PLEKHM1-RUN domain on hvps41-positive vesicle localization, we overexpressed either GFP alone, PLEKHM1-WT-GFP, GFP-RUN, or GFP-PH2 domain in U20S cells stably expressing mcherry-hvps41. Both PLEKHM1-WT and the RUN domain, but not PH2 domain, efficiently colocalized with mcherry-hvps41 (Figures 4E and S3C) and caused clustering of mcherry-hvps41-positive vesicles (Figure 4E, inset). Deletion of the RUN domain inhibited PLEKHM1-vesicle clustering in stable U20S cells 42 Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.

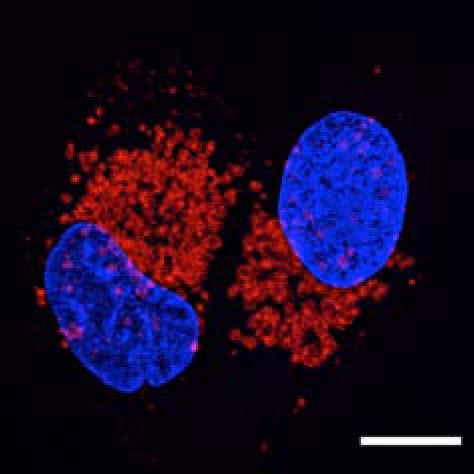
5 Figure 2. PLEKHM1 Localizes to the Limiting Membrane of Autolysosomes (A) Atg5 +/+ and Atg5 / MEFs were analyzed for total protein levels of Plekhm1. LC3B, p62, and Nbr1 were also probed using specific antibodies. Vinculin was used as a loading control. Densitometry analysis was used to measure the fold change in protein levels (Fold change versus Atg5 +/+, normalized to Vinculin). (B) HeLa cells transfected with PLEKHM1-GFP were treated with 100 nm Bafilomycin A1 for 5 hr. Cells were then harvested, pelleted, fixed in 4% paraformaldehyde (B) or 2.5% paraformaldehyde/0.2% glutaraldehyde in 0.1 M phosphate buffer (C), and then embedded in HM20 resin. Ultrathin sections (70nm) were immunolabeled with an anti-gfp antibody and protein A gold (10 nm) as described in Experimental Procedures. (D) HeLa cells were treated with either DMSO or Ku Chloroquine (Ku+CQ) for 4 hr; permeabilized with digitonin; and incubated in the buffer alone ( ), 100 ng/ml PK, or PK plus 0.1% Triton X-100 (PK+TX). Supernatant fraction (cytoplasmic release after digitonin treatment) is also shown. Samples were probed for endogenous PLEKHM1, p62, and Vps11, and percentage remaining was calculated based on buffer only sample. (E) WT MEFs were treated for 4 hr with Ku Chloroquine (Ku+CQ) and probed endogenous Plekhm1 (red), LAMP1 (magenta), and LC3B (green). For clarity, only Plekhm1 and LC3B signals are shown in the merged channel. Arrowheads indicate structures of interest. An enhancement of region in merge (E) is shown in (F). Scale bar is 20 mm unless otherwise stated. All images and blots are representative of at least n = 3 independent experiments. See also Figure S2. Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 43

6 Figure 3. PLEKHM1 Interacts Directly with the HOPS Complex (A) SILAC-based mass spectrometric analysis of PLEKHM1 interactome (red). Immunoprecipitation of GFP-tagged PLEKHM1 stably expressed in HeLa cells is shown. Bait-expressing cells were heavy labeled; cells expressing no bait were light labeled. Significantly enriched HOPS complex components are indicated by blue color dots. hvps18 (green) was below threshold. p value of 0.01 was used as threshold for significantly enriched interactors. PLEKHM1 is highlighted in orange. (legend continued on next page) 44 Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.
7 (Figure 4F). Further, LAMP2 + lysosome clustering, induced by serum starvation (Korolchuk et al., 2011), was not evident upon depletion of PLEKHM1 (Figures S3D and S3E). Since the RUN domain interacted with the HOPS complex, we concluded that PLEKHM1 could be one of the Rab7 effectors that recruit HOPS to the late endosomes and lysosomes and to test PLEKHM1 function in terminal degradation of endocytic and autophagic cargo. PLEKHM1 Loss Inhibits Ligand-Induced EGFR Degradation The interactions with the HOPS complex indicated that PLEKHM1 may also function in a classical endocytic pathway and is not only a specific autophagy adaptor. Perturbations within ligand-stimulated growth factor receptor endocytosis pathways commonly result in altered degradation dynamics (Sigismund et al., 2012). We therefore tested the effects of PLEKHM1 depletion on EGFR degradation. Both, stable (shrna; HeLa) and transient (sirna; Hke3) depletion of PLEKHM1 caused a significant time-dependent delay of EGFR degradation compared to controls (Figures 5A, 5B, S4A, and S4B). In addition, depletion of HOPS subunits hvps41 (Figure S4C) and hvps11 (Figure S4D) or Rab7 (Figure S4E) similarly impeded EGFR degradation (Figure S4F). Rhodamine-conjugated EGF (Rh-EGF) stimulation showed a similar delay in degradation dynamics in PLEKHM1-depleted cells (Figure S4G), while puncta of Rh-EGF at 120 min of internalization showed an increase in both number and size in PLEKHM1 knockdown cells compared to control (Figures 5C 5E). Upon PLEKHM1 depletion, Rh-EGF was retained within early endosomes, as demonstrated by colocalization with early endosomal antigen 1 (EEA1) (Figures 5F and 5G), while Transferrin receptor (TfnR) was unaffected by PLEKHM1 depletion (Figures S4H and S4I). Thus, we concluded that PLEKHM1 regulates a late stage of endosomal maturation and degradation of growth factor receptors similar to HOPS and Rab7, indicating a functional link between these molecules. Genetic Loss of Plekhm1 Inhibits Autophagy Flux In order to assess the effect of Plekhm1 loss on autophagy flux, we generated Plekhm1 KO MEFs. We confirmed the presence or absence of Plekhm1 by PCR (data not shown) and by western blot against Plekhm1 protein (Figures 6A and S2F). Plekhm1 +/+ and Plekhm1 / MEFs were immortalized using SV40 large T cell antigen (Tag), and we used them at an early passage number after immortalization for our autophagy flux experiments. First, we used several conditions to either stimulate autophagy (EBSS starvation media; Ku and Torin1) or to block autophagosome degradation (BafA1 or CQ). Under nonstimulated conditions (DMSO), we observed increased LC3B-II and p62 in Plekhm1 / MEFs compared to Plekhm1 +/+ (Figures 6A, 6B, and S5A), indicative of altered autophagic flux. Although we noticed a modest increase in LC3B- II in unstimulated and EBSS-treated Plekhm1 / MEFs, direct inhibition of mtor with either Ku or Torin1 resulted in a marked increase in LC3B-II and p62 levels compared to WT MEFs (Figure 6A, and quantified in Figure 6B). Treatment with either BafA1 (Figure S5A) or CQ (Figures 6A and 6B) resulted in equal levels of LC3B-II and p62 between the genotypes, indicating that Plekhm1 may influence the maturation of autophagosomes and not their formation. Next, we tested whether loss of Plekhm1 perturbs autophagosome to autolysosome maturation. Endogenous LC3B puncta increased in colocalization with LAMP1 under EBSS-mediated starvation conditions compared to nonstarved conditions (Figure 6C, upper panel and inset; Figure 6D, black bars). However, colocalization between LC3B and LAMP1 was significantly reduced in Plekhm1 / MEFs (Figure 6C, lower panels; Figure 6D, gray bars). Next, we used the tandem-fluorescence (mcherry-gfp) LC3B (tflc3b) as a readout of autophagy flux, where the GFP signal is quenched upon exposure to an acidic environment (lysosome). We quantified the number of red-positive/ green-negative puncta (autolysosome) over the red-positive/ green-positive puncta (autophagosome) and expressed it as percentage colocalization. Under autophagy-stimulated conditions (2 hr Ku ), Plekhm1 / MEFs displayed a decreased number of red only puncta (increased number of green+/red+ puncta; Figure 6E, and quantified in Figure 6F) compared to Plekhm1 +/+ MEFs. Thus, Plekhm1 is required for the terminal maturation of autophagosome to autolysosome, presumably through facilitating autophagosome-lysosome fusion. (B) HeLa-Flp-IN- TRex cells stably expressing PLEKHM1-GFP were induced for 16 hr and PLEKHM1-GFP was immunoprecipitated by GFP-Trap beads, and samples were immunoblotted for endogenous HOPS complex subunits (C E) (C) Immunoprecipitation of endogenous hvps11 from cells treated as described in (B). Sample was immunoblotted for endogenous HOPS subunits, and PLEKHM1-GFP (D) HEK293T cells were transfected with Flag-hVps39 (D) or Flag-hVps41 (E) together with PLEKHM1-GFP. Flag-tagged HOPS subunits were immunoprecipitated using Flag (M2) beads and samples were immunoblotted for PLEKHM1-GFP. Vinculin was used as a loading control and endogenous hvps11 as a marker for core HOPS complex. (F) Endogenous hvps11 was immunoprecipitated form HeLa cells under FM (10% serum FBS) or serum starvation (1% serum FBS) conditions. Samples were immunoblotted for endogenous PLEKHM1, hvps41, and hvps11 (G) Full-length MBP-tagged PLEKHM1 (MBP-PLEKHM1) or MBP alone was incubated with either hvps39 6x His-tagged N-terminal (aa 1 310) or C-terminal (aa ) fragments. Samples were blotted with Ant-His-tag, and the membrane was stained with Ponceau S. (H) Immunoprecipitation of GFP-tagged truncated versions of PLEKHM1 (aa 1 250; RUN domain [aa 1 198] and PH2 domain [aa ]) with either mcherryhvps41 (upper) or mcherry-hvps39 (lower). (I) Flag-hVps39 or Flag-hVps41 (J) subunits of HOPS were expressed in HEK293T cells, and lysates were incubated with bacterially purified GST alone, GST PLEKHM1-RUN domain (aa ), or GST-PLEKHM1-PH2 domain (aa ). Samples were immunoblotted with anti-flag antibody. (K) PLEKHM1-WT-Flag was either expressed alone (Ø) or coexpressed with GFP-tagged SNARE proteins (SNAP29, Vti1b, Syntaxin7 [STX7], VAMP7, or Syntaxin17 [STX17]). Cells were treated with either DMSO or Ku for 4 hr and anti-gfp Trap beads used to immunoprecipitate GFP-tagged SNAREs. Samples were probed with anti-flag, anti-gfp, and antivps41 for the presence of endogenous HOPS. Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 45
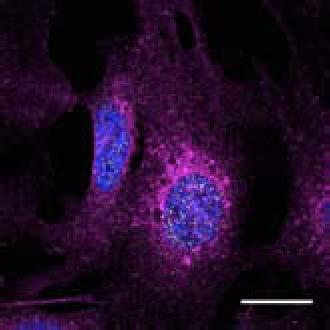
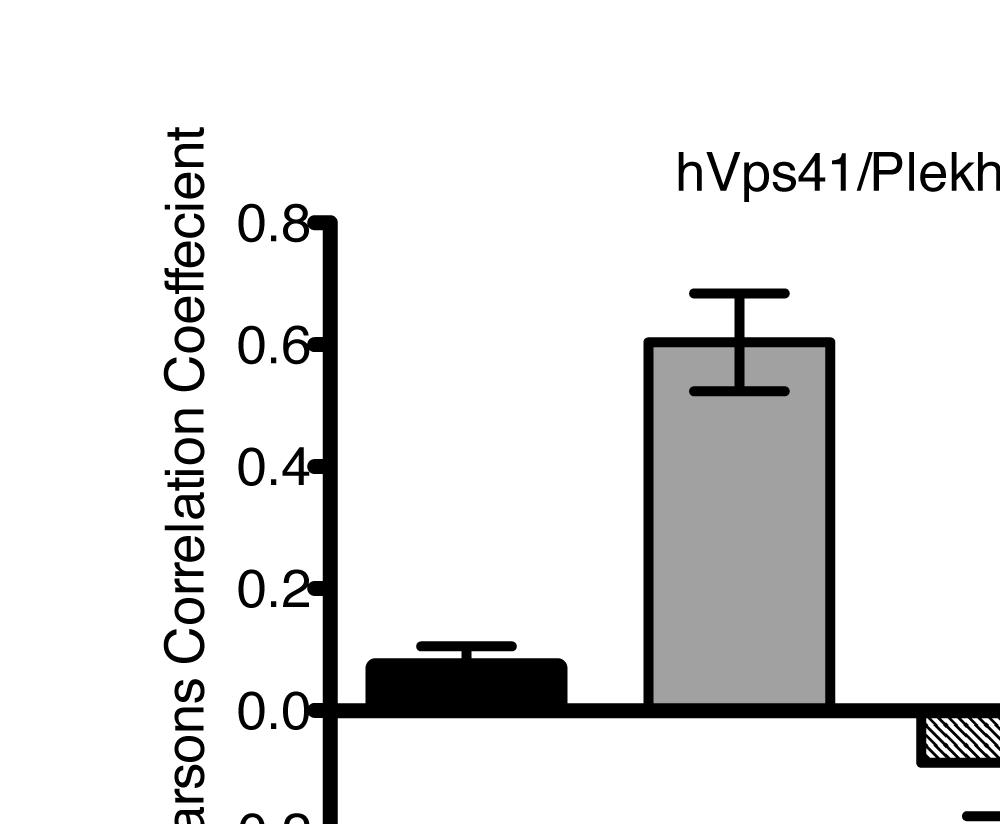
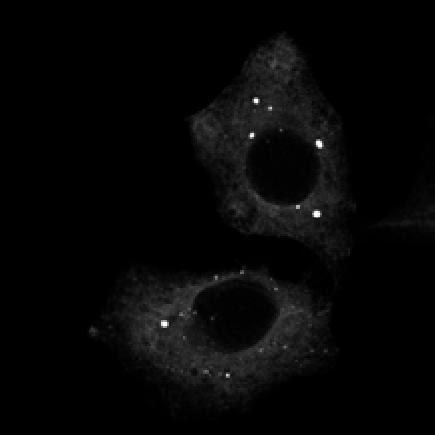
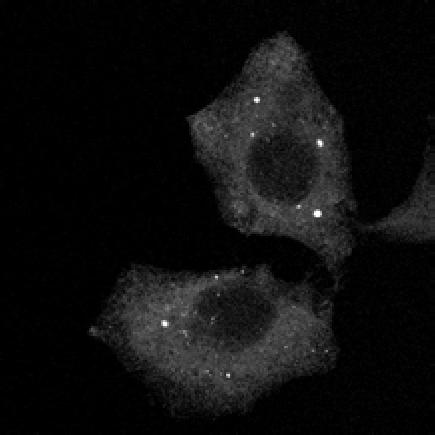
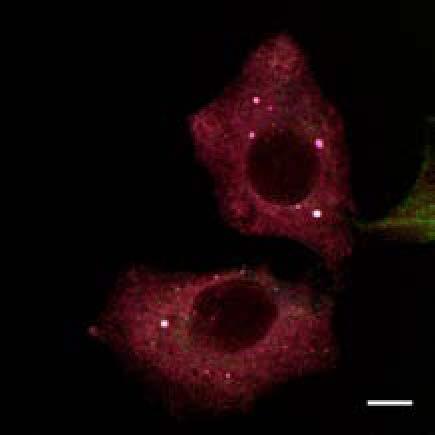
 U2OS cells stably expressing PLEKHM1-WT-GFP were fixed and stained for endogenous hvps41 and Rab7 proteins.")
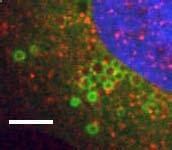
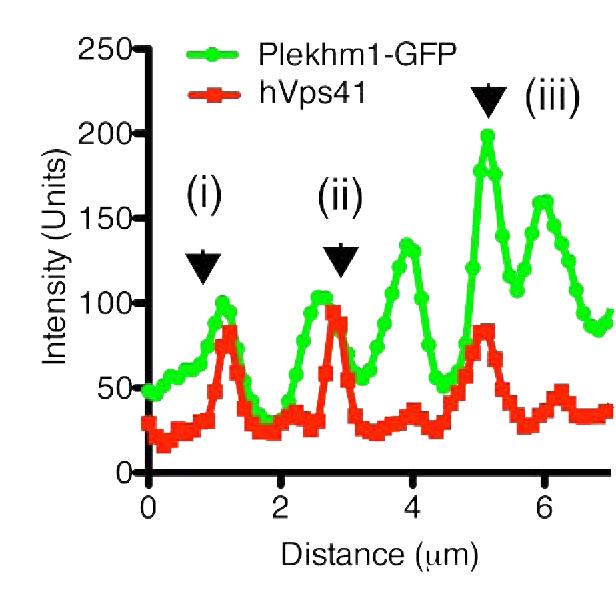
8 Figure 4. PLEKHM1 and HOPS Coordinate Vesicle Tethering and Fusion (A) U2OS cells stably expressing PLEKHM1-WT-GFP were fixed and stained for endogenous LAMP2 and Rab7 proteins. Arrow heads indicate regions of interest. Scale bar 10 mm. (B) U2OS cells stably expressing PLEKHM1-WT-GFP were fixed and stained for endogenous hvps41 and Rab7 proteins. Arrow heads indicate hvps41 puncta of interest. Scale bar, 10 mm. (C) Still image from time point 0 min of Movie S1 of PLEKHM1-GFP/mCherry-hVps41 in U2OS cells. Boxed region (i) indicates enhanced clusters of colocalized PLEKHM1 and hvps41; Box (Cii) represents the region imaged in Movie S2. Scale bar 10 mm. (D) Still frames of Movie S2 PLEKHM1-GFP/mCherry-hVps41 depicting PLEKHM1/hVps41-positive vesicles budding and fusing with a second vesicle. Similar behavior of vesicular structures was observed in n R 3 experiments. Scale bar, 1 mm. (E) Merged confocal micrographs of U20S cells stably expressing mcherry-hvps41 transfected either with GFP alone or PLEKHM1-GFP WT full-length (FL), GFP-PLEKHM1-RUN domain (aa 1 196), GFP-PLEKHM1-PH2 domain (aa ). Boxed regions indicate colocalization of proteins. Scale bars, 10 mm. (F) Localization of PLEKHM1-WT-GFP or PLEKHM1-DRUN-GFP stably expressed in U2OS cells. Scale bar, 10 mm. See also Figures S3. 46 Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.
 HeLa cells stably expressing shrna control oligo or oligo targeting PLEKHM1 were stimulated with 50 ng/ml EGF, and total cell lysates were")
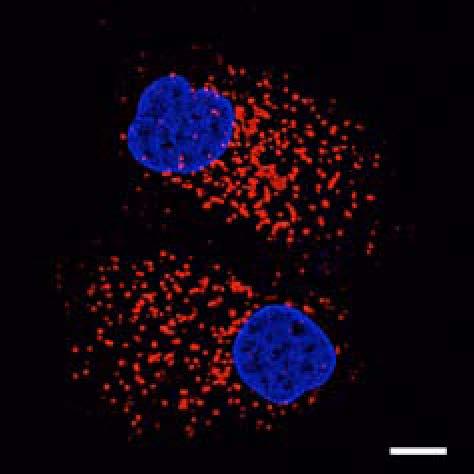
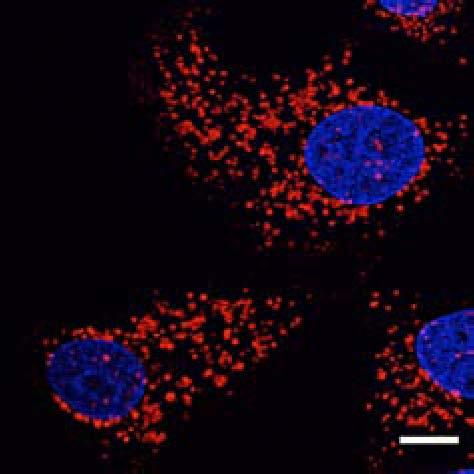
9 Figure 5. PLEKHM1 Depletion Inhibits EGFR Degradation (A) HeLa cells stably expressing shrna control oligo or oligo targeting PLEKHM1 were stimulated with 50 ng/ml EGF, and total cell lysates were immunoblotted for endogenous EGFR and PLEKHM1. Tubulin immunoblot represents loading control. Blots are representative of n = 3 separate experiments. (B) Hke2 cells were treated with sirna control oligo or oligo targeting PLEKHM1. Seventy-two hours post-sirna treatment, cells were stimulated with 50 ng/ml of EGF, and lysates were immunoblotted for endogenous EGFR and PLEKHM1. Blots are representative of n = 3 separate experiments. (C) HeLa cells stably expressing shrna control oligo or two different oligos targeting PLEKHM1 (shplek#1 and shplek#2) were stimulated with Rh-EGF (50 ng/ ml; white), fixed, and imaged by confocal microscope. Scale bars 10 mm unless otherwise stated. (legend continued on next page) Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 47
10 PLEKHM1 Is Required for Selective Autophagy Removal of Protein Aggregates Since PLEKHM1 regulated terminal autolysosome formation, we reasoned that its absence would also impair the degradation of protein aggregates. In a model antigen presentation system, autophagy inhibition causes the aggregation and accumulation of misfolded protein (GFP-SL8) in aggresome-like induced structures (ALIS) (Szeto et al., 2006). Aggregated GFP-SL8 is then rerouted to the proteasome, causing an influx of antigenic SL8 peptides through MHC class-i antigen presentation system that can be quantified by a specific antibody, 25D1 (Figure 7A) (Wenger et al., 2012). Depletion of PLEKHM1 resulted in the accumulation of GFP-SL8 in p62/ub-positive structures (Figures S6A and S6B) and a 35% increase in processed SL8 antigen presentation (Figure 7B, white bars). This was significantly enhanced upon the inhibition of autophagy by 3-MA (Figure 7B, gray bars) but was blocked by proteasomal inhibition (Figure 7B, black bars). This effect was comparable to silencing of ATG5 and NBR1 (Figure 7B) (Wenger et al., 2012). Puromycin-induced protein aggregates and p62/sqstm1- positive ALIS are known to be selectively recognized by ubiquitin binding autophagy receptors and subsequently removed via autophagy. Thus, we assessed the dynamics of puromycin-induced ubiquitin (Ub) and p62/sqstm1-positive ALIS clearance upon PLEKHM1 depletion. Seventy two hours posttransfection, control or PLEKHM1-deficient HeLa cells were left untreated in nutrient-rich media (no treatment; NT), treated with 5 mg/ml puromycin for 2 hr (Puro) or 2 hr Puro treated, washed, and allowed to recover for 3 or 6 hr in nutrient-rich media (Figure 7C). PLEKHM1 depletion caused the appearance of small Ub-positive (Ub+) puncta in untreated cells that were not present in control cells (Figure 7D; NT). Moreover, while control cells efficiently cleared puromycin-induced p62+/ub+ ALIS aggregates, recovery of PLEKHM1-deficient cells was not apparent even after 6 hr (Figures 7D and 7E), indicating that PLEKHM1 functions also during selective, ALISinduced autophagy. Finally, we assessed the functional role of the LIR in ALIS clearance. HeLa cells were treated as above (Figure 7A) with the addition that PLEKHM1-depleted HeLa cells were reconstituted with sirna-resistant PLEKHM1-WT (sir- WT) or sirna-resistant LIR mutant (sir-mutlir) 24 hr after sirna transfection and left for a further 48 hr. Cells were treated with puromycin for 2 hr or 2 hr puromycin and 6 hr recovery, and the resultant Ub + aggregate size was measured. Puromycininduced ALIS showed no difference in size in HeLa control or siplekhm1 cells (Figure 7F). After 6 hr recovery, the average aggregate size in control cells was approximately half the size of those still present in PLEKHM1-depleted cells (0.2mm 2 ± compared to 0.46 mm 2 ± 0.042; Figure 7F). PLEKHM1- WT was able to recapitulate the clearance of the aggregates but PLEKHM1-mutLIR was not ( ± compared to ± 0.013; Figures 7F and 7G). Thus, a lack of PLEKHM1 induces the formation of misfolded protein aggregates and significantly delays their removal by selective autophagy in a LIR-dependent manner. Taken together, these data demonstrate that PLEKHM1 is a lysosomal autophagy adaptor protein that links the vesicle fusion machinery to both selective and nonselective autophagy. DISCUSSION In this report, we characterized PLEKHM1, a Rab7 interacting protein that also binds components of the HOPS complex and the ubiquitin-like modifiers LC3/GABARAP. PLEKHM1 specifically functions at the late endosomes/lysosomes, being directly involved in the endocytic fusion events mediated by tethering complex HOPS. The defects that occur upon depletion of PLEKHM1 reflect an impairment not only of the endocytic pathway but also of autophagic-mediated degradation processes. The later phenomenon could be mediated by the ability of PLEKHM1 to bind to LC3 and promote fusion of LC3-positive autophagosomes with lysosomes. We show that PLEKHM1 sits at a point of convergence of both endocytic and autophagic pathways. Interestingly, the effects of PLEKHM1 loss have some resemblance to the effects of ESCRT component depletion, where enlarged amphisomes (enlarged vesicles formed by late endosome and autophagosome fusion) are formed with the accumulation of LC3, p62, and ALFY proteins occurring (Filimonenko et al., 2007). Further studies of the impact of PLEKHM1 loss on the endocytic pathway will help us fully understand the interplay between these convergent pathways. The small GTPase Rab7 is critical for the maturation of endosomes and delivery of the activated growth factor receptors to the lysosome. Consequently, Rab7 loss or mutation inhibits the final stages of EGFR degradation (Ceresa and Bahr, 2006; Vanlandingham and Ceresa, 2009). Additionally, autophagy is known to have a shared mode of final cargo degradation as endocytosis, where a close relationship of both processes is reflected in some of the key regulatory proteins. The common terminal end-point of autophagosome fusion with the lysosome is also dependent on Rab7 (Gutierrez et al., 2004; Jäger et al., 2004). As is the case for all GTPases within the cellular endomembrane system, Rab7 functions in its GTP-loaded form via specific recruitment of different effector proteins and fusion factors (Stenmark, 2009). The effectors that influence endocytic degradation appear not to be identical in the case of autophagy, and vice versa. For example, RILP and Rubicon were shown to have an opposing effect on EGFR degradation, as their depletion causes either accelerated (Rubicon; Sun et al., 2010; Tabata et al., 2010) or inhibited (RILP; Progida et al., 2007) degradation. However, RILP has not been reported to function in autophagy, while Rubicon (D and E) Number and size of Rh-EGF-positive puncta from experiment described in (C) was quantified in ImageJ software. T = 120 min. Number of Rh-EGFlabeled cells: n = 376 (shcntrl), n = 425 (shplek#1), and n = 283 (shplek#2) Rh-EGF analyzed in n = 2 experiments. Statistical significance was calculated in GraphPad Prism, using unpaired t test, and asterisk number assigned as follows: ***p < and **p < (F and G) Confocal micrographs of Rh-EGF colocalization with EEA1 and LAMP1 in HeLa cells treated with sicntrl (F) or (G) siplekhm1 after 120 min of stimulation. Colocalization of Rh-EGF with EEA1 in images shown was calculated by Volocity software (Perkin-Elmer) software and expressed as Pearson s correlation coefficient (R). See also Figures S4. 48 Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.
, Torin1 (250 nm, 2 hr), or Chloroquine (CQ) (2 hr. 20 mm). Lysates were immunoblotted for LC3B, p62, and Plekhm1.")
11 Figure 6. Genetic Loss of PLEKHM1 Inhibits Autolysosome Maturation (A) Plekhm1 +/+ (WT; +/+) and Plekhm1 / (KO; / ) MEFs were isolated at day E13.5, immortalized, and were treated as following: DMSO, Ku (Ku; 2 hr 10 mm), Torin1 (250 nm, 2 hr), or Chloroquine (CQ) (2 hr. 20 mm). Lysates were immunoblotted for LC3B, p62, and Plekhm1. Vinculin was used as a loading control, and anti-p-p70s6k (T389) was used as a marker for mtor activity. Densitometry analysis was used to calculate flux shown in (B). LC3B-II and p62 were normalized against Vinculin levels. mean ± SD n = 3 experiments. (C) Plekhm1 WT and KO MEFs were starved for 2 hr (EBSS), fixed, and immunolabeled with endogenous LAMP1 and LC3B antibodies. Boxed regions are enlarged in panels. Scale bar 20 mm. Images are representative of n = 3 independent experiments. (D) Pearson s correlation coefficient quantification of LC3B/LAMP1 colocalization in FM or EBSS-induced starvation conditions. Error bars represent mean ± SEM of >200 cells analyzed per each condition; n.s., not significant; **p = , unpaired t test. (legend continued on next page) Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 49
12 specifically associates with the autophagic PI3 kinase complex that contains Beclin1 and suppresses autophagosome maturation (Sun et al., 2010; Matsunaga et al., 2009; Zhong et al., 2009). Previous work has discounted a role of PLEKHM1 in the nonselective autophagy pathway on account of a minor effect on autophagy flux (LC3B lipidation) upon depletion (Tabata et al., 2010). In addition, the same report also shows that upon PLEKHM1 depletion, EGFR degradation is accelerated, as opposed to our study, where we show that EGFR is stabilized upon PLEKHM1 loss. The observed discrepancy could be attributed to the cell-type-specific effects, as Tabata et al. (2010) have employed HER-2 overexpressing A549 lung adenocarcinoma cells for their analysis. Interestingly, one of the key prognostic markers for both breast and lung cancer is the amplification of HER-2 on chromosome 17. Indeed, A549 cells contain four copies of chromosome 17 (Diaz et al., 2010), compared to HeLa, which only have three copies (Landry et al., 2013) and, due to its location on chromosome 17 (17q21) the PLEKHM1 gene, along with HER2 (17q12), would also be amplified in A549 cells compared to HeLa and MEFs (as used in this study). Conversely, the Chr17q21.31 region that contains the PLEKHM1 gene has recently been identified as a susceptibility locus for epithelial ovarian cancer due to an inversion polymorphism that results in significantly decreased PLEKHM1 expression (Permuth-Wey et al., 2013). Thus, the role of adaptor proteins, such as PLEKHM1, may be influenced by cell type, tissue specificity, copy number, and resident driver mutations within the various cancer cell lines commonly used in research where feedback loops of endocytic signaling networks may affect activation state of these adaptors and directly influence their role within the network. Therefore, further studies are required to assess how altered expression of PLEKHM1 in human and mice influences development of different tumor types. Protein aggregate accumulation is implicated in the pathogenesis of multiple diseases, such as Parkinson s, Alzheimer s, Huntington s, and amyotrophic lateral sclerosis, where mutant protein or misfolded proteins accumulate as cytotoxic aggregates. Autophagy has a critical role in the removal of these aggregates, as a compromised autophagy pathway leads to increased accumulation of aggregates, particularly in the neuronal cell types (Hara et al., 2006; Komatsu et al., 2006). Indeed, most of the proteins that accumulate are themselves autophagy substrates (Ravikumar et al., 2002; Webb et al., 2003) with the formation and removal of aggregates requiring autophagy receptors such as p62 and Alfy (Clausen et al., 2010; Filimonenko et al., 2010). Interestingly, two genome-wide association studies of Sporadic-Parkinson s disease (PD) and Familial-PD revealed that SNPs within the region of chromosome 17 that contains the PLEKHM1 gene were significantly linked to Familial-PD (Hill-Burns et al., 2014) and were a common risk factor for PD (Edwards et al., 2010). In another study, PLEKHM1 mrna was shown to be decreased in the cerebellum of multiple SNPs associated with Parkinson s disease (Nalls et al., 2014). We have shown that depletion of PLEKHM1 causes the formation of defective ribosomal products and is important for the removal of puromycin-induced ALIS in a LIR-dependent manner. Taken together, our data shed light on a potential mechanism that links PLEKHM1 to an increased risk of Parkinson s disease and may be an important regulator of protein aggregate removal that could have therapeutic and prognostic value for other proteinopathies. The fusion of the late endocytic/lysosomal compartments is driven by HOPS recruitment, tethering, and SNARE-mediated fusion (Balderhaar and Ungermann, 2013). The fusion of autophagosomes with lysosomes has recently been shown to be dependent upon the SNARE protein syntaxin 17 (STX17) (Itakura et al., 2012). This mechanism of fusion between autophagosome and lysosomes is conserved in Drosophila melanogaster, where both STX17 and SNAP29 are required for fusion (Takáts et al., 2013). SNAP29 and STX17 were both able to coprecipitate PLEKHM1; however, upon autophagy induction, PLEKHM1 coprecipitation with STX17 was enhanced along with endogenous hvps41. UVRAG, which was previously described to interact with the HOPS complex, does not coprecipitate STX17 (Jiang et al., 2014), and deletion of UVRAG/ Vps38 in Drosophila melanogaster does not impair autophagosome-lysosome fusion (Takáts et al., 2014). Thus, PLEKHM1 is potentially an upstream adaptor protein that interacts with Rab7, HOPS-SNARE complexes, and LC3/GABARAP proteins to facilitate autophagosome-lysosome fusion. Interestingly, a specialized process that requires extensive lysosome fusion is the formation of the ruffled membrane border of bone-resorbing osteoclasts. This process is essential for the polarized secretion of acid and proteolytic enzymes, thereby resulting in degradation of the bone matrix. A PLEKHM1 gene mutation in humans has been identified as a cause of osteopetrosis, a disease of high bone mass due to dysfunctional osteoclasts, where PLEKHM1 mutant osteoclasts fail to form the ruffled border and degrade bone (Van Wesenbeeck et al., 2007). Intriguingly, the PLEKHM1 mutation and Rab7 depletion both cause ruffled border loss and decreased bone resorption (Van Wesenbeeck et al., 2007; Zhao et al., 2001), and ruffled border formation is known to depend on LC3B and Atg5, essential autophagy proteins (DeSelm et al., 2011). Our data suggest a possible mechanistic explanation for the fusion defect observed in PLEKHM1 mutant osteoclasts that may have broader implications in the pathogenesis of human diseases such as osteopetrosis. Taken together, these data demonstrate that PLEKHM1 is a Rab7 effector protein that directly bridges autophagic and lysosomal membranes via binding to LC3 and Rab7/HOPS, facilitating fusion of these vesicles. PLEKHM1 appears to function as a platform that can link Rab7/HOPS to distinct cellular pathways, including endocytosis and autophagy, and has broad implications for the pathogenesis of cancer, proteopathies, and osteopetrosis. (E) Representative confocal micrographs of Plekhm1 WT (+/+) or KO ( / ) MEFs transfected with mcherry-gfp-tandem fluorescence-tagged LC3B after treatment for 2 hr with Ku (F) The percentages of colocalization where the number of red only puncta (autolysosomes) was quantified against Red + and Green + (autophagosomes; Red + / Red + +Green + ). Shown is mean ± SEM. n = 50 cells. p = , Student s t test. See also Figure S5 50 Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.
 Schematic representation of experimental setup for ihela model antigen presentation system (Wenger et")
13 Figure 7. PLEKHM1 Is Required for the Efficient Clearance of Protein Aggregates by Selective Autophagy (A) Schematic representation of experimental setup for ihela model antigen presentation system (Wenger et al., 2012). (B) ihela cells were transfected with sirna against PLEKHM1 (oligo #1 or #2), p62, NBR1, Atg5, or sirna control (sicntrl). GFP-Sl8 was induced, and cells were treated with 5 mm 3-MA (gray column) or 2 mm of MG132 (black column) or left untreated (white column) for the last 4 hr. SL-8 peptide presentation on H-2k b and (legend continued on next page) Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 51
14 EXPERIMENTAL PROCEDURES Immunoprecipitation For endogenous protein immunoprecipitation, supernatants (10 min, 13,000 3 g) from cells lysed in GTPase lysis buffer (50 mm Tris.HCl [ph 7.4], 150 mm NaCl, 5 mm MgCl 2, and 0.5% Triton X-100) were supplemented with Complete protease inhibitor cocktail (Roche), incubated with primary antibody overnight at 4 C rotation and protein A or G Sepharose (60min), centrifuged, and washed minimum of three times with lysis buffer. Precipitated proteins were detected by immunoblot analysis using chemiluminescence-based detection. For coprecipitation of epitope-tagged proteins, (3 ml anti-gfp beads; Chromotek or 5 ml anti-flagm2 beads, or 15 ml anti-ha beads) cells were incubated with lysates for a total of 60 min at 4 C rotation and pellets were washed three times with lysis buffer and four times with wash buffer (50 mm TRI.HCl [ph 7.4], 150 mm NaCl, 2 mm MgCl 2, and 0.1% Triton X-100) prior to loading onto SDS-PAGE gels for analysis. For immunoprecipitation of GFP-tagged SNARE proteins, cells were lysed in CHAPS lysis buffer (50 mm Tris [ph7.5], 20 mm CHAPS, 115 mm NaCl, and 5 mm MgCl 2 ) supplemented with EDTA-free protease inhibitor cocktail (Roche) and phosphatase inhibitor cocktail (Sigma). Cell suspensions were passed through a 27G needle twenty times, centrifuged at 16,000 3 g and the supernatant incubated with GFP-Trap (Chromotek) beads for 1 hr at 4C rotation. Samples were then washed five times in lysis buffer and boiled in presence of Laemlli buffer prior to SDS-PAGE analysis. Protein Aggregate Clearance Assay HeLa cells were transfected with either nontargeting sirna or two different sirnas against PLEKHM1 (siplek#1 and siplek#2). Puromycin treatment was carried out 72 hr posttransfection. In the case of reconstitution of sirna-depleted cells with either sirna#1-resistant PLEKHM1-WT or mutalir (sir-wt, sir-mutlir), sirna-resistant constructs were transfected 24 hr after sirna transfectetion and left for a further 48 hr prior to puromycin treatment. After treatment with 5 mg/ml puromycin for 2 hr, cells were either fixed in 4% PFA for 10 min or washed three times in FM without puromycin and incubated for a further 3 hr or 6 hr prior to fixation and immunofluorescence analysis. ImageJ Analyze particles function was used to analyze the size of protein aggregates of thresholded images. Immunogold Staining HeLa cells were transfected with PLEKHM1-GFP, GFP-Rab7, or GFP-LC3 and then 24 hr later were treated with 100 nm bafilomycin A1 for 5 hr. Cell pellets were prepared and fixed in 4% paraformaldehyde or 2.5% paraformaldehyde/0.2% glutaraldehyde in 0.1 M phosphate buffer (ph 7). Pellets were embedded in HM20, 70 nm sections cut, and transfected proteins detected by immunostaining with 0.5 mg/ml GFP antibody (Abcam; ab6556) and 1:200 Protein A-gold 10 nm (Utrecht Medical Centre, Netherlands). Sections were counterstained with uranyl acetate and lead citrate using a Leica EM AC20 autostainer and examined using a Philips CM10 transmission electron microscope equipped with a Gatan Bioscan Camera. Mouse Strain Maintenance and MEF Generation Plekhm1 gene trap mice (B6;129S5-Plekhm1 Gt(OST201689)Lex /Orl; LEXKO- 0354), were acquired from Lexicon Pharmaceuticals through the Wellcome Trust Access to Mutant Mouse Resources (UK). These mice had been generated by retrovirally transducing 129SV embryonic stem cells with a neomycin cassette targeted between exons 1 and 2 of Plekhm1 gene. The mice were subsequently backcrossed ten generations into C57BL/6. Mice were maintained at the Goethe University Frankfurt Medical School animal facility, as approved by the responsible Veterinary Officer of the City of Frankfurt. Animal welfare was approved by the Institutional Animal Welfare Officer (Tierschutzbeauftragter) in accordance with the Animal protection act (Tierschutzgesetz, TierSchG BGBI. IS. 1206, 1313 x7). Heterozygous mice (Plekhm1 +/ ) were mated and pregnant dams were euthanized humanely (isoflurane sedation followed by cervical dislocation). Embryos were isolated at E13.5 and sacrificed by decapitation and trypsinized. Isolated MEFs were then transfected with SV40 Tag and passaged until they were immortalized. Statistical Analysis Results were plotted and analyzed using unpaired t test using Graphpad (Prism) software. Detailed experimental procedures can be found in Supplemental Information. ACCESSION NUMBERS The atomic coordinates and structure factors have been deposited in the Protein Data Bank with accession code 3X0W. SUPPLEMENTAL INFORMATION Supplemental Information includes six figures, two tables, two movies, and Supplemental Experimental Procedures and can be found with this article online at ACKNOWLEDGMENTS We would like to acknowledge B. Richter, P. Grumati, A. Bremm, M. Frame, and K. Rajalingam for critical reading of the manuscript and valuable insights. We would like to thank W. Van Hul and B. Perdu for providing critical reagents and discussions and S. Wahl and J. Madlung for sample preparation and MS measurement. F.P.C. and M.H.H. would like to acknowledge TEM support from J. Greenhorn, the microscopy and histology core facility at the UoA, and research support from Arthritis Research UK (grant number 19379). D.M.S. was supported by a studentship (RHE/00092/S ) from the Nuffield Foundation s Oliver Bird Rheumatism Programme (UK). The anti-lamp2 antibody developed by J.T. August was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the expression level of model antigen were determined by FACS analysis (median of fluorescence intensity ± SD). One representative result out of three independent experiments is shown. The statistical analysis between siplekhm1 and sirna control were given by unpaired t test (*p < 0.05; **p < 0.01). (C) Schematic representation of experimental setup for clearance of puromycin (Puro)-induced protein aggregates in HeLa cells. (D) Merged representative confocal micrographs of sicntrl or siplekhm1 (siplek#1) HeLa cells treated as outlined in (A) fixed and immunolabeled using p62/ SQSTM1 and Ubiquitin antibodies. DAPI staining represents DNA (nucleus). White puncta pointed by arrowheads indicate colocalization between p62 and Ub. Scale bar, 10 mm. No treatment (NT), 2 hr puromycin (Puro); 2 hr puromycin followed by 3 hr recovery (3 hr off). (E) Quantification of confocal images of sicntrl, siplek#1, and siplek#2 after puromycin treatment. Zero hours represents HeLa cells treated with puromycin for 2 hr without recovery. Cells were scored as containing three or more p62/ub-positive aggregates or not and expressed as a percentage of total cells counted. Mean ± SEM. n = 3 independent experiments. ***p < unpaired t test. (F) HeLa cells were treated as in (A) with the exception of transfecting either sirna-resistant PLEKHM1-WT (sir-wt) or sirna-resistant PLEKHM1 mutant LIR (sir-mutlir; aa EDEWVNV/AAAAVNG) 48 hr prior to treatment with puromycin. Quantification of ubiquitin-positive (Ub+) aggregate size after 2 hr puromycin (2 hr puro) or 2 hr puro and 6 hr recovery (6 hr off) in sicntrl, siplek, siplek+sir-wt, and siplek+sir-mutlir. Values are mean ± SEM of aggregates from n > 25 cells per condition. ns = not significant; ***p < unpaired t test. (G) Representative confocal micrographs of reconstituted siplek cells with WT or mutlir. sir-plekhm1 (magenta) expressing cells 6 hr recovery after puromycin treatment. PLEKHM1-positive cells (dashed lines) and sirna-depleted cells (solid lines) are shown. Arrows indicate Ub (green) aggregates of interest. Scale bars, 20 mm. See also Figure S6. 52 Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.
15 the NICHD and maintained by The University of Iowa, Department of Biology. The GFP-SNARE constructs and Atg5 KO MEFs were a kind gift from N. Mizushima, and the constructs were acquired through Adgene. We would like to thank the beamline scientists and staff at BL-5A, Photon Factory, Tsukuba, Japan for helpful support during data collection. This work was supported by grants from Deutsche Forschungsgemeinschaft (DI 931/3-1; BE 4685/1-1), the Cluster of Excellence Macromolecular Complexes of the Goethe University Frankfurt (EXC115), LOEWE grant Ub-Net and LOEWE Centrum for Gene and Cell therapy Frankfurt, and the European Research Council/ERC grant agreement n LineUb to I.D. and XABA to C.B. Received: May 6, 2014 Revised: October 6, 2014 Accepted: November 5, 2014 Published: December 11, 2014 REFERENCES Balderhaar, H.J., and Ungermann, C. (2013). CORVET and HOPS tethering complexes - coordinators of endosome and lysosome fusion. J. Cell Sci. 126, Cantalupo, G., Alifano, P., Roberti, V., Bruni, C.B., and Bucci, C. (2001). Rabinteracting lysosomal protein (RILP): the Rab7 effector required for transport to lysosomes. EMBO J. 20, Ceresa, B.P., and Bahr, S.J. (2006). rab7 activity affects epidermal growth factor:epidermal growth factor receptor degradation by regulating endocytic trafficking from the late endosome. J. Biol. Chem. 281, Clausen, T.H., Lamark, T., Isakson, P., Finley, K., Larsen, K.B., Brech, A., Øvervatn, A., Stenmark, H., Bjørkøy, G., Simonsen, A., and Johansen, T. (2010). p62/sqstm1 and ALFY interact to facilitate the formation of p62 bodies/alis and their degradation by autophagy. Autophagy 6, DeSelm, C.J., Miller, B.C., Zou, W., Beatty, W.L., van Meel, E., Takahata, Y., Klumperman, J., Tooze, S.A., Teitelbaum, S.L., and Virgin, H.W. (2011). Autophagy proteins regulate the secretory component of osteoclastic bone resorption. Dev. Cell 21, Diaz, R., Nguewa, P.A., Parrondo, R., Perez-Stable, C., Manrique, I., Redrado, M., Catena, R., Collantes, M., Peñuelas, I., Díaz-González, J.A., and Calvo, A. (2010). Antitumor and antiangiogenic effect of the dual EGFR and HER-2 tyrosine kinase inhibitor lapatinib in a lung cancer model. BMC Cancer 10, 188. Edwards, T.L., Scott, W.K., Almonte, C., Burt, A., Powell, E.H., Beecham, G.W., Wang, L., Züchner, S., Konidari, I., Wang, G., et al. (2010). Genomewide association study confirms SNPs in SNCA and the MAPT region as common risk factors for Parkinson disease. Ann. Hum. Genet. 74, Filimonenko, M., Stuffers, S., Raiborg, C., Yamamoto, A., Malerød, L., Fisher, E.M., Isaacs, A., Brech, A., Stenmark, H., and Simonsen, A. (2007). Functional multivesicular bodies are required for autophagic clearance of protein aggregates associated with neurodegenerative disease. J. Cell Biol. 179, Filimonenko, M., Isakson, P., Finley, K.D., Anderson, M., Jeong, H., Melia, T.J., Bartlett, B.J., Myers, K.M., Birkeland, H.C., Lamark, T., et al. (2010). The selective macroautophagic degradation of aggregated proteins requires the PI3Pbinding protein Alfy. Mol. Cell 38, Gutierrez, M.G., Munafó, D.B., Berón, W., and Colombo, M.I. (2004). Rab7 is required for the normal progression of the autophagic pathway in mammalian cells. J. Cell Sci. 117, Hara, T., Nakamura, K., Matsui, M., Yamamoto, A., Nakahara, Y., Suzuki- Migishima, R., Yokoyama, M., Mishima, K., Saito, I., Okano, H., and Mizushima, N. (2006). Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441, Hill-Burns, E.M., Wissemann, W.T., Hamza, T.H., Factor, S.A., Zabetian, C.P., and Payami, H. (2014). Identification of a novel Parkinson s disease locus via stratified genome-wide association study. BMC Genomics 15, 118. Itakura, E., Kishi-Itakura, C., and Mizushima, N. (2012). The hairpin-type tailanchored SNARE syntaxin 17 targets to autophagosomes for fusion with endosomes/lysosomes. Cell 151, Jäger, S., Bucci, C., Tanida, I., Ueno, T., Kominami, E., Saftig, P., and Eskelinen, E.L. (2004). Role for Rab7 in maturation of late autophagic vacuoles. J. Cell Sci. 117, Jiang, P., Nishimura, T., Sakamaki, Y., Itakura, E., Hatta, T., Natsume, T., and Mizushima, N. (2014). The HOPS complex mediates autophagosome-lysosome fusion through interaction with syntaxin 17. Mol. Biol. Cell 25, Kirkin, V., Lamark, T., Sou, Y.S., Bjørkøy, G., Nunn, J.L., Bruun, J.A., Shvets, E., McEwan, D.G., Clausen, T.H., Wild, P., et al. (2009a). A role for NBR1 in autophagosomal degradation of ubiquitinated substrates. Mol. Cell 33, Komatsu, M., Waguri, S., Chiba, T., Murata, S., Iwata, J., Tanida, I., Ueno, T., Koike, M., Uchiyama, Y., Kominami, E., and Tanaka, K. (2006). Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 441, Korolchuk, V.I., Saiki, S., Lichtenberg, M., Siddiqi, F.H., Roberts, E.A., Imarisio, S., Jahreiss, L., Sarkar, S., Futter, M., Menzies, F.M., et al. (2011). Lysosomal positioning coordinates cellular nutrient responses. Nat. Cell Biol. 13, Kuma, A., Hatano, M., Matsui, M., Yamamoto, A., Nakaya, H., Yoshimori, T., Ohsumi, Y., Tokuhisa, T., and Mizushima, N. (2004). The role of autophagy during the early neonatal starvation period. Nature 432, Landry, J.J., Pyl, P.T., Rausch, T., Zichner, T., Tekkedil, M.M., Stutz, A.M., Jauch, A., Aiyar, R.S., Pau, G., Delhomme, N., et al. (2013). The genomic and transcriptomic landscape of a HeLa cell line. G3 (Bethesda) 3, Liang, C., Lee, J.S., Inn, K.S., Gack, M.U., Li, Q., Roberts, E.A., Vergne, I., Deretic, V., Feng, P., Akazawa, C., et al. (2008). Beclin1-binding UVRAG targets the class C Vps complex to coordinate autophagosome maturation and endocytic trafficking. Nat. Cell Biol. 10, Matsunaga, K., Saitoh, T., Tabata, K., Omori, H., Satoh, T., Kurotori, N., Maejima, I., Shirahama-Noda, K., Ichimura, T., Isobe, T., et al. (2009). Two Beclin 1-binding proteins, Atg14L and Rubicon, reciprocally regulate autophagy at different stages. Nat. Cell Biol. 11, Nalls, M.A., Saad, M., Noyce, A.J., Keller, M.F., Schrag, A., Bestwick, J.P., Traynor, B.J., Gibbs, J.R., Hernandez, D.G., Cookson, M.R., et al.; International Parkinson s Disease Genomics Consortium (IPDGC); Wellcome Trust Case Control Consortium 2 (WTCCC2); North American Brain Expression Consortium (NABEC); United Kingdom Brain Expression Consortium (UKBEC) (2014). Genetic comorbidities in Parkinson s disease. Hum. Mol. Genet. 23, Nixon, R.A. (2013). The role of autophagy in neurodegenerative disease. Nat. Med. 19, Novak, I., Kirkin, V., McEwan, D.G., Zhang, J., Wild, P., Rozenknop, A., Rogov, V., Löhr, F., Popovic, D., Occhipinti, A., et al. (2010). Nix is a selective autophagy receptor for mitochondrial clearance. EMBO Rep. 11, Pankiv, S., Alemu, E.A., Brech, A., Bruun, J.A., Lamark, T., Overvatn, A., Bjørkøy, G., and Johansen, T. (2010). FYCO1 is a Rab7 effector that binds to LC3 and PI3P to mediate microtubule plus end-directed vesicle transport. J. Cell Biol. 188, Permuth-Wey, J., Lawrenson, K., Shen, H.C., Velkova, A., Tyrer, J.P., Chen, Z., Lin, H.Y., Chen, Y.A., Tsai, Y.Y., Qu, X., et al. (2013). Identification and molecular characterization of a new ovarian cancer susceptibility locus at 17q Nat. Commun. 4, Popovic, D., and Dikic, I. (2014). TBC1D5 and the AP2 complex regulate ATG9 trafficking and initiation of autophagy. EMBO Rep. 15, Popovic, D., Akutsu, M., Novak, I., Harper, J.W., Behrends, C., and Dikic, I. (2012). Rab GTPase-activating proteins in autophagy: regulation of endocytic and autophagy pathways by direct binding to human ATG8 modifiers. Mol. Cell. Biol. 32, Press, B., Feng, Y., Hoflack, B., and Wandinger-Ness, A. (1998). Mutant Rab7 causes the accumulation of cathepsin D and cation-independent mannose Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc. 53
16 6-phosphate receptor in an early endocytic compartment. J. Cell Biol. 140, Progida, C., Malerød, L., Stuffers, S., Brech, A., Bucci, C., and Stenmark, H. (2007). RILP is required for the proper morphology and function of late endosomes. J. Cell Sci. 120, Ravikumar, B., Duden, R., and Rubinsztein, D.C. (2002). Aggregate-prone proteins with polyglutamine and polyalanine expansions are degraded by autophagy. Hum. Mol. Genet. 11, Rogov, V., Dötsch, V., Johansen, T., and Kirkin, V. (2014). Interactions between autophagy receptors and ubiquitin-like proteins form the molecular basis for selective autophagy. Mol. Cell 53, Sigismund, S., Confalonieri, S., Ciliberto, A., Polo, S., Scita, G., and Di Fiore, P.P. (2012). Endocytosis and signaling: cell logistics shape the eukaryotic cell plan. Physiol. Rev. 92, Stenmark, H. (2009). Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 10, Stolz, A., Ernst, A., and Dikic, I. (2014). Cargo recognition and trafficking in selective autophagy. Nat. Cell Biol. 16, Sun, Q., Westphal, W., Wong, K.N., Tan, I., and Zhong, Q. (2010). Rubicon controls endosome maturation as a Rab7 effector. Proc. Natl. Acad. Sci. USA 107, Szeto, J., Kaniuk, N.A., Canadien, V., Nisman, R., Mizushima, N., Yoshimori, T., Bazett-Jones, D.P., and Brumell, J.H. (2006). ALIS are stress-induced protein storage compartments for substrates of the proteasome and autophagy. Autophagy 2, Tabata, K., Matsunaga, K., Sakane, A., Sasaki, T., Noda, T., and Yoshimori, T. (2010). Rubicon and PLEKHM1 negatively regulate the endocytic/autophagic pathway via a novel Rab7-binding domain. Mol. Biol. Cell 21, Takáts, S., Nagy, P., Varga, A., Pircs, K., Kárpáti, M., Varga, K., Kovács, A.L., Heged}us, K., and Juhász, G. (2013). Autophagosomal Syntaxin17-dependent lysosomal degradation maintains neuronal function in Drosophila. J. Cell Biol. 201, Takáts, S., Pircs, K., Nagy, P., Varga, A., Kárpáti, M., Heged}us, K., Kramer, H., Kovács, A.L., Sass, M., and Juhász, G. (2014). Interaction of the HOPS complex with Syntaxin 17 mediates autophagosome clearance in Drosophila. Mol. Biol. Cell 25, Van Wesenbeeck, L., Odgren, P.R., Coxon, F.P., Frattini, A., Moens, P., Perdu, B., MacKay, C.A., Van Hul, E., Timmermans, J.P., Vanhoenacker, F., et al. (2007). Involvement of PLEKHM1 in osteoclastic vesicular transport and osteopetrosis in incisors absent rats and humans. J. Clin. Invest. 117, Vanlandingham, P.A., and Ceresa, B.P. (2009). Rab7 regulates late endocytic trafficking downstream of multivesicular body biogenesis and cargo sequestration. J. Biol. Chem. 284, Waterhouse, A.M., Procter, J.B., Martin, D.M., Clamp, M., and Barton, G.J. (2009). Jalview Version 2 a multiple sequence alignment editor and analysis workbench. Bioinformatics 25, Webb, J.L., Ravikumar, B., Atkins, J., Skepper, J.N., and Rubinsztein, D.C. (2003). Alpha-Synuclein is degraded by both autophagy and the proteasome. J. Biol. Chem. 278, Wenger, T., Terawaki, S., Camosseto, V., Abdelrassoul, R., Mies, A., Catalan, N., Claudio, N., Clavarino, G., de Gassart, A., Rigotti Fde, A., et al. (2012). Autophagy inhibition promotes defective neosynthesized proteins storage in ALIS, and induces redirection toward proteasome processing and MHCIrestricted presentation. Autophagy 8, Wild, P., Farhan, H., McEwan, D.G., Wagner, S., Rogov, V.V., Brady, N.R., Richter, B., Korac, J., Waidmann, O., Choudhary, C., et al. (2011). Phosphorylation of the autophagy receptor optineurin restricts Salmonella growth. Science 333, Wurmser, A.E., Sato, T.K., and Emr, S.D. (2000). New component of the vacuolar class C-Vps complex couples nucleotide exchange on the Ypt7 GTPase to SNARE-dependent docking and fusion. J. Cell Biol. 151, Zerial, M., and McBride, H. (2001). Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2, Zhao, H., Laitala-Leinonen, T., Parikka, V., and Väänänen, H.K. (2001). Downregulation of small GTPase Rab7 impairs osteoclast polarization and bone resorption. J. Biol. Chem. 276, Zhong, Y., Wang, Q.J., Li, X., Yan, Y., Backer, J.M., Chait, B.T., Heintz, N., and Yue, Z. (2009). Distinct regulation of autophagic activity by Atg14L and Rubicon associated with Beclin 1-phosphatidylinositol-3-kinase complex. Nat. Cell Biol. 11, Molecular Cell 57, 39 54, January 8, 2015 ª2015 Elsevier Inc.
17 Molecular Cell, Volume 57 Supplemental Information PLEKHM1 Regulates Autophagosome-Lysosome Fusion through HOPS Complex and LC3/GABARAP Proteins David G. McEwan, Doris Popovic, Andrea Gubas, Seigo Terawaki, Hironori Suzuki, Daniela Stadel, Fraser P. Coxon, Diana Miranda de Stegmann, Sagar Bhogaraju, Karthik Maddi, Anja Kirchof, Evelina Gatti, Miep H. Helfrich, Soichi Wakatsuki, Christian Behrends, Philippe Pierre, and Ivan Dikic







18 Figure S1 A 20kDa 20kDa 10% Input GST-Alone GST-PH1-LIR GST-PH2 GST-PH1-PH2 Flag-LC3A Flag-GABARAP B 15kDa 10% load GST GST-PH1 GST-PH1-LIR GST-PH2 GST-LIR GST-LIR Mut Flag- LC3B 20kDa Flag-GABARAP-L1 Ponceau S 75kDa 48kDa 37kDa Ponceau S 25kDa C D F E G



 B")


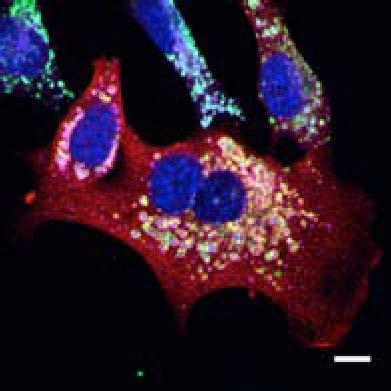
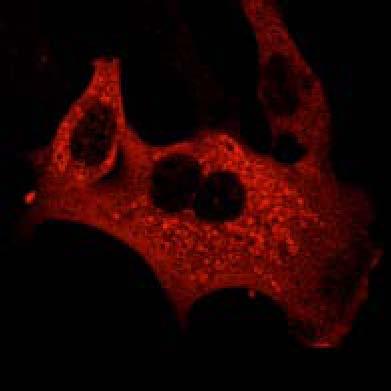
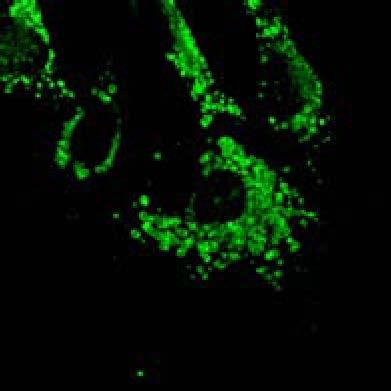


19 Plekhm1 +/+ MEFs Figure S2 A HeLa DMSO MG132 BafA1 EBSS Ku 130kDa PLEK/Vinculin PLEKHM1 (Low exposure) B HeLa-T-Rex-PLEKHM1-WT-2xFlag 130kDa DMSO MG132 BafA1 EBSS Ku PLEKHM1 (Low exposure) 130kDa 63kDa p62/vinculin 15kDa 10kDa 100kDa PLEKHM1 (high exposure) p62 I LC3B II Vinculin 130kDa 63kDa 15kDa 10kDa 100kDa PLEKHM1 (high exposure) p62 I II LC3B Vinculin C Myc-PLEKHM1 LAMP1 LC3B Merge Plekhm1 LAMP1 LC3B 100 FM Intensity (Units) D KU BafA1 Value (Units) Distance µm E MEFs MEFs Hke3 HCT116 Atg5+/+ Atg5 / Plekhm1+/+ Plekhm1 / sicntrl siplek shcntrl shplek#1 shplek#4 shplek#5 F Plekhm1/DAPI Plekhm1/DAPI/LAMP1 130kDa 100kDa Plekhm1 75kDa ROI 25kDa Vinculin Plekhm1 / MEFs ROI
20 Figure S3 A PLEKHM1-GFP/hVps41/DAPI B (i) (ii) (iii) C D 1% Serum Max Projection shcntrl shplek#1 shplek#2 E %Cells with clustered lysosomes shcntrl shplek#1 shplek#2 LAMP2/DAPI




21 Figure S4 A HeLa B HKE3 C 180kDa HKE3 sicntrl sivps min EGF EGFR D 180kDa HKE3 sicntrl sivps min EGF EGFR 100kDa Vps41 100kDa Vps11 100kDa E HKE3 sicntrl sirab Vinculin min EGF 100kDa F Vinculin 180kDa EGFR 100kDa Vinculin G sicntrl siplek min Rh-EGF 180kDa EGFR EGFR/Vinculin I 10% Serum 1% Serum 130kDa PLEKHM1 100kDa Vinculin sicntrl 10% serum 1% serum H 100kDa 75kDa 130kDa 100kDa sicntrl siplek sicntrl siplek TfnR PLEKHM1 Vinculin siplekhm1 Tfn Receptor/EEA1/DAPI
22 Figure S5 A Atg5 +/+ Atg5 / DMSO EBSS BafA1 +/+ / +/+ / +/+ / Plekhm1 Plekhm1 p62 (Low exposure) p62 (high exposure) I LC3B II I GABARAP II Vinculin



23 Figure S6 A p62 Ub GFP Merge sicntrl siplek#1 B p62 SL8 GFP Merge sicntrl siplek#1
GFP CCD2 GFP IP:GFP
 D1 D2 1 75 95 148 178 492 GFP CCD1 CCD2 CCD2 GFP D1 D2 GFP D1 D2 Beclin 1 IB:GFP IP:GFP Supplementary Figure 1: Mapping domains required for binding to HEK293T cells are transfected with EGFP-tagged mutant
D1 D2 1 75 95 148 178 492 GFP CCD1 CCD2 CCD2 GFP D1 D2 GFP D1 D2 Beclin 1 IB:GFP IP:GFP Supplementary Figure 1: Mapping domains required for binding to HEK293T cells are transfected with EGFP-tagged mutant
Coleman et al., Supplementary Figure 1
 Coleman et al., Supplementary Figure 1 BrdU Merge G1 Early S Mid S Supplementary Figure 1. Sequential destruction of CRL4 Cdt2 targets during the G1/S transition. HCT116 cells were synchronized by sequential
Coleman et al., Supplementary Figure 1 BrdU Merge G1 Early S Mid S Supplementary Figure 1. Sequential destruction of CRL4 Cdt2 targets during the G1/S transition. HCT116 cells were synchronized by sequential
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Legends for Supplementary Tables. Supplementary Table 1. An excel file containing primary screen data. Worksheet 1, Normalized quantification data from a duplicated screen: valid
SUPPLEMENTARY INFORMATION Legends for Supplementary Tables. Supplementary Table 1. An excel file containing primary screen data. Worksheet 1, Normalized quantification data from a duplicated screen: valid
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION Dynamic Phosphorylation of HP1 Regulates Mitotic Progression in Human Cells Supplementary Figures Supplementary Figure 1. NDR1 interacts with HP1. (a) Immunoprecipitation using
SUPPLEMENTARY INFORMATION Dynamic Phosphorylation of HP1 Regulates Mitotic Progression in Human Cells Supplementary Figures Supplementary Figure 1. NDR1 interacts with HP1. (a) Immunoprecipitation using
The Human Protein PRR14 Tethers Heterochromatin to the Nuclear Lamina During Interphase and Mitotic Exit
 Cell Reports, Volume 5 Supplemental Information The Human Protein PRR14 Tethers Heterochromatin to the Nuclear Lamina During Interphase and Mitotic Exit Andrey Poleshko, Katelyn M. Mansfield, Caroline
Cell Reports, Volume 5 Supplemental Information The Human Protein PRR14 Tethers Heterochromatin to the Nuclear Lamina During Interphase and Mitotic Exit Andrey Poleshko, Katelyn M. Mansfield, Caroline
Supplementary Table 1. The Q-PCR primer sequence is summarized in the following table.
 Supplementary Table 1. The Q-PCR primer sequence is summarized in the following table. Name Sequence (5-3 ) Application Flag-u ggactacaaggacgacgatgac Shared upstream primer for all the amplifications of
Supplementary Table 1. The Q-PCR primer sequence is summarized in the following table. Name Sequence (5-3 ) Application Flag-u ggactacaaggacgacgatgac Shared upstream primer for all the amplifications of
Figure S1. Figure S2. Figure S3 HB Anti-FSP27 (COOH-terminal peptide) Ab. Anti-GST-FSP27(45-127) Ab.
 Ab. Anti-GST-FSP27(45-127) Ab.") / 36B4 mrna ratio Figure S1 * 2. 1.6 1.2.8 *.4 control TNFα BRL49653 Figure S2 Su bw AT p iw Anti- (COOH-terminal peptide) Ab Blot : Anti-GST-(45-127) Ab β-actin Figure S3 HB2 HW AT BA T Figure S4 A TAG
/ 36B4 mrna ratio Figure S1 * 2. 1.6 1.2.8 *.4 control TNFα BRL49653 Figure S2 Su bw AT p iw Anti- (COOH-terminal peptide) Ab Blot : Anti-GST-(45-127) Ab β-actin Figure S3 HB2 HW AT BA T Figure S4 A TAG
PHT1;2-CFP YFP-PHF + PHT1;2-CFP YFP-PHF
 YFP-PHF1 CFP-PHT1;2 PHT1;2-CFP YFP-PHF + PHT1;2-CFP YFP-PHF + CFP-PHT1;2 Negative control!-gfp Supplemental Figure 1: PHT1;2 accumulation is PHF1 dependent. Immunoblot analysis on total protein extract
YFP-PHF1 CFP-PHT1;2 PHT1;2-CFP YFP-PHF + PHT1;2-CFP YFP-PHF + CFP-PHT1;2 Negative control!-gfp Supplemental Figure 1: PHT1;2 accumulation is PHF1 dependent. Immunoblot analysis on total protein extract
ASPP1 Fw GGTTGGGAATCCACGTGTTG ASPP1 Rv GCCATATCTTGGAGCTCTGAGAG
 Supplemental Materials and Methods Plasmids: the following plasmids were used in the supplementary data: pwzl-myc- Lats2 (Aylon et al, 2006), pretrosuper-vector and pretrosuper-shp53 (generous gift of
Supplemental Materials and Methods Plasmids: the following plasmids were used in the supplementary data: pwzl-myc- Lats2 (Aylon et al, 2006), pretrosuper-vector and pretrosuper-shp53 (generous gift of
7.06 Cell Biology EXAM #2 March 20, 2003
 7.06 Cell Biology EXAM #2 March 20, 2003 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #2 March 20, 2003 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
supplementary information
 DOI: 1.138/ncb1839 a b Control 1 2 3 Control 1 2 3 Fbw7 Smad3 1 2 3 4 1 2 3 4 c d IGF-1 IGF-1Rβ IGF-1Rβ-P Control / 1 2 3 4 Real-time RT-PCR Relative quantity (IGF-1/ mrna) 2 1 IGF-1 1 2 3 4 Control /
DOI: 1.138/ncb1839 a b Control 1 2 3 Control 1 2 3 Fbw7 Smad3 1 2 3 4 1 2 3 4 c d IGF-1 IGF-1Rβ IGF-1Rβ-P Control / 1 2 3 4 Real-time RT-PCR Relative quantity (IGF-1/ mrna) 2 1 IGF-1 1 2 3 4 Control /
Supplemental Movie Legend.
 Supplemental Movie Legend. Transfected T cells were dropped onto SEE superantigen-pulsed Raji B cells (approximate location indicated by circle). Maximum-intensity projections from Z-stacks (17 slices,
Supplemental Movie Legend. Transfected T cells were dropped onto SEE superantigen-pulsed Raji B cells (approximate location indicated by circle). Maximum-intensity projections from Z-stacks (17 slices,
Supplementary information to accompany: A novel role for the DNA repair gene Rad51 in Netrin-1 signalling
 Supplementary information to accompany: A novel role for the DNA repair gene Rad51 in Netrin-1 signalling Glendining KA 1, Markie D 2, Gardner RJM 4, Franz EA 3, Robertson SP 4, Jasoni CL 1 Supplementary
Supplementary information to accompany: A novel role for the DNA repair gene Rad51 in Netrin-1 signalling Glendining KA 1, Markie D 2, Gardner RJM 4, Franz EA 3, Robertson SP 4, Jasoni CL 1 Supplementary
University of Groningen
 University of Groningen Full length RTN3 regulates turnover of tubular endoplasmic reticulum via selective autophagy Grumati, Paolo; Morozzi, Giulio; Hölper, Soraya; Mari, Muriel; Harwardt, Marie-Lena
University of Groningen Full length RTN3 regulates turnover of tubular endoplasmic reticulum via selective autophagy Grumati, Paolo; Morozzi, Giulio; Hölper, Soraya; Mari, Muriel; Harwardt, Marie-Lena
SANTA CRUZ BIOTECHNOLOGY, INC.
 TECHNICAL SERVICE GUIDE: Western Blotting 2. What size bands were expected and what size bands were detected? 3. Was the blot blank or was a dark background or non-specific bands seen? 4. Did this same
TECHNICAL SERVICE GUIDE: Western Blotting 2. What size bands were expected and what size bands were detected? 3. Was the blot blank or was a dark background or non-specific bands seen? 4. Did this same
One-step split GFP staining for sensitive protein detection and localization in mammalian cells
 Supplementary Materials For: One-step split GFP staining for sensitive protein detection and localization in mammalian cells Lara Kaddoum 1,3, Eddy Magdeleine 1,3, Geoffrey S. Waldo 4, Etienne Joly 1,3,
Supplementary Materials For: One-step split GFP staining for sensitive protein detection and localization in mammalian cells Lara Kaddoum 1,3, Eddy Magdeleine 1,3, Geoffrey S. Waldo 4, Etienne Joly 1,3,
Assays for studying mitochondrial health and function
 APPLICATION NOTE Fluorescence labeling and detection Assays for studying mitochondrial health and function Introduction Mitochondria play a critical role in maintaining normal cellular activities. Mitochondria
APPLICATION NOTE Fluorescence labeling and detection Assays for studying mitochondrial health and function Introduction Mitochondria play a critical role in maintaining normal cellular activities. Mitochondria
Supplementary Figure S1. Immunodetection of full-length XA21 and the XA21 C-terminal cleavage product.
 Supplementary Information Supplementary Figure S1. Immunodetection of full-length XA21 and the XA21 C-terminal cleavage product. Total protein extracted from Kitaake wild type and rice plants carrying
Supplementary Information Supplementary Figure S1. Immunodetection of full-length XA21 and the XA21 C-terminal cleavage product. Total protein extracted from Kitaake wild type and rice plants carrying
7.06 Problem Set #3, Spring 2005
 7.06 Problem Set #3, Spring 2005 1. The Drosophila compound eye is composed of about 800 units called ommatidia. Each ommatidium contains eight photoreceptor neurons (R1 through R8), which develop in a
7.06 Problem Set #3, Spring 2005 1. The Drosophila compound eye is composed of about 800 units called ommatidia. Each ommatidium contains eight photoreceptor neurons (R1 through R8), which develop in a
GM130 Is Required for Compartmental Organization of Dendritic Golgi Outposts
 Current Biology, Volume 24 Supplemental Information GM130 Is Required for Compartmental Organization of Dendritic Golgi Outposts Wei Zhou, Jin Chang, Xin Wang, Masha G. Savelieff, Yinyin Zhao, Shanshan
Current Biology, Volume 24 Supplemental Information GM130 Is Required for Compartmental Organization of Dendritic Golgi Outposts Wei Zhou, Jin Chang, Xin Wang, Masha G. Savelieff, Yinyin Zhao, Shanshan
This Document Contains:
 This Document Contains: 1. In-Cell Western Protocol II. Cell Seeding and Stimulation Supplemental Protocol III. Complete Assay Example: Detailing the Seeding, Stimulation and Detection of the A431 Cellular
This Document Contains: 1. In-Cell Western Protocol II. Cell Seeding and Stimulation Supplemental Protocol III. Complete Assay Example: Detailing the Seeding, Stimulation and Detection of the A431 Cellular
Tracking Cellular Protein Localization and Movement in Cells with a Flexible Fluorescent Labeling Technology. Chad Zimprich January 2015
 Tracking Cellular Protein Localization and Movement in Cells with a Flexible Fluorescent Labeling Technology Chad Zimprich January 2015 Presentation verview HaloTag Fusion Technology Design Functionality
Tracking Cellular Protein Localization and Movement in Cells with a Flexible Fluorescent Labeling Technology Chad Zimprich January 2015 Presentation verview HaloTag Fusion Technology Design Functionality
CytoPainter Golgi Staining Kit Green Fluorescence
 ab139483 CytoPainter Golgi Staining Kit Green Fluorescence Instructions for Use Designed for the detection of Golgi bodies by microscopy This product is for research use only and is not intended for diagnostic
ab139483 CytoPainter Golgi Staining Kit Green Fluorescence Instructions for Use Designed for the detection of Golgi bodies by microscopy This product is for research use only and is not intended for diagnostic
The Two-Hybrid System
 Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine The Two-Hybrid System Carolina Vollert & Peter Uetz Institut für Genetik Forschungszentrum Karlsruhe PO Box 3640 D-76021 Karlsruhe
Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine The Two-Hybrid System Carolina Vollert & Peter Uetz Institut für Genetik Forschungszentrum Karlsruhe PO Box 3640 D-76021 Karlsruhe
Non-Organic-Based Isolation of Mammalian microrna using Norgen s microrna Purification Kit
 Application Note 13 RNA Sample Preparation Non-Organic-Based Isolation of Mammalian microrna using Norgen s microrna Purification Kit B. Lam, PhD 1, P. Roberts, MSc 1 Y. Haj-Ahmad, M.Sc., Ph.D 1,2 1 Norgen
Application Note 13 RNA Sample Preparation Non-Organic-Based Isolation of Mammalian microrna using Norgen s microrna Purification Kit B. Lam, PhD 1, P. Roberts, MSc 1 Y. Haj-Ahmad, M.Sc., Ph.D 1,2 1 Norgen
Supplement Figure 1. Plin5 Plin2 Plin1. KDEL-DSRed. Plin-YFP. Merge
 Supplement Figure 1 Plin5 Plin2 Plin1 KDEL-DSRed Plin-YFP Merge Supplement Figure 2 A. Plin5-Ab MitoTracker Merge AML12 B. Plin5-YFP Cytochrome c-cfp merge Supplement Figure 3 Ad.GFP Ad.Plin5 Supplement
Supplement Figure 1 Plin5 Plin2 Plin1 KDEL-DSRed Plin-YFP Merge Supplement Figure 2 A. Plin5-Ab MitoTracker Merge AML12 B. Plin5-YFP Cytochrome c-cfp merge Supplement Figure 3 Ad.GFP Ad.Plin5 Supplement
Nature Methods: doi: /nmeth Supplementary Figure 1. Retention of RNA with LabelX.
 Supplementary Figure 1 Retention of RNA with LabelX. (a) Epi-fluorescence image of single molecule FISH (smfish) against GAPDH on HeLa cells expanded without LabelX treatment. (b) Epi-fluorescence image
Supplementary Figure 1 Retention of RNA with LabelX. (a) Epi-fluorescence image of single molecule FISH (smfish) against GAPDH on HeLa cells expanded without LabelX treatment. (b) Epi-fluorescence image
Supplemental Online Material. The mouse embryonic fibroblast cell line #10 derived from β-arrestin1 -/- -β-arrestin2 -/-
 #1074683s 1 Supplemental Online Material Materials and Methods Cell lines and tissue culture The mouse embryonic fibroblast cell line #10 derived from β-arrestin1 -/- -β-arrestin2 -/- knock-out animals
#1074683s 1 Supplemental Online Material Materials and Methods Cell lines and tissue culture The mouse embryonic fibroblast cell line #10 derived from β-arrestin1 -/- -β-arrestin2 -/- knock-out animals
Description: Nuclear morphology and dynamics in nontargeting sirna transfected cells. HeLa Kyoto
 Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Tables Title of file for HTML: Supplementary Movie 1 Description: Nuclear morphology and dynamics
Title of file for HTML: Supplementary Information Description: Supplementary Figures and Supplementary Tables Title of file for HTML: Supplementary Movie 1 Description: Nuclear morphology and dynamics
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/323/5910/124/dc1 Supporting Online Material for Regulation of Neuronal Survival Factor MEF2D by Chaperone-Mediated Autophagy Qian Yang, Hua She, Marla Gearing, Emanuela
www.sciencemag.org/cgi/content/full/323/5910/124/dc1 Supporting Online Material for Regulation of Neuronal Survival Factor MEF2D by Chaperone-Mediated Autophagy Qian Yang, Hua She, Marla Gearing, Emanuela
Protocol for induction of expression and cell lysate production
 Protocol for induction of expression and cell lysate production AV-04 Doxycyclin induction and cell lysate 1.0 Introduction / Description This method is intended for the treatment of the previously transfected
Protocol for induction of expression and cell lysate production AV-04 Doxycyclin induction and cell lysate 1.0 Introduction / Description This method is intended for the treatment of the previously transfected
Purification of DNA from living cells
 Purification of DNA from living cells Total cell DNA & Plasmid DNA Grow and harvest bacterial culture Prepare cell extract Purify DNA from a cell extract Concentrate DNA samples Measure DNA concentration
Purification of DNA from living cells Total cell DNA & Plasmid DNA Grow and harvest bacterial culture Prepare cell extract Purify DNA from a cell extract Concentrate DNA samples Measure DNA concentration
1. Cross-linking and cell harvesting
 ChIP is a powerful tool that allows the specific matching of proteins or histone modifications to regions of the genome. Chromatin is isolated and antibodies to the antigen of interest are used to determine
ChIP is a powerful tool that allows the specific matching of proteins or histone modifications to regions of the genome. Chromatin is isolated and antibodies to the antigen of interest are used to determine
Supplementary Figure 1. RAD51 and RAD51 paralogs are enriched spontaneously onto
 Supplementary Figure legends Supplementary Figure 1. and paralogs are enriched spontaneously onto the S-phase chromatin during DN replication. () Chromatin fractionation was carried out as described in
Supplementary Figure legends Supplementary Figure 1. and paralogs are enriched spontaneously onto the S-phase chromatin during DN replication. () Chromatin fractionation was carried out as described in
Wnt16 smact merge VK/AB
 A WT Wnt6 smact merge VK/A KO ctrl IgG WT KO Wnt6 smact DAPI SUPPLEMENTAL FIGURE I: Wnt6 expression in MGP-deficient aortae. Immunostaining for Wnt6 and smooth muscle actin (smact) in aortae from 7 day
A WT Wnt6 smact merge VK/A KO ctrl IgG WT KO Wnt6 smact DAPI SUPPLEMENTAL FIGURE I: Wnt6 expression in MGP-deficient aortae. Immunostaining for Wnt6 and smooth muscle actin (smact) in aortae from 7 day
Protein Purification Products. Complete Solutions for All of Your Protein Purification Applications
 Protein Purification Products Complete Solutions for All of Your Protein Purification Applications FLAG-Tagged Protein Products EXPRESS with the pcmv-dykddddk Vector Set Fuse your protein of interest to
Protein Purification Products Complete Solutions for All of Your Protein Purification Applications FLAG-Tagged Protein Products EXPRESS with the pcmv-dykddddk Vector Set Fuse your protein of interest to
Quantitative analysis of recombination in YFP and CFP gene of FRET biosensor induced by lentiviral or retroviral gene transfer.
 Supplementary Information: Quantitative analysis of recombination in and gene of FRET biosensor induced by lentiviral or retroviral gene transfer. Akira T. Komatsubara, Michiyuki Matsuda,, and Kazuhiro
Supplementary Information: Quantitative analysis of recombination in and gene of FRET biosensor induced by lentiviral or retroviral gene transfer. Akira T. Komatsubara, Michiyuki Matsuda,, and Kazuhiro
Discovery and Humanization of Novel High Affinity Neutralizing Monoclonal Antibodies to Human IL-17A
 Discovery and Humanization of Novel High Affinity Neutralizing Monoclonal Antibodies to Human IL-17A Contacts: Marty Simonetti martysimonetti@gmail.com Kirby Alton kirby.alton@abeomecorp.com Rick Shimkets
Discovery and Humanization of Novel High Affinity Neutralizing Monoclonal Antibodies to Human IL-17A Contacts: Marty Simonetti martysimonetti@gmail.com Kirby Alton kirby.alton@abeomecorp.com Rick Shimkets
ab Hypoxic Response Human Flow Cytometry Kit
 ab126585 Hypoxic Response Human Flow Cytometry Kit Instructions for Use For measuring protein levels by flow cytometry: hypoxia-inducible factor 1-alpha (HIF1A) and BCL2/adenovirus E1B 19 kda proteininteracting
ab126585 Hypoxic Response Human Flow Cytometry Kit Instructions for Use For measuring protein levels by flow cytometry: hypoxia-inducible factor 1-alpha (HIF1A) and BCL2/adenovirus E1B 19 kda proteininteracting
TrkB knockdown cell lines (i.e., BBM1-KD and 361-KD cells) were prepared by
 were prepared by") Supplemental Information Cell Culture TrkB knockdown cell lines (i.e., BBM1-KD and 361-KD cells) were prepared by transducing BBM1 or 361 cells with a lentivirus encoding shrna for TrkB. Transduction was
Supplemental Information Cell Culture TrkB knockdown cell lines (i.e., BBM1-KD and 361-KD cells) were prepared by transducing BBM1 or 361 cells with a lentivirus encoding shrna for TrkB. Transduction was
by Neurobasal medium (supplemented with B27, 0.5mM glutamine, and 100 U/mL
 Supplementary Materials and methods Neuronal cultures and transfection The hippocampus was dissected from E8 rat embryos, dissociated, and neurons plated onto glass coverslips coated with poly-ornithine
Supplementary Materials and methods Neuronal cultures and transfection The hippocampus was dissected from E8 rat embryos, dissociated, and neurons plated onto glass coverslips coated with poly-ornithine
Autophagy Assays (LC3B immunofluorescence, LC3B western blot, acridine orange assay) Xin Zhang and Qingsong Liu *
 Xin Zhang and Qingsong Liu *") Autophagy Assays (LC3B immunofluorescence, LC3B western blot, acridine orange assay) Xin Zhang and Qingsong Liu * High Magnetic Field Laboratory, Chinese Academy of Sciences, Hefei, Anhui, China *For correspondence:
Autophagy Assays (LC3B immunofluorescence, LC3B western blot, acridine orange assay) Xin Zhang and Qingsong Liu * High Magnetic Field Laboratory, Chinese Academy of Sciences, Hefei, Anhui, China *For correspondence:
Supplemental Data Supplementary Figure Legends and Scheme Figure S1.
 Supplemental Data Supplementary Figure Legends and Scheme Figure S1. UTK1 inhibits the second EGF-induced wave of lamellipodia formation in TT cells. A and B, EGF-induced lamellipodia formation in TT cells,
Supplemental Data Supplementary Figure Legends and Scheme Figure S1. UTK1 inhibits the second EGF-induced wave of lamellipodia formation in TT cells. A and B, EGF-induced lamellipodia formation in TT cells,
Comparison of Commercial Transfection Reagents: Cell line optimized transfection kits for in vitro cancer research.
 Comparison of Commercial Transfection Reagents: Cell line optimized transfection kits for in vitro cancer research. by Altogen Labs, 11200 Manchaca Road, Suite 203 Austin TX 78748 USA Tel. (512) 433-6177
Comparison of Commercial Transfection Reagents: Cell line optimized transfection kits for in vitro cancer research. by Altogen Labs, 11200 Manchaca Road, Suite 203 Austin TX 78748 USA Tel. (512) 433-6177
The MAP Kinase Family
 The MAP Kinase Family Extracellular stimuli Classical MAP kinases Atypical MAP kinases MAPKKK MLK1/2/3/7; LZK RAF-1/A/B TAK1; TPL2 c-mos MEKK1-4; DLK ASK1/2; MLTK TAO1/2 ASK1 TAK1 MEKK1-4 MEKK2/3 TPL2???
The MAP Kinase Family Extracellular stimuli Classical MAP kinases Atypical MAP kinases MAPKKK MLK1/2/3/7; LZK RAF-1/A/B TAK1; TPL2 c-mos MEKK1-4; DLK ASK1/2; MLTK TAO1/2 ASK1 TAK1 MEKK1-4 MEKK2/3 TPL2???
Nature Neuroscience: doi: /nn Supplementary Figure 1
 Supplementary Figure 1 PCR-genotyping of the three mouse models used in this study and controls for behavioral experiments after semi-chronic Pten inhibition. a-c. DNA from App/Psen1 (a), Pten tg (b) and
Supplementary Figure 1 PCR-genotyping of the three mouse models used in this study and controls for behavioral experiments after semi-chronic Pten inhibition. a-c. DNA from App/Psen1 (a), Pten tg (b) and
SensoLyte Anti-alpha-Synuclein Quantitative ELISA Kit (Human/Mouse/Rat) *Colorimetric*
 *Colorimetric*") Catalog # Kit Size SensoLyte Anti-alpha-Synuclein Quantitative ELISA Kit (Human/Mouse/Rat) *Colorimetric* AS-55550 One 96-well strip plate This kit is optimized to detect human/mouse/rat alpha-synuclein
Catalog # Kit Size SensoLyte Anti-alpha-Synuclein Quantitative ELISA Kit (Human/Mouse/Rat) *Colorimetric* AS-55550 One 96-well strip plate This kit is optimized to detect human/mouse/rat alpha-synuclein
Isolation, culture, and transfection of primary mammary epithelial organoids
 Supplementary Experimental Procedures Isolation, culture, and transfection of primary mammary epithelial organoids Primary mammary epithelial organoids were prepared from 8-week-old CD1 mice (Charles River)
Supplementary Experimental Procedures Isolation, culture, and transfection of primary mammary epithelial organoids Primary mammary epithelial organoids were prepared from 8-week-old CD1 mice (Charles River)
MICB688L/MOCB639 Advanced Cell Biology Exam II
 MICB688L/MOCB639 Advanced Cell Biology Exam II May 10, 2001 Name: 1. Briefly describe the four major classes of cell surface receptors and their modes of action (immediate downstream only) (8) 2. Please
MICB688L/MOCB639 Advanced Cell Biology Exam II May 10, 2001 Name: 1. Briefly describe the four major classes of cell surface receptors and their modes of action (immediate downstream only) (8) 2. Please
sirna Transfection Into Primary Neurons Using Fuse-It-siRNA
 sirna Transfection Into Primary Neurons Using Fuse-It-siRNA This Application Note describes a protocol for sirna transfection into sensitive, primary cortical neurons using Fuse-It-siRNA. This innovative
sirna Transfection Into Primary Neurons Using Fuse-It-siRNA This Application Note describes a protocol for sirna transfection into sensitive, primary cortical neurons using Fuse-It-siRNA. This innovative
SUMOstar Gene Fusion Technology
 Gene Fusion Technology NEW METHODS FOR ENHANCING FUNCTIONAL PROTEIN EXPRESSION AND PURIFICATION IN INSECT CELLS White Paper June 2007 LifeSensors Inc. 271 Great Valley Parkway Malvern, PA 19355 www.lifesensors.com
Gene Fusion Technology NEW METHODS FOR ENHANCING FUNCTIONAL PROTEIN EXPRESSION AND PURIFICATION IN INSECT CELLS White Paper June 2007 LifeSensors Inc. 271 Great Valley Parkway Malvern, PA 19355 www.lifesensors.com
Toll Receptor-Mediated Hippo Signaling Controls Innate Immunity in Drosophila
 Cell Supplemental Information Toll Receptor-Mediated Hippo Signaling Controls Innate Immunity in Drosophila Bo Liu, Yonggang Zheng, Feng Yin, Jianzhong Yu, Neal Silverman, and Duojia Pan Supplemental Experimental
Cell Supplemental Information Toll Receptor-Mediated Hippo Signaling Controls Innate Immunity in Drosophila Bo Liu, Yonggang Zheng, Feng Yin, Jianzhong Yu, Neal Silverman, and Duojia Pan Supplemental Experimental
To isolate single GNS 144 cell clones, cells were plated at a density of 1cell/well
 Supplemental Information: Supplemental Methods: Cell culture To isolate single GNS 144 cell clones, cells were plated at a density of 1cell/well in 96 well Primaria plates in GNS media and incubated at
Supplemental Information: Supplemental Methods: Cell culture To isolate single GNS 144 cell clones, cells were plated at a density of 1cell/well in 96 well Primaria plates in GNS media and incubated at
mcherry Monoclonal Antibody (16D7) Catalog Number M11217 Product data sheet
 Catalog Number M11217 Product data sheet") Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 mcherry Monoclonal Antibody (16D7) Catalog Number M11217 Product data sheet Details
Website: thermofisher.com Customer Service (US): 1 800 955 6288 ext. 1 Technical Support (US): 1 800 955 6288 ext. 441 mcherry Monoclonal Antibody (16D7) Catalog Number M11217 Product data sheet Details
Biochimie II. Assistants Ben Brankatschk Eleonora Torti
 Biochimie II Purification de protéines exprimées dans des cellules humaines en culture Daniel Abegg Christophe Berthier Pauline Bonvin abegg6@etu.unige.ch berthie4@etu.unige.ch bonvinp0@etu.unige.ch Assistants
Biochimie II Purification de protéines exprimées dans des cellules humaines en culture Daniel Abegg Christophe Berthier Pauline Bonvin abegg6@etu.unige.ch berthie4@etu.unige.ch bonvinp0@etu.unige.ch Assistants
Short hairpin RNA (shrna) against MMP14. Lentiviral plasmids containing shrna
 against MMP14. Lentiviral plasmids containing shrna") Supplemental Materials and Methods Short hairpin RNA (shrna) against MMP14. Lentiviral plasmids containing shrna (Mission shrna, Sigma) against mouse MMP14 were transfected into HEK293 cells using FuGene6
Supplemental Materials and Methods Short hairpin RNA (shrna) against MMP14. Lentiviral plasmids containing shrna (Mission shrna, Sigma) against mouse MMP14 were transfected into HEK293 cells using FuGene6
BA, BSc, and MSc Degree Examinations
 Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Bacterial pathogenesis Time Allowed: 2 hours Marking Scheme: Total marks available
Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Bacterial pathogenesis Time Allowed: 2 hours Marking Scheme: Total marks available
PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%)
") 1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%) DESCRIPTION Nickel NTA Magnetic Agarose Beads are products that allow rapid and easy small-scale purification of
1 AFFINITY HIS-TAG PURIFICATION PROCEDURE FOR USE NICKEL NTA Magnetic Agarose Beads (5%) DESCRIPTION Nickel NTA Magnetic Agarose Beads are products that allow rapid and easy small-scale purification of
A subclass of HSP70s regulate development and abiotic stress responses in Arabidopsis thaliana
 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Journal of Plant Research A subclass of HSP70s regulate development and abiotic stress responses in Arabidopsis thaliana Linna Leng 1 Qianqian Liang
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Journal of Plant Research A subclass of HSP70s regulate development and abiotic stress responses in Arabidopsis thaliana Linna Leng 1 Qianqian Liang
Supplementary Figure Legend
 Supplementary Figure Legend Supplementary Figure S1. Effects of MMP-1 silencing on HEp3-hi/diss cell proliferation in 2D and 3D culture conditions. (A) Downregulation of MMP-1 expression in HEp3-hi/diss
Supplementary Figure Legend Supplementary Figure S1. Effects of MMP-1 silencing on HEp3-hi/diss cell proliferation in 2D and 3D culture conditions. (A) Downregulation of MMP-1 expression in HEp3-hi/diss
OPPF-UK Standard Protocols: Mammalian Expression
 OPPF-UK Standard Protocols: Mammalian Expression Joanne Nettleship joanne@strubi.ox.ac.uk Table of Contents 1. Materials... 3 2. Cell Maintenance... 4 3. 24-Well Transient Expression Screen... 5 4. DNA
OPPF-UK Standard Protocols: Mammalian Expression Joanne Nettleship joanne@strubi.ox.ac.uk Table of Contents 1. Materials... 3 2. Cell Maintenance... 4 3. 24-Well Transient Expression Screen... 5 4. DNA
Supplemental Information. The TRAIL-Induced Cancer Secretome. Promotes a Tumor-Supportive Immune. Microenvironment via CCR2
 Molecular Cell, Volume 65 Supplemental Information The TRAIL-Induced Cancer Secretome Promotes a Tumor-Supportive Immune Microenvironment via CCR2 Torsten Hartwig, Antonella Montinaro, Silvia von Karstedt,
Molecular Cell, Volume 65 Supplemental Information The TRAIL-Induced Cancer Secretome Promotes a Tumor-Supportive Immune Microenvironment via CCR2 Torsten Hartwig, Antonella Montinaro, Silvia von Karstedt,
Xeno-Free Systems for hesc & hipsc. Facilitating the shift from Stem Cell Research to Clinical Applications
 Xeno-Free Systems for hesc & hipsc Facilitating the shift from Stem Cell Research to Clinical Applications NutriStem Defined, xeno-free (XF), serum-free media (SFM) specially formulated for growth and
Xeno-Free Systems for hesc & hipsc Facilitating the shift from Stem Cell Research to Clinical Applications NutriStem Defined, xeno-free (XF), serum-free media (SFM) specially formulated for growth and
Alternative Cleavage and Polyadenylation of RNA
 Developmental Cell 18 Supplemental Information The Spen Family Protein FPA Controls Alternative Cleavage and Polyadenylation of RNA Csaba Hornyik, Lionel C. Terzi, and Gordon G. Simpson Figure S1, related
Developmental Cell 18 Supplemental Information The Spen Family Protein FPA Controls Alternative Cleavage and Polyadenylation of RNA Csaba Hornyik, Lionel C. Terzi, and Gordon G. Simpson Figure S1, related
Lecture 8: Affinity Chromatography-III
 Lecture 8: Affinity Chromatography-III Key words: Chromatography; Affinity chromatography; Protein Purification During this lecture, we shall be studying few more examples of affinity chromatography. The
Lecture 8: Affinity Chromatography-III Key words: Chromatography; Affinity chromatography; Protein Purification During this lecture, we shall be studying few more examples of affinity chromatography. The
Kinetics Review. Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets)
") Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
Quiz 1 Kinetics Review Tonight at 7 PM Phys 204 We will do two problems on the board (additional ones than in the problem sets) I will post the problems with solutions on Toolkit for those that can t make
TECHNICAL BULLETIN. MEK Activity Assay Kit. Product Code CS0490 Storage Temperature 20 C
 MEK Activity Assay Kit Product Code CS0490 Storage Temperature 20 C TECHNICAL BULLETIN Product Description The MAP kinase kinases (MAPKK, mitogen-activated protein kinase kinase, also termed MEK) are a
MEK Activity Assay Kit Product Code CS0490 Storage Temperature 20 C TECHNICAL BULLETIN Product Description The MAP kinase kinases (MAPKK, mitogen-activated protein kinase kinase, also termed MEK) are a
Xfect Protein Transfection Reagent
 Xfect Protein Transfection Reagent Mammalian Expression Systems Rapid, high-efficiency, low-toxicity protein transfection Transfect a large amount of active protein Virtually no cytotoxicity, unlike lipofection
Xfect Protein Transfection Reagent Mammalian Expression Systems Rapid, high-efficiency, low-toxicity protein transfection Transfect a large amount of active protein Virtually no cytotoxicity, unlike lipofection
3. Results. 3.1 Generation of HEK293 cell clones stably expressing ETA and ETB receptors
 3. Results 3.1 Generation of HEK293 cell clones stably expressing ETA and ETB receptors To investigate the dimerisation of the endothelin receptor subtypes HEK293 cells were stably transfected with plasmids
3. Results 3.1 Generation of HEK293 cell clones stably expressing ETA and ETB receptors To investigate the dimerisation of the endothelin receptor subtypes HEK293 cells were stably transfected with plasmids
Article. The Folliculin Tumor Suppressor Is a GAP. for the RagC/D GTPases That Signal. Amino Acid Levels to mtorc1
 Molecular Cell, Volume 52 Supplemental Information Article The Folliculin Tumor Suppressor Is a GAP for the RagC/D GTPases That Signal Amino Acid Levels to mtorc1 Zhi-Yang Tsun, Liron Bar-Peled, Lynne
Molecular Cell, Volume 52 Supplemental Information Article The Folliculin Tumor Suppressor Is a GAP for the RagC/D GTPases That Signal Amino Acid Levels to mtorc1 Zhi-Yang Tsun, Liron Bar-Peled, Lynne
ENCODE RBP Antibody Characterization Guidelines
 ENCODE RBP Antibody Characterization Guidelines Approved on November 18, 2016 Background An integral part of the ENCODE Project is to characterize the antibodies used in the experiments. This document
ENCODE RBP Antibody Characterization Guidelines Approved on November 18, 2016 Background An integral part of the ENCODE Project is to characterize the antibodies used in the experiments. This document
Supplemental Data. LMO4 Controls the Balance between Excitatory. and Inhibitory Spinal V2 Interneurons
 Neuron, Volume 61 Supplemental Data LMO4 Controls the Balance between Excitatory and Inhibitory Spinal V2 Interneurons Kaumudi Joshi, Seunghee Lee, Bora Lee, Jae W. Lee, and Soo-Kyung Lee Supplemental
Neuron, Volume 61 Supplemental Data LMO4 Controls the Balance between Excitatory and Inhibitory Spinal V2 Interneurons Kaumudi Joshi, Seunghee Lee, Bora Lee, Jae W. Lee, and Soo-Kyung Lee Supplemental
Supplemental Material for Xue et al. List. Supplemental Figure legends. Figure S1. Related to Figure 1. Figure S2. Related to Figure 3
 Supplemental Material for Xue et al List Supplemental Figure legends Figure S1. Related to Figure 1 Figure S. Related to Figure 3 Figure S3. Related to Figure 4 Figure S4. Related to Figure 4 Figure S5.
Supplemental Material for Xue et al List Supplemental Figure legends Figure S1. Related to Figure 1 Figure S. Related to Figure 3 Figure S3. Related to Figure 4 Figure S4. Related to Figure 4 Figure S5.
SUPPLEMENTAL MATERIALS
 SUPPLEMENL MERILS Eh-seq: RISPR epitope tagging hip-seq of DN-binding proteins Daniel Savic, E. hristopher Partridge, Kimberly M. Newberry, Sophia. Smith, Sarah K. Meadows, rian S. Roberts, Mark Mackiewicz,
SUPPLEMENL MERILS Eh-seq: RISPR epitope tagging hip-seq of DN-binding proteins Daniel Savic, E. hristopher Partridge, Kimberly M. Newberry, Sophia. Smith, Sarah K. Meadows, rian S. Roberts, Mark Mackiewicz,
QImaging Camera Application Notes Multicolor Immunofluorescence Imaging
 QImaging Camera Application Notes Multicolor Immunofluorescence Imaging In order to image localization of intracellular proteins with high specificity, it is frequently necessary to multiplex antibody
QImaging Camera Application Notes Multicolor Immunofluorescence Imaging In order to image localization of intracellular proteins with high specificity, it is frequently necessary to multiplex antibody
(A) Schematic illustration of sciatic nerve ligation. P, proximal; D, distal to the ligation site.
 Schematic illustration of sciatic nerve ligation. P, proximal; D, distal to the ligation site.") SUPPLEMENTRY INFORMTION SUPPLEMENTL FIGURES Figure S1. () Schematic illustration of sciatic nerve ligation. P, proximal; D, distal to the ligation site. () Western blot of ligated and unligated sciatic
SUPPLEMENTRY INFORMTION SUPPLEMENTL FIGURES Figure S1. () Schematic illustration of sciatic nerve ligation. P, proximal; D, distal to the ligation site. () Western blot of ligated and unligated sciatic
IMMUNOPRECIPITATION (IP)
") 1 IMMUNOPRECIPITATION (IP) Overview and Technical Tips 2 CONTENTS 3 7 8 9 12 13 17 18 19 20 Introduction Factors Influencing IP General Protocol Modifications Of IP Protocols Troubleshooting Contact Us
1 IMMUNOPRECIPITATION (IP) Overview and Technical Tips 2 CONTENTS 3 7 8 9 12 13 17 18 19 20 Introduction Factors Influencing IP General Protocol Modifications Of IP Protocols Troubleshooting Contact Us
Figure S1. Immunoblotting showing proper expression of tagged R and AVR proteins in N. benthamiana.
 SUPPLEMENTARY FIGURES Figure S1. Immunoblotting showing proper expression of tagged R and AVR proteins in N. benthamiana. YFP:, :CFP, HA: and (A) and HA, CFP and YFPtagged and AVR1-CO39 (B) were expressed
SUPPLEMENTARY FIGURES Figure S1. Immunoblotting showing proper expression of tagged R and AVR proteins in N. benthamiana. YFP:, :CFP, HA: and (A) and HA, CFP and YFPtagged and AVR1-CO39 (B) were expressed
SureSilencing sirna Array Technology Overview
 SureSilencing sirna Array Technology Overview Pathway-Focused sirna-based RNA Interference Topics to be Covered Who is SuperArray? Brief Introduction to RNA Interference Challenges Facing RNA Interference
SureSilencing sirna Array Technology Overview Pathway-Focused sirna-based RNA Interference Topics to be Covered Who is SuperArray? Brief Introduction to RNA Interference Challenges Facing RNA Interference
Antibody Structure. Antibodies
 Antibodies Secreted by B lymphocytes Great diversity and specificity: >10 9 different antibodies; can distinguish between very similar molecules Tag particles for clearance/destruction Protect against
Antibodies Secreted by B lymphocytes Great diversity and specificity: >10 9 different antibodies; can distinguish between very similar molecules Tag particles for clearance/destruction Protect against
Antibody Structure supports Function
 Antibodies Secreted by B lymphocytes Great diversity and specificity: >10 9 different antibodies; can distinguish between very similar molecules Tag particles for clearance/destruction Protect against
Antibodies Secreted by B lymphocytes Great diversity and specificity: >10 9 different antibodies; can distinguish between very similar molecules Tag particles for clearance/destruction Protect against
Supplemental Information. PARP1 Represses PAP and Inhibits Polyadenylation during Heat Shock
 Molecular Cell, Volume 49 Supplemental Information PARP1 Represses PAP and Inhibits Polyadenylation during Heat Shock Dafne Campigli Di Giammartino, Yongsheng Shi, and James L. Manley Supplemental Information
Molecular Cell, Volume 49 Supplemental Information PARP1 Represses PAP and Inhibits Polyadenylation during Heat Shock Dafne Campigli Di Giammartino, Yongsheng Shi, and James L. Manley Supplemental Information
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. Description of the observed lymphatic metastases in two different SIX1-induced MCF7 metastasis models (Nude and NOD/SCID). Supplementary Figure 2. MCF7-SIX1
Supplementary Figures Supplementary Figure 1. Description of the observed lymphatic metastases in two different SIX1-induced MCF7 metastasis models (Nude and NOD/SCID). Supplementary Figure 2. MCF7-SIX1
special offers from your protein biology resource
 special offers from your protein biology resource Pop open your cells, extract your proteins, purify, quantify and express them. Seeking knowledge about proteins with Thermo Scientific Protein Research
special offers from your protein biology resource Pop open your cells, extract your proteins, purify, quantify and express them. Seeking knowledge about proteins with Thermo Scientific Protein Research
Cytomics in Action: Cytokine Network Cytometry
 Cytomics in Action: Cytokine Network Cytometry Jonni S. Moore, Ph.D. Director, Clinical and Research Flow Cytometry and PathBioResource Associate Professor of Pathology & Laboratory Medicine University
Cytomics in Action: Cytokine Network Cytometry Jonni S. Moore, Ph.D. Director, Clinical and Research Flow Cytometry and PathBioResource Associate Professor of Pathology & Laboratory Medicine University
Learning Objectives. Define RNA interference. Define basic terminology. Describe molecular mechanism. Define VSP and relevance
 Learning Objectives Define RNA interference Define basic terminology Describe molecular mechanism Define VSP and relevance Describe role of RNAi in antigenic variation A Nobel Way to Regulate Gene Expression
Learning Objectives Define RNA interference Define basic terminology Describe molecular mechanism Define VSP and relevance Describe role of RNAi in antigenic variation A Nobel Way to Regulate Gene Expression
Supplemental Material to: SRam Sripad, Dongyoung Kim, Raimund Ober, E. Sally Ward
 Landes Bioscience www.landesbioscience.com Supplemental Material to: SRam Sripad, Dongyoung Kim, Raimund Ober, E. Sally Ward The level of HER2 expression is a predictor of antibody- HER2 trafficking behavior
Landes Bioscience www.landesbioscience.com Supplemental Material to: SRam Sripad, Dongyoung Kim, Raimund Ober, E. Sally Ward The level of HER2 expression is a predictor of antibody- HER2 trafficking behavior
The preparation of native chromatin from cultured human cells.
 Native chromatin immunoprecipitation protocol The preparation of native chromatin from cultured human cells. All solutions need to be ice cold. Sucrose containing solutions must be made up fresh on the
Native chromatin immunoprecipitation protocol The preparation of native chromatin from cultured human cells. All solutions need to be ice cold. Sucrose containing solutions must be made up fresh on the
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1: Vector maps of TRMPV and TRMPVIR variants. Many derivatives of TRMPV have been generated and tested. Unless otherwise noted, experiments in this paper use
Supplementary Figure 1 Supplementary Figure 1: Vector maps of TRMPV and TRMPVIR variants. Many derivatives of TRMPV have been generated and tested. Unless otherwise noted, experiments in this paper use
pdsipher and pdsipher -GFP shrna Vector User s Guide
 pdsipher and pdsipher -GFP shrna Vector User s Guide NOTE: PLEASE READ THE ENTIRE PROTOCOL CAREFULLY BEFORE USE Page 1. Introduction... 1 2. Vector Overview... 1 3. Vector Maps 2 4. Materials Provided...
pdsipher and pdsipher -GFP shrna Vector User s Guide NOTE: PLEASE READ THE ENTIRE PROTOCOL CAREFULLY BEFORE USE Page 1. Introduction... 1 2. Vector Overview... 1 3. Vector Maps 2 4. Materials Provided...
1. Introduction. T. Zavašnik-Bergant 1
 Application of monoclonal and ultrathin cryosections in immunogold electron microscopy: a study on recombinant human inhibitor expressed in bacterial cells T. Zavašnik-Bergant 1 1 Department of Biochemistry,
Application of monoclonal and ultrathin cryosections in immunogold electron microscopy: a study on recombinant human inhibitor expressed in bacterial cells T. Zavašnik-Bergant 1 1 Department of Biochemistry,
Product: Arrest-In TM Transfection Reagent for RNAi
 Product: Arrest-In TM Transfection Reagent for RNAi Catalog #: ATR1740, ATR1741, ATR1742, ATR1743 Product Description Arrest-In transfection reagent is a proprietary polymeric formulation, developed and
Product: Arrest-In TM Transfection Reagent for RNAi Catalog #: ATR1740, ATR1741, ATR1742, ATR1743 Product Description Arrest-In transfection reagent is a proprietary polymeric formulation, developed and
Blot: a spot or stain, especially of ink on paper.
 Blotting technique Blot: a spot or stain, especially of ink on paper. 2/27 In molecular biology and genetics, a blot is a method of transferring proteins, DNA or RNA, onto a carrier (for example, a nitrocellulose,pvdf
Blotting technique Blot: a spot or stain, especially of ink on paper. 2/27 In molecular biology and genetics, a blot is a method of transferring proteins, DNA or RNA, onto a carrier (for example, a nitrocellulose,pvdf
Purification of (recombinant) proteins. Pekka Lappalainen, Institute of Biotechnology, University of Helsinki
 proteins. Pekka Lappalainen, Institute of Biotechnology, University of Helsinki") Purification of (recombinant) proteins Pekka Lappalainen, Institute of Biotechnology, University of Helsinki Physical properties of proteins that can be applied for purification -size -charge (isoelectric
Purification of (recombinant) proteins Pekka Lappalainen, Institute of Biotechnology, University of Helsinki Physical properties of proteins that can be applied for purification -size -charge (isoelectric
Stabilization of a virus-like particle and its application as a nanoreactor at physiological conditions
 Supporting Information Stabilization of a virus-like particle and its application as a nanoreactor at physiological conditions Lise Schoonen, b Sjors Maassen, b Roeland J. M. Nolte b and Jan C. M. van
Supporting Information Stabilization of a virus-like particle and its application as a nanoreactor at physiological conditions Lise Schoonen, b Sjors Maassen, b Roeland J. M. Nolte b and Jan C. M. van
Post-expansion antibody delivery, after epitope-preserving homogenization.
 Supplementary Figure 1 Post-expansion antibody delivery, after epitope-preserving homogenization. (a, b) Wide-field fluorescence images of Thy1-YFP-expressing mouse brain hemisphere slice before expansion
Supplementary Figure 1 Post-expansion antibody delivery, after epitope-preserving homogenization. (a, b) Wide-field fluorescence images of Thy1-YFP-expressing mouse brain hemisphere slice before expansion
Masayoshi Honda, Jeehae Park, Robert A. Pugh, Taekjip Ha, and Maria Spies
 Molecular Cell, Volume 35 Supplemental Data Single-Molecule Analysis Reveals Differential Effect of ssdna-binding Proteins on DNA Translocation by XPD Helicase Masayoshi Honda, Jeehae Park, Robert A. Pugh,
Molecular Cell, Volume 35 Supplemental Data Single-Molecule Analysis Reveals Differential Effect of ssdna-binding Proteins on DNA Translocation by XPD Helicase Masayoshi Honda, Jeehae Park, Robert A. Pugh,
RNA oligonucleotides and 2 -O-methylated oligonucleotides were synthesized by. 5 AGACACAAACACCAUUGUCACACUCCACAGC; Rand-2 OMe,
 Materials and methods Oligonucleotides and DNA constructs RNA oligonucleotides and 2 -O-methylated oligonucleotides were synthesized by Dharmacon Inc. (Lafayette, CO). The sequences were: 122-2 OMe, 5
Materials and methods Oligonucleotides and DNA constructs RNA oligonucleotides and 2 -O-methylated oligonucleotides were synthesized by Dharmacon Inc. (Lafayette, CO). The sequences were: 122-2 OMe, 5
Supplementary Figure 1. Isolation of GFPHigh cells.
 Supplementary Figure 1. Isolation of GFP High cells. (A) Schematic diagram of cell isolation based on Wnt signaling activity. Colorectal cancer (CRC) cell lines were stably transduced with lentivirus encoding
Supplementary Figure 1. Isolation of GFP High cells. (A) Schematic diagram of cell isolation based on Wnt signaling activity. Colorectal cancer (CRC) cell lines were stably transduced with lentivirus encoding