MOLECULAR CELL BIOLOGY
|
|
|
- Roberta Jennings
- 5 years ago
- Views:
Transcription
1 Lodish Berk Kaiser Krieger scott Bretscher Ploegh Matsudaira MOLECULAR CELL BIOLOGY SEVENTH EDITION CHAPTER 19 The Eukaryotic Cell Cycle Copyright 2013 by W. H. Freeman and Company

2 Figure 19.1 The fate of a single parental chromosome throughout the eukaryotic cell cycle.
3 The fate of a single parental chromosome throughout the eukaryotic cell cycle 1. The cell cycle entails an ordered series of macromolecular events that lead to cell division and the production of two daughter cells each containing chromosomes identical to those of the parental cell. As illustrated in Figure 19-1, the cell cycle is divided into four major phases. In cycling (replicating) somatic cells, cells synthesize RNAs and proteins during the G1 phase, preparing for DNA synthesis and chromosome replication during the S (synthesis) phase. After progressing through the G2 phase, cells begin the complicated process of mitosis, also called the M (mitotic) phase, which is divided into several stages. In multicellular organisms, most differentiated cells exit the cell cycle and survive for days, weeks, or in some cases (e.g., nerve cells and cells of the eye lens) even the lifetime of the organism without dividing again. Such postmitotic cells generally exit the cell cycle in G1, entering a phase called G0 (Figure 19-1). Some G0 cells can return to the cell cycle and resume replicating; this reentry is regulated, thereby providing control of cell proliferation. 2. In vertebrates and diploid yeasts, cells in G1 have a diploid number of chromosomes (2n), one inherited from each parent. In haploid yeasts, cells in G1 have one of each chromosome (1n), the haploid number. Rapidly replicating human cells progress through the full cell cycle in about 24 hours: mitosis takes 30 minutes; G1, 9 hours; the S phase, 10 hours; and G2, 4.5 hours.

4 Figure 19.3 The budding yeast S. cerevisiae.

5 Figure 19.4 The fission yeast S. pombe.
6 Experimental Figure 19.5 Wild-type cell division cycle (CDC) genes can be isolated from a S. cerevisiae genomic library by functional complementation of cdc mutants.
7 Genetic Studies with S. cerevisiae and S. pombe 1. S. cerevisiae cells replicate by budding (Figure 19-3). Both mother and daughter cells remain in the G1 period of the cell cycle while growing, although it takes the initially larger mother cells a shorter time to reach a size compatible with cell division. When S. cerevisiae cells in G1 have grown sufficiently, they begin a program of gene expression that leads to entry into the S phase. If G1 cells are shifted from a rich medium to a medium low in nutrients before they reach a critical size, they remain in G1 and grow slowly until they are large enough to enter the S phase. However, once G1 cells reach the critical size, they become committed to completing the cell cycle, entering the S phase and proceeding through G2 and mitosis, even if they are shifted to a medium low in nutrients. The point in late G1 of growing S. cerevisiae cells when they become irrevocably committed to entering the S phase and traversing the cell cycle is called START. 2. Identification of the catalytic protein kinase subunit of MPF and further insight into its regulation came from genetic analysis of the cell cycle in the fission yeast S. pombe. This yeast grows as a rod-shaped cell that increases in length as it grows and then divides in the middle during mitosis to produce two daughter cells of equal size (Figure 19-4). 3. The phenotypic behavior of temperature-sensitive cdc28 mutants indicates that Cdc28 function is critical for entry into the S phase. When these mutants are shifted to the nonpermissive temperature, they behave like wild-type cells suddenly deprived of nutrients. That is, cdc28 mutant cells that have grown large enough to pass START at the time of the temperature shift continue through the cell cycle normally and undergo mitosis, whereas those that are too small to have passed START when shifted to the nonpermissive temperature do not enter the S phase even though nutrients are plentiful. Even though cdc28 cells blocked in G1 continue to grow in size at the nonpermissive temperature, they cannot pass START and enter the S phase. Thus they appear as large cells with no bud. The wild-type CDC28 gene was isolated by its ability to complement mutant cdc28 cells at the nonpermissive temperature. Note that in S. cerevisiae wild-type genes are indicated in italic capital letters (e.g., CDC28) and recessive mutant genes in italic lowercase letters (e.g., cdc28); the corresponding wildtype protein is written in Roman letters with an initial capital (e.g., Cdc28), similar to S. pombe proteins. Each type of yeast contains a single cyclin-dependent protein kinase (CDK), which can substitute for each other: Cdc2 in S. pombe and Cdc28 in S. cerevisiae
8

9 Figure Molecular Biology of the Cell ( Garland Science 2008)
10 Checkpoints in the cell-cycle control system. In most cells there are several points in the cell cycle, called checkpoints, at which the cycle can be arrested if previous events have not been completed. Entry into mitosis is prevented, for example, when DNA replication is not complete, and chromosome separation in mitosis is delayed if some chromosomes are not properly attached to the mitotic spindle. Progression through G1 and G2 is delayed by braking mechanisms if the DNA in the chromosomes is damaged by radiation or chemicals. Delays at these DNA damage checkpoints provide time for the damaged DNA to be repaired, after which the cellcycle brakes are released and progress resumes. Checkpoints are important in another way as well. They are points in the cell cycle at which the control system can be regulated by extracellular signals from other cells. These signals which can either promote or inhibit cell proliferation tend to act by regulating progression through a G1 checkpoint, using mechanisms discussed later in the chapter. (Figure Molecular Biology of the Cell -
11 Figure Regulation of cell cycle transitions.
12 Regulation of cell cycle transitions Figure outlines the role of the three major classes of cyclin-cdk complexes that control passage through the cell cycle: the G1, S-phase, and mitotic cyclin-cdk complexes. 1. When cells are stimulated to replicate, G1 cyclin-cdk complexes are expressed first. These prepare the cell for the S phase by activating transcription factors that promote transcription of genes encoding enzymes required for DNA synthesis and the genes encoding S-phase cyclins and CDKs. The activity of S-phase cyclin-cdk complexes is initially held in check by inhibitors. Late in G1, the G1 cyclin-cdk complexes induce degradation of the S-phase inhibitors by phosphorylating them and consequently stimulating their polyubiquitination by the multiprotein SCF ubiquitin ligase. Subsequent degradation of the polyubiquitinated S-phase inhibitor by proteasomes releases active S-phase cyclin-cdk complexes. 2. Once activated, the S-phase cyclin-cdk complexes phosphorylate regulatory sites in the proteins that form DNA pre-replication complexes, which are assembled on replication origins during G1. Phosphorylation of these proteins not only activates initiation of DNA replication but also prevents reassembly of new pre-replication complexes. 3. Mitotic cyclin-cdk complexes are synthesized during the S phase and G2, but their activities are held in check by phosphorylation at inhibitory sites until DNA synthesis is completed. Once activated by dephosphorylation of the inhibitory sites, mitotic cyclin-cdk complexes phosphorylate multiple proteins that promote chromosome condensation, retraction of the nuclear envelope, assembly of the mitotic spindle apparatus, and alignment of condensed chromosomes at the metaphase plate. During mitosis, the anaphasepromoting complex (APC), a multisubunit ubiquitin ligase, polyubiquitinates key regulatory proteins marking them for proteasomal degradation. Once all the chromosomes are aligned, the APC polyubiquitinates securin, leading to its proteasomal degradation and the subsequent degradation of the cross-linking proteins connecting sister chromatids. This sequence of events initiates anaphase by freeing sister chromatids to segregate to opposite spindle poles. Late in anaphase, the APC also directs polyubiquitination and subsequent proteasomal degradation of the mitotic cyclins. As a result, the now separated chromosomes decondense, the nuclear envelope re-forms around daughter-cell nuclei, and the Golgi apparatus reassembles during telophase; finally, the cytoplasm divides at cytokinesis, yielding the two daughter cells.

13 Figure Molecular Biology of the Cell ( Garland Science 2008)
14 Figure An overview of how CDKs regulate cell cycle progression.
15 Regulation of CDK activity 1. The concentrations of the cyclins, the regulatory subunits of the heterodimeric protein kinases that control cell-cycle events, increase and decrease as cells progress through the cell cycle. The catalytic subunits of these kinases, called cyclin-dependent kinases (CDKs), have no kinase activity unless they are associated with a cyclin. Each CDK can associate with different cyclins, and the associated cyclin determines which proteins are phosphorylated by a particular cyclin-cdk complex (Figure Molecular Biology of the Cell). 2. A key discovery in cell cycle studies was that cyclin-dependent kinases govern progression through the cell cycle. Three key features about these kinases are important to keep in mind throughout this chapter: Cyclin-dependent kinases (CDKs) are only active when bound to a regulatory cyclin subunit. Different types of cyclin-cdk complexes initiate different events. G1 CDKs and G1/S phase CDKs promote entry into the cell cycle, S phase CDKs trigger S phase, and mitotic CDKs initiate the events of mitosis (Figure 19-11). Multiple mechanisms are in place to ensure that the different CDKs are only active in the stages of the cell cycle they trigger.

16 Figure Structural models of human CDK2.
17 Structural models of human CDK2 Unlike both fission and budding yeasts, each of which produce just one CDK, vertebrates produce several CDKs. The three-dimensional structure of one human cyclin-dependent kinase (CDK2) has been determined and provides insight into how cyclin binding and phosphorylation of CDKs regulate their protein kinase activity Unphosphorylated, inactive CDK2 contains a flexible region, called the T-loop, that blocks access of protein substrates to the active site where ATP is bound (Figure 19-12a). Unphosphorylated CDK2 bound to one of its cyclin partners, cyclin A, has minimal but detectable protein kinase activity in vitro, although it may be essentially inactive in vivo. Extensive interactions between cyclin A and the T-loop cause a dramatic shift in the position of the T-loop, thereby exposing the CDK2 active site (Figure 19-12b). Binding of cyclin A also shifts the position of the 1 helix in CDK2, modifying its substrate-binding surface. Phosphorylation of the activating threonine, located in the T-loop, causes additional conformational changes in the cyclin A CDK2 complex that greatly increase its affinity for protein substrates (Figure 19-12c).

18 Experimental Figure Mitotic cyclins are rate limiting for mitosis.
19 Mitotic cyclins are rate limiting for mitosis In all cases, MPF activity and cyclin B concentration were determined at various times after addition of sperm nuclei to a Xenopus egg extract treated as indicated in each panel. 1. These studies showed that MPF activity rises and falls in synchrony with the concentration of cyclin B (Figure 19-13a). The early events of mitosis chromosome condensation and nuclear envelope disassembly occurred when MPF activity reached its highest levels in parallel with the rise in cyclin B concentration. Addition of cycloheximide, an inhibitor of protein synthesis, prevented cyclin B synthesis and also prevented the rise in MPF activity, chromosome condensation, and nuclear envelope disassembly. 2. To test the functions of cyclin B in these cell-cycle events, all mrnas in the egg extract were degraded by digestion with a low concentration of RNase, which then was inactivated by addition of a specific inhibitor. This treatment destroys mrnas without affecting the trnas and rrnas required for protein synthesis, since their degradation requires much higher concentrations of RNase. When sperm nuclei were added to the RNase-treated extracts, the 1n nuclei replicated their DNA, but the increase in MPF activity and the early mitotic events (chromosome condensation and nuclear envelope disassembly), which the untreated extract supports, did not occur (Figure 19-13b). 3. Addition of cyclin B mrna, produced in vitro from cloned cyclin B cdna, to the RNase-treated egg extract and sperm nuclei restored the parallel oscillations in the cyclin B concentration and MPF activity and the characteristic early and late mitotic events as observed with the untreated egg extract (Figure 19-13c). Since cyclin B is the only protein synthesized under these conditions, these results demonstrate that it is the crucial protein whose synthesis is required to regulate MPF activity and the cycles of chromosome condensation and nuclear envelope breakdown mediated by cycling Xenopus egg extracts. 4. In these experiments, chromosome decondensation and nuclear envelope assembly (late mitotic events) coincided with decreases in the cyclin B level and MPF activity. To determine whether degradation of cyclin B is required for exit from mitosis, researchers added a mutant mrna encoding a nondegradable cyclin B to a mixture of RNase-treated Xenopus egg extract and sperm nuclei. As shown in Figure 19-13d, MPF activity increased in parallel with the level of the mutant cyclin B, triggering condensation of the sperm chromatin and nuclear envelope disassembly (early mitotic events). However, the mutant cyclin B produced in this reaction never was degraded. As a consequence, MPF activity remained elevated, and the late mitotic events of chromosome decondensation and nuclear envelope formation were both blocked. This experiment demonstrates that the fall in MPF activity and exit from mitosis depend on degradation of cyclin B. The results of the two experiments with RNase-treated extracts show that entry into mitosis requires the accumulation

20
21 Cyclins determine the activity of CDKs 1. Cyclins are so named because their levels change during the cell cycle. They form a family of proteins that is defined by three key features: Cyclins bind to and activate CDKs. The activity and substrate specificity of any given CDK is primarily defined by the particular cyclin to which it is bound. Cyclins are only present during the cell cycle stage that they trigger and are absent in other cell cycle stage. Cyclins not only regulate a particular cell cycle stage but also set in motion a series of events in preparation for the next cell cycle stage. In this way, they propel the cell cycle forword. 2. Cyclins are divided into four classes defined by their presence and activity during the cell cycle: G1 cyclins, G1/S cyclins, S phase cyclins, and mitotic cyclins (Table 19-1).

22 Figure Molecular Biology of the Cell ( Garland Science 2008)
23 The activation of M-Cdk The activation of M-Cdk begins with the accumulation of M-cyclin (cyclin B in vertebrate cells). This increase in M-cyclin protein leads to a gradual accumulation of M-Cdk (the complex of Cdk1 and M-cyclin) as the cell approaches mitosis. Although the Cdk in these complexes is phosphorylated at an activating site by the enzyme CAK discussed earlier, it is held in an inactive state by inhibitory phosphorylation at two neighboring sites by the protein kinase Wee 1. Thus, by the time the cell reaches the end of G2, it contains an abundant stockpile of M-Cdk that is primed and ready to act, but the M-Cdk activity is repressed by the presence of two phosphate groups that block the active site of the kinase. What, then, triggers the activation of the M-Cdk stockpile? The crucial event is the activation in late G2 of the protein phosphatase Cdc25, which removes the inhibitory phosphates that restrain M-Cdk. At the same time, the activity of the inhibitory kinase Wee1 is also suppressed, further ensuring that M-Cdk activity increases abruptly. Two protein kinases activate Cdc25. One, known as Polo kinase, phosphorylates Cdc25 at one set of sites. The other activating kinase is M-Cdk itself, which phosphorylates a different set of sites on Cdc25. M-Cdk also phosphorylates and inhibits Wee1. (Figure Molecular Biology of the Cell -

24 CDK inhibitory protein (CKI) Figure Molecular Biology of the Cell ( Garland Science 2008)
25 CDK inhibitory protein (CKI) Cyclin-Cdk complexes can also be regulated by the binding of Cdk inhibitor proteins (CKIs). There are a variety of CKI proteins, and they are primarily employed in the control of G1 and S phase. The threedimensional structure of a cyclin-cdk-cki complex reveals that CKI binding dramatically rearranges the structure of the Cdk active site, rendering it inactive (Figure 17-19). The final layer of control that is of proteins known as CDK inhibitors, or CKIs, that directly bind to the cyclin-cdk complex and inhibit its activity. As we will see in Section 19.4, these proteins play an especially important role in the regulation of the G1-S phase transition and its integration with extracellular signals. It thus comes as no surprise that the genes encoding these CKIs are often found mutated in human cancers. (Figure Molecular Biology of the Cell -
26
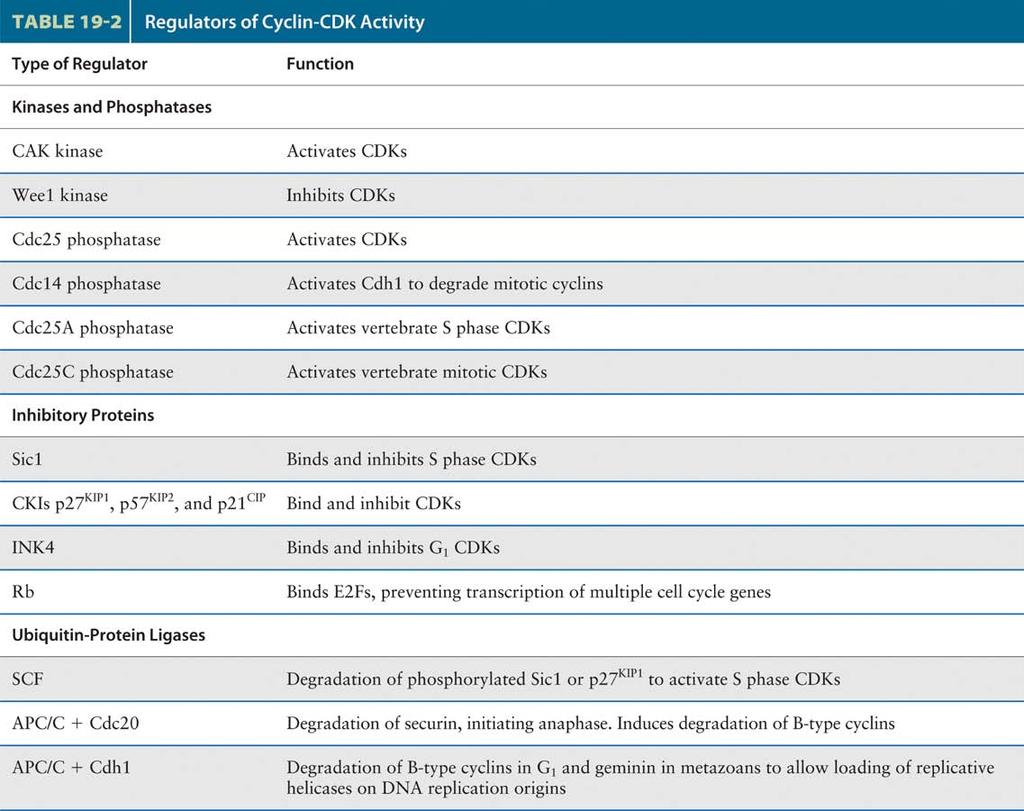
27 Regulators of Cyclin-CDK activity Multiple mechanisms ensure that CDKs are active in the right stage of the cell cycle. Table 19-2 lists these key regulators of CDKs. Four proteins regulate the protein kinase activity of the S. pombe CDK. First is Cdc13, the mitotic cyclin of S. pombe (equivalent to cyclin B in metazoans), which associates with the CDK to form MPF with extremely low activity. 1. Kinases and phosphatases Second is the Wee1 protein-tyrosine kinase, which phosphorylates an inhibitory tyrosine residue (Y15) in the CDK subunit. Third is another kinase, designated CDK-activating kinase (CAK), which phosphorylates an activating threonine residue (T161). When both residues are phosphorylated, MPF is inactive. Finally, the Cdc25 phosphatase removes the phosphate from Y15, yielding highly active MPF. 2. Inhibitory proteins As the S-phase cyclin-cdk heterodimers accumulate in late G1, they are immediately inactivated by binding of an inhibitor, called Sic1, that is expressed late in mitosis and in early G1. Because Sic1 specifically inhibits B-type cyclin-cdk complexes, but has no effect on the G1 cyclin-cdk complexes, it functions as an S-phase inhibitor. All eukaryotes habor CKIs involved in regulating S phase and mitotic CDKs. Although these inhibitors display little sequence similarity, they are all essential to prevent premature activation of S phase and M phase CDKs. Inhibitors of G1 CDKs play an essential role in mediating a G1 arrest in response to proliferation inhibitory signals. A three proteins-p21cip, p27kip1, and p57kip2. These CKIs and must be degraded before DNA replication can begin. A class of CKIs called INK4s (inhibitors of kinase 4) includes several small, closely related proteins that interact only with the G1 CDKs. Binding of INK4s to CDK4 and CDK6 blocks their interaction with cyclin D and hence their protein kinase activity.
28

29 Mitotic CDK (mitosis-promoting factor (MPF), CDK1/cycinB) Figure Fundamental processes in the eukaryotic cell cycle.
 chromosome")
30 MPF 1) chromosome condensation 2) nuclear envelope breakdown 3) chromosome segregation
31 Mitotic CDK (mitosis-promoting factor (MPF), CDK1/cycinB) A heterodimer composed of a mitotic cyclin and CDK is commonly referred to as a mitosis-promoting factor (MPF). As we will see shortly, MPF turned out to be the key factor that regulates the initiation of mitosis in all eukaryotic cells. Presumably, the entry into mitosis is a consequence of the phosphorylation of specific proteins by the protein kinase activity of MPF. Although many of the critical substrates of MPF remain to be identified, we now know of examples that show how regulation by MPF phosphorylation mediates many of the early events of mitosis leading to metaphase: chromosome condensation, formation of the mitotic spindle, and disassembly of the nuclear envelope. Recall that a decrease in mitotic cyclins and the associated inactivation of MPF coincides with the later stages of mitosis. Just before this, in early anaphase, sister chromatids separate and move to opposite spindle poles. During telophase, microtubule dynamics return to interphase conditions, the chromosomes decondense, the nuclear envelope re-forms, the Golgi complex is remodeled, and cytokinesis occurs. Some of these processes are triggered by dephosphorylation; others, by protein degradation.

32 Figure The nuclear lamina and its regulation by phosphorylation. Figure Nuclear envelope proteins phosphorylated by mitotic CDKs.


33 Figure The nuclear lamina and its regulation by phosphorylation.
34 Figure Nuclear envelope proteins phosphorylated by mitotic CDKs.
35 Nuclear envelope breakdown The nuclear envelope is a double-membrane extension of the rough endoplasmic reticulum containing many nuclear pore complexes. The lipid bilayer of the inner nuclear membrane is supported by the nuclear lamina, a meshwork of lamin filaments located adjacent to the inside face of the nuclear envelope (Figure 19-22a). The three nuclear lamins (A, B, and C) present in vertebrate cells belong to the class of cytoskeletal proteins, the intermediate filaments, that are critical in supporting cellular membranes. All three nuclear lamins form dimers containing a rodlike α-helical coiled-coil central section and globular head and tail domains; polymerization of these dimers through head-to-head and tail-to-tail associations generates the intermediate filaments that compose the nuclear lamina. 1. Early in mitosis, MPF phosphorylates specific serine residues in all three nuclear lamins, causing depolymerization of the lamin intermediate filaments (Figure 19-22b). The phosphorylated lamin A and C dimers are released into solution, whereas the phosphorylated lamin B dimers remain associated with the nuclear membrane via their isoprenyl anchor. Depolymerization of the nuclear lamins leads to disintegration of the nuclear lamina meshwork and contributes to disassembly of the nuclear envelope. Disassembly of the nuclear envelope, which normally occurs early in mitosis, depends on phosphorylation of lamin A. 2. In addition, MPF-catalyzed phosphorylation of specific nucleoporins causes nuclear pore complexes to dissociate into subcomplexes during prophase. Similarly, phosphorylation of integral membrane proteins of the inner nuclear membrane is thought to decrease their affinity for chromatin and contribute to disassembly of the nuclear envelope. The weakening of the associations between the inner nuclear membrane and the nuclear lamina and chromatin may allow sheets of inner nuclear membrane to retract into the endoplasmic reticulum, which is continuous with the outer nuclear membrane (Figure 19-23).
36
37 Metaphase-to-anaphase transition Figure 19.2 The stages of mitosis.
 Figure 19.")
38 Figure Molecular Biology of the Cell ( Garland Science 2008) Figure Regulation of cohesin cleavage.
39 Figure Regulation of cohesin cleavage.
40 3. Because Cdc20 the specificity factor that directs APC to securin is activated before Cdh1 the specificity factor that directs APC to mitotic cyclins MPF activity does not decrease until after the chromosomes have segregated. As a result of this temporal order in the activation of the two APC specificity factors (Cdc20 and Cdh1), the chromosomes remain in the condensed state and reassembly of the nuclear envelope does not occur until chromosomes are moved to the proper position (Figure Molecular Biology of the Cell). Chromosome segregation 1. After M-Cdk has triggered the complex rearrangements that occur in early mitosis, the cell cycle reaches its culmination with the separation of the sister chromatids at the metaphase-toanaphase transition. As mentioned earlier, each sister chromatid of a metaphase chromosome is attached to microtubules via its kinetochore, a complex of proteins assembled at the centromere. The opposite ends of these kinetochore microtubules associate with one of the spindle poles. At metaphase, the spindle is in a state of tension with forces pulling the two kinetochores toward the opposite spindle poles balanced by forces pushing the spindle poles apart. Sister chromatids do not separate because they are held together at their centromeres and multiple positions along the chromosome arms by multiprotein complexes called cohesins. Among the proteins composing the cohesin complexes are members of the SMC protein family. 2. Recent genetic studies in the budding yeast S. cerevisiae have led to the model depicted in Figure for how the APC regulates sister chromatid separation to initiate anaphase. Cohesin SMC proteins bind to each sister chromatid; other subunits of cohesin, including Scc1, then link the SMC proteins, firmly associating the two chromatids. The cross-linking activity of cohesin depends on securin, which is found in all eukaryotes. Prior to anaphase, securin binds to and inhibits separase, a ubiquitous protease related to the caspase proteases that regulate programmed cell death. Once all chromosome kinetochores have attached to spindle microtubules, the APC is directed by a specificity factor called Cdc20 to polyubiquitinate securin, leading to the onset of anaphase. (This specificity factor is distinct from Cdh1, which directs the APC to polyubiquitinate B-type cyclins.) Polyubiquitinated securin is rapidly degraded by proteasomes, thereby releasing separase. Free from its inhibitor, separase cleaves Scc1, breaking the protein cross-link between sister chromatids. Once this link is broken, the poleward force exerted on kinetochores can move sister chromatids toward the opposite spindle poles.
 Nat Rev Cancer (2006)")
41 Anaphase-promoting complex/cyclosome (APC/C) Nat Rev Cancer (2006) 6(5):
42 Anaphase-promoting complex/cyclosome (APC/C) 1. Levels of cyclin expression during cell division are periodic. This is the result of a constant synthetic rate coupled with a defined window in the cycle of specific proteolysis, which is executed by the ubiquitin proteasome system (UPS). CKIs, negative-regulators of cyclin CDK kinase complexes, are also targeted for degradation by the UPS. Therefore, the cell cycle is predominantly regulated by two types of posttranslational protein modification phosphorylation and ubiquitylation. The UPS comprises two discrete steps: the covalent attachment of multiple ubiquitin molecules to the protein substrate and degradation of the polyubiquitylated protein by the 26S proteasome complex3 (FIG. 1). The first step is mediated by at least three enzymes: a ubiquitin-activating enzyme (E1), a ubiquitin-conjugating enzyme (E2) and a ubiquitin ligase (E3). An involvement of E1 in cancer has not been described, and only a few reports have linked E2 to cancer development4. By contrast, a large amount of evidence indicates that deregulation of E3 ligase, which is involved in many biological systems, often results in cancer development. 2. Overview of the ubiquitin-proteasome pathway. Ubiquitin (Ub) is a small 8-kDa protein, which is first transferred to the ubiquitin-activating enzyme, E1, in an ATPdependent manner. This activated ubiquitin is then transferred to the ubiquitin conjugating enzyme, E2. Finally, the ubiquitin is covalently attached to the target protein by E3 ubiquitin ligase, leading to the formation of a polyubiquitin chain. The polyubiquitylated protein is recognized by the 26S proteasome, and is destroyed in an ATP-dependent manner. There are many E3 ubiquitin ligases, which are categorized into four major classes: HECT-type, RING-finger-type, U- box-type and PHD-finger-type. RING-finger-type E3s are further divided into subfamilies, including cullinbased E3s, which constitute one of the largest classes of E3s. There are seven cullin-based E3s including the SKP1 CUL1 F-box-protein (SCF) complex and the anaphase-promoting complex/cyclosome (APC/C) (Nat Rev Cancer (2006) 6(5): ).
43 Nat Rev Cancer (2006) 6(5):
44 The structural similarity of SCF and APC/C SCF and APC/C: similar but distinct. E3 components in the UPS are thought to be primarily responsible for the specific recognition of a large number of target proteins. This requires both specificity and versatility, which are provided by the existence of 500 1,000 different E3 ligases. These are now categorized into four major classes on the basis of their specific structural motif: HECT-type, RING finger-type, U-box-type or PHD-finger-type. RING-finger-type E3s are thought to be the largest family and are further divided into subfamilies; one of these, the cullin-based E3 subfamily, is one of the largest single classes of E3. There are seven cullin-based E3s, including the SKP1 CUL1 F-box-protein (SCF) complex and the anaphasepromoting complex/cyclosome (APC/C), both of which are involved in the proteolysis of core components of the cell-cycle machinery. The SCF complex consists of three invariable components RBX1 (RING-finger protein), CUL1 (scaffold protein), and SKP1 (adaptor protein) as well as one variable component, known as an F-box protein, that binds through its F-box motif to SKP1 and is responsible for substrate recognition (FIG. 2). The APC/C consists of the invariable core components APC1 (RBX1-related RING-finger protein), APC2 (CUL1-related scaffold protein) and at least 11 other components, the functions of which are not fully understood. In addition, there is the variable activator subunit, cell division cycle 20 (CDC20) or CDH1 (also known as HCT1), both of which confer substrate specificity in the same way as F-box proteins do in the SCF complex. Known substrates of each variable subunit in SCF and APC/C are shown (Nat Rev Cancer (2006) 6(5): )
45 Figure Fundamental processes in the eukaryotic cell cycle.
46 APC/C-CDC20/proteasome degrades securin APC/C-CDC20 contributes to the proteolysis of securin, and thereby triggers chromosomal separation at anaphase. After replication, sister chromatids are accompanied until anaphase by the multiprotein complex cohesin. Separase cleaves cohesin, but its activity is suppressed by securin. The degradation of securin by APC/C-CDC20 activates separase, resulting in the cleavage of cohesin and separation of sister chromatids (Figure 19-30, Nat Rev Cancer (2006) 6(5): ). Because Cdc20 the specificity factor that directs APC to securin is activated before Cdh1 the specificity factor that directs APC to mitotic cyclins MPF activity does not decrease until after the chromosomes have segregated (Figure 19-30). As a result of this temporal order in the activation of the two APC specificity factors (Cdc20 and Cdh1), the chromosomes remain in the condensed state and reassembly of the nuclear envelope does not occur until chromosomes are moved to the proper position.
47
 reformation of interphase MT array")
48 chromosome decondensation Cyclin B should be degraded for 1) chromosome decondensation 2) reformation of interphase MT array MPF
49 Nat Rev Cancer (2006) 6(5):
50 Figure Fundamental processes in the eukaryotic cell cycle.
51 APC/C-Cdh1/proteasome degrades mitotic cyclins 1. We saw earlier that in late anaphase, polyubiquitination of mitotic cyclin by the anaphase-promoting complex (APC) leads to the proteasomal destruction of this cyclin. Additional experiments with Xenopus egg extracts provided evidence that degradation of cyclin B, the Xenopus mitotic cyclin, and the resulting decrease in MPF activity are required for chromosome decondensation but not for chromosome segregation. The fall in MPF activity in telophase allows constitutive protein phosphatases to remove the regulatory phosphates from condensin, lamins, nucleoporins, and other nuclear membrane proteins, permitting the decondensation of chromosomes and the reassembly of the nuclear membrane, nuclear lamina, and nuclear pore complexes. When mitosis is complete, the spindle disassembles and the interphase microtubule network re-forms (Figure 18-34a part 2 6 th ed). 2. The APC/C is structurally similar to the SCF complex, and consists of invariable core components APC11 (RBX1-related RING-finger protein), APC2 (CUL1-related scaffold protein) and at least 11 other components without a defined role as well as a variable component known as an activator. There are two such variable components in mitotically cycling cells cell division cycle 20 (CDC20) and CDH1 (also known as HCT1) and they confer substrate specificity in the same way that F-box proteins do in the SCF complex. In late M phase, CDC20 is ubiquitylated, and therefore degraded, by APC/C-CDH1 or by itself, leading to the transition from APC/C-CDC20 to APC/C-CDH1 (Nat Rev Cancer (2006) 6(5): ). 3. Cyclin B, the primary mitotic cyclin in multicellular animals (metazoans), is synthesized throughout the cell cycle from a stable mrna. The observed fall in its concentration in late anaphase results from its APCstimulated degradation at this point in the cell cycle. Genetic studies with yeast led to identification of an APC specificity factor, called Cdh1, that binds to APC and directs it to polyubiquitinate mitotic cyclins. This specificity factor is active only in late anaphase when the segregating chromosomes have moved far enough apart in the dividing cell to assure that both daughter cells will contain one complete set of chromosomes. Phosphorylation of Cdh1 by other cyclin-cdk complexes during G1 inhibits its association with the APC and thus degradation of mitotic cyclin (Figure 19-30). This inhibition permits the gradual rise in mitotic cyclin levels observed throughout interphase of the next cell cycle.
52 Figure Regulation of S phase and mitotic cyclin levels in budding yeast.
53 Regulation of S phase and mitotic cyclin levels in budding yeast One of the important substrates of the G1/S phase cyclin-cdk complexes is Cdh1. During late anaphase, this substrate targeting factor directs the APC/C to ubiquitinylate substrate protein including S phase and mitotic cyclins, marking them for proteolysis by proteasomes. The APC/C-Cdh1 remains active throughout G1, preventing the premature accumulation of S phase and mitotic cyclins. 1. Phosphorylation of Cdh1 by G1/S cyclin-cdks causes it to dissociate from the APC/C complex, inhibiting further ubiquitinylation of S phase and mitotic cyclins during late G1. This, combined with the induced transcription of S phase cyclins during late G1, allows S phase cyclin proteins to accumulate as G1/S cyclin-cdks levels rise. Later in the cell cycle, S phase and mitotic CDKs take over to maintain Cdh1 in the phosphorylated and hence inactive state. 2. Only as mitotic CDKs decline and a protein phosphatase known as Cdc14 is activated are these inhibitory phosphates removed from Cdh1, leading to its reactivation. In mammalian cells similar mechanisms are responsible for stabilizing S phase and mitotic cyclins, but the phosphatase involved in dephosphorylation of Cdh1 has not been identified (Figure 19-16).
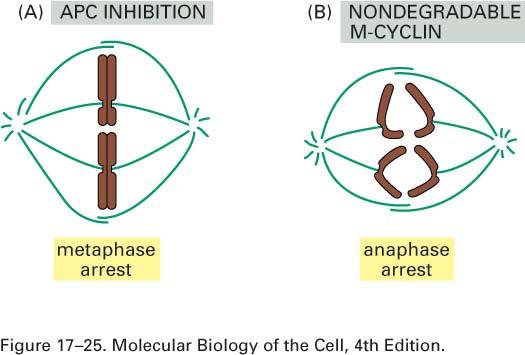
54
55 Two experiments that demonstrate the requirement for protein degradation to exit from mitosis Anaphase begins with a sudden disruption of the cohesion between sister chromatids, which allows them to separate and move to opposite poles of the spindle. This process is initiated by a remarkable cascade of signaling events. The sister-chromatid separation requires the activation of the APC enzyme complex, suggesting that proteolysis is central to the process. 1. An APC inhibitor was added to frog egg extracts undergoing mitosis in vitro. The inhibitor arrested mitosis at metaphase, indicating that proteolysis is required for the separation of sister chromatids at the metaphase-to-anaphase transition. A similar arrest occurs in budding yeasts with mutations in components of the APC. 2. A nondegradable mutant form of M-cyclin was added to mitotic frog egg extracts. This addition arrested mitosis after sister-chromatid separation, indicating that destruction of M-cyclin is not required for sisterchromatid separation but is required for the subsequent exit from mitosis. (Figure Molecular Biology of the Cell -
56


57 Figure 17-20b Molecular Biology of the Cell ( Garland Science 2008)
58 The cell-cycle control system depends on cyclical proteolysis 1. Cell-cycle control depends crucially on at least two distinct enzyme complexes that act at different times in the cycle to cause the proteolysis of key proteins of the cell-cycle control system, thereby inactivating them. Most notably, cyclin-cdk complexes are inactivated by regulated proteolysis of cyclins at certain cellcycle stages. This cyclin destruction occurs by a ubiquitin-dependent mechanism, like that involved in the proteolysis of many other intracellular proteins. An activated enzyme complex recognizes specific aminoacid sequences on the cyclin and attaches multiple copies of ubiquitin to it, marking the protein for complete destruction in proteasomes. 2. The rate-limiting step in cyclin destruction is the final ubiquitin-transfer reaction catalyzed by enzymes known as ubiquitin ligases. Two ubiquitin ligases are important in the destruction of cyclins and other cell-cycle regulators. 1 2 In G1 and S phase, an enzyme complex called SCF (after its three main protein subunits) is responsible for the ubiquitylation and destruction of G1/S-cyclins and certain CKI proteins that control S-phase initiation. In M phase, the anaphase-promoting complex (APC) is responsible for the ubiquitylation and proteolysis of M-cyclins and other regulators of mitosis. 3. These two large, multisubunit complexes contain some related components, but they are regulated in different ways. SCF activity is constant during the cell cycle. Ubiquitylation by SCF is controlled by changes in the phosphorylation state of its target proteins: only specifically phosphorylated proteins are recognized, ubiquitylated, and destroyed. APC activity, by contrast, changes at different stages of the cell cycle. APC is turned on mainly by the addition of activating subunits to the complex. (Figure 17-20b Molecular Biology of the Cell -
59
60 Figure Fundamental processes in the eukaryotic cell cycle. G1 CDK
61
62
63 G1 CDK 1. When cells are stimulated to replicate, G1 cyclin-cdk complexes are expressed first. These prepare the cell for the S phase by activating transcription factors that promote transcription of genes encoding enzymes required for DNA synthesis and the genes encoding S-phase cyclins and CDKs. During early G1 of the next cell cycle, phosphatases dephosphorylate the proteins that form pre-replication complexes. These proteins had been phosphorylated by S-phase cyclin-cdk complexes during the previous S phase, and their phosphorylation was maintained during mitosis by mitotic cyclin-cdk complexes. As a result of their dephosphorylation in G1, new pre-replication complexes are able to reassemble at replication origins in preparation for the next S phase (Figure 19-30). Phosphorylation of Cdh1 by G1 cyclin-cdk complexes in late G1 inactivates it, allowing accumulation of S-phase and mitotic cyclins during the ensuing cycle. The activity of S-phase cyclin-cdk complexes is initially held in check by inhibitors. Late in G1, the G1 cyclin-cdk complexes induce degradation of the S-phase inhibitors by phosphorylating them and consequently stimulating their polyubiquitination by the multiprotein SCF ubiquitin ligase. Subsequent degradation of the polyubiquitinated S-phase inhibitor by proteasomes releases active S-phase cyclin-cdk complexes. 2. Cyclin D and E are the mammalian G1 cyclins. Experiments in which cultured mammalian cells were microinjected with anti-cyclin D antibody at various times after addition of growth factors demonstrated that cyclin D is essential for passage through the restriction point. Figure (6 th ed.) presents a current model for the periods of the cell cycle in which different cyclin-cdk complexes act in G0-arrested mammalian cells stimulated to divide by the addition of growth factors. In the absence of growth factors, cultured G0 cells express neither cyclins nor CDKs; the absence of these critical proteins explains why G0 cells do not progress through the cell cycle and replicate.
64 Figure Control of the G 1 S phase transition
65 Control of the G1 S phase transition 1. In yeast, the G1 cyclin gene is called CLN3 (Figure 19-15a). Its mrna is produced at a nearly constant level throughout the cell cycle, but its translation is regulated in response to nutrient levels and, as we will see shortly, CLN3 is a linchpin in coupling cell cycle entry to nutrient signals. Once sufficient Cln3 is synthesized from its mrna, Cln3-CDK complexes phosphorylate and inactivate the transcriptional repressor Whi5. Phosphorylation of Whi5 promotes its export out of the nucleus, allowing the transcription factor complex SBF to induce transcription of the G1/S phase cyclin genes CLN1 and CLN2 as well as other gene important for DNA replication. Once produced Cln1/2-CDKs contribute to further Whi5 phosphorylation. This positive feedback loop ensures the rapid accumulation of G1/S phase CDKs. Once a critical level of Cln1/2-CDKs is reached, these G1/S phase CDKs promote bud formation, entry into S phase, and the duplication of the centroseom (also known as the spindle pole body, which later in the cell cycle will organize the mitotic spindle). This state of G1/S phase CDK activity, sufficient to initiate S phase, bud formation, and centrosome duplication, is the molecular definition of START. 2. Some members of a small family of related transcription factors, referred to collectively as E2F factors, are encoded by delayed-response genes. These transcription factors activate genes encoding many of the proteins involved in DNA and deoxyribonucleotide synthesis. They also stimulate transcription of genes encoding the late-g1 cyclin (cyclin E), the S-phase cyclin (cyclin A), and the S-phase CDK (CDK2). Thus the E2Fs function in late G1 similarly to the S. cerevisiae transcription factors SBF and MBF. In addition, E2Fs auto stimulate transcription of their own genes. E2Fs function as transcriptional repressors when bound to Rb protein, which in turn binds histone deacetylase complexes. Transcription of a gene is highest when the associated histones are highly acetylated; histone deacetylation causes chromatin to assume a more condensed, transcriptionally inactive form. Rb protein is one of the most significant substrates of mammalian G1 cyclin-cdk complexes. Phosphorylation of Rb protein at multiple sites prevents its association with E2Fs, thereby permitting E2Fs to activate transcription of genes required for entry into S phase. As shown in Figure 19-15b, phosphorylation of Rb protein is initiated by cyclin D-CDK4 and cyclin D- CDK6 in mid G1.
66
67 Regulation of Rb and E2F activities 1. Mitogen withdrawal prior to passage through the restriction point leads to accumulation of p16. p16 binds specifically to CDK4 and CDK6, thereby inhibiting their kinase activity and causing G1 arrest. Under normal circumstances, phosphorylation of Rb protein is initiated midway through G1 by active cyclin D- CDK4 and cyclin D-CDK6 complexes. Rb phosphorylation is completed by other cyclin-cdk complexes in late G1, allowing activation of E2F transcription factors, which stimulate transcription of genes encoding proteins required for DNA synthesis. The complete phosphorylation of Rb irreversibly commits the cell to DNA synthesis. Most tumors contain an oncogenic mutation that causes overproduction or loss of one of the components of this pathway such that the cells are propelled into the S phase in the absence of the proper extracellular growth signals. 2. Once cyclin E and CDK2 are induced by phosphorylation of some Rb, the resulting cyclin E-CDK2 further phosphorylates Rb in late G1. When cyclin E-CDK2 accumulates to a critical threshold level, further phosphorylation of Rb by cyclin E-CDK2 continues even when cyclin D-CDK4/6 activity is removed. This is one of the principle biochemical events responsible for passage through the restriction point. At this point, further phosphorylation of Rb by cyclin E-CDK2 occurs even when mitogens are withdrawn and cyclin D and CDK4/6 levels fall. Since E2F stimulates its own expression and that of cyclin E and CDK2, positive cross-regulation of E2F and cyclin E-CDK2 produces a rapid rise of both activities in late G1. As they accumulate, S-phase cyclin-cdk and mitotic cyclin-cdk complexes maintain Rb protein in the phosphorylated state throughout the S, G2, and early M phases. After cells complete anaphase and enter early G1 or G0, the fall in cyclin-cdk levels leads to dephosphorylation of Rb by unopposed phosphatases. As a consequence, hypophosphorylated Rb is available to inhibit E2F activity during early G1 of the next cycle and in G0-arrested cells (Figure th ed).
68

69 Figure (part 3 of 3) Molecular Biology of the Cell ( Garland Science 2008)
70 Figure Fundamental processes in the eukaryotic cell cycle.
71 Mechanisms controlling S-phase initiation in animal cells The best understood effects of G 1 -Cdk activity in animal cells are mediated by a gene regulatory protein called E2F. It binds to specific DNA sequences in the promoters of many genes that encode proteins required for S-phase entry, including G 1 /S-cyclins and S-cyclins. E2F function is controlled primarily by an interaction with the retinoblastoma protein (Rb), an inhibitor of cell-cycle progression. During G 1, Rb binds to E2F and blocks the transcription of S-phase genes. When cells are stimulated to divide by extracellular signals, active G 1 -Cdk accumulates and phosphorylates Rb, reducing its affinity for E2F. The Rb then dissociates, allowing E2F to activate S-phase gene expression. This transcriptional control system, like so many other control systems that regulate the cell cycle, includes feedback loops that sharpen the G1/S transition : The liberated E2F increases the transcription of its own gene. E2F-dependent transcription of G1/S-cyclin and S-cyclin genes leads to increased G1/S-Cdk and S- Cdk activities, which in turn increase Rb phosphorylation and promote further E2F release, forming a positive feedback loop. The increase in G1/S-Cdk and S-Cdk activities enhances the phosphorylation of Hct1 and p27, leading to their inactivation or destruction. The resulting appearance of G1/S-Cdk and S-Cdk activities further enhances Rb phosphorylatione2f acts back to stimulate the transcription of its own gene, forming another positive feedback loop (Figure (part 3 of 3) Molecular Biology of the Cell -

72 Figure Control of S phase onset in S. cerevisiae by regulated proteolysis of the S phase inhibitor, Sic1.
73 Figure Six suboptimal G 1 /S phase CDK phosphorylation sites in Sic1 create a switch-like cell cycle entry.
74
75 Figure Fundamental processes in the eukaryotic cell cycle.
76 S phase inhibitor, Sic1 1. We can now see that regulated proteasomal degradation directed by two ubiquitin ligase complexes, SCF and APC, controls three major transitions in the cell cycle: onset of the S phase through degradation of Sic1, the beginning of anaphase through degradation of securin, and exit from mitosis through degradation of B-type cyclins. The APC is directed to polyubiquitinate securin, which functions as an anaphase inhibitor, by the Cdc20 specificity factor. Another specificity factor, Cdh1, targets APC to B-type cyclins. The SCF is directed to polyubiquitinate Sic1 by a different mechanism, namely, phosphorylation of Sic1 by a G1 cyclin-cdk (Figure 19-17, 19-30). This difference in strategy probably occurs because the APC has several substrates, including securin and B-type cyclins, which must be degraded at different times in the cycle. In contrast, entry into the S phase requires the degradation of only a single protein, the Sic1 inhibitor (Table 19-2). An obvious advantage of proteolysis for controlling passage through these critical points in the cell cycle is that protein degradation is an irreversible process, ensuring that cells proceed irreversibly in one direction through the cycle. 2. Degradation of Sic1 is induced by its phosphorylation by G1/S phase CDKs (Figure 19-17). It must be phosphorylated at least six sites, which are relatively poor substrates for the G1/S phase CDKs, before it is bound sufficiently well by SCF to be ubiquitinylated. Multiple, poor G1/S phase CDK phosphorylation sites lead to an ultrasensitive, switch-like response in Sic1 degradation and hence precipitous activation of S phase CDKs (Figure 19-18). If Sic1 were inactivated following the phosphorylation of a single site, Sic1 molecules would start to get phosphorylated as soon as the levels of G1/S phase CDK activity begin to rise, leading to a gradual decrease in Sic1 levels. In contrast, when several sites need to be phosphorylated, at low levels of G1/S phase CDK activity only a few sites are phosphorylated and Sic1 is not destroyed. Only when G1/S phase CDK levels are high is Sic1 sufficiently phosphorylated at multiple sites to target it for degradation. Thus Sic1 degradation occurs only when G1/S phase CDK activity has reached its peak and virtually all other G1/S phase CDK substrates have been phophorylated.
77 Figure The molecular mechanisms governing the initiation of DNA replication.
78 The molecular mechanisms governing the initiation of DNA replication 1. Eukaryotic chromosomes are replicated from multiple origins. Yeast replication origins contain an 11-bp conserved core sequence to which is bound a hexameric protein, the originrecognition complex (ORC), required for initiation of DNA synthesis. Cdc6 and Cdt1 are required to load opposing MCM helicase complex on the origin (Figure 19-19, step 1). The MCM helicase function to unwind the DNA during the initiation of DNA replication. To ensure that origins fire only once at the beginning of S phase, MCM helicase complex loading and its activation occur at two opposing phosphorylation states. 1 The MCM helicases can only load onto the DNA in a state of low CDK activity that occurs when CDKs are inactivated during exit from mitosis and during early G1. In other words, MCM helicases load onto DNA when they are unphosphorylated. 2 In contrast, activation of MCM helicases and the recruitment of DNA polymerases to the unwound origin DNA are triggered by S phase CDKs. Recall that S phase CDKs only become active when G1/S phase CDK levels reach their peak and CKIs of S phase CDKs and a second heterodimeric protein kinase, DDK, activates the MCM helicase and recruits the polymerases to the sites of replication initiation (Figure 19-19, steps 2 and 3). 2. So how do S phase CDKs and DDK collaborate to initiate DNA replication? 1 ORC and the two other initiation factors, Cdc6 and Cdt1, recruit the MCM helicases to sites of replication initiation during G1, when CDK activity is low (Figure 19-19, step 1). 2 When DDK and S phase CDKs are activated in late G1, DDK phosphorylates two subunits of the MCM helicase. The S phase CDKs phosphorylate two proteins called Sld2 and Sld3. These phosphorylation events have an activating effect, promoting the recruitment of MCM helicase activators to sites of replication initiation (the phosphorylation events shown in green in Figure 19-19, steps 2 and 3). 3 The helicase activators are called the Cdc45-Sld3 complex and the GINS complex. Exactly how they promote activation of the MCM helicases is not yet clear. In addition to activating the MCM helicase to unwind DNA, the Cdc45-Sld3 complex and the GINS complex recruit polymerases to the DNA polymerase ε to synthesize the leading strand and polymerase δ to synthesize the lagging strand (see Figure 19-19, step 3). The replication machinery then initiates DNA synthesis.

79 Figure Model for establishing cohesin linkage of sister chromatids.
80 Model for establishing cohesin linkage of sister chromatids The linkages between sister chromatids established during S phase will be essential for their accurate segregation during mitosis. The protein complexes that establish cohesion between sister chromatids are called cohesins. Some details of how cohesins are loaded onto DNA to mediate sister chromatid cohesion are being unraveled. We know that establishment of cohesion between sister chromatids is tightly tied to DNA replication. Cohesins associate with chromosomes during G1 (Figure 19-20, step 1). During DNA replication, they are loaded onto chromosomes in such a way that they can hold sister chromatids together. Most likely this occurs as replication forks replicate the DNA (step 2). Converting DNA-bound G1 cohesins into cohesive complexes requires several cohesin loading factors-including a protein complex related to proteins that load the sliding clamp at the replication fork-and the acetylation of the Smc3 subunit. 1. Step1: Cohesins are loaded onto chromosomes during G1, but they do not posses cohesive properties (indicated as cohesins laterally associated with chromosomes). 2. Step2: Concomitant with DNA replication, and most likely closely behind the replication fork, cohesins are converted into cohesive molecules, able to hold sister chromatids together (indicated as cohesin rings encircling the replicated sister chromatids). This conversion into cohesive cohesins requires cohesin loading factors. During G2, sister chromatids are replicated and linked along their entire length by cohesins. During this time, the Mei-S332/Sgo proteins recruit the protein phosphatase 2A (PP2A) to centromeric regions. 3. Step3: In vertebrate cells, cohesins are released from chromosome arms during prophase and early metaphase by the action of Polo kinase and Aurora B kinase. By the end of metaphase, cohesins are retained only in the region of the centromere, where PP2A prevents cohesin phosphorylation and hence dissociation through recruiting PP2A.
81
82 Figure Phosphorylation on the CDK subunit restrains mitotic CDK activity during S phase and G 2.
83 Phosphorylation on the CDK subunit restrains mitotic CDK activity during S phase and G2 Mitotic CDKs initiate mitosis. Whereas levels of the catalytic CDK subunit are constant throughout the cell cycle, mitotic cyclins gradually accumulate during S phase. Most eukaryotes contain multiple mitotic cyclins, which for historical reasons are called cyclin A and cyclin B. As they assemble, mitotic CDK complexes are maintained in an inactive state through inhibitory phosphorylation on the CDK subunit. Two highly conserved tyrosine and threonine residues in mammalian CDKs are subject to regulated phosphorylation. In CDK1, the mitotic CDK, phosphorylation of tyrosine 15 and threonine 14 maintains mitotic cyclin-cdk complexes in an inactivate state. The phosphorylation state of T14 and T15 is controlled by a dual-specificity protein kinase known as Wee1 and a dual-specificity phosphatase Cdc25 (Figure 19-21). The regulation of mitotic CDKs by these activities underlies the abrupt activation of their kinase activity at the G2-M phase transition and explains the observation that although mitotic cyclins gradually accumulate during S phase and G2, mitotic CDKs are not active until cells enter mitosis. Activation of mitotic CDKs is the consequence of rapid inactivation of Wee1 and activation of Cdc25. Central to this rapid transition are feedback loops, where mitotic CDKs activate Cdc25 and inactivate Wee1 (Figure 19-21). Phosphorylation of Cdc25 by mitotic CDKs stimulates its phosphatase activity; phosphorylation of Wee1 by mitotic CDKs inhibits its kinase activity. Ongoing DNA replication inhibits Cdc25 activity. A critical question that we know little about is how this positive feedback loop is started once DNA replication has been completed. CDKs that function earlier in the cell cycle have been suggested to start the positive feedback loop.
84
85 Figure Overview of DNA damage checkpoint controls in the cell cycle.
86 DNA damage checkpoint controls in the cell cycle 1. The complete and accurate duplication of the genetic material is essential for cell division. If cells enter mitosis when DNA is not completely replicated or otherwise damaged, genetic changes occur. In many instances, those changes will lead to cell death, they can also lead to genetic alterations that result in loss of growth and division control and eventually cancer. The enzymes that replicate DNA are highly accurate, but this exactness is not enough to ensure complete accuracy during DNA synthesis. Furthermore, environmental insults such as x-rays and UV light can cause DNA damage, and this damage must be repaired before a cell's entry into mitosis. Cells have a DNA damage response system in place that senses many different types of DNA damage and responds by activating repair pathways and halting cell cycle progression until the damage has been repaired. Cell cycle arrest can occur either in G1, S phase, or G2, depending on whether DNA damage occurred before cell cycle entry or during DNA replication. 2. Chk1 and Chk2 halt the cell cycle. The protein kinases phosphorylate Cdc25, thereby inactivating it. When the DNA damage occurs during G1, Cdc25A inhibition results in inhibition of G1/S phase CDKs and S phase CDKs (Figure 19-34). 1 As a result, these kinases cannot initiate DNA replication. When the DNA damage occurs during S phase or in G2, Cdc25C inhibition by Chk1/2 results in the inhibtion of mitotic CDKs and hence arrest in G2. 2 Active DNA replication also inhibits entry into mitosis. ATR continues to inhibit Cdc25C via Chk1 until all replication forks complete DNA replication and disassemble. This mechanism makes the initiation of mitosis dependent on the completion of chromosome replication. 3 Finally, cells also sense DNA replication stress that results in stalled or slowing of replication forks. It triggers activation of the ATR-Chk1 checkpoint pathway and results in down-regulation of S phase CDK activity and prevents firing of latereplicating origins. 4 Chk1-mediated inhibition of the Cdc25 family of phosphatases is not the only mechanism whereby DNA damage or incomplete replication inhibits cell cycle progression. DNA damage leads to the activation of the transcription factor p53, which transcribes the CDK inhibitor p21. p21 binds to and inhibits all metazoan cyclin-cdk complexes. As a result, cells are arrested in G1 and G2 (Figure 19-34). Under some circumstances, such as when DNA damage is extensive, p53 also activates expression of genes that lead to apoptosis, the process of programmed cell death that normally occurs in specific cells during the development of multicellular animals.
87 Figure The DNA damage response system.
88 The DNA damage response system 1. DNA damage exists in many different forms and ranges in severity. A break of the DNA helix, known as a double-strand break, is perhaps the most severe form of damage since such a lesion would almost certainly lead to DNA loss if mitosis ensued in its presence. More subtle defects include single-strand breaks, structural changes in nucleotides, or DNA mismatches. For our discussion here, it is important to note that cells have sensors for all these different types of damage. These sensors scan the genome and, when they detect a lesion, assemble signaling and repair factors onto the site of the lesion. 2. Central to the detection of the different lesions is a pair of homologous protein kinase called ATM and ATR. These protein kinases get recruited to sites of DNA damage. They then initiate the sequential recruitment of adapter proteins and another set of protein kinases called Chk1 and Chk2. These kinases then activate repair mechanisms and cause cell cycle arrest or apoptosis in animals (Figure 19-33). ATR and ATM recognize different types of DNA damage. ATM is very specialized in that it only responds to double-strand breaks. ATR is able to recognize more diverse types of DNA damage, such as stalled replication forks, damaged nucleotides, and double-strand breaks. Stalled replication forks, for example, are recognized by ATR. The association of ATR with stalled forks is thought to activate its protein kinase activity, leading to the recruitment of adapter proteins whose function is to recruit and help activate the Chk1 kinase. Active Chk1 then induces repair pathways and inhibits cell cycle progression. 3. A key effector of the DNA damage response in metazoan cells is the transcription factor p53 (Figure 19-33). It is known as a tumor suppressor because its normal function is to limit cell proliferation in the face of DNA damage. The protein is extremely unstable and generally does not accumulate to high enough levels to stimulate transcription under normal conditions. The instability of p53 results from its unbiquitinylation by a ubiquitin-protein ligase called Mdm2 and subsequent proteasomal degradation. The rapid degradation of p53 is inhibited by ATM and ATR, which phophorylate p53 at a site that interferes with Mdm2 binding. This and other modifications of p53 in response to DNA damage greatly increase its ability to activate transcription of specific genes that help the cell cope with DNA damage. One of these genes encodes the CKI p21.
89 Figure Molecular Biology of the Cell ( Garland Science 2008)
90 How DNA damage arrests the cell cycle in G1 1. When chromosomes are damaged, as can occur after exposure to radiation or certain chemicals, it is essential that they be repaired before the cell attempts to duplicate or segregate them. The cell-cycle control system can readily detect DNA damage and arrest the cycle at DNA damage checkpoints. Most cells have at least two such checkpoints one in late G1, which prevents entry into S phase, and one in late G2, which prevents entry into mitosis. The G2 checkpoint depends on a mechanism similar to the one discussed earlier that delays entry into mitosis in response to incomplete DNA replication. When cells in G2 are exposed to damaging radiation, for example, the damaged DNA sends a signal to a series of protein kinases that phosphorylate and inactivate the phosphatase Cdc25. This blocks the dephosphorylation and activation of M-Cdk, thereby blocking entry into mitosis. When the DNA damage is repaired, the inhibitory signal is turned off, and cell-cycle progression resumes. The G1 checkpoint blocks progression into S phase by inhibiting the activation of G1/S-Cdk and S-Cdk complexes. In mammalian cells, for example, DNA damage leads to the activation of the gene regulatory protein p53, which stimulates the transcription of several genes. One of these genes encodes a CKI protein called p21, which binds to G1/S-Cdk and S-Cdk and inhibits their activities, thereby helping to block entry into S phase. 2. DNA damage activates p53 by an indirect mechanism. In undamaged cells, p53 is highly unstable and is present at very low concentrations. This is because it interacts with another protein, Mdm2, that acts as a ubiquitin ligase that targets p53 for destruction by proteasomes. DNA damage activates protein kinases that phosphorylate p53 and thereby reduce its binding to Mdm2. This decreases p53 degradation, which results in a marked increase in p53 concentration in the cell. In addition, the decreased binding to Mdm2 enhances the ability of p53 to stimulate gene transcription. (Figure Molecular Biology of the Cell -
")
91 Nat Rev Cancer (2007) 7(11):
Cell Cycle. Trends in Cell Biology
 Cell Cycle Trends in Cell Biology Cyclic proteolysis Two distinct enzyme complexes proteolysis of cyclin-cdk complexes SCF (Skp1-Cul1-F box) Ubiquitylation and destruction of G1/S-cyclins and CKI proteins
Cell Cycle Trends in Cell Biology Cyclic proteolysis Two distinct enzyme complexes proteolysis of cyclin-cdk complexes SCF (Skp1-Cul1-F box) Ubiquitylation and destruction of G1/S-cyclins and CKI proteins
Section 10. Junaid Malek, M.D.
 Section 10 Junaid Malek, M.D. Cell Division Make sure you understand: How do cells know when to divide? (What drives the cell cycle? Why is it important to regulate this?) How is DNA replication regulated?
Section 10 Junaid Malek, M.D. Cell Division Make sure you understand: How do cells know when to divide? (What drives the cell cycle? Why is it important to regulate this?) How is DNA replication regulated?
7.06 Problem Set Four, 2006
 7.06 Problem Set Four, 2006 1. Explain the molecular mechanism behind each of the following events that occur during the cell cycle, making sure to discuss specific proteins that are involved in making
7.06 Problem Set Four, 2006 1. Explain the molecular mechanism behind each of the following events that occur during the cell cycle, making sure to discuss specific proteins that are involved in making
Chapter 3. DNA Replication & The Cell Cycle
 Chapter 3 DNA Replication & The Cell Cycle DNA Replication and the Cell Cycle Before cells divide, they must duplicate their DNA // the genetic material DNA is organized into strands called chromosomes
Chapter 3 DNA Replication & The Cell Cycle DNA Replication and the Cell Cycle Before cells divide, they must duplicate their DNA // the genetic material DNA is organized into strands called chromosomes
Human Anatomy & Physiology I Dr. Sullivan Unit IV Cellular Function Chapter 4, Chapter 27 (meiosis only)
") Human Anatomy & Physiology I Dr. Sullivan Unit IV Cellular Function Chapter 4, Chapter 27 (meiosis only) I. Protein Synthesis: creation of new proteins a. Much of the cellular machinery is devoted to synthesizing
Human Anatomy & Physiology I Dr. Sullivan Unit IV Cellular Function Chapter 4, Chapter 27 (meiosis only) I. Protein Synthesis: creation of new proteins a. Much of the cellular machinery is devoted to synthesizing
There are four phases: M = mitosis is when chromosomes become condensed and separate and the cell divides: visible changes. ~ 1 hr.
 The cell cycle in eukaryotes Jason Kahn, Biochem 465 Spring 2006 (Mostly from Alberts et al., Molecular Biology of the Cell, 2002). The book is available for free on line at http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4
The cell cycle in eukaryotes Jason Kahn, Biochem 465 Spring 2006 (Mostly from Alberts et al., Molecular Biology of the Cell, 2002). The book is available for free on line at http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4
Regulation of the cell cycle
 Regulation of the cell cycle Dr. F. Neuschäfer-Rube Cyclin-dependent Kinases (CDKs) Cyclin-dependent kinases: Motors and switches of the cell cycle Dr. F. Neuschäfer-Rube The cell cyle M S The cell cyle
Regulation of the cell cycle Dr. F. Neuschäfer-Rube Cyclin-dependent Kinases (CDKs) Cyclin-dependent kinases: Motors and switches of the cell cycle Dr. F. Neuschäfer-Rube The cell cyle M S The cell cyle
CELL BIOLOGY - CLUTCH CH CELL DIVISION.
 !! www.clutchprep.com CONCEPT: OVERVIEW OF THE CELL CYCLE There are three classes of cells based on whether they divide CELL TYPE Cells that do not divide Cells that normally do not divide, but can when
!! www.clutchprep.com CONCEPT: OVERVIEW OF THE CELL CYCLE There are three classes of cells based on whether they divide CELL TYPE Cells that do not divide Cells that normally do not divide, but can when
CHAPTER 16 THE CYTOSKELETON
 CHAPTER 16 THE CYTOSKELETON THE SELF-ASSEMBLY AND DYNAMIC STRUCTURE OF CYTOSKELETAL FILAMENTS HOW CELLS REGULATE THEIR CYTOSKELETAL FILAMENTS MOLECULAR MOTORS THE CYTOSKELETON AND CELL BEHAVIOR THE SELF-ASSEMBLY
CHAPTER 16 THE CYTOSKELETON THE SELF-ASSEMBLY AND DYNAMIC STRUCTURE OF CYTOSKELETAL FILAMENTS HOW CELLS REGULATE THEIR CYTOSKELETAL FILAMENTS MOLECULAR MOTORS THE CYTOSKELETON AND CELL BEHAVIOR THE SELF-ASSEMBLY
NUCLEUS. Fig. 2. Various stages in the condensation of chromatin
 NUCLEUS Animal cells contain DNA in nucleus (contains ~ 98% of cell DNA) and mitochondrion. Both compartments are surrounded by an envelope (double membrane). Nuclear DNA represents some linear molecules
NUCLEUS Animal cells contain DNA in nucleus (contains ~ 98% of cell DNA) and mitochondrion. Both compartments are surrounded by an envelope (double membrane). Nuclear DNA represents some linear molecules
Cell cycle oscillations
 Positive and negative feedback produce a cell cycle Fast Slow Cell cycle oscillations Active Cdk1-Cyclin Inactive Cdk1-Cyclin Active APC 1 Ubiquitin mediated proteolysis Glycine Isopeptide bond Lysine
Positive and negative feedback produce a cell cycle Fast Slow Cell cycle oscillations Active Cdk1-Cyclin Inactive Cdk1-Cyclin Active APC 1 Ubiquitin mediated proteolysis Glycine Isopeptide bond Lysine
Judy Wieber. Department of Computational Biology. May 28, 2008
 Review III: Cellular Processes Judy Wieber BBSI @ Pitt 2008 Department of Computational Biology University it of Pittsburgh School of Medicine i May 28, 2008 Outline Metabolism Cell cycle Transcription
Review III: Cellular Processes Judy Wieber BBSI @ Pitt 2008 Department of Computational Biology University it of Pittsburgh School of Medicine i May 28, 2008 Outline Metabolism Cell cycle Transcription
Vocab Word 1: Interphase
 Vocab Word 1: Interphase Interphase is the phase of the cell cycle in which a typical cell spends most of its life. During this phase, the cell copies its DNA in preparation for mitosis. Interphase is
Vocab Word 1: Interphase Interphase is the phase of the cell cycle in which a typical cell spends most of its life. During this phase, the cell copies its DNA in preparation for mitosis. Interphase is
Cell cycle: Cell growth and division (when the replication and segregation of chromosomes occurs). Interphase: Interphase Chromosomes
. Interphase: Interphase Chromosomes") Chromosomes exist in different states throughout the life of a cell Cell cycle: Cell growth and division (when the replication and segregation of chromosomes occurs). Interphase: Interphase Chromosomes
Chromosomes exist in different states throughout the life of a cell Cell cycle: Cell growth and division (when the replication and segregation of chromosomes occurs). Interphase: Interphase Chromosomes
Cell cycle. Chen Li. Department of cellular and genetic medicine
 Cell cycle Chen Li Department of cellular and genetic medicine 13 223 chenli2008@fudan.edu.cn Outline A. Historical background B. Phases of cell cycle C. DNA replication D. Telomere & telomerase E. DNA
Cell cycle Chen Li Department of cellular and genetic medicine 13 223 chenli2008@fudan.edu.cn Outline A. Historical background B. Phases of cell cycle C. DNA replication D. Telomere & telomerase E. DNA
Prof. R. V. Skibbens. Cell Cycle, Cell Division and Cancer
 Prof. R. V. Skibbens November 18, 2010 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer 4 Stages of the cell cycle: Genome segregation Interphase DNA Microtubules Microtubules
Prof. R. V. Skibbens November 18, 2010 BIOS 10: BioScience in the 21 st Century Cell Cycle, Cell Division and Cancer 4 Stages of the cell cycle: Genome segregation Interphase DNA Microtubules Microtubules
CELLULAR PROCESSES; REPRODUCTION. Unit 5
 CELLULAR PROCESSES; REPRODUCTION Unit 5 Cell Cycle Chromosomes and their make up Crossover Cytokines Diploid (haploid diploid and karyotypes) Mitosis Meiosis What is Cancer? Somatic Cells THE CELL CYCLE
CELLULAR PROCESSES; REPRODUCTION Unit 5 Cell Cycle Chromosomes and their make up Crossover Cytokines Diploid (haploid diploid and karyotypes) Mitosis Meiosis What is Cancer? Somatic Cells THE CELL CYCLE
Plant Molecular and Cellular Biology Lecture 9: Nuclear Genome Organization: Chromosome Structure, Chromatin, DNA Packaging, Mitosis Gary Peter
 Plant Molecular and Cellular Biology Lecture 9: Nuclear Genome Organization: Chromosome Structure, Chromatin, DNA Packaging, Mitosis Gary Peter 9/16/2008 1 Learning Objectives 1. List and explain how DNA
Plant Molecular and Cellular Biology Lecture 9: Nuclear Genome Organization: Chromosome Structure, Chromatin, DNA Packaging, Mitosis Gary Peter 9/16/2008 1 Learning Objectives 1. List and explain how DNA
Genetics and Genomics in Medicine Chapter 1. Questions & Answers
 Genetics and Genomics in Medicine Chapter 1 Multiple Choice Questions Questions & Answers Question 1.1 In a DNA double helix each type of base forms a stable base pair with only one type of base. When
Genetics and Genomics in Medicine Chapter 1 Multiple Choice Questions Questions & Answers Question 1.1 In a DNA double helix each type of base forms a stable base pair with only one type of base. When
Chapter 13. The Nucleus. The nucleus is the hallmark of eukaryotic cells; the very term eukaryotic means having a "true nucleus".
 Chapter 13 The Nucleus The nucleus is the hallmark of eukaryotic cells; the very term eukaryotic means having a "true nucleus". Fig.13.1. The EM of the Nucleus of a Eukaryotic Cell 13.1. The Nuclear Envelope
Chapter 13 The Nucleus The nucleus is the hallmark of eukaryotic cells; the very term eukaryotic means having a "true nucleus". Fig.13.1. The EM of the Nucleus of a Eukaryotic Cell 13.1. The Nuclear Envelope
CELL BIOLOGY - CLUTCH CH. 7 - GENE EXPRESSION.
 !! www.clutchprep.com CONCEPT: CONTROL OF GENE EXPRESSION BASICS Gene expression is the process through which cells selectively to express some genes and not others Every cell in an organism is a clone
!! www.clutchprep.com CONCEPT: CONTROL OF GENE EXPRESSION BASICS Gene expression is the process through which cells selectively to express some genes and not others Every cell in an organism is a clone
Control of Gene Expression
 Control of Gene Expression 1 How Gene Regulation Works 2 Control of Gene Expression Controlling gene expression is often accomplished by controlling transcription initiation Regulatory proteins bind to
Control of Gene Expression 1 How Gene Regulation Works 2 Control of Gene Expression Controlling gene expression is often accomplished by controlling transcription initiation Regulatory proteins bind to
PRINCIPLES O F NUCLEAR STRUCTUR E AND FUNCTIO N. Peter R. Cook
 PRINCIPLES O F NUCLEAR STRUCTUR E AND FUNCTIO N Peter R. Cook Preface xii i Acknowledgments xv 1. SOME PRINCIPLES 1 Overview of the Cell Nucleus 1 Box 1-1. Discovery of Cells, Nuclei, and DNA 2 A Sense
PRINCIPLES O F NUCLEAR STRUCTUR E AND FUNCTIO N Peter R. Cook Preface xii i Acknowledgments xv 1. SOME PRINCIPLES 1 Overview of the Cell Nucleus 1 Box 1-1. Discovery of Cells, Nuclei, and DNA 2 A Sense
Chapter 18: Regulation of Gene Expression. 1. Gene Regulation in Bacteria 2. Gene Regulation in Eukaryotes 3. Gene Regulation & Cancer
 Chapter 18: Regulation of Gene Expression 1. Gene Regulation in Bacteria 2. Gene Regulation in Eukaryotes 3. Gene Regulation & Cancer Gene Regulation Gene regulation refers to all aspects of controlling
Chapter 18: Regulation of Gene Expression 1. Gene Regulation in Bacteria 2. Gene Regulation in Eukaryotes 3. Gene Regulation & Cancer Gene Regulation Gene regulation refers to all aspects of controlling
1. I can describe the stages of the cell cycle.
 Unit 5 Study Guide Cell Cycle pg. 1 1. I can describe the stages of the cell cycle. Interphase = period in between division G1 = growth phase S = DNA replication G2 = Preparation for division (extra copies
Unit 5 Study Guide Cell Cycle pg. 1 1. I can describe the stages of the cell cycle. Interphase = period in between division G1 = growth phase S = DNA replication G2 = Preparation for division (extra copies
Lecture 6. Chromosome Structure and Function in Mitosis
 Lecture 6 Chromosome Structure and Function in Mitosis Outline: Chromosome Organization and Function in Interphase Chromatin effects on replication Effects of nuclear organization on gene expression Chromosomes
Lecture 6 Chromosome Structure and Function in Mitosis Outline: Chromosome Organization and Function in Interphase Chromatin effects on replication Effects of nuclear organization on gene expression Chromosomes
1. I can describe the stages of the cell cycle.
 Unit 5 Study Guide Cell Cycle pg. 1 1. I can describe the stages of the cell cycle. Interphase = period in between division G1 = growth phase S = DNA replication G2 = Preparation for division (extra copies
Unit 5 Study Guide Cell Cycle pg. 1 1. I can describe the stages of the cell cycle. Interphase = period in between division G1 = growth phase S = DNA replication G2 = Preparation for division (extra copies
Lecture 4 04/12/2018
 Lecture 4 04/12/2018 Communicating cells may be close (local) or far (distant) Local signaling: a chemical signal which communicates between 2 nearby cells. Paracrine signaling: a cell secretes the signal
Lecture 4 04/12/2018 Communicating cells may be close (local) or far (distant) Local signaling: a chemical signal which communicates between 2 nearby cells. Paracrine signaling: a cell secretes the signal
BIOLOGY 111. CHAPTER 6: DNA: The Molecule of Life
 BIOLOGY 111 CHAPTER 6: DNA: The Molecule of Life Chromosomes and Inheritance Learning Outcomes 6.1 Describe the structure of the DNA molecule and how this structure allows for the storage of information,
BIOLOGY 111 CHAPTER 6: DNA: The Molecule of Life Chromosomes and Inheritance Learning Outcomes 6.1 Describe the structure of the DNA molecule and how this structure allows for the storage of information,
Chapter 17 Lecture. Concepts of Genetics. Tenth Edition. Regulation of Gene Expression in Eukaryotes
 Chapter 17 Lecture Concepts of Genetics Tenth Edition Regulation of Gene Expression in Eukaryotes Chapter Contents 17.1 Eukaryotic Gene Regulation Can Occur at Any of the Steps Leading from DNA to Protein
Chapter 17 Lecture Concepts of Genetics Tenth Edition Regulation of Gene Expression in Eukaryotes Chapter Contents 17.1 Eukaryotic Gene Regulation Can Occur at Any of the Steps Leading from DNA to Protein
GENETICS. Chapter 1: Cell cycle. Thème 1 : La Terre dans l Univers A. Expression, stabilité et variation du patrimoine génétique.
 Introduction: GENETICS 3M = first look at genetics (study of inheritance, discovery of chromosomes, genes, dominant and recessive alleles and the DNA molecule within chromosomes) 2D = not much in fact,
Introduction: GENETICS 3M = first look at genetics (study of inheritance, discovery of chromosomes, genes, dominant and recessive alleles and the DNA molecule within chromosomes) 2D = not much in fact,
Section C: The Control of Gene Expression
 Section C: The Control of Gene Expression 1. Each cell of a multicellular eukaryote expresses only a small fraction of its genes 2. The control of gene expression can occur at any step in the pathway from
Section C: The Control of Gene Expression 1. Each cell of a multicellular eukaryote expresses only a small fraction of its genes 2. The control of gene expression can occur at any step in the pathway from
CHAPTER 17 FROM GENE TO PROTEIN. Section C: The Synthesis of Protein
 CHAPTER 17 FROM GENE TO PROTEIN Section C: The Synthesis of Protein 1. Translation is the RNA-directed synthesis of a polypeptide: a closer look 2. Signal peptides target some eukaryotic polypeptides to
CHAPTER 17 FROM GENE TO PROTEIN Section C: The Synthesis of Protein 1. Translation is the RNA-directed synthesis of a polypeptide: a closer look 2. Signal peptides target some eukaryotic polypeptides to
Control of Eukaryotic Gene Expression (Learning Objectives)
") Control of Eukaryotic Gene Expression (Learning Objectives) 1. Compare and contrast chromatin and chromosome: composition, proteins involved and level of packing. Explain the structure and function of
Control of Eukaryotic Gene Expression (Learning Objectives) 1. Compare and contrast chromatin and chromosome: composition, proteins involved and level of packing. Explain the structure and function of
B. Incorrect! Centromeric DNA is largely heterochromatin, which is inactive DNA.
 MCAT Biology - Problem Drill 06: Molecular Biology of Eukaryotes Question No. 1 of 10 1. Which type of DNA would have the highest level of expression? Question #01 (A) Heterochromatin. (B) Centromeric
MCAT Biology - Problem Drill 06: Molecular Biology of Eukaryotes Question No. 1 of 10 1. Which type of DNA would have the highest level of expression? Question #01 (A) Heterochromatin. (B) Centromeric
Lab 5 Cell Biology II
 Lab 5 Cell Biology II Learning Objectives Describe the structures and processes required for protein synthesis. Define a gene. Compare chromosomes and chromatin. Describe the process of DNA replication.
Lab 5 Cell Biology II Learning Objectives Describe the structures and processes required for protein synthesis. Define a gene. Compare chromosomes and chromatin. Describe the process of DNA replication.
AP Biology. The BIG Questions. Chapter 19. Prokaryote vs. eukaryote genome. Prokaryote vs. eukaryote genome. Why turn genes on & off?
 The BIG Questions Chapter 19. Control of Eukaryotic Genome How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions?
The BIG Questions Chapter 19. Control of Eukaryotic Genome How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions?
7.06 Cell Biology EXAM #2 March 20, 2003
 7.06 Cell Biology EXAM #2 March 20, 2003 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #2 March 20, 2003 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
The Nucleus, DNA and Chromatin Structure
 The Nucleus, DNA and Chromatin Structure Size of the human genom: The diploid human genome contains approximately 6 billion base pairs of DNA packaged into 23 chromosome pairs. Each diploid cell therefore
The Nucleus, DNA and Chromatin Structure Size of the human genom: The diploid human genome contains approximately 6 billion base pairs of DNA packaged into 23 chromosome pairs. Each diploid cell therefore
Y1 Biology 131 Syllabus - Academic Year
 Y1 Biology 131 Syllabus - Academic Year 2016-2017 Monday 28/11/2016 DNA Packaging Week 11 Tuesday 29/11/2016 Regulation of gene expression Wednesday 22/9/2014 Cell cycle Sunday 4/12/2016 Tutorial Monday
Y1 Biology 131 Syllabus - Academic Year 2016-2017 Monday 28/11/2016 DNA Packaging Week 11 Tuesday 29/11/2016 Regulation of gene expression Wednesday 22/9/2014 Cell cycle Sunday 4/12/2016 Tutorial Monday
MCB130 midterm exam. 1) The exam has 6 pages (not including this page). Check that you have all 6 pages
 The exam has 6 pages (not including this page). Check that you have all 6 pages") MCB130 midterm exam 1) The exam has 6 pages (not including this page). Check that you have all 6 pages 2) Please write your name and SID on the top of each page. The pages will be separated for grading
MCB130 midterm exam 1) The exam has 6 pages (not including this page). Check that you have all 6 pages 2) Please write your name and SID on the top of each page. The pages will be separated for grading
Bio Study Guide. What is the structure of a DNA. A nitrogen base (ATCG/AUCG) connected to a sugar and a phosphate. nucleotide?
 connected to a sugar and a phosphate. nucleotide?") What are nucleic acids? The largest organic molecules. Master organisms, they are in control of cell functions. They are found in all living organisms, and they are two types of them- DNA and RNA. They
What are nucleic acids? The largest organic molecules. Master organisms, they are in control of cell functions. They are found in all living organisms, and they are two types of them- DNA and RNA. They
Cell Nucleus. Chen Li. Department of Cellular and Genetic Medicine
 Cell Nucleus Chen Li Department of Cellular and Genetic Medicine 13 223 chenli2008@fudan.edu.cn Outline A. Historical background B. Structure of the nucleus: nuclear pore complex (NPC), lamina, nucleolus,
Cell Nucleus Chen Li Department of Cellular and Genetic Medicine 13 223 chenli2008@fudan.edu.cn Outline A. Historical background B. Structure of the nucleus: nuclear pore complex (NPC), lamina, nucleolus,
Cells and Tissues. Overview CELLS
 Cells and Tissues WIll The basic unit of structure and function in the human body is the cell. Each of a cell's parts, or organelles, as well as the entire cell, is organized to perform a specific function.
Cells and Tissues WIll The basic unit of structure and function in the human body is the cell. Each of a cell's parts, or organelles, as well as the entire cell, is organized to perform a specific function.
1. What is DNA? 2. List anything you know about DNA. (from readings, class, TV?)
") 1. What is DNA? 2. List anything you know about DNA (from readings, class, TV?) Before we begin, let s investigate the way DNA molecules are set up! http://learn.genetics.utah.edu/content/molec ules/builddna/
1. What is DNA? 2. List anything you know about DNA (from readings, class, TV?) Before we begin, let s investigate the way DNA molecules are set up! http://learn.genetics.utah.edu/content/molec ules/builddna/
Quick Review of Protein Synthesis
 Collin College BIOL. 2401 Quick Review of Protein Synthesis. Proteins and Protein Synthesis Proteins are the molecular units that do most of the work in a cell. They function as molecular catalysts, help
Collin College BIOL. 2401 Quick Review of Protein Synthesis. Proteins and Protein Synthesis Proteins are the molecular units that do most of the work in a cell. They function as molecular catalysts, help
Chapter 12: Molecular Biology of the Gene
 Biology Textbook Notes Chapter 12: Molecular Biology of the Gene p. 214-219 The Genetic Material (12.1) - Genetic Material must: 1. Be able to store information that pertains to the development, structure,
Biology Textbook Notes Chapter 12: Molecular Biology of the Gene p. 214-219 The Genetic Material (12.1) - Genetic Material must: 1. Be able to store information that pertains to the development, structure,
1. What is DNA? 2. List anything you know about DNA. (from readings, class, TV?)
") 1. What is DNA? 2. List anything you know about DNA (from readings, class, TV?) Before we begin, let s investigate the way DNA molecules are set up! http://learn.genetics.utah.edu/content/molec ules/builddna/
1. What is DNA? 2. List anything you know about DNA (from readings, class, TV?) Before we begin, let s investigate the way DNA molecules are set up! http://learn.genetics.utah.edu/content/molec ules/builddna/
DNA is the genetic material found in cells Stands for: Deoxyribonucleic Acid Is made up of repeating nucleic acids It s the Unit of Heredity
 What is DNA? DNA is the genetic material found in cells Stands for: Deoxyribonucleic Acid Is made up of repeating nucleic acids It s the Unit of Heredity DNA is found in the cytoplasm of prokaryotes and
What is DNA? DNA is the genetic material found in cells Stands for: Deoxyribonucleic Acid Is made up of repeating nucleic acids It s the Unit of Heredity DNA is found in the cytoplasm of prokaryotes and
Exam notes. Taxol stabilizes MTs and prevents cell division. Testing the role of MTs using pharmacological agents - inhibitors
 Exam notes Means are 76, 70 and? Mean of means is Approximate grades after 3 exams Testing the role of MTs using pharmacological agents - inhibitors Screens for anti-cancer agents have identified a variety
Exam notes Means are 76, 70 and? Mean of means is Approximate grades after 3 exams Testing the role of MTs using pharmacological agents - inhibitors Screens for anti-cancer agents have identified a variety
1. What is DNA? 2. List anything you know about DNA. (from readings, class, TV?)
") 1. What is DNA? 2. List anything you know about DNA (from readings, class, TV?) Before we begin, let s investigate the way DNA molecules are set up! http://learn.genetics.utah.edu/content/molec ules/builddna/
1. What is DNA? 2. List anything you know about DNA (from readings, class, TV?) Before we begin, let s investigate the way DNA molecules are set up! http://learn.genetics.utah.edu/content/molec ules/builddna/
REVIEW SHEET: Units 9 & 10 Cell Cycle, DNA, & Gene Expression
 REVIEW SHEET: Units 9 & 10 Cell Cycle, DNA, & Gene Expression HONORS BIOLOGY Textbook Reading: Cell Cycle (Ch. 10.1 and 10.2), DNA (Ch. 12), and Gene Expression (Ch. 13) Handouts:! Online Tutorial: Cell
REVIEW SHEET: Units 9 & 10 Cell Cycle, DNA, & Gene Expression HONORS BIOLOGY Textbook Reading: Cell Cycle (Ch. 10.1 and 10.2), DNA (Ch. 12), and Gene Expression (Ch. 13) Handouts:! Online Tutorial: Cell
Cell Division. embryo: an early stage of development in organisms
 Over the past several years, a debate has been brewing over the use of stem cells. Stem cells can be used to treat certain diseases and conditions such as spinal cord injuries, diabetes, arthritis, and
Over the past several years, a debate has been brewing over the use of stem cells. Stem cells can be used to treat certain diseases and conditions such as spinal cord injuries, diabetes, arthritis, and
Cell cycle oscillations. Active Cdk1-Cyclin Inactive Cdk1-Cyclin Active APC
 Cell cycle oscillations Active Cdk1-Cyclin Inactive Cdk1-Cyclin Active APC 20 Ubiquitin mediated proteolysis Glycine Isopeptide bond Lysine Many biological processes are regulated by controlling the stability
Cell cycle oscillations Active Cdk1-Cyclin Inactive Cdk1-Cyclin Active APC 20 Ubiquitin mediated proteolysis Glycine Isopeptide bond Lysine Many biological processes are regulated by controlling the stability
Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Chapter 18 Regulation of Gene Expression Edited by Shawn Lester PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley
Unit IX Problem 3 Genetics: Basic Concepts in Molecular Biology
 Unit IX Problem 3 Genetics: Basic Concepts in Molecular Biology - The central dogma (principle) of molecular biology: Information from DNA are transcribed to mrna which will be further translated to synthesize
Unit IX Problem 3 Genetics: Basic Concepts in Molecular Biology - The central dogma (principle) of molecular biology: Information from DNA are transcribed to mrna which will be further translated to synthesize
Chapter 11: Regulation of Gene Expression
 Chapter Review 1. It has long been known that there is probably a genetic link for alcoholism. Researchers studying rats have begun to elucidate this link. Briefly describe the genetic mechanism found
Chapter Review 1. It has long been known that there is probably a genetic link for alcoholism. Researchers studying rats have begun to elucidate this link. Briefly describe the genetic mechanism found
Lecture Overview. Overview of the Genetic Information. Marieb s Human Anatomy and Physiology. Chapter 3 DNA & RNA Protein Synthesis Lecture 6
 Marieb s Human Anatomy and Physiology Marieb Hoehn Chapter 3 DNA & RNA Protein Synthesis Lecture 6 Lecture Overview The Genetic Information Structure of DNA/RNA DNA Replication Overview of protein synthesis
Marieb s Human Anatomy and Physiology Marieb Hoehn Chapter 3 DNA & RNA Protein Synthesis Lecture 6 Lecture Overview The Genetic Information Structure of DNA/RNA DNA Replication Overview of protein synthesis
Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site.
 Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site. Still having trouble understanding the material? Check
Find this material useful? You can help our team to keep this site up and bring you even more content consider donating via the link on our site. Still having trouble understanding the material? Check
Cell Division. Use Target Reading Skills. This section explains how cells grow and divide.
 Name Date Class Cell Processes Guided Reading and Study Cell Division This section explains how cells grow and divide. Use Target Reading Skills As you read, make a cycle diagram that shows the events
Name Date Class Cell Processes Guided Reading and Study Cell Division This section explains how cells grow and divide. Use Target Reading Skills As you read, make a cycle diagram that shows the events
ZOO-302CR:(1.4)CELL DIVISION AND CELL CYCLE
CELL DIVISION AND CELL CYCLE") ZOO-302CR:(1.4)CELL DIVISION AND CELL CYCLE Cell Division, Mitosis, and Meiosis https://bio100.class.uic.edu/lecturesf04am/lect16.htm Cell Division Functions in Reproduction, Growth, and Repair Cell division
ZOO-302CR:(1.4)CELL DIVISION AND CELL CYCLE Cell Division, Mitosis, and Meiosis https://bio100.class.uic.edu/lecturesf04am/lect16.htm Cell Division Functions in Reproduction, Growth, and Repair Cell division
Genetics and Genomics in Medicine Chapter 1 Questions
 Genetics and Genomics in Medicine Chapter 1 Questions Multiple Choice Questions Question 1.1 In a DNA double helix each type of base forms a stable base pair with only one type of base. When bases on an
Genetics and Genomics in Medicine Chapter 1 Questions Multiple Choice Questions Question 1.1 In a DNA double helix each type of base forms a stable base pair with only one type of base. When bases on an
CHAPTER 18 LECTURE NOTES: CONTROL OF GENE EXPRESSION PART B: CONTROL IN EUKARYOTES
 CHAPTER 18 LECTURE NOTES: CONTROL OF GENE EXPRESSION PART B: CONTROL IN EUKARYOTES I. Introduction A. No operon structures in eukaryotes B. Regulation of gene expression is frequently tissue specific.
CHAPTER 18 LECTURE NOTES: CONTROL OF GENE EXPRESSION PART B: CONTROL IN EUKARYOTES I. Introduction A. No operon structures in eukaryotes B. Regulation of gene expression is frequently tissue specific.
Cells: The Living Units
 Chapter 3 Part D Cells: The Living Units Annie Leibovitz/Contact Press Images PowerPoint Lecture Slides prepared by Karen Dunbar Kareiva Ivy Tech Community College 3.10 Cell Cycle Series of changes a cell
Chapter 3 Part D Cells: The Living Units Annie Leibovitz/Contact Press Images PowerPoint Lecture Slides prepared by Karen Dunbar Kareiva Ivy Tech Community College 3.10 Cell Cycle Series of changes a cell
Chromosomes. M.Sc. Biotechnology. Hawler Medical University, Iraq
 Chromosomes Bashdar Mahmud Hussen M.Sc. Biotechnology Hawler Medical University, Iraq bashdar@res.hmu.edu.iq bmhscience@yahoo.com History of Chromosome Karl Nagali (1842) E. Russow (1872) first description
Chromosomes Bashdar Mahmud Hussen M.Sc. Biotechnology Hawler Medical University, Iraq bashdar@res.hmu.edu.iq bmhscience@yahoo.com History of Chromosome Karl Nagali (1842) E. Russow (1872) first description
Control of Eukaryotic Genes. AP Biology
 Control of Eukaryotic Genes The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions? Evolution
Control of Eukaryotic Genes The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions? Evolution
Human Biology 175 Lecture Notes: Cells. Section 1 Introduction
 Human Biology 175 Lecture Notes: Cells Section 1 Introduction A) 1) All organisms are composed of at least one cell. a) Unicellular example: b) Multicellular example(s): 2) All cells are similar in their
Human Biology 175 Lecture Notes: Cells Section 1 Introduction A) 1) All organisms are composed of at least one cell. a) Unicellular example: b) Multicellular example(s): 2) All cells are similar in their
Chapter 2. An Introduction to Genes and Genomes
 PowerPoint Lectures for Introduction to Biotechnology, Second Edition William J.Thieman and Michael A.Palladino Chapter 2 An Introduction to Genes and Genomes Lectures by Lara Dowland Chapter Contents
PowerPoint Lectures for Introduction to Biotechnology, Second Edition William J.Thieman and Michael A.Palladino Chapter 2 An Introduction to Genes and Genomes Lectures by Lara Dowland Chapter Contents
Protein Synthesis
 HEBISD Student Expectations: Identify that RNA Is a nucleic acid with a single strand of nucleotides Contains the 5-carbon sugar ribose Contains the nitrogen bases A, G, C and U instead of T. The U is
HEBISD Student Expectations: Identify that RNA Is a nucleic acid with a single strand of nucleotides Contains the 5-carbon sugar ribose Contains the nitrogen bases A, G, C and U instead of T. The U is
Control of Eukaryotic Genes. AP Biology
 Control of Eukaryotic Genes The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions? Evolution
Control of Eukaryotic Genes The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions? Evolution
These molecules make up the ladder of the DNA Bound by weak hydrogen bonds. 4 Different Types (2 specific matches) look at the
 look at the") B.A.T. Review DNA & Cell Cycle Test is 11/3/16 NAME: PERIOD DNA STRUCTURE VOCABULARY Mitosis Chromosomes Interphase G1,S,G2,M Prophase Metaphase Anaphase Telophase Cytokinesis Cleavage Furrow Spindle Fibers
B.A.T. Review DNA & Cell Cycle Test is 11/3/16 NAME: PERIOD DNA STRUCTURE VOCABULARY Mitosis Chromosomes Interphase G1,S,G2,M Prophase Metaphase Anaphase Telophase Cytokinesis Cleavage Furrow Spindle Fibers
DNA STRUCTURE. Nucleotides: Nitrogenous Bases (Carry the Genetic Code) Expectation Sheet: DNA & Cell Cycle. I can statements: Basic Information:
 Expectation Sheet: DNA & Cell Cycle. I can statements: Basic Information:") Expectation Sheet: DNA & Cell Cycle NAME: Test is 11/8/17 I can statements: I can discuss how DNA is found in all organisms and that the structure is common to all living things. I can diagram and label
Expectation Sheet: DNA & Cell Cycle NAME: Test is 11/8/17 I can statements: I can discuss how DNA is found in all organisms and that the structure is common to all living things. I can diagram and label
Practice Problems 5. Location of LSA-GFP fluorescence
 Life Sciences 1a Practice Problems 5 1. Soluble proteins that are normally secreted from the cell into the extracellular environment must go through a series of steps referred to as the secretory pathway.
Life Sciences 1a Practice Problems 5 1. Soluble proteins that are normally secreted from the cell into the extracellular environment must go through a series of steps referred to as the secretory pathway.
amino acid nucleic acid nucleotide DNA/RNA enzymes lock and key model catalyst carbohydrate monosaccharide glucose
 Unit 1: Biomolecules I. Terms You Should Know lipid fatty acid & glycerol monomer biomolecule protein amino acid nucleic acid nucleotide DNA/RNA enzymes lock and key model catalyst carbohydrate monosaccharide
Unit 1: Biomolecules I. Terms You Should Know lipid fatty acid & glycerol monomer biomolecule protein amino acid nucleic acid nucleotide DNA/RNA enzymes lock and key model catalyst carbohydrate monosaccharide
CELL BIOLOGY: DNA. Generalized nucleotide structure: NUCLEOTIDES: Each nucleotide monomer is made up of three linked molecules:
 BIOLOGY 12 CELL BIOLOGY: DNA NAME: IMPORTANT FACTS: Nucleic acids are organic compounds found in all living cells and viruses. Two classes of nucleic acids: 1. DNA = ; found in the nucleus only. 2. RNA
BIOLOGY 12 CELL BIOLOGY: DNA NAME: IMPORTANT FACTS: Nucleic acids are organic compounds found in all living cells and viruses. Two classes of nucleic acids: 1. DNA = ; found in the nucleus only. 2. RNA
Biology Lecture 2 Genes
 Genes Definitions o Gene: DNA that codes for a single polypeptide/mrna/rrna/trna o Euchromatin: region of DNA containing genes being actively transcribed o Heterochromatin: region of DNA containing genes
Genes Definitions o Gene: DNA that codes for a single polypeptide/mrna/rrna/trna o Euchromatin: region of DNA containing genes being actively transcribed o Heterochromatin: region of DNA containing genes
Chromatin. Structure and modification of chromatin. Chromatin domains
 Chromatin Structure and modification of chromatin Chromatin domains 2 DNA consensus 5 3 3 DNA DNA 4 RNA 5 ss RNA forms secondary structures with ds hairpins ds forms 6 of nucleic acids Form coiling bp/turn
Chromatin Structure and modification of chromatin Chromatin domains 2 DNA consensus 5 3 3 DNA DNA 4 RNA 5 ss RNA forms secondary structures with ds hairpins ds forms 6 of nucleic acids Form coiling bp/turn
Genetics Lecture Notes Lectures 6 9
 Genetics Lecture Notes 7.03 2005 Lectures 6 9 Lecture 6 Until now our analysis of genes has focused on gene function as determined by phenotype differences brought about by different alleles or by a direct
Genetics Lecture Notes 7.03 2005 Lectures 6 9 Lecture 6 Until now our analysis of genes has focused on gene function as determined by phenotype differences brought about by different alleles or by a direct
Unit 4: Cell Development and Replication, Part I: The Cell Cycle
 Name: Block: PACKET #9 Reading: Unit 4: Cell Development and Replication, Part I: The Cell Cycle Objectives: By the conclusion of this unit you will be able to: Date: Introduction to the cell cycle 1.
Name: Block: PACKET #9 Reading: Unit 4: Cell Development and Replication, Part I: The Cell Cycle Objectives: By the conclusion of this unit you will be able to: Date: Introduction to the cell cycle 1.
Chapter 18. Regulation of Gene Expression
 Chapter 18 Regulation of Gene Expression 2007-2008 Control of Prokaryotic (Bacterial) Genes 2007- Bacterial metabolism Bacteria need to respond quickly to changes in their environment STOP GO if they have
Chapter 18 Regulation of Gene Expression 2007-2008 Control of Prokaryotic (Bacterial) Genes 2007- Bacterial metabolism Bacteria need to respond quickly to changes in their environment STOP GO if they have
Chapter 18: Regulation of Gene Expression. Gene Regulation. Transcription Factors 3/21/2017
 Chapter 18: Regulation of Gene Expression 1. Gene Regulation in Bacteria 2. Gene Regulation in Eukaryotes 3. Gene Regulation & Cancer Gene Regulation Gene regulation refers to all aspects of controlling
Chapter 18: Regulation of Gene Expression 1. Gene Regulation in Bacteria 2. Gene Regulation in Eukaryotes 3. Gene Regulation & Cancer Gene Regulation Gene regulation refers to all aspects of controlling
Chapter 14 Regulation of Transcription
 Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
Chapter 14 Regulation of Transcription Cis-acting sequences Distance-independent cis-acting elements Dissecting regulatory elements Transcription factors Overview transcriptional regulation Transcription
Translation Mechanisms
 Translation Mechanisms Biology I Hayder A. Giha Translation The translation is the process of protein synthesis, where information in nucleotides sequences of a mrna is translated into amino acids sequence
Translation Mechanisms Biology I Hayder A. Giha Translation The translation is the process of protein synthesis, where information in nucleotides sequences of a mrna is translated into amino acids sequence
4.1 CELL DIVISION AND GENETIC MATERIAL
 4.1 CELL DIVISION AND GENETIC MATERIAL GENETICS Field of biology Study how genetic information is passed from one generation of organism/cells to the next THE CELL THEORY developed in mid-1800s 1. All
4.1 CELL DIVISION AND GENETIC MATERIAL GENETICS Field of biology Study how genetic information is passed from one generation of organism/cells to the next THE CELL THEORY developed in mid-1800s 1. All
- Does not stop at the formation of a mature organism but continues in certain tissues throughout
 CHAPTER 14: CELL DIVISION Cell division new cells originate only from other living cell - Does not stop at the formation of a mature organism but continues in certain tissues throughout THE CELL CYCLE
CHAPTER 14: CELL DIVISION Cell division new cells originate only from other living cell - Does not stop at the formation of a mature organism but continues in certain tissues throughout THE CELL CYCLE
5. Which of the following enzymes catalyze the attachment of an amino acid to trna in the formation of aminoacyl trna?
 Sample Examination Questions for Exam 3 Material Biology 3300 / Dr. Jerald Hendrix Warning! These questions are posted solely to provide examples of past test questions. There is no guarantee that any
Sample Examination Questions for Exam 3 Material Biology 3300 / Dr. Jerald Hendrix Warning! These questions are posted solely to provide examples of past test questions. There is no guarantee that any
Unit 1 Human cells. 1. Division and differentiation in human cells
 Unit 1 Human cells 1. Division and differentiation in human cells Stem cells Describe the process of differentiation. Explain how differentiation is brought about with reference to genes. Name the two
Unit 1 Human cells 1. Division and differentiation in human cells Stem cells Describe the process of differentiation. Explain how differentiation is brought about with reference to genes. Name the two
GENE EXPRESSSION. Promoter sequence where RNA polymerase binds. Operator sequence that acts as a switch (yellow) OPERON
 OPERON") GENE EXPRESSSION 1 GENE REGULATION IN PROKARYOTES Bacteria can turn genes on or off depending on their environment Prokaryotes have operons clusters of related genes and regulatory sequences Promoter sequence
GENE EXPRESSSION 1 GENE REGULATION IN PROKARYOTES Bacteria can turn genes on or off depending on their environment Prokaryotes have operons clusters of related genes and regulatory sequences Promoter sequence
Biotechnology Unit 3: DNA to Proteins. From DNA to RNA
 From DNA to RNA Biotechnology Unit 3: DNA to Proteins I. After the discovery of the structure of DNA, the major question remaining was how does the stored in the 4 letter code of DNA direct the and of
From DNA to RNA Biotechnology Unit 3: DNA to Proteins I. After the discovery of the structure of DNA, the major question remaining was how does the stored in the 4 letter code of DNA direct the and of
Characterizing the Relationship Between Cell-Cycle Progression. and a Transcriptional Oscillator
 Characterizing the Relationship Between Cell-Cycle Progression and a Transcriptional Oscillator by Sara Lynn Bristow University Program in Genetics and Genomics Duke University Date: Approved: Steven Haase,
Characterizing the Relationship Between Cell-Cycle Progression and a Transcriptional Oscillator by Sara Lynn Bristow University Program in Genetics and Genomics Duke University Date: Approved: Steven Haase,
Control of Eukaryotic Genes
 Control of Eukaryotic Genes 2007-2008 The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions?
Control of Eukaryotic Genes 2007-2008 The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions?
Figure S1, related to Figure 1. Characterization of biosensor behavior in vivo.
 Developmental Cell, Volume 23 Supplemental Information Separase Biosensor Reveals that Cohesin Cleavage Timing Depends on Phosphatase PP2A Cdc55 Regulation Gilad Yaakov, Kurt Thorn, and David O. Morgan
Developmental Cell, Volume 23 Supplemental Information Separase Biosensor Reveals that Cohesin Cleavage Timing Depends on Phosphatase PP2A Cdc55 Regulation Gilad Yaakov, Kurt Thorn, and David O. Morgan
Unit 7. Genetic Regulation, Development, and Biotechnology. AP Biology
 Unit 7 Genetic Regulation, Development, and Biotechnology The BIG Questions How are genes turned on & off in eukaryotes and prokaryotes? How do cells with the same genes differentiate to perform completely
Unit 7 Genetic Regulation, Development, and Biotechnology The BIG Questions How are genes turned on & off in eukaryotes and prokaryotes? How do cells with the same genes differentiate to perform completely
GENE REGULATION slide shows by Kim Foglia modified Slides with blue edges are Kim s
 GENE REGULATION slide shows by Kim Foglia modified Slides with blue edges are Kim s 2007-2008 Bacterial metabolism Bacteria need to respond quickly to changes in their environment STOP GO if they have
GENE REGULATION slide shows by Kim Foglia modified Slides with blue edges are Kim s 2007-2008 Bacterial metabolism Bacteria need to respond quickly to changes in their environment STOP GO if they have
Epigenetics in. Saccharomyces cerevisiae. Chapter 4 2/4/14
 Epigenetics in Saccharomyces cerevisiae Chapter 4 2/4/14 The budding yeast - Saccharomyces cerevisiae The fission yeast - Schizosaccharomyces pombe The budding yeast, Saccharomyces cerevisiae and the fission
Epigenetics in Saccharomyces cerevisiae Chapter 4 2/4/14 The budding yeast - Saccharomyces cerevisiae The fission yeast - Schizosaccharomyces pombe The budding yeast, Saccharomyces cerevisiae and the fission
Control of Eukaryotic Genes. AP Biology
 Control of Eukaryotic Genes The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions? Evolution
Control of Eukaryotic Genes The BIG Questions How are genes turned on & off in eukaryotes? How do cells with the same genes differentiate to perform completely different, specialized functions? Evolution
October 24, BIOS 95: Cell Division and Chromosome Dynamics
 October 24, 2008 BIOS 95: Cell Division and Chromosome Dynamics Cancer unregulated cell growth and migration Aneuploidy errors in chromosome segregation and genome maintenance living with an altered genome
October 24, 2008 BIOS 95: Cell Division and Chromosome Dynamics Cancer unregulated cell growth and migration Aneuploidy errors in chromosome segregation and genome maintenance living with an altered genome
Section 10.3 Outline 10.3 How Is the Base Sequence of a Messenger RNA Molecule Translated into Protein?
 Section 10.3 Outline 10.3 How Is the Base Sequence of a Messenger RNA Molecule Translated into Protein? Messenger RNA Carries Information for Protein Synthesis from the DNA to Ribosomes Ribosomes Consist
Section 10.3 Outline 10.3 How Is the Base Sequence of a Messenger RNA Molecule Translated into Protein? Messenger RNA Carries Information for Protein Synthesis from the DNA to Ribosomes Ribosomes Consist
DNA. Griffith s Transforming Principle Experiment 11/30/2006 DNA 2
 DNA Griffith s Transforming Principle Experiment 11/30/2006 DNA 2 1 Avery, McCarty, & MacLeod 1944 Extended Griffith s work 16 years later Search for the transforming factor Live rough cells + Protein
DNA Griffith s Transforming Principle Experiment 11/30/2006 DNA 2 1 Avery, McCarty, & MacLeod 1944 Extended Griffith s work 16 years later Search for the transforming factor Live rough cells + Protein
Part2. University of Technology Applied Sciences Department Biotechnology Division. Genetics. 2 nd class. Produced by L. Wasnaa H.
 University of Technology Applied Sciences Department Biotechnology Division 2 nd class Produced by L. Wasnaa H. Mohammad Part 2 2013 Cell Division or cell cycle When cells reproduce, the genetic material
University of Technology Applied Sciences Department Biotechnology Division 2 nd class Produced by L. Wasnaa H. Mohammad Part 2 2013 Cell Division or cell cycle When cells reproduce, the genetic material